Effects of 10 Hz rTMS on the Neural
Efficiency of Working Memory
Gilbert Preston1, Erik Anderson1, Claudio Silva1
Terry Goldberg2, and Eric M. Wassermann3
Abstrakt
■ Working memory ( WM) has been described as short-term
retention of information that is no longer accessible in the en-
vironment, and the manipulation of this information for sub-
sequent use in guiding behavior. WM is viewed as a cognitive
process underlying higher-order cognitive functions. Beweis
supports a critical role for PFC in mediating WM performance.
Studies show psychomotor processing speed and accuracy
account for considerable variance in neural efficiency (Ne). Das
study compared the relative effects of active and sham 10 Hz
rTMS applied to dorsolateral prefrontal cortex (DLPFC) on indi-
ces of Ne in healthy participants performing a WM paradigm
that models the association between WM load and task be-
havior [Sternberg, S. High-speed scanning in human memory.
Wissenschaft, 153, 652–654, 1966]. Previous studies identified a rela-
tionship between diminished Ne and impaired WM across a
broad array of clinical disorders. In der vorliegenden Studie, Die
authors predicted there would be a main effect of stimulation
Gruppe (STM) on accuracy (SCR) and processing speed (RT),
somit, Ne. We observed a main effect of STM for RT without an
effect on SCR; even so, there was a robust effect of STM on Ne. ■
EINFÜHRUNG
Working memory ( WM) has been described as short-
term retention of information that is no longer accessible
in the environment, and the manipulation of this informa-
tion for subsequent use in guiding behavior (DʼEsposito,
Postle, & Rypma, 2000). WM is widely viewed as a cognitive
process that underlies an array of higher order cogni-
tive functions, such as reasoning (Rypma, Prabhakaran,
Desmond, & Gabriela, 2001), planning (Goel & Grafman,
1995), and problem solving (Duncan & Owen, 2000), für
Beispiel. Over the past few decades, such studies have de-
veloped a substantial body of convergent evidence that
supports a critical role for prefrontal cortex (PFC) in medi-
ating WM performance. Zum Beispiel, in a study aimed to
identify the relationship between WM demand and task
Leistung, Rypma, Berger, and DʼEsposito (2002) found
that task factors exert their effects largely during WM
encoding. Subject factorsʼ influence on task performance
occurred mainly during WM retrieval. In an fMRI study
(Rypma et al., 2002), subjects performed a WM task that
required them to maintain from one to eight letters over
a brief delay. Neural activity was measured during en-
coding, Wartung, and retrieval task phases. With in-
creasing memory load, the researchers observed that RT
increased and accuracy (SCR) verringert. Zusätzlich, A
decline in ventrolateral PFC (VLPFC) activation was seen
1University of Utah, 2Hillside-Zucker Hospital, Glen Oaks, New York, 3Bereits-
tional Institute of Neurological Disorders and Stroke, Bethesda, MD
during encoding, whereas dorsolateral PFC (DLPFC) ac-
tivation increased during maintenance and retrieval. Der
authors concluded that VLPFC mediates WM storage and
that DLPFC mediates memory organization processes that
facilitate supra-capacity WM storage. Interessant, hoch-
performing subjects in toto showed less activation than
low-performing subjects, suggesting that high performers
utilized fewer neural resources in resolving task require-
gen. In comparison to low performers, such task perfor-
mance could plausibly be seen as an example of greater
neural efficiency (Ne). Auch so, higher-scoring performers
showed activation increases with increasing memory load
during maintenance and retrieval in lateral PFC. Gesamt,
low-performing subjects showed more activation than
high-performing subjects, but high performers showed
minimal activation increases in DLPFC with increasing
memory load. The authors suggested that their results in-
dicate that individual differences in Ne and cognitive strat-
egy mediate individual differences in WM performance
(Rypma et al., 2002). In a subsequent study comparing
WM performance in younger and older adults, Rypma,
Berger, Genova, Rebbechi, and DʼEsposito (2005) gebraucht
event-related fMRI to measure brain activity while younger
and older adults performed an item-recognition task with
set size varying between one and eight letters. Both groups
experienced a decline in Ne indexed as RT increases and
accuracy decreases with increasing memory set size. Der
authors also found related differences in neural activation
with increasing memory set size in PFC. Analysis of in-
dividual subjectsʼ performance relative to cortical activity
© 2009 Massachusetts Institute of Technology
Zeitschrift für kognitive Neurowissenschaften 22:3, S. 447–456
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
F
/
.
.
.
T
Ö
N
1
8
M
A
j
2
0
2
1
suggested that speed and accuracy accounted for con-
siderable variance in dorsal and ventral PFC activity during
encoding and retrieval. The authors concluded that their
results suggested support to a model of cognitive slowing
in which processing rate is related to Ne (Rypma et al.,
2005). In ähnlicher Weise, Altamura et al. (2007) used fMRI
to dissociate the effects of Sternberg WM demands on
DLPFC. The effect of varying loads of three, five, or eight
letters on WM maintenance was determined by using two
time delays (1 sec and 6 Sek) between the letter set and
the probe stimuli. The authors observed strongly activated
DLPFC with load manipulation. Vor allem, regions of right
DLPFC were exclusively affected by load. The authors con-
cluded that their results suggested the possibility that top–
down modulation of cognitive control during encoding or
response to probe stimulus could be mediated by these
Bereiche (Altamura et al., 2007).
Increasingly, transcranial magnetic stimulation (TMS) Ist
used as a research tool for mapping brain function and
modification of neural processes during WM tasks that en-
gage PFC. TMS has been shown to be a safe, noninvasive
means of stimulating the awake and alert human cere-
bral cortex in carefully screened subjects ( Wassermann,
1998). Zum Beispiel, Koch et al. (2005) used a repetitive
TMS (rTMS) approach to disambiguate the spatial distribu-
tion and reciprocal interactions of different regions of the
parieto-frontal network in healthy human participants per-
forming a spatial WM task. These researchers compared
the effect of 25 Hz rTMS on neural activity within PFC
and posterior parietal cortex (PPC) during the delay and
decision phases of the task. Trains of rTMS at 25 Hz were
applied to PPC, premotor cortex, and right DLPFC alterna-
tively during the two phases. They found that TMS dur-
ing the delay phase in both parietal cortex and DLPFC
interfered with performance. When 25-Hz rTMS trains
were applied during the decision phase, they observed in-
terference in DLPFC, but not in PPC or premotor cortex. A
local neural network subserving decisional processes and
a second neural population functionally interconnected
with PPC that was activated when spatial information was
maintained temporarily in WM (Koch et al., 2005). Brandt,
Ploner, Meyer, Leistner, and Villringer (1998) investigated
the role of DLPFC and PPC in a visuospatial delayed-
response task. Using high-frequency rTMS, the researchers
interfered temporarily with cortical activity in DLPFC and
PPC during the maintenance period in order to determine
whether rTMS to DLPFC or PPC during maintenance af-
fects accuracy of memory-guided saccades. Stimulation
over DLPFC significantly impaired accuracy of amplitude
and direction of saccades. They concluded that, innerhalb
this network, DLPFC mediates the mnemonic representa-
tion and PPC with the sensory representation of spatially
defined perceptual information (Brandt et al., 1998).
Studies of the chronometrics of WM phase evolution have
used TMS in conjunction with fMRI to induce a temporary
lesion that is intended to interfere with task performance,
somit, identifying a causal role for the targeted cortical
region in mediating WM task performance. Such studies
have shown that the effects of rTMS on cognitive perfor-
mance are dependent on domain, experimental design,
cortical target, stimulation frequency, zeitliche Koordinierung, and duration
of TMS application relative to the stage of task processing.
Folglich, most studies of WM and the role of PFC
have been aimed to identify whether and precisely when
functional disruption of PFC would degrade WM perfor-
Mance. Zum Beispiel, Mull and Seyal (2001) aimed to deter-
mine whether transient functional disruption of DLPFC
would impair performance in a WM task in which par-
ticipants were shown sequences of letters and asked to
decide if the letter just displayed was the same as the letter
presented three trials back. Single-pulse TMS was applied
over DLPFC between letter presentations. TMS to left
DLPFC caused increased errors relative to controls. Der
authors concluded that their results supported the idea
that DLPFC played a crucial role in this paradigm. Delete-
rious effects of TMS to LPFC on WM were also observed
by Osaka et al. (2007) in a study using paired-pulse TMS.
Albeit fewer in number, studies showing beneficial effects
of TMS on WM have also observed a critical role for stim-
ulation frequency and timing relative to the phase of
task evolution. Zum Beispiel, individual power and peak
frequency attributes of alpha band oscillations have been
shown to predict performance on a WM task (Luber et al.,
2007; Klimesch, Sauseng, & Gerloff, 2003). In a previous
Studie, the present authors designed a combined EEG–
TMS study of the interaction between WM Ne, alpha band
oscillations, Und 10 Hz rTMS applied to DLPFC prior to
the onset of a Sternberg task (Anderson, Preston, & Silva,
2007). We predicted first that compared with sham active
10 Hz rTMS would significantly reduce psychomotor pro-
cessing speed indexed as RT. Zweite, we predicted that
improvement in RT would be associated with a 10-Hz
rTMS induced increase in pretask alpha power and pretask
alpha band phase synchrony. We identified a robust as-
sociation between 10 Hz rTMS perturbed individual mean
peak alpha frequency and alpha spectral phase synchrony
on task performance.
Frequency and target-specific effects have also been
observed in studies of cognitive domains other than WM:
Zum Beispiel, enhancement of picture naming and posterior
superior temporal cortex (Mottaghy, Sparing, & Topper,
2006); enhancement of episodic memory and left inferior
PFC (Kohler, Paus, Buckner, & Milner, 2004); analogic rea-
soning and left DLPFC (Boroojerdi et al., 2001); mental
rotation of 3-D objects and mesial frontal and right parietal
Kortex (Klimesch et al., 2003). Endlich, Luber et al. (2007)
showed that TMS could enhance WM. They predicted
that healthy subjects’ performance on a delayed match-to-
sample task would be enhanced when TMS of 1, 5, oder 20 Hz
was applied to either left dorsolateral prefrontal or midline
parietal cortex during the delay phase of the task. Sie
found 5 Hz, but not 1 Hz or 20 Hz, stimulation to the
midline parietal site during delay resulted in a significant de-
crease in RT without a corresponding decrease in accuracy
448
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 3
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
T
.
.
.
F
/
Ö
N
1
8
M
A
j
2
0
2
1
(SCR). In a second experiment, 5 Hz rTMS was applied to
the parietal site during the delay or during presentation of
the probe. RT enhancement occurred only with stimulation
during the delay phase. The authors concluded TMS could
enhance WM performance under specific conditions of
Zeit, target, and stimulation frequency.
The present study was designed to determine the effects
von 10 Hz rTMS directed to right or left DLPFC on psycho-
motor processing speed (RT), accuracy (SCR), and indices
of Ne, on a WM task (Sternberg, 1966). In view of the
numerous studies that showed a critical role for DLPFC
in processing WM tasks, yet a minority of studies that show
performance enhancement when rTMS was directed to
DLPFC during evolutionary stages of a WM task, we enter-
tained the notion that subject distraction might play a role
in such an apparent discrepancy. Relevant in this regard
is the study of Abler et al. (2005). In order to determine
the relation between TMS-induced distraction and perfor-
Mance, Abler et al. asked healthy subjects to evaluate the
discomfort caused by TMS during a verbal WM task. Sub-
jects were first studied using fMRI to identify specific cor-
tical regions engaged in task processing. Subsequently,
left DLPFC, inferior frontal, parietal, and temporo-parietal
cortices were identified and stimulated during the delay
phase of a delayed match-to-sample task. Task perfor-
mance and subjective disturbance due to side effects were
monitored. Subjectsʼ level of discomfort was correlated
with their error rates: The higher the discomfort, the more
errors they made. Abler et al. concluded that TMS sensory–
motor side effects induced during task evolution may
bias task performance and lead to misinterpretation of
the results. Our predictions were also influenced by earlier
work that showed DLPFC plays an important role in WM.
Zum Beispiel, Hamidi, Tononi, and Postle (2009) used high-
frequency rTMS to evaluate the role of DLPFC in memory-
guided response to two different types of spatial WM tasks:
one requiring a recognition decision about a probe stimu-
lus using a YES–NO button press, another requiring recall
of the stimulus location. In half the trials, randomly dis-
tributed rTMS was applied to DLPFC; in a separate session,
the same stimulation was directed to the superior parietal
lobule (SPL), a brain area implicated in spatial WM stor-
Alter. In each condition, a 3-sec train of 10 Hz at 110% von
motor threshold was delivered at the onset of the response
Zeitraum. DLPFC, but not SPL, rTMS affected performance;
rTMS to right DLPFC decreased SCR of delayed-recall trials;
rTMS to left DLPFC enhanced SCR on delayed-recognition
Versuche. The authors concluded that their results suggested
DLPFC plays an important role in memory-guided re-
sponse and that the nature of such a role varies with the
processes required for making a response (Hamidi et al.,
2009).
The concept of Ne in human studies is largely derived
from fMRI studies of the relationship between cognitive
task performance and neural activation indexed by the
signal magnitude of blood oxygen level-dependent BOLD
utilization. An array of similar investigations lends sup-
port to the finding that as item-load increases, BOLD
signal amplitude increases. Such increases have been re-
peatedly shown to be inversely associated with task per-
Form. Zum Beispiel, Gould, Braun, Owen, ffytche, Und
Howard (2003) used event-related fMRI to measure PFC
activity while younger and older adults performed an
item-recognition task in which the memory set size varied
between one and eight letters. RT increased and accuracy
decreased with increasing memory set size. Variance in RT
and BOLD signal in dorsal and ventral PFC accounted
for most of the variance. The authors concluded that their
results support a model of cognitive slowing with age in
which a decline in processing speed is related to a decline
in Ne (Gould et al., 2003).
In the present rTMS study, we predicted first that com-
pared with sham active 10 Hz rTMS applied to either right
or left DLPFC 10 sec prior to task onset would enhance the
neural efficiency (Ne) of WM operationally indexed by the
relationship between RT and SCR, whereby Ne = work/
Zeit; somit, Ne = accuracy/RT. Guided, in part, by earlier
work that suggested the auditory and sensory stimulation
of rTMS could degrade cognitive performance in partici-
pants who are distracted by such extraneous stimuli (Abler
et al., 2005), we directed 10 Hz rTMS to either right or left
DLPFC serially 10 sec prior to task onset with the intention
of allowing time for resolution of the putative disruptive
effects of stimulation. Zweite, we posited that if sensori-
motor side effects of rTMS biased our results, we would
see equal or better performance in the sham-stimulated
group compared with the actively stimulated group.
METHODEN
Experimental Design
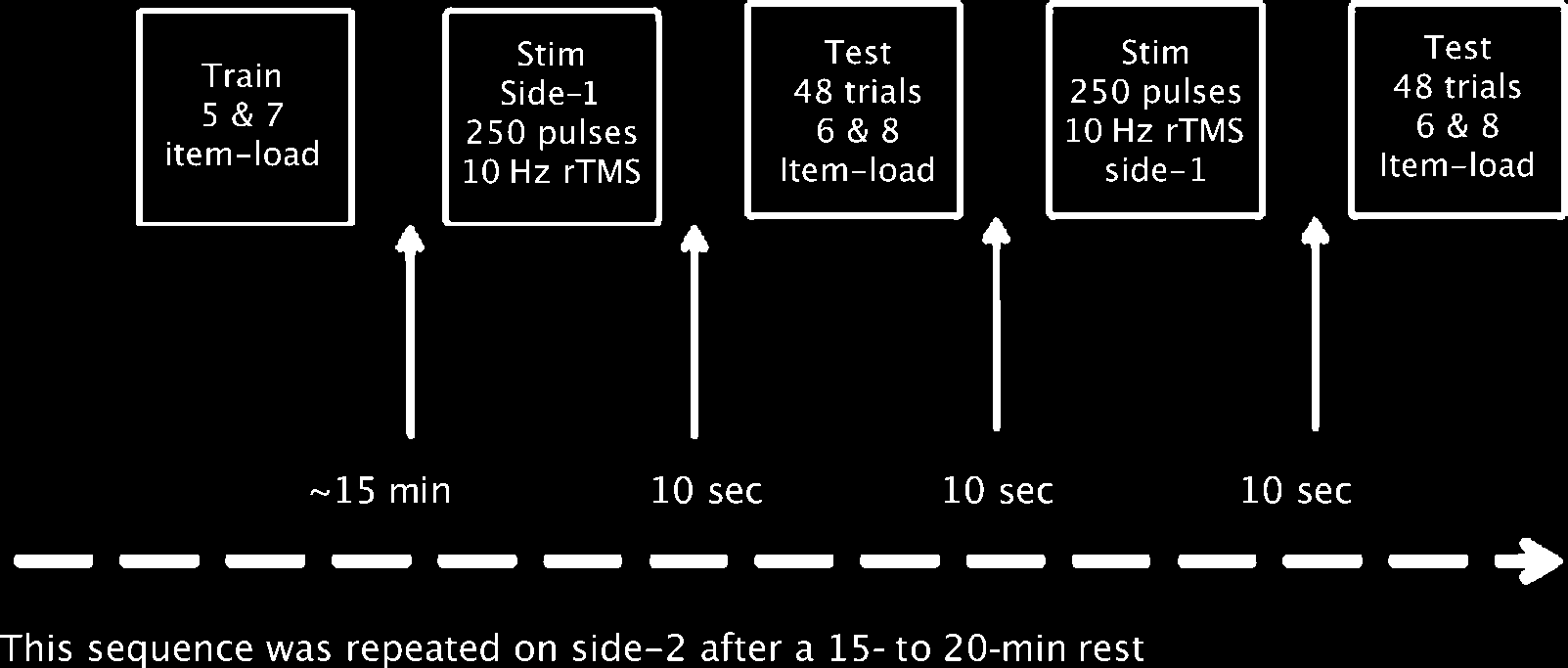
This was a single-blind, sham-controlled study (Figur 1).
In 32 healthy participants, active or sham 10 Hz rTMS
was directed to either right or left DLPFC 10 sec prior
to the onset of a Sternberg short-term verbal recognition
memory paradigm. Five 5-sec trains of 10 Hz rTMS (for a
total of 250 pulses) were applied to DLPFC ending 10 Sek
prior to task onset. Stimulation was followed by 48 single
trials of 6- and 8-item strings of capital letters that were
randomized and counterbalanced. Nächste, 250 zusätzlich
pulses of 10 Hz rTMS were delivered to DLPFC of the same
Seite, followed by a second set of 48 single trials of 6- Und
8-item strings of capital letters. After a 15- to 20-min rest,
the same sequence of events was repeated on side-2.
Outcome measures were pre- and poststimulation accu-
racy (SCR), RT, and Ne by stimulation group (STM).
Teilnehmer
The Investigational Review Boards for the University of
Utah and Veterans Health Affairs of the Salt Lake City
Health Care System approved this study. Thirty-four medi-
cally healthy, right-handed participants, ages 18–55 years,
Preston et al.
449
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
/
.
T
F
.
.
Ö
N
1
8
M
A
j
2
0
2
1
from the University of Utah and the Salt Lake City commu-
nity were admitted to the study free of exclusionary condi-
tions for TMS (Wassermann, 1998) and without a past or
present psychiatric disorder. All participants had a negative
screen for drugs of abuse on the morning of the study.
Participants were instructed not to drink caffeinated bev-
erages for 12 hours prior to testing. Participants were com-
pensated for their time. Group assignment to stimulation
(active or sham) and side first stimulated (S-1) were pseudo-
randomized and counterbalanced.
Paradigm Training
Participants were trained and tested between 11:00 Bin.
Und 1:00 Uhr. They were familiarized with the task and
response procedures by practicing to use their right first
finger to press one of two buttons (YES or NO) on a hand-
held keypad as comfortably quick as possible while attend-
ing to a fixation cue at the center of a 19-inch computer
monitor. Baseline values of SCR and RT were derived from
20 randomized and counterbalanced 5- and 7-item single
trials of the Sternberg task.
The Sternberg Paradigm
Transcranial Magnetic Stimulation
Task stimuli and computation of RTs and accuracy (SCR)
were derived using Presentation 9.90 Software (Neuro-
behavioral Systems, San Francisco, CA). A single trial of
this task was composed of four phases: stimulus (A 5-
or 7-item string of uppercase consonants and vowels
was presented at the center of a 19-inch video monitor
für 1.5 Sek), delay (participants focused on a fixation marker
appearing at the center of the computer monitor while
maintaining the previous string in memory for 1.5 Sek),
probe (a single uppercase letter appeared at center screen
für 3 Sek), und Reaktion (participants were instructed to
decide as quickly and accurately as possible whether the
probe was present in the previous string, and to respond
by pressing one of two keypad buttons; if there was a re-
sponse within 3 Sek, it was recorded, and the next trial
began. If there was no response, the trial was marked as
an error, and the next trial began). A minimum baseline
SCR of 80% on the 5-item, Und 70% on the more difficult
7-item task was required to continue. If necessary, partici-
pants were allowed to repeat each of the training para-
digms once and were disqualified if they failed to make
the minimum SCR. Training and testing phase item-strings
had equal numbers of randomized and counterbalanced
true positive and true negative probes.
During the experiment, participants were comfortably
seated in an individually adjustable chair. Participantsʼ
single-pulse TMS resting motor threshold (RMT) War
determined by identification of the lowest level of TMS
machine output that would induce a visible contraction
of the first dorsal interosseous muscle of the dominant
hand on 5 von 10 pulses delivered with a 10-sec interpulse
interval. A MagStimTM Super Rapid stimulator with physi-
cally identical sham and active air-cooled figure-of-eight
coils (Magstim, Whitland, Dyfed UK) was used to deliver
TMS pulses directed to either left or right DLPFC. Der
sound generated by capacitor discharge was identical in
the two coils; the sham coil did not generate a sensori-
motor stimulus. A custom-designed mechanical coil holder
was used to maintain contact between the midpoint of
the 70-mm figure-eight coil and the location of the fron-
tal scalp overlying right or left DLPFC. To determine the
scalp correlates for targeting of DLPFC, we used the mea-
sures 10–20 EEG system. Positioning of the coil on the
scalp was determined by the measured position of EEG
sensors F3 (links) and F4 (Rechts). Participants used dis-
posable in-ear sound protection during the experiment.
Side stimulated first (S-1) was randomly assigned and
counterbalanced.
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
F
.
.
.
/
T
Ö
N
1
8
M
A
j
2
0
2
1
Figur 1. Experimentelles Design.
450
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 3
Subject Stimulation and Testing
A brief period of acclimation to the laboratory environment
was followed by the onset of five 5-sec trains of 10 Hz rTMS
with an intertrain interval of 10 sec at 100% of RMT for
insgesamt 250 pulses of 10 Hz rTMS. Stimulation ended
10 sec prior to the onset of the first 48 single trials of the
Aufgabe. In order to minimize the effects of practice on per-
Form, single trials in the testing phase were composed
of more difficult 6- or 8-item uppercase letters pseudo-
randomly presented and counterbalanced for true positive
and true negative probes.
This sequence was repeated once more on S-1. Following
A 15- to 20-min rest period, the entire sequence was re-
peated on S-2; somit, the study totaled 192 single trials
Und 1000 pulses of 10 Hz rTMS.
Planned Analyses
Skewness and kurtosis in the distribution of dependent
variables were identified and normalized with square
root transformation. Repeated measures ANOVA with
Bonferroni correction for multiple comparisons was used
to identify the main effects of STM, S-1, and interactions
between STM and S-1. Dependent variables were covaried
with RMT and age. Pairwise comparisons were used to de-
termine the within-subject effects of active and sham 10 Hz
rTMS on SCR and RT by task load.
ERGEBNISSE
Thirty-four right-handed men and 12 women recruited from
the Salt Lake City, Utah, community participated in this study.
Two subjects in the actively stimulated group were unable to
complete the study due to the development of moderate
muscle contraction headaches. In both cases, the headaches
spontaneously resolved. There were no adverse events in the
sham group. All participants achieved the minimum 5- Und
7-item training SCR on their first attempt.
Demographics
There was no difference in mean age between stimulation
groups or by sex. Mean age by Stimulation group: active =
28.6 (SD = 5.68); sham = 30.88 (SD = 11.2); independent-
samples two-tailed t test for equality of means; equal
variance not assumed: T(31) = −0.707, p = .478. Mean
age by Sex: male = 28.75 Jahre; female = 29.50 Jahre;
zweiseitiger t-Test: T(30) = −0.25, p = .80. There was equal
distribution of the sexes between groups [Pearson un-
corrected chi-square test: χ2(1, n = 32) = 0.139, p = .710].
Resting Motor Threshold
The groups were evenly matched for the intensity of stim-
ulation indexed as RMT and expressed as a percent of max-
imum machine output [active RMT = 67%, sham RMT =
66%; two-tailed t(34) = 0.252, p = .4484].
Results of the Repeated Measures ANOVA
There was no main effect of S-1, and no interaction be-
tween S-1 and STM. Measures of dependent variables did
not covary with RMT, Sex, or age. In the absence of a main
effect of S-1, prestimulation and poststimulation values of
dependent variables were collapsed across side for subse-
quent analysis. There were no prestimulation differences
between groups regarding SCR, RT, or Ne. There was, Wie-
immer, a robust main effect of STM on poststimulation RT
and Ne, but not SCR, as presented in Table 1.
In der vorliegenden Studie, the authors directed 10 Hz rTMS
to right or left DLPFC 10 sec prior to task onset, predicting
there would be a main effect of STM on SCR, RT, and Ne.
We observed a robust main effect of STM in the compari-
son of poststimulation RT and Ne, but not SCR. The mean
poststimulation RT in the active group was 219 ms
(±0.16) faster than the prestimulation baseline, wohingegen
the mean poststimulation RT in the sham group was
30 ms (±0.16) slower than the mean baseline RT. Pair-
wise comparisons of pre- and poststimulation RT, SCR, Und
Ne were computed by comparing the relatively easier 5-
and 7-item (prestimulation) measures with the 6- Und
8-item (poststimulation) messen. Pairwise comparisons
of the 5- and 7-item training RT data (Tisch 2) show a small
but nonsignificant within-subject difference in both groups
for this comparison. Speziell, pairwise comparisons in
the actively treated group showed that compared with the
larger 6-item and 8-item poststimulation loads, the 5-item
prestimulation RT was slower when the converse would
be expected. Similar results were seen when the compari-
son of the prestimulation 7-item RT was compared with
the poststimulation 8-item RT. In the sham group, the same
comparisons failed to show a difference for pre- or post-
measures of RT, SCR, or Ne (Figuren 2 Und 3).
DISKUSSION
The present study compared the relative effects of active
and sham 10 Hz rTMS directed to the right and left DLPFC
on psychomotor processing speed and accuracy, indices
of Ne, in healthy participants performing the Sternberg
Aufgabe, a short-term verbal recognition memory paradigm
that has been shown to model the association between
WM load and task behavior (Sternberg, 1966). Previous
studies of the role of DLPFC in WM have been designed
to identify the temporal and spatial dynamics of the com-
putational role of DLPFC in a WM network. Zum Beispiel,
Osaka et al. (2007) investigated the role of executive func-
tion in human left DLPFC in eight normal participants
using low-frequency TMS after fMRI activation confirmed
a role for left DLPFC. TMS was applied to left DLPFC im-
mediately after subjects finished reading sentences from
the reading span test, a measure of verbal WM. They found
Preston et al.
451
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
T
.
/
F
.
.
Ö
N
1
8
M
A
j
2
0
2
1
Tisch 1. Effects of 10 Hz rTMS on the Neural Efficiency of
Working Memory
Descriptive Statistics
Active
Sham
Mean
SD
Mean
SD
Pre-10 Hz rTMS SCRa
Post-10 Hz rTMS SCRa
Pre-10 Hz rTMS RTb
Post-10 Hz rTMS RTb
Pre-10 Hz rTMS NEc
Post-10 Hz rTMS NEc
Mean % Change NEc
0.920
0.870
1.117
0.874
0.832
0.995
1.200
0.07
0.970
0.03
0.06
0.860
0.05
0.08
1.010
0.10
0.05
1.014
0.11
0.17
0.969
0.21
0.13
0.866
0.20
0.30
0.900
0.10
Repeated Measures ANOVA
F (df = 1, 30)
Sig.
Pre-10 Hz rTMS SCRa
Post-10 Hz rTMS SCRa
Pre-10 Hz rTMS RTb
Post-10 Hz rTMS RTb
Pre-10 Hz rTMS NEc
Post-10 Hz rTMS NEc
0.019
0.012
3.170
7.380
1.628
5.344
Mean % Change NEc
14.253
.892
.915
.086
.011
.212
.028
.001
aAccuracy.
bReaction time.
cNeural efficiency.
a significant deterioration of performance in this WM task,
somit, supporting a crucial role for DLPFC in WM. Koch
et al. (2005) used an rTMS approach to investigate the
temporal and spatial dynamics of the parieto-frontal net-
work in normal subjects performing a spatial WM task.
They aimed to compare neural activity in the relevant
areas during the delay and decision phases of the task.
Trains of rTMS at 25 Hz were directed to PPC and right
DLPFC during the two phases. Premotor cortex was used
as a spatial control. They observed that TMS interfered
with performance during the delay phase in the posterior
parietal and DLPFC sites. There was no effect for the con-
trol site. When rTMS trains were applied during the re-
trieval phase, interference was observed only in DLPFC.
The authors concluded that their study identified an exam-
ple of parallel processing in the parieto-frontal network of
spatial WM during the delay phase. The results of Koch
et al. represent additional evidence of the critical role of
DLPFC during both the delay and the retrieval phases of
a WM task. Mottaghy, Gangitano, Krause, and Pascual-
Leone (2003) observed significant interference with WM
RT after 180 msec of left PFC stimulation. Results in a simi-
lar vein were observed by other researchers (Postle et al.,
2006; Mull & Seyal, 2001; Mottaghy et al., 2000).
The present study was designed, in part, to identify a
putative role for DLPFC in the Ne of a WM task. Accuracy
(SCR) declined to a similar extent in both groups, likely
due to the effects of memory load, hence supporting the
conclusion of previous studies that also found no effect
of TMS on WM task accuracy (Luber et al., 2007, 2008;
Desmond, Chen, & Shieh, 2005). Im Gegenzug, such findings
suggest that the effect of 10 Hz rTMS on RT when di-
rected to DLPFC 10 sec prior to task onset mediates the
enhancement of Ne. Zusätzlich, these data suggest that
the retrieval phase of this WM task may be the temporal
boundary of the observed effects.
Results of the present study are in line with Rypma
et al. (2006), who set out to identify the neural bases of
interindividual differences in cognitive performance. Sie
had participants perform a simple speed of processing task
during fMRI scanning. In certain prefrontal cortical regions
(PFC), faster performers used fewer neural resources than
slower performers, whereas in other PFC and parietal
Regionen, they used more. These results were interpreted
to suggest that a critical determinant of interindividual
differences in cognitive performance was the efficiency of
interactions between brain regions. In order to perform
well, slower individuals may utilize more prefrontal re-
sources than faster individuals (Rypma et al., 2006).
Taking into account the lack of difference between
groups in baseline RT (Tisch 1), together with a robust
RT enhancement with active stimulation, and consider-
ing that participants in both groups had equal practice ex-
perience over the course of experiment, one might
plausibly argue that our data support the conclusion that
the poststimulation difference in task performance be-
tween groups is best explained as a difference between ac-
tive and sham stimulation (Tisch 3). Auch so, we recognize
valid challenges to these conclusions. Our results were
Tisch 2. Pairwise Comparisons: Effects of 10 Hz rTMS Pre- Und
Post-10 Hz rTMS
T
P ( Two-Tailed)
Active
Pre vs. Post SCR
Pre vs. Post RT
Pre vs. Post Efficiency
Sham
Pre vs. Post SCR
Pre vs. Post RT
Pre vs. Post Efficiency
2.362
4.417
−4.993
3.113
−1.264
0.986
.0340
.0010
.0002
.008
.228
.342
452
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 3
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
.
/
T
.
.
F
Ö
N
1
8
M
A
j
2
0
2
1
with Klimesch et al. (2003), who showed that mean peak
alpha spectral frequency (IAF) is related to memory per-
Form. Klimesch et al. showed that compared with at-
tentional demands, memory performance exerted the
strongest effect on IAF. The difference in IAF between
good and bad memory performers peaked when subjects
retrieved information from memory. During retrieval, Die
IAF of good performers was 1.25 Hz higher than that of
bad performers.
Brignani, Manganotti, Rossini, and Miniussi (2008) found
that low-frequency TMS over primary motor cortex in-
duced a synchronization of the background oscillatory
activity in the stimulated region, hence suggesting that
TMS effects may involve short-term modification of the
neural circuitry sustaining motor behavior (Brignani et al.,
2008; Leiberg, Lutzenberger, & Kaiser, 2006; Klimesch,
1997). Emergent evidence suggests that a diminished
capacity to synchronize distributed neural assemblies me-
diating WM may be a critical and enduring underlying
mechanism of impaired WM in a broad array of seemingly
disparate neuropsychiatric disorders such as schizophre-
nia (Ford, Krystal, & Mathalon, 2007; Andreasen et al.,
1999; Friston, 1999), depression (Linkenkaer-Hansen
et al., 2005), Alzheimerʼs disease (Yener, Güntekin, Oniz,
& Basar, 2007; Osipova, Ahveninen, Jensen, Ylikoski, &
Pekkonen, 2005), and autism (Uhlhaas & Singer, 2007). Von-
spite such recognition, there is no treatment for impaired
memory in any such disorder. TMS has been shown to
mediate events at the neural synapse (Fuggetta, Pavone,
Fiaschi, & Manganotti, 2008). In light of previous work, Das
suggests that such effects may be related to 10 Hz rTMS in-
duced phase-state perturbation (Anderson et al., 2007); Wenn
Also, it may also suggest a role for TMS as a clinical tool for
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
.
F
.
/
.
T
Ö
N
1
8
M
A
j
2
0
2
1
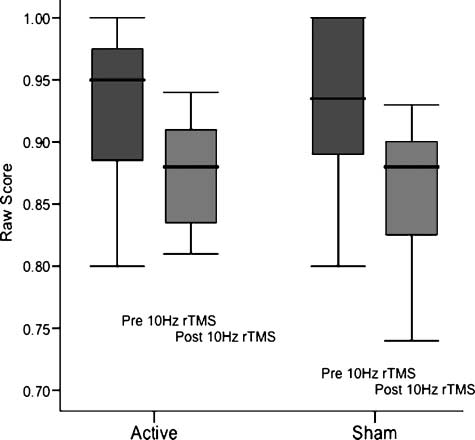
Figur 3. Lack of effect of 10 Hz rTMS on accuracy within or between
Fächer. The difference in accuracy within and between groups pre-
and poststimulation was not significant.
Preston et al.
453
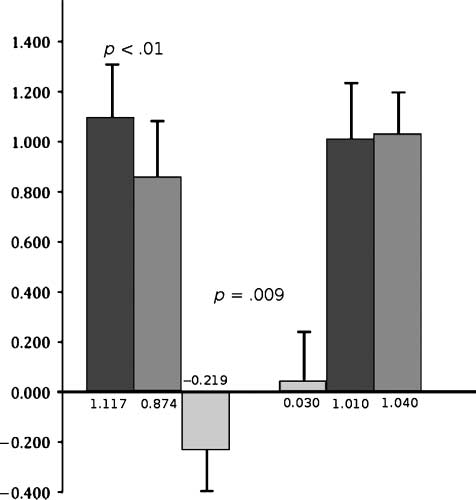
Figur 2. Comparative effects of active and sham 10 Hz rTMS on
working memory reaction time. Left-side sequence: Pre-10 Hz rTMS
RT*; Post-10 Hz rTMS RT; RT difference. Right-side sequence: RT
difference; Pre-10 Hz rTMS RT*; Post-10 Hz rTMS RT. Compared
with sham, the group treated with active 10 Hz rTMS applied to DLPFC
10 sec before onset experienced a 219-msec enhancement of RT.
specific to SCR, RT, and Ne; even so it is possible that the
effects were due to a priming effect of rTMS on psychomo-
tor functions that are also mediated through prefrontal net-
funktioniert (Rounis, Yarrow, & Rothwell, 2007; Vogt et al., 2007).
Although we would suggest that it is unlikely the observed
effects are strictly due to learning, inasmuch as both the
sham and active rTMS groups had equal opportunity to
üben, we recognize there may have been an interaction
zwischen 10 Hz rTMS and the learning of a response strat-
egy that results in faster but not more accurate responses.
An additional challenge to our conclusions is related to the
capacity limits of short-term memory (Marois & Ivanoff,
2005; Cowan, 2001). To overcome such limitations, Studie
subjects may devise strategies, such as chunking to suc-
cessfully perform at supra-capacity limits, which can affect
neural activity in a complex way, leading to potential com-
plications in the interpretation of results. In view of the
high-capacity nature of the task we used in this study, es ist
possible that chunking or other mnemonic strategies may
have been used to manage supra-capacity aspects of task
Leistung.
The underlying neurobiological mechanisms subserving
the effects of TMS on WM Ne are unclear. In a previous ex-
ploratory study, the present authors found that 10 Hz rTMS
applied to DLPFC in healthy humans resulted in a signifi-
cantly enhanced measure of individual mean peak alpha
frequency, and enhanced within band 10–12 Hz phase syn-
chrony (Anderson et al., 2007). These results are in line
Tisch 3. Pairwise Comparisons of Reaction Time × Memory
Load for Each Group
Group
RT (ICH)
RT (J)
Difference (I−J)
Mean
Active
5-item 6-item
8-item
7-item 6-item
8-item
Sham
5-item 6-item
8-item
7-item 6-item
8-item
0.278
0.252
0.167
0.141
−0.046
−0.101
0.098
0.052
SEM
0.07
0.07
0.03
0.03
Sig.
.0060
.0170
.0001
.0005
0.03
1.0000
0.04
0.04
.1930
.1450
0.05
1.0000
In the actively stimulated group, the 5-item prestimulation RT is slower
than the 6- and 8-item RT. The same relationship is true for comparison
of the 7- with the 8-item RT; in the sham group, Die 6- and 8-item RTs
are slower than the 5-item RT, as would be expected.
neural circuit repair or enhancement (Plewnia et al., 2008;
Sauseng & Klimesch, 2008). In a study with 24 Fächer,
Hamidi et al. (2009) used high-frequency rTMS to evaluate
the role of DLPFC in memory-guided response to two dif-
ferent types of spatial WM tasks: one requiring a recognition
decision about a probe stimulus (operationalized with a
yes/no button press), and another requiring direct recall
of the memory stimulus by moving a cursor to the remem-
bered location. In half the trials, randomly distributed, rTMS
was applied to DLPFC, and in a separate session, to the SPL,
a brain area implicated in spatial WM storage. A 10-Hz (3 Sek,
110% of motor threshold) train of rTMS was delivered at the
onset of the response period. They found that only rTMS
applied to DLPFC significantly affected performance. Wann
10 Hz rTMS was directed to right DLPFC, accuracy declined
on delayed-recall trials. Wann 10 Hz rTMS was directed to
left DLPFC, accuracy on the delayed-recognition trials was en-
hanced. These findings support DLPFCʼs role in memory-
guided response and suggest that the nature of this role
varies depending on the processes required for making a
response. Koch et al. (2005) used an rTMS approach to in-
vestigate the temporal dynamics and reciprocal interactions
of different regions of the parieto-frontal network in normal
subjects performing a spatial WM task. They aimed to com-
pare neural activity in these regions in the delay and deci-
sion phases of the task. Trains of rTMS at 25 Hz were
delivered over PPC, premotor cortex (SFG), and DLPFC
of the right hemisphere alternatively during the two phases.
They observed TMS interference during the delay phase for
PPC and DLPFC sites of stimulation, with no effect on the
control site. When rTMS trains were applied during the re-
trieval phase, task interference was limited to DLPFC. Der
authors concluded their study supported the existence of
a parallel processing network that was active during the
delay phase of the task. They concluded that in DLPFC,
two task-specific networks coexisted: a local neural network
subserving the decisional processes, and a second neural
population functionally connected to PPC that was activated
when spatial-specific information maintained in memory
was made available for use.
Luber et al. (2007) designed a delayed match-to-sample
task in which rTMS at 1, 5, oder 20 Hz was applied to either
left DLPFC or midline parietal cortex during the delay
phase of the task. Nur 5 Hz stimulation to the parietal site
resulted in a significant decrease in RT with no effect on
SCR. In a second experiment, 5 Hz rTMS was directed to
the parietal site during either the maintenance or retrieval
phase of the recognition probe. RT enhancement occurred
in the retention phase but not the probe phase. They con-
cluded their results suggest that 5 Hz rTMS may improve
WM performance when stimulation of a specific frequency
was timed to a specific phase of task performance.
Earlier work to show that TMS may enhance prefrontal
oscillatory synchrony in spectral bands subserving WM in-
formation processing (Anderson et al., 2007), as noted
über, lends support to the idea of developing a neural
circuit-based model for the treatment of memory impair-
ment using noninvasive brain stimulation (NIBS) Methoden
such as TMS. Presently, evidence to support an NIBS-based
approach is supported by work to show improved clinical
status in depression (OʼReardon et al., 2007), hallucina-
tions in schizophrenia (Aleman, Sommer, & Kahn, 2007;
Hoffman et al., 2005), and posttraumatic stress disorder
(Osuch et al., 2009; Grisaru, Amir, Cohen, & Kaplan, 1998).
Danksagungen
This study was supported by the NARSAD Young Investigator
Award (to G. P.), the L. C. Tanner Family Foundation (to G. P.),
and Intramural NINDS, DARPA (to E. W.). National Science Foun-
dation (IIS-0513692 to C. S., CCF-0401498 to C. S., EIA-0323604 Zu
C. S., CNS-0514485 to C. S., IIS-0534628 to C. S., CNS-0528201 Zu
C. S., OISE-0405402 to C. S.); Department of Energy (to C. S.);
IBM Faculty Award (to C. S.).
Reprint requests should be sent to Dr. Gilbert Preston, Depart-
ment of Psychiatry, University of Utah School of Medicine, 30
Norden 1900 Ost, Salt Lake City, Utah 84132-2502, oder per E-Mail:
gilbert.preston@hsc.utah.edu or gilbert.preston@med.va.gov.
VERWEISE
Abler, B., Walter, H., Wunderlich, A., Grothe, J., Schonfeldt-
Lecuona, C., Spitzer, M., et al. (2005). Side effects of
transcranial magnetic stimulation biased task performance
in a cognitive neuroscience study. Brain Topography, 17,
193–196.
Aleman, A., Sommer, ICH. E., & Kahn, R. S. (2007). Efficacy of
slow repetitive transcranial magnetic stimulation in the
treatment of resistant auditory hallucinations in
schizophrenia: A meta-analysis. Journal of Clinical
Psychiatrie, 68, 416–421.
Altamura, M., Elvevag, B., Blasi, G., Bertolino, A., Callicott, J. H.,
Weinberger, D. R., et al. (2007). Dissociating the effects of
Sternberg working memory demands in prefrontal cortex.
Psychiatry Research, 154, 103–114.
454
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 3
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
F
T
/
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
Anderson, E. W., Preston, G. A., & Silva, C. T. (2007).
Towards development of a circuit based treatment for
impaired memory: A multidisciplinary approach. Verfahren
of the 3rd International IEEE EMBS Conference on
Neural Engineering (S. 302–305).
Andreasen, N. C., Nopoulos, P., OʼLeary, D. S., Müller, D. D.,
Wassink, T., & Flaum, M. (1999). Defining the phenotype
of schizophrenia: Cognitive dysmetria and its neural
mechanisms. Biologische Psychiatrie, 46, 908–920.
Boroojerdi, B., Phipps, M., Kopylev, L., Wharton, C. M., Cohen,
L. G., & Grafman, J. (2001). Enhancing analogic reasoning
with rTMS over the left prefrontal cortex. Neurologie, 56,
526–528.
Brandt, S. A., Ploner, C. J., Meyer, B. U., Leistner, S., &
Villringer, A. (1998). Effects of repetitive transcranial
magnetic stimulation over dorsolateral prefrontal and
posterior parietal cortex on memory-guided saccades.
Experimentelle Hirnforschung, 118, 197–204.
Brignani, D., Manganotti, P., Rossini, P. M., & Miniussi, C.
(2008). Modulation of cortical oscillatory activity during
transcranial magnetic stimulation. Kartierung des menschlichen Gehirns,
29, 603–612.
Cowan, N. (2001). The magical number 4 in short-term
Erinnerung: A reconsideration of mental storage capacity.
Behavioural Brain Science, 24, 87–114; Diskussion
114–185.
Desmond, J. E., Chen, S. H., & Shieh, P. B. (2005). Cerebellar
transcranial magnetic stimulation impairs verbal working
Erinnerung. Annals of Neurology, 58, 553–560.
DʼEsposito, M., Postle, B. R., & Rypma, B. (2000). Prefrontal
cortical contributions to working memory: Beweis
from event-related fMRI studies. Experimental Brain
Forschung, 133, 3–11.
Duncan, J., & Owen, A. M. (2000). Common regions of
the human frontal lobe recruited by diverse cognitive
demands. Trends in den Neurowissenschaften, 23, 475–483.
Ford, J. M., Krystal, J. H., & Mathalon, D. H. (2007). Neuronal
synchrony in schizophrenia: From networks to new
treatments. Schizophrenia Bulletin, 33, 848–852.
Friston, K. J. (1999). Schizophrenia and the disconnection
Hypothese. Acta Psychiatrica Scandinavica Supplementum,
395, 68–79.
Fuggetta, G., Pavone, E. F., Fiaschi, A., & Manganotti, P. (2008).
Acute modulation of cortical oscillatory activities during
short trains of high-frequency repetitive transcranial
magnetic stimulation of the human motor cortex: A
combined EEG and TMS study. Kartierung des menschlichen Gehirns,
29, 1–13.
Goel, V., & Grafman, J. (1995). Are the frontal lobes implicated
in “planning” functions? Interpreting data from the Tower
of Hanoi. Neuropsychologie, 33, 623–642.
Gould, R. L., Braun, R. G., Owen, A. M., ffytche, D. H., &
Howard, R. J. (2003). fMRI BOLD response to increasing
task difficulty during successful paired associates learning.
Neurobild, 20, 1006–1019.
Grisaru, N., Amir, M., Cohen, H., & Kaplan, Z. (1998). Effect
of transcranial magnetic stimulation in posttraumatic stress
disorder: A preliminary study. Biologische Psychiatrie,
44, 52–55.
Hamidi, M., Tononi, G., & Postle, B. R. (2009). Evaluating the
role of prefrontal and parietal cortices in memory-guided
response with repetitive transcranial magnetic stimulation.
Neuropsychologie, 47, 295–302.
Hoffman, R. E., Gueorguieva, R., Hawkins, K. A., Varanko, M.,
Boutros, N. N., Wu, Y. T., et al. (2005). Temporoparietal
transcranial magnetic stimulation for auditory hallucinations:
Safety, efficacy and moderators in a fifty patient sample.
Biologische Psychiatrie, 58, 97–104.
Klimesch, W. (1997). EEG-alpha rhythms and memory processes.
International Journal of Psychophysiology, 26, 319–340.
Klimesch, W., Sauseng, P., & Gerloff, C. (2003). Enhancing
cognitive performance with repetitive transcranial magnetic
stimulation at human individual alpha frequency.
Europäisches Journal für Neurowissenschaften, 17, 1129–1133.
Koch, G., Oliveri, M., Torriero, S., Carlesimo, G. A., Turriziani,
P., & Caltagirone, C. (2005). rTMS evidence of different
delay and decision processes in a fronto-parietal neuronal
network activated during spatial working memory.
Neurobild, 24, 34–39.
Kohler, S., Paus, T., Buckner, R. L., & Milner, B. (2004). Effects
of left inferior prefrontal stimulation on episodic memory
Formation: A two-stage fMRI–rTMS study. Zeitschrift für
Cognitive Neuroscience, 16, 178–188.
Leiberg, S., Lutzenberger, W., & Kaiser, J. (2006). Effects of
memory load on cortical oscillatory activity during auditory
pattern working memory. Gehirnforschung, 1120, 131–140.
Linkenkaer-Hansen, K., Monto, S., Rytsälä, H., Suominen, K.,
Isometsä, E., & Kähkönen, S. (2005). Breakdown of
long-range temporal correlations in theta oscillations in
patients with major depressive disorder. Zeitschrift für
Neurowissenschaften, 25, 10131–10137.
Luber, B., Kinnunen, L. H., Rakitin, B. C., Ellsasser, R., Stern, Y.,
& Lisanby, S. H. (2007). Facilitation of performance in
a working memory task with rTMS stimulation of the
precuneus: Frequency- and time-dependent effects.
Gehirnforschung, 1128, 120–129.
Luber, B., Stanford, A. D., Bulow, P., Nguyen, T., Rakitin, B. C.,
Habeck, C., et al. (2008). Remediation of sleep-deprivation-
induced working memory impairment with fMRI-guided
transcranial magnetic stimulation. Hirnrinde,
18, 2077–2085.
Marois, R., & Ivanoff, J. (2005). Capacity limits of information
processing in the brain. Trends in den Kognitionswissenschaften, 9,
296–305.
Mottaghy, F. M., Gangitano, M., Krause, B. J., & Pascual-Leone, A.
(2003). Chronometry of parietal and prefrontal activations
in verbal working memory revealed by transcranial magnetic
stimulation. Neurobild, 18, 565–575.
Mottaghy, F. M., Krause, B. J., Kemna, L. J., Töpper, R.,
Tellmann, L., Beu, M., et al. (2000). Modulation of the
neuronal circuitry subserving working memory in healthy
human subjects by repetitive transcranial magnetic
stimulation. Neurowissenschaftliche Briefe, 280, 167–170.
Mottaghy, F. M., Sparing, R., & Topper, R. (2006). Enhancing
picture naming with transcranial magnetic stimulation.
Behavioural Neurology, 17, 177–186.
Mull, B. R., & Seyal, M. (2001). Transcranial magnetic
stimulation of left prefrontal cortex impairs working
Erinnerung. Clinical Neurophysiology, 112, 1672–1675.
OʼReardon, J. P., Solvason, H. B., Janicak, P. G., Sampson, S.,
Isenberg, K. E., Nahas, Z., et al. (2007). Efficacy and safety
of transcranial magnetic stimulation in the acute treatment
of major depression: A multisite randomized controlled
trial. Biologische Psychiatrie, 62, 1208–1216.
Osaka, N., Otsuka, Y., Hirose, N., Ikeda, T., Mima, T., Fukuyama,
H., et al. (2007). Transcranial magnetic stimulation (TMS)
applied to left dorsolateral prefrontal cortex disrupts verbal
working memory performance in humans. Neurowissenschaften
Letters, 418, 232–235.
Osipova, D., Ahveninen, J., Jensen, O., Ylikoski, A., & Pekkonen,
E. (2005). Altered generation of spontaneous oscillations
in Alzheimerʼs disease. Neurobild, 27, 835–841.
Osuch, E. A., Benson, B. E., Luckenbaugh, D. A., Geraci, M.,
Post, R. M., & McCann, U. (2008). Repetitive TMS combined
with exposure therapy for PTSD: A preliminary study.
Journal of Anxiety Disorders, 23, 54–59.
Preston et al.
455
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
T
.
.
F
.
/
Ö
N
1
8
M
A
j
2
0
2
1
Plewnia, C., Rilk, A. J., Soekadar, S. R., Arfeller, C., Huber, H. S.,
Sauseng, P., et al. (2008). Enhancement of long-range EEG
coherence by synchronous bifocal transcranial magnetic
stimulation. Europäisches Journal für Neurowissenschaften, 27,
1577–1583.
Postle, B. R., Ferrarelli, F., Hamidi, M., Feredoes, E., Massimini,
M., Peterson, M., et al. (2006). Repetitive transcranial
magnetic stimulation dissociates working memory
manipulation from retention functions in the prefrontal,
but not posterior parietal, Kortex. Zeitschrift für Kognition
Neurowissenschaften, 18, 1712–1722.
Rounis, E., Yarrow, K., & Rothwell, J. C. (2007). Effects of
rTMS conditioning over the fronto-parietal network on
motor versus visual attention. Zeitschrift für Kognition
Neurowissenschaften, 19, 513–524.
Rypma, B., Berger, J. S., & DʼEsposito, M. (2002). The influence
of working-memory demand and subject performance on
prefrontal cortical activity. Zeitschrift für Kognition
Neurowissenschaften, 14, 721–731.
Rypma, B., Berger, J. S., Genova, H. M., Rebbechi, D., &
DʼEsposito, M. (2005). Dissociating age-related changes
in cognitive strategy and neural efficiency using
event-related fMRI. Kortex, 41, 582–594.
Rypma, B., Berger, J. S., Prabhakaran, V., Bly, B. M., Kimberg,
D. Y., Biswal, B. B., et al. (2006). Neuronale Korrelate von
cognitive efficiency. Neurobild, 33, 969–979.
Rypma, B., Prabhakaran, V., Desmond, J. E., & Gabriela, J. D.
(2001). Age differences in prefrontal cortical activity in
Arbeitsgedächtnis. Psychologie und Altern, 16, 371–384.
Sauseng, P., & Klimesch, W. (2008). What does phase
information of oscillatory brain activity tell us about
cognitive processes? Neuroscience and Biobehavioral
Rezensionen, 32, 1001–1013.
Sternberg, S. (1966). High-speed scanning in human
Erinnerung. Wissenschaft, 153, 652–654.
Uhlhaas, P. J., & Singer, W. (2007). What do disturbances
in neural synchrony tell us about autism? Biologisch
Psychiatrie, 62, 190–191.
Vogt, S., Wellhornschnecke, G., Wohlschlager, A. M., Canessa, N., Shah,
N. J., Zilles, K., et al. (2007). Prefrontal involvement in
imitation learning of hand actions: Effects of practice
and expertise. Neurobild, 37, 1371–1383.
Wassermann, E. M. (1998). Risk and safety of repetitive
transcranial magnetic stimulation: Report and suggested
guidelines from the International Workshop on the Safety
of Repetitive Transcranial Magnetic Stimulation, June 5–7,
1996. Electroencephalography and Clinical Neurophysiology,
108, 1–16.
Yener, G. G., Güntekin, B., Oniz, A., & Basar, E. (2007). Increased
frontal phase-locking of event-related theta oscillations in
Alzheimer patients treated with cholinesterase inhibitors.
International Journal of Psychophysiology, 64, 46–52.
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
3
e
–
4
P
4
D
7
F
1
/
9
2
3
2
8
/
4
3
6
/
1
4
Ö
4
C
7
N
/
1
2
0
7
0
6
9
9
2
2
8
1
7
2
0
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
0
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
.
.
F
.
T
/
Ö
N
1
8
M
A
j
2
0
2
1
456
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 3