Dynamics of White Matter Plasticity Underlying Working
Memory Training: Multimodal Evidence from
Diffusion MRI and Relaxometry
Claudia Metzler-Baddeley1, Sonya Foley1, Silvia de Santis2, Cyril Charron1,
Adam Hampshire3, Karen Caeyenberghs4, and Derek K. Jones1,4
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
Abstrakt
■ Adaptive working memory (WM) training may lead to cogni-
tive benefits that are associated with white matter plasticity in
parietofrontal networks, but the underlying mechanisms remain
poorly understood. We investigated white matter microstructural
changes after adaptive WM training relative to a nonadaptive
comparison group. Microstructural changes were studied in the
superior longitudinal fasciculus, the main parietofrontal connec-
tion, and the cingulum bundle as a comparison pathway. MRT-
based metrics were the myelin water fraction and longitudinal
relaxation rate R1 from multicomponent relaxometry (captured
with the mcDESPOT approach) as proxy metrics of myelin, Die
restricted volume fraction from the composite hindered and re-
stricted model of diffusion as an estimate of axon morphology,
and fractional anisotropy and radial diffusivity from diffusion ten-
sor imaging. PCA was used for dimensionality reduction. Adaptive
training was associated with benefits in a “WM capacity” compo-
nent and increases in a microstructural component (increases in
R1, restricted volume fraction, fractional anisotropy, and reduced
radial diffusivity) that predominantly loaded on changes in the
right dorsolateral superior longitudinal fasciculus and the left
parahippocampal cingulum. Im Gegensatz, nonadaptive comparison
activities were associated with the opposite pattern of reductions
in WM capacity and microstructure. No group differences were
observed for the myelin water fraction metric suggesting that R1
was a more sensitive “myelin” index. These results demonstrate
task complexity and location-specific white matter microstructural
changes that are consistent with tissue alterations underlying
myelination in response to training. ■
EINFÜHRUNG
Training schedules that adapt task difficulty to optimally
challenge a trainee have been shown to maximize train-
ing benefits and plasticity (Metzler-Baddeley & Baddeley,
2009; Smith et al., 2009). Zum Beispiel, Arbeitsgedächtnis
( WM) Kapazität, our ability to temporarily maintain and
manipulate information (Baddeley & Hitch, 1974), can be
enhanced with adaptive training (Melby-Lervåg & Hulme,
2013; Shipstead, Redick, & Engle, 2012; Morrison & Chein,
2011), and such WM benefits are associated with plastic
changes in parietofrontal networks (Caeyenberghs,
Metzler-Baddeley, Foley, & Jones, 2016; Metzler-Baddeley,
Caeyenberghs, Foley, & Jones, 2016; Takeuchi, Solch, &
Kawashima, 2010; Olesen, Westerberg, & Klingberg, 2004).
The neural substrates underpinning such plastic changes,
Jedoch, remain poorly understood.
Kürzlich, we compared the effects of 2 months of adap-
tive WM training (Cogmed, 2012; Klingberg et al., 2005)
with a nonadaptive comparison activity that trained the
same tasks but on three item spans only. Subtle changes
1Cardiff University, Brain Research Imaging Centre (CUBRIC),
2University Miguel Hernandez, Alicante, Spanien, 3Imperial College
London, 4Australian Catholic University
across a number of gray matter regions including increased
cortical thickness in the right frontal cortex and increased
volume of the left pallidum were associated with adaptive
WM training, whereas reductions in cortical thickness in
the right pars triangularis were associated with repeated
unchallenging comparison activities (Metzler-Baddeley
et al., 2016). Using graph theoretical analysis (GTA) von
white matter microstructural metrics, we found improved
global integration within the right parietofrontal network
after adaptive WM training (Caeyenberghs et al., 2016).
This increase in network global efficiency was best cap-
tured by MR relaxation rates, notably the longitudinal
relaxation rate R1 and was positively correlated with WM
benefits.
Although GTA is helpful in understanding the effects of
training at the global network level, this method con-
founds differences in the connectivity profile with dif-
ferences in the microstructure of those connections.
Darüber hinaus, the global approach precludes the interroga-
tion of individual “edges” in the graph. Daher, on the basis
of GTA alone, it can be difficult to infer about the nature
and location of microstructural alterations within the net-
arbeiten, and subtle activity-related changes may be missed.
The aim of this study was therefore to explore the neural
substrates underpinning adaptive training-induced white
© 2017 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 3.0 Unportiert (CC BY 3.0) Lizenz.
Zeitschrift für kognitive Neurowissenschaften 29:9, S. 1509–1520
doi:10.1162/jocn_a_01127
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
F
.
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
matter plasticity on the local level within parietofrontal
white matter of the superior longitudinal fasciculus
(SLF).
The SLF is the largest intrahemispheric parietofrontal
connection and comprises dorsal-superior (SLF1), zentral
(SLF2), and ventral-inferior (SLF3) parietofrontal white
matter (Thiebaut de Schotten et al., 2011; Makris et al.,
2005; Figur 1). We expected plastic changes in the SLF
because this bundle connects parietofrontal cortical re-
gions that are known to be important for WM functions
such as action control and organization (Dosenbach, Fair,
Cohen, Schlaggar, & Petersen, 2008; Rizzolatti & Matelli,
2003) and have been shown to change with WM training
(Takeuchi et al., 2011; Takeuchi, Sekiguchi, et al., 2010;
McNab et al., 2009; Olesen et al., 2004). To assess the
specificity of SLF changes, the subgenual (SGC), retro-
splenial (RSC), and parahippocampal (PHC) portions of
the cingulum bundle ( Jones, Christiansen, Chapman, &
Aggleton, 2013; Mufson & Mesulam, 1982) were also re-
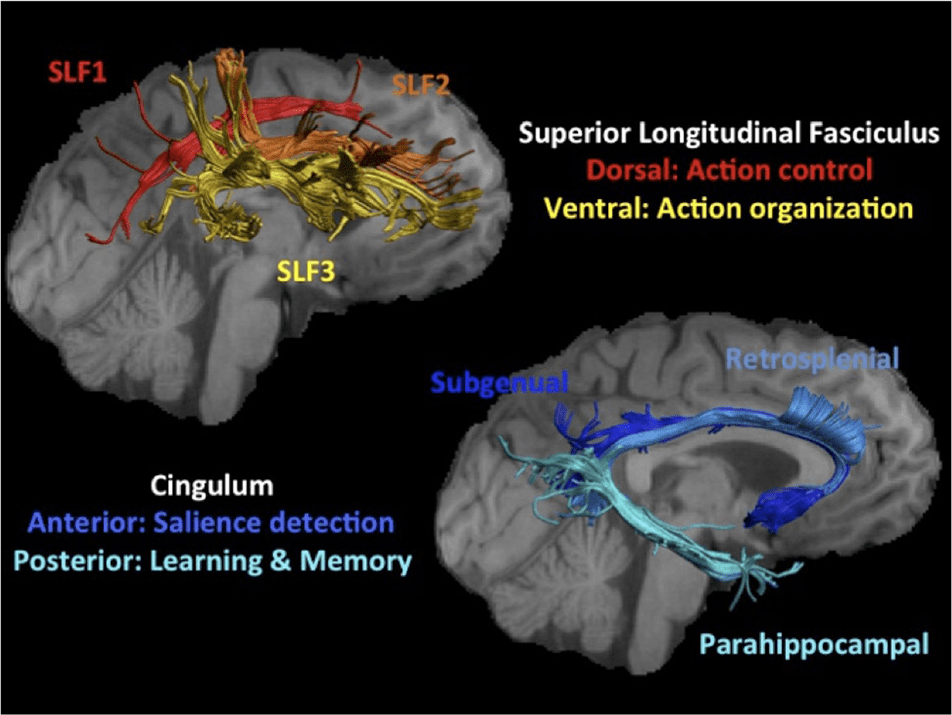
Figur 1. The top left hand visualizes the three subfascicles of the
right SLF. The tracts were reconstructed for one participant and were
displayed on their T1-weighted image visualizing the lateral view of
the right hemisphere. The SLF1 (Rot) connects the dorsal-superior
parietofrontal regions of the visual attention network proposed to be
important for action control in WM (Rizzolatti & Matelli, 2003). SLF3
(Gelb) connects ventral-inferior parietofrontal regions thought to be
important for action organization and recognition (Rizzolatti & Matelli,
2003). SLF2 (orange) comprises central parietofrontal white matter that
has been suggested to allow the cross-communication between dorsal
and ventral visual attention networks (Thiebaut de Schotten et al.,
2011). Parietofrontal cortical regions have been reported to undergo
structural and functional changes after WM training (Takeuchi et al.,
2011; Takeuchi, Sekiguchi, et al., 2010; McNab et al., 2009; Olesen et al.,
2004). The lower right image displays the three subfascicles of the
cingulum bundle (SGC in dark blue, RSC in blue, PHC in light blue)
reconstructed for the same participant and displayed on a medial
view of the right hemisphere. The cingulum bundle was chosen as
comparison pathway for the SLF because SGC and RSC maintain
projections to and from anterior salience network regions, welche haben
been proposed to support WM action control by detecting salient
stimuli in the environment (Dosenbach et al., 2008). The PHC forms
part of the medial-temporal lobe network known to be important for
learning and episodic memory.
constructed as comparison pathways. SGC and RSC main-
tain anterior cingulate projections and form part of the
salience network (Dosenbach et al., 2008), wohingegen
PHC forms part of the extended medial-temporal lobe
memory network ( Jones, Christiansen, et al., 2013).
White matter plasticity is thought to be largely driven
by axon myelination (Fields et al., 2014; Zatorre, Fields, &
Johansen-Berg, 2012; Fields 2010), which regulates salta-
tory conduction and has been linked to neuronal activity
(Gibson et al., 2014) and to the learning of new motor
skills in animals (Fields et al., 2014; McKenzie et al.,
2014; Sampaio-Baptista et al., 2013). The formation and
remodeling of myelin is associated with changes in the
biochemical features of brain tissue, such as alterations
in water, lipids, proteins, and iron content within oligo-
dendrocytes (Alexander et al., 2011). Although white mat-
ter plasticity in the human brain has been predominantly
studied with diffusion tensor MRI (DT-MRI)-based metrics
of fractional anisotropy (FA) or diffusivities (Pierpaoli &
Basser, 1996), these indices are not specific to any white
matter property and therefore difficult to interpret in
terms of biological changes (De Santis, Drakesmith, Bells,
Assaf, & Jones, 2014).
In this study, we therefore applied the myelin water
fraction (MWF) and the longitudinal relaxation rate R1
from the multicomponent-driven equilibrium single-
pulse observation of T1 and T2 (mcDESPOT; Deoni &
Kolind, 2015; Deoni, 2011B) as proxy metrics of myelin
and the restricted volume fraction (RVF) from the com-
posite hindered and restricted model of diffusion
(CHARMED; Assaf & Basser, 2005; Assaf, Freidlin, Rohde,
& Basser, 2004) as a proxy metric of axon morphology.
These measures were combined with FA for the purposes
of comparability with previous training studies that re-
ported increases in FA (Zatorre et al., 2012; Lövdén et al.,
2010; Takeuchi, Sekiguchi, et al., 2010; Scholz, Klein,
Behrens, & Johansen-Berg, 2009) and with radial diffusivity
(RD). Following evidence showing a link between RD and
myelin in coaxially aligned fibers in the mouse brain (Song
et al., 2005), RD is often interpreted as a metric of myelin
despite the inherent problems of interpreting DT-MRI
measures in terms of specific biological white matter
properties (Wheeler-Kingshott & Cercignani, 2009).
Assuming that white matter plasticity would be driven
by myelin-related tissue changes, we hypothesized in-
creases in MWF and R1 relaxation time due to reductions
in T1 and T2 components with increasing myelination
(Barkhof & van Walderveen, 1999). We also expected
training-related increases in RVF due to plasticity-related
changes in glia cell morphology (Tavor, Hofstetter, &
Assaf, 2013) as well as increased FA and reduced RD, welche
have previously been linked to myelin plasticity (Sampaio-
Baptista et al., 2013; Zatorre et al., 2012). Training-induced
microstructural changes were expected in parietofrontal
SLF connections, whereas no specific alterations were
hypothesized for the cingulum bundle since the extent
to which salience network and limbic memory regions
1510
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
/
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
may alter with WM trainining remains a matter of debate
( Jolles, van Buchem, Crone, & Rombouts, 2013; Rose,
Olsen, Craik, & Rosenbaum, 2012). Endlich, we explored
whether training-induced microstructural changes would
correlate with cognitive benefits ( Valkanova, Rodriguez,
& Ebmeier, 2014).
METHODEN
Detailed descriptions of the training procedure and cog-
nitive outcome assessments can be found in Caeyenberghs
et al. (2016) and Metzler-Baddeley et al. (2016) and are
only briefly summarized here.
Teilnehmer
The study was approved by the Cardiff University School
of Psychology ethics committee. Forty-eight healthy
Erwachsene (19–40 years) participated and gave informed writ-
ten consent. Participants were randomly allocated to the
adaptive training or the comparison group with the pro-
vision that both groups were matched for age and sex.
Participants were blind to their training condition. Four
participants in the training and four in the comparison
group dropped out because of time commitments, leav-
ing 20 individuals in each group who completed the
Studie. The two groups were comparable in their demo-
graphics and baseline cognitive performance (Tisch 1).
WM Training
Participants performed computerized exercises of verbal
and spatial span tasks (Cogmed, 2012).1 Training was ac-
cessed via the Internet from home, and participants had
to practice five times per week for 8 weeks (40 Ausbildung
sessions of about 30 hr in total). Die Nummer, frequency,
and order of training tasks were identical for all partici-
Hose. Training progress was monitored, and participants
received weekly feedback by e-mail. In the adaptive train-
ing condition, task difficulty was altered depending on
the trainee’s level of performance to ensure that partici-
pants exercised at their maximum level of WM capacity.
Participants in the comparison group trained on level
three item spans throughout all training sessions. Beide
groups completed the same number of training sessions
Aber, on average, the comparison group spent 7 min less
per session because each trial was on average shorter
(Tisch 1).
Cognitive Assessment
Participants were tested before and after the training with
a previously validated battery of computerized assess-
ment tests from Cambridge Brain Sciences (www.
cambridgebrainsciences.com; Hampshire, Highfield,
Parkin, & Owen, 2012). WM capacity was tested with
forward and backward digit span and spatial span, dis-
tractor suppression with an adapted version of the
Stroop test (Double Trouble), problem solving with a
Tisch 1. Summary of Demographic Variables and Mean (SD) Performance in WM and Executive Function Benchmark Tests of the
Two Groups at Baseline
N
Alter (Jahre)
Female
Right-handed
Forward digit span
Backward digit span
Spatial span
Stroop (double trouble)
Grammatical reasoning
Tree task
Odd-one-out
Self-ordered search
Symmetry span
Number of training sessions
Training
20
26 (6.2)
11
19
5.3 (0.8)
4 (1.4)
5 (0.5)
22.8 (13.6)
0.79 (0.2)
23.7 (8.7)
9.5 (3.2)
6.2 (1.1)
25.3 (6.5)
40
Training time per session (min)
42.7 (4.65)
Controls
20
27 (6.8)
10
20
5.2 (0.7)
4 (1.4)
4.9 (0.5)
25.9 (15.4)
0.73 (0.2)
19.8 (7.2)
9.1 (4.3)
5.5 (1.4)
22.6 (7.9)
39.9 (0.44)
36.31 (6.15)
T(38)
–
0.44
–
–
0.67
0.01
0.97
0.69
0.97
0.15
0.37
0.18
0.12
1.00
3.75
P
–
.67
–
–
.51
.99
.34
.49
.34
.93
.71
.07
.25
.32
.001
Metzler-Baddeley et al.
1511
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
.
T
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
version of the Tower of London task (the Tree task),
abstract reasoning with grammatical reasoning and the
odd-one-out task, and the ability to manipulate and orga-
nize spatial information with a self-ordered spatial span
Aufgabe. Multitasking abilities were tested with the automated
symmetry span task (Unsworth, Heitz, Schrock, & Engle,
2005).
MRT-Datenerfassung
MRI data were acquired on a 3-T General Electric HDx
MRI system (GE Medical Systems, Milwaukee, WI) verwenden
an eight-channel receiver-only head RF coil at the Cardiff
University Brain Research Imaging Centre. MRI sessions
were interleaved for both groups to avoid confounds be-
tween the experimental conditions and any potential
scanner-related changes in data acquisition (Thomas &
Bäcker, 2013). T1-weighted anatomical FSPGR images
(256 × 256 acquisition matrix, repetition time [TR] =
7.8 ms, Echozeit [DER] = 2.9 ms, flip angle = 20, 172
Scheiben, 1 mm slice thickness, Sichtfeld = 23 cm) war
acquired. Diffusion data were collected with a spin-echo
echo-planar high-angular resolution diffusion imaging
(Tuch et al., 2002) sequence with diffusion encoded
along 60 isotropically distributed orientations according
to an optimized gradient vector scheme (Jones, Horsfield,
& Simmons, 1999) and six nondiffusion weighted scans
(TR/TE = 87 ms, b value = 1200 sec/mm2, 60 Scheiben,
96 × 96 acquisition matrix, Sichtfeld = 230 ×
230 mm, 2.4 mm slice thickness, reconstructed spatial
Auflösung 1.8 × 1.8 × 2.4 mm). Data acquisition was
peripherally gated to the cardiac cycle with a total acqui-
sition time of ∼30 min. To gain RVF, data were acquired
with the CHARMED protocol (TE = 126 ms, TR =
17,000 ms, 45 gradient orientations distributed on four
shells, Scheibendicke = 2.4 mm, maximum b value =
8700 sec/mm2, spatial resolution 2.4 mm isotropic, Erwerb
Zeit 13 min). To gain MWF and R1 maps, data were
acquired with the mcDESPOT protocol (spoiled gradient
recalled echo [SPGR] acquisitions: TE = 2.1 ms, TR =
4.7 ms, flip angles = [3°, 4°, 5°, 6°, 7°, 9°, 13°, 18°];
balanced steady-state free precession [bSSFP] acquisitions:
TE = 1.6 ms, TR = 3.2 ms, flip angles = [10.6°, 14.1°,
18.5°, 23.8°, 29.1°, 35.3°, 45°, 60°], spatial resolution 1.7 mm
isotropic, acquisition time 12 min; Deoni, Rutt, Arun,
Pierpaoli, & Jones, 2008). bSSFP acquisitions were repeated
with and without 180° RF phase alteration to remove SSFP
banding artifacts, and SPGR and IR-SPGR acquisitions were
used to correct B0- and B1-induced errors in the derived
MWF estimates (Deoni, 2011A).
MRI Data Processing
The diffusion-weighted data were corrected for distor-
tions induced by the diffusion-weighted gradients, arti-
facts due to head motion and EPI-induced geometrical
distortions by registering each image volume to the T1-
weighted anatomical images (Irfanoglu, Walker, Sarlls,
Marenco, & Pierpaoli, 2012), with appropriate reorienta-
tion of the encoding vectors (Leemans & Jones, 2009) In
ExploreDTI ( Version 4.8.3; Leemans, Jeurissen, Sijbers, &
Jones, 2009). A two compartment model using the free
water elimination (FWE) Ansatz (Pasternak, Sochen,
Gur, Intrator, & Assaf, 2009) was then fitted to derive
maps of (CSF partial volume-corrected) FA and RD in each
voxel (Metzler-Baddeley, O’Sullivan, Bells, Pasternak, &
Jones, 2012). CHARMED data were corrected for motion
and distortion artifacts according to the extrapolation
method of Ben-Amitay, Jones, and Assaf (2012) and cor-
rected for CSF partial volume with the FWE approach.
The number of distinct fiber populations (1, 2, oder 3) In
each voxel was obtained using a model selection approach
(De Santis et al., 2014) and RVF, das ist, the fraction of the
signal assigned to restricted diffusion was calculated per
voxel with in-house software (De Santis et al., 2014) coded
in MATLAB (The MathWorks, Natick, MA).
The SPGR and bSSFP images acquired as part of the
mcDESPOT pipeline (Deoni, Rutt, Arun, et al., 2008;
Deoni, Rutt, & Jones, 2008) were corrected for motion
using the FMRIB Linear Image Registration Tool (Schmied,
De Stefano, Jenkinson, & Matthews, 2001) to align all
images to the first in the acquisition series. The mcDESPOT
model was fitted to the data using in-house software coded
in C++ (Deoni, Rutt, Arun, et al., 2008; Deoni, Rutt, &
Jones, 2008) to obtain maps of MWF and of the intrinsic
relaxation times T1 and T2. All quantitative maps were
coregistered to the T1-weighted anatomical images. RVF
maps were coregistered to the anatomical image using
the Elastix registration toolbox (Klein, Staring, Murphy,
Viergever, & Pluim, 2010) whereas MWF maps (derived
from mcDespot) were coregistered using the FMRIB non-
linear registration tool FNIRT (Andersson, Jenkinson, &
Schmied, 2010). Tract-specific MWF indices were corrected
for partial volume artifacts by normalizing these values
by their tissue volume fraction from the FWE approach
(Bells, Deoni, Pasternak, & Jones, 2011).
Whole-brain Tractography
Whole-brain tractography was performed for each data set
using the damped Richardson-Lucy algorithm (Dell’acqua
et al., 2010), which was calculated with in-house software
coded in MATLAB. The damped Richardson-Lucy tracking
algorithm estimated peaks in the fiber orientation density
Funktion (fODF) by selecting seed points at the vertices
of a 2 × 2 × 2 mm grid superimposed over the image
and propagated in 0.5-mm steps along these axes re-
estimating the fODF peaks at each new location (Jeurissen,
Leemans, Jones, Tournier, & Sijbers, 2011). Tracks were
terminated if the fODF threshold fell below 0.05 oder der
direction of pathways changed through an angle greater
than 45°. This procedure was then repeated by tracking
in the opposite direction from the initial seed point.
Three-dimensional fiber reconstructions of the SLF
1512
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
and cingulum subfascicles were achieved by applying
waypoint ROI gates (“AND”, “OR,” and “NOT” gates fol-
lowing Boolean logic) to isolate specific tracts from the
whole-brain tractography data. ROIs were drawn manually
by three operators blind to the experimental group and
time of assessment of each data set on color-coded
fiber orientation maps in native space following previ-
ously validated anatomical landmark protocols ( Jones,
Christiansen, et al., 2013; Thiebaut de Schotten et al.,
2011; Makris et al., 2005). Pairs of baseline and outcome
data sets from the different experimental conditions
were counterbalanced across the three operators.
Reconstructions of SLF and Cingulum
Bundle Subfascicles
All tracts were reconstructed separately for each hemi-
Kugel. The reconstructions of the three SLF subfascicles
followed the protocol by Thiebaut de Schotten et al.
(2011). A coronal seed ROI was placed at the level of
the posterior commissure around the parietal lobe and
an axial “NOT” ROI was placed at the level of the lateral
sulci around the temporal lobes. SLF1 was located by
placing a coronal “AND” ROI at the level with the anterior
commissure around the superior frontal gyrus, SLF2 by
placing the ROI around the middle frontal gyrus, Und
SLF3 by drawing the ROI around the inferior frontal gyrus.
The reconstructions of the cingulum fascicles followed
the protocol by Jones, Christiansen, et al. (2013). SGC
was located by placing two coronal “AND” ROIs: eins
around the SGC below the corpus callosum (CC) und ein-
other one around the cingulum anterior to the midline of
the CC (identified on the sagittal plane). The RSC was re-
constructed by placing two “AND” ROIs around the cin-
gulum: one coronal ROI posteriorly to the CC midline
and one axial “retrosplenial” ROI dorsal to the ventral lim-
it of the splenium. The PHC was located by drawing the
“retrosplenial” ROI and a second axial “AND” ROI at the
level of the cerebral peduncle. Figur 1 displays reconstruc-
tions of the SLF and cingulum subfascicles for one repre-
sentative data set. Average values of all microstructural
metrics were obtained for each white matter tract. Der
reliability of the metrics derived from the tracts across
the three operators was assessed with intraclass correlation
coefficient (ICC) for six randomly chosen data sets. MWF,
R1, and RVF had reliable ICCs of >.8 in all tracts, Und
FA and RD had ICCs of >.8 for the majority of tracts.
Statistical Analyses
Statistical analyses were carried out in SPSS Version 20.0
(IBM, 2011). All data were inspected for outliers, defined
as values more than three times the standard deviation
from the average cognitive or microstructural index for
each time point and group. The left SLF1 pathway could
not be reconstructed for two adaptive baseline data sets,
one adaptive outcome data set, one comparison baseline
data set, and three comparison outcome data sets. Rechts
SLF1 subfascicles could not be reconstructed for three
baseline and two outcome comparison data sets. Daher,
these data were missing from the microstructural analy-
ses. Because of a technical difficulty, four symmetry span
data sets were lost during data acquisition (one training
and three control). For all remaining data, changes in
cognitive and microstructural metrics were calculated
for each participant as difference scores between post-
and pretraining values.
Because performance measures in WM and executive
function tasks have been shown to share underlying cog-
nitive structures (Testa, Bennett, & Ponsford, 2012) Und,
similarly, microstructural metrics are known to correlate
between white matter pathways (Penke et al., 2010),
PCA was employed to reduce the complexity of the cog-
nitive change scores in the nine benchmark tests and
Die 60 microstructural change scores (5 metrics ×
6 pathways × 2 hemispheres). PCAs were run on change
Tisch 2. Rotated Component Loadings on Change in the Cognitive Benchmark Tests
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
.
/
Cognitive Change
Forward digit span
Backward digit span
Spatial span
Stroop (double trouble)
Grammatical reasoning
Tree task
Odd-one-out
Self-ordered search
Symmetry span
Loadings >0.5 are highlighted in bold.
Executive
−0.017
0.230
0.428
0.793
−0.746
0.102
−0.072
0.568
0.038
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
WM Capacity
Problem Solving
0.588
0.244
0.670
−0.190
−0.278
0.220
0.470
0.068
0.653
0.177
0.677
0.167
0.019
−0.032
−0.801
0.514
−0.01
−0.24
Metzler-Baddeley et al.
1513
Tisch 3. Rotated Component Loadings on Change in White Matter Microstructure
MWF–R1
Left SLF1–Left SGC
Right SLF1–Left PHC
Right SGC–RSC
Myelin Water Fraction (MWF)
SGC
RSC
PHC
SLF1
SLF2
SLF3
L
R
L
R
L
R
L
R
L
R
L
R
0.421
0.483
0.767
0.303
0.624
0.508
0.333
0.274
0.525
0.624
0.733
0.654
Longitudinal Relaxation Rate R1
SGC
RSC
PHC
SLF1
SLF2
SLF3
L
R
L
R
L
R
L
R
L
R
L
R
0.064
0.518
0.304
0.073
0.175
0.078
−0.178
−0.065
0.130
0.583
0.525
0.516
Restricted Volume Fraction (RVF)
SGC
RSC
PHC
SLF1
SLF2
L
R
L
R
L
R
L
R
L
R
−0.025
−0.199
0.509
−0.072
0.437
0.150
−0.116
−0.111
−0.378
0.385
0.425
−0.319
0.004
0.458
0.291
−0.325
0.620
0.425
0.175
−0.107
−0.001
−0.045
0.531
0.086
0.107
0.512
0.339
−0.358
0.689
0.385
0.265
−0.069
−0.034
0.070
0.588
−0.230
0.127
0.159
0.336
−0.242
0.694
0.149
0.487
−0.023
−0.175
−0.323
0.129
−0.025
0.337
0.340
−0.190
0.243
−0.492
−0.387
−0.092
−0.40
0.125
−0.035
0.526
0.397
0.701
0.351
−0.034
0.512
−0.482
−0.158
0.212
−0.115
0.127
−0.008
0.121
−0.221
0.193
−0.179
−0.284
0.611
−0.146
−0.069
0.015
0.506
−0.146
0.380
−0.057
0.024
−0.117
0.357
−0.025
0.142
−0.109
−0.204
−0.089
0.605
−0.127
0.363
−0.090
−0.074
−0.109
0.379
−0.067
0.136
−0.193
−0.259
0.166
0.533
0.146
0.433
0.034
−0.028
−0.040
0.443
0.300
0.190
1514
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Tisch 3. (Fortsetzung )
SLF3
L
R
Fractional Anisotropy (FA)
SGC
RSC
PHC
SLF1
SLF2
SLF3
L
R
L
R
L
R
L
R
L
R
L
R
Radial Diffusivity (RD)
SGC
RSC
PHC
SLF1
SLF2
SLF3
L
R
L
R
L
R
L
R
L
R
L
R
MWF–R1
−0.047
0.229
−0.067
0.05
−0.287
−0.073
0.15
0.378
0.071
−0.186
−0.05
0.211
−0.228
0.494
−0.059
−0.061
0.131
−0.006
0.118
−0.389
−0.034
0.246
0.042
−0.317
−0.372
−0.514
Loadings >0.5 are highlighted in bold.
Left SLF1–Left SGC
Right SLF1–Left PHC
Right SGC–RSC
−0.136
0.285
0.532
−0.149
−0.176
0.044
0.128
−0.47
0.729
0.31
0.218
0.284
0.018
−0.187
−0.502
0.189
0.254
−0.069
−0.019
0.527
−0.692
−0.226
−0.369
−0.194
−0.059
0.172
0.323
−0.263
0.055
0.147
0.236
−0.038
0.577
0.414
−0.304
0.584
−0.715
−0.281
0.264
−0.062
−0.125
−0.130
−0.354
−0.164
−0.675
−0.109
0.374
−0.593
0.753
0.287
−0.094
0.078
−0.077
0.189
0.128
0.835
−0.186
0.709
−0.127
−0.017
−0.136
0.450
−0.030
0.388
0.071
0.153
−0.025
−0.827
−0.059
−0.603
0.030
−0.061
0.166
−0.413
−0.030
−0.438
0.269
−0.131
scores for all participants across both groups. Performance
Veränderungen in der 11 trained Cogmed tasks were not in-
cluded in the PCA since due to the nature of the com-
parison activities (three-item span practice only) alle
control participants had zero change scores for all trained
tasks. Participants in the adaptive training group, Jedoch,
showed significant improvements in all trained Cogmed tasks
(siehe Abbildung 3 und Tisch 3 in Metzler-Baddeley et al., 2016).
Given the relatively small sample size for PCA, Wir
followed recommendations to limit the number of ex-
tracted components as much as possible (de Winter,
Dodou, & Wieringa, 2009; Preacher & MacCallum, 2002).
Choosing the number of components for data summary
is always a compromise between selecting too few com-
ponents that may miss important structures and too many
components that reflect noise. Since there is no single
recommended method available, we adopted a threefold
Ansatz: Erste, we employed the SPSS default of the
Kaiser criterion of including all components with an eigen-
value of >1 (IBM, 2011). Zweite, we inspected Cattell’s
Metzler-Baddeley et al.
1515
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
.
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
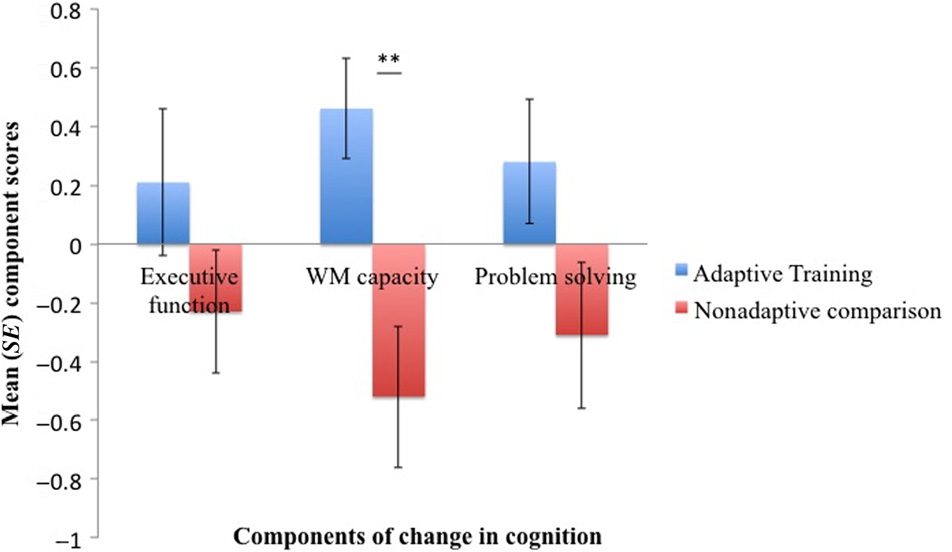
component loaded on performance changes in the for-
ward digit span, spatial span, and symmetry span task
and was therefore labeled “WM capacity” component.
The third component loaded on performance changes
in backward digit span, the tree task, and the odd-one-
out task and was labeled “problem-solving” component.
To find out if the two groups differed in change in cog-
Nation, independent t tests were carried out on the scores
for the three extracted components. The adaptive train-
ing group differed significantly from the comparison group in
the WM capacity component scores, T(34) = 3.33, p = .002,
but not in the executive, T(34) = 1.3, p = .19, or the problem-
solving component, T(34) = 1.8, p = .08 (Figur 2).
Training-related Changes in White
Matter Microstructure
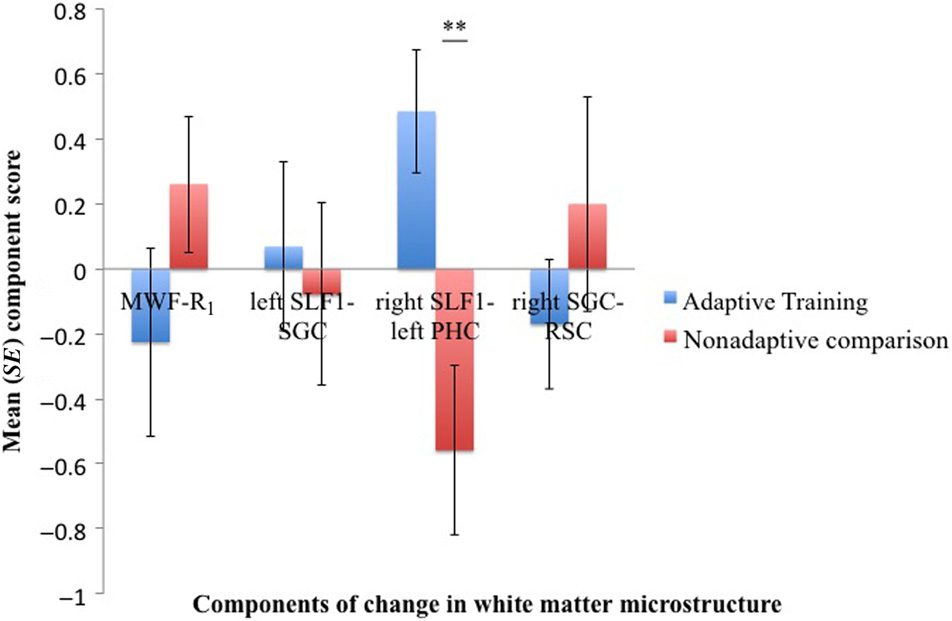
Four components of change in white matter microstruc-
ture were extracted, and they explained together 45% von
the variability in the data (Tisch 3). The first component
loaded (>0.5) predominantly on change in MWF and R1
(“MWF–R1” component), the second component on
change in the left SLF1 and the left SGC (“left SLF1–
SGC” component), the third component on changes in
the right SLF1, the left PHC, and the left SLF2 (“right
SLF1–left PHC”), and the fourth on changes in the right
SGC and right RSC (“right SGC–RSC” component).
To assess if there were any group differences in the
change in microstructure, independent t tests were car-
ried out on the scores for the four extracted components.
A significant group difference was present for the “right
SLF1–left PHC” component, T(26) = 3.2, p = .004, Aber
not for any of the other components (MWF–R1, p = .2; links
SLF1–SGC, p = .7; right SGC–RSC, p = .33; Figur 3).
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
F
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Figur 3. The bar charts display the mean component scores for
the four microstructural components extracted from the change scores
of average MWF, longitudinal relaxation rate R1, RVF, FA, and RD across
the SLF and the cingulum bundle. The adaptive training group (Blau)
differed significantly from the control group (Rot) in the “right SLF1–left
PHC”: Adaptive training was associated with positive change in this
component, whereas control activities were associated with negative
ändern. No differences were observed for the other three components.
SE = standard error. **p = .004.
Figur 2. The bar charts display the mean component scores for
the “executive function,” “WM capacity,” and “problem-solving”
components for the adaptive training group (Blau) and the nonadaptive
comparison group (Rot). Components were extracted from change
scores of the nine cognitive benchmark tests. The adaptive training
group differed significantly from the comparison group in the “WM
capacity” component: Adaptive training was associated with positive
ändern, whereas control activities were associated with negative
ändern. No difference was observed for the “executive” and the
“problem-solving” components. SE = standard error. **p = .002.
scree plots (Cattell, 1952) to identify the minimal number
of components that accounted for most of the variability
in the data. Dritte, we assessed each component with re-
gard to their interpretability. We used a PCA procedure
with orthogonal Varimax rotation of the component
Matrix. Tables 2 Und 3 summarize the component load-
ings for the cognitive and microstructural variables, Re-
spectively. Loadings that exceeded a value of 0.5 war
considered as significant.
Group differences in the component scores were then
assessed with independent t tests. Pearson correlation
coefficient was calculated between those cognitive and
microstructural components that showed significant group
differences to assess whether microstructural changes
were related to any cognitive benefits of the training.
All statistical tests were corrected for multiple comparison
errors with the Bonferroni correction with a family-wise
alpha level of 5% (two-tailed) leading to a corrected p value
von <.0163 for three independent t tests on cognitive
change, p < .0123 for four independent t tests on micro-
structural change, and p < .05 for one correlation coefficient.
RESULTS
Training-related Changes in Cognition
Three components that accounted for 55% of the variance
of performance changes in the cognitive benchmark tests
were extracted (Table 2). The first component loaded
highly (>0.5) on performance changes in the Stroop,
grammatical reasoning, and self-ordered search. Weil
these tasks all rely on a variety of executive functions
including focused attention, distractor suppression, orga-
nization, planning, and reasoning, the first component
was labeled “executive function” component. The second
1516
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 9
Correlation between Cognitive and
Microstructural Changes
To ascertain whether changes in microstructure were
related to changes in cognition, Pearson correlation co-
efficient was calculated between the “WM capacity” and
the “right SLF1–left PHC network” component scores.
No correlation was observed between these component
scores (r = .1, p = .68).
DISKUSSION
Nach 2 months of adaptive WM training, participants’ WM
capacity was improved compared with comparison
volunteers who practiced the same tasks but at a non-
challenging level of three items only (Figur 2).
The two groups also differed in a component that
loaded highly on microstructural changes in the right
SLF1, the left PHC, and the left SLF2. Adaptive training
was associated with a positive change in this component,
whereas comparison activities were associated with a
negative change (Figur 3). From the direction of the
component loadings (Tisch 3), we can infer that adap-
tive training led to increases in R1, RVF, and FA (positive
loadings) and to reductions in RD (negative loadings) In
the right SLF1 that connects superior parietal with supe-
rior and dorsal prefrontal cortical regions (Makris et al.,
2005). These changes were in the expected direction and
are consistent with previous reports of right-lateralized
changes in parietofrontal attention networks (Caeyenberghs
et al., 2016; Metzler-Baddeley et al., 2016; Takeuchi, Sekiguchi,
et al., 2010; Olesen et al., 2004). We also observed in-
creases in R1 and FA and reductions in RD in the left PHC
with projections from the posterior cingulate cortex, pari-
etal cortical regions, and the occipital lobe to the medial-
temporal lobes ( Jones, Christiansen, et al., 2013). Es ist
therefore likely that alterations in the PHC reflect learning,
Erinnerung, and visual modality-related plastic changes due
to the engagement with the adaptive training schedule.
In contrast to the PHC, we found no evidence for changes
in anterior portions of the cingulum bundle, notably the
SGC (with the exception of the “right SLF1-left PHC” com-
ponent loading on R1 in the left RSC), suggesting that the
ACC of the salience network was not significantly involved
in mediating training effects.
In contrast to the microstructural changes in the right
SLF1 and left PHC, we also observed reductions in FA
(negative loadings) and increases in RD (positive loading)
in the left central parietofrontal white matter of the SLF2.
These changes were only observed for FA and RD and are
likely the result of differences in fiber complexity and ori-
entation ( Jones, Knösche, & Turner, 2013). SLF fibers
cross with the corona radiata, an ascending white matter
bundle that fans out toward superior cortical regions.
Microstructural changes within SLF fibers relative to the
corona radiata may therefore have caused opposing effects
on diffusion metrics depending on the relative volume
fractions of the two white matter pathways. In superior
parietofrontal voxels, one would expect the relative vol-
ume fraction of the corona radiata to be smaller than the
one from the SLF1; somit, training-related increases in
microstructural properties of the SLF1 ought to result
in increased FA and reduced RD. In contrast in central
parietofrontal voxels the relative volume fractions of the
SLF2 and the corona radiata may be equal or even larger
for the corona radiata; somit, training-related changes in
SLF2 fibers might have paradoxically caused a reduction
in FA and increases in RD (De Santis et al., 2014). This ex-
ample highlights the importance of interpreting changes
in DT-MRI-based metrics not only in terms of biological
white matter properties but also in light of their geo-
metrical and architectural features.
Although fiber complexity-related effects may explain
the observed changes in the left SLF2, they cannot ac-
count for the opposing group differences in microstructure
im Laufe der Zeit (Figur 3). Figuren 2 Und 3 demonstrate a clear
separation between the two groups: Whereas adaptive
training was associated with positive changes in the micro-
structural and WM capacity component, comparison
activities were associated with negative changes in micro-
structure and cognition.
This pattern of opposing results was unexpected but
was observed across a number of modalities. Utilizing
morphological data, we recently found similar results of
increased cortical thickness for the adaptive training
group and reduced thickness for the comparison group
in right pFC regions (Metzler-Baddeley et al., 2016) Und
also observed opposing effects on global efficiency in the
parietofrontal network across the groups (Caeyenberghs
et al., 2016). We interpret these observations in light of
models proposing that activity levels in brain networks
are regulated in response to environmental demands with
the overall aim to minimize energy consumption (Laughlin
& Sejnowski, 2003). Adaptive training may have triggered
increased neural and hence axonal activation in parieto-
frontal WM networks, whereas comparison activities, fällig
to their repetitive and nonchallenge nature, may have
resulted in a down-regulation of signaling in these net-
works since they are not needed for the completion of
low-demanding tasks. Evidence from studies into the
effects of job demands suggests that prolonged un-
challenging activities may adversely affect cognition and
brain function. Gajewski et al. (2010) found reductions in
WM capacity and electroencephalography in older but
not younger assembly line employees compared with
age-matched managers. Ähnlich, Suo et al. (2012) Re-
ported that supervisory and managerial experience in
midlife was the largest predictor of total gray matter vol-
ume in the medial-temporal lobes in a group of older
Erwachsene. There is a clear need for replicating our findings
in a future training study that compares the effects of
nonadaptive activities with a passive nonintervention
Kontrolle. Together these results point to the possibility
though that the nature of prolonged activities may
Metzler-Baddeley et al.
1517
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
T
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
significantly impact on an individual’s brain structure
and cognition.
This study adopted three non-DT-MRI microstructural
indices, MWF and R1 from mcDESPOT as proxy estimates
of axon myelin and RVF from CHARMED as a proxy metric
of axon morphology with the aim to find out more about
the underpinnings of white matter plasticity. On the basis
of accumulating evidence suggesting that axonal activa-
tion may trigger biochemical processes in surrounding
glia cells that alter water, lipid, protein, and iron con-
centrations to induce myelination (Fields et al., 2014;
Fields, 2010), we expected to find adaptive training-
induced increases in MWF, R1, RVF, and FA and reductions
in RD. Although the expected changes were observed for
R1, RVF, FA, and RD, we did not find any evidence for
training-related changes in the MWF metric that was de-
rived from the two water pool mcDESPOT model. Der
latter has recently been found to provide insufficiently
precise parameter estimates to allow the unambiguous
estimation of specific tissue properties such as myelin
(Lankford & Does, 2013). Insbesondere, the two-pool model
may underestimate MWF in voxels affected by partial vol-
ume (Deoni, Matthews, & Kolind, 2013). In this study,
DT-MRI and CHARMED indices were corrected for CSF-
based partial volume artifacts with the FWE approach by
Pasternak et al. (2009), and tract-specific MWF indices
were corrected for partial volume by normalizing these
values by their tissue volume fraction from the FWE
method (Bells et al., 2011). The latter approach may not
have sufficiently corrected for partial volume in the MWF
metrics. Future studies should therefore apply Deoni
et al.’s (2013) novel three-component model that adds to
the myelin-associated water pool and the intra/extracellular
water pool a third “free water” component to model CSF-
based partial volume effects. This three-pool model might
provide MWF metrics that are more sensitive to subtle
training-induced changes in myelin.
A critical question for all training studies relates to the
functional significance of any observed plastic changes. In
this study, we did not observe any beneficial effects of
WM training on cognitive domains other than specific
WM capacity improvements assessed with verbal and spa-
tial span tasks. There is an ongoing debate in the training
literature regarding the far transfer effects of WM training
(Au et al., 2015; Corbett et al., 2015; Melby-Lervag &
Hulme, 2016; Owen et al., 2010), which goes beyond
the scope of the current article. Our results suggest that
WM training does not lead to generalization effects in
healthy adults, but we cannot rule out that transfer effects
may be measurable in considerably larger sample sizes
(Corbett et al., 2015). Zusätzlich, we did not observe a
correlation between microstructural and WM capacity
component scores. A lack of correlation between structural
and functional changes after training has been observed in a
number of training studies and may suggest that these pro-
cesses follow different time courses and may occur in differ-
ent brain regions (Valkanova, Rodriguez, & Ebmeier, 2013).
Zusammenfassend, we report activity- and location-dependent
plastic changes in the microstructure of parietofrontal and
parahippocampal white matter after adaptive versus non-
adaptive WM training. Microstructural changes were
captured by alterations in R1, RVF, FA, and RD and were
likely a result of biochemical changes related to myelin
remodeling.
Danksagungen
This research was made possible by a Wellcome Trust New
Investigator Award to D. K. J. (096646/Z/11/Z). We would like
to thank Greg Parker, Cardiff, for his assistance with scripting
the CHARMED analysis pipeline, Sonya Bells, Toronto, for her
assistance with the mcDESPOT processing pipeline and the
Elastix coregistration, Flavio dell’Acqua and Michel Thibaut de
Schotten, London, for their advice regarding SLF subfascicle
reconstructions, and Naomi Warne, Cardiff, for help with some
of the tract reconstruction.
Reprint requests should be sent to Claudia Metzler-Baddeley,
Cardiff University Brain Research Imaging Centre (CUBRIC),
School of Psychology, Cardiff University, Cardiff, CF24 4HQ, Vereinigtes Königreich, oder
via e-mail: Metzler-BaddeleyC@cardiff.ac.uk.
Notiz
1. Cogmed and Cogmed Working Memory Training are trade-
marks, in the US and/or other countries, of Pearson Education,
Inc. or its affiliate(S).
VERWEISE
Alexander, A. L., Hurley, S. A., Samsonov, A. A., Adluru, N.,
Hosseinbor, A. P., Mossahebi, P., et al. (2011). Characterization
of cerebral white matter properties using quantitative
magnetic resonance imaging stains. Gehirnkonnektivität, 1,
423–446.
Andersson, J., Jenkinson, M., & Schmied, S. (2010). Non-linear
registration, aka spatial normalisation. FMRIB technical
report TR07JA2.
Assaf, Y., & Basser, P. J. (2005). Composite hindered and
restricted model of diffusion (CHARMED) MR imaging of
das menschliche Gehirn. Neurobild, 27, 48–58.
Assaf, Y., Freidlin, R. Z., Rohde, G. K., & Basser, P. J. (2004).
New modeling and experimental framework to characterize
hindered and restricted water diffusion in brain white matter.
Magnetic Resonance in Medicine, 52, 965–978.
Au, J., Sheehan, E., Tsai, N., Duncan, G. J., Buschkuehl, M., &
Jaeggi, S. M. (2015). Improving fluid intelligence with
training on working memory: A meta-analysis. Psychonomic
Bulletin & Rezension, 22, 366–377.
Baddeley, A. D., & Hitch, G. J. (1974). Working memory. In
G. Bower (Ed.), Recent Advances in Learning and
Motivation Vol. VIII (S. 47–90). New York: Academic Press.
Barkhof, F., & van Walderveen, M. (1999). Characterization of
tissue damage in multiple sclerosis by nuclear magnetic
resonance. Philosophical Transactions of the Royal Society
of London, Serie B, Biological Sciences, 354, 1675–1686.
Bells, S., Deoni, S., Pasternak, O., & Jones, D. (2011). Partial
volume corrections of myelin water fraction values. In Proc
ISMRM 19th Annual Meeting, Montreal (S. 8076).
Ben-Amitay, S., Jones, D. K., & Assaf, Y. (2012). Motion
correction and registration of high b-value diffusion
weighted images. Magnetic Resonance in Medicine, 67,
1694–1702.
1518
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
F
T
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Caeyenberghs, K., Metzler-Baddeley, C., Foley, S., & Jones, D. K.
IBM. (2011). SPSS statistics, Ausführung 20.0. Armonk, New York: IBM
(2016). Dynamics of the human structural connectome
underlying working memory training. Zeitschrift für Neurowissenschaften,
36, 4056–4066.
Cattell, R. B. (1952). Factor analysis. New York: Harper.
Cogmed Working Memory Training. (2012). Pearson Education
(www.cogmed.com).
Corbett, A., Owen, A., Hampshire, A., Grahn, J., Stenton, R.,
Dajani, S., et al. (2015). The effect of an online cognitive
training package in healthy older adults: An online
randomized controlled trial. Journal of the American
Medical Directors Association, 16, 990–997.
De Santis, S., Drakesmith, M., Bells, S., Assaf, Y., & Jones, D. K.
(2014). Why diffusion tensor MRI does well only some of
die Zeit: Variance and covariance of white matter tissue
microstructure attributes in the living human brain.
Neurobild, 89, 35–44.
de Winter, J. C., Dodou, D., & Wieringa, P. A. (2009). Exploratory
factor analysis with small sample sizes. Multivariate
Behavioral Research, 44, 147–181.
Dell’acqua, F., Scifo, P., Rizzo, G., Catani, M., Simmons, A.,
Scotti, G., et al. (2010). A modified damped Richardson-Lucy
algorithm to reduce isotropic background effects in
spherical deconvolution. Neurobild, 49, 1446–1458.
Deoni, S. C. (2011A). Correction of main and transmit
magnetic field (B0 and B1) inhomogeneity effects in
multicomponent-driven equilibrium single-pulse observation
of T1 and T2. Magnetic Resonance in Medicine, 65,
1021–1035.
Deoni, S. C. (2011B). Magnetic resonance relaxation and
quantitative measurement in the brain. Methods in Molecular
Biology, 711, 65–108.
Deoni, S. C., & Kolind, S. H. (2015). Investigating the stability of
mcDESPOT myelin water fraction values derived using a
stochastic region contraction approach. Magnetic Resonance
in Medicine, 73, 161–169.
Deoni, S. C., Matthews, L., & Kolind, S. H. (2013). Eins
component? Two components? Drei? The effect of including
a nonexchanging “free” water component in multicomponent
driven equilibrium single pulse observation of T1 and T2.
Magnetic Resonance in Medicine, 70, 147–154.
Deoni, S. C., Rutt, B. K., Arun, T., Pierpaoli, C., & Jones, D. K.
(2008). Gleaning multicomponent T1 and T2 information
from steady-state imaging data. Magnetic Resonance in
Medicine, 60, 1372–1387.
Deoni, S. C., Rutt, B. K., & Jones, D. K. (2008). Investigating
exchange and multicomponent relaxation in fully-balanced
steady-state free precession imaging. Journal of Magnetic
Resonance Imaging, 27, 1421–1429.
Dosenbach, N. U., Fair, D. A., Cohen, A. L., Schlaggar, B. L.,
& Petersen, S. E. (2008). A dual-networks architecture of
top–down control. Trends in den Kognitionswissenschaften, 12, 99–105.
Fields, R. D. (2010). Change in the brain’s white matter.
Wissenschaft, 330, 768–769.
Fields, R. D., Araque, A., Johansen-Berg, H., Lim, S. S., Lynch,
G., Nave, K. A., et al. (2014). Glial biology in learning and
Erkenntnis. Neuroscientist, 20, 426–431.
Gajewski, P. D., Wild-Wall, N., Schapkin, S. A., Erdmann, U.,
Freude, G., & Falkenstein, M. (2010). Effects of aging and
job demands on cognitive flexibility assessed by task
switching. Biological Psychology, 85, 187–199.
Gibson, E. M., Purger, D., Mount, C. W., Goldstein, A. K., Lin,
G. L., Holz, L. S., et al. (2014). Neuronal activity promotes
oligodendrogenesis and adaptive myelination in the
mammalian brain. Wissenschaft, 344, 1252304.
Hampshire, A., Highfield, R. R., Parkin, B. L., & Owen, A. M.
(2012). Fractionating human intelligence. Neuron, 76,
1225–1237.
Corp.
Irfanoglu, M. O., Walker, L., Sarlls, J., Marenco, S., & Pierpaoli,
C. (2012). Effects of image distortions originating from
susceptibility variations and concomitant fields on diffusion
MRI tractography results. Neurobild, 61, 275–288.
Jeurissen, B., Leemans, A., Jones, D. K., Tournier, J. D., &
Sijbers, J. (2011). Probabilistic fiber tracking using the
residual bootstrap with constrained spherical deconvolution.
Kartierung des menschlichen Gehirns, 32, 461–479.
Jolles, D. D., van Buchem, M. A., Crone, E. A., & Rombouts, S. A.
(2013). Functional brain connectivity at rest changes after
working memory training. Kartierung des menschlichen Gehirns, 34,
396–406.
Jones, D. K., Christiansen, K. F., Chapman, R. J., & Aggleton,
J. P. (2013). Distinct subdivisions of the cingulum bundle
revealed by diffusion MRI fibre tracking: Implications for
neuropsychological investigations. Neuropsychologie, 51,
67–78.
Jones, D. K., Horsfield, M. A., & Simmons, A. (1999). Optimal
strategies for measuring diffusion in anisotropic systems by
magnetic resonance imaging. Magnetic Resonance in
Medicine, 42, 515–525.
Jones, D. K., Knösche, T. R., & Turner, R. (2013). White
matter integrity, fiber count, and other fallacies: Der
do’s and don’ts of diffusion MRI. Neurobild, 73,
239–254.
Klein, S., Staring, M., Murphy, K., Viergever, M. A., & Pluim, J. P.
(2010). elastix: A toolbox for intensity-based medical image
registration. IEEE Transactions on Medical Imaging, 29,
196–205.
Klingberg, T., Fernell, E., Olesen, P. J., Johnson, M., Gustafsson,
P., Dahlström, K., et al. (2005). Computerized training of
working memory in children with ADHD—A randomized,
controlled trial. Journal of the American Academy of Child &
Adolescent Psychiatry, 44, 177–186.
Lankford, C., & Does, M. (2013). On the inherent precision of
mcDESPOT [electronic version]. Magnetic Resonance in
Medicine, 69, 127–136.
Laughlin, S. B., & Sejnowski, T. J. (2003). Communication in
neuronal networks. Wissenschaft, 301, 1870–1874.
Leemans, A., Jeurissen, B., Sijbers, J., & Jones, D. K. (2009).
ExploreDTI: A graphical toolbox for processing, analyzing,
and visualizing diffusion MR data. Paper presented at the
17th Annual Meeting of International Society for Magnetic
Resonance in Medicine.
Leemans, A., & Jones, D. K. (2009). The B-matrix must be
rotated when correcting for subject motion in DTI data.
Magnetic Resonance in Medicine, 61, 1336–1349.
Lövdén, M., Bodammer, N. C., Kühn, S., Kaufmann, J., Schütze,
H., Tempelmann, C., et al. (2010). Experience-dependent
plasticity of white-matter microstructure extends into old
Alter. Neuropsychologie, 48, 3878–3883.
Makris, N., Kennedy, D. N., McInerney, S., Sorensen, A. G.,
Wang, R., Caviness, V. S., et al. (2005). Segmentation of
subcomponents within the superior longitudinal fascicle in
humans: A quantitative, in vivo, DT-MRI study. Zerebral
Kortex, 15, 854–869.
McKenzie, ICH. A., Ohayon, D., Li, H., de Faria, J. P., Emery, B.,
Tohyama, K., et al. (2014). Motor skill learning requires
active central myelination. Wissenschaft, 346, 318–322.
McNab, F., Varrone, A., Ausgeliefert, L., Jucaite, A., Bystritsky, P.,
Forssberg, H., et al. (2009). Changes in cortical dopamine
D1 receptor binding associated with cognitive training.
Wissenschaft, 323, 800–802.
Melby-Lervåg, M., & Hulme, C. (2013). Is working memory
training effective? A meta-analytic review. Developmental
Psychologie, 49, 270–291.
Metzler-Baddeley et al.
1519
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
.
F
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Melby-Lervag, M., & Hulme, C. (2016). There is no convincing
evidence that working memory training is effective: A reply
to Au et al. (2014) and Karbach and Verhaegen (2014).
Psychonomic Bulletin & Rezension, 23, 324–330.
Metzler-Baddeley, C., & Baddeley, R. (2009). Does adaptive
training work? Applied Cognitive Psychology, 23, 254–266.
Metzler-Baddeley, C., Caeyenberghs, K., Foley, S., & Jones, D. K.
(2016). Task complexity and location specific changes of
cortical thickness in executive and salience networks after
working memory training. Neurobild, 130, 48–62.
Metzler-Baddeley, C., O’Sullivan, M. J., Bells, S., Pasternak, O.,
& Jones, D. K. (2012). How and how not to correct for CSF-
contamination in diffusion MRI. Neurobild, 59, 1394–1403.
Morrison, A. B., & Chein, J. M. (2011). Does working memory
training work? The promise and challenges of enhancing
cognition by training working memory. Psychonomic
Bulletin & Rezension, 18, 46–60.
Mufson, E. J., & Mesulam, M. M. (1982). Insula of the old world
Affe. II: Afferent cortical input and comments on the
claustrum. Zeitschrift für Vergleichende Neurologie, 212, 23–37.
Olesen, P. J., Westerberg, H., & Klingberg, T. (2004). Increased
prefrontal and parietal activity after training of working
Erinnerung. Naturneurowissenschaften, 7, 75–79.
Owen, A. M., Hampshire, A., Grahn, J. A., Stenton, R., Dajani, S.,
Verbrennungen, A. S., et al. (2010). Putting brain training to the test.
Natur, 465, 775–778.
Pasternak, O., Sochen, N., Gur, Y., Intrator, N., & Assaf, Y.
(2009). Free water elimination and mapping from diffusion
MRT. Magnetic Resonance in Medicine, 62, 717–730.
Penke, L., Muñoz Maniega, S., Murray, C., Gow, A. J.,
Hernández, M. C., Clayden, J. D., et al. (2010). A general
factor of brain white matter integrity predicts information
processing speed in healthy older people. Zeitschrift für
Neurowissenschaften, 30, 7569–7574.
Pierpaoli, C., & Basser, P. J. (1996). Toward a quantitative
assessment of diffusion anisotropy. Magnetic Resonance in
Medicine, 36, 893–906.
Preacher, K. J., & MacCallum, R. C. (2002). Exploratory factor
analysis in behavior genetics research: Factor recovery with
small sample sizes. Behavior Genetics, 32, 153–161.
Rizzolatti, G., & Matelli, M. (2003). Two different streams form
the dorsal visual system: Anatomy and functions.
Experimentelle Hirnforschung, 153, 146–157.
Rose, N. S., Olsen, R. K., Craik, F. ICH., & Rosenbaum, R. S. (2012).
Working memory and amnesia: The role of stimulus novelty.
Neuropsychologie, 50, 11–18.
Sampaio-Baptista, C., Khrapitchev, A. A., Foxley, S., Schlagheck,
T., Scholz, J., Jbabdi, S., et al. (2013). Motor skill learning
induces changes in white matter microstructure and
myelination. Zeitschrift für Neurowissenschaften, 33, 19499–19503.
Scholz, J., Klein, M. C., Behrens, T. E., & Johansen-Berg, H.
(2009). Training induces changes in white-matter
architecture. Naturneurowissenschaften, 12, 1370–1371.
Shipstead, Z., Redick, T. S., & Engle, R. W. (2012). Is working
memory training effective? Psychological Bulletin, 138,
628–654.
Schmied, G. E., Housen, P., Yaffe, K., Ruff, R., Kennison, R. F.,
Mahncke, H. W., et al. (2009). A cognitive training program
based on principles of brain plasticity: Results from the
Improvement in Memory with Plasticity-based Adaptive
Cognitive Training (IMPACT) Studie. Journal of the American
Geriatrics Society, 57, 594–603.
Schmied, S. M., De Stefano, N., Jenkinson, M., & Matthews, P. M.
(2001). Normalized accurate measurement of longitudinal
brain change. Journal of Computer Assisted Tomography,
25, 466–475.
Song, S. K., Yoshino, J., Le, T. Q., Lin, S. J., Sun, S. W., Cross,
A. H., et al. (2005). Demyelination increases radial diffusivity
in corpus callosum of mouse brain. Neurobild, 26,
132–140.
Suo, C., León, ICH., Brodaty, H., Trollor, J., Wen, W., Sachdev, P.,
et al. (2012). Supervisory experience at work is linked to
low rate of hippocampal atrophy in late life. Neurobild, 63,
1542–1551.
Takeuchi, H., Sekiguchi, A., Solch, Y., Yokoyama, S., Yomogida,
Y., Komuro, N., et al. (2010). Training of working memory
impacts structural connectivity. Zeitschrift für Neurowissenschaften,
30, 3297–3303.
Takeuchi, H., Solch, Y., & Kawashima, R. (2010). Effects of
working memory training on cognitive functions and neural
Systeme. Reviews in the Neurosciences, 21, 427–449.
Takeuchi, H., Solch, Y., Sassa, Y., Hashizume, H., Sekiguchi, A.,
Fukushima, A., et al. (2011). Working memory training
using mental calculation impacts regional gray matter of
the frontal and parietal regions. PLoS One, 6, e23175.
Tavor, ICH., Hofstetter, S., & Assaf, Y. (2013). Micro-structural
assessment of short term plasticity dynamics. Neurobild,
81, 1–7.
Testa, R., Bennett, P., & Ponsford, J. (2012). Factor analysis of
nineteen executive function tests in a healthy adult population.
Archives of Clinical Neuropsychology, 27, 213–224.
Thiebaut de Schotten, M., Dell’Acqua, F., Forkel, S. J., Simmons,
A., Vergani, F., Murphy, D. G., et al. (2011). A lateralized
brain network for visuospatial attention. Naturneurowissenschaften,
14, 1245–1246.
Thomas, C., & Bäcker, C. ICH. (2013). Teaching and adult brain new
tricks: A critical review of evidence for training-dependent
structural plasticity in humans. Neurobild, 73, 225–236.
Tuch, D. S., Reese, T. G., Wiegell, M. R., Makris, N., Belliveau,
J. W., & Wedeen, V. J. (2002). High angular resolution
diffusion imaging reveals intravoxel white matter fiber
heterogeneity. Magnetic Resonance in Medicine, 48,
577–582.
Unsworth, N., Heitz, R. P., Schrock, J. C., & Engle, R. W. (2005).
An automated version of the operation span task. Behavior
Research Methods, 37, 498–505.
Valkanova, V., Rodriguez, R. E., & Ebmeier, K. P. (2013). Geist
over matter—What do we know about neuroplasticity in
Erwachsene? International Psychgeriatrics, 26, 891–909.
Wheeler-Kingshott, C. A., & Cercignani, M. (2009). About “axial”
and “radial” diffusivities. Magnetic Resonance in Medicine,
61, 1255–1260.
Zatorre, R. J., Fields, R. D., & Johansen-Berg, H. (2012).
Plasticity in gray and white: Neuroimaging changes in
brain structure during learning. Naturneurowissenschaften, 15,
528–536.
1520
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
9
2
9
1
/
5
9
0
/
9
1
1
5
9
0
5
9
2
/
8
1
9
7
5
8
Ö
6
C
5
N
8
_
5
A
/
_
J
0
Ö
1
C
1
N
2
7
_
A
P
_
D
0
1
B
1
j
2
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
T
.
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1