Distractors Selectively Modulate Electrophysiological
Markers of Perceptual Decisions
Shou-Han Zhou1, Gerard Loughnane2, Redmond O’Connell1,3,
Mark A. Bellgrove1*, and Trevor T.-J. Chong1*
Abstrakt
■ Current models of perceptual decision-making assume that
choices are made after evidence in favor of an alternative accu-
mulates to a given threshold. This process has recently been
revealed in human EEG recordings, but an unresolved issue is
how these neural mechanisms are modulated by competing,
yet task-irrelevant, Reize. In this study, we tested 20 healthy par-
ticipants on a motion direction discrimination task. Teilnehmer
monitored two patches of random dot motion simultaneously
presented on either side of fixation for periodic changes in an
upward or downward motion, which could occur equiprobably
in either patch. On a random 50% of trials, these periods of co-
herent vertical motion were accompanied by simultaneous task-
irrelevant, horizontal motion in the contralateral patch. Our data
showed that these distractors selectively increased the amplitude
of early target selection responses over scalp sites contralateral to
the distractor stimulus, without impacting on responses ipsilat-
eral to the distractor. Wichtig, this modulation mediated a
decrement in the subsequent buildup rate of a neural signature
of evidence accumulation and accounted for a slowing of RTs.
These data offer new insights into the functional interactions be-
tween target selection and evidence accumulation signals, Und
their susceptibility to task-irrelevant distractors. More broadly,
these data neurally inform future models of perceptual
decision-making by highlighting the influence of early process-
ing of competing stimuli on the accumulation of perceptual
evidence. ■
EINFÜHRUNG
Canonical models of perceptual decision-making propose
that decisions are formed after perceptual evidence in
favor of a given response accumulates to a given thresh-
alt (z.B., Ratcliff, Schmied, Braun, & McKoon, 2016).
Neurophysiological signatures of this evidence accumula-
tion process have been observed in several areas within
the rodent, Affe, and human brains (O’Connell,
Shadlen, Wong-Lin, & Kelly, 2018; Shadlen & Kiani, 2013;
Ding & Gold, 2011; Huk & Shadlen, 2005). Typically,
Jedoch, these processes are examined in the context of
perceptual decisions focused on a single stimulus. In con-
trast, perceptual decisions in daily life are often made in
the presence of numerous stimuli that compete for pro-
cessing resources and may impair the discrimination of
the primary target. Although an established literature on
selective attention and attentional capture has shown that
distractor stimuli can interfere with target processing
(Eimer, 2014; Luck, 2012; Forster & Lavie, 2011; Pashler,
1998; Posner & Petersen, 1990), an unanswered question
is how the neural mechanisms underlying perceptual
decision-making are modulated by competing, yet task-
irrelevant, distractors.
1Monash University, Australia, 2National College of Ireland,
3Trinity College Dublin
*These authors declare equal contribution.
© 2021 Massachusetts Institute of Technology
Sequential sampling models of evidence accumulation
have provided accurate accounts of perceptual decision-
making behavior (Ratcliff et al., 2016; Forstmann,
Wagenmakers, Eichele, Braun, & Serences, 2011), Und
neural signals exhibiting the predicted dynamics of evi-
dence accumulation have recently been isolated in humans
using EEG (Murphy, Boonstra, & Nieuwenhuis, 2016;
Philiastides, Heekeren, & Sajda, 2014; de Lange, Rahnev,
Donner, & Lau, 2013; O’Connell, Dockree, & Kelly, 2012).
Insbesondere, recent studies have established that the centro-
parietal positivity (CPP) builds at an evidence-dependent,
RT-predictive rate that peaks immediately before response
execution and exhibits amplitude modulations consistent
with boundary adjustments (Kelly, Corbett, & O’Connell,
2019; Steinemann, O’Connell, & Kelly, 2018; Newman,
Loughnane, Kelly, O’Connell, & Bellgrove, 2017; Kelly &
O’Connell, 2013; O’Connell et al., 2012). A notable feature
of the CPP is that its representation of cumulative evidence
is abstracted from the specific sensory or motor require-
ments of the task (Twomey, Kelly, & O’Connell, 2016;
O’Connell et al., 2012).
Although studies on perceptual decision-making have
focused on how evidence accumulation modulates proper-
ties of the CPP, much less is known about the role of earlier
target selection signals. Shortly after target onset, but be-
fore the onset of evidence accumulation, recent reports
have shown an early, negative deflection over posterior
Elektroden. This N2 response is not merely a sensory-
Zeitschrift für kognitive Neurowissenschaften 33:6, S. 1020–1031
https://doi.org/10.1162/jocn_a_01703
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
J
Ö
C
N
_
A
_
0
1
7
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
evoked response—rather, previous studies show that it is
elicited only by goal-relevant sensory changes (Newman
et al., 2017; Loughnane et al., 2016). The N2 exhibits distinct
components over both hemispheres. When target stimuli are
lateralized, there is an enhancement of the N2 contralateral
to the target (the N2c), which predicts RT via a relationship
with the onset and buildup rate of the CPP (Loughnane et al.,
2016). Zusätzlich, lateralized targets are also accompanied
by an N2 waveform in the ipsilateral hemisphere (the N2i)
—the N2i has been shown to be sensitive to the salience
of the target stimulus, but its relationship to behavior re-
mains obscure (Loughnane et al., 2016).
Wichtig, analogous N2 waveforms have been exten-
sively described in the literature on attentional capture
(Luck & Hillyard, 1994B). In diesem Kontext, the N2 is typically
probed with rapidly appearing, perceptually salient stimuli
(Luck et al., 2006; Luck & Hillyard, 1994A). Such experi-
mental designs have been useful in delineating the re-
sponse properties of N2 signals in visual search, showing
a positive relationship between search set size and N2 sig-
nal amplitude (Luck, 2012; Mazza, Turatto, & Caramazza,
2009). Jedoch, these studies were not optimized to iso-
late and measure the gradual accumulation of sensory ev-
idence that characterizes a perceptual decision, at least in
part because of the overlap between target selection sig-
nals and visual-evoked potentials triggered by abrupt stim-
ulus onsets. Außerdem, target processing in studies on
attentional capture is often focused on the difference
between contralateral and ipsilateral N2 waveforms (Die
classic “N2pc”). Such studies usually demonstrate an en-
hancement of the N2pc in the presence of nontarget stim-
uli in visual search tasks (Eimer, 1996). Jedoch, A
limitation of such an approach is that it may overlook later-
alisiert, hemisphere-specific contributions to target and
distractor processing (Schneider, Beste, & Wascher,
2012; Hickey, McDonald, & Theeuwes, 2006).
Gesamt, although previous studies have shown that the
N2pc is modulated by distractors, the mechanism underly-
ing this effect remains unclear. Traditional studies on atten-
tional capture are limited in their capacity to examine the
independent contributions of the ipsilateral and contra-
lateral N2 in the absence of overlapping visual-evoked po-
tentials and have not examined how distractor-mediated
effects on the N2 impact on subsequent evidence accumu-
lation. Hier, we examined the effects of distractor stimuli
on electrophysiological markers of human perceptual deci-
sionen. Participants discriminated the direction of coherent
dot motion in one hemifield, in the presence or absence
of task-irrelevant distractor motion in the contralateral
hemifield. We measured neural activity with EEG and asked
how distractor stimuli modulated critical components of
perceptual decisions, including the initial detection of the
target stimulus (as measured by the N2) and/or the onset
or rate of evidence accumulation (the slope of the CPP).
Notably in our task, transitions from random to coherent
motion occurred seamlessly, without any abrupt or net
changes in the energy of low-level stimulus features. Das
allowed us to dissect decision-related ERPs in the presence
of balanced displays of target and distractor stimuli, while
eliminating large sensory-evoked potentials that may have
otherwise masked or confounded these signals.
METHODEN
Teilnehmer
We tested 20 Teilnehmer (10 Frauen; Durchschnittsalter = 22.6 ±
4.8 Jahre, range = 19–32 years; all right-handed), mit
normal or corrected-to-normal vision and no history of neu-
rological or psychiatric disease. Participants were recruited
and tested at Monash University, Australia, in accordance
with an experimental protocol approved by Monash
University Human Research Ethics Committee.
Experimental Design
We examined perceptual decision-making in the context of
a prototypical motion direction discrimination task
(Loughnane et al., 2016; Britten, Shadlen, Newsome, &
Movshon, 1992; Newsome, Britten, & Movshon, 1989;
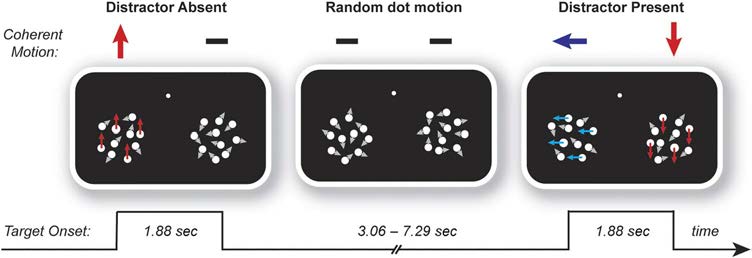
Figur 1. Task design.
Participants monitored two
patches of dots in the left and
right hemifields for intermittent
periods of coherent vertical
motion in one of those patches.
The task was to discriminate the
direction of motion (upward or
downward) of coherent vertical
motion whenever it occurred.
These target stimuli were
accompanied in the alternate
patch by either persistent
random dot motion (“distractor
absent”) or synchronous
periods of coherent motion in the orthogonal, horizontal plane (“distractor present”). Participants were instructed to provide fast, accurate responses
to the direction of vertical target motion (red arrows) and to ignore the presence of any horizontal distractor motion (blue arrows), which was
entirely task irrelevant. The location of targets (left/right), the presence of distractors (present/absent), the vertical direction of target motion
(up/down), and the horizontal direction of distractor motion when present (left/right) were equiprobable.
Zhou et al.
1021
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
J
Ö
C
N
_
A
_
0
1
7
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 1). Participants sat in a darkened room, 70 cm from a
27-In. LCD monitor (Dell S2716DG running at 120 Hz,
Auflösung: 1024 × 768). Responses were registered by
button press on a Cedrus Response Pad (Model RB-840),
and eye movements were monitored with an SR Research
Eyelink eye tracker (DM890, Eyelink 1000 Plus Version 5.09;
SR Research Ltd/SMI). The paradigm was implemented on
the Psychophysics Toolbox (Brainard & Spatial Vision,
1997) run in MATLAB (MathWorks, Inc.).
Participants fixated centrally while monitoring two
patches of random dot motion on either side of fixation.
On all trials, dots in one patch transitioned to coherent
vertical (d.h., upward or downward) Bewegung. Teilnehmer
were instructed to respond to the direction of target mo-
tion as quickly and accurately as possible with a left or right
index finger button press (with response mappings coun-
terbalanced across participants). The location (left vs.
right hemifield) and direction (upward vs. downward) von
target motion were equiprobable. On a random 50% of tri-
als, the target was presented alone, in which case random
dot motion persisted in the alternate patch (“distractor-
absent” trials). On the remaining 50% of trials, the target
motion was accompanied in the alternate patch by tempo-
rally coincident, horizontal distractor motion (leftward
or rightward; “distractor present”). The direction of hori-
zontal distractor motion was random, and participants
were informed that the horizontal distractor motion was
entirely uninformative and should be ignored.
Dot stimuli flickered at a rate of 30 Hz within the two
patches, which were centered at 10° on either side of fix-
ation and 4° below the horizontal meridian. Each patch
subtended 8° of visual angle and comprised 150 Weiß
dots (jede 6 × 6 pixels) against a black background.
Each trial consisted of a pretarget period of random
Bewegung, dauerhaft 3.06, 5.17, oder 7.29 Sek, during which dots
were positioned randomly throughout each patch on
each frame. During each trial, there was then a seamless
step change from random motion to coherent motion,
Wann 40% of the dots were randomly selected on each
frame to be displaced by 9° s−1 on the following frame
in the upward/downward direction (Und, in the case of
distractor-present trials, in the left/right direction in
the alternate patch). Periods of coherent motion lasted
1.88 Sek. Participants completed 16 experimental blocks
von 24 Versuche, and trials were presented continuously with-
out an intertrial interval. Trials that contained eye blinks
or breaks of fixation of >3° during the entire epoch were
repeated at the end of the block in random order.
EEG Acquisition and Preprocessing
Continuous EEG was acquired from 64 scalp electrodes
using a Brain Products BrainVision actiCHAMP system dig-
itized at 500 Hz. Data were processed using a combination
of custom scripts and EEGLAB routines implemented in
MATLAB (Delorme & Makeig, 2004). A 35-Hz fourth-order
Butterworth low-pass filter was applied offline; noisy
channels were interpolated using spherical spline, Und
the data were rereferenced to the average reference.
Epochs were extracted from the continuous data from
−700 to 1880 msec around the target onset (start of the
coherent motion) and were baselined with respect to
−100 to 0 msec before the target onset. To limit the
potential effects of volume conduction between adjacent
Elektroden, we transformed all signals using a current
source density transformation with the CSD Toolbox for
MATLAB (Kayser & Tenke, 2006).
We analyzed ERP data from the posterior N2 waveforms
(N2c, N2i) and the CPP. The N2c and N2i components
were measured contralateral and ipsilateral to the target
patch location, jeweils, from electrodes P7 and P8
(Newman et al., 2017; Loughnane et al., 2016). The CPP
was measured at peak electrode CPz (Steinemann et al.,
2018; Newman et al., 2017; Kelly & O’Connell, 2013).
The N2pc was computed as the difference between the
N2c and the N2i (N2c > N2i; Luck, 2012).
Statistical Analysis
Trials were excluded if (A) RTs were faster than 200 ms
(classified as a preemptive response) or slower than
1880 ms (d.h., after coherent motion offset; classified as a
missed response) oder (B) recordings from any channel ex-
ceeded ±100 μV from 100 msec before the target onset to
100 msec after the response. Behavioral performance was
quantified by fitting RT data to an ex-Gaussian distribution
using the retimes package in R v4.0.3 (R Foundation for
Statistical Computing, Vienna, Österreich). Accuracy data rep-
resent the percentage of correct trials for those trials made
within the acceptable response window. These measures,
as well as the ERP components, were compared between
the distractor-present and distractor-absent conditions
across participants using standard parametric statistics
(paired t tests, ANOVAs). In addition to these frequentist
Analysen, we performed the Bayesian equivalents using
the BayesFactor package in R v4.0.3. Bayesian analyses
specified a multivariate Cauchy prior on the effects, mit
a distribution centered around zero and a width parameter
von 0.707. Bayes factors were used to quantify evidence
in favor of each hypothesis and interpreted based on
Jeffreys (1961). We included subject as a random intercept
in all models. Daher, the null model for all comparisons was
a model including not only the grand mean but also sub-
ject as an additive factor. For the Bayesian equivalent
ANOVAs, we compared the null model against the full
model space of all simple effects and their interactions.
ERP analyses were performed on data from both correct
and incorrect trials. All waveforms were averaged for each
distractor condition. N2 peak amplitudes were defined as
the most negative amplitudes after target onset within
their characteristic time windows (N2c: 150–400 msec;
N2i: 150–450 msec; Newman et al., 2017; Loughnane
et al., 2016). In the case of the N2pc, many participants
had a discernible N2pc only for the distractor-absent,
1022
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 6
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
J
Ö
C
N
_
A
_
0
1
7
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
and not the distractor-present, condition. Daher, to statisti-
cally analyze the effect of distractors on the N2pc, we took
the N2pc peak amplitude in the distractor-absent condi-
tion for each participant and compared it against the
N2pc amplitude at that same time point for that partici-
pant in the distractor-present condition.
The CPP onsets for each participant were computed by
running sample-point-by-sample-point t tests of each
individual’s stimulus-locked CPP amplitudes against zero.
For each participant, we analyzed the distribution of
CPP amplitudes across all trials, within a 100-msec window
centered on each time point. The onset was defined as the
first time at which the distribution of CPP amplitudes was
significantly greater than zero, at a threshold of p = .05, für
15 or more consecutive samples from 100 Zu 400 ms
after stimulus onset (Newman et al., 2017; Loughnane
et al., 2016; O’Connell et al., 2012). The CPP slope was
defined as the gradient of a straight line, fit from 450 Zu
50 msec before the response-locked CPP waveform, Zu
analyze effects distal to the time interval for the N2 mea-
surements (Loughnane et al., 2016). The CPP amplitude
was measured as the mean amplitude of a 100-msec
window, from −50 to + 50 msec relative to the response.
Endlich, we performed a path analysis to determine whether
distractor-evoked changes in the N2 and/or CPP wave-
forms mediated participants’ behavior (Loughnane,
Newman, Tamang, Kelly, & O’Connell, 2018). The depen-
dent variable was the RT, and the predictor variable was
the absence or presence of a distractor (0 oder 1). Mediating
variables included the ERP components that were found
to be significantly affected by the presence of a distractor
in the aforementioned analyses (d.h., the N2i peak ampli-
tude and the CPP slope). These variables were individually
z scored across distractor condition. We then performed
a robust path analysis with the Yuan–Bentler method
and Huber–White standard errors (Ullman & Bentler,
2013). This analysis was implemented in R with the SEM
package.
ERGEBNISSE
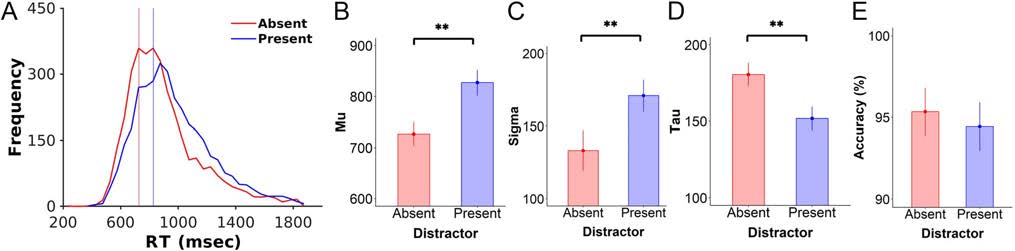
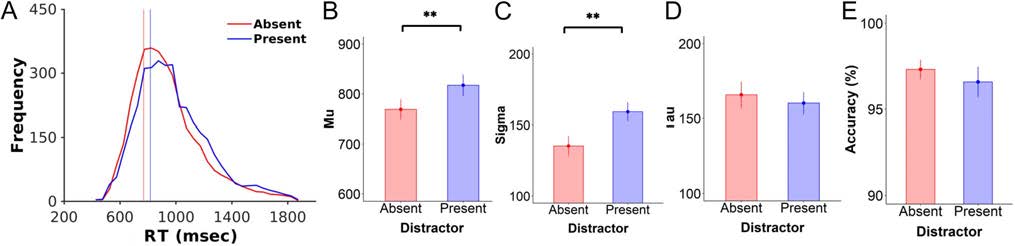
Behavioral Data
We examined the effect of distractors on behavioral perfor-
mance by fitting RT data to an ex-Gaussian distribution. Das
revealed a significant effect of distractors on all RT parame-
ters. Wie erwartet, der Mittelwert (M) of the RT distributions was
significantly greater when distractors were present versus
absent (present vs. absent: mean = 827 ± 25.0 vs. 727 ±
23.2 ms), T(19) = 8.48, P < .001, BF10 = 7.56 × 104
(decisive evidence). In addition, the presence of distractors
led to a higher standard deviation (σ) and a lower exponen-
tial decay parameter (τ) than the distractor-absent condition
(σ: present vs. absent, 171 ± 11.0 vs. 133 ± 14.0 msec; t(19) =
5.52, p < .001, BF10 = 493, decisive evidence; τ: present vs.
absent, 152 ± 7.72 vs. 181 ± 7.53 msec; t(19) = 4.44, p <
.001, BF10 = 94, very strong evidence). Response accuracy
was numerically lower when distractors were present,
but the effect did not reach statistical significance, with
Bayesian analyses indicating anecdotal evidence in favor
of a null effect (present vs. absent, 94.4 ± 1.48% vs. 95.3 ±
1.46%), t(19) = 1.67, p = .11, BF10 = 0.83 (Figure 2).
EEG Data
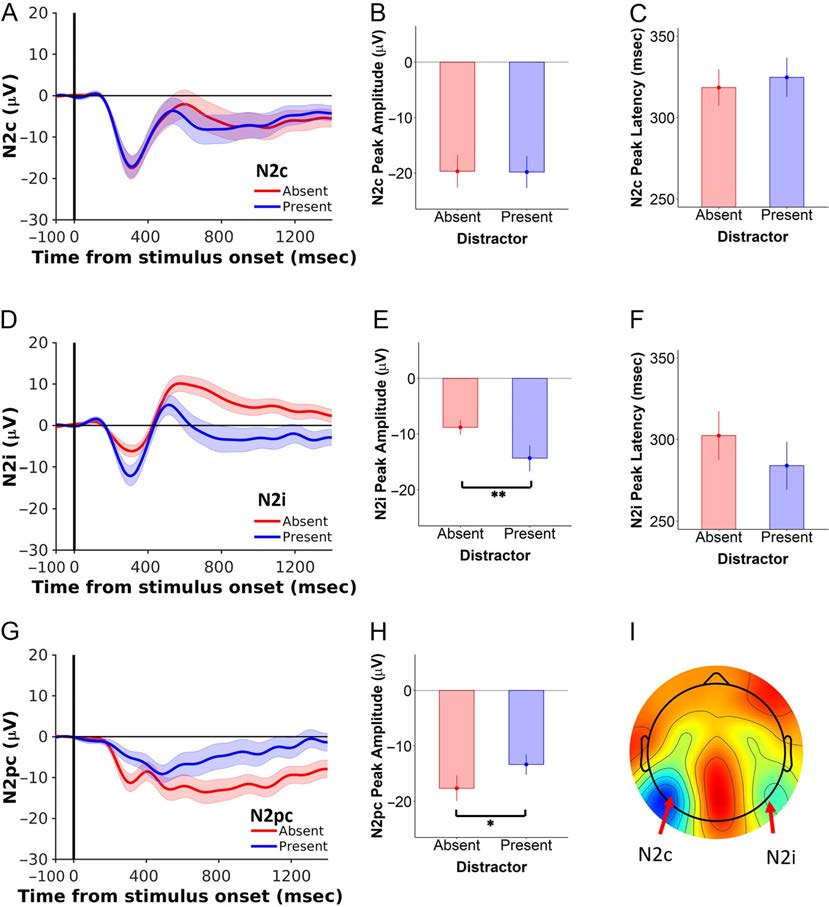
Distractors Selectively Increased the Peak Negativity of
the N2i but Not the N2c
First, we considered the effect of distractors on the peak
amplitudes of the N2c and N2i (Figure 3). Note that the
“contralateral” (N2c) and “ipsilateral” (N2i) waveforms
are assigned relative to the position of the target stimulus.
We performed a 2 × 2 repeated-measures ANOVA on the
factors of Distractor (present, absent) and N2 waveform
(N2c, N2i). This revealed a significant main effect of
Distractor and N2, which was qualified by a significant
Distractor × N2 interaction (Distractor: F(1, 19) = 11.9,
p = .003; N2: F(1, 19) = 17.8, p < .001; interaction: F(1,
19) = 15.1, p = .001). Importantly, decomposing this
Figure 2. RTs were slower in the presence versus absence of distractors. (A) RT distributions for the distractor-absent (red) and distractor-present
(blue) conditions. Vertical lines indicate mean RTs. (B–D) RT data were fit to an ex-Gaussian distribution to derive parameters for the (B) mean (μ),
(C) variance (σ), and (D) exponential decay (τ) of each condition. Data showed that distractors resulted in significantly longer RTs, with greater
variance and less decay than when no distractors were present. (E) Accuracy data represent the percentage of correct trials for those trials made
within the acceptable response window. There were no significant differences in accuracy as a function of distractor condition. Error bars indicate
1 SEM. **p < .001.
Zhou et al.
1023
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
j
o
c
n
_
a
_
0
1
7
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
j
o
c
n
_
a
_
0
1
7
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Distractors had a significant effect on the N2i but not the N2c. (A) The effect of distractors on the N2c waveform. ERPs are time-locked to
target onset. Distractor-absent data are in red, and distractor-present data are in blue. There were no effects of distractors on mean (B) N2c peak
amplitude or (C) N2c peak latency. (D) The effect of distractors on the N2i waveform. (E) Distractors led to more negative (greater amplitude) N2i
peak amplitudes, (F) but there were no effects on N2i peak latency. (G, H) Distractors suppressed the N2pc peak amplitude. (I) Topographic
distributions of the N2i and N2c, representing the average ERP between 200 and 400 msec after stimulus onset. The topographic plot is oriented to
depict targets appearing in the right visual hemifield. Note that the N2 waveforms are defined relative to the target hemifield (i.e., the N2c is
contralateral, and the N2i is ipsilateral, to the target stimulus). Thus, in our design, the N2i waveform comprises both the potential evoked by the
ipsilateral target and the “N2c-like” potential evoked by the contralateral distractor. Error bars indicate 1 SEM. *p < .05, **p < .001.
interaction with post hoc Bonferroni-corrected pairwise
comparisons revealed that N2i peak amplitudes were greater
(i.e., more negative) in the presence versus absence of dis-
tractors (present vs. absent, −14.06 ± 1.95 vs. −8.23 ±
1.68 μV; p < .001), but there was no such effect on the
N2c (present vs. absent, −19.3 ± 3.06 vs. −19.3 ± 2.71 μV;
p = 1.0).
The analogous Bayesian repeated-measures ANOVA re-
vealed a similar pattern of results, showing that the data
were best represented by a model that included both
simple effects (Distractor and N2 waveform) and their
interaction (posterior probability of .57; BF10 = 2.05 ×
105). We then performed post hoc comparisons for this
model (Bayesian t tests corrected for multiplicity). As in
the frequentist analyses, the adjusted posteriors demon-
strated very strong evidence for a difference in the N2i
waveforms as a result of distractors (BF10 = 96.9), together
with moderate evidence that the N2c waveforms were
unaffected (BF10 = 0.23).
There was no evidence of distractor modulation of the
peak latencies of the N2 waveforms. The analogous
Distractor × N2 waveform ANOVA did not reveal any
1024
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
j
o
c
n
_
a
_
0
1
7
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
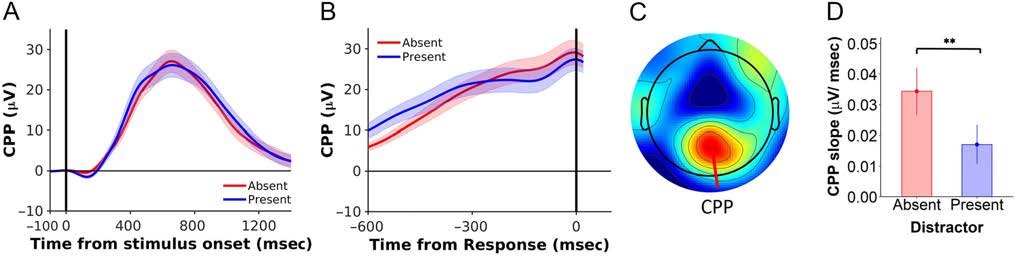
Figure 4. Distractors reduced the slope of the CPP. The CPP waveform (mean ± 1 SEM ), time-locked to (A) the stimulus onset and (B) the time of
response. (C) Topographic distribution of the CPP. (D) The CPP slope was significantly lower in the presence of distractors. Distractor-absent data are
in red, and distractor-present data are in blue. Error bars indicate 1 SEM. **p < .001.
significant effects of distractors on N2 peak latencies
(Distractor: F(1, 19) = 0.913, p = .35: N2, F(1, 19) =
2.39, p = .14; Distractor × N2: F(1, 19) = 3.47, p = .08).
Bayesian analyses showed that the data were best fit
by a model that only included the simple effect of N2
waveform, excluding the simple effect of Distractor or
its interaction with N2 waveform (posterior probability
of .60; BF10 = 3.67).
Prior studies on target discrimination in the context of
visual search have described an N2pc waveform (Luck,
2012; Eimer, 1996), which recent studies show can be
derived as a difference wave between the N2c and the
N2i (Loughnane et al., 2016). To interpret our findings in
the context of this work, we derived an N2pc difference
wave from our recordings (N2c > N2i) and performed a
paired t test on the effect of distractors on the N2pc peak
Amplitude (Figur 3). As expected from the lateralized N2
responses reported above, distractors led to a lower N2pc
peak amplitude than when no distractors were present
(present vs. absent, −12.7 ± 1.85 vs. −16.9 ± 2.35 μV),
T(19) = 10.2, p = .024, BF10 = 2.41. In the context of the
preceding analyses, this difference was clearly driven by
the distractor-sensitive N2i component.
Distractors Reduced the CPP Slope
Nächste, we considered the effects of distractors on the rate of
evidence accumulation, operationalized here as the slope
of the CPP (Figur 4). We measured the CPP slope in the
time window 50–450 msec before response, to analyze
effects distal to the time interval for the N2 measurements
(Loughnane et al., 2016). Wichtig, there was decisive
evidence that the CPP slope was shallower in the presence
versus absence of distractors (present vs. absent, 17.1 ×
10−3 ± 6.30 × 10−3 vs. 34.3 × 10−3 ± 7.77 × 10−3 μV
msec−1), T(19) = 5.07, P < .001, BF10 = 219. There was
no significant effect of distractors on stimulus-locked
CPP onsets, t(19) = 1.95, p = .07, BF10 = 1.73. There
was a borderline effect of distractors on reducing CPP
peak amplitudes (t(19) = 2.23, p = .04, BF10 = 3.34),
but this effect was not replicable in a follow-up experiment
(see Appendix).
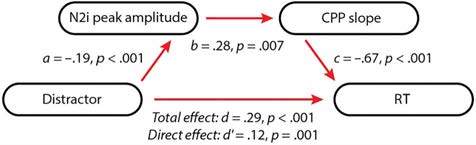
The Effects of Distractors on Behavior Were Significantly
Mediated by N2i Peak Amplitude and CPP Slope
To summarize, the preceding analyses demonstrated that
distractors significantly slowed RT and that this change in
performance was accompanied first by higher amplitude
N2i waveforms, followed by shallower response-locked
CPP slopes. To determine whether the N2i amplitudes
and CPP slopes were significant mediating variables for
the effects of distractors on behavior, we performed a
robust path analysis, with distractor as the predictor and
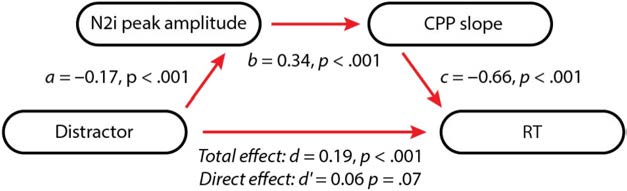
RT as the outcome (Figure 5). This analysis revealed signif-
icant path coefficients between successive variables. As ex-
pected, the relationship between distractors and the N2i
peak amplitude was significant in the negative direction,
indicating that distractors resulted in greater negative
N2i amplitudes (a = −0.19, p < .001). Critically, N2i peak
amplitudes had a positive relationship with the CPP slope,
indicating that higher amplitude (more negative) N2i am-
plitudes resulted in shallower CPP slopes (b = 0.28, p =
.007). Finally, the shallower the CPP slope, the longer
the RT (c = −0.67, p < .001). Overall, this analysis
revealed that N2i peak amplitude and CPP slope partially
mediated the effect of distractors on RT (total distractor–
RT effect, d = 0.29, p < .001; direct distractor–RT effect,
0 = 0.12, p = .001).
d
Figure 5. Path analysis showing that the effect of distractors on RT was
partially mediated by the N2i peak amplitude and the CPP slope.
Zhou et al.
1025
Results Were Replicated in a Separate Task Using
Reduced Dot Densities
Finally, we examined whether these effects generalize
across different viewing conditions. In a separate replication
study, we asked whether our results hold when both target
and distractor motion are more discriminable. Reducing the
dot density of a patch of coherent motion has been shown
to increase the discriminability of motion direction (Eagle
& Rogers, 1997; Ramachandran & Anstis, 1983). We
therefore performed a similar study to that described
here, but with reduced dot densities in both target and
distractor patches. This study involved a separate group
of 20 healthy individuals, and the data revealed a similar
pattern of results to those above. Details of this study are
provided in the Appendix.
DISCUSSION
Current frameworks broadly consider perceptual
decision-making to comprise several stages, from the ini-
tial representation of the sensory stimulus to the forma-
tion of a decision variable (O’ Connell et al., 2018;
Heekeren, Marrett, & Ungerleider, 2008). An unresolved
issue is how, and at which of these stages, distractors influ-
ence a perceptual decision. We addressed this question by
focusing on the rarely studied functional interactions be-
tween early target selection and evidence accumulation
processes. Our key findings were that (1) distractor stimuli
were accompanied by higher N2i waveforms (i.e., those
ipsilateral to the target), relative to when no distractors
were present; (2) these larger amplitude N2i waveforms
mediated downstream effects on the CPP slope; and (3)
together, these effects on the N2i and CPP mediated the
effects of distractors on slowing RTs. Together, these data
suggest that distractors slow perceptual decisions through
selective effects on distractor processing and on the rate
of evidence accumulation.
EEG markers of perceptual decision-making have typi-
cally been described in the absence of competing distrac-
tor stimuli (Kelly & O’Connell, 2013; O’Connell et al.,
2012). These tasks have reported an early N2c waveform,
the peak amplitude of which consistently mediates sensory
evidence accumulation and behavioral performance. This
has led to the proposal that the N2c is a signal reflect-
ing early target selection (van Kempen et al., 2019;
Newman et al., 2017; Loughnane et al., 2016). Our
data replicate these previous findings by demonstrating
a clear N2c waveform—in both the presence and absence
of distractors—the peak negativity of which occurred
∼300 msec after stimulus onset. In addition, however,
our data add to these earlier findings by showing that
the N2c waveforms are relatively insensitive to competing,
concurrently presented distractor stimuli.
In contrast, distractor stimuli had robust effects on the
N2i. Although the N2i has been consistently reported in
perceptual decision-making tasks (van Kempen et al.,
2019; Newman et al., 2017; Loughnane et al., 2016) and
is sensitive to properties of the target stimulus (e.g., its co-
herence), the N2i does not typically account for behavior
itself (e.g., RT; Loughnane et al., 2016). An outstanding
issue, therefore, has been the role of the N2i in gating
the information that feeds into later stages of decision-
making. The lateralized nature of our design was able to
shed light on this issue. Distractors, when they appeared,
were necessarily located in the hemifield contralateral
to the target itself. The fact that distractors specifically
enhanced the N2i, with a significantly less impact on the
N2c, suggests that the N2 components generated by each
hemisphere are likely to be independent.
What might this distractor-induced increase in N2i
peak amplitude represent? Behaviorally, we found that
distractors had a robust effect on the variance of the RT
distributions, indicating a likely effect of distractors on
a critical element of the decision-making process (e.g.,
drift rate), rather than merely a shift in nondecision
time, which would have simply served to increase the
mean, but not necessarily the variance, of the RT distribu-
tions. Furthermore, our key EEG result is that the
distractor-mediated increases in N2i peak amplitudes led
to a reduction in the rate of evidence accumulation as
measured by the slope of the CPP. Importantly, this im-
plies that higher amplitude N2i waveforms in fact related
to less efficient target processing.
One explanation is that the N2i may indirectly reflect
the resources consumed by an incompletely suppressed
distractor stimulus—resources that are then unable to
be assigned to enhance the processing of the target itself.
Indeed, the nature of our distractors was such that they
were events in a sensory stream that could potentially
contain a target but happened not to be a target by virtue
of its motion direction. Thus, it is conceivable that distrac-
tors first had to be strategically attended before being
dismissed. By this account, the N2i response could repre-
sent an “N2c-like” signal evoked by the distractor stimuli
and reflect the processing resources consumed by those
distractors. Such an account would be consistent with
perceptual load theories of attention, which propose that
distractor stimuli may involuntarily capture attentional
resources that are not currently allocated to target pro-
cessing (Chong, Cunnington, Williams, & Mattingley,
2009; Lavie, Hirst, De Fockert, & Viding, 2004).
This interpretation might also explain our finding that
distractors led to a suppression of the N2c > N2i differ-
ence wave, rather than the typical N2pc enhancement
associated with distractors in visual search paradigms
(Luck, 2005, 2012; Eimer, 1996). One key difference
between our paradigm and visual search tasks is that the
latter typically involve more perceptually salient distractor
stimuli with abrupt onsets, which might require greater
attentional suppression. Such suppression may result in
an attenuation of the N2i waveform, leading to a net higher
amplitude N2pc. This suppression may also result in a
separate signal related to distractor suppression, Die
1026
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 6
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
J
Ö
C
N
_
A
_
0
1
7
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
distractor positivity, as has been found in other salient
stimulus paradigms (Hickey, Di Lollo, & McDonald,
2009). Although we replicated the primary effects of our
study with stimulus patches of reduced dot density, Es
may be that the discriminability of even these stimuli
was lower than that of the unmasked, highly perceptually
salient stimuli typically used in visual search tasks. Das ist
suggested by the average RTs in our study (>720 msec),
which were considerably longer than the mean RTs in
typical visual search paradigms used to examine the N2pc
(z.B., ∼580 msec; Mazza et al., 2009). This speculation
would need to be confirmed by future work, aber die
overall conclusion from our data emphasizes the unique
importance of posterior N2 waveforms in distractor process-
ing and in mediating the subsequent accumulation of percep-
tual evidence.
Zusammenfassend, this study provides a first demonstration of
the selective effects of task-irrelevant distractors on the
neural mechanisms of evidence accumulation. Speziell,
our data show how early neural processes, as measured by
the N2 waveforms, are modulated by task-irrelevant dis-
tractor stimuli, and we provide evidence of the broader
role of the N2i in influencing downstream perceptual evi-
dence accumulation. Such findings invite future studies to
develop more refined computational models of percep-
tual processing—models that may be neurally informed by
electrophysiological parameters such as those described
Hier (O’Connell et al., 2018; Purcell et al., 2010).
Teilnehmer
We tested a separate group of 20 Teilnehmer (12 Frauen;
Durchschnittsalter = 25.05 ± 5.0 Jahre, range = 19–35 years; alle
right-handed), with normal or corrected-to-normal vision
and no history of neurological or psychiatric disease.
Experimental Design
The task was similar to that reported in the main text. Der
only difference was that each patch comprised reduced
dot densities of 10 dots per patch, instead of 150 im
main experiment. The analysis approach was identical to
the primary analyses reported in the main text.
Ergebnisse
RT data revealed the same pattern of effects as in the
primary task, with distractors resulting in slower and more
variable RTs relative to the distractor-absent condition (M:
present vs. absent, 817 ± 21.6 vs. 769 ± 20.2 ms, T(19) =
4.55, P < .001, BF10 = 87; σ: present vs. absent, 159 ± 6.55
vs. 135 ± 6.97 msec, t(19) = 4.79, p < .001, BF10 = 150).
However, the decay parameter was similar in both distrac-
tor conditions (τ: present vs. absent, 160 ± 7.34 vs. 166 ±
8.97 msec; t(19) = 0.92, p = .37, BF10 = 0.41). Accuracy
data again showed no evidence of a speed–accuracy
trade-off (present vs. absent, 96.6 ± 0.88% vs. 97.3 ±
0.57%; t(19) = 1.21, p = .24, BF10 = 0.53; Figure A1).
APPENDIX: REPLICATION EXPERIMENT WITH
REDUCED PATCH DENSITIES
EEG Data
In a separate replication study, we asked whether the
effects reported in the main text generalize across different
viewing conditions. This replication study was identical to
the main study, except that we increased the discrimina-
bility of target and distractor motion by reducing the dot
density of target and distractor patches (Eagle & Rogers,
1997; Ramachandran & Anstis, 1983). The results of this
task replicated the pattern of results from the main study.
Distractors Selectively Increased the Peak Negativity of
the N2i, but not the N2c
The corresponding Distractor (present, absent) × N2
Waveform (N2c, N2i) repeated-measures ANOVA revealed
significant main effects of Distractor and N2, which was qual-
ified by a significant Distractor × N2 interaction (Distractor:
F(1, 19) = 17.44, p < .001; N2: F(1, 19) = 11.12, p = .003;
interaction: F(1, 19) = 7.69, p = .012; Figure A2). This full
Figure A1. RTs were slower in the presence versus absence of distractors. (A) RT distributions for the distractor-absent (red) and distractor-present
(blue) conditions. Vertical lines indicate mean RTs. As in the primary study, distractors led to higher (B) mu and (C) sigma parameters. (D) However,
the tau parameter was no different between distractor conditions. (E) There was no significant different in accuracy between conditions. Error bars
indicate 1 SEM. **p < .001.
Zhou et al.
1027
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
j
o
c
n
_
a
_
0
1
7
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
model of simple effects and the interaction was also the best
fitting model in the analogous Bayesian repeated-measures
ANOVA (posterior probability = 0.62; BF10 = 3161).
Decomposing this interaction revealed that N2i peak
amplitudes were greater (i.e., more negative) in the
presence versus absence of distractors (present vs. absent,
−16.57 ± 3.99 vs. −7.69 ± 2.22 μV; p < .001, BF10 = 36.6).
However, there was no evidence for a similar effect on the
N2c (present vs. absent, −20.13 ± 3.94 vs. −18.34 ± 4.21 μV;
p = 1.0, BF10 = 0.66).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
j
o
c
n
_
a
_
0
1
7
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure A2. Distractors had a significant effect on the N2i, but not the N2c. (A) The effect of distractors on the N2c waveform. ERPs are time-locked to
target onset. Distractor-absent data are in red; and distractor-present data, in blue. There were no effects of distractors on mean (B) N2c peak
amplitude or (C) N2c peak latency. (D) The effect of distractors on the N2i waveform. (E) Distractors led to more negative (greater amplitude) N2i
peak amplitudes, (F) but there were no effects on N2i peak latency. (G, H) Distractors suppressed the N2pc peak amplitude. (I) Topographic
distributions of the N2i and N2c, representing the average ERP between 200 and 400 msec poststimulus onset. Error bars indicate 1 SEM. *p < .05,
**p < .001.
1028
Journal of Cognitive Neuroscience
Volume 33, Number 6
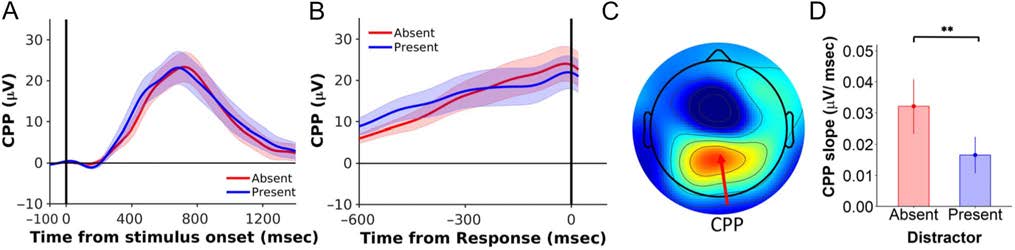
Figure A3. Distractors reduced the slope of the CPP. The CPP waveform (mean ± 1 SEM), time-locked to (A) the stimulus onset and (B) the time of
response. (C) Topographic distribution of the CPP. (D) The CPP slope was significantly lower in the presence of distractors. Distractor-absent data are
in red; and distractor-present data, in blue. Error bars indicate 1 SEM. **p < .001.
Figure A4. Path analysis
showing that the effect of
distractors on RT was fully
mediated by the N2i peak
amplitude and CPP slope.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
j
o
c
n
_
a
_
0
1
7
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
There was no evidence of distractor modulation of the
peak latencies of the N2 waveforms. The Distractor × N2
ANOVA did not reveal effects of distractors on N2 peak la-
tencies (Distractor: F(1, 19) = 1.65, p = .21; N2: F(1, 19) =
4.37, p = .05; Distractor × N2: F(1,19) = 0.003, p = .95).
This was reaffirmed by the Bayesian analyses, which
showed that the data were best fit by a model that included
only the simple effect of N2 waveform. Analyses of the
N2pc again suggested a lower N2pc peak amplitude when
distractors were absent versus present (present vs. absent,
−8.03 ± 2.17 vs. −13.3 ± 2.71 μV; t(19) = 2.21, p = .04,
BF10 = 1.83).
Distractors Reduced the CPP Slope
As in the main study, distractors resulted in a significantly shal-
lower response-locked CPP slope relative to the distractor-
absent condition (present vs. absent, 16.6 × 10−3 ± 5.86 ×
10−3 vs. 32.1 × 10−3 ± 8.74 × 10−3 μV/msec; t(19) = 3.40,
p = .003, BF10 = 11; strong evidence for distractor effect;
Figure A3). There was again no significant evidence that
distractors had an effect on CPP onsets, t(19) = 1.45, p =
.16, BF10 = 0.68, and there was no significant evidence that
distractors affected the CPP peak amplitudes, t(19) = 1.84,
p = .08, BF10 = 1.03.
The Effects of Distractors on Behavior Were Significantly
Mediated by N2i Peak Amplitude and CPP Slope
The analogous path analysis to the main study revealed a
similar pattern of effects, but with a more complete
mediation effect of the N2i peak amplitude and CPP slope
on RT (Figure A4). There were significant relationships be-
tween distractors and the N2i peak amplitude (a = −0.17,
p < .001), N2i peak amplitudes and the CPP slope (b =
0.34, p < .001), and CPP slope and RT (c = −0.66, p <
.001). Overall, this analysis revealed that N2i peak ampli-
tude and CPP slope fully mediated the effect of distractors
on RT (total distractor–RT effect: d = 0.19, p < .001; direct
distractor–RT effect: d′ = 0.06, p = .07).
Reprint requests should be sent to Shou-Han Zhou or Trevor T.-J.
Chong, Turner Institute for Brain and Mental Health, Monash
University, VIC 3800, Australia, or via e-mail: shou-han.zhou
@monash.edu or trevor.chong@monash.edu.
Author Contributions
Shou-Han Zhou: Data curation; Formal analysis; Investi-
gation; Methodology; Project administration; Validation;
Visualization; Writing – review & editing. Gerard
Loughnane: Conceptualization; Formal analysis; Investi-
gation; Methodology; Validation; Visualization; Writing –
review & editing. Redmond O’Connell: Conceptualization;
Formal analysis; Investigation; Methodology; Validation;
Visualization; Writing – review & editing. Mark A.
Bellgrove: Conceptualization; Funding acquisition; Inves-
tigation; Methodology; Project administration; Resources;
Software; Supervision; Validation; Writing – original draft;
Writing – review & editing. Trevor T.-J. Chong: Concep-
tualization; Data curation; Formal analysis; Funding
acquisition; Investigation; Methodology; Project admin-
istration; Resources; Software; Supervision; Validation;
Zhou et al.
1029
Visualization; Writing – original draft; Writing – review &
editing.
Funding Information
Redmond O’Connell: H2020 European Research Council
(http://dx.doi.org/10.13039/100010663), grant number:
63829; Office of Naval Research Global (http://dx.doi
.org/10.13039/100007297). Mark A. Bellgrove: Australian
R e s e a r c h C o u n c i l ( h t t p : / / d x . d o i . o r g / 1 0 . 1 3 0 3 9
/501100000923), grant number: DP180102066, National
Health and Medical Research Council of Australia
(http://dx.doi.org/10.13039/501100000925); Office of
Naval Research Global (http://dx.doi.org/10.13039
/100007297). Trevor T.-J. Chong: Australian Research
Council (http://dx.doi.org/10.13039/501100000923),
grant numbers: DP 180102383 and DE 180100389, the
Judith Jane Mason and Harold Stannett Williams
Memorial Foundation (http://dx.doi.org/10.13039
/100008718), the Brain Foundation (http://dx.doi.org
/10.13039/501100000942), the Rebecca L. Cooper
Medical Research Foundation (http://dx.doi.org
/10.13039/501100001061), and the Society for Mental
H e a l t h R e s e a r c h ( h t t p : / / d x . d o i . o r g / 1 0 . 1 3 0 3 9
/501100006624), Office of Naval Research Global
(http://dx.doi.org/10.13039/100007297).
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by estimated
gender identification of first author/last author) pub-
lishing in the Journal of Cognitive Neuroscience ( JoCN)
during this period were M(an)/M = .408, W(oman)/M =
.335, M/W = .108, and W/W = .149, the comparable pro-
portions for the articles that these authorship teams cited
were M/M = .579, W/M = .243, M/W = .102, and W/W =
.076 (Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently,
JoCN encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender cita-
tion balance.
REFERENCES
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision,
10, 433–436. DOI: https://doi.org/10.1163/156856897X00357,
PMID: 9176952
Britten, K. H., Shadlen, M. N., Newsome, W. T., & Movshon, J. A.
(1992). The analysis of visual motion: A comparison of
neuronal and psychophysical performance. Journal of
Neuroscience, 12, 4745–4765. DOI: https://doi.org/10.1523
/JNEUROSCI.12-12-04745.1992, PMID: 1464765, PMCID:
PMC6575768
Chong, T. T.-J., Cunnington, R., Williams, M. A., & Mattingley, J. B.
(2009). The role of selective attention in matching observed
and executed actions. Neuropsychologia, 47, 786–795. DOI:
https://doi.org/10.1016/j.neuropsychologia.2008.12.008,
PMID: 19124033
de Lange, F. P., Rahnev, D. A., Donner, T. H., & Lau, H. (2013).
Prestimulus oscillatory activity over motor cortex reflects
perceptual expectations. Journal of Neuroscience, 33,
1400–1410. DOI: https://doi.org/10.1523/JNEUROSCI.1094
-12.2013, PMID: 23345216, PMCID: PMC6618755
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. DOI: https://doi.org/10.1016/j.jneumeth
.2003.10.009, PMID: 15102499
Ding, L., & Gold, J. I. (2011). Neural correlates of perceptual
decision making before, during, and after decision commitment
in monkey frontal eye field. Cerebral Cortex, 22, 1052–1067.
DOI: https://doi.org/10.1093/cercor/bhr178, PMID: 21765183,
PMCID: PMC3328342
Eagle, R. A., & Rogers, B. J. (1997). Effects of dot density, patch
size and contrast on the upper spatial limit for direction
discrimination in random-dot kinematograms. Vision
Research, 37, 2091–2102. DOI: https://doi.org/10.1016
/S0042-6989(96)00153-8
Eimer, M. (1996). The N2pc component as an indicator of
attentional selectivity. Electroencephalography and Clinical
Neurophysiology, 99, 225–234. DOI: https://doi.org/10.1016
/0013-4694(96)95711-9
Eimer, M. (2014). The time course of spatial attention: Insights from
event related brain potentials. In A. Nobre & S. Kastner (Eds.),
The Oxford handbook of attention (1st ed., pp. 289–317).
Oxford: Oxford University Press. DOI: https://doi.org/10
.1093/oxfordhb/9780199675111.013.006
Forster, S., & Lavie, N. (2011). Entirely irrelevant distractors can
capture and captivate attention. Psychonomic Bulletin &
Review, 18, 1064–1070. DOI: https://doi.org/10.3758/s13423
-011-0172-z, PMID: 21989770, PMCID: PMC3219870
Forstmann, B. U., Wagenmakers, E. J., Eichele, T., Brown, S., &
Serences, J. T. (2011). Reciprocal relations between cognitive
neuroscience and formal cognitive models: Opposites attract?
Trends in Cognitive Sciences, 15, 272–279. DOI: https://doi
.org/10.1016/j.tics.2011.04.002, PMID: 21612972, PMCID:
PMC3384559
Heekeren, H. R., Marrett, S., & Ungerleider, L. G. (2008). The
neural systems that mediate human perceptual decision making.
Nature Reviews Neuroscience, 9, 467. DOI: https://doi.org
/10.1038/nrn2374, PMID: 18464792
Hickey, C., Di Lollo, V., & McDonald, J. J. (2009). Electrophysiological
indices of target and distractor processing in visual search.
Journal of Cognitive Neuroscience, 21, 760–775. DOI:
https://doi.org/10.1162/jocn.2009.21039, PMID: 18564048
Hickey, C., McDonald, J. J., & Theeuwes, J. (2006).
Electrophysiological evidence of the capture of visual
attention. Journal of Cognitive Neuroscience, 18, 604–613.
DOI: https://doi.org/10.1162/jocn.2006.18.4.604, PMID:
16768363
Huk, A. C., & Shadlen, M. N. (2005). Neural activity in macaque
parietal cortex reflects temporal integration of visual motion
signals during perceptual decision making. Journal of
Neuroscience, 25, 10420–10436. DOI: https://doi.org/10
.1523/JNEUROSCI.4684-04.2005, PMID: 16280581, PMCID:
PMC6725829
Jeffreys, H. (1961). Theory of probability (3rd ed.). Oxford:
Oxford University Press.
Kayser, J., & Tenke, C. E. (2006). Principal components analysis
of laplacian waveforms as a generic method for identifying
erp generator patterns: I. evaluation with auditory oddball
tasks. Clinical Neurophysiology, 117, 348–368. DOI: https://
doi.org/10.1016/j.clinph.2005.08.034, PMID: 16356767
1030
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
j
o
c
n
_
a
_
0
1
7
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Kelly, S. P., Corbett, E. A., & O’Connell, R. G. (2020).
Neurocomputational mechanisms of prior-informed perceptual
decision-making in humans. Nature Human Behaviour, 5,
467–481 DOI: https://doi.org/10.1038/s41562-020-00967-9
Kelly, S. P., & O’Connell, R. G. (2013). Internal and external
influences on the rate of sensory evidence accumulation in
the human brain. Journal of Neuroscience, 33, 19434–19441.
DOI: https://doi.org/10.1523/JNEUROSCI.3355-13.2013,
PMID: 24336710, PMCID: PMC6618757
Lavie, N., Hirst, A., De Fockert, J. W., & Viding, E. (2004). Load
theory of selective attention and cognitive control. Journal
of Experimental Psychology: General, 133, 339. DOI:
https://doi.org/10.1037/0096-3445.133.3.339, PMID:
15355143
Loughnane, G. M., Newman, D. P., Bellgrove, M. A., Lalor, E. C.,
Kelly, S. P., & O’Connell, R. G. (2016). Target selection signals
influence perceptual decisions by modulating the onset and
rate of evidence accumulation. Current Biology, 26, 496–502.
DOI: https://doi.org/10.1016/j.cub.2015.12.049, PMID:
26853360
Loughnane, G. M., Newman, D. P., Tamang, S., Kelly, S. P., &
O’Connell, R. G. (2018). Antagonistic interactions between
microsaccades and evidence accumulation processes during
decision formation. Journal of Neuroscience, 38, 2163–2176.
DOI: https://doi.org/10.1523/JNEUROSCI.2340-17.2018,
PMID: 29371320, PMCID: PMC6596275
Luck, S. J. (2005). The operation of attention—Millisecond by
millisecond—Over the first half second. In H. Ogmen & B. G.
Breitmeyer (Eds.), The first half second: The microgenesis
and temporal dynamics of unconscious and conscious
visual processes. Cambridge, MA: MIT Press.
Luck, S. J. (2012). Electrophysiological correlates of the focusing of
attention within complex visual scenes: N2pc and related ERP
components. In S. J. Luck & E. S. Kappenman (Eds.), Oxford
handbook of ERP components. New York: Oxford University
Press. DOI: https://doi.org/10.1093/oxfordhb/9780195374148
.013.0161
Luck, S. J., Fuller, R. L., Braun, E. L., Robinson, B., Summerfelt,
A., & Gold, J. M. (2006). The speed of visual attention in
schizophrenia: Electrophysiological and behavioral evidence.
Schizophrenia Research, 85, 174–195. DOI: https://doi.org
/10.1016/j.schres.2006.03.040, PMID: 16713184
Luck, S. J., & Hillyard, S. A. (1994a). Electrophysiological correlates
of feature analysis during visual search. Psychophysiology,
31, 291–308. DOI: https://doi.org/10.1111/j.1469-8986.1994
.tb02218.x, PMID: 8008793
Luck, S. J., & Hillyard, S. A. (1994b). Spatial-filtering during
visual-search: Evidence from human electrophysiology.
Journal of Experimental Psychology: Human Perception
and Performance, 20, 1000–1014. DOI: https://doi.org
/10.1037/0096-1523.20.5.1000
Mazza, V., Turatto, M., & Caramazza, A. (2009). Attention selection,
distractor suppression and N2pc. Cortex, 45, 879–890. DOI:
https://doi.org/10.1016/j.cortex.2008.10.009, PMID: 19084218
Murphy, P. R., Boonstra, E., & Nieuwenhuis, S. (2016). Global
gain modulation generates time-dependent urgency during
perceptual choice in humans. Nature Communications, 7,
13526. DOI: https://doi.org/10.1038/ncomms13526, PMID:
27882927, PMCID: PMC5123079
Newman, D. P., Loughnane, G. M., Kelly, S. P., O’Connell, R. G.,
& Bellgrove, M. A. (2017). Visuospatial asymmetries arise
from differences in the onset time of perceptual evidence
accumulation. Journal of Neuroscience, 37, 3378–3385.
DOI: https://doi.org/10.1523/JNEUROSCI.3512-16.2017,
PMID: 28242798, PMCID: PMC6596774
Newsome, W. T., Britten, K. H., & Movshon, J. A. (1989). Neuronal
correlates of a perceptual decision. Nature, 341, 52–54. DOI:
https://doi.org/10.1038/341052a0, PMID: 2770878
O’Connell, R. G., Dockree, P. M., & Kelly, S. P. (2012). A
supramodal accumulation-to-bound signal that determines
perceptual decisions in humans. Nature Neuroscience,
15, 1729. DOI: https://doi.org/10.1038/nn.3248, PMID:
23103963
O’Connell, R. G., Shadlen, M. N., Wong-Lin, K., & Kelly, S. P.
(2018). Bridging neural and computational viewpoints on
perceptual decision-making. Trends in Neurosciences, 41,
838–852. DOI: https://doi.org/10.1016/j.tins.2018.06.005,
PMID: 30007746, PMCID: PMC6215147
Pashler, H. (1998). Attention. London: Psychology Press.
Philiastides, M. G., Heekeren, H. R., & Sajda, P. (2014). Human
scalp potentials reflect a mixture of decision-related signals
during perceptual choices. Journal of Neuroscience, 34,
16877–16889. DOI: https://doi.org/10.1523/JNEUROSCI
.3012-14.2014, PMID: 25505339, PMCID: PMC4261107
Posner, M. I., & Petersen, S. E. (1990). The attention system of
the human brain. Annual Review of Neuroscience, 13,
25–42. DOI: https://doi.org/10.1146/annurev.ne.13.030190
.000325, PMID: 2183676
Purcell, B. A., Heitz, R. P., Cohen, J. Y., Schall, J. D., Logan, G. D.,
& Palmeri, T. J. (2010). Neurally constrained modeling of
perceptual decision making. Psychological Review, 117,
1113–1143. DOI: https://doi.org/10.1037/a0020311, PMID:
20822291, PMCID: PMC2979343
Ramachandran, V. S., & Anstis, S. M. (1983). Displacement
thresholds for coherent apparent motion in random-dot
patterns. Vision Research, 23, 1719–1724. DOI: https://doi
.org/10.1016/0042-6989(83)90188-8
Ratcliff, R., Smith, P. L., Brown, S. D., & McKoon, G. (2016).
Diffusion decision model: Current issues and history. Trends in
Cognitive Sciences, 20, 260–281. DOI: https://doi.org/10.1016/j
.tics.2016.01.007, PMID: 26952739, PMCID: PMC4928591
Schneider, D., Beste, C., & Wascher, E. (2012). Attentional
capture by irrelevant transients leads to perceptual errors in
a competitive change detection task. Frontiers in Psychology,
3, 164. DOI: https://doi.org/10.3389/fpsyg.2012.00164, PMID:
22654780, PMCID: PMC3360465
Shadlen, M. N., & Kiani, R. (2013). Decision making as a window
on cognition. Neuron, 80, 791–806. DOI: https://doi.org
/10.1016/j.neuron.2013.10.047, PMID: 24183028, PMCID:
PMC3852636
Steinemann, N. A., O’Connell, R. G., & Kelly, S. P. (2018).
Decisions are expedited through multiple neural adjustments
spanning the sensorimotor hierarchy. Nature Communications,
9, 3627. DOI: https://doi.org/10.1038/s41467-018-06117-0,
PMID: 30194305, PMCID: PMC6128824
Twomey, D. M., Kelly, S. P., & O’Connell, R. G. (2016). Abstract
and effector-selective decision signals exhibit qualitatively
distinct dynamics before delayed perceptual reports. Journal
of Neuroscience, 36, 7346–7352. DOI: https://doi.org/10
.1523/JNEUROSCI.4162-15.2016, PMID: 27413146, PMCID:
PMC4945659
Ullman, J. B., & Bentler, P. M. (2013). Structural equation modeling.
In J. A. Schinka, W. F. Velicer, & I. B. Weiner (Eds.), Handbook
of psychology: Research methods in psychology (2nd ed.,
pp. 661–690). Hoboken, NJ: John Wiley & Sons Inc.
van Kempen, J., Loughnane, G. M., Newman, D. P., Kelly, S. P.,
Thiele, A., O’Connell, R. G., et al. (2019). Behavioural and
neural signatures of perceptual decision-making are modulated
by pupil-linked arousal. eLife, 8, e42541. DOI: https://doi.org
/10.7554/elife.42541
Zhou et al.
1031
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
6
1
0
2
0
1
9
1
3
6
4
7
/
/
j
o
c
n
_
a
_
0
1
7
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3