Corticostriatal Contributions to Musical
Expectancy Perception
Carol A. Seger, Brian J. Spiering*, Anastasia G. Sares
Sarah I. Quraini, Catherine Alpeter, James David,
and Michael H. Thaut
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
Abstrakt
■ This study investigates the functional neuroanatomy of
harmonic music perception with fMRI. We presented short
pieces of Western classical music to nonmusicians. The ending
of each piece was systematically manipulated in the following
four ways: Standard Cadence (expected resolution), Deceptive
Cadence (moderate deviation from expectation), Modulated
Cadence (strong deviation from expectation but remaining
within the harmonic structure of Western tonal music), Und
Atonal Cadence (strongest deviation from expectation by leav-
ing the harmonic structure of Western tonal music). Music com-
pared with baseline broadly recruited regions of the bilateral
superior temporal gyrus (STG) and the right inferior frontal
gyrus (IFG). Parametric regressors scaled to the degree of de-
viation from harmonic expectancy identified regions sensitive
to expectancy violation. Areas within the BG were significantly
modulated by expectancy violation, indicating a previously un-
appreciated role in harmonic processing. Expectancy violation
also recruited bilateral cortical regions in the IFG and anterior
STG, previously associated with syntactic processing in other
domains. The posterior STG was not significantly modulated
by expectancy. Granger causality mapping found functional
connectivity between IFG, anterior STG, posterior STG, Und
the BG during music perception. Our results imply the IFG,
anterior STG, and the BG are recruited for higher-order har-
monic processing, whereas the posterior STG is recruited for
basic pitch and melodic processing. ■
EINFÜHRUNG
Music follows a complex syntactic structure. Multiple ele-
gen (z.B., rhythm, melody, and harmony) are ordered
by rules governing the sequential (horizontal) and simul-
taneous (vertical) organization of sound patterns. Diese
syntactic rules are implicitly acquired during develop-
ment via environmental exposure (Pearce, Ruiz, Kapasi,
Wiggins, & Bhattacharya, 2009) and differ between mu-
sical systems; Western tonal music, one such musical sys-
tem, underlies the forms of classical and popular music
originating from Europe. We investigated harmonic pro-
cessing by systematically manipulating musical pieces to
follow or violate Western tonal conventions. These mu-
sical pieces were presented to musically untrained young
adults raised listening to Western tonal music. Our pri-
mary goal was to examine BG involvement in processing
harmonic expectancies. We predicted a role for the BG in
harmonic processing based on functional neuroanatomy,
computational considerations, and research in other cog-
nitive domains demonstrating the BGʼs involvement in
processing expectancies. Our second goal was to identify
Colorado State University
*Present address: Abteilung für Psychologie, The University of
Maryland.
© 2013 Massachusetts Institute of Technology
cortical regions involved in harmonic processing, specifi-
cally within the inferior frontal and superior temporal
lobes. Our third goal was to examine how these cortical
regions functionally interact with each other and with the
BG during music perception using Granger causality
mapping (GCM).
The role of the BG in harmonic processing has not
been studied, although BG recruitment has been reported
in some previous studies (z.B., Kölsch, Fritz, & Schlaug,
2008). Research focusing specifically on the BG and music
has more commonly explored rhythm and tempo pro-
Abschließen (Schwartze, Keller, Patel, & Kotz, 2011; Grahn &
Rowe, 2009; Chen, Penhune, & Zatorre, 2008). Jedoch,
there are a number of theoretical reasons to predict that
the BG may play an important role in harmonic processing
based on their functional neuroanatomy and findings from
research in other cognitive domains. Neuroanatomically,
the BG are a subcortical structure that interacts with cere-
bral cortex via recurrent corticostriatal “loops.” The BG
modulate cortex activity via separate pathways (usually re-
ferred to as direct, indirect, and hyperdirect) by affecting
inhibition and excitation of cortical representations, daher
allowing for selection or gating of specific representations,
inhibition of alternative representations, and switching
between representations (for more detailed reviews, sehen
Braunlich & Seger, 2013; Seger, 2008). The BG are able
Zeitschrift für kognitive Neurowissenschaften 25:7, S. 1062–1077
doi:10.1162/jocn_a_00371
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
to learn from experience which representations should be
selected or inhibited via a dopaminergic feedback, a reward
signal from the midbrain, that affects the degree to which
corticostriatal synapses are strengthened (long-term po-
tentiation) or weakened (long-term depression; Tritsch &
Sabatini, 2012). The dopamine reward signal indicates
unexpected reward or omission of expected reward
(Bromberg-Martin, Matsumoto, & Hikosaka, 2010), welche
creates the potential conditions for learning. Reinforcement
learning methods developed in the computer science field
offer formal methods to characterize and quantify both
measures of expectation (d.h., reward prediction) und vio-
lations of expectation (d.h., reward prediction error;
Glimcher, 2011; Sutton & Barto, 1998). As a primary target
of dopaminergic projections, the BG play an important role
in coding these reward predictions and reward prediction
Fehler. There are regional differences within the striatum.
The dorsal striatum shows relatively greater activity for
reward prediction, whereas the ventral striatum shows
the strongest reward prediction error activity (Lee, Seo,
& Jung, 2012; Seger, Peterson, Cincotta, Lopez-Paniagua,
& Anderson, 2010).
We predicted the BG would be involved in harmonic
Verarbeitung, given the strong parallels between the BGʼs
role in forming predictions and assessing prediction error
and the similar functions inherent in musical harmony.
One fundamental element of music is the manipulation
of harmonic expectancy and violation of expectancy
across sequences of musical events within a specific tonal
System. Expectations are manipulated to create patterns
of tension (when violated) and resolution (when the ex-
pected occurs). For example in the Western tonal system,
moving away from a tonic chord develops tension but re-
turning to the tonic creates release (Huron, 2006; Meyer,
1989). This idea led to the prediction that the BG would
have a role in processing predictions and violation of pre-
dictions in music, similar to their role in other cognitive
domains (Kranjec, Cardillo, Schmidt, Lehet, & Chatterjee,
2012; Zacks, Kurby, Eisenberg, & Haroutunian, 2011;
Bahlmann, Schubotz, Mueller, Koester, & Friederici,
2009; Lohrenz, McCabe, Camerer, & Montague, 2007).
Außerdem, theories of music learning argue tonal sys-
tems are primarily learned through experience (Krumhansl
& Cuddy, 2010; Pearce et al., 2009; Tillman, 2008), welches ist
consistent with the plasticity of the BG.
Much of the research establishing the role of the stria-
tum in reward prediction and prediction error was per-
formed using classical or instrumental conditioning tasks
and their homologs (for review of these tasks, see Seger,
2009). These tasks typically use primary (z.B., food or
Wasser) or secondary (z.B., money or explicit verbal feed-
back) rewards. Jedoch, recent research has found that
explicit external rewards are not necessary for prediction
and prediction error processing: Similar activity occurs
when expectations are formed and confirmed or violated
even if the expected situation is not a primary or secondary
reward. Zum Beispiel, in sequence processing, reward pre-
diction error occurs when an unexpected item (z.B., a let-
ter in an artificial grammar learning experiment) occurs
(Bahlmann et al., 2009). This type of sequence processing
is similar to harmonic processing in music; harmonic fea-
tures are not primary or secondary rewards, yet they are
subject to expectancy and violation of expectancy. Empiri-
cal studies have demonstrated that the BG are critical for
developing expectations and responding to violations of
those expectations across numerous additional domains,
including social cognition (Schiffer & Schubotz, 2011),
causal reasoning (Kranjec et al., 2012), counterfactual
“might-have been” reasoning (Lohrenz et al., 2007), Und
event perception (Zacks et al., 2011).
The striatum is not a unitary structure: BG and cortex
interact with each other within several dissociable cortico-
striatal “loops” (Haber, 2003; Alexander, DeLong, & Strick,
1986) that connect different regions of the cortex to
different regions of the BG. We predict the BG will perform
similar processes of prediction and prediction error pro-
cessing within these loops, but the specific types of pre-
dictions will depend on the type of the representations
within each respective cortical regions within a given loop.
Although the corticostriatal system has no distinct borders
separating loops, it is useful for heuristic purposes to
conceptualize the system as containing at least four pri-
mary loops connecting striatum and cortex (Seger, 2008;
Lawrence, Sahakian, & Robbins, 1998). The “motor loop”
connects the putamen with motor and premotor regions
of the frontal and parietal lobes. The “executive loop” con-
nects the head of the caudate with lateral prefrontal and
inferior parietal cortex. The “visual loop” connects poste-
rior regions of the body and tail of the caudate with regions
of extrastriate occipital and temporal lobes. The “motiva-
tional” loop connects the ventral striatum with OFC and
other regions mediating reward and emotion. As described
below, we predicted that harmonic expectancy might mod-
ulate activity in two important cortical regions: the inferior
frontaler Gyrus (IFG) and superior temporal gyrus (STG).
Speziell, we predicted the IFG and the head and body
of the caudate would show similar harmonic expectancy
recruitment and would functionally interact. This is based
on the well-established anatomical connections between
IFG and the anterior caudate (Haber, 2003; Alexander
et al., 1986) and is consistent with Koelsch and colleaguesʼ
arbeiten (2008), which reported head of the caudate activity in
a chord harmonic processing task that also activated the
IFG. The connections between auditory regions of the
superior temporal cortex and BG have been less well stud-
ied than other cortical regions. Yeterian and Pandya (1998)
reported projections from the anterior STG to the dorsal
anterior putamen and head of the caudate and projections
from the posterior STG to the posterior putamen and body
and tail of the caudate nucleus, and we therefore predicted
that these corresponding regions would show similar re-
cruitment by harmonic expectancy.
In conjunction with the BG, we were also interested in ex-
amining the role of the STG and IFG in harmonic processing.
Seger et al.
1063
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
T
.
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
parison with the isolated chords used in previous studies
described above. Two authors with extensive professional
musical training and composition experience ( J.D. Und
M.T.) selected pieces and composed alternative endings
that deviated to varying degrees from Western musical con-
Erfindungen. A total of 15 pieces written by European classical
Komponisten (z.B., J.S. Bach and Ludwig van Beethoven)
were taken from keyboard instruction sourcebooks. Pieces
varied in length from 10 Zu 24 sec when played. Popular
pieces and excerpts from major compositions were avoided
to minimize the chance of familiarity with the pieces among
the participants.
Western music is based on tonality within key centers.
The fundamental or starting pitch serves as the tonal center
for a given key. Relationships in tonal space between keys
can be determined by various systems of analysis (Krumhansl
& Cuddy, 2010; Lerdahl & Jackendoff, 1996). We chose to
use the circle of fifths, one of the most common systems,
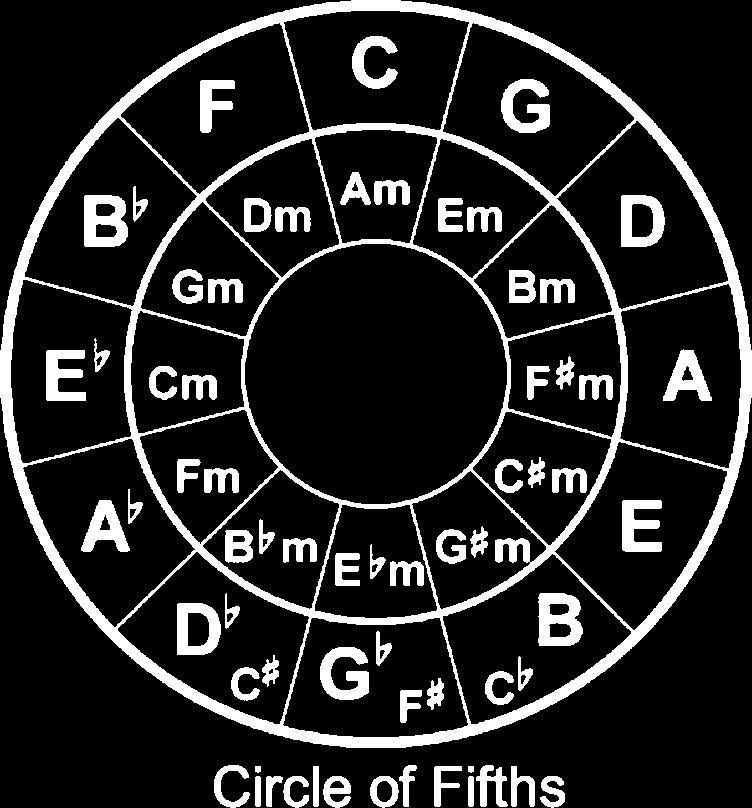
to characterize the degree of deviation in the stimuli. Der
circle of fifths is depicted in Figure 1. Janata et al. (2002)
found different voxels within cortical regions were sensitive
to different keys, indicating this tonal structure has some
neural validity. Maps of tonal key space have been identi-
fied within the rostromedial pFC (Menschen, 2009; Janata et al.,
2002), the left IFG (Menschen, 2009), and superior frontal gyrus
(Alluri et al., 2012).
Cadences, a progression of notes that concludes a
Phrase, section, or piece of music, are one of the most cen-
tral syntactic components in Western tonal music theory
and composition; they enforce the sense of a central pitch
or key center in a musical passage. One of the most com-
mon cadence configurations used to create a sense of
closure to a passage, or a whole piece, is presenting the
penultimate chord on the fifth scale degree (the dominant)
above the fundamental pitch (the tonic) before proceeding
to the tonic (z.B., G chord to C chord in the key of C). Der
The STG has been associated with processing sequences of
tones and melodic structures (Klein & Zatorre, 2011; Lee,
Menschen, Frost, Hanke, & Granger, 2011; Rogalsky, Rong,
Saberi, & Hickok, 2011). Musical processing in the IFG over-
laps with regions involved in motor and linguistic sequenc-
ing (Streuer & Tirovolas, 2009; Menschen & Grafton, 2003), Und
researchers have argued that this region contributes to syn-
tactic and hierarchical processing in music (Patel, 2003). Pre-
vious research investigating musical harmonic expectancy
has compared unexpected tonal chords (z.B., Neapolitan
subdominants) to standard chords within a chord progres-
sion context. Neapolitan subdominants lead to an early right
anterior negativity when studied using electroencephalogra-
phy (Kölsch, Gunter, Wittfoth, & Sammler, 2005) and mag-
netoencephalography (Maess, Kölsch, Gunter, & Friederici,
2001) and activity in the right inferior frontal lobe when
studied using fMRI (Kölsch, Fritz, Schulze, Alsop, &
Schlaug, 2005; Koelsch et al., 2002). On the basis of this re-
suchen, we predicted the IFG would be sensitive to musical
expectancy. We did not make any predictions about the STG
because previous research was unclear about its role in har-
monic processing, with some researchers arguing that STG
is limited to tonal processing, and others indicating a poten-
tial role of anterior STG regions in harmonic processing (sehen
Kölsch, 2011, für eine Rezension).
To summarize, we examined, for the first time, the role of
the BG in higher-order harmonic processing. We further
investigated the role of the IFG and STG in harmony pro-
cessing and examined interactions between these cortical
regions using GCM. We predicted that the BG and IFG
would be sensitive to harmonic expectancy violation, Das
sensitivity would be greatest in regions of the BG (d.h., Kopf
and body of the caudate) that interact with the IFG, Und
these regions would show functional connectivity during
music perception. Zusätzlich, we predicted STG recruit-
ment during music perception, along with regions of the
BG (d.h., the putamen and posterior caudate) that interact
with STG.
METHODEN
Teilnehmer
Eleven participants (six men, five women) were recruited
from the University of Colorado School of Medicine, Denver
Gemeinschaft. All participants met the criteria for MRI scan-
ning (d.h., no metallic implants, no claustrophobia, and no
known neurological injury or disease). All participants re-
ported normal hearing and no history of neurological dis-
orders or impairments that might affect musical processing.
Zusätzlich, all participants reported no formal music theory
training and no professional musical employment.
Stimuli
The stimuli were short, intact musical pieces. Choosing
intact musical pieces improves ecological validity, in com-
Figur 1. Circle of fifths.
1064
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 7
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
F
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
penultimate chord creates a strong sense of expectation
in Western music for the tonic to occur. Each stimulus
pieceʼs ending (the final cadence) closely followed stan-
dard tonal convention. Zusätzlich, each piece had a half
cadence in the middle of the piece, followed by a repeti-
tion of the first half of the music that ended with a full
cadence.
Three different types of musically realistic alternative
cadences were written for each piece, each diverging to
differing degrees from the cadence expectancies of
Western tonal music. In addition to the Standard Cadence
(d.h., original), we developed a Deceptive Cadence, A
Modulated Cadence, and an Atonal Cadence. In the Stan-
dard Cadence, the ending cadence remained in the same
key and resolved to the tonic chord; this is the most com-
mon resolution in Western tonal music and has a high
degree of expectancy. In the Deceptive cadence, Die
piece modulated to a closely related key built on the sub-
mediant (the sixth scale degree from the tonic note);
deceptive cadences are frequently used as a “surprise”
device by replacing the expected final cadence, thus pro-
viding for the continuation of the piece. Deceptive ca-
dences occur significantly less frequently than standard
cadences and are not used to end a piece (Huron, 2006).
In the Modulated Cadence, the piece modulated from the
original key (z.B., C) to a new distant key via varying
harmonic devices (d.h., a German sixth chord). Diese
chords can be inserted within the middle of pieces to build
additional compositional complexity but are never used as
the ending cadence in Western tonal pieces. This cadence
therefore has a lower expectancy than Deceptive Cadence.
Jedoch, the piece remains within the harmonic space of
Western tonal music. In the Atonal Cadence, the piece
shifted into a series of chords that are not part of the
“common practice” structure of Western tonal music; Die
final chord did not have a key center. All four versions of
each piece were equated for number of notes and rhythmic
properties. Thus to the greatest degree possible, only the
harmonic structure of the piece was altered across ca-
dences. An example piece with each of the four cadences
is notated in Figure 2.
Alle 60 stimuli were validated by a separate group of
27 Teilnehmer, who judged “How consistent is the end-
ing?” on a scale from 1 (sehr) Zu 8 (not very). These partic-
ipants were undergraduate students who participated in
partial fulfillment of a research requirement for introduc-
tory psychology and were not selected on the basis of
musical experience. Across all subjects and all pieces,
the Standard, Deceptive, Modulated, and Atonal cadences
received average ratings of 2.1, 4.4, 5.3, Und 7.2 bzw-
aktiv, mit 1 as the minimum and 8 as the maximum. A
one-way within-subject ANOVA performed on the aver-
age ratings of each cadence for each participant indicated
a significant effect of cadence, F(3, 78) = 255, P < .0001.
Post hoc tests indicated that Standard received significantly
lower ratings than Deceptive, t(26) = 13.0, p < .0001, De-
ceptive significantly lower than Modulated, t(26) = 5.37,
Figure 2. Top: A full piece with the Standard cadence (dashed box).
Bottom: Deceptive, Modulated, and Atonal cadences for same piece.
p < .0001, and Modulated significantly lower than Atonal,
t(26) = 12.68, p < .0001.
Procedure
Participants listened to a different musical piece on each
of the 60 trials (i.e., each stimulus was presented only
once). The order of the musical pieces was randomized
with the restriction that same piece or cadence type
would not be presented on two consecutive trials. Each
piece was prerecorded and played on a digitally synthe-
sized piano. When played, pieces ranged in duration from
10 to 24 sec. After the auditory presentation of each mu-
sical piece, the participants were visually probed with “Is
the music pleasant? Yes or No.” The probe question en-
couraged the participant to stay alert. Participants re-
sponded with their right index and middle fingers. A
period of time was added after the probe question, during
which no music or visual input was presented, to make the
total duration of each trial 30 sec.
Auditory stimuli were presented to participants through
a pair of magnet compatible headphones. Visual stimuli
were presented to participants using a magnet-compatible
projector that projects visual images onto a mirror attached
to the radio-frequency (RF) head coil. Responses were
Seger et al.
1065
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
o
9
c
1
n
2
_
8
a
/
_
j
0
o
0
c
3
n
7
1
_
a
p
_
d
0
0
b
3
y
7
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
recorded via a magnet-compatible button box. A computer
running E-Prime 2.0 experiment software (Psychology
Software Tools, Inc., Pittsburgh, PA) was used to control
stimulus presentation and interface with the magnet-
compatible response boxes. Head movement was mini-
mized using small foam pads placed on each side of the
head inside the RF head coil.
fMRI Data Acquisition and Processing
Imaging was obtained with a whole-body 3.0-T MRI scan-
ner (GE Healthcare, Milwaukee, WI) at the Brain Imaging
Center at the University of Colorado Denver (Aurora, CO).
The scanner was equipped with an eight-channel, high-
resolution phased array head coil using GEʼs Array Spatial
Sensitivity Encoding Technique (ASSET) software. Ana-
tomical images were collected using a T1-weighted spoiled
gradient recall sequence (minimal repetition time [TR],
minimal echo time, inversion time = 450 msec, flip angle =
10°, field of view = 220 nm, 256 × 256 coronal matrix,
166 1.2-mm slices). The structural images were used to
verify proper slice selection and to determine the sites of
functional activation (i.e., voxels that were found to be sig-
nificantly activated during the functional scan were over-
laid on the high-resolution structural images). Functional
images were reconstructed from 32 axial oblique slices
obtained using a T2*-weighted EPI-gradient recalled echo
sequence (TR = 2000 msec, echo time = 28 msec, flip
angle = 70°, field of view = 220 nm, 64 × 64 matrix,
40-mm slices, no interslice gap) to measure BOLD signal
change. In addition, the first three volumes, recorded be-
fore longitudinal magnetization reached a steady state,
were discarded.
We chose to record BOLD signal continuously, and as
a result, participants were exposed to scanner noise at
the same time as the musical stimuli. However, we utilized
high-quality magnet-compatible headphones, and all par-
ticipants reported that they were able to hear the musical
stimuli clearly above the scanner noise. We chose not to
use a sparse sampling sequence, in which active scanning
is limited to pauses between auditory stimuli, because it
would have imposed constraints on our analysis. To model
the BOLD signal both for the beginning of the piece
(Music Baseline condition) and the endings (Standard,
Deceptive, Modulated, and Atonal Cadence conditions),
it was essential to record the BOLD signal during the
entire stimulus presentation. A recent study comparing
continuous scanning with sparse acquisition techniques
during music perception reported comparable results for
both methods in the regions of the brain we examined
(Mueller et al., 2011).
Imaging preprocessing was performed using Brain
Voyager QX 1.1 (Brain Innovation, Maastricht, the
Netherlands). The functional data were first preprocessed
(i.e., three-dimensional motion correction, slice time cor-
rection, temporal data smoothing with a high-pass filter,
and linear trend removal). Each participantʼs high-resolution
anatomical image was normalized to the Talairach and
Tournoux (1998) brain template. The normalization pro-
cess consisted of two steps: first, an initial rigid body
translation into AC–PC plane and, second, an elastic defor-
mation into the standard space performed on 12 individ-
ual subvolumes. The resulting set of transformations
was applied to the participantʼs functional image volumes
to form volume time course representations to be used in
subsequent statistical analysis. Lastly, the volume time
course representations were spatially smoothed with a
Gaussian kernel FWHM of 6.0 mm.
Whole-brain Analyses
Brain Voyager QX 2.3 was used to analyze contrasts be-
tween conditions (Goebel, Esposito, & Formisano, 2006).
First, a prototypical hemodynamic response function was
convolved with the time course of the condition to create
condition-specific models. Then, the condition-specific
models were compared using the general linear model
(GLM) with participants treated as random effects. We cor-
rected for multiple comparisons using the cluster-size thresh-
olding procedure developed by Forman et al. (1995),
extended to 3-D maps, and implemented in the Brain
Voyager Cluster Threshold plug-in (Goebel et al., 2006).
An initial map was formed using an uncorrected voxelwise
threshold of p < .005. Then, the minimum cluster size (on
the basis of an alpha level of .05) was set by a 1000-iteration
Monte Carlo simulation, simulating the stochastic process
of image generation. Afterward, spatial correlations be-
tween neighboring voxels were calculated. Then, voxel in-
tensity thresholds were calculated, and the corrected map
was formed. The following six conditions were defined:
Music Baseline, Silence Baseline, Standard Cadence,
Deceptive Cadence, Modulated Cadence, and Atonal
Cadence. The Music Baseline epochs began at the start
of each musical piece and extended for a total of 3 TR
(6 sec). The Silence Baseline epochs were defined as the
last 2 TR (4 sec) of every trial. All cadence epochs (i.e.,
Standard, Deceptive, Modulated, and Atonal) were defined
as the final 1–2 TR (2–4 sec) of music for each respective
piece, depending on the specific length of the cadence
within a given piece.
ROI Analysis
Our goal was an unbiased and conservative analysis to
assess BG contributions to expectancy processing. To this
end, we adopted a set of anatomically defined ROIs. These
ROIs were previously defined for a different experiment
(Peterson & Seger, in press), thus completely avoiding
the possibility of biasing our anatomical ROI definitions
by the functional analysis results. This ensured ROI defini-
tion independence and avoided issues of “double dipping,”
the statistically improper method of defining ROIs on the
basis of a functional task and then further analyzing the
1066
Journal of Cognitive Neuroscience
Volume 25, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
o
9
c
1
n
2
_
8
a
/
_
j
0
o
0
c
3
n
7
1
_
a
p
_
d
0
0
b
3
y
7
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
data within these ROIs (Kriegeskorte, Simmons, Bellgowan,
& Baker, 2009; Poldrack & Mumford, 2009).
ROIs within the BG were defined a priori for the ven-
tral striatum and three subregions of the dorsal striatum:
the putamen, head of the caudate, and body of the cau-
date. These ROIs were hand drawn on an averaged image
formed from the normalized high-resolution anatomical
images from 10 participants. We manually verified the
BG in this averaged anatomical image were comparable
in size and shape to the averaged anatomical BG image
from the participants in the present experiment. Care
was taken to ensure only gray matter within each region
was selected; any surrounding white matter or gray matter
structures (e.g., thalamus) were excluded. The head of
the caudate was defined as extending posteriorly through
y = 3; the body of the caudate was defined as extending
anteriorly through y = 0. A 2-voxel-wide gap was in-
cluded between the ROIs to ensure independence. The
ventral striatal ROI included the ventral caudate (nucleus
accumbens) and ventral putamen, with the dividing line
between the ventral and dorsal striatum extending along
a diagonal from z = 5 in the most medial portions, down
to z = −4 in the most lateral portions. All ROIs were then
translated horizontally across the x axis to create comple-
mentary ROIs for the right hemisphere. These ROIs are
illustrated in Figure 5. The ROI random effects GLM tool
of BrainVoyager QX was used to analyze contrasts be-
tween conditions separately within each ROI.
Parametric Regressors
An a priori linear parametric regressor was defined to re-
flect the degree to which each cadence deviated from ex-
pected Western harmonic structure. We used the circle of
fifths (see Figure 1) to determine the numerical distance
between the original key and each modulated cadence.
The distance was computed as the smallest number of
steps between the roots of the original key and the final
cadence within the tonal space of 12 major and 12 minor
keys and then scaled to a range of 0.1 to 0.99, with the
Standard cadence at .1 and the Atonal Cadence at .99.
The Deceptive Cadence, modulating to a closely related
key, resulted in a numerical value of 0.2. The Modulated
Cadence, modulating out of original key via an aug-
mented sixth chord, resulted in a value of 0.5. This para-
metric regressor was implemented in the same manner
as the GLM used to perform the whole-brain analysis,
described above.
We also formed a parametric regressor based on the
average consistency ratings in the pilot testing of the
pieces (described above), scaled to a 0 to .99 range. This
resulted in weights of 0.16, 0.49, 0.62, and 0.89 for the
Standard, Deceptive, Modulated, and Atonal cadences,
respectively. Overall, these estimates are very close to
those formed using the circle of fifths method. The larg-
est discrepancy is for the Deceptive cadences, which
were relatively close in tonal space (scaled value of 0.2)
yet received relatively high ratings for being inconsistent
(scaled value of 0.49). Recent research in music is devel-
oping more sophisticated methods for calculating expec-
tancy based on psychoacoustic methods, and future
research may fruitfully incorporate these methods (Alluri
et al., 2012; Janata, 2009).
GCM
GCM was used to explore effective connectivity between
the striatum and other brain regions. The causality maps
were created using Roebroeck, Formisano, and Goebelʼs
(2005) procedure, as implemented within BrainVoyager
QX 2.3. The first step in GCM is identifying reference re-
gions, sometimes referred to as seed regions. Target re-
gions were defined as any voxel not included in the
reference region ( y). Influence measures were then calcu-
lated from the reference to target region (FX→Y), target to
reference region (FY→X), and total linear dependence be-
tween the reference and target regions (FX,Y) by repeatedly
pairing the time course maps of each voxel in these re-
gions. Time-course data were sampled from all trials.
GCM analyses were performed on the preprocessed data,
which included spatial smoothing. Directed influences to
and from the reference region were calculated by subtract-
ing the influence of the target to reference region from the
influence from the reference to the target region (FX→Y −
FY→X) for every voxel to calculate a difference (dGCM).
Thus, effective connectivity was described as dGCM =
FX→Y − FY→X (see Roebroeck et al., 2005, for details). A
positive difference value indicates FX→Y (reference →
volume) influence, whereas negative difference values de-
pict FY→X (volume → reference) influence.
We focused on three cortical (IFG, anterior STG, and
posterior STG) and two BG (caudate and putamen) seed
regions. The cortical regions were functionally defined
based on the All Cadences > Music Baseline contrast,
shown in Table 2. We choose right hemisphere ROIs
because they were more consistently active than the left
hemisphere ROIs; Jedoch, exploratory analyses of left
hemisphere ROIs found similar patterns. The right ante-
rior putamen was functionally defined based on the All
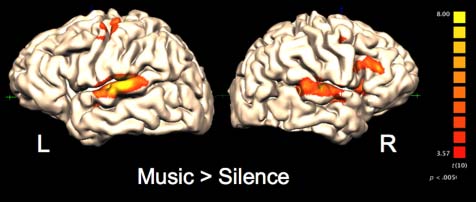
Figur 3. Regions with greater activity for Music Baseline than Silence
Baseline. Positive t values are shown in orange–yellow scale. Corrected for
multiple comparisons using the cluster level threshold method with a
voxelwise threshold of p < .005 and cluster threshold of p < .05.
Seger et al.
1067
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
o
9
c
1
n
2
_
8
a
/
_
j
0
o
0
c
3
n
7
1
_
a
p
_
d
0
0
b
3
y
7
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 1. Regions of Activity for Music
Music Baseline > Silence Baseline
R STG
L STG
R IFG
Medial frontal/supplementary motor area
R postcentral gyrus
L postcentral gyrus
X
52
−53
45
2
49
−48
j
−20
−20
14
5
−2
−14
z
5
9
21
53
42
55
Voxels
16556
15444
10231
3411
956
2332
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
X, j, z: Talairach coordinates of the central voxel within the activated cluster. Voxels: Size of cluster in voxels. R= right; L= left. Cluster size threshold
minimum of 82 voxels based on corrected alpha p < .05, voxelwise alpha p < .005.
Cadences > Music Baseline contrast. The right caudate
ROI was functionally defined based the results of the
parametric regressor.
The right IFG ROI was centered at x = 45, y = 23, z =
−4 and encompassed a total of 454 voxels. The right
anterior STG ROI was centered at x = 40, y = 17, z =
−17 and encompassed at total of 394 voxels. The right
posterior STG ROI was centered at x = 55, y = −20,
z = 0 and totaled 2502 voxels.1 The putamen ROI was
centered at x = 17, y = 2, z = 6 and encompassed a total
von 314 voxels. The caudate ROI was centered at x = 10,
y = −1, z = 15 and encompassed a total of 425 voxels.
Individual effective connectivity maps were computed for
each ROI for each participant, then activation across
maps were compared using a voxelwise t test examining
whether activity was significantly different from zero, at a
threshold of p < .05.
RESULTS
Overall Music Perception
As shown in Figure 3, extensive activations for music
(Music Baseline > Silence Baseline) were found bilat-
erally in secondary auditory cortical regions extending
along the STG. Music also recruited large regions of the
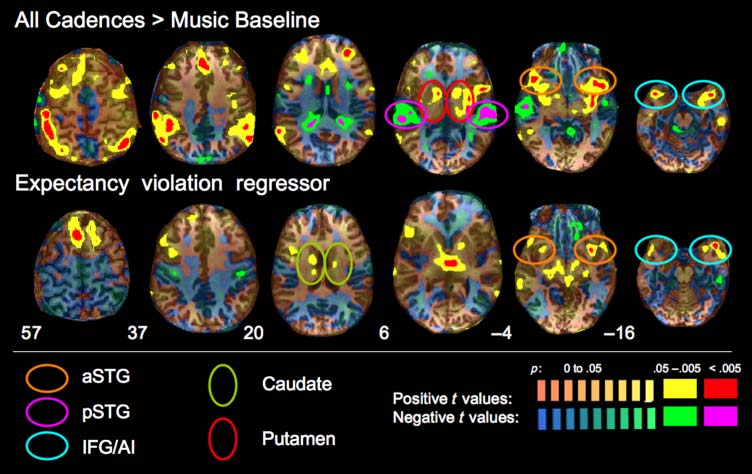
Figur 4. Top: Regions
of activity for all Cadences
compared with the Music
Baseline (initial segments of
musical pieces). Bottom:
Regions of activity during
cadence perception that were
modulated by the parametric
regressor scaled to cadence
expectancy. Positive t values
are coded by the red–yellow–
orange color scale and indicate
greater activity for Cadences
than Musical Baseline (top)
or activity that was positively
predicted by the expectancy
regressor (greater activity for
greater violation of expectancy,
bottom). Negative t values
are coded in the blue–green–
magenta color scale and
indicate the opposite contrasts:
greater activity for Musical
Baseline than the Cadences
(top) or activity negatively
predicted by the expectancy regressor (less activity for a greater violation of expectancy, bottom). Multiple thresholds are indicated via different color
Waage. For positive t values, red indicates regions of activity meeting a voxelwise threshold of p < .005, corrected for multiple comparisons with
the cluster level threshold method (cluster threshold p < .05); yellow indicates regions of activity meeting a voxelwise threshold of p < .05,
uncorrected. For negative t values, these cluster thresholds are indicated by magenta and bright green, respectively. The transparent red–orange
and blue–green color scales show unthresholded maps indicating t values that correspond to probability values ranging from p = 0 to p < .05.
Ovals indicate corresponding regions recruited in the two analyses. Orange= anterior STG; magenta= posterior STG; cyan= inferior frontal/anterior
insula; red= putamen; green= caudate.
1068
Journal of Cognitive Neuroscience
Volume 25, Number 7
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
o
9
c
1
n
2
_
8
a
/
_
j
0
o
0
c
3
n
7
1
_
a
p
_
d
0
0
b
3
y
7
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 2. Regions of Activation for Cadences
All Cadences > Music Baseline
Cortical regions
L superior frontal gyrus
Medial frontal gyrus
Medial frontal gyrus/supplementary motor area
L anterior STG
R anterior STG
L IFG/anterior insula
R IFG
L superior parietal lobe
R inferior and superior parietal lobe
L inferior parietal lobule
Subcortical regions
L cerebellum
R anterior putamen
L putamen/insula
Music Baseline > All Cadences
R posterior STG
R posterior STG
L posterior STG
R caudate (tail)
X
−27
2
−5
−39
39
−44
45
−37
50
−51
−41
9
−32
58
52
−53
16
j
48
38
24
14
16
14
22
−58
−40
−41
−32
5
−8
−27
−15
−21
−42
z
Voxels
22
34
48
−18
−18
−1
−3
59
44
37
−32
6
2
8
2
8
22
506
1228
192
307
219
1057
217
1207
6521
222
73
15
1220
595
625
2384
497
X, j, z: Talairach coordinates of the central voxel within the activated cluster. Voxels: Size of cluster in voxels. R= right; L= left. Cluster size threshold
minimum of 7 voxels based on corrected alpha p < .05, voxelwise alpha p < .005.
right IFG, including the right hemisphere homolog of
Brocaʼs area. In addition, music recruited neural regions
associated with motor processing, including the bilateral
sensorimotor cortex in the precentral and postcentral
gyrus and SMA. The recruitment of both motor planning
and execution regions is consistent with previous music
and motor studies (Zatorre, Chen, & Penhune, 2007;
Janata & Grafton, 2003). A complete list of active clusters
can be found in Table 1. We also found regions of activity
that were significantly more active during Silence Baseline
than Music Baseline in medial regions of the frontal and
parietal lobes (posterior cingulate/precuneus) and across
higher-order visual cortical regions. The former regions
are commonly active during relatively unstructured control
conditions in neuroimaging studies and participate in a
neural system often referred to as the “default network”
(Andrews-Hanna, Reidler, Huang, & Buckner, 2010). Rela-
tive decreases in visual cortex activity are often reported
when participants attend to an auditory task (Langner
et al., 2012).
Cadence Processing
To identify neural regions associated with ending cadence-
specific musical processing, we compared all cadence
epochs (i.e., Standard Cadence, Deceptive Cadence, Mod-
ulated Cadence, and Atonal Cadence) with Music Baseline
epochs (i.e., the first 6 sec of each piece). As shown in
Figure 4 (top row) and Table 2, there was significant acti-
vation in several regions of the pFC, including the bilateral
IFG. There was also activity in the bilateral anterior STG, a
region that is known to have strong interconnections with
the IFG and which is implicated in linguistic syntax pro-
cessing (Friederici, 2009). There was also significant activity
in parietal lobe regions and in the medial frontal gyrus. Sub-
cortically, this contrast recruited regions of the cerebellum,
the anterior putamen and head of the caudate (bilaterally),
and the left posterior putamen and insula. The reverse con-
trast (Music Baseline > All Cadences) found increased pos-
terior STG and tail of the caudate activity during the
beginnings of the musical pieces in contrast with the
Seger et al.
1069
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
F
/
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
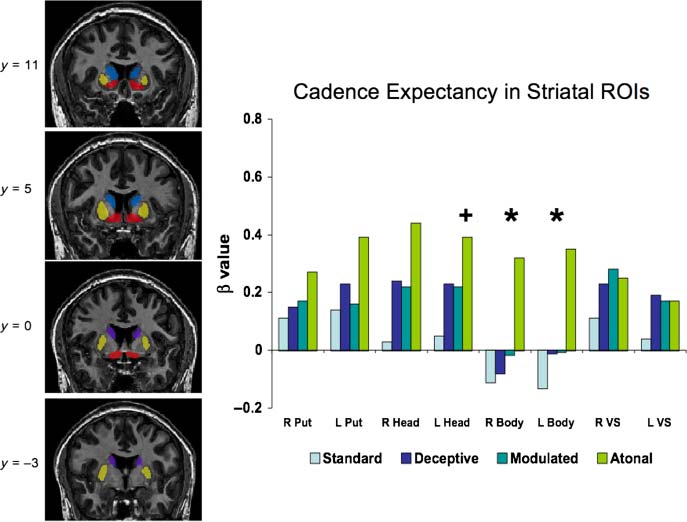
Figur 5. (Links) BG ROIs
used in the parametric analysis.
Yellow= putamen; blue= head
of the caudate; purple= body
of the caudate; red= ventral
striatum. (Rechts) Beta weights
within the BG ROIs for Standard
Cadence, Deceptive Cadence,
Modulated Cadence, and Atonal
Cadence. *regions that were
significantly modulated by the
parametric regressor, P < .05;
+regions with a trend toward
modulation by the parametric
regressor, p < .1. L= left;
R= right; Put= putamen;
Head= head of the caudate
nucleus; Body= body of the
caudate nucleus; VS= ventral
striatum. See text for
analysis details.
cadences. This unexpected pattern could reflect a variety of
mechanisms, including increased analytical demands for
processing tones at the beginning of a piece or habituation
during extended musical processing.
Parametric Analysis of Cadence Expectancy
To identify neural regions sensitive to the degree of expec-
tancy violation within the cadences, we utilized the para-
metric regressor based on the circle of fifths described
above. Within the anatomically defined striatal ROIs, the re-
gressor significantly predicted both right and left body of
the caudate activity (right: t(10) = 2.4, p = .03; left: t(10) =
2.1, p = .05), with a trend toward predicting activity in the
right dorsal head of caudate (t(10) = 2.0, p = .07). This
finding is illustrated in Figure 5.
We also incorporated the regressor in a whole-brain
analysis to identify regions outside the BG significantly
modulated by expectancy. This analysis indicated signifi-
cant activity in the following cortical regions: the left ante-
rior STG, the left IFG bordering on the anterior insula,
and the supplementary motor region. This finding is illus-
trated in Figure 4 (bottom row), and significant clusters
are listed in Table 3. Specific beta values within each of
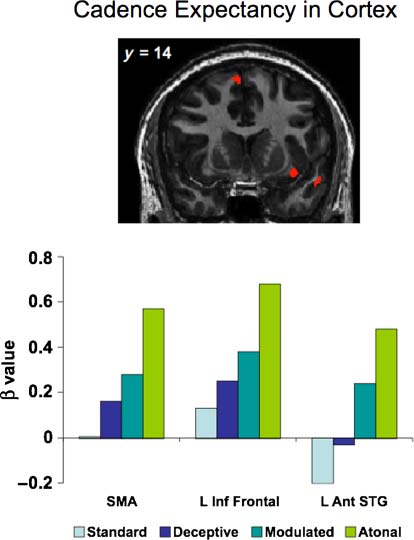
these three regions are shown in Figure 6. As shown in
Figure 4, there was substantial activity in right hemisphere
homologous regions at a lower statistical threshold. Over-
all, similar patterns of recruitment in bilateral IFG and
Table 3. Regions of Activity Predicted by Cadence Expectancy (Parametric Regressor)
x
y
Positive Relation to Parametric Regressor (Increased Activity with Increased Expectancy Violation)
Bilateral thalamus/midbrain
Medial frontal gyrus/supplementary motor area
L IFG/anterior insula
L anterior STG
0
8
−32
−48
−14
16
12
14
Negative Relation to Parametric Regressor (Decreased Activity with Increased Expectancy Violation)
Medial parietal/paracentral lobule
R inferior temporal gyrus
−3
−53
−30
−31
z
5
60
6
−12
50
−22
Voxels
1211
437
181
488
501
723
x, y, z: Talairach coordinates of the central voxel within the activated cluster. Voxels: Size of cluster in voxels. R= right; L= left. Cluster size threshold
minimum of 7 voxels, corrected alpha p < .05, voxelwise alpha p < .005.
1070
Journal of Cognitive Neuroscience
Volume 25, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
o
9
c
1
n
2
_
8
a
/
_
j
0
o
0
c
3
n
7
1
_
a
p
_
d
0
0
b
3
y
7
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
bilateral anterior STG, as well as in the putamen, can be
seen by comparing the top (Cadences > Music Baseline
Kontrast) rows to the bottom (the parametric regressor)
rows of Figure 4. We found head and body of the caudate
activity at the lenient threshold, consistent with the ana-
tomical ROI analysis (siehe Abbildung 4, bottom row, slice at z =
20). There was a striking difference between the parietal
and superior temporal lobes. The parietal cortex, bilater-
ally, was more active for cadences than early musical seg-
gen, yet there was little indication that it was modulated
by expectancy. Andererseits, the posterior STG were
more active for early music than for cadences and also
showed little evidence of sensitivity to the expectancy vio-
lation regressor. Supplementary motor region recruitment
for expectancy was not predicted, although a similar region
was identified by Alluri et al. (2012) as negatively correlated
with key clarity during music perception. It may reflect
general processes of conflict detection and resolution
within the medial frontal cortex (Nee, Kastner, & Braun,
2011).
In addition to the circle of fifths-derived regressor, Wir
also implemented a parametric regressor based on the
mean expectancy ratings made by a separate group of
27 Teilnehmer (see above). Gesamt, the two sets of regres-
sor weights were very similar: 0.1, 0.2, 0.5, Und 0.99 für die
circle of fifths weights, compared with 0.16, 0.48, 0.62, Und
0.89 for the participant ratings weights. This regressor re-
sulted in almost identical patterns of activity as found when
using the circle of fifths derived regressor.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
/
T
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Seger et al.
1071
Figur 6. Activity within cortical regions identified in the whole-brain
parametric analysis as significantly sensitive to cadence expectancy.
Top: Regions overlaid on a coronal image at y = 14. Beta weights
are plotted for the Standard Cadence, Deceptive Cadence, Modulated
Cadence, and Atonal Cadence. SMA= supplementary motor area.
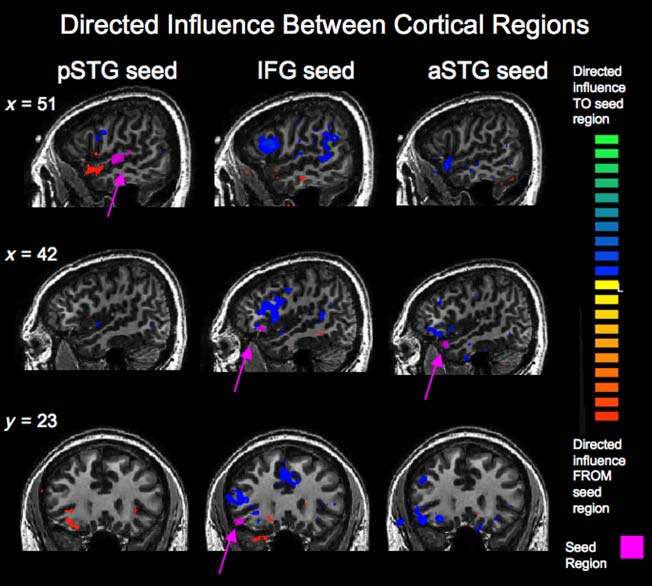
Figur 7. Directed influences
to and from cortical seed
regions in the right posterior
STG (pSTG), right IFG, Und
right anterior STG (aSTG) als
measured by GCM. Seed
regions are shown magenta.
Regions that the seed region
exerted directed influence
on are shown in warm colors,
and regions exerting directed
influence on the seed region are
shown in cool colors. Gesamt,
the pSTG exerted directed
influence onto the IFG and
aSTG (left column). The IFG
(middle column) received
directed influence from the
pSTG and nearby regions of the
frontal cortex. The aSTG (Rechts
column) also received directed
influence from nearby regions
of the frontal cortex.
between right posterior STG and the BG. Speziell, Dort
was directed influence from the putamen (Figur 8,
bottom left) and directed influence to the ventral striatum
(not illustrated).
There was directed influence onto the right IFG re-
gion from adjacent regions of the IFG and middle fron-
tal gyrus in the right hemisphere as well as homologous
regions in the left hemisphere. The right IFG showed
directed influence both from and onto small regions
of the anterior STG in both hemispheres. There was di-
rected influence from the posterior STG bilaterally, con-
sistent with posterior STG seed results. There was also
directed influence from the bilateral inferior parietal re-
gions around the angular gyrus, which is consistent with
the known interactions between inferior parietal and
IFG during verbal and auditory working memory (Burzynska
et al., 2011). Endlich, there were directed influences from
the putamen and onto the caudate (Figur 8, bottom
center).
The right anterior STG seed region exerted directed
influence onto adjacent regions of the anterior STG and
inferior and middle frontal regions in both hemispheres
(Figur 7, right column). Interactions between right ante-
rior STG and the BG were similar to those for the IFG seed:
The anterior STG received directed influence from the
putamen and exerted directed influence on the caudate
(Figur 8, bottom right).
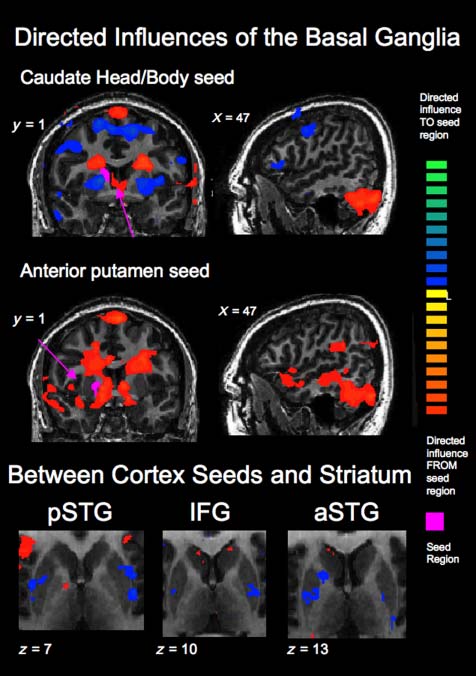
The patterns of directed influences between cortex
and each of the two BG seed regions differed substan-
tially, as shown in the top two rows of Figure 8. Der
putamen seed region exerted directed influence on
the posterior and anterior regions of the superior tem-
poral lobe, the TPJ, and the caudate nucleus. The cau-
date seed region, which was at the junction of the head
and the body of the caudate, showed directed influence
from several frontal cortex regions, including the bilat-
eral IFG, bilateral dorsolateral pFC, and bilateral medial
pFC. Within the striatum, the putamen seed exerted
directed influence on the caudate nucleus, and the cau-
date seed received directed influence from the putamen
and exerted directed influence on other regions of the
Schwanz. The directed influence from putamen onto the
caudate has been found previously in our lab in category
learning tasks (Lopez-Paniagua & Seger, 2011; Seger,
Dennison, Lopez-Paniagua, Peterson, & Roark, 2011;
Seger et al., 2010), but this is the first time this putamen–
caudate interaction has been reported in another cogni-
tive domain.
Both the putamen and caudate seed regions received
directed influence from large regions of the cerebellum; Es
is unclear what functional role these interactions may have
in music perception. There is a growing literature concern-
ing the roles of the cerebellum in music perception, mit
many studies indicating a role in temporal properties of
Musik (Lebrun-Guillaud, Tillmann, & Justus, 2008), inkl-
ing rhythm processing (Chen et al., 2008) and auditory–
motor synchronization (Thaut et al., 2009). It is less clear
Figur 8. (Top) Directed influences to and from the anatomically
defined R body of the caudate seed region (see top left slice for seed
region location). The body of the caudate interacted with other regions
of the striatum in that it received influence from the putamen and
exerted influence on other parts of the caudate nucleus (best illustrated
in the top left image; the top right and bottom left also show the
directed influence on the head and tail of the caudate, jeweils).
The body of the caudate also received directed influence from the
posterior STG and IFG. (Bottom) Directed influences to and from
the cortical seed regions shown in Figure 7 within the caudate and
putamen. The putamen exerted directed influence on all three seed
Regionen, whereas the caudate received directed influence from the
IFG and anterior STG.
GCM
We examined patterns of directed influence from the
following five seed regions in the right hemisphere: Die
posterior STG, the IFG, the anterior STG, the anterior
putamen, and the caudate. The defining procedures for
these seed regions were discussed above. Although we
focused on the right hemisphere seed regions, ähnlich
directed influence patterns were found for left hemi-
sphere seed regions. We found directed influence from
the right posterior STG onto the IFG and adjacent re-
gions of the STG, including the anterior STG (Figur 7,
left column). There was also directed influence from
small regions of bilateral primary motor cortex and to the
left anterior STG and cerebellum. We found interaction
1072
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 7
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
F
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
whether the cerebellum plays a role in tonal properties of
Musik, though studies have found a potential role in pitch
perception (Parsons, Petacchi, Schmahmann, & Bower,
2009).
DISKUSSION
We found regions of the BG, in particular the anterior
caudate nucleus, and cortex, in particular the IFG and
anterior STG, were modulated by expectancy violation
during music perception. The bilateral posterior STG
were active for music but were not sensitive to expecta-
tion violation. Our results are consistent with music the-
ories that argue music requires processes of structural
development in which expectancy increases and resolu-
tion in which the expectancies are confirmed (Huron,
2006; Meyer, 1989).
The BG in Harmonic Expectancy Processing
BGʼs sensitivity to musical expectancy violation was pre-
dicted by reinforcement learning theories that propose
BG networks represent both reward prediction and pre-
diction error (Glimcher, 2011). This is consistent with
previous findings of BG recruitment for prediction and
prediction error in other cognitive domains, including rea-
soning (Kranjec et al., 2012; Lohrenz et al., 2007), sequence
Verarbeitung (Bahlmann et al., 2009), and event perception
(Zacks et al., 2011). Our results provide convergent evi-
dence from music for the general role of the BG in both
reward prediction and reward prediction error.
Within the BG, both the anatomical ROI analysis and
whole-brain analysis indicated significant recruitment dur-
ing expectancy violation was localized to the body and
head of the caudate. Our ROI analysis further showed a
common, but nonsignificant, general pattern of expec-
tancy sensitivity across all regions of the striatum (sehen
Figur 5). The parallel pattern of activation of head and
body region of the caudate along with the IFG is con-
sistent with the known interaction pattern within the
“executive” corticostriatal loop. Zusätzlich, GCM showed
directed influence from frontal cortical regions to the
right caudate seed region. The putamen showed a similar
pattern of activity: activation in the All Cadences > Music
Baseline contrast and modulated by the parametric re-
gressor at a lenient threshold. Jedoch, GCM found a
very different pattern of functional connectivity for the
putamen than the caudate. The putamen interacted with
the anterior and posterior superior temporal regions,
consistent with anatomical connections between these
Regionen (Yeterian & Pandya, 1998). Endlich, the All
Cadences > Music Baseline contrast revealed regions of
the tail of the caudate that were more active for the
beginnings of the pieces of music than the ending ca-
dences but was not notably affected by the expectancy
regressor; this same pattern of activity was found in the
posterior STG, known to project to the tail of the caudate
(Yeterian & Pandya, 1998).
We did not find significant modulation of the ventral
striatum by expectancy (although Figure 5 does show a
small trend toward less activity for the Standard cadences
compared with the other cadences). This result is in con-
trast to studies that have found ventral striatal activity
associated with the rewarding and pleasurable aspects
of music (Montag, Reuter, & Axmacher, 2011; Wow &
Kringelbach, 2010) and that found dopamine release in
the ventral striatum associated with listening to musical
segments selected by subjects individually as being in-
tensely rewarding and emotional (Salimpore, Benovoy,
Larcher, Dagher, & Zatorre, 2011).
Our findings of BG recruitment during harmonic pro-
cessing have implications for both the fields of BG re-
search and musical neuroscience. BG research would
benefit from the continued exploration of music as a spe-
cific instantiation of the more general prediction functions
of the BG. Examination of expectancies across musical
domains (z.B., rhythm, melody, and harmony) may provide
insights into the roles of different corticostriatal loops.
Given the BGʼs highly plastic nature, they play an im-
portant role beyond representing expectations within the
current environment to learning what to expect in the fu-
tur (Frank, 2011; Ashby, Turner, & Horvitz, 2010; Seger,
2009), which may have implications for theories of music
Erwerb.
The Inferior Frontal Lobe and Anterior STG
in Harmonic Processing
The IFG has three main subregions: the frontal operculum
bordering on the anterior insula (classic Brocaʼs area, often
identified as BA 44), an immediately anterior region (often
identified as BA 45), and an immediately inferior region
(often identified as BA 47). All three of these areas, In
the right hemisphere, were active when comparing music
to silence, but their activity was limited to the inferior re-
gion in our analyses of cadences and cadence expectancy.
Our results are dissimilar from previous studies, comparing
standard chord progressions with unexpected Neapolitan
subdominant chord progressions, which found broad
inferior frontal lobe recruitment for unexpected chord pro-
gressions (Kölsch, Fritz, et al., 2005). This inferior frontal
lobe recruitment was associated with ERAN (Early Right
Anterior Negativity) modulation, an ERP shown to originate
in right frontal cortex (Garza Villarreal, Brattico, Leino,
Ostergaard, & Wow, 2011; Maidhof & Kölsch, 2011;
Kölsch, Gunter, et al., 2005; Maess et al., 2001). Neapolitan
subdominants rarely appear in Western tonal music, Und
although they are consistent with its tonal structure, Sie
require a specific harmonic context to make syntactic sense.
It is still unclear how inferior frontal regions differ in
their contributions to music processing. Both natural lan-
guage and artificial grammar studies found evidence for
a posterior (less complex hierarchy) to anterior (mehr
Seger et al.
1073
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
/
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
complex hierarchy) processing gradient within the inferior
frontal region (Perani et al., 2011; Santi & Grodzinsky,
2010). Zusätzlich, more anterior regions are recruited to
process syntactic dependencies across longer lags (Bahlmann
et al., 2009; Bahlmann, Schubotz, & Friederici, 2008; Opitz
& Friederici, 2007). BA 47 was recruited during polyrhythm
hierarchical processing (Wow, Wallentin, Mouridsen,
Ostergaard, & Roepstorff, 2011). The inferior frontal region
is important for verbal working memory and the articu-
latory loop (Groussard et al., 2010), and it is possible that
cadences with higher degree of expectation violation have
increased working memory demands. The IFG is active
during melodic working memory tasks (Jerde, Childs, Handy,
Nagode, & Pardo, 2011; Schulze, Zysset, Mueller, Friederici,
& Kölsch, 2010). Jedoch, language studies found this
region makes contributions to syntax that extend beyond
Arbeitsgedächtnis (Makuuchi, Bahlmann, Anwander, &
Friederici, 2009).
A similar sensitivity to cadence expectancy was found in
both anterior STG and IFG. Previous music studies have
not distinguished between the contributions from these
two regions. In the domain of language, the anterior STG
has been associated with syntactic processing (Brennan
et al., 2012). Anatomically, the anterior STG region is highly
interconnected with the IFG (Friederici, 2009). Some stud-
ies (Friederici, 2009), aber nicht alles (Wilson et al., 2011), have
related individual differences in syntactic processing ability
to differences in white matter connectivity within this path-
Weg. Our results imply that the anterior STG may play a role
in processing musical as well as linguistic syntax and the
musical processing functions of the anterior STG differ
from those of the posterior STG, which was not sensitive
to harmonic expectancy in the current study.
Functional Connectivity across Frontal,
Temporal, and Striatal Regions
We found that both anterior STG and IFG were sensitive to
expectancy. GCM provided convergent evidence for the
presence of this functional interaction: Seed regions in
both anterior STG and IFG indicated functional connectiv-
ity between these two regions. GCM also indicated func-
tional connectivity between the posterior STG and IFG
and anterior STG. Interactions between IFG and the STG
are consistent with studies of congenital amusia that find
decreased gray matter in both regions along with de-
creased white matter in the tracts connecting the two re-
gions (Hyde, Zatorre, & Peretz, 2011; Peretz, Brattico,
Järvenpää, & Tervaniemi, 2009). Zusätzlich, white matter
connectivity in this pathway is associated with the ability
to learn new syntactic structures in the auditory domain
(Lopez-Barroso et al., 2011; Louie, Li, & Schlaug, 2011) Und
increased white matter connectivity is found in these tracts
after intensive melodic intonation therapy for nonfluent
aphasia (Schlaug, Marchina, & Norton, 2009).
GCM also supports the presence of functional inter-
action between anterior STG, posterior STG, IFG, Und
BG during music perception. Gesamt, these interactions
were consistent with the known anatomical connections
between cortex and BG within corticostriatal loops. Der
putamen interacted with the STG, whereas the caudate
interacted with the IFG.
GCM results should be considered within the method-
ologyʼs limitations. Erste, GCM influence measures do not
necessarily reflect a direct anatomic connection between
Regionen; GCM does not discriminate between direct con-
nections and indirect influences through a third region.
Zweite, recent fMRI simulations have indicated that all
functional connectivity techniques, including GCM, have
limited ability to accurately determine the direction of
influences (z.B., whether region A is causing activity in
region B or vice versa; Smith et al., 2011).
Characterizing the Roles of Frontal and
Temporal Regions
The current study contributes to the emerging view that
early music perception recruits the posterior STG and later
perception recruits anterior regions of the anterior STG
and IFG. In addition to the GCM results, this view is sup-
ported by ERP studies finding an early MMN in temporal
sites and a later ERAN in frontal sites (Garza Villarreal
et al., 2011; Doeller et al., 2003). The posterior STG ap-
pears to be recruited for individual pitch processing and
chord processing (Fujisawa & Cook, 2011; Klein & Zatorre,
2011) and simple pitch sequence processing in melody
(Rogalsky et al., 2011; Stewart, Overath, Warren, Foxton,
& Griffiths, 2008; Brattico, Tervaniemi, Näätänen, & Peretz,
2006; Patterson, Uppenkamp, Johnsrude, & Griffiths, 2002).
Acquired musical perception deficits are associated with
primarily posterior STG damage (Stewart, von Kriegstein,
Warren, & Griffiths, 2006), and musical training is asso-
ciated with changes in this region (Hyde et al., 2009).
These results are consistent with our finding that the pos-
terior STG was recruited for musical processing overall but
not differentially recruited for different cadence types.
Anterior regions of the STG and IFG are typically recruited
for chord progressions and natural music with harmonic
content (d.h., pieces with multiple voices; Caria, Venuti, &
de Falco, 2011; Kölsch, Fritz, et al., 2005). Harmonized
music differs from simple melody and isolated chords in
a number of salient ways. It requires processing more
acoustic information overall, tracking tones both hori-
zontally (across time) and vertically (simultaneously in
Zeit), and interpreting the piece in light of previously
learned harmonic structure of the musical system. Re-
cruitment of anterior STG for harmonic processing is
consistent with studies showing hierarchical organiza-
tion of the STG, with anterior regions representing more
complex information (Chevillet, Riesenhuber, &
Rauschecker, 2011). Garza Villarreal et al. (2011) charac-
terized the difference between regions as the posterior
STG detecting sequential scale regularities and the IFG
parsing hierarchical regularities.
1074
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 7
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
.
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Abschluss
Our study contributes to the understanding of the neural
bases of music by mapping, for the first time, a cortico-BG
network underling harmonic expectancy processing. Es
contributes to the understanding of music as embodying
development of tension and resolution and relates these
functions in a broader context, as one of many several types
of cognitive expectancy. Speziell, this study provides
evidence for the BGʼs role in processing musical expecta-
tion violation. It supports the view that inferior frontal and
anterior superior temporal regions, previously linked to
processing linguistic syntax, are also involved in processing
musical syntax, and suggests differences in the contribu-
tions of the anterior and posterior STG to music percep-
tion. Endlich, our study contributes a broader trend within
cognitive neuroscience toward an increased appreciation
of the predictive nature of many neural systems (Friston
& Kiebel, 2009; Summerfield & Egner, 2009).
Reprint requests should be sent to Carol A. Seger, Department
of Psychology, 1876 Campus Delivery, Colorado State University,
Fort Collins, CO 80523, oder per E-Mail: Carol.Seger@colostate.edu.
Notiz
1. This region was more active overall in Music Baseline > All
Cadences contrast, which was the opposite direction of activity
than the other two ROIs.
VERWEISE
Alexander, G. E., DeLong, M. R., & Strick, P. L. (1986). Parallel
organization of functionally segregated circuits linking basal
ganglia and cortex. Jahresrückblick auf die Neurowissenschaften, 9,
357–381.
Alluri, V., Hoffnungsvoll, P., Jääskeläinen, ICH. P., Glerean, E., Sams, M.,
& Brattico, E. (2012). Large-scale brain networks emerge from
dynamic processing of musical timbre, key and rhythm.
Neurobild, 59, 3677–3689.
Andrews-Hanna, J. R., Reidler, J. S., Huang, C., & Buckner,
R. L. (2010). Evidence for the default networkʼs role
in spontaneous cognition. Journal of Neurophysiology,
104, 322–335.
Ashby, F. G., Turner, B. O., & Horvitz, J. C. (2010). Cortical and
basal ganglia contributions to habit learning and automaticity.
Trends in den Kognitionswissenschaften, 14, 208–215.
Bahlmann, J., Schubotz, R. ICH., & Friederici, A. D. (2008).
Hierarchical artificial grammar processing engages Brocaʼs
Bereich. Neurobild, 42, 525–534.
Bahlmann, J., Schubotz, R. ICH., Mueller, J. L., Koester, D., &
Friederici, A. D. (2009). Neural circuits of hierarchical
visuo-spatial sequence processing. Gehirnforschung,
1298C, 161–170.
Brattico, E., Tervaniemi, M., Näätänen, R., & Peretz, ICH. (2006).
Musical scale properties are automatically processed in the
human auditory cortex. Gehirnforschung, 1117, 162–174.
Braunlich, K., & Seger, C. A. (2013). Basal ganglia.
Wiley Interdisciplinary Reviews: Cognitive Science, 4,
135–148.
Brennan, J., Niv, Y., Hasson, U., Malach, R., Heeger, D. J., &
Pylkkänen, L. (2012). Syntactic structure building in the
anterior temporal lobe during natural story listening.
Brain and Language, 120, 163–173.
Bromberg-Martin, E. S., Matsumoto, M., & Hikosaka, Ö. (2010).
Dopamine in motivational control: Rewarding, aversive, Und
alerting. Neuron, 68, 815–834.
Burzynska, A. Z., Nagel, ICH. E., Preuschhof, C., Li, S. C.,
Lindenberger, U., Backmann, L., et al. (2011). Microstructure
of frontoparietal connections predicts cortical responsivity
and working memory performance. Hirnrinde, 21,
2261–2271.
Caria, A., Venuti, P., & de Falco, S. (2011). Functional and
dysfunctional brain circuits underlying emotional processing
of music in autism spectrum disorders. Hirnrinde,
21, 2838–2849.
Chen, J. L., Penhune, V. B., & Zatorre, R. J. (2008). Listening
to musical rhythms recruits motor regions of the brain.
Hirnrinde, 18, 2844–2854.
Chevillet, M., Riesenhuber, M., & Rauschecker, J. P. (2011).
Functional correlates of the anterolateral processing
hierarchy in human auditory cortex. Zeitschrift für
Neurowissenschaften, 31, 9345–9352.
Doeller, C. F., Opitz, B., Mecklinger, A., Krick, C., Reith, W., &
Schröger, E. (2003). Prefrontal cortex involvement in
preattentive auditory deviance detection: Neuroimaging
and electrophysiological evidence. Neurobild, 20,
1270–1282.
Forman, S. D., Cohen, J. D., Fitzgerald, M., Eddy, W. F.,
Mintun, M. A., & Noll, D. C. (1995). Improved assessment
of significant activation in functional magnetic resonance
Bildgebung (fMRT): Use of a cluster-size threshold. Magnetic
Resonance in Medicine, 33, 636–647.
Frank, M. J. (2011). Computational models of motivated action
selection in corticostriatal circuits. Aktuelle Meinung in
Neurobiology, 21, 381–386.
Friederici, A. D. (2009). Pathways to language: Fiber tracts in
das menschliche Gehirn. Trends in den Kognitionswissenschaften, 13, 175–181.
Friston, K., & Kiebel, S. (2009). Predictive coding under the
free-energy principle. Philosophical Transactions of the
Royal Society of London, Serie B, Biological Sciences,
364, 1211–1221.
Fujisawa, T. X., & Cook, N. D. (2011). The perception of
harmonic triads: Eine fMRT-Studie. Brain Imaging and
Behavior, 5, 109–125.
Garza Villarreal, E. A., Brattico, E., Leino, S., Ostergaard, L., &
Wow, P. (2011). Distinct neural responses to chord
violations: A multiple source analysis study. Gehirn
Forschung, 1389, 103–114.
Glimcher, P. W. (2011). Understanding dopamine and
reinforcement learning: The dopamine reward prediction
error hypothesis. Proceedings of the National Academy
of Sciences, USA., 108(Suppl. 3), 15647–15654.
Goebel, R., Esposito, F., & Formisano, E. (2006). Analysis of
functional image analysis contest (FIAC) data with
brainvoyager QX: From single-subject to cortically aligned
group general linear model analysis and self-organizing group
independent component analysis. Kartierung des menschlichen Gehirns,
27, 392–401.
Grahn, J. A., & Rowe, J. B. (2009). Feeling the beat: Premotor
and striatal interactions in musicians and nonmusicians
during beat perception. Zeitschrift für Neurowissenschaften, 29,
7540–7548.
Groussard, M., Rauchs, G., Landeau, B., Viader, F., Desgranges,
B., Eustache, F., et al. (2010). The neural substrates of
musical memory revealed by fMRI and two semantic tasks.
Neurobild, 53, 1301–1309.
Haber, S. N. (2003). The primate basal ganglia: Parallel and
integrative networks. Journal of Chemical Neuroanatomy,
26, 317–330.
Seger et al.
1075
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
/
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Huron, D. (2006). Sweet anticipation: Music and the
psychology of expectation. Cambridge, MA: MIT Press.
Hyde, K. L., Lerch, J., Norton, A., Forgeard, M., Gewinner, E.,
Evans, A. C., et al. (2009). Musical training shapes structural
brain development. Zeitschrift für Neurowissenschaften, 29, 3019–3025.
Hyde, K. L., Zatorre, R. J., & Peretz, ICH. (2011). Functional MRI
evidence of an abnormal neural network for pitch processing
in congenital amusia. Hirnrinde, 21, 292–299.
Menschen, P. (2009). Die neuronale Architektur musikevozierter Musik
autobiografische Erinnerungen. Hirnrinde, 19, 2579–2594.
Menschen, P., Birk, J. L., Van Horn, J. D., Leman, M., Tillmann, B.,
& Bharucha, J. J. (2002). The cortical topography of tonal
structures underlying Western music. Wissenschaft, 298,
2167–2170.
Menschen, P., & Grafton, S. T. (2003). Swinging in the brain: Shared
neural substrates for behaviors related to sequencing and
Musik. Naturneurowissenschaften, 6, 682–687.
Jerde, T. A., Childs, S. K., Handy, S. T., Nagode, J. C., & Pardo,
J. V. (2011). Dissociable systems of working memory for
rhythm and melody. Neurobild, 57, 1572–1579.
Klein, M. E., & Zatorre, R. J. (2011). A role for the right superior
temporal sulcus in categorical perception of musical chords.
Neuropsychologie, 49, 878–887.
Kölsch, S. (2011). Toward a neural basis of music perception—
A review and updated model. Grenzen in der Psychologie,
2, 110.
Kölsch, S., Fritz, T., & Schlaug, G. (2008). Amygdala activity
can be modulated by unexpected chord functions during
Musik hören. NeuroReport, 19, 1815–1819.
Kölsch, S., Fritz, T., Schulze, K., Alsop, D., & Schlaug, G.
(2005). Adults and children processing music: Eine fMRT-Studie.
Neurobild, 25, 1068–1076.
Kölsch, S., Gunter, T. C., von Cramon, D. Y., Zysset, S.,
Lohmann, G., & Friederici, A. D. (2002). Bach speaks: A
cortical “language-network” serves the processing of music.
Neurobild, 17, 956–966.
Kölsch, S., Gunter, T. C., Wittfoth, M., & Sammler, D. (2005).
Interaction between syntax processing in language and in
Musik: An ERP study. Zeitschrift für kognitive Neurowissenschaften,
17, 1565–1577.
Kranjec, A., Cardillo, E. R., Schmidt, G. L., Lehet, M., &
Chatterjee, A. (2012). Deconstructing events: The neural
bases for space, Zeit, and causality. Zeitschrift für Kognition
Neurowissenschaften, 24, 1–16.
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., & Bäcker,
C. ICH. (2009). Circular analysis in systems neuroscience:
The dangers of double dipping. Naturneurowissenschaften,
12, 535–540.
Krumhansl, C. L., & Cuddy, L. L. (2010). A theory of tonal
hierarchies in music. In M. R. Jones et al. (Hrsg.), Musik
perception, Springer handbook of auditory research (P. 36).
New York: Springer.
Langner, R., Kellermann, T., Eickhoff, S. B., Boers, F.,
Chatterjee, A., Willmes, K., et al. (2012). Staying responsive to
die Welt: Modality-specific and −nonspecific contributions
to speeded auditory, tactile, and visual stimulus detection.
Kartierung des menschlichen Gehirns, 33, 398–418.
Lawrence, A. D., Sahakian, B. J., & Robbins, T. W. (1998).
Cognitive functions and corticostriatal circuits: Insights
from Huntingtonʼs disease. Trends in den Kognitionswissenschaften,
2, 379–388.
Lebrun-Guillaud, G., Tillmann, B., & Justus, T. (2008).
Perception of tonal and temporal structures in chord
sequences by patients with cerebellar damage. Musik
Wahrnehmung, 25, 271–283.
Lee, D., Seo, H., & Jung, M. W. (2012). Neural basis of
Lee, Y. S., Menschen, P., Frost, C., Hanke, M., & Granger, R. (2011).
Investigation of melodic contour processing in the brain
using multivariate pattern-based fMRI. Neurobild, 57,
293–300.
Lerdahl, F., & Jackendoff, R. S. (1996). A generative theory
of tonal music. Cambridge, MA: MIT Press.
Streuer, D. J., & Tirovolas, A. K. (2009). Current advances in the
cognitive neuroscience of music. Annalen von New York
Akademie der Wissenschaften, 1156, 211–231.
Lohrenz, T., McCabe, K., Camerer, C. F., & Montague, P. R.
(2007). Neural signature of fictive learning signals in a
sequential investment task. Verfahren des Nationalen
Akademie der Wissenschaften, USA., 104, 9493–9498.
Lopez-Barroso, D., de Diego-Balaguer, R., Cunillera, T., Camara, E.,
Münte, T. F., & Rodriguez-Fornells, A. (2011). Language
learning under working memory constraints correlates with
microstructural differences in the ventral language pathway.
Hirnrinde, 21, 2742–2750.
Lopez-Paniagua, D., & Seger, C. A. (2011). Interactions within
and between corticostriatal loops during component processes
of category learning. Zeitschrift für kognitive Neurowissenschaften, 23,
3068–3083.
Louie, P., Li, H. C., & Schlaug, G. (2011). White matter integrity
in right hemisphere predicts pitch-related grammar learning.
Neurobild, 55, 500–507.
Maess, B., Kölsch, S., Gunter, T. C., & Friederici, A. D. (2001).
Musical syntax is processed in Brocaʼs area: An MEG study.
Naturneurowissenschaften, 4, 540–545.
Maidhof, C., & Kölsch, S. (2011). Effects of selective attention
on syntax processing in music and language. Zeitschrift für
Cognitive Neuroscience, 23, 2252–2267.
Makuuchi, M., Bahlmann, J., Anwander, A., & Friederici, A. D.
(2009). Segregating the core computational faculty of
human language from working memory. Proceedings of
the National Academy of Sciences, USA., 106, 8362–8367.
Meyer, L. (1989). Style and music: Theory, Geschichte, Und
ideology. Chicago: University of Chicago Press.
Montag, C., Reuter, M., & Axmacher, N. (2011). How oneʼs
favorite song activates the reward circuitry of the brain:
Personality matters! Behavioural Brain Research, 225,
511–514.
Mueller, K., Mildner, T., Fritz, T., Lepsien, J., Schwarzbauer, C.,
Schroeter, M. L., et al. (2011). Investigating brain response
to music: A comparison of different fMRI acquisition
schemes. Neurobild, 54, 337–343.
Nee, D. E., Kastner, S., & Braun, J. W. (2011). Funktional
heterogeneity of conflict, Fehler, task-switching, Und
unexpectedness effects within medial prefrontal cortex.
Neurobild, 54, 528–540.
Opitz, B., & Friederici, A. D. (2007). Neural basis of processing
sequential and hierarchical syntactic structures. Human
Brain Mapping, 28, 585–592.
Parsons, L. M., Petacchi, A., Schmahmann, J. D., & Bower, J. M.
(2009). Pitch discrimination in cerebellar patients: Beweis
for a sensory deficit. Gehirnforschung, 1303, 84–96.
Patel, A. D. (2003). Language, Musik, syntax and the brain.
Naturneurowissenschaften, 6, 674–681.
Patterson, R. D., Uppenkamp, S., Johnsrude, ICH. S., &
Griffiths, T. D. (2002). The processing of temporal pitch
and melody information in auditory cortex. Neuron, 36,
767–776.
Pearce, M. T., Ruiz, M. H., Kapasi, S., Wiggins, G. A., &
Bhattacharya, J. (2009). Unsupervised statistical learning
underpins computational, behavioural, and neural
manifestations of musical expectation. Neurobild, 50,
302–313.
reinforcement learning and decision making. Annual Review
der Neurowissenschaften, 35, 287–308.
Perani, D., Saccuman, M. C., Scifo, P., Awander, A., Spada, D.,
Baldoli, C., et al. (2011). Neural language networks at birth.
1076
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 7
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
T
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Verfahren der Nationalen Akademie der Wissenschaften, USA.,
108, 16056–16061.
Peretz, ICH., Brattico, E., Järvenpää, M., & Tervaniemi, M. (2009).
The amusic brain: In tune, out of key, and unaware. Gehirn,
132, 1277–1286.
Peterson, E. J., & Seger, C. A. (in press). Many hats: Intra-trial
and reward-level dependent BOLD activity in the striatum
and premotor cortex. Journal of Neurophysiology.
Poldrack, R. A., & Mumford, J. A. (2009). Independence in
ROI analysis: Where is the voodoo? Social Cognitive and
Affective Neuroscience, 4, 208–213.
Roebroeck, A., Formisano, E., & Goebel, R. (2005). Mapping
directed influence over the brain using Granger causality
and fmri. Neurobild, 25, 230–242.
Rogalsky, C., Rong, F., Saberi, K., & Hickok, G. (2011).
Functional anatomy of language and music perception:
Temporal and structural factors investigated using
functional magnetic resonance imaging. Zeitschrift für
Neurowissenschaften, 31, 3843–3852.
novelty and subjective judgments. Neurobild, 55,
1739–1753.
Seger, C. A., Peterson, E. J., Cincotta, C. M., Lopez-Paniagua, D.,
& Anderson, C. W. (2010). Dissociating the contributions of
independent corticostriatal systems to visual categorization
learning through the use of reinforcement learning modeling
and Granger causality modeling. Neurobild, 50, 644–656.
Schmied, S. M., Müller, K. L., Salimi-Khorshidi, G., Webster, M.,
Beckmann, C. F., Nichols, T. E., et al. (2011). Netzwerk
modelling methods for fMRI. Neurobild, 54, 875–891.
Stewart, L., Overath, T., Warren, J. D., Foxton, J. M., & Griffiths,
T. D. (2008). fMRI evidence for a cortical hierarchy of pitch
pattern processing. Plos One, 3, e1470.
Stewart, L., von Kriegstein, K., Warren, J. D., & Griffiths, T. D.
(2006). Music and the brain: Disorders of musical listening.
Gehirn, 129, 2533–2553.
Summerfield, C., & Egner, T. (2009). Expectation (Und
attention) in visual cognition. Trends in den Kognitionswissenschaften,
13, 403–409.
Salimpore, V. N., Benovoy, M., Larcher, K., Dagher, A., &
Sutton, R. S., & Barto, A. G. (1998). Reinforcement learning:
Zatorre, R. J. (2011). Anatomisch unterschiedliche Dopaminausschüttung
während der Vorfreude und dem Erleben von Höhepunktgefühlen
Musik. Naturneurowissenschaften, 14, 257–262.
Santi, A., & Grodzinsky, Y. (2010). fMRI adaptation dissociates
syntactic complexity dimensions. Neurobild, 51, 1285–1293.
Schiffer, A. M., & Schubotz, R. ICH. (2011). Caudate nucleus signals
for breaches of expectation in a movement observation
paradigm. Grenzen der menschlichen Neurowissenschaften, 5, 38.
Schlaug, G., Marchina, S., & Norton, A. (2009). Evidence for
plasticity in white-matter tracts of patients with chronic Brocaʼs
aphasia undergoing intense intonation-based speech therapy.
Annalen der New York Academy of Sciences, 1169, 385–394.
Schulze, K., Zysset, S., Mueller, K., Friederici, A. D., & Kölsch, S.
(2010). Neuroarchitecture of verbal and tonal working
memory in nonmusicians and musicians. Menschliches Gehirn
Mapping, 32, 771–783.
Schwartze, M., Keller, P. E., Patel, A. D., & Kotz, S. A. (2011).
The impact of basal ganglia lesions on sensorimotor
synchronization, spontaneous motor tempo, und das
detection of tempo changes. Behavioural Brain Research,
216, 685–691.
Seger, C. A. (2008). How do the basal ganglia contribute to
categorization? Their roles in generalization, response
Auswahl, and learning via feedback. Neurowissenschaften und
Bioverhaltensberichte, 32, 265–278.
Seger, C. A. (2009). The involvement of corticostriatal loops
in learning across tasks, Spezies, and methodologies.
In H. J. Groenewegen, P. Voorn, H. W. Berendse, A. B. Mulder,
A. R. Cools (Hrsg.), Basal ganglia IX: Proceedings of
the International Basal Ganglia Society (S. 25–40).
New York, New York: Springer.
Seger, C. A., Dennison, C. S., Lopez-Paniagua, D., Peterson, E. J.,
& Roark, A. A. (2011). Dissociating hippocampal and basal
ganglia contributions to category learning using stimulus
An introduction. Cambridge, MA: MIT Press.
Talairach, J., & Tournoux, P. (1998). Koplanare Stereotaxie
Atlas des menschlichen Gehirns: 3-Dimensional proportional
system—An approach to cerebral imaging. New York: Thieme.
Thaut, M. H., Stephan, K. M., Wunderlich, G., Schicks, W.,
Tellmann, L., Herzog, H., et al. (2009). Distinct cortico-
cerebellar activations in rhythmic auditory motor
synchronization. Kortex, 45, 44–53.
Tillman, B. (2008). Music Cognition: Learning, Wahrnehmung,
Expectations. In R. Kronland-Martinet, S. Ystad, and K. Jensen
(Hrsg.), CMMR 2007, LNCS 4969, S. 11–33.08
Tritsch, N. X., & Sabatini, B. L. (2012). Dopaminergic
modulation of synaptic transmission in cortex and
striatum. Neuron, 76, 33–50.
Wow, P., & Kringelbach, M. L. (2010). The pleasure of making
sense of music. Interdisciplinary Science Reviews, 35,
166–182.
Wow, P., Wallentin, M., Mouridsen, K., Ostergaard, L., &
Roepstorff, A. (2011). Tapping polyrhythms in music activates
language areas. Neurowissenschaftliche Briefe, 494, 211–216.
Wilson, S. M., Galantucci, S., Tartaglia, M. C., Rising, K., Patterson,
D. K., Henry, M. L., et al. (2011). Syntactic processing
depends on dorsal language tracts. Neuron, 72, 397–403.
Yeterian, E. H., & Pandya, D. N. (1998). Corticostriatal connections
of the superior temporal region in rhesus monkeys. Zeitschrift
of Comparative Neurology, 399, 384–402.
Zacks, J. M., Kurby, C. A., Eisenberg, M. L., & Haroutunian, N.
(2011). Prediction error associated with the perceptual
segmentation of naturalistic events. Zeitschrift für Kognition
Neurowissenschaften, 23, 4057–4066.
Zatorre, R. J., Chen, J. L., & Penhune, V. B. (2007). When the
brain plays music: Auditory-motor interactions in music
perception and production. Nature Reviews Neurowissenschaften,
8, 547–558.
Seger et al.
1077
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
7
2
5
1
/
0
7
6
/
2
1
1
0
9
6
4
2
5
/
2
1
8
7
7
7
Ö
9
C
1
N
2
_
8
A
/
_
J
0
Ö
0
C
3
N
7
1
_
A
P
_
D
0
0
B
3
j
7
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
.
T
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1