Capturing Cognitive Events Embedded in the Real World
Using Mobile Electroencephalography and Eye-Tracking
Simon Ladouce1
, Magda Mustile2
, Magdalena Ietswaart2, and Frédéric Dehais1,3
Abstrakt
■ The study of cognitive processes underlying natural behav-
iors implies departing from computerized paradigms and artifi-
cial experimental probes. The present study aims to assess the
feasibility of capturing neural markers (P300 ERPs) of cognitive
processes evoked in response to the identification of task-
relevant objects embedded in a real-world environment. To this
end, EEG and eye-tracking data were recorded while partici-
pants attended stimuli presented on a tablet and while they
searched for books in a library. Initial analyses of the library data
revealed that P300-like features shifted in time. A Dynamic Time
Warping analysis confirmed the presence of P300 ERP in the
library condition. Library data were then lag-corrected based
on cross-correlation coefficients. Zusammen, these approaches
uncovered P300 ERP responses in the library recordings. Diese
findings highlight the relevance of scalable experimental
designs, joint brain and body recordings, and template-
matching analyses to capture cognitive events during natural
behaviors. ■
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
9
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
EINFÜHRUNG
The emergence of mobile brain and body imaging (MoBI;
Gramann et al., 2011) research methods provides the
unprecedented opportunity to depart from artificial
laboratory-based settings to study cognitive processes
directly in real-world environments (De Vos, Gandras, &
Debener, 2014; Gramann, Jung, Ferris, Lin, & Makeig,
2014; Makeig, Gramann, Jung, Sejnowski, & Poizner,
2009). Over the last decade, technical advances have been
made toward the miniaturization of sensors, improving
the portability of research-grade body and neuroimaging
hardware (Mcdowell et al., 2013), thus allowing to record
brain data outside of the laboratory over long periods
(Hölle, Meekes, & Bleichner, 2021). More pointedly,
mobile EEG and mobile eye-tracking (E-T) open new
research avenues to better understand how people think
and act in the real world. The exciting prospects offered by
the exploitation of such mobile research methods have
sparked interest in the development of novel signal pro-
cessing approaches (Reis, Hebenstreit, Gabsteiger, von
Tscharner, & Lochmann, 2014). Taken together, diese
developments enable to investigate human cognition
directly in naturalistic settings (Ladouce, Donaldson,
Dudchenko, & Ietswaart, 2017) to tackle fundamental
and applied questions across a wide range of research
fields such as sport science (Park, Fairweather, & Donaldson,
2015), architecture (Djebbara, Fich, & Gramann, 2019) Und
Stadtplanung (Birenboim, Helbich, & Kwan, 2021), neu-
roergonomics (Gramann et al., 2021; Dehais, Karwowski,
1Institut Supérieur de l’Aéronautique et de l’Espace (ISAE),
Toulouse, Frankreich, 2University of Stirling, Großbritannien,
3Drexel University, Philadelphia, PA
& Ayaz, 2020; Ayaz & Dehais, 2018), spatial navigation
(Do, Lin, & Gramann, 2021; Miyakoshi, Gehrke, Gramann,
Makeig, & Iversen, 2021), perception of art, architecture
and neuroaesthetics (Djebbara, Jensen, Parada, & Gramann,
2022; King & Parada, 2021; Djebbara, Fich, Petrini, &
Gramann, 2019), and the development of assessment
and rehabilitation methods for neurocognitive disorders
(Lau-Zhu, Lau, & McLoughlin, 2019; Kranczioch, Zich,
Schierholz, & Sterr, 2014). As elegantly articulated by
Parada (2018), the overarching challenges lying ahead of
the MoBI approach to reach its full potential imply a pro-
gressive transition from highly controlled laboratory set-
tings to the study of cognitive phenomena in real-world
environments with high ecological validity. Initiating
such an incremental approach, a series of influential
out-of-laboratory studies have revisited experimental par-
adigms commonly used in neuroimaging research and
performed them in naturalistic contexts. The following
sections present this body of research that established
the foundations upon which the present study is based.
Measuring Brain Activity during Motion and
Outside of the Laboratory
In a seminal study, Gramann, Gwin, Bigdely-Shamlo, Ferris,
and Makeig (2010) revealed that transient brain responses
to the presentation of stimuli could be extracted from sur-
face EEG data acquired while participants walked on a
treadmill. Genauer, the authors examined the
impact of walking speed (Stehen, walking, and walking
briskly) on the P300 component amplitude, which is a
widely studied feature of EEG signals whose robustness
has established it as a gold standard of EEG research
© 2022 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 International (CC BY 4.0) Lizenz.
Zeitschrift für kognitive Neurowissenschaften 34:12, S. 2237–2255
https://doi.org/10.1162/jocn_a_01903
(Polich, 2007). The P300 component is a positive deflection
in the time domain of the EEG signal occurring 300 ms
after the presentation of infrequent or task-related stimuli
(typically within the frame of an oddball paradigm) reflec-
tive of selective attention processes. These properties
established the P300 as a relevant measure to assess the
validity and quality of data acquired while participants were
in motion or went outside of the laboratory.
Taking the EEG outside of the laboratory, Debener,
Minow, Emkes, Gandras, and de Vos (2012) demonstrated
the feasibility of recording the P300 component elicited
through an auditory oddball paradigm when participants
were walking outside versus sitting inside. The neural
responses typically elicited by the presentation of target
auditory stimuli were observed in both experimental con-
ditions although they were attenuated in the outdoor-
walking condition. In a follow-up study (De Vos et al.,
2014), the authors controlled for the environmental factor
by having the participants perform the same auditory odd-
ball task while sitting and walking outdoor. Similar to the
previous study, an attenuation of the P300 effect was
observed for the walking condition, which was interpreted
as reflecting either a lower signal-to-noise ratio, potentially
related to the presence of motion artifacts contaminating
the walking data, or a reallocation of attentional resources
during walking. These early studies were hinting toward
important distinctions between how the mind works
under artificial and natural conditions. Circumventing
signal-to-noise issues relative to gait-related artifacts, zwei
cycling studies (Scanlon, Redman, Kuziek, & Mathewson,
2020; Scanlon, Townsend, Cormier, Kuziek, & Mathewson,
2019; Zink, Hunyadi, Huffel, & Vos, 2016) confirmed
that the attenuation previously reported was partly attrib-
utable to the physical activity related to cycling but also,
and more importantly, to the higher cognitive demands
of being outdoor. Ladouce, Donaldson, Dudchenko, Und
Ietswaart (2019) further specified the nature of the reallo-
cation of cognitive resources underlying the P300 attenu-
ation by demonstrating that it is not the locomotor
demands themselves that take cognitive resources away
from the task. In der Tat, this research pinpointed that it is
the displacement through space that is substantially taxing
in terms of cognitive resources. The reallocation of cogni-
tive resources is therefore because of the increased flow of
vestibular and visual information that needs to be pro-
cessed during locomotion. Consistently, Liebherr et al.
(2021) reported a reduction of P300 amplitude when stu-
dents performed an oddball task while finding their way
through a university campus as compared with simply
walking around a sports field, highlighting the cost of inte-
grating sensorimotor information. Ähnlich, two studies
conducted in real-flight conditions using a passive
(Dehais, Rida, et al., 2019) and active (Dehais, Duprès,
et al., 2019) auditory oddball paradigms disclosed lower
P300 amplitude when participants faced challenging flying
conditions involving an increased flow of visual informa-
tion to be processed. The MoBI approach has further been
applied to investigate embodied aspects of perception
and attentional processes during natural movement
(Cao, Chen, & Haendel, 2020; Cao & Händel, 2019; Reiser,
Wascher, & Arnau, 2019; Benjamin, Wailes-Newson, Ma-
Wyatt, Bäcker, & Wade, 2018; Schmidt-Kassow, Heinemann,
Abel, & Kaiser, 2013), as well as learning and memory
(Schmidt-Kassow, Deusser, et al., 2013). Taken together,
these findings further underlined that cognitive functions
are altered when people are immersed in an ever-
changing, dynamic, and complex environment.
To date, out-of-the-laboratory studies have mainly
resorted to the presentation of artificial stimuli through
computerized paradigms (z.B., visual or auditory oddball
tasks), interactions with artificial apparatus taking place
within highly controlled environments, or even experi-
mental protocols characterized by the repetition of proto-
typical behaviors. An important step toward realizing the
vision of studying human cognition in the real world
would imply to progressively transitioning from comput-
erized paradigms to the study of cognitive processes in
relation to embodied experiences that are grounded in
the real world. Such a goal remains challenging as the
study of cognitive phenomena in the real world prevents
the use of external and artificial experimental probes. In
the absence of these experimental markers, Fragen
such as when do cognitive processes happen and how to
timestamp and segment the data accordingly become
nontrivial issues that need to be addressed to accurately
extract cognitive events. In contrast to computerized par-
adigms in which the timing of cognitive events is dictated
by the course of the experiment and can therefore be
derived from event markers with very high temporal accu-
racy, the definition of cognitive events in the real world
poses several conceptual and technical questions.
Capturing Cognitive Events in the Real World
A relevant approach to segment and extract meaningful
information from continuous brain recordings is to con-
textualize the data based on physiological and behavioral
data recorded simultaneously. An example of such a mul-
timodal approach can be found in a study carried out by
Mavros, Austwick, and Smith (2016) in which Global
Positioning System (GPS) tracking and brain data were
combined to study spatial perception and cognitive expe-
riences related to different types of urban environments.
Banaei, Hatami, Yazdanfar, and Gramann (2017) used vir-
tual reality environments and mobile EEG to study the
impact of architectural design on spatial representations
when individuals walked through them. Durch Gebrauch von
of head orientation information provided by the virtual
reality system, the brain signals could be segmented to
extract brain dynamics related to the embodied experi-
ence of the virtual environment. Another example of con-
textual information being used to retrieve experimental
events from continuous brain imaging recordings is illus-
trated by Mustile et al.’s (2021) study in which infrared
2238
Zeitschrift für kognitive Neurowissenschaften
Volumen 34, Nummer 12
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
9
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
motion sensors were placed in a real-world environment
to detect the passage of individuals over obstacles and
extract neural dynamics reflecting motor planning. Hölle,
Blum, Kissner, Debener, and Bleichner (2022) imple-
mented a microphone and a dedicated processing pipe-
line to detect the onset of natural sounds and lock the
associated EEG analyses to unveil auditory attention pro-
Prozesse. In a series of studies looking at spatial knowledge
acquisition in natural environments (Wunderlich & Gramann,
2021), the experimenter was following the participants
navigating through an urban environment and encoded
the timing of experimental events manually through a
smartphone interface that was synchronized to the EEG
System. These manually encoded timestamps, although
arguably not of the highest temporal precision, provide
nevertheless a basis for the extraction of experimental
Veranstaltungen. The accuracy in the definition of experimental
event timings, Jedoch, has important implications for
later EEG analyses. In der Tat, the millisecond scale of ERP
components makes their analysis particularly sensitive to
temporal lags. The presence of variance across trials in the
latency of the responses will likely result in a smearing of
the averaged ERP waveform amplitude or the contamina-
tion of ERP components at neighboring latencies.
In natural settings, first-person video recording coupled
with eye movements that can be both recorded using
head-mounted E-T glasses can serve as contextual refer-
ence points to segment and analyze the EEG signal to cap-
ture neural dynamics of visual processing. Erste, the video
recording can be used to flag and label experimental
events in the continuous EEG data by providing informa-
tion about the onset and end of a visual search task. Der
E-T data provide a second layer of contextual information
to the scene capture, increasing the temporal resolution
of the experimental event segmentation. In der Tat, Die
superposition of gaze dynamics to the video recording
allows to extract temporal information about the timing
of visual events more precisely (Hayhoe & Ballard, 2005).
Gaze dynamics are composed of a sequence of discrete
fixations whose features (z.B., Dauer, pattern, previous
saccadic distance) provide information about the timing
and depth of visual and attentional processes. The timing
information of the initial fixations on experimental
objects to extract fixation-related potentials (FRPs; Baccino
& Manunta, 2005) has been widely adopted to investigate
neural underpinnings of reading and visual search.
Applying FRP analysis approach to free viewing visual
search paradigm, Brouwer, Reuderink, Vincent, van Gerven,
and van Erp (2013) demonstrated that the P300 ERP com-
ponent can be used to infer whether participants are look-
ing at target or nontarget stimuli. Kaunitz et al. (2014) Auch
observed the emergence of sensory and attentional com-
ponents in the FRP associated with the detection of target
faces in a scene crowded with distractor faces. Der
authors further contrasted the free visual search task to
a control fixation task and revealed differences in terms
of the topography and latency of the P300 component.
The free viewing paradigm elicited P300 responses that
were most prominent over centro-parietal electrode sites,
whereas the traditional oddball paradigm exhibited an ini-
tial earlier peak at frontal sites followed by later peaks
over parietooccipital sites. These results further demon-
strated that the P300 component is robustly elicited upon
target detection during visual exploration of natural
scenes. Kamienkowski, Ison, Quiroga, and Sigman
(2012) reported similar FRP components in a free viewing
search task and a replay task during which individual ele-
ments of the scene are presented as discrete sequences. Es
Ist, Jedoch, important to note that in the context of free
viewing search task paradigms, participants are typically
instructed to keep fixating at the target object when they
identify them for an extended period of time. This prac-
tice aims to facilitate later ERP analyses by avoiding eye
movements artifacts contaminating the EEG signals
time-locked to the fixations on targets. Applying such
Methoden, Roberts et al. (2018) and Soto et al. (2018)
explored neural markers of faces and economic value pro-
cessing related to the viewing of visual elements (pictures
presented on panels) placed in the real-world using
mobile E-T and wireless EEG. In both studies, the panels
comprised a central fixation cross to which participants
had to return their gaze to after looking at each individual
element for a minimum of a few seconds. Despite the arti-
ficial nature of the task, these studies demonstrated the
feasibility of capturing P300 FRP in a real-world environ-
ment. It however remains unclear whether such approach
applies in the context of a naturalistic behavior.
Aim of the Present Study
The present proof-of-concept study assesses the feasibility
of capturing neural markers of visual processing of objects
embedded in (d.h., being an integral part of ) a real-world
Umfeld. To achieve this, the present study applies
the concept of scalable experimental design (Parada,
2018; Ayaz & Dehais, 2018) according to which similar
cognitive phenomena are studied over a spectrum of
experiments ranging from highly controlled to naturalis-
tic environments. Inspired by this approach, the present
study contrasts two experimental conditions. Der erste
condition consists of a classic computerized paradigm
to elicit neural responses related to the processing of
visual information. For this purpose, abstract visual stim-
uli are presented on a screen and participants are
instructed to count the number of occurrences of a cer-
tain type of stimuli (d.h., targets). The artificial (als
opposed to natural) elements of this condition make it
far removed from experiences taking place in the real
Welt. Im Gegensatz, the second condition maximizes eco-
logical validity (d.h., the applicability of research findings
to real-life contexts) by getting as close as possible to the
recording of a natural behavior taking place in the real
Welt. Zu diesem Zweck, the second condition consists of
the performance of a scripted (semistructured) Aber
Ladouce et al.
2239
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
9
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
realistic behavior (d.h., searching for a book) grounded in
a real-world environment (d.h., a library). Through this
leap across the ecological validity continuum, differences
related to embodied experiences can be explored
whereas conceptual and methodological gaps pertaining
to the study of cognitive processes in the real-world can be
uncovered. An elegant example of how to fill the ecologi-
cal validity gap is provided by the work of Chen, Cao, Und
Haendel (2022), in which translucent augmented reality
glasses were used to superpose visual stimuli on a dynam-
ically changing real-world environment as participants
walked around. The authors report an amplification of
early sensory responses (N1 component) to visual stimuli
during locomotion. Interessant, later EEG dynamics
(N2pc ERP component and alpha oscillations) reflective
of cognitive processes related to stimulus discrimination
were not altered by the act of walking in the real world.
By enabling the presentation of experimental stimuli
superposed to participants’ natural field of view in a timely
manner, augmented reality technology offers a practical
solution to retain experimental control while studying
visual experiences in real-world environments. Im Gegensatz,
the present study sets out to address whether neural
markers of visual cognitive processes elicited through
computerized paradigms are also naturally present (d.h.,
embedded) in the real world. This aim not only implies
methodological capacities in terms of recording mean-
ingful neural data during real-world behaviors (welche
have been demonstrated by the aforementioned MoBI
body of research) but also novel solutions to extract
experiment-related neural signals from the continuous
recordings of dynamically and ever-changing environment
without resorting to the introduction of artificial/
extraneous stimuli (d.h., through computerized paradigms).
The visual search task in the library and its high degree of
ecological validity offer the opportunity to identify and
address the challenges related to the capture of cognitive
events as they occur naturally during real-world behaviors.
METHODEN
Teilnehmer
Twenty-four participants took part in the study. The partic-
ipants were exempt from any motor, visuell, and cognitive
Beeinträchtigung. All the participants were provided with
detailed information regarding the experimental protocol
and were introduced to EEG and E-T recording proce-
dures. Inclusion and exclusion criteria were checked
through the completion of a questionnaire by the partici-
Hose. The participants gave their written informed con-
sent to take part in the study. The study was reviewed by
a local research ethics committee, complied with data pro-
tection laws, and was carried out in accordance with the
principles delineated in the declaration of Helsinki. Der
order of the conditions (tablet, library) was counterbal-
anced across participants to control for potential biases
related to fatigue and training. Inconsistencies between
synchronization pulses timing sent to the EEG and E-T
data streams led to the exclusion of two data sets. Der
average gaze tracking proportion across participants was
über 80% for the remaining participants, with most of
the missing gaze data coinciding with displacements
across the library. Gesamt, these missing data points were
therefore not consequential with regard to analyses that
concerned periods during which participants were static
(d.h., standing in front of the library shelves). Jedoch,
three additional data sets were excluded from the study
because of their insufficient proportion of the E-T data
containing pupil position (42%, 55%, Und 58%). In those
data sets, a substantial portion of gaze data were missing
when the participants were scanning through the shelves.
In the absence of gaze information at key moments of the
experimental paradigm (d.h., when a participant locates a
target book cover), it was therefore impossible to retrieve
an estimation of experimental events timing. As a conse-
quence of the aforementioned technical issues, nur 19
out of the 24 initial recordings were included in the
reported analyses. The remaining number of data sets
complied with the minimal sample size of 16 individual
data sets estimated based on previous mobile P300 ERP
Studien (Ladouce et al., 2019; Reiser et al., 2019; Debener
et al., 2012).
Paradigm
Tablet Condition
Following a standard visual P300 elicitation oddball para-
digm, infrequent target stimuli were presented within a
series of frequent nontarget stimuli (at a 1:4 Verhältnis). Der
visual stimuli were presented on a Windows Surface tablet
positioned on a library shelf at 60-cm distance from partic-
ipants’ eye level as they were standing still. The target
stimuli consisted of red circles and nontarget stimuli were
blue squares of matching areas (circle diameter = 5.1 cm,
square length = 4.52 cm). daher, visual stimuli width
was maintained throughout the tablet condition at 4° of
visual angle. All stimuli were presented in the center of
the screen for a 200-msec period, which was followed
by an 800-msec interstimulus interval. The participants
were instructed to mentally count the number of target
stimuli presented. A total of 300 (260 nontargets, 40 tar-
gets) stimuli were presented through a Python-based pro-
gram operating on a 10.8-in. tablet (60-Hz refresh rate,
Windows 10). The communication rate of devices and
software used to send event markers to the amplifier
was measured through blackbox testing (BBTK v2 system
from Black Box ToolKit Ltd.). This approach measures
the magnitude and variance of the interval difference
between the local timing of stimulus onset on the exter-
nal device (tablet running the visual presentation pro-
Gramm) and the registration of the triggers received on
the amplifier end that will be interpreted as timestamps
2240
Zeitschrift für kognitive Neurowissenschaften
Volumen 34, Nummer 12
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
9
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
of experimental events in the EEG trace. The accuracy of
stimuli presentation timing was assessed over an hour-
long recording. The test revealed a 50-msec difference
between stimulus presentation onset and the EEG time-
stamp, with low variance throughout the recording (SD =
2 ms). Entsprechend, event marker latencies were cor-
rected to account for the measured delay by subtracting
50 ms (implemented at the beginning of the process-
ing pipeline).
Library Condition
The library condition consisted of a natural visual search
task and was designed as an analogy to the tablet para-
digm previously described. Teilnehmer (all native English
speakers) were instructed to search for 40 books (whose
titles were all written in modern English) in the library.
Target visual objects were the books that the participants
were instructed to find in the library, whereas nontarget
objects were defined as the four book covers preceding
the initial visual exploration of a target Book Cover 1. Jede
trial started at a specific location in the library where par-
ticipants received instructions regarding the title and the
location (aisle of the library where the bookshelf was
located, and on which shelve the book could be found)
of the target book on a sheet of paper that they carried
with them. Once a book was found, participants returned
to the starting position to pick up the next sheet provid-
ing them with instructions about the next book to find. In
contrast to the tablet condition in which distance from
stimuli presented on the tablet was fixed, the natural
exploration of the library was completely free of restric-
tionen. The lack of information regarding eye-to-stimulus
distance and its potential trial-to-trial (and even moment-
to-moment) variance complicates the computation of
visual angle measures for the tablet condition. Assuming
that participants maintained a distance ranging from
60 cm to 1 m away from the shelves while searching for
book covers, the visual angle can be estimated as a range
between 2°510 and 4°. This approximation, Jedoch, does
not allow for a valid definition of the different subfields
of central vision as further explained in the Discussion
section.
Several factors were considered for the selection of tar-
get books. All the target books selected were placed on
shelves that were at the eye level of the participants to
reduce the contamination of the EEG signal by artifacts
related to neck movements and other muscular activity.
Early works on visual attention have revealed the
bottom–up influences of low-level visual features (d.h.,
intensity, Kontrast, and edge density) on the initial visual
exploration of a scene (Peters, Iyer, Itti, & Koch, 2005).
The confrontation of computational models inspired by
this visual saliency hypothesis with experimental data
has later revealed that such bottom–up influences,
although partially accounting for the visual exploration
pattern, Sind, Jedoch, not sufficient to accurately predict
visual exploration within the frame of complex scenes
(Henderson, Brockmole, Castelhano, & Mack, 2007). Fur-
thermore, empirical evidence from scene perception and
visual search experiments highlighted the major contri-
bution of top–down processes (z.B., use of prior seman-
tic information to orient visual search) in how complex
scenes are explored and perceived (Birmingham, Bischof,
& Kingstone, 2009; Underwood, 2009). As both stimulus-
driven and cognitive-driven processes interact and influ-
ence how information embedded in complex scenes is
perceived and processed, several dispositions were taken
to ensure consistency in bottom–up and top–down influ-
ences across trials. The top–down influence was con-
trolled through the homogeneity of books within a
shelve in terms of their semantic field. In der Tat, Das
homogeneity does not favor top–down driven explora-
tion strategies such as parsing and skipping book covers
based on prior semantic information gathered (z.B., Die
target book is more likely to be surrounded by books of
the same semantic field). As a consequence of this con-
sideration, shelves with semantically homogeneous
books (d.h., related to the same lexical field) war
included in the experiment. Darüber hinaus, the classification
system adopted by the library used as an experimental
environment was not based on alphabetical order but
followed a systematic catalog arrangement (d.h., books
sorted in accordance to the domain and types of publica-
tionen). This nonalphabetical ordering makes the filtering
of content during the initial stage of a bookshelf explora-
tion more difficult. The systematic catalog system could
nevertheless be leveraged for semantic parsing of book-
shelf content, but it requires metaknowledge regarding
the classification system used itself and domain-specific
knowledge to navigate and parse effectively sections that
do not correspond with the title of the target book. Der
wide variety (Genre, types) of the books selected across
trials further discouraged the adoption of such domain-
specific search strategies by the participants over the
course of the experiment. To minimize the bottom–up
influences of objects whose visual properties make them
stand out from the rest of a visual scene, the size and
color of book covers were taken into consideration in
the design of the experiment. In der Tat, shelves containing
books whose covers were particularly salient were not
included in the experiment. The position of the book rel-
ative to the edge of a bookshelf was another factor taken
into consideration for the selection of target books.
In der Tat, a qualitative inspection of preliminary E-T data
confirmed that participants mainly scan through the
shelves using an initial reading-like approach (left to right
and top to bottom) as a default exploration strategy.
daher, to gather sufficient fixations on individual
books preceding the first fixation on the target book to
allow for an analogous analysis, the position of the target
book relative to the edges of the bookshelf was purpose-
fully central.
Ladouce et al.
2241
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
9
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
EEG Data Recording and Processing
EEG data were recorded from 32 sensors fitted in an elastic
cap following the International 10–20 system, die Waren
tethered to a portable amplifier (eego sports from
ANTNeuro) recording data at a sampling rate of 500 Hz
(with a 0.1- to 250-Hz on-line bandpass filter). The ampli-
fier was fitted in an ergonomic backpack carried by the
Teilnehmer. The data were initially referenced to channel
Cpz with the ground placed at the Afz electrode site. Elektr-
trode impedance was measured before each recording
session, and each channel was maintained below 5 kΩ
using electrode gel. EEG data were downsampled to
250 Hz, and mastoid electrodes (M1 and M2) were dis-
carded. The data were then rereferenced to the average
of all remaining electrodes.
Continuous EEG data from both recording conditions
were processed jointly using the EEGLAB (Delorme &
Makeig, 2004) open-source toolbox and custom MATLAB
scripts (Version R2019b 9.7.0, The MathWorks Inc.). Als
initial preprocessing step, the continuous data were visu-
ally examined and the portions of the EEG displaying
extreme levels of noise (z.B., channel disconnections)
were manually discarded. Following this manual data
rejection preprocessing, the processing pipeline was
divided into two stages. In the first stage, the data sets
were filtered with a low-pass filter of 20 Hz and a high-pass
filter of 1 Hz with a −6-dB cutoff and a filter order of 1650.
Dann, the continuous EEG was split into consecutive
epochs of 1 Sek. Epochs presenting abnormal values were
pruned based on standard statistical criteria (mehr als
3 SDs from the mean).
Following the initial filtering and removal of noisy data,
the first stage of artifact removal was carried out. Ein
extended infomax Independent Component Analysis
(ICA; Glocke & Sejnowski, 1995) was performed on the
remaining data, and the resulting independent compo-
nen (ICs) decomposition matrices were saved. In a sec-
ond stage, the IC features obtained during the first stage of
the processing procedure were back-projected to the orig-
inal filtered data. An automatic classification algorithm
(ICLabel) was used to classify ICs (Pion-Tonachini,
Kreutz-Delgado, & Makeig, 2019). The results of this clas-
sification were examined, and ICs identified as artifactual
(d.h., ocular and cardiac components) were confirmed
manually. The weights of ICs reflecting common artifacts
such as eye blinks, eye movements, and heartbeats were
subtrahiert. After this ICA-based data pruning, an average
von 58% (SD = 8.2%) of the initial ICs remained across par-
ticipants. This proportion of remaining components is in
line with the guidelines proposed by Klug and Gramann
(2021). The ICA-pruned continuous data sets were then
epoched around the onset of experimental events
(−2000 msec to 2000 ms). Epoched data were then split
into the experimental paradigm (tablet and library) Und
stimulus type (target and nontarget) conditions and base-
line corrected (the mean voltage recorded within the
200-msec prestimulus period was subtracted from the
signal for each electrode and each trial). Averaging across
epochs resulted in the obtention of ERP waveforms for
each condition. The P300 effect amplitude was computed
as the voltage difference (in microVolts) between target
and nontarget ERP waveforms within the a priori time
window ranging from 250 Zu 500 msec after stimulus
onset. The P300 latency was extracted based on the max-
imal value recorded within the a priori time window on a
single-trial basis.
Although not subjected to statistical analyses, Ereignis-
related spectral features were nevertheless examined.
Time–frequency decomposition of the epoched data were
performed through complex Morlet wavelet convolution.
Wavelet frequency ranged from 1 Zu 20 Hz in 38 linearly
spaced steps with the number of wavelet cycles increasing
aus 3 Zu 16 following a 0.8-step increase. Frequency-
specific power was baseline-corrected using a decibel
(dB) transform for each time point of the epoched data
relative to the mean spectral activity recorded during the
prestimulus period (−200 to 0 msec relative to stimulus
onset) on a single-trial basis. Relative power change was
averaged over time points within the data-driven time
window used for P300 ERP analysis (300–500 msec).
E-T Data Recording and Processing
Gaze dynamics were recorded using a portable Tobii Pro
Glasses 2 E-T system (Tobii Pro AB). E-T data were
acquired from four eye cameras tracking pupil position
and corneal reflection binocularly at a sampling rate of
100 Hz. Built-in parallax and slippage compensation
methods were performed to maintain E-T tracking accu-
racy during movement. The calibration procedure con-
sisted in presenting a target placed at 1-, 3-, and 5-m
distance from the participants to ensure reliable tracking
at different fixation depths. The E-T apparatus comprises
the camera-equipped glasses and a recording unit to
which the glasses were connected through HDMI. Der
recording unit was fitted in the backpack with the mobile
EEG amplifier to which it was connected through micro
USB to the 8-bit trigger input. A timestamp was gener-
ated by the E-T every 5 sec and sent to the EEG amplifier
for synchronization purposes. Before the study, the accu-
racy of the synchronization triggers has been extensively
tested over hours-long recordings. The delay between
recording systems remained below 10 msec and was con-
sistent throughout the testing recordings. The raw E-T
data were then reviewed visually, and periods character-
ized by poor tracking accuracy were recalibrated using
known fixation points (z.B., participants were instructed
to look at a fixation point at the beginning of each trial).
Missing gaze samples were interpolated (using a moving
median of five samples) if the gap between retrieved
samples was lesser than 75 ms, otherwise the samples
were considered lost. The proportion of gaze samples
retrieved throughout the recording (expressed in
2242
Zeitschrift für kognitive Neurowissenschaften
Volumen 34, Nummer 12
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
9
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
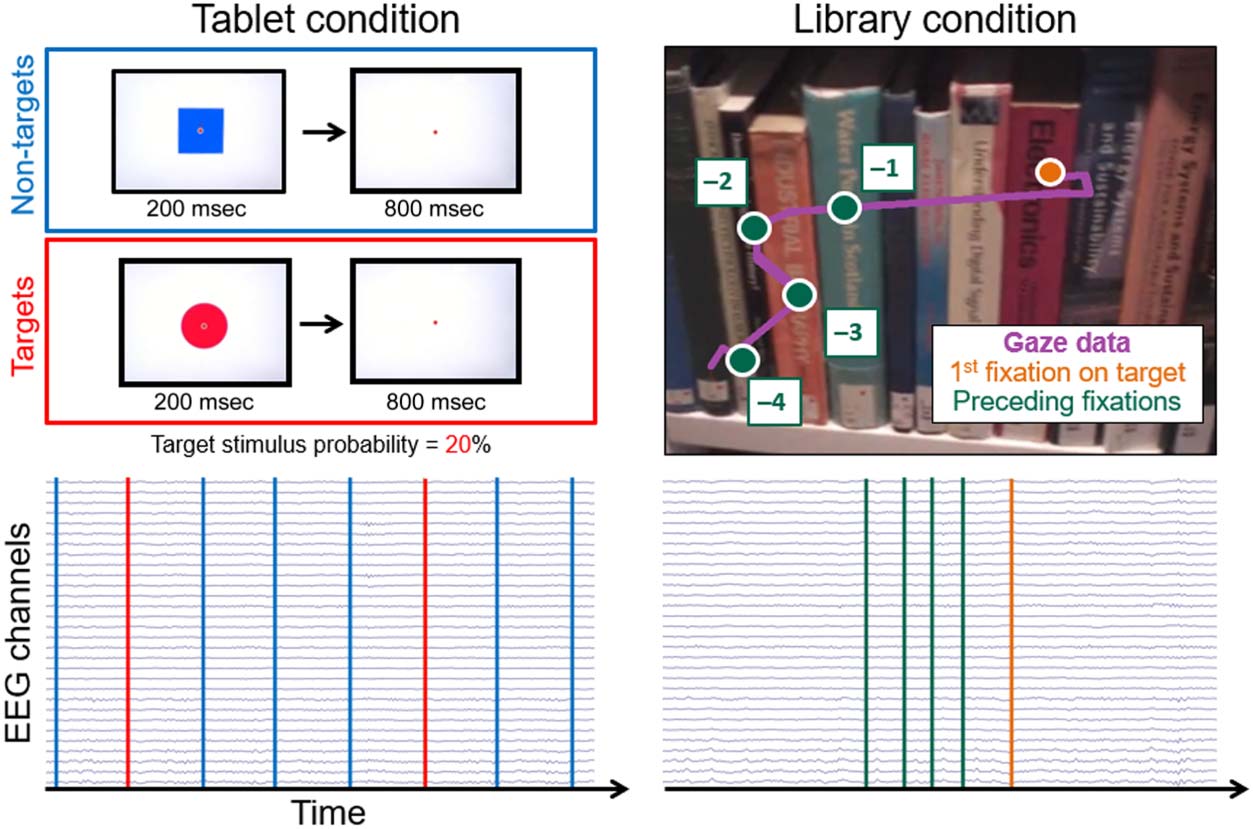
Figur 1. Example of the two
types of experimental events
(target and nontarget stimuli)
for both conditions (tablet on
the left and library on the right
Seite). For the tablet condition, A
series of discrete visual stimuli
were presented through a
computerized paradigm. Der
infrequent target stimuli were
red circles, and the frequent
stimuli were blue squares
(1:4 Verhältnis). The stimuli were
presented for 200 ms
followed by an 800-msec
interstimulus period. The onset
of visual stimuli was used to
define experimental events’
zeitliche Koordinierung. As can be observed from
the continuous 32 Kanäle
EEG data, event markers were
equally spaced over the time
Serie. An example of a participant’s gaze during the task is represented by the red dot. For the library condition, gaze data and first-person video
recording were reviewed by the experimenter. The experimental event timing was defined as the onset of a fixation on the target book cover for
target trials and as the first fixation on the four preceding book covers for the nontarget trials. The variability in terms of fixation and saccade duration
results in an irregular temporal distribution of visual events as denoted by the lower right plot.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
9
0
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Prozentsatz) was over 75% for the majority of partici-
Hose. Wie oben erwähnt, two outlying data sets had
to be excluded because of their low proportion of E-T
samples recorded. The continuous data were then sub-
jected to a noise reduction function based on a non-
weighted moving median filter with a window size of
three samples. A classification algorithm was then per-
formed on the raw E-T data to identify fixations. Der
built-in Tobii I-VT Fixation Filter was used with a velocity
(expressed in visual degrees per second) threshold of
30°/sec over 20-msec window length. Gaze samples
above the velocity threshold were classified as saccade
Proben. Short fixations lasting less than 50 msec were
discarded. Adjacent short fixations were merged when
their interfixation (saccade) duration was lower than
75 msec or that the visual angle difference between these
fixations was lower than 0.5°. Henderson and Luke
(2014) have reported that the mean fixation time was
around 250 msec during complex scene visual search
tasks and that this fixation duration, although prone to
intersubject variability, was stable within and between
sessions. Based on these findings, a lower threshold of
200 msec was used for the definition of visual fixations.
Event Extraction
The processed E-T data were then visually inspected by
the experimenter, and experimental events timings were
manually annotated using the Tobii Pro Lab software.
The onset of the initial fixation on a target book cover
was used as a timestamp for the definition of a target trial.
The timings of the onset of preceding fixations on four
distinct book covers were used to retrieve nontarget
experimental events. This approach to the definition of
the library experimental events timing was adopted to
allow for comparisons with the tablet condition in which
target stimuli were presented in the midst of nontarget
stimuli with a 1:4 Verhältnis. It should be noted that an auto-
matic solution for event extraction was available at the
time of the study. This approach, Jedoch, required plac-
ing QR-code probes in the vicinity of experimental objects.
The high contrast and odd nature of these probes with
regard to a library environment yield strong bottom–up
influences on visual attention. Not only those probes
would act as cues heavily orienting participants’ visual
exploration of the environment, but the number of probes
required to assign every potential experimental object
(book cover) made this solution inadequate within the
frame of the present experimental design (Figur 1).
Statistical Analysis
Statistical analyses were performed on the mean ampli-
tudes within the P300 time window (300–500 msec)
recorded at Pz electrode site where the P300 is most
prominent (Polich et al., 1997; Alexander et al., 1996).
Repeated-measures ANOVA and paired-samples t tests
were performed on the extracted amplitude features. To
ensure that parametric analysis was appropriate, a nor-
mality test was carried out to ensure that the data
followed a normal Gaussian distribution. Zusätzlich,
Holm-Bonferroni correction for multiple comparisons
was applied for all post hoc t tests. Partial eta squared
(η2) and Cohen’s d measures of effect sizes are reported
for ANOVA and t tests, jeweils.
Ladouce et al.
2243
ERGEBNISSE
P300 ERP Analyses
A repeated-measures ANOVA was carried out on ERP fea-
tures with the Experimental Paradigm (tablet, library) Und
Stimulus Type (target, nontarget) as factors. Post hoc
paired-samples t tests were carried out to explore the main
Effekte.
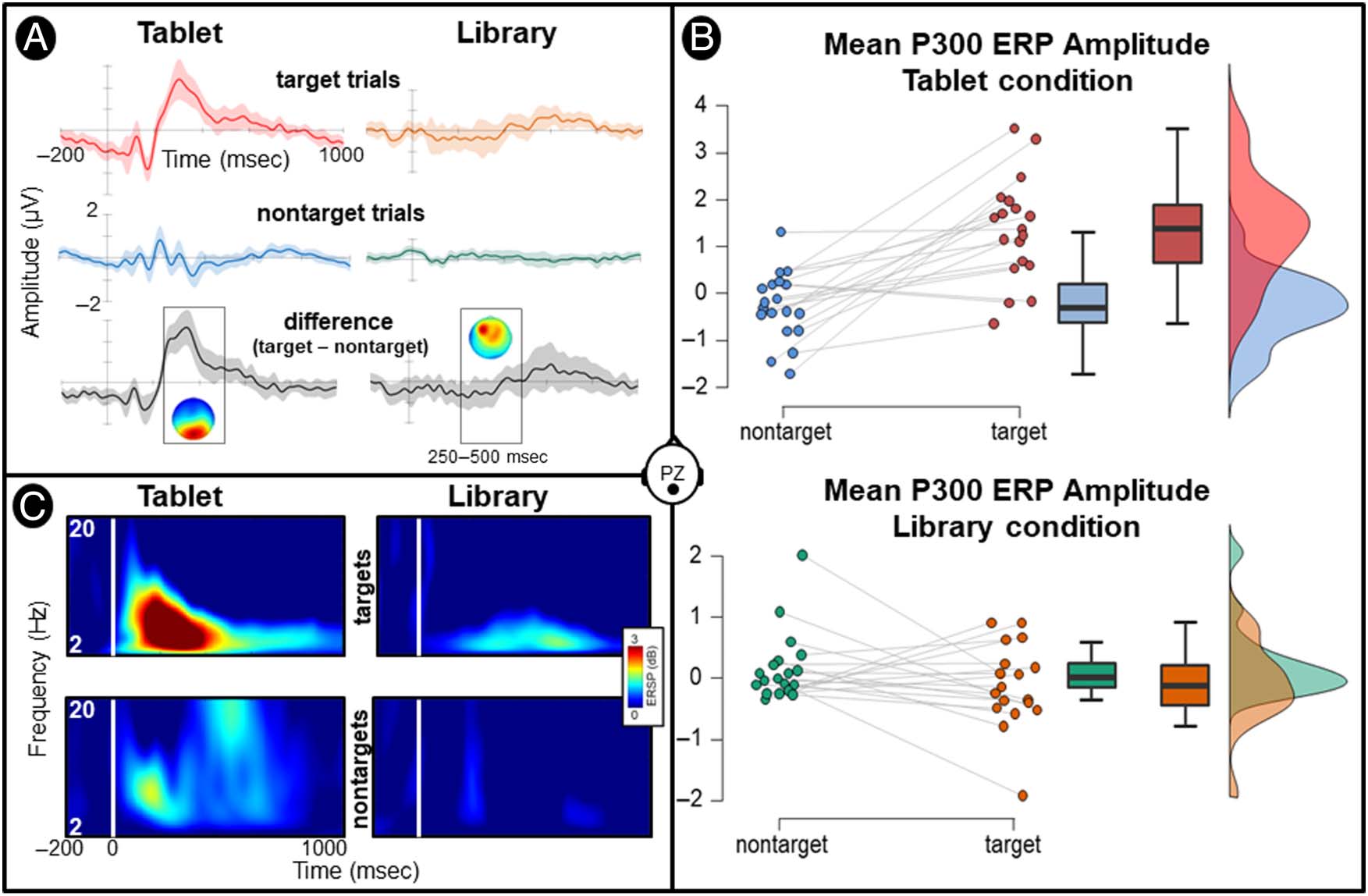
The repeated-measures ANOVA revealed that both the
paradigm, F(1, 18) = 8.98, P < .01, η2 = .33, and the stim-
ulus type, F(1, 18) = 17.62, p < .001, η2 = .49, had a main
effect on P300 ERP amplitude. Moreover, an interaction,
F(1, 18) = 19.98, p < .001, η2 = .52, between the two
factors was found.
Post hoc comparisons revealed that target stimuli elic-
ited P300 ERP responses of significantly higher amplitude
than nontarget stimuli for the tablet condition, t(18) = 5.2,
p < .001, Cohen’s d = 1.19, but not for the library condi-
tion, t(18) = 1.33, p = .209, Cohen’s d = .29. The present
results indicate that a P300 ERP response was consistently
elicited in the tablet condition, whereas the effect was not
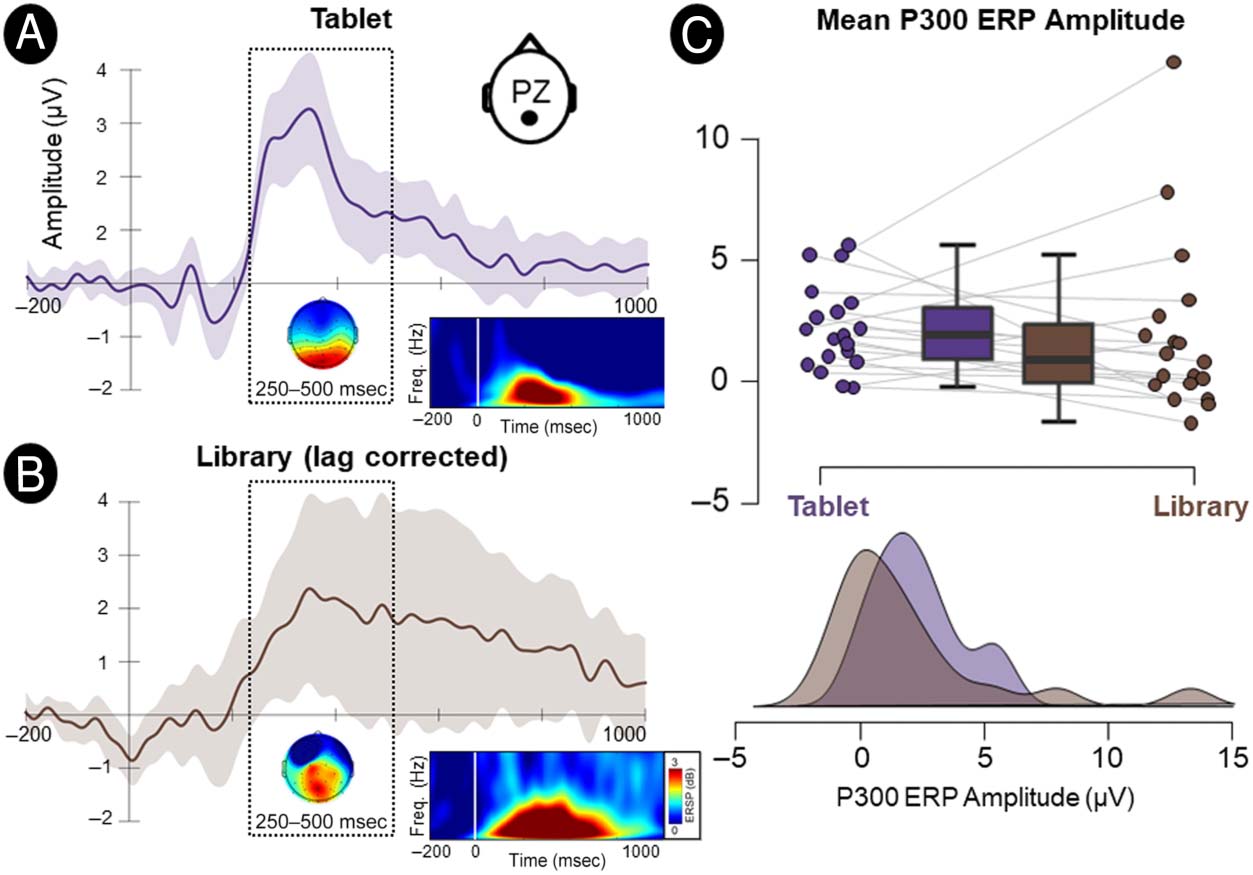
observed in the library data (as illustrated in Figure 2).
Interim Discussion
As discussed in the previous sections, there are many
unknown variables affecting the definition of the onset
of a cognitive event in the real world. The a priori
approaches based on gaze data (i.e., fixations on experi-
mental objects) may not coincide with the actual onset
of cognitive processing of that particular visual informa-
tion. The cognitive processes related to a visual fixation
may precede or follow the initial fixation. The temporal
gaps in either of those scenarios would introduce variance
in the latency of ERP responses, which would not survive
averaging processes. The inspection of gaze data provided
striking evidence that both scenarios (i.e., fixation-event
related potentials (fERP)-based definition of visual pro-
cesses onset being late or early) were commonly found
within single recordings and across participants.
Indeed, the first fixation on the target book may already
be relatively late with regard to the overall temporal course
of the visual processing of that information, which may
have already started when the information entered the
peripheral visual field. This possibility is illustrated by
the gaze pattern of the participants preceding target iden-
tification: The visual exploration strategy typically shifts
from an orderly scan of the book covers to a sudden bal-
listic saccade toward the target. The large angular distance
of these saccades further suggests that the book covers
present in the periphery are already being processed
semantically. In this scenario, the ERP responses related
to the visual processing of these target trials would pre-
cede the initial fixations and, therefore, the ERP would
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Grand average (n = 19) ERPs recorded at the parietal electrode (Pz). (A) Grand-average ERP waveforms elicited by target and nontarget
visual stimuli for both the tablet (left) and library (right) conditions. The difference ERP waveform (target − nontarget) is plotted below with a scalp
map presenting the topographical distribution of activity recorded within the P300 time window (250–500 msec). (B) Distribution of mean P300 ERP
amplitude recorded at Pz electrode across participants. (C) Time–frequency plots of mean event-related changes in spectral power (decibel scale)
elicited by target and nontarget stimuli for each experimental condition (tablet and library).
2244
Journal of Cognitive Neuroscience
Volume 34, Number 12
be temporally shifted in time in what is commonly called
the prestimulus period. This shift is particularly problem-
atic for the application of baseline correction approaches
that aim to detrend the data using this prestimulus period.
If the prestimulus period contains the signal of interest,
then subtracting it from later time-series signals would
introduce antagonist artifactual effects.
In contrast, all recordings contained several trials in
which the individuals visually explored the target book
cover without identifying them as targets and continued
their exploration of the shelves. The object may have only
been identified as a target after having been fixated on. In
that second scenario, using the fixation on the object as a
timestamp for the onset of visual processing introduces a
delay in the responses shifting the neural signals at a later
point in the time series. This second scenario is more
likely to occur when the target object is less salient
and/or there is more competing visual information in the
visual field such that the bottom–up influences of the non-
target objects counteract top–down strategies. Although
these “missed” trials were relatively infrequent (their num-
ber was too low to allow for a dedicated ERP analysis), aver-
aging them with actual target identification trials would
lower the signal-to-noise ratio of ERP responses of the lat-
ter. The strict definition of the first fixation on a target
book cover was therefore relaxed, and the fERP onset for
such trials was changed to the first fixation on the target
preceding their actual identification (i.e., the participant
stops scanning the shelves and returns to the starting
point).
Both of these phenomena are likely to occur in real-
world environments where the objects and their sur-
roundings’ visual features are variables. Not only the cover
of the target books may have been more or less salient, but
the density of the books on the shelves, visual properties
of the books’ covers (i.e., colors, width, orientation), the
lighting in different parts of the library, and other priming
effects through semantic association (e.g., shared lexical
field) induced by surrounding cover titles are as many fac-
tors that may affect the timing between early visual and
later attentional processing of target objects and their
visual exploration. Therefore, the naive fixation-based
approach for the timestamping of experimental events is
inherently limited to capturing accurately the onset of cog-
nitive processing of visual information embedded in the
real world.
Another source of temporal imprecision comes from
the relatively low sampling rate of the eye-tracker scene
camera. Indeed, the scene recording is captured at a rate
of 30 frames per second. There is therefore a 33-msec gap
between every frame. This gap means that the accuracy of
visual events timing achieved by reviewing the video
recording frame by frame is limited by the temporal reso-
lution of the scene capture. In addition to this temporal
variance, the gaze data superposed to the scene recording
is acquired at a higher rate (100 Hz) and is smoothed to
match both recordings. Considering the additional
degrees of freedom that apply to a fully mobile E-T system,
it is sensible to assume that the matching between the
scene recording and gaze data points may be subject to
some imprecision, especially during head movements
and even more so during whole-body movements. Any
incoherence between the data streams comes, however,
at the price of an additional 33-msec variance added on
top of the original 33 msec. The millisecond scale of ERP
components and the averaging process usually applied to
uncover the signal from background activity make them
particularly sensitive to subtle temporal variations. Taken
together, the aforementioned considerations suggest that
mobile E-T data, while providing contextual information
for the definition of experimental events, may, however,
not provide a temporal estimate sufficiently precise to per-
form ERP analyses. The direct consequence of any of these
sources of temporal imprecision is that there would be
important variations across trials in terms of ERP latencies,
essentially leading up to smearing effects (Ouyang,
Herzmann, Zhou, & Sommer, 2011) or even canceling out
potential ERP components through averaging procedures.
Although it appears plausible that the absence of ERP
components time-locked to the initial fixation on experi-
mental objects observed in the library condition may be
caused by intertrial latency variability, it is nevertheless
important to consider that such brain signals may simply
not be present during the library visual search task.
Indeed, the P300 ERP component could be an artificial
response evoked by computerized paradigms that do
not transfer to the real world. The former hypothesis
implies that ERP components would manifest at the
single-trial level but would be shifted in the time domain,
whereas the latter hypothesis implies the total absence of
such ERP components for the library condition. To eluci-
date these competing assumptions, single-trial ERP
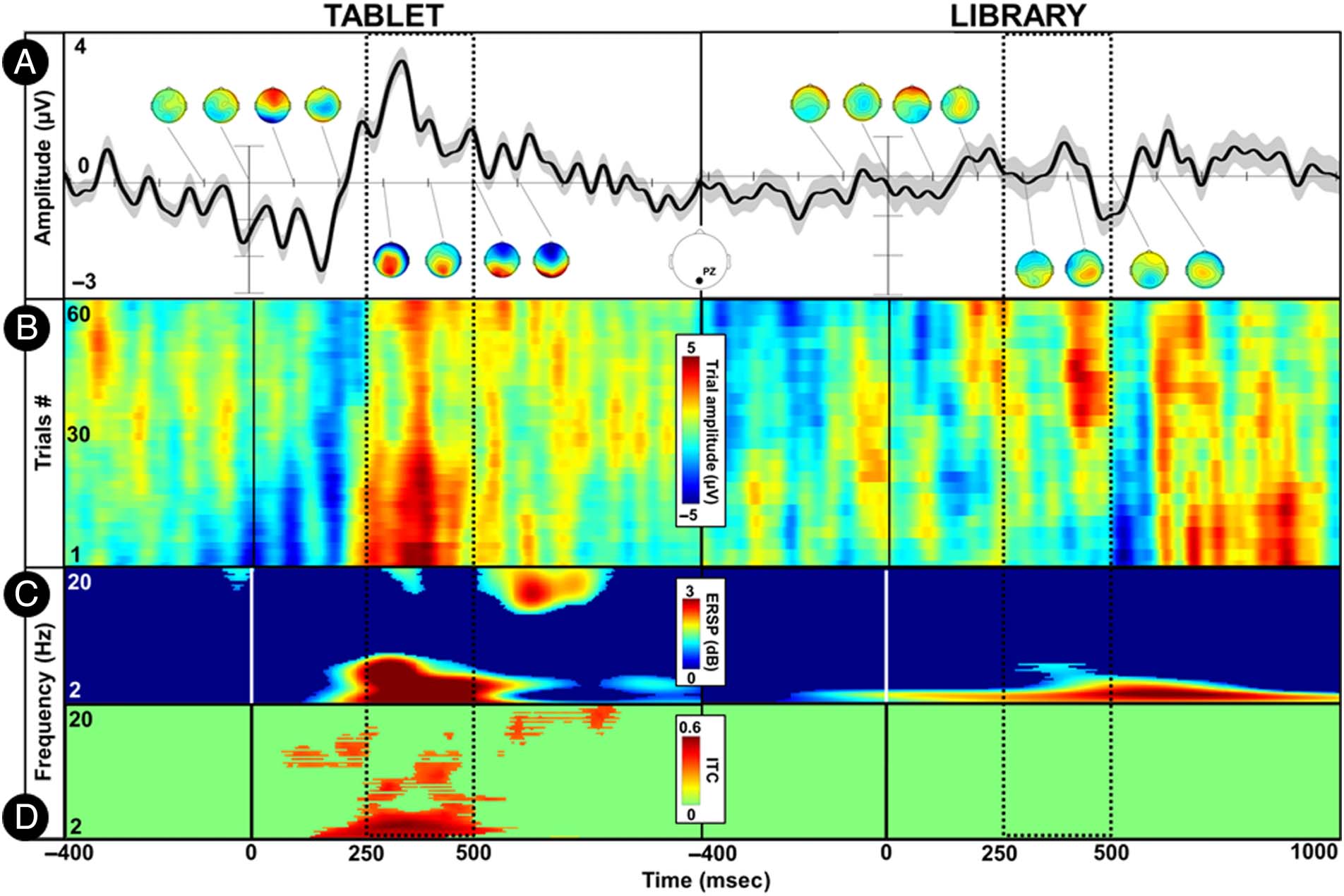
responses to target book covers were inspected over a
larger time around their fixation onset. As can be observed
in Figure 3, signals sharing spatial (parietal topographical
distribution) and spectral features (delta and theta band
activation) of the P300 ERP are present at the single-trial
level. The variance in the latency of such signals is, how-
ever, important, spanning across both the prefixation
and postfixation periods, with a wide temporal distribu-
tion over the latter. These observations suggest that the
P300 ERP response may be present in the library condition
recordings but have substantial variance in their latency.
The FRP approach applied to define the onset of a cogni-
tive event may not be valid in the context of the present
real-world data. It remains unknown whether the P300-like
signals observed at the single-trial level are effectively a
specific response to target stimuli. To address this ques-
tion, library epoched data of both target and nontarget
stimuli were compared with a subject-specific template
of P300 ERP response based on the tablet ERP average
waveform. A higher similarity between target waveforms
than between the template and nontarget stimuli would
provide further evidence, suggesting that the library target
Ladouce et al.
2245
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Illustrative single-subject (Participant 5) fERP and ERP channel data (Pz electrode) comparing temporal dynamics across the library and
tablet conditions. The dotted windows indicate the a priori time window commonly used to extract P300 ERP features in experiments using
computerized paradigms (250–500 msec). (A) The uppermost plots present the grand average ERP waveforms (shaded area indicates standard error)
around the onset of experimental events, which were defined as the first fixation on the target book cover for the library condition and the onset of
stimulus presentation for the tablet condition. A series of topographical scalp maps indicate the spatial distribution of ERP responses over time (from
−100 to 600 msec around event onset, 100-msec incremental step). (B) ERP-image plot presenting single-trial broadband ERP responses stacked over
the y axis. This plot reveals a delayed amplitude increase in the case of the library condition in comparison to the prototypical P300 ERP response
latency observed in the tablet condition. (C) Average time–frequency activity relative to baseline period (−200 to 0 msec) in decibels (dB). The P300
ERP response is mainly arising from delta and theta-band (1–8 Hz) activity. (D) Intertrial coherence (ITC) measures of time–frequency dynamics. A
statistical mask with an alpha of .05 was applied (permutation t tests, FDR correction). The tablet condition exhibits a strong coherence across trials
within the P300 ERP time window and frequency range whereas no coherence is found in time–frequency dynamics for the library condition. Taken
together, these results highlight that the tablet condition reliably evoked P300 ERP responses that were time- and phase-locked to the onset of
stimulus presentation. In contrast, the library condition is characterized by EEG signals that share P300 characteristic features but that are distributed
over a wide time range. ESRP = Event-Related Spectral Perturbations.
responses are effectively reflecting time-shifted P300 ERP
responses.
Assessing Similarity between Library and
Tablet Signals
A template matching method accounting for temporal
shifts is required to assess whether the P300-like signals
observed across the library EEG data are similar to time-
locked ERP responses recorded in the tablet condition.
Dynamic Time-Warping (DTW) algorithms allow to com-
pute measures of similarity between time series that may
vary in speed and consequently be shifted in time. DTW is
an ubiquitous approach commonly applied to speech rec-
ognition to handle variations in speaking speed. DTW
algorithms compute an optimal match between two time
series, in the present case a template based on the average
ERP waveform of the tablet condition and single-trial ERP
waveforms of the library condition. The sequences are
nonlinearly warped (i.e., shifted) in the time domain,
every data point of each sequence being matched with
at least one data point from the other time series. A cost
measure is computed as the sum of absolute distance
values of each matched pair of indices. The optimal match
is selected on a minimal cost basis. The similarity (some-
times referred as dissimilarity depending on applications)
measure provided by DTW accounts for amplitude dif-
ferences between the signals following the nonlinear
warping.
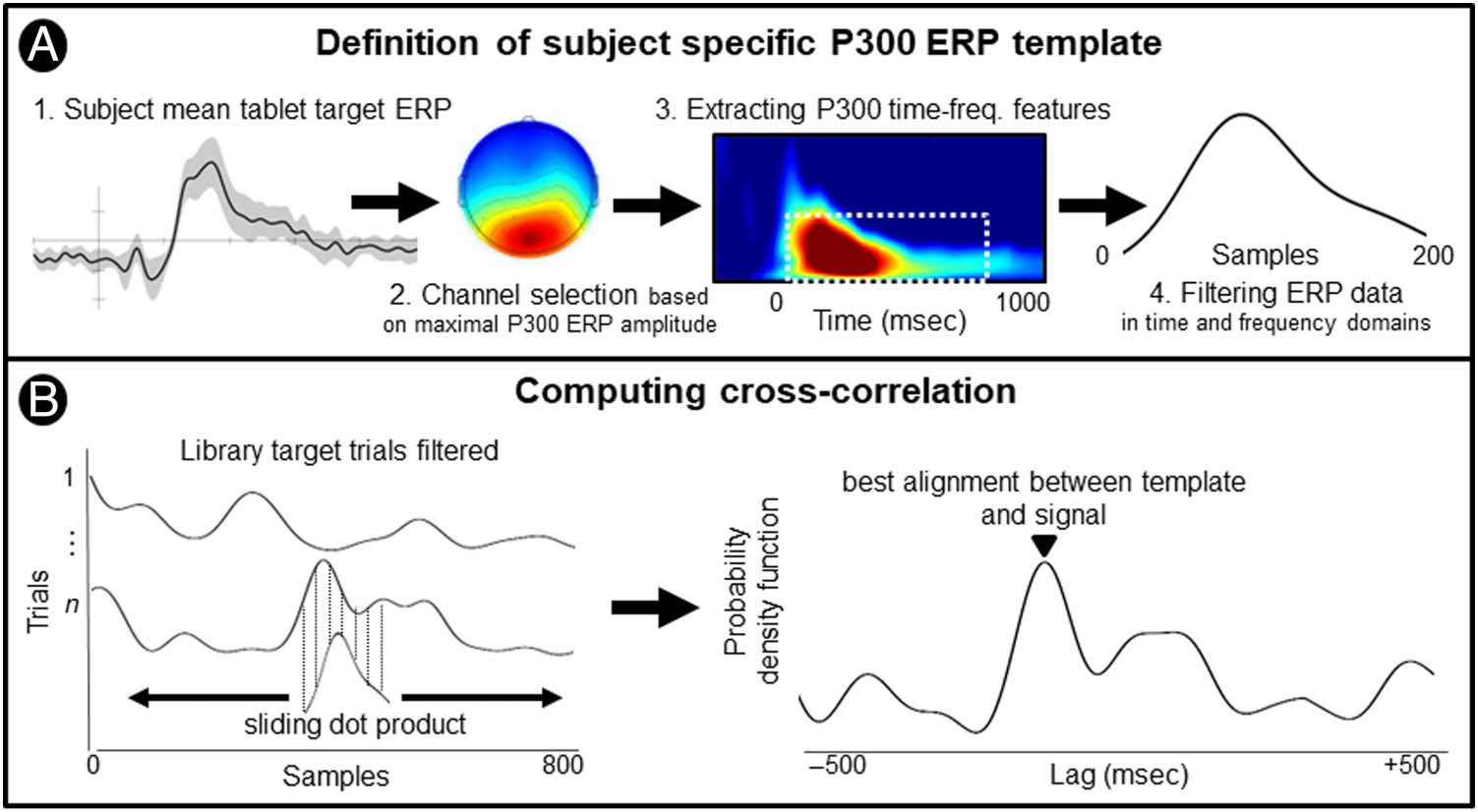
For each participant, a template was defined as the aver-
age ERP waveform (1- to 8-Hz bandpass filtered) of target
stimuli during the tablet condition (see Figure 4A). DTW
2246
Journal of Cognitive Neuroscience
Volume 34, Number 12
similarity measures were computed between the template
and single-trial time series (1- to 8-Hz bandpass filter) for
both target and nontarget stimuli of the library EEG data.
The library target trials were significantly more similar to
the template than the nontarget trials, t(18) = 4.58, p <
.001, Cohen’s d = 1.05, following nonlinear warping in
the time domain. This similarity further support that
P300 ERP responses are present in EEG data epoched
around the first fixation on the target book covers as these
time series are more to typical P300 ERP waveform elicited
by a computerized paradigm.
Alignment of Real-world ERP Responses
The previous observations and analyses have provided evi-
dence that P300 ERP features are present around the first
fixations on target book covers but these signals are not
time-locked to the fixation onset. To perform ERP analyses
on the library data, it is critical to address the latency var-
iability of its ERP responses. Although DTW measures offer
a metric to assess the similarity between temporal
sequences, it cannot be applied to correct latency because
of its nonlinearity (further developed in the Discussion
section). The cross-correlation method computes the sim-
ilarity between two time series that are shifted along each
other. The result of this convolution is a sliding dot prod-
uct whose maximum value informs about the lag between
the two series that optimizes similarity (see Figure 4B).
This method is useful to search for known features within
long signals. Following a similar implementation than
DTW, a subject-specific template of the tablet P300 ERP
is slid over every library EEG data epochs. The temporal
lag maximizing the similarity between the time series will
then be used to align the library single-trial data. The aver-
age temporal lag between the tablet P300 template and
library single-trial data was −80 msec (SD = 344 msec).
The first fixation on target stimuli timing used for epoch
extraction was corrected on a single-trial basis. The contin-
uous EEG data were then epoched around lag-corrected
markers. The following sections present ERP analyses per-
formed on time-corrected epoched data.
Lag-corrected ERP Analyses
The repeated-measures ANOVA revealed that Condition
did not have a main effect on P300 ERP amplitude, F(1,
18) = 0.44, p = .51, η2 = .007, anymore, whereas the
Stimulus Type, F(1, 18) = 16.82, p < .001, η2 = .27, had
a main effect on P300 ERP amplitude. There was no inter-
action, F(1, 18) = .996, p = .33, η2 = .007, between the
two factors on P300 ERP amplitude. Post hoc comparisons
revealed that target stimuli elicited P300 ERP responses of
significantly higher amplitude than nontarget stimuli in
the lag-corrected library condition, t(18) = 2.403, p =
.027, Cohen’s d = 0.551. Interestingly, no significant dif-
ference was found in P300 ERP amplitude between the
lag-corrected library and tablet conditions, t(18) = .454,
Figure 4. Implementation of cross-correlation measures for the time-domain alignment of P300 ERP responses recorded in the library condition. (A)
First, a subject-specific template was computed as the mean ERP response elicited by target trials of the tablet condition (shaded area indicates the
standard error). Second, the channel exhibiting the maximal ERP amplitude within the P300 time window (250–500 msec) was selected. Third, a
time–frequency decomposition of the ERP was then performed on the selected channel data; the frequencies and temporal features of the P300 ERP
responses were extracted. Last, the selected channel data were then filtered at the frequencies contributing the most to the P300 ERP, and data points
included in the temporal window previously defined were then used to create a filtered P300 ERP template. (B) The library target data were filtered
accordingly to the spectral and spatial filters applied to the template data. Cross-correlations between the template and the signal data were
performed for each target trial of the library condition. The resulting sliding dot product was used to create a probability density function whose
maximal value indicates the best alignment between the template and the signal. The lag associated with the maximal value was then used to correct
the single-trial onset timing.
Ladouce et al.
2247
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Grand-average (n =
19) ERP difference (targets −
nontargets) waveforms
recorded at parietal electrode
(Pz). (A) Grand-average ERP
waveforms for the tablet
condition (shaded area
indicates the standard error).
The topographical distribution
within the P300 ERP time
window (250–500 msec, see
dotted frame) is presented as a
scalp map. A time–frequency
plot presents mean event-
related changes in spectral
power (decibel relative to
−200- to 0-msec prestimulus
baseline). (B) Grand-average
ERP waveforms, scalp map, and
time–frequency plot for the
library condition after lag
correction. (C) Graphical
visualizations of mean P300 ERP
amplitude distribution across
experimental conditions.
p = .655, Cohen’s d = 0.104, as can be observed in
Figure 5.
DISCUSSION
The present study was designed to assess the feasibility of
capturing neural markers of visual attention related to the
processing of objects embedded in the real world.
Inspired by the scalable experimental design approach,
two conditions at both ends of the experimental control
and ecological validity continuum were contrasted: a clas-
sic visual oddball paradigm running on a tablet and a nat-
uralistic visual search task of book covers in a library.
Whereas the tablet paradigm presented a series of discrete
stimuli whose onset was timestamped in the EEG data,
the library EEG data were epoched around visual fixations
on experimental objects.
In the initial analyses, the presence of the P300 ERP
response was only found for the tablet condition. The
inspection of single-trial data recorded in the library
revealed signals whose features were similar to the P300
ERP found for the tablet condition. Such signals appeared
to be shifted in time around the initial fixations on target
book covers. Temporal jitters have important implications
for analyses based on the averaging of brain signals
acquired over multiple experimental trials. Indeed, averag-
ing time-shifted signals may lead to a smearing of the
resulting averaged waveform. This issue raises the ques-
tion regarding how to determine the timing of the pro-
cessing of the visual objects (i.e., books) embedded in
the real world. Such a definition is highly dependent on
the validity of the measure used. Should that definition
be based on the visual information entering the field of
view of the individual already raises issues regarding the
very definition of this visual field. Is the phenomenological
experience of a visual object bound to its entrance into the
foveal spotlight? In this case, then the initial fixation on a
visual object (or the onset of the preceding saccade)
appears a valid approach to extract brain dynamics reflec-
tive of such a visual cognitive experience. This assump-
tion, however, does not take into account empirical
evidence from reading research that visual information
entering parafoveal fields are already processed at sensory
but also semantic levels (Pan, Frisson, & Jensen, 2021).
Using the entrance within the parafoveal field of view to
define the onset of a visual event, however, requires
computing the angular distance between the individual’s
retina and the visual object.
In the absence of a continuous measure of object-to-eye
distance over the course of the experiment added to var-
iability in object dimensions, the visual subfield delimita-
tion approach was not possible. As an alternative solution,
we decided to apply template-matching approaches to
investigate the presence of P300 ERP in the library data.
We performed DTW similarity measures using the individ-
ual subject template of the P300 response based on the
tablet data that we compared with signals recorded in
the library. The similarity measures with the tablet tem-
plate were substantially higher for signals around visual fix-
ations on target book covers than on nontarget book
covers, suggesting the presence of time-shifted P300
ERP that are specific to target trials in the library condition.
Although DTW is a powerful method to evaluate the like-
ness of ERP responses between library and tablet condi-
tions, it does not provide a measurement of how much
time series have to be warped in the time domain to match
the template. Moreover, the nonlinear warping may intro-
duce distortions of the time series and the resulting
warped series may be of different lengths (i.e., different
number of data points), which may bias further statistical
2248
Journal of Cognitive Neuroscience
Volume 34, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
analyses (Zhao, Xu, Li, & Wu, 2020). Another approach was
therefore needed to quantitatively characterize ERP fea-
tures while accounting for temporal lags. The cross-
correlation coefficient was therefore used to find the lag
that would maximize the similarity between the tablet tem-
plate and the library data upon which target events latency
was then lag corrected.
The realignment of the library signals uncovered a P300
ERP response. The present findings suggest that neural
markers of visual attention elicited by computerized para-
digms are also present in a naturalistic environment.
These results demonstrate the potential of the joint
recording of EEG and E-T data to study visual cognition
in the real world. Moreover, retrieval of cognitive events
latency through analyses carried upon EEG data can in
turn be used to gain insight into gaze dynamics involved
in object recognition and processing (see Figure 4C).
These results also highlight the relevance of data-driven
approaches such as template-matching to recover neural
signals of interest in real-world data. However, these find-
ings should be interpreted cautiously. For instance, did
the P300 evoked response to account for the same cogni-
tive phenomena in the laboratory and real-world scenar-
ios? Is it possible to compare the amplitude and the latency
of this latter ERP across the two experimental conditions
and draw a conclusion about the effect of environmental
factors and task difficulty previously reported in several
studies (Cortney Bradford, Lukos, Passaro, Ries, & Ferris,
2019; Dehais, Rida, et al., 2019; Ladouce et al., 2019)? The
following sections will address critical differences between
the two experimental settings (tablet and library condi-
tions) and discuss the implications of such discrepancies.
Differences between Free-viewing and
Gaze-fixed Paradigms
In a gaze-fixed scenario such as the tablet paradigm, no
particular predictions of information localization or eye
movements are required. In contrast, the active free-
viewing search task in the library implies an exploration
of the visual environment to identify the identity of a stim-
ulus (target or nontarget). The decision of where to look
next at any given time is therefore critical. Because of the
high degree of freedom inherent in real-world settings
such as in the library condition, our participants’ cognitive
experiences were more complex, dynamic, and multidi-
mensional. Indeed, at any given time, there is a wide range
of multimodal sensory information to process, with each
of this information having a certain number of potential
states. Based on this information input, the nervous sys-
tem works toward building an understanding of the pres-
ent state of the environment and attempts to predict
future states. These differences have several implications.
First, the higher-order cognitive processes involved in the
free-viewing library task to update contextual information
and guide the visual exploration of the environment do
not correspond to those involved in the gaze-fixed tablet
condition. Second, the EEG data recorded in a free-
viewing task inherently includes both neural activity
related to experimental events but also ocular artifacts
(especially within the frame of FRP analyses; see Ehinger
& Dimigen, 2019)). The temporal overlap between neural
responses evoked by consecutive fixations is another crit-
ical issue related to free-viewing conditions. Indeed, the
series of rapid eye movements that typically precede the
initial fixation on an experimental object may all bring their
respective neural signatures, which could be confounded
for the visual ERPs as their delayed latencies and their
short temporal spacing may overlap with the latter.
Although later components such as the P300 are less
prone to this issue than early sensory processing compo-
nents (i.e., N1 and P1; Dandekar, Privitera, Carney, &
Klein, 2012), it is worth noting that regression-based anal-
yses approaches have been proposed to disentangle the
contribution of consecutive eye movements on EEG sig-
nals based on E-T information (Dimigen & Ehinger,
2021). Furthermore, the presence of neighboring visual
information requires increasing cognitive demands to
select the target information and inhibit the exploration
and processing of concurrent information (Hillyard, Hink,
Schwent, & Picton, 1973). In the case of the free-viewing
library task, top–down cognitive processes direct atten-
tional resources toward the detection and identification
of the target while inhibiting bottom–up influences of dis-
tracting information. The nature of this competition for
attentional resources does not apply to the same extent
to the tablet condition. Future studies applying a scalable
experimental design coupled with template-matching
approaches such as presented here should try to design
the laboratory condition to be as close to the real-world
condition as possible. In the case of the present study, a
free-viewing paradigm where participants look for an
object embedded in natural scenes could have been an
intermediate condition between the tablet and the library
condition.
Permanency and Uniqueness of Real-world
Visual Experiences
The perceptual differences between target stimuli in the
experimental tasks are another factor that could lead to
differences in terms of neural responses recorded
between the tablet and library conditions. It has been
largely documented in traditional P300 experiments that
a higher contrast between target and nontarget stimuli
leads to an increase in P300 amplitude. The saliency of tar-
get stimuli has been shown to largely contribute to this
effect (Luo & Ding, 2020). The tablet experiment uses var-
iations of low-level perceptual properties of the stimuli
such as shapes and colors to make the target and nontar-
get highly distinguishable from each other. The target
book covers used in this experiment were all different,
and so were their respective neighboring book covers. More
than shapes and colors, the shelves density and the width of
Ladouce et al.
2249
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
covers systematically varied across trials recorded in the
naturalistic library setting. This means that each trial was
singularly different from another in terms of how target
and nontarget stimuli were contrasted and how relatively
salient the target stimuli were. The objective of the present
study was to dive directly into the investigation of neural
dynamics during free visual exploration of a real-world
environment. For this reason, the experimental setup took
place in a naturalistic environment that was not altered by
the introduction of experimental probes. Nevertheless, the
additional degrees of freedom pertaining to the use of vari-
ous books as target stimuli in the library condition could,
however, be circumvented through the use of identical
book covers that would be placed in the vicinity of other
book covers that would share similar low-level perceptual
properties. Although such an approach is not an ideal solu-
tion to achieve the vision of studying cognitive experiences
directly in the real world and may be impractical to set up, it
could, however, help reconcile the measures by reducing
potential differences related to low-level visual processes
between experimental conditions. Eventually, the per-
manence of the visual information is yet another difference
between the two experimental conditions. What is meant by
permanence here is that the visual objects are always present
in the environment, before and after being experienced by
the user. This is in contrast to a discrete visual event such
as a sudden flash of light or images appearing on a moni-
tor. In the latter cases, the phenomenal visual experience
can be timely defined coincidentally to the onset and
offset of the physical apparition of the visual stimuli.
A distinction should nevertheless be made between exper-
imental events and cognitive phenomena. Computerized
paradigms use the former to elicit the latter. In the real
world, how such cognitive phenomena are expressed and
how they can be captured remain unknown. Therefore, a
sensible question is whether neural responses typically
observed in the laboratory such as the P300 ERP are merely
just a by-product of the specific way sequences of stimuli
are presented and are therefore not present in everyday
life experiences.
Considerations Regarding Brain and Body Imaging
in the Real World
Template Matching Approaches: Advantages
and Limitations
This study clearly illustrates how capturing neural
responses to visual objects embedded in the real world con-
stitutes a substantial leap (both technically and conceptu-
ally) from recording evoked potentials elicited under highly
controlled laboratory settings. These challenges were
addressed through a combination of novel methodologies
ranging from the experimental design to the recording and
analysis of the data. However, the template-matching
approaches (DTW and cross-correlation) applied to correct
the library data event latencies present several pitfalls that
are crucial to consider to interpreting the results. First, by
comparing the library EEG signals recorded a few seconds
before and after the first fixation on the target with a pro-
totypical signal representing a P300 effect recorded using a
computerized paradigm, a strong assumption regarding
the expression of the visual cognitive event in the library
is made. Although this assumption is directly derived from
the original hypotheses of the study and therefore pro-
vides a sound basis for the application of such a method,
the results should nevertheless be interpreted with cau-
tion. Indeed, by identifying features in the library data that
resemble the template, the alignment resulting from the
correction may produce a waveform whose features are
similar to the template but whose origin may be different
(i.e., brain signals not related to the task or even noise). In
the present case, other peaks related to earlier or later
components reflecting distinct cognitive processes might
be picked up and realigned as if they were a P300.
Although this possibility cannot be entirely dismissed, it
should be noted that the relative proximity of the cor-
rected responses (indicated by the low lag values around
the original fixation point) and the presence of other com-
ponents that were not included in the template and its
localization (as indicated by topographical maps) together
suggest that the latency-corrected signals share properties
with the tablet signals that go beyond mere spectral and
temporal domains similarities. Second, the application of
such an approach to the library data not only provides a
means to correct the latency of the effect (provided that
the previous assumption is valid) but this approach also
artificially reduces the temporal variance of such signals
in the corrected data by imposing strong temporal con-
straints that are based on the template data. This issue
has implications for the interpretation of both the ampli-
tude and latency of the P300 ERP. Indeed, the strong time-
locking and phase-locking induced artificially by the
realignment of the EEG signals will exhibit minimal tempo-
ral variance that may inflate the amplitude of the averaged
waveform as it is not subject to the same smearing that
would naturally occur in relation to variance in the P300
latency across trials. Moreover, variations in P300 latency
may be particularly informative within the frame of real-
world experiments as several aspects of the visual stimuli
(i.e., book cover more or less salient) but also environ-
mental factors (i.e., competition of surrounding visual
information) could have an impact on cognitive process-
ing speed. Although the cross-correlation-based template
matching approach is not an ideal solution, the corrected
latencies derived from the EEG signal template provide
information that opens a new range of analyses to be car-
ried out on the multimodal data.
Intrusiveness of MoBI Devices: Toward
Transparent Solutions
The participants were equipped with a research-grade
mobile EEG system that comprised an EEG cap tethered
to an amplifier that was fitted in a backpack and E-T glasses
2250
Journal of Cognitive Neuroscience
Volume 34, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
also tethered to a recording unit. Although the E-T glasses
have been deemed as relatively unobtrusive following a
short adaptation period, wearing the EEG cap in a public
space such as a university library was reportedly a self-
conscious experience for the participants. Although data
quality is critical for the selection of EEG systems, the
degree of comfort and discretion are also key aspects to
consider in the context of real-world studies. An elegant
solution to this issue could be found in minimalist EEG
devices. For example, around-the-ear electrode arrays
are discreet, quick to set up, and can be worn comfortably
for extended periods (Debener, Emkes, De Vos, & Bleichner,
2015). The minimalist and ear-centered montage does not,
however, offer the spatial resolution of a whole-head EEG
system and therefore may not be sensitive to far-field
potentials (Meiser, Tadel, Debener, & Bleichner, 2020).
Nonetheless, around-the-ear arrays have proven to be an
effective solution to record neural markers of visual
processes (Pacharra, Debener, & Wascher, 2017) and
selective attention to auditory information (Mirkovic,
Bleichner, De Vos, & Debener, 2016; Bleichner et al.,
2015; Debener et al., 2015). This comfortable solution is
particularly appealing for the study of everyday-life human
cognition but also offers promising medical use cases
such as sleep staging and long-term monitoring of epilep-
tiform activity (Bleichner & Debener, 2017). The future of
MoBI research field is linked to the development of
inconspicuous and comfortable recording devices as they
will enable the range of research to expand to social
interactions in real-life situations.
Ethics of Real-world Brain and Body Imaging Data
A wide range of personal information is gathered from per-
vasive devices (i.e., mobile phones, smartwatches). The
recording of GPS, heart rate, and accelerometer data can
be used to track where and what an individual is doing at
various times of the day. Although privacy concerns are
raised sporadically, the quantity and variety of data col-
lected from individuals have been steadily on the rise.
More pointedly, instrumentations that were limited to
medical and research purposes are now ported into
consumer-grade devices. As this new generation of
devices find their way to households and their daily usage
becomes widely adopted by the population, it will be con-
ceivable that the general public might be desensitized to
the intimate nature of the data collected. Gaze data are a
striking example of how personal preferences and other-
wise covert information regarding one’s experience of its
environment can be effectively derived. Indeed, metrics
such as fixation duration and relative number of saccades
when exploring a visual object provide objective measures
of how much attention has been paid to the object and
indicate the depth of cognitive processing of that informa-
tion, revealing interest and preferences. Advances in the
fields of computer vision are providing efficient ways to
label visual objects in video recordings in an automatic
and increasingly reliable manner. The extraction of neural
markers in relation to labeled visual data adds another
layer of insight into how an individual’s attention, seman-
tic processing, and decision-making operates. The fusion
of gaze and brain data offers novel opportunities to extend
Brain–Computer Interface applications beyond the pre-
sentation of artificial interfaces and paradigms. It is
through the acquisition of such contextual information
that neural data will deliver its full potential for real-world
applications. Aside from the exciting prospects offered by
the exploitation of multimodal data for everyday-life appli-
cations, it is critical to reflect on the sensitive nature of
such data. Indeed, once equipped with sensors that will
become ever more transparent, data will be acquired con-
tinuously without the user being conscious of it. The
covert aspects of continuous physiological data recordings
pose questions regarding the consent of the user for such
information to be exploited at any given moment. To be
clear, multimodal brain and body imaging data pose novel
ethical issues regarding data privacy. It falls upon
researchers the responsibility to ensure that data privacy
remains a priority by setting precedents of high standards
in how such multimodal data are handled.
Conclusion
The challenges related to the study of human cognition in
everyday life contexts are numerous. Abandoning comput-
erized paradigm lessens experimental control over a wide
range of variables that can add variance to participants’
behavior and cognitive processes involved in a certain
task. In the context of EEG analyses, this high degree of
freedom poses several conceptual and technical issues,
notably related to the timing of experimental and cognitive
events. The present findings provide evidence that such
challenges may be overcome through a combination of
scalable experimental design, recording of multimodal
brain and body imaging data, and the application of state
of the art signal processing and template matching
methods. By applying these approaches, neural markers
of cognitive processes related to visual information
embedded in a real-world environment could be captured.
These encouraging results further highlight the relevance
of scalable experiments to study human cognition in real-
world contexts. Indeed, recording neural responses elic-
ited by the discrete presentation of visual stimuli through
a computerized paradigm (i.e., the tablet condition) was
instrumental to create a template that could be used to
search for and extract similar responses in a more natural-
istic, and therefore less controlled, recording setting such
as the library condition. Moreover, the extended range of
analyses enabled by the joint recording of gaze and brain
dynamics showcases the complementarity of MoBI
methods. The E-T data provided contextual information
regarding the occurrence of experimental events that
enabled time-domain analyses to be performed on the
EEG data. Although the information provided about
Ladouce et al.
2251
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
experimental event timing may not have the temporal res-
olution required to perform time-domain analyses at the
milliseconds scale, they can, however, serve as an initial
estimate upon which template matching approaches are
applied to extract ERP features. In conclusion, adopting
a scalable approach to experimental design and leveraging
the potential of multimodal recording methodologies are
important steps toward enabling the study of embodied
aspects of human cognition in naturalistic environments.
Acknowledgments
The authors thank Catriona Bruce and Stephen Stewart for the
technical assistance in setting up the experiment. Thank you
to Ludovic Darmet for his advice on data analysis.
Reprint requests should be sent to Simon Ladouce, PhD, Depart-
ment for Aerospace Vehicles Design and Control, Institut
Supérieur de l’Aéronautique et de l’Espace (ISAE), Toulouse,
Haute-Garonne, 31055, France, or via e-mail: simon.ladouce@isae
-supaero.fr.
Data Availability Statement
Original EEG data with experimental events annotations
along with the code used for processing and analysis of
the data are available on the following online repository:
https://osf.io/zhcr7/.
Funding Information
This work was supported by the Agence Innovation
Defense - AID (RAPID Neurosynchrone project). Frédéric
Dehais is a ANITI (Artificial and Natural Intelligence Tou-
louse Institute) chairholder. Simon Ladouce and Magda
Mustile were supported by scholarships from the Univer-
sity of Stirling and a research grant from SINAPSE (Scottish
Imaging Network: A Platform for Scientific excellence).
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance.
REFERENCES
Alexander, J. E., Bauer, L. O., Kuperman, S., Morzorati, S.,
O’Connor, S. J., Rohrbaugh, J., et al. (1996). Hemispheric
differences for P300 amplitude from an auditory oddball task.
International Journal of Psychophysiology, 21, 189–196.
https://doi.org/10.1016/0167-8760(95)00047-X, PubMed:
8792206
Ayaz, H., & Dehais, F. (2018). Neuroergonomics: The brain at
work and in everyday life. Elsevier - Academic Press.
Baccino, T., & Manunta, Y. (2005). Eye-fixation-related
potentials: Insight into parafoveal processing. Journal of
Psychophysiology, 19, 204–215. https://doi.org/10.1027/0269
-8803.19.3.204
Banaei, M., Hatami, J., Yazdanfar, A., & Gramann, K. (2017).
Walking through architectural spaces: The impact of interior
forms on human brain dynamics. Frontiers in Human
Neuroscience, 11, 477. https://doi.org/10.3389/fnhum.2017
.00477, PubMed: 29033807
Bell, A. J., & Sejnowski, T. J. (1995). An information-
maximization approach to blind separation and blind
deconvolution. Neural Computation, 7, 1129–1159. https://
doi.org/10.1162/neco.1995.7.6.1129, PubMed: 7584893
Benjamin, A. V., Wailes-Newson, K., Ma-Wyatt, A., Baker, D. H.,
& Wade, A. R. (2018). The effect of locomotion on early visual
contrast processing in humans. Journal of Neuroscience, 38,
3050–3059. https://doi.org/10.1523/JNEUROSCI.1428-17
.2017, PubMed: 29463642
Birenboim, A., Helbich, M., & Kwan, M. P. (2021). Advances in
portable sensing for urban environments: Understanding
cities from a mobility perspective. Computers, Environment
and Urban Systems, 88, 101650. https://doi.org/10.1016/j
.compenvurbsys.2021.101650
Birmingham, E., Bischof, W. F., & Kingstone, A. (2009). Saliency
does not account for fixations to eyes within social scenes.
Vision Research, 49, 2992–3000. https://doi.org/10.1016/j
.visres.2009.09.014, PubMed: 19782100
Bleichner, M. G., & Debener, S. (2017). Concealed, unobtrusive
ear-centered EEG acquisition: cEEGrids for transparent
EEG. Frontiers in Human Neuroscience, 11, 163.
https://doi.org/10.3389/fnhum.2017.00163, PubMed:
25847919
Bleichner, M. G., Lundbeck, M., Selisky, M., Minow, F., Jäger,
M., Emkes, R., et al. (2015). Exploring miniaturized EEG
electrodes for brain–computer interfaces. An EEG you do not
see? Physiological Reports, 3, e12362. https://doi.org/10
.14814/phy2.12362, PubMed: 25847919
Brouwer, A. M., Reuderink, B., Vincent, J., van Gerven, M. A., &
van Erp, J. B. (2013). Distinguishing between target and
nontarget fixations in a visual search task using
fixation-related potentials. Journal of Vision, 13, 17. https://
doi.org/10.1167/13.3.17, PubMed: 23863335
Cao, L., Chen, X., & Haendel, B. F. (2020). Overground walking
decreases alpha activity and entrains eye movements in
humans. Frontiers in Human Neuroscience, 14, 561755.
https://doi.org/10.3389/fnhum.2020.561755, PubMed:
33414709
Cao, L., & Händel, B. (2019). Walking enhances peripheral
visual processing in humans. PLoS Biology, 17, e3000511.
https://doi.org/10.1371/journal.pbio.3000511, PubMed:
31603894
Chen, X., Cao, L., & Haendel, B. F. (2022). Differential effects of
walking across visual cortical processing stages. Cortex, 149,
16–28. https://doi.org/10.1016/j.cortex.2022.01.007, PubMed:
35182897
Cortney Bradford, J., Lukos, J. R., Passaro, A., Ries, A., & Ferris,
D. P. (2019). Effect of locomotor demands on cognitive
processing. Scientific Reports, 9, 9234. https://doi.org/10
.1038/s41598-019-45396-5, PubMed: 31239461
Dandekar, S., Privitera, C., Carney, T., & Klein, S. A. (2012).
Neural saccadic response estimation during natural viewing.
Journal of Neurophysiology, 107, 1776–1790. https://doi.org
/10.1152/jn.00237.2011, PubMed: 22170971
2252
Journal of Cognitive Neuroscience
Volume 34, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
De Vos, M., Gandras, K., & Debener, S. (2014). Towards a truly
mobile auditory brain-computer interface: Exploring the P300
to take away. International Journal of Psychophysiology, 91,
46–53. https://doi.org/10.1016/j.ijpsycho.2013.08.010,
PubMed: 23994208
Debener, S., Emkes, R., De Vos, M., & Bleichner, M. (2015).
Unobtrusive ambulatory EEG using a smartphone and flexible
printed electrodes around the ear. Scientific Reports, 5,
16743. https://doi.org/10.1038/srep16743, PubMed: 26572314
Debener, S., Minow, F., Emkes, R., Gandras, K., & de Vos, M.
(2012). How about taking a low-cost, small, and wireless
EEG for a walk? Psychophysiology, 49, 1617–1621.
https://doi.org/10.1111/j.1469-8986.2012.01471.x, PubMed:
23013047
Dehais, F., Duprès, A., Blum, S., Drougard, N., Scannella, S.,
Roy, R. N., et al. (2019). Monitoring pilot’s mental
workload using ERPs and spectral power with a
six-dry-electrode EEG system in real flight conditions.
Sensors, 19, 1324. https://doi.org/10.3390/s19061324,
PubMed: 30884825
Dehais, F., Karwowski, W., & Ayaz, H. (2020). Brain at work and
in everyday life as the next frontier: Grand field challenges for
neuroergonomics. Frontiers in Neuroergonomics, 1, 583733.
https://doi.org/10.3389/fnrgo.2020.583733
Dehais, F., Rida, I., Roy, R. N., Iversen, J., Mullen, T., & Callan,
D. (2019). A pBCI to predict attentional error before
it happens in real flight conditions. In 2019 IEEE
international conference on systems, man and cybernetics
(SMC) (pp. 4155–4160). https://doi.org/10.1109/SMC.2019
.8914010
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003
.10.009, PubMed: 15102499
Dimigen, O., & Ehinger, B. V. (2021). Regression-based analysis
of combined EEG and eye-tracking data: Theory and
applications. Journal of Vision, 21, 3. https://doi.org/10.1167
/jov.21.1.3, PubMed: 33410892
Djebbara, Z., Fich, L. B., & Gramann, K. (2019). Brain art.
Springer International Publishing.
22, 593–608. https://doi.org/10.1515/RNS.2011.047, PubMed:
22070621
Gramann, K., Jung, T. P., Ferris, D. P., Lin, C. T., & Makeig, S.
(2014). Toward a new cognitive neuroscience: Modeling
natural brain dynamics. Frontiers in Human Neuroscience,
8, 444. https://doi.org/10.3389/fnhum.2014.00444, PubMed:
24994978
Gramann, K., McKendrick, R., Baldwin, C., Roy, R. N., Jeunet, C.,
Mehta, R. K., et al. (2021). Grand field challenges for
cognitive neuroergonomics in the coming decade. Frontiers
in Neuroergonomics, 2, 643969. https://doi.org/10.3389
/fnrgo.2021.643969
Hayhoe, M., & Ballard, D. (2005). Eye movements in natural
behavior. Trends in Cognitive Sciences, 9, 188–194. https://
doi.org/10.1016/j.tics.2005.02.009, PubMed: 15808501
Henderson, J. M., Brockmole, J. R., Castelhano, M. S., &
Mack, M. (2007). Visual saliency does not account for eye
movements during visual search in real-world scenes. In
R. P. G. van Gompel, M. H. Fischer, W. S. Murray, & R. L. Hill
(Eds.), Eye movements: A window on mind and brain
(pp. 537–562). Elsevier. https://doi.org/10.1016/B978
-008044980-7/50027-6
Henderson, J. M., & Luke, S. G. (2014). Stable individual
differences in saccadic eye movements during reading,
pseudoreading, scene viewing, and scene search. Journal of
Experimental Psychology: Human Perception and
Performance, 40, 1390–1400. https://doi.org/10.1037
/a0036330, PubMed: 24730735
Hillyard, S. A., Hink, R. F., Schwent, V. L., & Picton, T. W. (1973).
Electrical signs of selective attention in the human brain.
Science, 182, 177–180. https://doi.org/10.1126/science.182
.4108.177, PubMed: 4730062
Hölle, D., Blum, S., Kissner, S., Debener, S., & Bleichner, M. G.
(2022). Real-time audio processing of real-life soundscapes
for EEG analysis: ERPs based on natural sound onsets.
Frontiers in Neuroergonomics, 3, 793061. https://doi.org/10
.3389/fnrgo.2022.793061
Hölle, D., Meekes, J., & Bleichner, M. G. (2021). Mobile ear-EEG
to study auditory attention in everyday life. Behavior
Research Methods, 53, 2025–2036. https://doi.org/10.3758
/s13428-021-01538-0, PubMed: 33721208
Djebbara, Z., Fich, L. B., Petrini, L., & Gramann, K. (2019).
Kamienkowski, J. E., Ison, M. J., Quiroga, R. Q., & Sigman, M.
Sensorimotor brain dynamics reflect architectural
affordances. Proceedings of the National Academy of
Sciences, U.S.A., 116, 14769–14778. https://doi.org/10.1073
/pnas.1900648116, PubMed: 31189596
Djebbara, Z., Jensen, O. B., Parada, F. J., & Gramann, K. (2022).
Neuroscience and architecture: Modulating behavior through
sensorimotor responses to the built environment.
Neuroscience and Biobehavioral Reviews, 138, 104715.
https://doi.org/10.1016/j.neubiorev.2022.104715, PubMed:
35654280
Do, T. T. N., Lin, C. T., & Gramann, K. (2021). Human brain
dynamics in active spatial navigation. Scientific Reports, 11,
13036. https://doi.org/10.1038/s41598-021-92246-4, PubMed:
34158525
Ehinger, B. V., & Dimigen, O. (2019). Unfold: An integrated
toolbox for overlap correction, non-linear modeling, and
regression-based EEG analysis. PeerJ, 7, e7838. https://doi.org
/10.7717/peerj.7838, PubMed: 31660265
Gramann, K., Gwin, J. T., Bigdely-Shamlo, N., Ferris, D. P., &
Makeig, S. (2010). Visual evoked responses during standing
and walking. Frontiers in Human Neuroscience, 4, 202.
https://doi.org/10.3389/fnhum.2010.00202, PubMed:
21267424
Gramann, K., Gwin, J. T., Ferris, D. P., Oie, K., Jung, T. P., Lin,
C. T., et al. (2011). Cognition in action: Imaging brain/body
dynamics in mobile humans. Reviews in the Neurosciences,
(2012). Fixation-related potentials in visual search: A
combined EEG and eye tracking study. Journal of Vision,
12, 4. https://doi.org/10.1167/12.7.4, PubMed: 22776848
Kaunitz, L. N., Kamienkowski, J. E., Varatharajah, A., Sigman, M.,
Quiroga, R. Q., & Ison, M. J. (2014). Looking for a face in the
crowd: Fixation-related potentials in an eye-movement visual
search task. Neuroimage, 89, 297–305. https://doi.org/10
.1016/j.neuroimage.2013.12.006, PubMed: 24342226
King, J. L., & Parada, F. J. (2021). Using mobile brain/body imaging
to advance research in arts, health, and related therapeutics.
European Journal of Neuroscience, 54, 8364–8380. https://
doi.org/10.1111/ejn.15313, PubMed: 33999462
Klug, M., & Gramann, K. (2021). Identifying key factors for
improving ICA-based decomposition of EEG data in mobile
and stationary experiments. European Journal of
Neuroscience, 54, 8406–8420. https://doi.org/10.1111/ejn
.14992, PubMed: 33012055
Kranczioch, C., Zich, C., Schierholz, I., & Sterr, A. (2014).
Mobile EEG and its potential to promote the theory and
application of imagery-based motor rehabilitation.
International Journal of Psychophysiology, 91, 10–15.
https://doi.org/10.1016/j.ijpsycho.2013.10.004, PubMed:
24144637
Ladouce, S., Donaldson, D. I., Dudchenko, P. A., & Ietswaart, M.
(2017). Understanding minds in real-world environments:
Toward a mobile cognition approach. Frontiers in Human
Ladouce et al.
2253
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neuroscience, 10, 694. https://doi.org/10.3389/fnhum.2016
.00694, PubMed: 28127283
Ladouce, S., Donaldson, D. I., Dudchenko, P. A., & Ietswaart, M.
(2019). Mobile EEG identifies the re-allocation of attention
during real-world activity. Scientific Reports, 9, 15851. https://
doi.org/10.1038/s41598-019-51996-y, PubMed: 31676780
Lau-Zhu, A., Lau, M., & McLoughlin, G. (2019). Mobile EEG in
research on neurodevelopmental disorders: Opportunities
and challenges. Developmental Cognitive Neuroscience, 36,
100635. https://doi.org/10.1016/j.dcn.2019.100635, PubMed:
30877927
Liebherr, M., Corcoran, A. W., Alday, P. M., Coussens, S., Bellan,
V., Howlett, C. A., et al. (2021). EEG and behavioral correlates
of attentional processing while walking and navigating
naturalistic environments. Scientific Reports, 11, 22325.
https://doi.org/10.1038/s41598-021-01772-8, PubMed:
34785702
Luo, C., & Ding, N. (2020). Visual target detection in a
distracting background relies on neural encoding of both
visual targets and background. Neuroimage, 216, 116870.
https://doi.org/10.1016/j.neuroimage.2020.116870, PubMed:
32339773
Makeig, S., Gramann, K., Jung, T. P., Sejnowski, T. J., & Poizner,
H. (2009). Linking brain, mind and behavior. International
Journal of Psychophysiology, 73, 95–100. https://doi.org/10
.1016/j.ijpsycho.2008.11.008, PubMed: 19414039
Mavros, P., Austwick, M. Z., & Smith, A. H. (2016). Geo-EEG:
Towards the use of EEG in the study of urban behaviour.
Applied Spatial Analysis and Policy, 9, 191–212. https://doi
.org/10.1007/s12061-015-9181-z
Mcdowell, K., Lin, C.-T., Oie, K. S., Jung, T.-P., Gordon, S.,
Whitaker, K. W., et al. (2013). Real-world neuroimaging
technologies. IEEE Access, 1, 131–149. https://doi.org/10.1109
/ACCESS.2013.2260791
Meiser, A., Tadel, F., Debener, S., & Bleichner, M. G. (2020).
The sensitivity of ear-EEG: Evaluating the source-sensor
relationship using forward modeling. Brain Topography, 33,
665–676. https://doi.org/10.1007/s10548-020-00793-2,
PubMed: 32833181
Mirkovic, B., Bleichner, M. G., De Vos, M., & Debener, S.
(2016). Target speaker detection with concealed EEG around
the ear. Frontiers in Neuroscience, 10, 349. https://doi.org/10
.3389/fnins.2016.00349, PubMed: 27512364
Miyakoshi, M., Gehrke, L., Gramann, K., Makeig, S., & Iversen, J.
(2021). The AudioMaze: An EEG and motion capture study of
human spatial navigation in sparse augmented reality.
European Journal of Neuroscience, 54, 8283–8307. https://
doi.org/10.1111/ejn.15131, PubMed: 33497490
Mustile, M., Kourtis, D., Ladouce, S., Learmonth, G., Edwards,
M. G., Donaldson, D. I., et al. (2021). Mobile EEG reveals
functionally dissociable dynamic processes supporting
real-world ambulatory obstacle avoidance: Evidence for early
proactive control. European Journal of Neuroscience, 54,
8106–8119. https://doi.org/10.1111/ejn.15120, PubMed:
33465827
Ouyang, G., Herzmann, G., Zhou, C., & Sommer, W. (2011).
Residue iteration decomposition (RIDE): A new method
to separate ERP components on the basis of latency
variability in single trials. Psychophysiology, 48, 1631–1647.
https://doi.org/10.1111/j.1469-8986.2011.01269.x, PubMed:
21895682
Pacharra, M., Debener, S., & Wascher, E. (2017). Concealed
around-the-ear EEG captures cognitive processing in a visual
Simon task. Frontiers in Human Neuroscience, 11, 290.
https://doi.org/10.3389/fnhum.2017.00290, PubMed:
28642695
Pan, Y., Frisson, S., & Jensen, O. (2021). Neural evidence for
lexical parafoveal processing. Nature Communications, 12,
5234. https://doi.org/10.1038/s41467-021-25571-x, PubMed:
34475391
Parada, F. J. (2018). Understanding natural cognition in
everyday settings: 3 pressing challenges. Frontiers in Human
Neuroscience, 12, 386. https://doi.org/10.3389/fnhum.2018
.00386, PubMed: 30319381
Park, J. L., Fairweather, M. M., & Donaldson, D. I. (2015).
Making the case for mobile cognition: EEG and sports
performance. Neuroscience and Biobehavioral Reviews, 52,
117–130. https://doi.org/10.1016/j.neubiorev.2015.02.014,
PubMed: 25735956
Peters, R. J., Iyer, A., Itti, L., & Koch, C. (2005). Components of
bottom–up gaze allocation in natural images. Vision
Research, 45, 2397–2416. https://doi.org/10.1016/j.visres.2005
.03.019, PubMed: 15935435
Pion-Tonachini, L., Kreutz-Delgado, K., & Makeig, S. (2019).
ICLabel: An automated electroencephalographic
independent component classifier, dataset, and website.
Neuroimage, 198, 181–197. https://doi.org/10.1016/j
.neuroimage.2019.05.026, PubMed: 31103785
Polich, J. (2007). Updating P300: An integrative theory of P3a
and P3b. Clinical Neurophysiology, 118, 2128–2148.
https://doi.org/10.1016/j.clinph.2007.04.019, PubMed:
17573239
Polich, J., Alexander, J. E., Bauer, L. O., Kuperman, S., Morzorati,
S., O’Connor, S. J., et al. (1997). P300 topography of
amplitude/latency correlations. Brain Topography, 9,
275–282. https://doi.org/10.1007/BF01464482, PubMed:
9217986
Reis, P. M., Hebenstreit, F., Gabsteiger, F., von Tscharner, V., &
Lochmann, M. (2014). Methodological aspects of EEG and
body dynamics measurements during motion. Frontiers in
Human Neuroscience, 8, 156. https://doi.org/10.3389/fnhum
.2014.00156, PubMed: 24715858
Reiser, J. E., Wascher, E., & Arnau, S. (2019). Recording mobile
EEG in an outdoor environment reveals cognitive-motor
interference dependent on movement complexity. Scientific
Reports, 9, 13086. https://doi.org/10.1038/s41598-019-49503-4,
PubMed: 31511571
Roberts, H., Soto, V., Tyson-Carr, J., Kokmotou, K., Cook, S.,
Fallon, N., et al. (2018). Tracking economic value of products
in natural settings: A wireless EEG study. Frontiers in
Neuroscience, 12, 910. https://doi.org/10.3389/fnins.2018
.00910, PubMed: 30618548
Scanlon, J. E. M., Redman, E. X., Kuziek, J., & Mathewson, K. E.
(2020). A ride in the park: Cycling in different outdoor
environments modulates the auditory evoked potentials.
International Journal of Psychophysiology, 151, 59–69.
https://doi.org/10.1016/j.ijpsycho.2020.02.016, PubMed:
32119887
Scanlon, J. E. M., Townsend, K. A., Cormier, D. L., Kuziek, J., &
Mathewson, K. E. (2019). Taking off the training wheels:
Measuring auditory P3 during outdoor cycling using an active
wet EEG system. Brain Research, 1716, 50–61. https://doi.org
/10.1016/j.brainres.2017.12.010, PubMed: 29248602
Schmidt-Kassow, M., Deusser, M., Thiel, C., Otterbein, S.,
Montag, C., Reuter, M., et al. (2013). Physical exercise during
encoding improves vocabulary learning in young female
adults: A neuroendocrinological study. PLoS One, 8, e64172.
https://doi.org/10.1371/journal.pone.0064172, PubMed:
23700461
Schmidt-Kassow, M., Heinemann, L. V., Abel, C., & Kaiser, J.
(2013). Auditory-motor synchronization facilitates attention
allocation. Neuroimage, 82, 101–106. https://doi.org/10.1016
/j.neuroimage.2013.05.111, PubMed: 23732882
Soto, V., Tyson-Carr, J., Kokmotou, K., Roberts, H., Cook, S.,
Fallon, N., et al. (2018). Brain responses to emotional faces in
natural settings: A wireless Mobile EEG recording study.
2254
Journal of Cognitive Neuroscience
Volume 34, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Frontiers in Psychology, 9, 2003. https://doi.org/10.3389
/fpsyg.2018.02003, PubMed: 30410458
Underwood, G. (2009). Cognitive processes in eye guidance:
Algorithms for attention in image processing. Cognitive
Computation, 1, 64–76. https://doi.org/10.1007/s12559-008
-9002-7
Wunderlich, A., & Gramann, K. (2021). Landmark-based
navigation instructions improve incidental spatial knowledge
acquisition in real-world environments. Journal of
Environmental Psychology, 77, 101677. https://doi.org/10
.1016/j.jenvp.2021.101677
Zhao, W., Xu, Z., Li, W., & Wu, W. (2020). Modeling and
analyzing neural signals with phase variability using fisher-Rao
registration. Journal of Neuroscience Methods, 346, 108954.
https://doi.org/10.1016/j.jneumeth.2020.108954, PubMed:
32950555
Zink, R., Hunyadi, B., Huffel, S. V., & Vos, M. D. (2016).
Mobile EEG on the bike: Disentangling attentional and
physical contributions to auditory attention tasks.
Journal of Neural Engineering, 13, 046017.
https://doi.org/10.1088/1741-2560/13/4/046017, PubMed:
27351459
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
3
7
2
0
6
5
1
4
3
/
/
j
o
c
n
_
a
_
0
1
9
0
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Ladouce et al.
2255