Autobiographical Memory and Patterns of Brain
Atrophy in Frontotemporal Lobar Degeneration
Margaret C. McKinnon1,2, Elena I. Nica3, Pheth Sengdy3,
Natasa Kovacevic3, Morris Moscovitch3,4, Morris Freedman3,4,7,8,
Bruce L. Miller4, Sandra E. Black3,4,5, and Brian Levine3,4,6
D
Ö
w
N
l
Ö
A
D
e
D
Abstrakt
& Autobiographical memory paradigms have been increasingly
used to study the behavioral and neuroanatomical correlates of
human remote memory. Although there are numerous func-
tional neuroimaging studies on this topic, relatively few studies
of patient samples exist, with heterogeneity of results owing to
methodological variability. In this study, frontotemporal lobar
degeneration (FTLD), a form of dementia affecting regions cru-
cial to autobiographical memory, was used as a model of auto-
biographical memory loss. We emphasized the separation of
episodic (recollection of specific event, perceptual, and mental
state information) from semantic (factual information unspecific
in time and place) autobiographical memory, derived from a
reliable method for scoring transcribed autobiographical pro-
tocols, the Autobiographical Interview [Levine, B., Svoboda, E.,
Hay, J., Winocur, G., & Moscovitch, M. Aging and autobio-
graphical memory: Dissociating episodic from semantic retrieval.
Psychologie und Altern, 17, 677–689, 2002]. Patients with the
frontotemporal dementia (FTD) and mixed frontotemporal
and semantic dementia (FTD/SD) variants of FTLD were im-
paired at reconstructing episodically rich autobiographical mem-
ories across the lifespan, with FTD/SD patients generating an
excess of generic semantic autobiographical
Information. Pa-
tients with progressive nonfluent aphasia were mildly impaired
for episodic autobiographical memory, but this impairment was
eliminated with the provision of structured cueing, likely reflect-
ing relatively intact medial temporal lobe function, whereas the
same cueing failed to bolster the FTD and FTD/SD patients’
performance relative to that of matched comparison subjects.
The pattern of episodic, but not semantic, autobiographical im-
pairment was enhanced with disease progression on 1- to 2-year
follow-up testing in a subset of patients, supplementing the
cross-sectional evidence for specificity of episodic autobiograph-
ical impairment with longitudinal data. This behavioral pattern
covaried with volume loss in a distributed left-lateralized pos-
terior network centered on the temporal lobe, consistent with
evidence from other patient and functional neuroimaging stud-
ies of autobiographical memory. Frontal lobe volumes, Jedoch,
did not significantly contribute to this network, suggesting that
frontal contributions to autobiographical episodic memory may
be more complex than previously appreciated. &
EINFÜHRUNG
lobar degeneration (FTLD) is a com-
Frontotemporal
mon form of dementia and a leading cause of early-onset
Demenz, along with Alzheimer’s disease (Ratnavalli,
Brayne, Dawson, & Hodges, 2002). Although everyday
memory has been considered relatively spared in FTLD
(Neary et al., 1998), there is evidence that this disease
affects performance on laboratory tests of memory
(Simons, Graham, & Hodges, 2002). An emerging liter-
ature suggests that this deficit extends to the autobio-
graphical domain (Piolino et al., 2003; Nestor, Graham,
Bozeat, Simons, & Hodges, 2002; Hodges & Gurd, 1994).
FTLD is uniquely positioned as a model for testing
hypotheses concerning autobiographical memory. Gen-
1McMaster University, 2St. Joseph’s Healthcare, 3Baycrest Centre,
4Universität von Toronto, 5University of California at San Francisco,
6Sunnybrook and Women’s College Health Sciences Centre,
7Baycrest Centre for Geriatric Care, 8University Health Network
erally speaking, memory impairments in FTLD follow
from degeneration of the frontal and temporal cortex
associated with this disease. The temporal lobes, partic-
ularly the medial temporal regions, are classically asso-
ciated with mnemonic operations. The prefrontal cortex
is involved in higher-order mnemonic retrieval opera-
tionen (Fletcher & Henson, 2001; Stuss & Benson, 1986)
including strategic retrieval and monitoring within auto-
biographical memory (McKinnon, Svoboda, & Levine,
2007; Conway & Pleydell-Pearce, 2000). Genauer,
FTLD comprises three distinct subtypes: frontotemporal
Demenz (FTD), progressive nonfluent aphasia (PNFA),
and semantic dementia (SD) (Neary et al., 1998). FTD is
associated with unilateral or bilateral prefrontal degen-
eration with personality change, social comportment def-
icits, and impaired self-regulation. SD involves unilateral
or bilateral degeneration of the anterior and inferior
temporal cortex accompanied by a central semantic def-
icit and behavioral changes. PNFA involves left inferior
D 2008 Massachusetts Institute of Technology
Zeitschrift für kognitive Neurowissenschaften 20:10, S. 1839–1853
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
0
l
e
1
0
–
P
1
D
8
F
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
Ö
3
C
9
N
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
8
e
.
S
T
2
Ö
0
N
1
2
0
6
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
/
F
T
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
prefrontal degeneration and insidious and progressive
language abnormality, including reduced phrase length,
agrammatism, and effortful and halting speech. Contrast-
ing performance across these subtypes can illuminate
the contribution of damage centered on different brain
regions to autobiographical memory.
In contrast to standard laboratory testing of memory,
assessment of autobiographical memory is affected by
lack of control over encoding and the inherently per-
sonal nature of the memoranda. Autobiographical as-
sessment methods, daher, are more heterogeneous
than laboratory memory tasks, confounding interpreta-
tion of autobiographical memory findings across stud-
ies. In der vorliegenden Studie, we used advanced assessment
methods to assess autobiographical memory in patients
with FTLD. These allowed us to assess several hypoth-
eses concerning the behavioral and neuroanatomical
substrates of autobiographical memory.
The first hypothesis concerned the degree to which
episodic versus semantic autobiographical memory is
affected in FTLD. Episodic autobiographical memory re-
fers to recollection for events specific in time and place,
informa-
with accompanying mental and emotional
tion, including a subjective sense of personal continuity
across time (Tulving, 2002). This latter element is of par-
ticular relevance in FTLD where behavioral disturbances
suggest altered self-awareness (Miller et al., 2001). Se-
mantic autobiographical memory refers to recall of per-
sonal information not specific in time or place, wie zum Beispiel
where one was born, or repeated events, such as yearly
vacations at a particular location. Generally speaking,
studies have shown that patients with FTLD are im-
paired on measures of episodic autobiographical mem-
ory while they are preserved on semantic autobiographical
Erinnerung (Matuszewski et al., 2006; Piolino et al., 2003;
Hodges & Gurd, 1994).
Although episodic and semantic autobiographical
memory occur simultaneously during natural discourse
(Levine, Svoboda, Hay, Winocur, & Moscovitch, 2002), In-
struments for the assessment of autobiographical mem-
ory assume that memories can be wholly classified as
either ‘‘episodic’’ or ‘‘semantic.’’ Thus, these two ele-
ments of autobiographical memory are assessed sepa-
rately using interviews unmatched for content, difficulty,
and psychometric characteristics (z.B., Kopelman, Wilson,
& Baddeley, 1990) or tallied by sorting memories into
episodic or semantic categories (Piolino et al., 2007). Epi-
sodic memory is further quantified on the basis of ordinal
scale ratings that encompass both generic (z.B., repeated
or not temporally specific) and specific autobiographical
events that are subject to contamination by semantic
Verarbeitung.
In this study, we used the Autobiographical Interview
(Levine et al., 2002), which assumes that episodic and
semantic memory reflect distinct information processing
streams that can be simultaneously active during narra-
tive recall. This instrument yields independent, paramet-
ric estimates of episodic and semantic autobiographical
memory derived from within each memory at the time
of scoring rather than with separate interviews at the
time of testing or by wholesale assignment of recalled
narrative events to episodic or semantic categories. Das
measure has proven useful in dissociating episodic from
semantic memory in the elderly (St.-Jacques & Levine, 2007;
Levine et al., 2002),
in patients with medial temporal
lobe damage (Rosenbaum et al., 2008; Addis, Moscovitch,
& McAndrews, 2007; Steinvorth, Levine, & Corkin, 2005),
and in patients with semantic dementia (McKinnon, Black,
Müller, Moscovitch, & Levine, 2006). By quantifying inde-
pendent categories of autobiographical details, the Auto-
biographical Interview permits the assessment of profiles
of autobiographical content, including elevation of non-
episodic details extraneous to the central event as docu-
mented in aging (Levine et al., 2002), semantic dementia
(McKinnon et al., 2006), and frontal dysfunction (Levine,
2004). Based on these studies, we predicted an elevation
of nonepisodic autobiographical content (semantic and
other extraneous details) in FTLD, in addition to a decre-
ment of episodic content. As FTLD is a progressive dis-
Befehl, we assessed this pattern not just cross-sectionally,
but longitudinally, by retesting patients after 1 Zu 2 Jahre.
Decline in episodic, but not semantic, autobiographical
memory over time would suggest that the former is spe-
cifically affected by progressive volume loss in regions af-
fected by FTLD.
The second issue addressed in this study is the role of
retrieval support in autobiographical memory. Retrieval
support bolsters autobiographical memory recall in healthy
Erwachsene (Levine et al., 2002) and in patients with FTLD
(McKinnon et al., 2006; Moss, Kopelman, Cappelletti, Von
Mornay Davies, & Jaldow, 2003; Hodges & Gurd, 1994),
possibly by compensating for executive deficits affecting
strategic search and retrieval of remote information (z.B.,
Craik & McDowd, 1987). Methods of evoking auto-
biographical memories vary widely in the amount of re-
trieval support they provide, confounding comparison of
findings across studies. In this study, we assessed auto-
biographical memory under both low and high levels of
retrieval support to allow direct comparison of the effects
of this manipulation. We also examined the relationship
of neuropsychological test performance to autobiograph-
ical retrieval at these different levels of retrieval support.
To our knowledge, there are no studies of autobiog-
raphical memory in PNFA. Patients with PNFA compose
an interesting comparison group as this condition is less
likely to be associated with disturbances of the self such
as loss of insight and social conduct disorder than are
FTD or SD (Rosen et al., 2006). Given the relationship
between self-related information processing and auto-
biographical memory (Tulving, 2002; Conway & Pleydell-
Pearce, 2000), it was predicted that PNFA patients would
show preservation of episodic autobiographical memory
relative to the other FTLD patients. Andererseits,
the left inferior frontal region affected in PNFA is part
1840
Zeitschrift für kognitive Neurowissenschaften
Volumen 20, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
0
l
e
1
0
–
P
1
D
8
F
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
Ö
3
C
9
N
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
8
e
.
S
T
2
Ö
0
N
1
2
0
6
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
T
.
.
F
/
.
Ö
N
1
8
M
A
j
2
0
2
1
of a core autobiographical network in the functional
neuroimaging literature (Svoboda, McKinnon, & Levine,
2006). Given this region’s association with strategic re-
trieval operations (Simons & Spiers, 2003), deficits in
PNFA patients, if present, should be limited to the low
retrieval support condition that places greater demands
on strategic retrieval.
Endlich, we investigated individual differences in auto-
biographical memory performance as a factor of regional
atrophy quantified from patients’ high-resolution struc-
tural magnetic resonance imaging (MRT) scans con-
current to testing. Such data can provide important
adjunctive information to functional neuroimaging stud-
ies by identifying regions necessary for autobiographical
Erinnerung, as opposed to those that are simply engaged
by it. Few studies have assessed quantified cortical dam-
age effects on remote autobiographical memory, mit
results implicating regions across the cortical mantle
(z.B., McKinnon et al., 2007; Eustache et al., 2004;
Kopelman et al., 2003; Eslinger, 1998; Rubin & Greenberg,
1998),
lobe structures
(Rosenbaum et al., 2008; Moscovitch et al., 2005), al-
though this is disputed (Squire & Bayley, 2007).
including medial
zeitlich
In FTLD, the integrity of the medial temporal lobes
has been related to mnemonic processing (So¨derlund,
Black, Müller, Freedman, & Levine, 2008; Simons, Graham,
et al., 2002; Simons, Verfaellie, et al., 2002). Very general
evidence in support of a role for the frontal lobes has
been derived from neuropsychological test data and
diagnostic grouping (Matuszewski et al., 2006; Simons,
Verfaellie, et al., 2002). In a recent study of frontal-variant
FTD patients, florodeoxyglucose (FDG) uptake in the left
orbito-frontal and anterior temporal cortex was correlated
with autobiographical specificity (Piolino et al., 2007).
In der vorliegenden Studie, we used multivariate statistical
analyses of quantitated regional volumes derived from
structural MRI taken concurrent to testing to identify pat-
terns of regional volume loss across the brain associated
with indices of episodic and semantic autobiographical
memory in FTLD. In line with evidence supporting a role
for the medial temporal lobes and associated structures in
episodic autobiographical memory retrieval (Svoboda
et al., 2006; Moscovitch et al., 2005), we predicted that
volume loss in these regions would be strongly and spe-
cifically associated with lifespan retrieval of episodic
autobiographical details. We also predicted distributed
involvement of other regions (particularly frontal and
posterior regions given the strategic and visuospatial ele-
ments of autobiographical memory).
METHODEN
Subjects
FTLD Patients
Patients were recruited from dementia clinics at Baycrest
Centre and Sunnybrook Health Sciences Centre in Tor-
onto and at the University of California at San Francisco
Medical Center. FTLD diagnosis followed the consensus
criteria set out by Neary et al. (1998), which delineates
FTD, PNFA , and SD. Patients with significant aphasia,
neglect, or other focal neurological disturbance or se-
vere cognitive or physical disability that interfered with
testing were excluded. PNFA patients were in the early
stages of disease and, daher, had sufficient residual
speech capacity to participate.
We tested 22 Patienten. Eight met criteria for FTD and
five met criteria for PNFA. Nine patients met criteria for
both FTD and SD (Snowden, Neary, & Mann, 2007) Und
are therefore designated as FTD/SD. Two patients meet-
ing criteria for SD have been reported as part of a sep-
arate study (McKinnon et al., 2006). To date, 10 Patienten
in our sample have come to autopsy. These cases con-
firmed the presence of pathology consistent with FTLD,
including ubiquitin-positive, tau-negative inclusions with
or without degeneration of the motor neurons in some
Fälle, tau-positive Pick bodies inclusions with or with-
out a-synuclein inclusions, or progressive supranuclear
palsy and cortical basal degeneration, which can also
manifest as tauopathy in other cases (McKhann et al.,
2001). There were no significant differences across
groups for age, Ausbildung, duration of illness, or Mini
Mental State Examination (siehe Tabelle 1). Seventeen of
our patients (6 FTD, 7 FTD/SD, Und 4 PNFA) received
high-resolution structural MRI scans as part of the test-
ing protocol. The background characteristics of these
patients were similar to the full sample. Follow-up test-
ing was conducted on eight patients (5 FTD, 1 FTD/SD,
Und 2 PNFA) 1 Zu 2 years after initial assessment. Seven
of these eight patients received repeat MRI scans at the
same time as their follow-up testing.
Comparison Groups
A comparison group for behavioral assessment was
zusammengesetzt aus 16 healthy adults with no history of neu-
rological or psychiatric illness and free from medica-
tion known to affect cognitive functioning. They were
matched to the FTLD patients in terms of age (M =
58, SD = 9.2) and education (M = 16.4, SD = 3.0).
A separate scanning comparison group (n = 10), Auch
matched to the FTLD patients in terms of age (M = 63,
SD = 10.5) and education (M = 17, SD = 3.6), was used
for the purposes of assessing volume reductions in our
FTLD sample and for constructing an MRI template (sehen
below). These subjects were not tested behaviorally and
they were not included with the patients in the brain–
behavior analyses. Exclusion criteria included prior neu-
rological or systemic disease that could affect cogni-
tion, prior psychiatric hospitalization or treatment with
psychiatric medication for greater than 6 weeks, prior
significant alcohol/drug abuse, and significant develop-
mental disabilities.
McKinnon et al.
1841
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
0
l
e
1
0
–
P
1
D
8
F
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
Ö
3
C
9
N
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
8
e
.
S
T
2
Ö
0
N
1
2
0
6
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
F
T
.
.
.
/
Ö
N
1
8
M
A
j
2
0
2
1
Tisch 1. Characteristics of FTLD Patients and Comparison Subjects
N
Ageb
Comparison
FTD
16 (5 M)
58 (9.2)
8 (5 M)
59 (6.0)
FTD/SD
9 (2 M)
59 (9.4)
PNFA
5 (3 M)
66 (10.4)
Education (Jahre)
16.4 (3.0)
15.5 (4.0)
16.5 (3.0)
17.3 (3.0)
Duration of illness (Jahre)
MMSE
N/A
N/A
3.0 (2–10)
4.0 (2–6)
2.5 (1–5)
25.9 (3.3)
28.0 (1.9)
27.8 (1.5)
Follow-up FTLDa
8 (5 M)
61 (6.2)
15.1 (3.8)
3.0 (2–7)
24.4 (3.4)
MMSE = Mini Mental State Examination.
aThese patients comprise five FTD, one FTD/SD, and two PNFA patients that were retested 1 Zu 2 years after their initial assessment.
bStatistics are means and standard deviations, except for duration of illness, where median and range are reported.
Verfahren
The Autobiographical Interview
Event selection and instructions. The Autobiographi-
cal Interview was administered as described by Levine
et al. (2002), with slight modifications. Subjects were
asked to provide a detailed description of a significant
personal event from each of five life periods: early child-
hood (to age 11 Jahre), teenage years (ages 11–17 years),
early adulthood (ages 18–35 years), middle age (35–
55 Jahre), and the past year. Subjects were instructed to
recall an event that occurred at a specific time and place.
In cases where subjects were unable to generate a specific
life events (z.B.,
event independently, a list of typical
losing something important) was presented to assist in
event retrieval.
In order to examine
Conditions of retrieval support.
facilitative effects of retrieval support on memory, Wir
manipulated the level of structure available to subjects
across three conditions: recall, general probe, and spe-
cific probe (Levine et al., 2002). At recall, subjects spoke
about the event extemporaneously without any inter-
ruption from the examiner, continuing until it was evi-
dent that they had reached a natural ending point. Nach
an event was recalled, general probes were used to clar-
ify instructions and to encourage greater recall of details.
If general probing did not elicit a specific event, Die
subject was given the option of selecting a different
event that was more likely to result in successful recall.
General probes were limited to nonspecific statements
or repetitions of the instructions. At the specific probe
Phase, a structured interview was administered that was
designed to elicit additional contextual details. In order
to prevent the specific probe process from contaminat-
ing recall of subsequent memories, specific probing was
administered after all five events were recounted under
the recall and general probe conditions. Subjects’ de-
scriptions of the selected events were audio-recorded
for later transcription and analysis.
Following probing, subjects were asked to rate the fol-
lowing on a 6-point scale: importance (both at the time
of the event and at the time of testing), visualization,
experienced emotion at the time of the event, and fre-
quency of reactivation (thinking or talking about the
Ereignis). These ratings were unavailable for five FTD pa-
tients. There were no significant group differences in
Bewertungen; these data will not be discussed further.
Text segmentation and categorization. Following
transcription, each memory was segmented into infor-
mational bits or details. Each detail was then classified
according to the procedure outlined in Levine et al.
(2002). Briefly, details were defined as ‘‘internal’’ or epi-
sodic and assigned to one of five categories (Ereignis, Ort,
Zeit, perceptual, and emotion/thought) if they were
related directly to the main event described, were spe-
cific to time and place, and conveyed a sense of episodic
re-experiencing. Ansonsten, details were considered ‘‘ex-
ternal,’’ and consisted of semantic facts (factual infor-
mation or extended events that did not require
recollection of a specific time and place), autobiographical
events tangential or unrelated to the main event, repe-
titions, or other metacognitive statements (‘‘I can’t re-
member.’’) or editorializing (‘‘It was the best of times.’’).
Details were tallied for each category and summed to
form internal and external composites, which were the
main variables of interest in the present study. Scoring
was done separately for each condition (recall, allgemein
probe, specific probe), but scores were analyzed cumu-
latively across levels of recall with general probe and
specific probe details added to details generated from
the prior condition.
To avoid bias in scoring, subjects’ memories were placed
in a common pool and scored at random by seven expe-
rienced scorers who had achieved high interrater reliability
(see Levine et al., 2002) and who were blind to group.
Neuropsychological test results are presented in Table 2.
All neuropsychological tests showed a significant effect
of group (Fs ranged from 4.1 Zu 11.3, ps ranged from
<.001 to <.05). The pattern of group differences was
consistent with expectation given the literature on neu-
ropsychological differences across subgroups of patients
with FTLD. Patients with FTD were impaired on tests
of executive functioning and inventories of behavioral
change due to frontal damage while being preserved on
1842
Journal of Cognitive Neuroscience
Volume 20, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
f
.
.
t
/
.
o
n
1
8
M
a
y
2
0
2
1
Table 2. Neuropsychological Test Performancea
FTD (n = 8)
FTD/SD (n = 9)
PNFA (n = 5)
Comparison (n = 16)
Trail Making Test, Part Ab
Trail Making Test, Part B
Phonemic word list generation (FAS)
WCSTc perseverations
WCST categories
Dysexecutive questionnaired
Frontal Behavior Inventorye
Pyramids and Palm Treesf
Boston Naming Testg
11.4 (6.7)
5.4 (3.6)**
9.3 (2.4)
29 (11)*
59 (48)*
5 (2.5)*
24 (11)*
49 (34)
2.8 (4.0)*
2.2 (2.5)*
23 (20)*
43 (10.8)
28 (22)*
32 (13)
47.3 (57.2)
39 (12)****
50 (9)
31 (14.5)****
9.6 (3.7)
9.8 (4.9)
25 (13)*
26 (14)
6.2 (2.5)
12.3 (2.2)
10.4 (3.5)
46 (16)
21 (10)
6.9 (3.1)
(cid:2)4.4 (10.7)***
(cid:2)3.2 (7.8)
11 (1.8)***
51 (1)
54 (7)
N/A
N/A
N/A
aMeans and standard deviations are reported.
bFour patients received the Trail Making Test from the Delis–Kaplan Executive Function System (D-KEFS; Delis, Kaplan, & Kramer, 2001). The
remaining patients and comparison subjects received the standard Trail Making Test (Army Individual Test Battery, 1944). All scores were converted
to standard scores using the MOANS normative data (Ivnik, 1996) or the D-KEFS manual. Trails A: Control, n = 15. Trails B, FTD, n = 7.
cWisconsin Card Sorting Test (Milner, 1963).
dBurgess et al., 1996. Other score minus self score (higher scores indicate greater executive problems with less insight).
eFrontal Behavior Inventory (Kertesz, Davidson, & Fox, 1997).
fPyramids and Palm Trees—picture version. FTD/SD: n = 4.
gBoston Naming Test—when necessary, scores were prorated from 15- or 30-item version. FTD, n = 5. FTD/SD, n = 6. PNFA, n = 4.
*Significantly different from comparison group.
**Significantly different from comparison and FTD groups.
***Significantly different from FTD and FTD/SD groups.
****Significantly different from FTD and PNFA.
tasks of semantic retrieval and naming. FTD/SD patients
were also impaired on executive tasks and behavioral in-
ventories, with additional evidence of semantic and lexi-
cal retrieval deficits (although these data are limited by
low n) and speeded information processing deficits.
PNFA patients show low verbal fluency and preservation
on executive tasks and behavioral inventories.
Statistical Analyses
FTLD patients’ data for certain Autobiographical Inter-
view indices were markedly positively skewed. This skew-
ness could not be corrected through transformation. We
therefore applied a Winsorization procedure to the data
by which outliers (i.e., scores exceeding 1.5(cid:1) the intra-
quartile range above the third quartile or below the first
quartile) were rescaled to be ±2.5 SD from the mean
(calculated excluding outliers), allowing the mainte-
nance of extreme observations without unduly influenc-
ing statistical estimates or sample sizes.
We next examined detail composite production across
each of the five time periods tested using a 4 (cid:1) 2 (cid:1) 5
mixed-design analysis of variance that treated group
(FTD, FTD/SD, PNFA, and comparison) as a between-
subjects factor and detail composite (internal, external)
and lifetime period (early childhood, teenage years, early
adulthood, middle age and past year) as within-subjects
factors. There were no interactions involving the internal
and external composites when these were incorporated
into the lifetime period analyses. We found main effects
of lifetime period reflecting a recency effect (i.e., greater
recall from the past year; Rubin & Schulkind, 1997). As this
effect did not interact with group, it will not be discussed
further (for a similar finding, see Piolino et al., 2003).
A second analysis examined performance on the
category-specific measures of autobiographical retrieval
using a 4 (cid:1) 9 mixed-design analysis of variance that
treated group (FTD, FTD/SD, PNFA, and comparison) as
a between-subjects factor and category (internal event,
time, place, perceptual and emotion/thought details;
external event, other, and semantic details and repeti-
tions) as a within-subjects variable. To examine inter-
val change in Autobiographical Interview performance
among patients retested after 1 to 2 years, we used a
2 (cid:1) 2 repeated measures design treating detail type (in-
ternal, external) and test session (Session 1, Session 2)
as within-subjects factors.
Tukey’s Honestly Significant Difference post hoc test
was used for follow-up pairwise comparisons of between-
subjects variables. Where required, the Greenhouse–Geisser
correction was applied to effects involving repeated mea-
sures. All analyses were performed with alpha set at .05.
McKinnon et al.
1843
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
t
/
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Correlations between neuropsychological assessment
test scores and performance on the internal and exter-
nal detail composites were assessed nonparametrically
(Spearman’s (cid:1) [rho]) due to nonnormality of the data.
MRI Scan Acquisition
Patients and comparison subjects were scanned on 1.5-T
scanners (Toronto: Signa, General Electric Medical Sys-
tems, Waukesha, WI; San Francisco: Magnetom VISION
system, Siemens, Iselin, NJ ) with similar in-plane reso-
lution. The Toronto protocol involved axial acquisitions
using spoiled gradient-echo T1-weighted 3-D volume im-
aging (TR/TE/flip angle = 35 msec/5 msec/358, 1.0 NEX,
acquisition matrix = 256 * 256; 124 slices, slice thickness =
1.3 mm; FOV = 22 cm), as well as spin echo, proton
density, and T2-weighted images (TR/TE = 3000 msec/
30 msec, 80 msec, 0.5 NEX, acquisition matrix 256 (cid:1) 192,
slice thickness = 3 mm; FOV = 20 cm). The San Francisco
protocol also applied a double spin-echo sequence (TR/
TE1/TE2 = 5000/20/85 msec, 51 contiguous 3 mm axial
slices covering the entire brain and angulated (cid:2)108 from
the AC–PC line). Volumetric T1-weighted gradient-echo
MRI were achieved using the MP-RAGE sequence (TR/TE/
TI = 10/4/300 msec, 158 flip angle, 1.5 mm slab thickness)
in coronal orientation perpendicular to the double spin-
echo sequence.
Image Processing
Brain MRI data were analyzed via an updated version
of our previously reported image processing pipeline
(Dade et al., 2004; Kovacevic et al., 2002). The main
modification to this protocol involves template match-
ing, allowing for comparison of individual images to a
standard image and facilitating automation of previously
semi-automated steps. The first step in the pipeline was
to create an unbiased nonlinear average of T1-weighted
images from the age- and education-matched scanning
comparison group using a modification of an algorithm
previously developed for mouse brain MRI (Kovacevic
et al., 2005). Each subject’s T1-weighted image was
then registered to the template brain (Woods, Grafton,
Holmes, Cherry, & Mazziotta, 1998; Woods, Grafton,
Watson, Sicotte, & Mazziotta, 1998) preserving the orig-
inal size of the brain while standardizing the position
and orientation. Images were resampled into template
space using windowed sinc interpolation. Template
matching was accomplished via nonlinear registration
of T1-weighted images to the template image (Collins
& Evans, 1997). Removal of nonbrain tissue from the
image incorporated thresholding information derived
from the PD- and T2-weighted images, facilitating the
distinction between the dura mater and the gray matter
(Kovacevic et al., 2002). This is contrasted to methods of
brain extraction on the basis of the T1-weighted image
that emphasize the cortical surface, inconsistently preserv-
ing subdural cerebrospinal fluid (CSF). The voxels on the
T1-image were then classified as representing gray matter,
white matter, or CSF using an automated tissue classifica-
tion method that corrects for radio-frequency inhomoge-
neity inherent to MR scanning (Kovacevic et al., 2002).
A modified Semi-Automated Brain Region Extraction
(SABRE) (Dade et al., 2004) method was then used to
create ROIs on the template brain. This method involves
manual identification of 15 landmarks and tracing of the
cingulate gyrus on the template brain. Based on iden-
tification of the edges of the brain and the anterior
and posterior commissures, a Talairach-like (Talairach
& Tournoux, 1988) grid is automatically created. The
algorithm uses this grid along with the landmark co-
ordinates to divide the brain into 38 regions (19 per
hemisphere; see Figure 1). Nonlinear deformation field
matching of the template to individual images was used
to customize these regions to fit each subject’s brain
anatomy (as opposed to transforming images to fit the
template, which can distort interindividual topographi-
cal variability). Regional gray matter, white matter, and
CSF volumes were adjusted for total intracranial capacity
using a regression-based method (Arndt, Cohen, Alliger,
Swayze, & Andreasen, 1991). As our segmentation pro-
tocol is flexible across different T1-weighted contrasts,
no adjustment was necessary to accommodate images
acquired from different scanners.
Our tissue compartment segmentation and SABRE
software are particularly well suited to analysis of brains
with atrophy, as they do not require spatial transforma-
tion that can distort interindividual topographical vari-
ability. They have thus been successfully applied across
a variety of patient samples (e.g., Levine et al., 2008;
Bocti, Rockel, Roy, Gao, & Black, 2006; Gilboa et al.,
2005; Feinstein et al., 2004).
Image Analysis
Partial least squares (PLS) is a flexible multivariate tech-
nique that has been extensively applied to brain imag-
ing data (McIntosh, Chau, & Protzner, 2004; McIntosh,
Bookstein, Haxby, & Grady, 1996). In general terms, PLS
is a multivariate analysis technique for relating two sets
of variables to each other. In the present application, it
was used to identify patients’ patterns of volume loss re-
lated to measures derived from the Autobiographical
Interview. It is unbiased in that there are no a priori as-
sumptions about structure–function correlations. Because
PLS considers the brain as a whole, it is well suited to the
detection of distributed patterns of volume loss that co-
vary with test performance.
In the first step of the PLS analyses, correlations were
computed between the brain imaging data (i.e., regional
gray matter, white matter, and CSF volumes) for the full
sample of patients (without respect to diagnosis) and
Autobiographical Interview category scores (i.e., five in-
ternal detail categories and four external detail catego-
1844
Journal of Cognitive Neuroscience
Volume 20, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
t
.
/
.
f
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
/
t
.
.
f
o
n
1
8
M
a
y
2
0
2
1
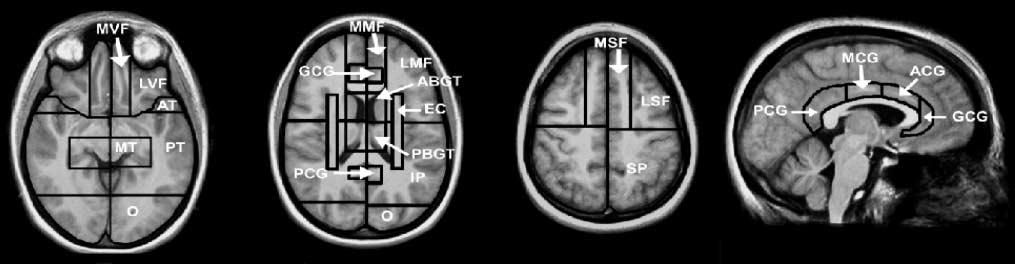
Figure 1. SABRE regional cortical divisions in axial and sagittal views. LSF = lateral superior frontal; MSF = medial superior frontal; LMF =
lateral middle frontal; MMF = medial middle frontal; LVF = lateral ventral frontal; MVF = medial ventral frontal; GCG = genual cingulate
gyrus; ACG = anterior cingulate; MCG = middle cingulate gyrus; PCG = posterior cingulate gyrus/retrosplenial; AT = anterior temporal;
MT = medial temporal; PT = posterior temporal; O = occipital; ABGT = anterior basal ganglia/thalamus; PBGT = posterior basal ganglia/
thalamus; EC = external capsule/corona radiata; IP = inferior parietal; SP = superior parietal.
ries) for both recall and specific probe, collapsed across
time period. The goal of this analysis was to assess
brain–behavior correlations in FTLD by treating quanti-
fied regional brain atrophy as an independent variable,
rather than diagnosis, which is an imprecise proxy for
regional brain changes.
Singular value decomposition was then applied to the
correlation matrix to identify latent variables [LVs] that
indicated optimal relations between patterns of regional
brain volume loss and test performance. Saliences (sim-
ilar to factor loadings in factor analyses) reflected the
contribution of individual brain volumes and behavioral
measures to the LV (McIntosh et al., 1996, 2004). Mul-
tiplication of each region’s salience with its volume and
summing over all volumes gives a ‘‘brain score’’ for each
patient on a given LV that indicates the degree to which
the pattern of volumetric changes identified by the LV is
expressed in each patient. Similarly, each patient’s con-
tribution to the behavioral aspect of the LV (‘‘behavior
score’’) is derived by multiplying the salience for each
test by the patient’s test score. The relationship be-
tween the two aspects of the LV (patterns of regional
brain volume loss and test performance) can be exam-
ined by plotting the brain scores against the behavior
scores on a patient-by-patient basis, allowing us to ex-
amine the contribution of diagnosis to the pattern of
brain–behavior relationships (see Figure 5C).
The statistical significance of each LV was assessed by
1500 permutation tests (Edgington, 1980), in which the
observations are randomly reordered without replace-
ment to calculate the probability of each LV having oc-
curred by chance. The stability of each brain region’s
salience contribution to the LV was determined through
bootstrap resampling (subjects were resampled 500 times;
McIntosh et al., 1996; Wasserman & Bockenholt, 1989).
Brain regions in the singular images were considered
reliable if they had a ratio of salience to standard error
(hereafter referred to as the boostrap ratio), interpreted
similar to a Z-score (McIntosh et al., 2004; Efron &
Tibshirani, 1986) greater than 3, corresponding to 99.9%
confidence limits. The bootstrap procedure yields 99%
confidence intervals around the correlations between test
scores and the pattern of regional brain volume changes.
Because image-wide statistical assessment is done in a sin-
gle analytic step, no correction for multiple comparisons
across brain regions is required.
For the purposes of characterizing the degree of brain
atrophy in our patient sample, the 19 SABRE regions per
hemisphere were reduced to eight (ventral frontal, dorsal
frontal, anterior, medial, and posterior temporal, inferior
parietal, superior parietal, and occipital). Reduction in
parenchymal volumes (gray + white matter) were as-
sessed via planned comparisons between the FTD, FTD/
SD, and PNFA subgroups and the 10 matched scanning
comparison subjects. Interval change between the first
and second scans for the seven rescanned FTLD patients
was also assessed via planned comparisons.
RESULTS
Patterns of Brain Atrophy in FTLD Patients
frontal regions bilaterally,
FTLD patients showed a characteristic pattern of signif-
icantly reduced parenchymal volumes maximal over the
frontal and temporal lobes. For illustration, the left dor-
solateral frontal and posterior temporal volumes are
displayed in Figure 2. The FTD group had volume loss
over the dorsolateral
left
( p < .009) greater than right ( p < .05), as well as sig-
nificant volume loss in the right posterior temporal re-
gion ( p < .005). The FTD/SD group had marked volume
loss over anterior, posterior, and medial temporal lobe
sectors ( ps ranging from .0001 to .02) as well as all fron-
tal sectors ( ps ranging from .001 to .007). Volume loss in
PNFA patients was more restricted, maximal in the left
dorsolateral region ( p < .001), but also in the right pos-
terior temporal and right inferior parietal regions ( ps <
.05). It should be noted that only four PNFA patients
McKinnon et al.
1845
of autobiographical memory. A significant Detail cate-
gory (cid:1) Group interaction [F(24, 272) = 7.03, p < .001]
was due to lower production of internal details in the
FTD and the FTD/SD groups than in comparison subjects
for all internal detail categories ( ps < .001–.05) except for
time, which was uniquely impaired in the FTD/SD group
( p < .05), and for place, which was uniquely impaired in
the FTD group ( p < .05). The FTD/SD group produced
more semantic details than did the comparison group
( p < .05). Significant differences between the PNFA pa-
tients and comparison subjects were limited to the internal
perceptual and thought/emotion categories ( ps < .05).
Specific Probe
Following specific probing, the significant Detail type (cid:1)
Group interaction remained [F(3, 128) = 8.38, p < .001],
with greater production of internal details by the com-
parison group as compared to the FTD and FTD/SD
groups ( p < .01; see Figure 3). Whereas the PNFA group
produced significantly fewer internal details in the recall
condition than did comparison subjects, after specific
probing, the PNFA group’s production of internal details
differed neither from the comparison subjects nor from
the FTD group. There were again no group differences
for external details. The main effect of group was not
significant, indicating that all groups generated proto-
cols of similar length.
The Detail category (cid:1) Group interaction [F(24, 272) =
4.49, p < .001] remained significant at specific probe.
Whereas the FTD/SD group produced fewer internal de-
tails than did the comparison group for all internal detail
categories ( ps < .001–.01) except for time, where this
group was unimpaired, the FTD group was impaired rel-
ative to the comparison group for the perceptual and
thought/emotion internal categories only ( ps < .01).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
.
f
.
t
/
o
n
1
8
M
a
y
2
0
2
1
Figure 3. Mean number of internal (episodic) and external
(nonepisodic) details generated by comparison subjects, FTD, FTD/SD,
PNFA, on the Autobiographical Interview (specific probe phase).
FTD (n = 8) and FTD/SD (n = 9) patients generated fewer internal
details and, in the case of FTD/SD, more external (semantic) details,
whereas PNFA patients (n = 5) were not significantly different
from comparison subjects. *Significantly different from comparison
subjects, p < .01.
Figure 2. Regional parenchymal volumes in FTLD patients and
comparison subjects in left dorsolateral prefrontal and in left posterior
temporal regions. Comp. = comparison subjects. Vertical lines
indicate 95% confidence intervals. Group means are indicated by
bold horizontal lines. Relative to comparison subjects, patients in
all groups showed significant decline in left dorsolateral prefrontal
volume, whereas only the FTD/SD subgroup had significant volume
loss in the left posterior temporal region.
were scanned, one of whom was left-handed with pri-
marily right-sided degeneration, suggesting crossed lat-
eralization of language. Interval change for the seven
rescanned FTLD patients was statistically significant over
all regions of interest ( ps ranged from .0005 to .03).
Autobiographical Memory
Recall
A significant interaction between detail type and group
[F(3, 132) = 14.18, p < .01], during the free recall phase
of the Autobiographical Interview, was due to reduced
production of internal details in the FTD, FTD/SD and
PNFA groups relative to comparison subjects ( ps < .001,
.001, and .05 for the FTD, FTD/SD, and PNFA groups,
respectively); there were no group differences for exter-
nal details. The main effect of group was not significant
( p > .05), indicating that the patients’ reduction in in-
ternal details was not attributable to an overall reduction
in protocol length among the patient groups.
Analysis of individual detail categories provided more
information concerning group differences in the elements
1846
Zeitschrift für kognitive Neurowissenschaften
Volumen 20, Nummer 10
The FTD/SD group continued to produce more seman-
tic details than did the comparison group ( P < .05).
Significant differences between the PNFA patients
and comparison subjects were limited to the thought/
emotion category ( p < .01).
One-year Follow-up Testing
Among the eight patients retested 1 year later, there was
a significant Test session (cid:1) Detail type interaction at
specific probe [F(1, 7) = 8.77, p < .05] that was due to a
reduction of internal ( p < .005) details across the test
sessions (see Figure 4). External details increased across
test sessions, although this difference fell short of statis-
tical significance due to high variability. There were no
significant main effects involving test session or detail
type at recall or specific probe. The Test session (cid:1) Detail
type interaction at recall was not significant.
Relation of Autobiographical Memory to
Neuropsychological Test Scores
Both FTLD patients and comparison subjects showed a
positive relation between performance on the phonemic
word list generation and scores on the internal detail
composite at recall [(cid:1) (21, 16) = 0.54 and 0.64, for pa-
tients and comparison subjects, respectively, ps < .02
and .01]. There was a correlation between Trail Making,
Part A and internal details at recall
in FTLD patients
[(cid:1) (21) = 0.45, p < .05]. There were no significant cor-
relations between neuropsychological test performance
and internal details at specific probe. External details at
specific probe were negatively correlated with Trail Mak-
ing, Part B in patients [(cid:1) (20) = (cid:2)0.47, p < .05]; there
were no other significant correlations between neuro-
psychological tests and the external detail composites.
Figure 4. Effects of FTLD (n = 8) progression on internal (episodic)
and external (nonepisodic) autobiographical details from the
Autobiographical Interview (specific probe phase). *Significantly
different from Session 1, p < .005. Error bars represent standard
error of the mean.
Relation to Regional Brain Atrophy
The PLS analysis allowed us to examine the pattern of
regional volume loss associated with patients’ impaired
performance on the Autobiographical Interview. One LV
was identified ( p = .05) by permutation test. This LV
accounted for 41% of the covariance between test scores
and regional volumes. The pattern of Autobiographical
Interview scores associated with this LV indicated a sepa-
ration of episodic from nonepisodic details concentrated
at specific probe (see Figure 5A). Specifically, increased
event, time, place, and perceptual details at specific
probe and event details at recall were related to increased
parenchymal volumes across the identified brain regions,
whereas increased semantic details and repetitions at
specific probe were related to decreased volumes.
Figure 5B depicts significant negative bootstrap ratios
for CSF (i.e., indicating increased parenchymal volume
positively associated with internal details and negatively
associated with external details) with foci
in the left
posterior and anterior temporal lobes and the medial
temporal lobes bilaterally. The left inferior parietal lobe,
the left posterior cingulate/retrosplenial region, and the
bilateral posterior basal ganglia regions also emerged as
significant in this analysis. There were additional signif-
icant positive bootstrap ratios for gray and white matter
(not displayed in the figure) that mirrored those de-
picted in Figure 5: left posterior temporal and left oc-
cipital gray and white matter, bilateral posterior basal
ganglia white matter, and left anterior basal ganglia
white matter. Significant negative bootstrap ratios were
observed for the left anterior cingulate gyrus and bilat-
eral internal capsule/corona radiata gray matter regions.
In Figure 5C, the brain scores (reflecting each pa-
tient’s contribution to the pattern of volume loss iden-
tified by the LV) and the behavior scores (reflecting each
patient’s contribution to the pattern of test scores iden-
tified by the LV) are plotted against each other. Al-
though the sample is small and there is overlap across
it can be seen that the lower
the diagnostic groups,
ends of the behavioral and brain score distributions
are mainly occupied by patients in the FTD/SD group,
whereas FTD and PNFA patients occupy the upper end.
DISCUSSION
Lifespan autobiographical episodic recall was impaired
in patients with FTLD relative to comparison subjects,
whereas semantic autobiographical memory was spared
or elevated in patients with FTLD. Narrative episodic
specificity was associated with the integrity of a left-
lateralized posterior network centered on the temporal
lobe. The specificity of the episodic relative to semantic
impairment was further supported by longitudinal data.
When the medial temporal lobes are spared, as in PNFA,
retrieval support is effective in increasing autobiograph-
ical recall to normal levels. Contrary to expectation, we
McKinnon et al.
1847
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
/
.
.
.
t
f
o
n
1
8
M
a
y
2
0
2
1
Figure 5. LV from PLS analysis
indicating the association
of performance on the
Autobiographical Interview
with patterns of regional brain
volume changes. (A) The
pattern of Autobiographical
Interview measures associated
with the LV, expressed as
correlations between test scores
and the pattern of brain volume
changes depicted in B. Error
bars represent 99% confidence
intervals. Measures with error
bars crossing the horizontal axis
did not significantly contribute
to the LV. This panel shows that
internal event details at recall
and internal event, place,
time, and perceptual details at
specific probe were positively
correlated with the pattern of
brain volume changes depicted
in B, whereas external semantic
and repetition details at the
specific probe phase were
negatively correlated with
the pattern of brain volume
changes. Ed = event details
(note that event details can be
either internal or external,
depending on whether they
concern the main event);
Perc = perceptual details;
Tho = thought details; Sem =
semantic details; Rep =
repetitions; Oth = other
details (e.g., metacognitive or
editorializing). (B) The pattern
of regional CSF volume
(expressed as color-coded
bootstrap ratios) associated
with the pattern of test scores
depicted in the top panel. The
color bar indicates the coding
scheme according to the
level of the bootstrap ratio,
interpreted similar to a Z-score.
Lower values (darker blue)
correspond to brain volume
decreases (i.e., less CSF, greater
parenchyma) associated with
higher internal details and lower
external details. Images were
thresholded at a bootstrap
ratio of 3.0, corresponding
approximately to p < .001.
Axial images are displayed in
radiological convention (right
hemisphere displayed on left
side of image). The right and
left cingulate volumes are
displayed on the right and left
side of the images, respectively.
Not pictured: Bootstrap ratios
for gray and white matter (see
text). (C) A scatterplot of individual patients’ brain scores (indicating the degree to which the pattern of atrophy identified by the LV is expressed in each
patient) and behavior scores (indicating the degree to which the pattern of test scores identified by the LV is expressed in each patient), with each
patient color-coded according to diagnosis, allowing for the appreciation of the how FTLD diagnosis contributes to the brain–behavior relationships.
1848
Journal of Cognitive Neuroscience
Volume 20, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
t
.
.
/
f
o
n
1
8
M
a
y
2
0
2
1
found no significant relation of frontal lobe volumes to
autobiographical memory retrieval. Overall, these find-
ings support a behavioral and neuroanatomical distinc-
tion between episodic and semantic autobiographical
memory. They further suggest that, contrary to standard
diagnostic criteria, memory function can be significantly
affected in FTLD.
Dissociating Episodic and Semantic
Autobiographical Memory across FTLD Diagnoses
Prior results indicating impaired episodic autobiograph-
ical memory and spared semantic autobiographical
memory in FTLD (Piolino et al., 2003; Nestor et al.,
2002) are based on the use of ordinal ratings that can be
elevated by inclusion of nonepisodic details that are re-
lated to the event, but do not require recreation of tem-
poral, spatial, and other contextual details. One approach
to this issue is to apply a binary transformation to the or-
dinal data that divides those receiving the highest ratings
(considered ‘‘strictly episodic’’) from those receiving
lower ratings (Matuszewski et al., 2006; Piolino et al.,
2003). Our approach is to extract estimates of episodic
and semantic elements from within each memory, under
the assumption that such elements occur simultaneously
to varying degrees within all naturalistic autobiographical
narratives and that these can be independently affected
by normal or pathological intersubject variables (e.g., ag-
ing, brain disease; Rosenbaum et al., 2008; Addis et al.,
2007; Steinvorth et al., 2005; Levine, 2004; Levine et al.,
2002) or by intrasubject manipulations (e.g., retrieval sup-
port, emotional state; St.-Jacques & Levine, 2007; Levine
et al., 2002).
The PNFA group was less impaired than the FTD and
FTD/SD groups. Indeed, retrieval support selectively
bolstered recall of episodic autobiographical details in
patients with PNFA such that they were not statistically
differentiated from comparison subjects. Mild episodic
autobiographical deficits in patients with PNFA are there-
fore likely attributable to strategic retrieval or speech ini-
tiation deficits that are ameliorated by retrieval support.
Patients with FTD and FTD/SD remained impaired rela-
tive to comparison subjects after retrieval support, al-
though effects across individual categories of episodic
details were more reliable among those with FTD/SD.
As described in more detail below, this may be attribut-
able to the constraining effects of medial temporal lobe
damage on episodic autobiographical recall. These re-
sults cannot be attributed to reduced speech output, as
there were no effects of overall detail production across
groups.
Patterns of spared and impaired autobiographical
memory across these groups are further clarified by con-
sideration of nonepisodic (external) autobiographical
details. Although there was no overall effect on external
details, detail category analyses revealed that semantic
autobiographical details were uniquely elevated in the
FTD/SD group, indicating that these patients produce an
excess of generic autobiographical information in addi-
tion to reduced richness of happenings, spatial referen-
ces, perceptual information, and mental states.
Patients with PNFA are distinguished from patients
with FTD by a relative preservation of insight and social
and personal conduct (Neary et al., 1998), an observa-
tion confirmed by these patients’ normal scores on in-
ventories of behavioral and social cognitive changes.
Although deficits in these areas are not core to the diag-
nosis of SD in its original formulation, there is evidence
that behavioral features in SD and FTD significantly over-
lap (Rosen et al., 2006; Bozeat, Gregory, Ralph, & Hodges,
2000). Our findings are consistent with the notion that
self-referential processing is fundamental to the capacity
for episodic autobiographical memory (Tulving, 2002;
Conway & Pleydell-Pearce, 2000), with both processes
spared in PNFA relative to FTD and SD.
It is acknowledged that interpretation of results may
be limited by low power as there were only five patients
in the PNFA groups. Yet these patients were differenti-
ated from the FTD and FTD/SD groups not just by de-
gree, but by their pattern of distribution of details across
internal and external categories, which paralleled that of
the comparison group. Although there were no signifi-
cant group differences for disease duration, PNFA pa-
tients may have tested at an earlier stage of disease
progression as those with significant aphasia were ex-
cluded. We directly assessed the effects of disease pro-
gression by retesting patients after 1 to 2 years,
where the pattern of reduced episodic but not semantic
autobiographical memory was enhanced, reinforcing the
specificity of episodic autobiographical memory loss in
FTLD. We did not have sufficient data to differentiate
disease progression effects according to diagnosis. Given
evidence that differences across FTLD diagnostic entities
blur with disease progression (Kertesz, McMonagle,
Blair, Davidson, & Munoz, 2005), it is likely that the au-
tobiographical profile in late PNFA is similar to that of
FTD and SD.
Relation of Autobiographical Memory
Performance to Patterns of Regional Volume Loss
For the purposes of analyzing the effects of diagnosis,
we sought to define groups with distinct clinical syn-
dromes. In our brain–behavior analysis, we directly as-
sessed the effects of individual differences in patterns of
brain atrophy across clinical diagnoses, rather than
inferring lesion location from diagnosis. The dissociation
between episodic and semantic autobiographical mem-
ory was further reinforced by the brain–behavior analy-
ses. Critically, the patterns of regional atrophy that
emerged in relation to this dissociation were not spec-
ified in a priori planned contrasts (as is typically done in
brain–behavior analyses, for example, by contrasting
groups with presumed focal patterns of damage), but
McKinnon et al.
1849
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
t
/
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
rather from unrestricted multivariate analyses of brain–
behavior correlations.
Regional volumes in a distributed network over the
temporal lobes, posterior subcortical regions, and left
inferior parietal and occipital regions were positively
related to four out of five indices of episodic autobio-
graphical memory at specific probe, as well as one (event
details) at recall. Volumes in these same regions were
negatively related to semantic details and repetitions at
specific probe. Thus, in FTLD, specificity and richness of
narrative production of lifespan autobiographical memory
are related to the integrity of a left-lateralized posterior
network centered on the temporal lobes.
Our findings are consistent with other patient and
functional neuroimaging studies emphasizing the di-
verse mnemonic, attentional, and multimodal sensory
processes that enable autobiographical re-experiencing
(Svoboda et al., 2006; Kopelman et al., 2003). In inter-
preting the brain behavior analysis, we emphasize CSF
volumes (i.e., the inverse of parenchymal volume). Gray
and white matter volume paralleled the CSF effects, al-
though not all regions identified in the CSF emerged as
significant in the gray and white matter analyses, and
some regions (left occipital lobe, left anterior basal gan-
glia) emerged in gray and white matter analyses, but not
CSF analyses. Although these were consistent with the
overall CSF pattern, the left anterior cingulate and in-
ternal capsule/corona radiata gray matter showed a re-
lationship opposite to the direction of the other regions
(i.e., negatively correlated with internal details, posi-
tively correlated with external details). As these regions
have relatively small volumes (e.g., mean gray matter
volume for the left anterior cingulate gyrus: 1777 mm3
vs. 41,166 mm3 for the left posterior temporal lobe),
they may be relatively unstable and will not be inter-
preted further.
Right and left medial temporal lobe volumes were re-
lated to autobiographical memory specificity. Our medial
temporal
lobe region included the hippocampus and
surrounding structures, regarded as critical for the bind-
ing of features required for recall of contextual details
(Eichenbaum, 2000), a distinction that holds within auto-
biographical memory (Spiers, Maguire, & Burgess, 2001;
Kapur, 1999; Cermak, 1985). The dependence of episodic
autobiographical memory on the medial temporal lobes
is reinforced by recent studies of patients with focal dam-
age in these regions and selective impairment on episodic
details as assessed by the Autobiographical Interview
(Rosenbaum et al., 2008; Addis et al., 2007; Steinvorth
et al., 2005; but see Bright et al., 2006; Kirwan, Bayley,
Galvan, & Squire, 2008; and Squire & Bayley, 2007, for
different results), as well as functional neuroimaging data
(see also Svoboda et al., 2006; Moscovitch et al., 2005).
The left posterior cingulate/retrosplenial region was
also significantly related to autobiographical memory
specificity. This region is connected to the hippocampus
and the prefrontal cortex (Morris, Pandya, & Petrides,
1999; Rosene & Van Hoesen, 1977), facilitating integra-
tion of mnemonic and higher level cognitive processing.
It is also directly connected to multiple thalamic nuclei
(Morris, Petrides, & Pandya, 1999) including the anterior
thalamus (Vogt, Pandya, & Rosene, 1987) that is part of a
midline diencephalic system involved in episodic mem-
ory (Aggleton & Brown, 1999). Damage to this region,
which is frequently activated in functional neuroimaging
studies of autobiographical memory (Svoboda et al.,
2006), can cause retrograde amnesia (Valenstein et al.,
1987).
The left anterior and posterior temporal regions were
strongly associated with episodic autobiographical mem-
ory retrieval, along with the left inferior parietal region.
These regions are also strongly associated with autobio-
graphical memory in functional neuroimaging studies
(Svoboda et al., 2006) due to retrieval of specific factual
information (anterior temporal cortex), semantic pro-
cessing (posterior temporal cortex), and visuospatial
processing (temporo-parietal junction). Occipital involve-
ment (derived from analyses of gray and white matter) is
consistent with the importance of visual processes in rec-
ollection (Rubin & Greenberg, 1998). The left-lateralization
of neocortical activation associated with autobiographical
memory and the present findings are inconsistent with
evidence of asymmetric right lateral temporal lobe in-
volvement in episodic autobiographical memory (Gilboa
et al., 2005; Eustache et al., 2004; Kopelman et al., 2003;
Eslinger, 1998; for exception, see Piolino et al., 2007).
Our findings cannot be explained by asymmetric damage
in our patients, as the left and right temporal regions
were similarly affected. As an alternative account, it is
noted that our findings were concentrated at the specific
probe phase as contrasted to prior studies that used the
autobiographical memory interview (AMI) (or variants
thereof ) that is more comparable to procedures in our
recall phase. Conversely, AMI probing of semantic auto-
biographical memory, which is highly structured, has
been related to left temporal volumes (Gilboa et al.,
2005; Eslinger, 1998). Thus, the left-lateralized findings
may indicate lack of access to queried specific autobio-
graphical details. Our patients with compromised ability
to retrieve such details produced an excess of nonspe-
cific semantic autobiographical details, reflecting com-
pensation, disinhibition, and impaired monitoring of
mnemonic output. The semantic autobiographical de-
tails elevated in association with left posterior pathology
are not equivalent to the central semantic concepts
affected by temporal lobe damage in SD. Rather, they
reflect autobiographical information not specific in time
and place that can be preserved in association with left
lateral temporal damage (Westmacott & Moscovitch,
2003; Snowden, Griffiths, & Neary, 1994).
The regions related to autobiographical memory per-
formance composed only part of the overall pattern of
volume loss in our patients, which included the signif-
icant frontal lobe changes. Given the established role
1850
Journal of Cognitive Neuroscience
Volume 20, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
/
.
t
.
f
.
o
n
1
8
M
a
y
2
0
2
1
of the prefrontal cortex in autobiographical memory
(McKinnon et al., 2007; Svoboda et al., 2006; Kopelman
et al., 2003), it is surprising these regions did not emerge
as related to behavior in our analysis. SABRE-defined
frontal volumes also were not related to autobiograph-
ical memory performance in patients with Alzheimer’s
disease (Gilboa et al., 2005). On the other hand, left
orbito-frontal FDG uptake was related to autobiograph-
ical memory performance in patients with frontal-variant
FTD (Piolino et al., 2007; for a related study in Alzheimer
patients, see Eustache et al., 2004), a discrepancy at-
tributable to interstudy differences in patient selection,
assessment methods, and imaging platform. Although
performance on neuropsychological tests of executive
functioning has been related to autobiographical mem-
ory performance in FTLD (Matuszewski et al., 2006), we
found that speeded neuropsychological tests (phonemic
word list generation and Trail Making, Part A) were
related to episodic autobiographical memory, but not
a widely used untimed test of executive functioning, the
Wisconsin Card Sorting Test. These correlations were
limited to the free recall phase of the Autobiographical
Interview. These findings suggest a relation between
lexical semantic retrieval and generalized cognitive func-
tioning and strategic autobiographical memory retrieval,
perhaps due to the cognitive demands involved in cue
specification at recall, whereas the structured cueing at
the specific probe phase lessens the demands on these
processes. At specific probe, the negative relationship
between Trail Making, Part B and external details sug-
gests that an excess of nonepisodic autobiographical
information may be related to higher-level attentional
deficits, especially among FTD/SD patients, who per-
formed poorly on Trail Making, Part B and had signifi-
cantly elevated semantic details.
In another study using an overlapping sample of FTLD
patients (So¨derlund et al., 2008), performance on labo-
ratory tasks of episodic memory, including source mem-
ory and remember/know judgments, was associated with
the integrity of the left temporal lobe and not the frontal
lobes. These findings suggest that although retrieval
can be reliably associated with temporal lobe damage
in FTLD, the functional localization of advanced states
of mnemonic consciousness may be more distributed,
without a specific mapping to prefrontal regions, at least
in FTLD, although functional changes in frontal regions,
possibly reflecting alternations in frontal–posterior net-
works, cannot be ruled out as contributing to patients’
altered autobiographical memory function.
As noted above, the brain–behavior relationships were
most strongly evident at the specific probe phase, where
prefrontally mediated executive-retrieval mechanisms
might be less in demand (Matuszewski et al., 2006). This
may reflect the fact that, in the context of significant
temporal lobe damage limiting access to episodic auto-
biographical information, prefrontal function does not con-
tribute to autobiographical retrieval, even with retrieval
support (Rosenbaum, McKinnon, Levine, & Moscovitch,
2004; Kopelman et al., 2003). This also may explain why
patients with PNFA, with less medial temporal
lobe
damage, benefited from retrieval support, whereas those
with FTD/SD did not.
Conclusion
Autobiographical memory has recently received increas-
ing attention by cognitive neuroscientists interested in
human remote memory processes. Although it is clear
that autobiographical memory is multifactorial, with an
accordingly distributed functional neuroanatomy, the
delineation of specific brain–behavior relationships has
been elusive, likely owing to significant heterogeneity in
behavioral measures, imaging methods, and neuropsy-
chological populations.
In this study, FTLD, a form of presenile dementia
affecting key regions in the autobiographical memory
network, was used as a model for autobiographical
memory impairment. With the Autobiographical Inter-
view, we localized this impairment to episodic autobio-
graphical memory in both cross-sectional and longitudinal
analyses. Contrasting FTLD subtypes further specified pat-
terns of altered autobiographical memory. Despite their
mild speech abnormalities, PNFA patients were the least
impaired and were unique among subgroups in their ca-
pacity to benefit from retrieval support. FTD/SD patients
were the most impaired, failed to benefit from retrieval
support, and generated an excess of semantic autobio-
graphical details. FTD patients occupied a middle posi-
tion, impaired episodic, but not semantic autobiographical
memory, and a failure to benefit from retrieval support.
The identified pattern of impaired episodic but not se-
mantic autobiographical memory was related to volume
loss in a distributed network over the temporal lobes,
posterior subcortical regions, and left inferior parietal
and occipital regions. The capacity of patients with PNFA
to benefit from specific cueing may relate to these pa-
tients’ relative sparing of the medial temporal regions.
The lack of relation between frontal lobe volumes and
autobiographical memory performance suggests that
this brain–behavior correlation may be more complex
than previously recognized.
Acknowledgments
We thank the patients and their families for their assistance,
and Ann Campbell, Marina Mandic, Charlene O’Connor,
Colleen O’Toole, Joel Ramirez, Adriana Restagno, Gary Turner,
and Eva Svoboda for technical assistance. This research was
supported by grants from the Canadian Institutes of Health
Research (Grant nos. MT-14744, MOP-37535, and MOP-108540
to B. L., and MT-13129 to S. E. B.), and the NIH-NICHD (Grant
no. HD42385-01) to B. L.
Reprint requests should be sent to Brian Levine, Rotman Research
Institute, Baycrest Centre, 3560 Bathurst Street, Toronto, ON,
Canada M6A 2E1, or via e-mail: blevine@rotman-baycrest.on.ca.
McKinnon et al.
1851
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
f
/
t
.
.
o
n
1
8
M
a
y
2
0
2
1
REFERENCES
Addis, D. R., Moscovitch, M., & McAndrews, M. P. (2007).
Consequences of hippocampal damage across the
autobiographical memory network in left temporal lobe
epilepsy. Brain, 130, 2327–2342.
Aggleton, J. P., & Brown, M. W. (1999). Episodic memory, amnesia,
and the hippocampal–anterior thalamic axis. Behavioral and
Brain Sciences, 22, 425–444; discussion 444–489.
Arndt, S., Cohen, G., Alliger, R. J., Swayze, V. W., II, &
Andreasen, N. C. (1991). Problems with ratio and proportion
measures of imaged cerebral structures. Psychiatry
Research, 40, 79–89.
Army Individual Test Battery. (1944). Manual of directions
and scoring. Washington, D.C.: War Department, Adjutant
General’s Office.
Bocti, C., Rockel, C., Roy, P., Gao, F., & Black, S. E. (2006).
Topographical patterns of lobar atrophy in frontotemporal
dementia and Alzheimer’s disease. Dementia and Geriatric
Cognitive Disorders, 21, 364–372.
Bozeat, S., Gregory, C. A., Ralph, M. A., & Hodges, J. R. (2000).
Which neuropsychiatric and behavioural features distinguish
frontal and temporal variants of frontotemporal dementia
from Alzheimer’s disease? Journal of Neurology,
Neurosurgery and Psychiatry, 69, 178–186.
Bright, P., Buckman, J., Fradera, A., Yoshimasu, H., Colchester,
A. C., & Kopelman, M. D. (2006). Retrograde amnesia in
patients with hippocampal, medial temporal, temporal lobe,
or frontal pathology. Learning and Memory, 13, 545–557.
Burgess, P. W., Alderman, N., Evans, J. J., Wilson, B. A., &
Emslie, H. (1996). The Dysexecutive Questionnaire. In B. A.
Wilson, N. Alderman, P. W. Burgess, H. Emslie, & J. J. Evans
(Eds.), Behavioral assessment of the dysexecutive
syndrome. Bury St. Edmunds: Thames Valley Test Company.
Cermak, L. S. (1985). The episodic–semantic distinction in
amnesia. In L. R. Squire & N. Butters (Eds.), Neuropsychology
of memory (pp. 55–62). New York: Guilford Press.
Collins, D., & Evans, A. (1997). ANIMAL: Validation and
applications of non-linear registration based segmentation.
International Journal of Pattern Recognition, 11, 1271–1294.
Conway, M. A., & Pleydell-Pearce, C. W. (2000). The
construction of autobiographical memories in the self-
memory system. Psychological Review, 107, 261–288.
Craik, F. I. M., & McDowd, J. M. (1987). Age differences in
recall and recognition. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 13, 474–479.
Dade, L. A., Gao, F. Q., Kovacevic, N., Roy, P., Rockel, C., O’Toole,
C. M., et al. (2004). Semiautomatic brain region extraction: A
method of parcellating brain regions from structural magnetic
resonance images. Neuroimage, 22, 1492–1502.
Delis, D. C., Kaplan, E., & Kramer, J. H. (2001). Delis-Kaplan
Executive Functioning System (D-KEFS). San Antonio: The
Psychological Corporation.
Edgington, E. S. (1980). Randomization tests. New York:
Marcel Dekker.
Efron, B., & Tibshirani, R. (1986). Bootstrap methods for
standard errors, confidence intervals and other measures
of statistical accuracy. Statistical Sciences, 1, 54–77.
Eichenbaum, H. (2000). A cortical–hippocampal system for
declarative memory. Nature Reviews Neuroscience, 1, 41–50.
Eslinger, P. J. (1998). Autobiographical memory after temporal
lobe lesions. Neurocase, 4, 481–495.
Eustache, F., Piolino, P., Giffard, B., Viader, F., De La Sayette,
V., Baron, J. C., et al. (2004). ‘‘In the course of time’’: A PET
study of the cerebral substrates of autobiographical amnesia
in Alzheimer’s disease. Brain, 127, 1549–1560.
Feinstein, A., Roy, P., Lobaugh, N., Feinstein, K., O’Connor, P.,
& Black, S. (2004). Structural brain abnormalities in multiple
sclerosis patients with major depression. Neurology, 62,
586–590.
Fletcher, P. C., & Henson, R. N. (2001). Frontal lobes and
human memory: Insights from functional neuroimaging.
Brain, 124, 849–881.
Gilboa, A., Ramirez, J., Kohler, S., Westmacott, R., Black, S. E., &
Moscovitch, M. (2005). Retrieval of autobiographical memory
in Alzheimer’s disease: Relation to volumes of medial temporal
lobe and other structures. Hippocampus, 15, 535–550.
Hodges, J. R., & Gurd, J. M. (1994). Remote memory and lexical
retrieval in a case of frontal Pick’s disease. Archives of
Neurology, 51, 821–827.
Ivnik, R. J., Malec, J. F., Smith, G. E., Tangalos, E. G., &
Peterson, R. C. (1996). Neuropsychological tests’ norms
above age 55: COWAT, BNT, MAE token, WRAT-R Reading,
AMNART, STROOP, TMT and JLO. The Clinical
Neuropsychologist, 10, 262–278.
Kapur, N. (1999). Syndromes of retrograde amnesia: A
conceptual and empirical synthesis. Psychological Bulletin,
125, 800–825.
Kertesz, A., Davidson, W., & Fox, H. (1997). Frontal behavioral
inventory: Diagnostic criteria for frontal lobe dementia.
Canadian Journal of Neurological Sciences, 24, 29–36.
Kertesz, A., McMonagle, P., Blair, M., Davidson, W., & Munoz,
D. G. (2005). The evolution and pathology of
frontotemporal dementia. Brain, 128, 1996–2005.
Kirwan, C. B., Bayley, P. J., Galvan, V. V., & Squire, L. R. (2008).
Detailed recollection of remote autobiographical memory
after damage to the medial temporal lobe. Proceedings of
the National Academy of Sciences, U.S.A., 105, 2676–2680.
Kopelman, M. D., Lasserson, D., Kingsley, D. R., Bello, F., Rush, C.,
Stanhope, N., et al. (2003). Retrograde amnesia and the volume
of critical brain structures. Hippocampus, 13, 879–891.
Kopelman, M. D., Wilson, B. A., & Baddeley, A. D. (1990). The
autobiographical memory interview. Bury St. Edmunds:
Thames Valley Test Company.
Kovacevic, N., Henderson, J. T., Chan, E., Lifshitz, N., Bishop, J.,
Evans, A. C., et al. (2005). A three-dimensional MRI atlas of
the mouse brain with estimates of the average and
variability. Cerebral Cortex, 15, 639–645.
Kovacevic, N., Lobaugh, N. J., Bronskill, M. J., Levine, B.,
Feinstein, A., & Black, S. E. (2002). A robust method for
extraction and automatic segmentation of brain images.
Neuroimage, 17, 1087–1100.
Levine, B. (2004). Autobiographical memory and the self in
time: Brain lesion effects, functional neuroanatomy, and
lifespan development. Brain and Cognition, 55, 54–68.
Levine, B., Kovacevic, N., Nica, I., Cheung, G., Schwartz, M. L., &
Black, S. E. (2008). The Toronto traumatic brain injury study:
Injury severity and quantified MRI. Neurology, 70, 771–778.
Levine, B., Svoboda, E., Hay, J., Winocur, G., & Moscovitch, M. (2002).
Aging and autobiographical memory: Dissociating episodic from
semantic retrieval. Psychology and Aging, 17, 677–689.
Matuszewski, V., Piolino, P., de la Sayette, V., Lalevee, C., Pelerin,
A., Dupuy, B., et al. (2006). Retrieval mechanisms for
autobiographical memories: Insights from the frontal variant of
frontotemporal dementia. Neuropsychologia, 44, 2386–2397.
McIntosh, A. R., Bookstein, F. L., Haxby, J. V., & Grady, C. L.
(1996). Spatial pattern analysis of functional brain images
using partial least squares. Neuroimage, 3, 143–157.
McIntosh, A. R., Chau, W. K., & Protzner, A. B. (2004).
Spatiotemporal analysis of event-related fMRI data using
partial least squares. Neuroimage, 23, 764–775.
McKhann, G. M., Albert, M. S., Grossman, M., Miller, B.,
Dickson, D., & Trojanowski, J. Q. (2001). Clinical and
pathological diagnosis of frontotemporal dementia: Report
of the Work Group on Frontotemporal Dementia and Pick’s
Disease. Archives of Neurology, 58, 1803–1809.
1852
Journal of Cognitive Neuroscience
Volume 20, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
f
/
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
McKinnon, M. C., Black, S. E., Miller, B., Moscovitch, M., & Levine,
B. (2006). Autobiographical memory in semantic dementia:
Implication for theories of limbic–neocortical interaction in
remote memory. Neuropsychologia, 44, 2421–2429.
McKinnon, M. C., Svoboda, E., & Levine, B. (2007). The frontal
lobes and autobiographical memory. In B. L. Miller & J. L.
Cummings (Eds.), The human frontal lobes, functions and
disorders (pp. 227–248). New York: Guilford Press.
Miller, B. L., Seeley, W. W., Mychack, P., Rosen, H. J., Mena, I., &
Boone, K. (2001). Neuroanatomy of the self: Evidence from
patients with frontotemporal dementia. Neurology, 57,
817–821.
Milner, B. (1963). Effects of different brain lesions on card
sorting: The role of the frontal lobes. Archives of Neurology,
9, 100–110.
Morris, R., Pandya, D. N., & Petrides, M. (1999). Fiber system
linking the mid-dorsolateral frontal cortex with the
retrosplenial/presubicular region in the rhesus monkey.
Journal of Comparative Neurology, 407, 183–192.
Morris, R., Petrides, M., & Pandya, D. N. (1999). Architecture
and connections of retrosplenial area 30 in the rhesus
monkey (Macaca mulatta). European Journal of
Neuroscience, 11, 2506–2518.
Moscovitch, M., Rosenbaum, R. S., Gilboa, A., Addis, D. R.,
Westmacott, R., Grady, C., et al. (2005). Functional
neuroanatomy of remote episodic, semantic and spatial
memory: A unified account based on multiple trace theory.
Journal of Anatomy, 207, 35–66.
Moss, H. E., Kopelman, M., Cappelletti, M., De Mornay Davies,
P., & Jaldow, E. (2003). Lost for words or loss of memories?
Autobiographical memory in semantic dementia. Cognitive
Neuropsychology, 20, 703–732.
Rubin, D. C., & Greenberg, D. L. (1998). Visual memory-deficit
amnesia: A distinct amnesic presentation and etiology.
Proceedings of the National Academy of Sciences, U.S.A., 95,
5413–5416.
Rubin, D. C., & Schulkind, M. D. (1997). The distribution of
autobiographical memories across the lifespan. Memory &
Cognition, 25, 859–866.
Simons, J. S., Graham, K. S., & Hodges, J. R. (2002). Perceptual
and semantic contributions to episodic memory: Evidence
from semantic dementia and Alzheimer’s disease. Journal of
Memory and Language, 47, 197–213.
Simons, J. S., & Spiers, H. J. (2003). Prefrontal and medial
temporal lobe interactions in long-term memory. Nature
Reviews Neuroscience, 4, 637–648.
Simons, J. S., Verfaellie, M., Galton, C. J., Miller, B. L., Hodges,
J. R., & Graham, K. S. (2002). Recollection-based memory
in frontotemporal dementia: Implications for theories of
long-term memory. Brain, 125, 2523–2536.
Snowden, J., Griffiths, H., & Neary, D. (1994). Semantic
dementia: Autobiographical contribution to preservation of
meaning. Cognitive Neuropsychology, 11, 265–288.
Snowden, J., Neary, D., & Mann, D. (2007). Frontotemporal
lobar degeneration: Clinical and pathological relationships.
Acta Neuropathologica (Berlin), 114, 31–38.
So¨derlund, H., Black, S. E., Miller, B. L., Freedman, M., &
Levine, B. (2008). Episodic memory and regional atrophy
in frontotemporal lobar degeneration. Neuropsychologia,
46, 127–136.
Spiers, H. J., Maguire, E. A., & Burgess, N. (2001). Hippocampal
amnesia. Neurocase, 7, 357–382.
Squire, L. R., & Bayley, P. J. (2007). The neuroscience of remote
memory. Current Opinion in Neurobiology, 17, 185–196.
Neary, D., Snowden, J. S., Gustafson, L., Passant, U., Stuss,
St.-Jacques, P., & Levine, B. (2007). Aging and emotional
D. T., Black, S., et al. (1998). Frontotemporal lobar
degeneration: A consensus on clinical diagnostic criteria.
Neurology, 51, 1546–1554.
Nestor, P. J., Graham, K. S., Bozeat, S., Simons, J. S., & Hodges,
J. R. (2002). Memory consolidation and the hippocampus:
Further evidence from studies of autobiographical memory
in semantic dementia and frontal variant frontotemporal
dementia. Neuropsychologia, 40, 633–654.
Piolino, P., Chetelat, G., Matuszewski, V., Landeau, B., Mezenge,
F., Viader, F., et al. (2007). In search of autobiographical
memories: A PET study in the frontal variant of
frontotemporal dementia. Neuropsychologia, 45, 2730–2743.
Piolino, P., Desgranges, B., Belliard, S., Matuszewski, V.,
Lalevee, C., De la Sayette, V., et al. (2003). Autobiographical
memory and autonoetic consciousness: Triple dissociation
in neurodegenerative diseases. Brain, 126, 2203–2219.
Ratnavalli, E., Brayne, C., Dawson, K., & Hodges, J. R. (2002).
The prevalence of frontotemporal dementia. Neurology, 58,
1615–1621.
Rosen, H. J., Allison, S. C., Ogar, J. M., Amici, S., Rose, K.,
Dronkers, N., et al. (2006). Behavioral features in semantic
dementia vs other forms of progressive aphasias. Neurology,
67, 1752–1756.
Rosenbaum, R. S., McKinnon, M. C., Levine, B., & Moscovitch,
M. (2004). Visual imagery deficits, impaired strategic
retrieval, or memory loss: Disentangling the nature of an
amnesic person’s autobiographical memory deficit.
Neuropsychologia, 42, 1619–1635.
Rosenbaum, R. S., Moscovitch, M., Foster, J. K., Verfaellie, M., Gao,
F. Q., Black, S. E., et al. (2008). Patterns of autobiographical
memory loss in medial temporal lobe amnesic patients.
Journal of Cognitive Neuroscience, 20, 1490–1506.
Rosene, D. L., & Van Hoesen, G. W. (1977). Hippocampal
efferents reach widespread areas of cerebral cortex and
amygdala in the rhesus monkey. Science, 198, 315–317.
autobiographical memory. Memory, 15, 129–144.
Steinvorth, S., Levine, B., & Corkin, S. (2005). Medial temporal
lobe structures are needed to re-experience remote
autobiographical memories: Evidence from H.M. and W.R.
Neuropsychologia, 43, 479–496.
Stuss, D. T., & Benson, D. F. (1986). The frontal lobes. New
York: Raven Press.
Svoboda, E., McKinnon, M. C., & Levine, B. (2006). The
functional neuroanatomy of autobiographical memory: A
meta-analysis. Neuropsychologia, 44, 2189–2208.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
atlas of the human brain. Stuttgart: Georg Thieme Verlag.
Tulving, E. (2002). Episodic memory: From mind to brain.
Annual Review of Psychology, 53, 1–25.
Valenstein, E., Bowers, D., Verfaellie, M., Heilman, K. M., Day,
A., & Watson, R. T. (1987). Retrosplenial amnesia. Brain,
110, 1631–1646.
Vogt, B. A., Pandya, D. N., & Rosene, D. L. (1987). Cingulate
cortex of the rhesus monkey: I. Cytoarchitecture and
thalamic afferents. Journal of Comparative Neurology, 262,
256–270.
Wasserman, S., & Bockenholt, U. (1989). Bootstrapping:
Applications to psychophysiology. Psychophysiology, 26,
208–221.
Westmacott, R., & Moscovitch, M. (2003). The contribution of
autobiographical significance to semantic memory. Memory
& Cognition, 31, 761–774.
Woods, R. P., Grafton, S. T., Holmes, C. J., Cherry, S. R., &
Mazziotta, J. C. (1998). Automated image registration: I.
General methods and intrasubject, intramodality validation.
Journal of Computer Assisted Tomography, 22, 139–152.
Woods, R. P., Grafton, S. T., Watson, J. D., Sicotte, N. L., &
Mazziotta, J. C. (1998). Automated image registration: II.
Intersubject validation of linear and nonlinear models.
Journal of Computer Assisted Tomography, 22, 153–165.
McKinnon et al.
1853
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
e
1
0
-
p
1
d
8
f
3
/
9
2
1
0
9
/
3
1
6
0
6
/
1
1
0
8
o
3
c
9
n
/
2
1
0
7
0
6
8
4
4
2
8
0
1
2
2
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
2
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
.
t
/
.
f
o
n
1
8
M
a
y
2
0
2
1