A Surface-based Analysis of Language Lateralization
and Cortical Asymmetry
Douglas N. Greve1, Lise Van der Haegen2, Qing Cai2,3,4,
Steven Stufflebeam1, Mert R. Sabuncu1,5,
Bruce Fischl1,5, and Marc Brysbaert2
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
Abstrakt
■ Among brain functions, language is one of the most lateral-
isiert. Cortical language areas are also some of the most asymmet-
rical in the brain. An open question is whether the asymmetry in
function is linked to the asymmetry in anatomy. Um das zu erwähnen
question, we measured anatomical asymmetry in 34 Teilnehmer
shown with fMRI to have language dominance of the left hemi-
Kugel (LLD) Und 21 participants shown to have atypical right
hemisphere dominance (RLD). All participants were healthy
and left-handed, and most (80%) were female. Gray matter
(GM) volume asymmetry was measured using an automated
surface-based technique in both ROIs and exploratory analyses.
In the ROI analysis, a significant difference between LLD and
RLD was found in the insula. No differences were found in planum
temporale (PT), pars opercularis (POp), pars triangularis (PTr),
or Heschlʼs gyrus (HG). The PT, POp, insula, and HG were all
significantly left lateralized in both LLD and RLD participants.
Both the positive and negative ROI findings replicate a pre-
vious study using manually labeled ROIs in a different cohort
[Keller, S. S., Roberts, N., Garcia-Finana, M., Mohammadi, S.,
Ringelstein, E. B., Knecht, S., et al. Can the language-dominant
hemisphere be predicted by brain anatomy? Zeitschrift für Kognition
Neurowissenschaften, 23, 2013–2029, 2011]. The exploratory analysis
was accomplished using a new surface-based registration that
aligns cortical folding patterns across both subject and hemi-
Kugel. A small but significant cluster was found in the superior
temporal gyrus that overlapped with the PT. A cluster was also
found in the ventral occipitotemporal cortex corresponding to
the visual word recognition area. The surface-based analysis also
makes it possible to disentangle the effects of GM volume, dick-
ness, and surface area while removing the effects of curvature. Für
both the ROI and exploratory analyses, the difference between
LLD and RLD volume laterality was most strongly driven by differ-
ences in surface area and not cortical thickness. Gesamt, Dort
were surprisingly few differences in GM volume asymmetry
between LLD and RLD indicating that gross morphometric asym-
metry is only subtly related to functional language laterality. ■
EINFÜHRUNG
It has long been known that speech production is later-
alized in the brain, with the left side being dominant in
most people. On the basis of a review of the existing clin-
ical data, Benson and Geschwind (1985) estimated that
60% of right-handed patients with unilateral left hemi-
Kugel (LH) lesions developed aphasia, whereas only
2% of right-handed patients with right hemisphere (RH)
damage did. For left-handed patients, the figures were,
jeweils, 32% Und 24%. Other clinical evidence came
from the Wada test (Wada & Rasmussen, 1960). In diesem
test, sodium amytal is injected unilaterally to anesthetize
a hemisphere, and the effects on speech production are
monitored. The test was administered to determine
speech dominance in epileptic patients before surgery.
Loring et al. (1990) described the outcome of one of
the best controlled studies. A total of 103 Patienten (91
right-handed and 12 links- or mixed-handed) were tested.
1Harvard Medical School, 2Ghent University, 3East China Nor-
mal University, 4INSERM, Cognitive Neuroimaging Unit, Frankreich,
5Massachusetts Institute of Technology
© 2013 Massachusetts Institute of Technology
Of these, 79 had exclusively LH language representation
(73 right-handers) involving both production (counting)
and comprehension (following simple commands). Nur
two patients had exclusive RH language representation
(1 right-hander). The remaining 22 Teilnehmer (17 Rechts-
handers) had performance decrements after injection to
each hemisphere.
In den 1990ern, brain imaging started to replace the Wada
test. Classifications with these techniques were shown to
be nearly identical to those of the Wada test (z.B., Binder
et al., 1996) and made possible research on healthy indi-
viduals. Using transcranial Doppler sonography, Knecht
and colleagues reported that 95% of right-handers had
a larger increase in blood flow in the LH when silently
generating words starting with a particular letter. Für
the left-handers, the percentage of left dominance was
75–90%, depending on the degree of handedness (Van der
Haegen, Cai, Seurinck, & Brysbaert, 2011; Knecht et al.,
2000; Loring et al., 1990).
Kürzlich, researchers have started to investigate what
consequences atypical right speech lateralization has for
the lateralization of other brain functions. Erste, es war
Zeitschrift für kognitive Neurowissenschaften 25:9, S. 1477–1492
doi:10.1162/jocn_a_00405
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
T
F
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
shown that right lateralization of Brocaʼs area is mostly,
but not always, accompanied by atypical lateralization of
the occipito-temporal regions involved in word reading
(Van der Haegen, Cai, & Brysbaert, 2012; Seghier & Price,
2011; Cai, Paulignan, Brysbaert, Ibarrola, & Nazir, 2010;
Cai, Lavidor, Brysbaert, Paulignan, & Nazir, 2008). Zweite,
it was observed that all participants with RH speech
dominance were right dominant for tool use as well
(Vingerhoets et al., 2013), independent of handed-
ness. Good performance on this task involved the SMA
(BA 6), Brocaʼs area (BA 44/45), the dorsolateral pFC
(BA 9/46), and the posterior parietal cortex (BA 40/7).
Endlich, Cai, Van der Haegen, and Brysbaert (2013) Re-
ported that all participants with right speech lateralization
had atypical, LH lateralization of the frontoparietal network
involved in visuospatial processing (the Landmark task).
This network involved the inferior parietal sulcus and the
superior parietal lobule (BA 39/40) together with the FEF
(the intersection of BA 4/6/8) and the inferior frontal sulcus
(BA 44/45) in the nondominant hemisphere. Daher, Es
appears that atypical speech lateralization has profound
implications for functional lateralization throughout the
brain.1
In this study, we examine how atypical speech domi-
nance is related to the structural asymmetries of the
cerebral cortex. Given the large functional implications,
one might expect that atypical speech lateralization is
accompanied by substantial differences in gray matter
(GM). A dominant hypothesis is that the usual LH language
dominance may be the result of the LH being larger in
brain areas related to language processing. In der Tat, beide
the planum temporale (PT), involved in auditory language
Verarbeitung, and the opercular part of Brocaʼs area are
known to show a leftward volume asymmetry, welches ist
larger in right-handers than in left-handers (Toga &
Thompson, 2003). This discovery has led to a number
of studies in which functional laterality scores were corre-
lated with anatomical laterality scores (z.B., Josse, Kherif,
Flandin, Seghier, & Price, 2009; Josse, Mazoyer, Crivello,
& Tzourio-Mazoyer, 2003; Tzourio, Nkanga-Ngila, &
Mazoyer, 1998). A limitation of these studies, Jedoch,
was that they usually involved small numbers of partici-
pants who were not selected for hemisphere dominance.
Given the rareness of atypical right speech laterality, es ist
unlikely that enough participants with truly inversed
speech dominance were included to draw valid conclu-
sionen (see Cai et al., 2013, for more information on the
problem of drawing conclusions on the basis of samples
mainly consisting of left language dominant participants).
Arguably the best study comparing left language domi-
nant (LLD) participants with right language dominant
(RLD) participants was Keller et al. (2011). The authors
compared the anatomical MRI images of 15 LLD partici-
pants with those of 10 RLD participants (language dom-
inance determined by transcranial Doppler sonography
or fMRI during a silent word generation [WG]). To their
surprise, Keller et al. observed no relationship between
volume asymmetry of Brocaʼs area or the PT and lan-
guage dominance (the trends were even in the opposite
Richtung, with larger leftward asymmetries for RLD par-
ticipants than for LLD participants). The only region for
which they found a robust relationship between volume
asymmetry and language dominance was the insula, mit
a tendency toward rightward asymmetry for the RLD par-
ticipants, making the authors conclude that the insular
morphology should be given more importance in studies
on the anatomical correlates of human language laterali-
zation. This study performs an analysis similar to Keller
et al. (2011) to see whether the surprising findings can
be replicated on new groups of LLD and RLD participants
and to what extent the results depend on the methodol-
ogy used.
An important issue in studying the relationship be-
tween structure and function is the delineation of mor-
phological features. Typically, cortical structures known
to be involved in language are manually segmented
based on landmarks to obtain measures of volume, Bereich,
Länge, and/or angulation. Jedoch, the cortical surface
is highly folded with variable thickness, and there is a
large degree of anatomical variability across participants.
This variability makes it difficult to consistently and accu-
rately define and quantify structures of interest. Manual
labeling is also a tedious procedure that is not feasible
in the large data sets required for testing hypotheses in
human studies.
Even in the case where morphological features are well
defined and quantified, they may still be somewhat arbi-
trary relative to the underlying function. The anatomical
manifestation of a functional asymmetry may not confine
itself neatly within a morphological feature chosen by an
anatomist. This can cause a loss of statistical power if the
entire feature is considered. In this case, exploratory
methods may be more powerful because they allow the
data itself to determine what the underlying morphologi-
cal feature should be. Voxel-based morphometry ( VBM;
Aschenbrenner & Friston, 2000) is often used in this type of
application, including brain asymmetry (Luders, Gaser,
Jancke, & Schlaug, 2004; Watkins et al., 2001). Dort
has only been one study using VBM to assess structural
asymmetry differences in LLD and RLD (Dorsaint-Pierre
et al., 2006). This VBM study was in an epilepsy pre-
surgical cohort involving a small number of patients (11 von
whom were RLD), which may not reflect the general
Bevölkerung. Zusätzlich, VBM has two other short-
kommt. Erste, it relies on volume-based registration to
align brains across participants, which is often inaccurate
when attempting to align particular cortical folds (Fischl
et al., 2008). Zweite, when differences are found with
VBM, the results cannot be refined to determine
whether the underlying cause of the difference was be-
cause of differences in folding pattern, GM volume, cor-
tical surface, or cortical thickness.
This study addresses the limitations of Keller et al.
(2011) in three important ways. Erste, it uses a relatively
1478
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
large sample of healthy participants who have been ver-
ified to be strongly RLD based on fMRI (n = 21 instead of
10), thereby considerably increasing the power of the
Studie. This increase was made practical through the
use of a low-cost and noninvasive screening procedure
developed previously (Van der Haegen et al., 2011).
Zweite, it uses computer-generated mesh models of
the cortical surface to automatically find and accurately
quantify standard morphological features, einschließlich
those associated with language. Endlich, it uses surface-
based interparticipant and interhemispheric registration
to align folding patterns so that the cortical volume,
surface area, and thickness can be consistently evaluated
across language dominance group, Thema, and hemi-
Kugel. This registration allows for an exploratory analy-
sis in which the data dictate the anatomical boundaries
of areas that show consistent structural and functional
asymmetries.
METHODEN
Laterality Index
The laterality index (LI) is used to quantify the difference
between left and right while removing the effects of brain
Größe. We use the formula:
LI ¼
ðL − RÞ
ðL þ RÞ
ð1Þ
where L is the value from the left side and R is the value
from the right side. LI has a range from −1 (vollständig
right lateralized) Zu +1 (completely left lateralized). Das
formula can be applied to any metric including volume,
Bereich, Dicke, volume of functional activation, or behav-
ioral measure.
Participants and fMRI Analysis
Full details of subject recruitment, handedness assess-
ment, screening procedures, fMRI analysis, and language
LI calculation are provided in Van der Haegen et al.
(2011, 2012). For completeness, we summarize them
Hier. All participants signed an informed consent form
according to the guidelines of the Ethics Committee of
the Ghent University Hospital. A total of 269 partici-
pants2 were accepted to the initial screening based
on the criteria that they wrote and drew with their
left hand to increase the likelihood of atypical language
dominance. Handedness was later assessed with the
Edinburgh Handedness Inventory (Oldfield, 1971) modi-
fied to have answers in the range of −3 to −1 (degree of
left-handedness) oder +1 Zu +3 (right-handedness). Most
participants underwent two visual half field ( VHF) tasks
(Hunter & Brysbaert, 2008) in which they were asked to
name words and pictures presented to the left visual field
(LVF) or to the right visual field (RVF). LIs were calculated
by subtracting the mean RT to stimuli in RVF from the
mean RT to stimuli in LVF.
Sixty-five participants were invited (and willing) to take
part in the fMRI study. Twenty-five were expected to be
LLD on the basis of their VHF scores; the remaining 40
had an LVF advantage on one of the VHF tasks and were
hoped to be RLD (Van der Haegen et al., 2011; Hunter &
Brysbaert, 2008).3 The fMRI task consisted of silent WG
(Hunter & Brysbaert, 2008; Knecht et al., 2000; Pujol,
Deus, Losilla, & Capdevila, 1999). Participants were asked
to silently think of as many words as possible, beginning
with a cued letter. The control/baseline condition was
silent repetition of the nonword “baba.” SPMs were gen-
erated based on target letter versus nonword contrast.
The functional LIs were computed in areas approximately
corresponding to Brocaʼs area (d.h., BA 44 and BA 45; AAL
Vorlage; Tzourio-Mazoyer et al., 2002). These regions
were chosen because they are the most active areas in
the silent WG task and are known to be involved in many
linguistic functions (Heim, Eickhoff, & Amunts, 2008;
Amunts et al., 2004).
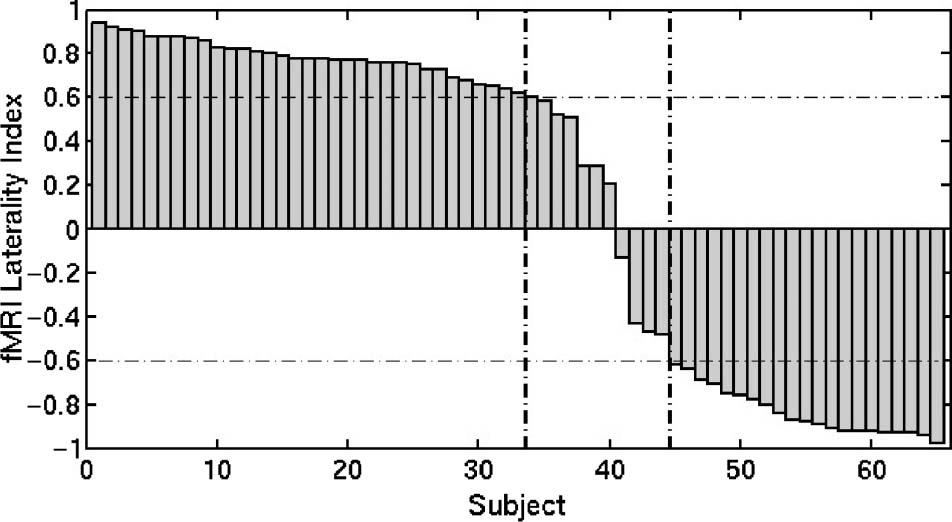
For statistical analysis, Die 65 participants were cate-
gorized into three groups based on the functional WG
LI scores: LLD if LI > 0.6, RLD if LI < −0.6, and bilateral
language dominant (BLD) otherwise. This categorization
was used to make a clear separation between the RLD
group and the LLD group (see the Discussion for the rea-
soning behind this model). Figure 1 shows the distribu-
tion of the fMRI LIs of all participants. The demographics
and mean LI for the three groups are shown in Table 1.
The handedness scores for all groups were less than −2,
indicating strong left handedness (−3 would be the most
extreme for left handers). The groups did not signifi-
cantly differ in handedness ( p > .55) or age ( p > .48).
The sample was recruited from a wide range of courses at
university or higher education schools. As female stu-
dents seemed to be more willing to take part, Sie
formed the majority of participants.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
/
.
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Figur 1. Distribution of the functional LI from the WG task for
all participants. The horizontal dashed lines indicate the threshold of
±0.6. The vertical dashed lines indicate the categorical boundaries
of LLD, BLD, and RLD.
Greve et al.
1479
Tisch 1. Participant Demographics
Group
Total
Male
Female
Alter (SD)
Handedness (SD)
fMRI LI (SD)
Template
LLD
RLD
BLD
34
21
10
8
3
4
26
18
6
20.4 (2.6)
20.9 (2.8)
21.0 (1.7)
−2.12 (1.06)
−2.35 (0.78)
−2.01 (1.36)
+0.78 (.09)
−0.84 (.11)
+0.14 (.43)
21
21
0
A second fMRI task (lexical decision task, LDT) War
also collected on these participants (Van der Haegen
et al., 2012). The LDT assesses the lateralization of word
reading by looking at activity in the vOT. Stimuli con-
sisted of high- and low-frequency words, consonant
strings, and pixel-scrambled words. Participants were re-
quired to respond with button press as to whether the
stimulus was a word or nonword. Cerebral dominance
was determined on the basis of the WG task and not
the LDT, in line with clinical practice (Spreer et al.,
2002; Benson et al., 1999; Springer et al., 1999), welche
uses speech production lateralization as an index of lan-
guage laterality. The brain areas involved in speech pro-
duction are the most lateralized, arguably because fluent
speaking requires a single control center (Kosslyn, 1987).
MRI Acquisition
Images were acquired on a 3-T Siemens Trio MRI scanner
at Ghent University (Siemens Medical Systems, Erlangen,
Deutschland) with an eight-channel radiofrequency head
coil. A high-resolution anatomical image was collected
using a T1-weighted 3-D MPRAGE sequence. All images
war 256 × 256 × 176 with voxel size = 0.9 × 0.9 ×
0.9 mm3. The data were acquired with either one of
two sets of pulse sequence parameters: repetition time =
1550 ms, Echozeit = 2.39 ms, flip angle = 9°, inver-
sion time = 900 ms, pixel frequency = 180 Hz; oder
repetition time = 2530 ms, Echozeit = 2.58 ms,
flip angle = 7°, inversion time = 1100 ms, pixel fre-
quency = 190 Hz. Most (n = 54) were acquired with
the first set. The proportion of each parameter set was
the same (85% Zu 15%) in the LLD and RLD groups.
Anatomical Analysis
All participants were analyzed in FreeSurfer (www.surfer.
nmr.mgh.harvard.edu, Ausführung 5.1) to provide detailed
anatomical information customized for each participant
(Dale, Fischl, & Sereno, 1999; Fischl, Sereno, & Dale,
1999). The FreeSurfer analysis stream includes intensity
bias field removal, skull stripping, and assigning a neuro-
anatomical label (z.B., hippocampus, amygdala, usw.) Zu
each voxel (Segonne et al., 2004; Fischl et al., 2002). In
addition to the volume-based analysis, FreeSurfer con-
structs models of the cortical surface. A surface model
consists of a mesh of triangles. The location of the mesh
is controlled by adjusting the location of the vertices. A
vertex is the place where the points of neighboring tri-
angles meet (typically about 1 mm apart). The vertex
positions are adjusted such that the surface follows the
T1 intensity gradient between cortical white matter
(WM) and cortical GM. Smoothness constraints allow the
surface to cut through a voxel to model partial volume
effects and provide subvoxel accuracy of the location of
the surface. This highly folded surface can be “inflated”
(Figur 2) to see inside the sulci. A second surface is also
fit to the outside of the brain (between the GM and the
pia). The first surface is called the “white” surface, und das
second is called the “pial” surface (Figures 3A and 4A show
pial surfaces). The LH and RH are modeled separately. Alle
surfaces are constructed in the native anatomical space.
Cortical GM Metrics
In FreeSurfer, the distance between the white and pial
surfaces at a vertex is defined to be the cortical thickness
at that vertex (Fischl & Dale, 2000). The area of a vertex is
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
/
F
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
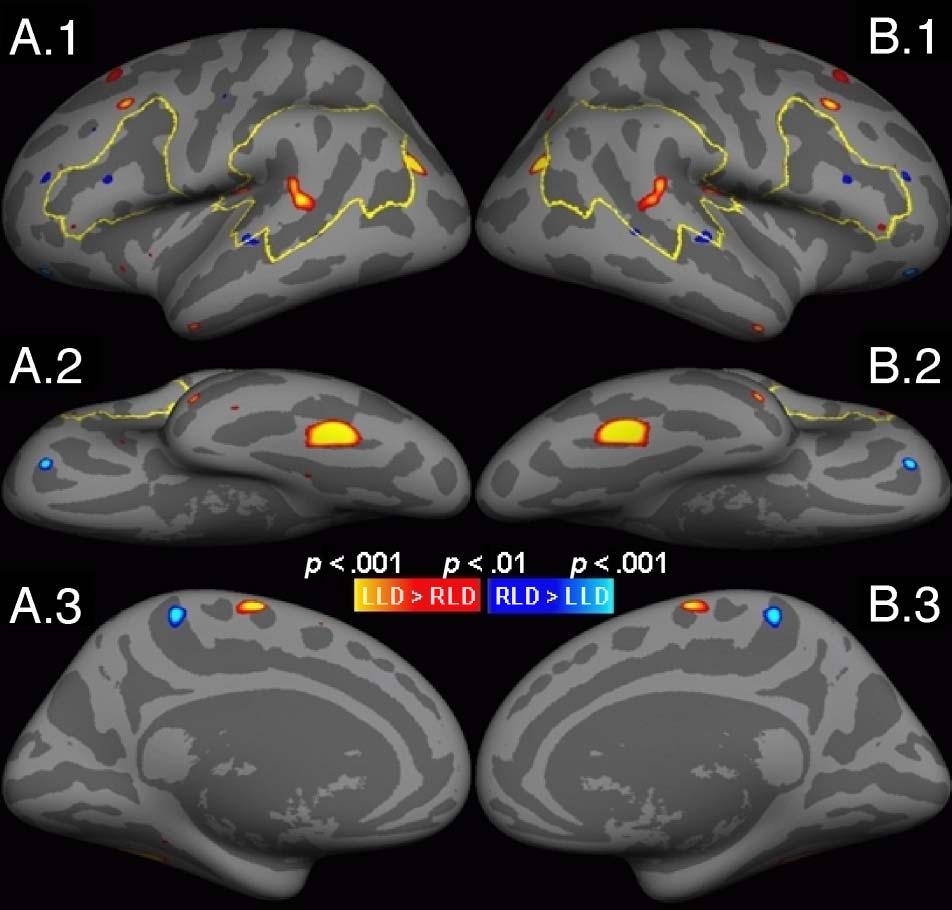
Figur 2. Significance map of volume LI difference between LLD
and RLD rendered on the inflated cortical surface of the symmetric
Vorlage (uncorrected vertex-wise threshold p < .01). Red/yellow
indicates that LLD > RLD; blue/cyan indicates that LLD < RLD.
The yellow outlines indicate the boundaries of the a priori ROIs
(ALA and PLA). (A) Results from the template initialized with the LH.
(B) Results from the template initialized with the RH. From top to
bottom: (1) lateral view, (2) inferior view, (3) medial view.
1480
Journal of Cognitive Neuroscience
Volume 25, Number 9
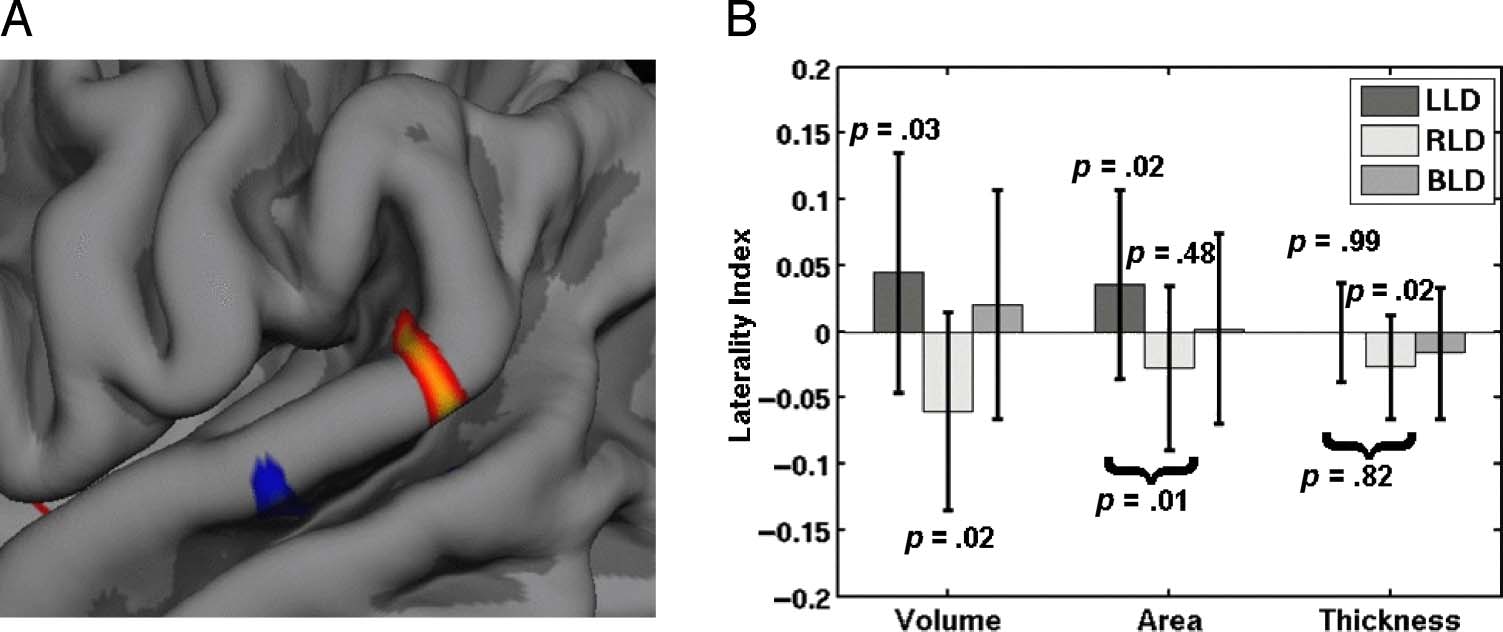
Figure 3. (A) View of the
left posterior sylvian fissure
including the STG cluster on
the folded symmetric template
pial surface (same data as in
Figure 2A1). (B) LI results
broken down by subject group
and volume, area, and thickness
(with standard deviation bars
across subject) based upon
averages from within the
exploratory STG cluster. The
p values are for post hoc tests
of laterality or differences in
laterality between LLD and
RLD. The p values have been
corrected for the eight
post hoc tests. All p values
are based on a two-sided t test
with 53 DOF.
defined as the average area of the triangles of which the
vertex is a member. The GM volume of a vertex is defined
as the area times the thickness. The surface area of a
region can be computed by adding up the area of the ver-
tices in that region, and the same can be done to com-
pute the GM volume. The surface curvature at a vertex
can be computed based on its spatial relationship to
neighboring vertices. The curvature is a quantification
of the folding patterns. Thus, at each point along the sub-
jectʼs surface, the thickness, area, volume, and curvature
can be quantified, all with subvoxel accuracy.
rate for cortical areas (Fischl et al., 2008). The thickness, area,
and volume can be resampled into the standard space so that
vertex-wise comparisons across subjects can be made in an
exploratory analysis. The area and volume resampling in-
cludes a “jacobian correction“ (similar to that described in
Winkler et al., 2012) to account for any stretching or compres-
sion in the registration. This correction is not needed for
thickness because it is measured along a vector normal to
any stretching or compression.
Cross-hemisphere Symmetric
Registration Template
Surface-based Intersubject Registration
The curvature is used to drive a nonlinear surface-based inter-
subject registration procedure that aligns the cortical folding
patterns of each subject to a standard surface space (Fischl,
Sereno, Tootell, & Dale, 1999). This approach is similar to
performing a volume-based registration to Talairach or
Montreal Neurological Institute space but is much more accu-
There are two standard space surface registration templates
in FreeSurfer, one for each hemisphere. Simply registering
a hemisphere to a contralateral template can create biases,
that is, one will get different results depending upon
whether one registers both hemispheres to the LH template
or to the RH template. A symmetric template is needed,
which can be generated in one of two ways: (1) regenerate
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
o
9
c
4
n
2
_
5
a
/
_
j
0
o
0
c
4
n
0
5
_
a
p
_
d
0
0
b
4
y
0
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
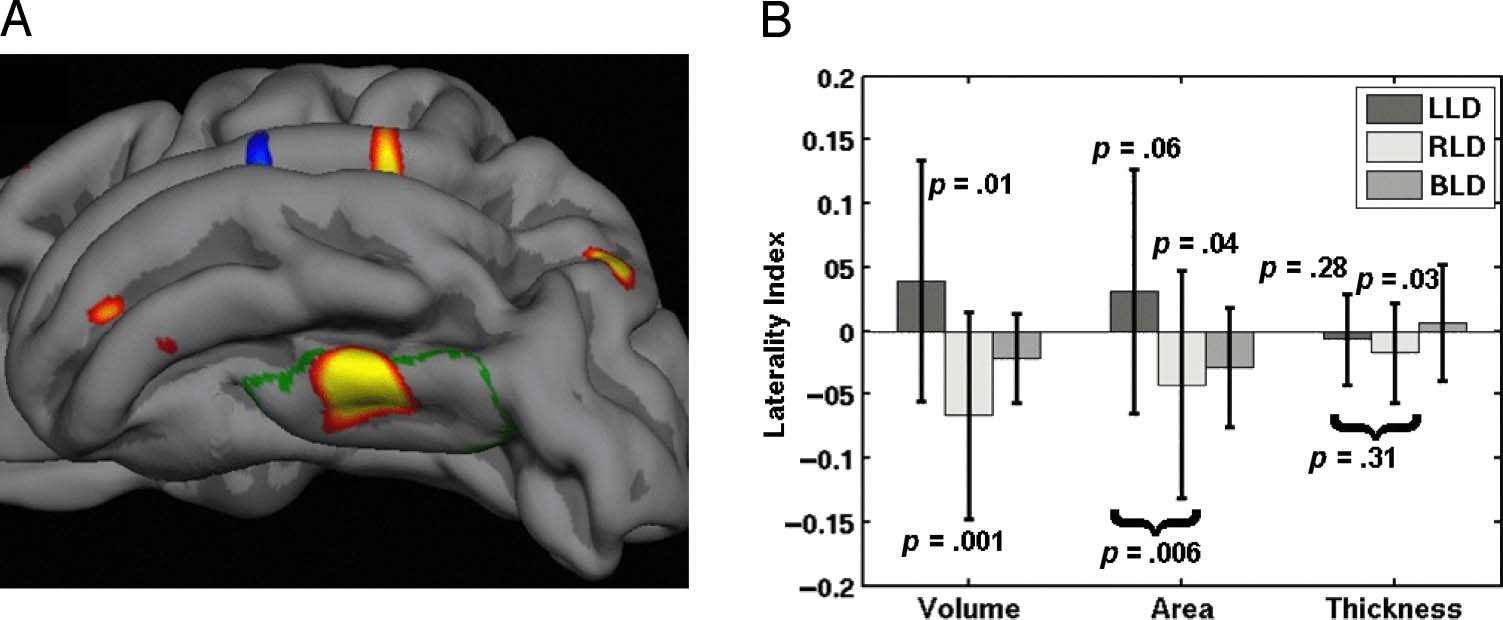
Figure 4. (A) vOT cluster
shown on the pial surface of
the LH (view is inferior and
slightly lateral). The green
outline is the lateral occipito-
temporal gyrus from the
Destrieux atlas. This is the
same data as in Figure 2A2.
(B) LI results broken down
by subject group and volume,
area, and thickness (with
standard deviation bars across
subject) based upon averages
from within the vOT cluster.
The p values are for post hoc
tests of laterality or differences
in laterality between LLD and
RLD. The p values have been
corrected for the eight post hoc
tests. All p values are based on
a two-sided t test with 53 DOF.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Greve et al.
1481
the template from scratch by left–right reversing the sub-
jects used in the template and reregistering with all data
(i.e., treating the reversed subjects as new, additional sub-
jects) or (2) left–right reversing the template itself and aver-
aging with the non reversed atlas. This second approach is
easy to implement and dominants in the field ( Josse et al.,
2009; Eckert et al., 2008; Watkins et al., 2001). However, this
approach has several drawbacks, namely, a blurrier atlas is
created because features are washed out, and there is no
way to keep track of the amount of variance because of
variability across hemispheres. For these reasons, we cre-
ated a new surface template from scratch using an iterative
technique described in Fischl, Sereno, Tootell, et al. (1999)
modified to yield a symmetric template. The symmetric
template was created using 42 participants from this study
consisting of an equal number of LLD and RLD, each group
represented by 3 men and 18 women. No BLD were used in
the template construction. First, an initial template was cre-
ated from only the LHs of these participants based on the
registration to the LH standard template. Both LHs and
RHs from all participants were aligned with this initial LH
template. A new template was then created from these 84
surfaces, and the surfaces were reregistered to it. This
new template is a mixture of LH and RH and so less biased
than the initial LH-only template. This process was repeated
35 times to remove the influence of initializing with LH. To
test whether this procedure leads to a symmetrical tem-
plate, the entire template creation procedure was replicated
initialing with the RH. The exploratory analysis described
below was performed using both the LH- and RH-initialized
templates. Similarity in the results is evidence of a sym-
metric template.
Anatomically Defined Cortical ROIs
FreeSurfer has the ability to automatically label the cortex
in a way intended to replicate the labeling of a trained
anatomist (Destrieux, Fischl, Dale, & Halgren, 2010;
Desikan et al., 2006; Fischl et al., 2004). This labeling
goes beyond simply the mapping of an ROI atlas into
the subject space because the boundaries of the labels
are customized to each participant based on curvature
statistics stored in the ROI atlas. Note that, whereas an
anatomist uses explicit rules and landmarks to define
specific areas, the automatic procedure relies only on
implicit rules generated during the training process. This
method has been validated using a jackknifing proce-
dure (Fischl et al., 2004). We used the Destrieux atlas
(Destrieux et al., 2010), which has 74 ROIs on each hemi-
sphere and was derived from 12 participants not involved
in this study. We selected five language-related ROIs from
which to report the volume, area, and thickness (see
Destrieux et al., 2010, for details on the landmarks used
to define these ROIs): (1) opercular part of the inferior
frontal gyrus (pars opercularis, POp), (2) triangular part of
the inferior frontal gyrus (pars triangularis; PTr), (3) PT, (4)
anterior transverse temporal gyrus (Heschlʼs gyrus, HG),
(5) insula composed of superior, anterior, and inferior
segments of the circular sulcus of the insula, short and long
insular gyri, and the central sulcus of the insula. The PT is
shown in an inflated hemisphere in Figure 5. We emphasize
that the anatomical ROIs are independent of the interhemi-
spheric registration. Note that the Destrieux PT definition
includes both the posterior horizontal segment and the
posterior ascending ramus (sometimes referred to as the
planum parietale or PP). These ROIs were chosen for
several reasons. POp and PTr approximate Brocaʼs area.
PT has prominent anatomic asymmetries and is an area of
language function. The insula has been found to have
an interaction between anatomy and language function
(Keller et al., 2011). HG was chosen because of known
anatomical asymmetry (Dorsaint-Pierre et al., 2006;
Penhune, Cismaru, Dorsaint-Pierre, Petitto, & Zatorre,
2003; Penhune, Zatorre, MacDonald, & Evans, 1996;
Rademacher, Caviness, Steinmetz, & Galaburda, 1993).
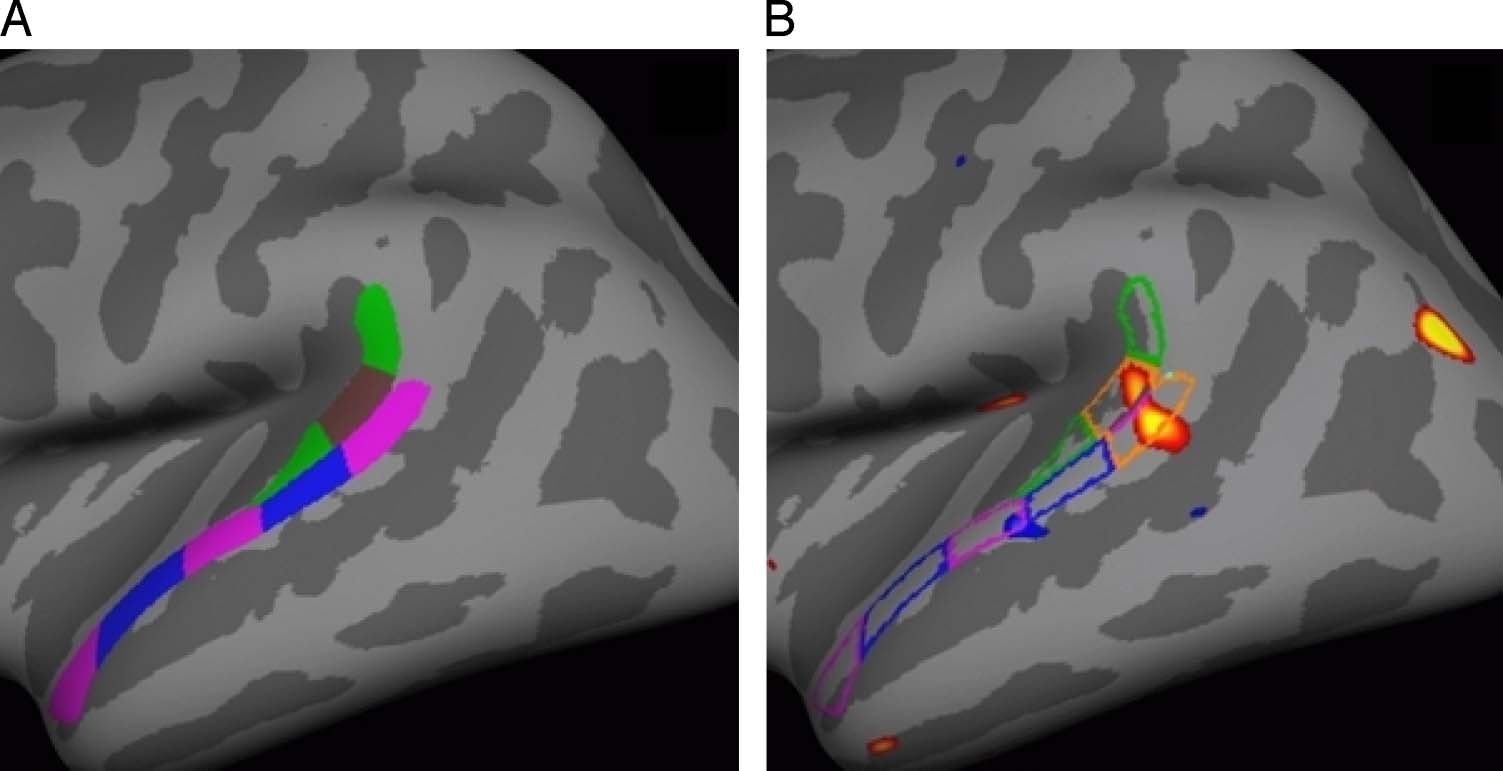
Figure 5. Method used to
geometrically construct an
ROI to cover the STG cluster.
(A) The PT was divided into
three sections along its long
axis (green–brown–green
labels). The lateral aspect of
the STG (LASTG) was divided
into five sections (pink–blue–
pink labels). (B) The second
section of the PT and the
first section of the LASTG
are combined to cover the
STG cluster fairly well (orange
outline). The significance
map is the same as in
Figure 2A1 and Figure 3.
1482
Journal of Cognitive Neuroscience
Volume 25, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
o
9
c
4
n
2
_
5
a
/
_
j
0
o
0
c
4
n
0
5
_
a
p
_
d
0
0
b
4
y
0
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Performing an anatomical ROI analysis also provides a
means through which the FreeSurfer ROI results can be
compared with those of other studies as well as with the
exploratory analysis.
Exploratory Spatial Analysis
Both hemispheres of all 65 participants were registered
to the symmetric template, and the vertex-wise cortical
GM volume maps were mapped into the symmetric stan-
dard space. These were then surface-smoothed by 10 mm
FWHM. For each participant, the volume LI at each vertex
was computed as per Equation 1. A statistical analysis was
performed at each vertex to evaluate the difference be-
tween the LLD and RLD participants using a two-group un-
signed t test with 53 degrees of freedom (DOF). Clusters
were defined as groups of contiguous vertices with vertex-
wise p < .01 (the cluster forming threshold). The p value
for a cluster was determined through Monte Carlo simula-
tion in which white Gaussian noise was repeatedly syn-
thesized on the surface, spatially smoothed, thresholded,
and clustered to determine the distribution of cluster sizes
under the null hypothesis (Hagler, Saygin, & Sereno,
2006). The cluster search space was constrained by two a
priori regions corresponding to an anterior language area
(ALA) that surrounds Brocaʼs area and a posterior language
area (PLA) that surrounds Wernickeʼs area, which are
often mentioned as traditional language areas (Toga &
Thompson, 2003). We defined the ALA by combining four
of the Destrieux ROIs: (1) POp, (2) PTr, (3) horizontal and
vertical ramus of the anterior segment of the lateral sulcus,
and (4) inferior part of the precentral sulcus. We defined
the PLA by combining Destrieux ROIs: (1) angular gyrus,
(2) supramarginal gyrus, (3) PT, (4) transverse temporal
sulcus, (5) posterior ramus of the lateral sulcus, (6) lateral
aspect of the superior temporal gyrus (STG; posterior
half ), (7) STS (posterior two thirds), and (8) sulcus inter-
medius primus of Jensen. The resulting ALA and PLA are
shown in yellow outline in Figure 2. Because this is an
exploratory analysis, the boundaries for these areas were
selected to be rather large so as to reduce the chance of
missing clusters. The ALA and PLA are larger and different
than the anatomical ROIs above so that the exploratory
analysis might allow the data to define new ROIs that are
not necessarily defined by landmarks visible to an anato-
mist. A cluster analysis using the whole hemisphere was
also performed to search for any clusters that might be
outside of traditional language areas.
RESULTS
Anatomically Defined ROIs
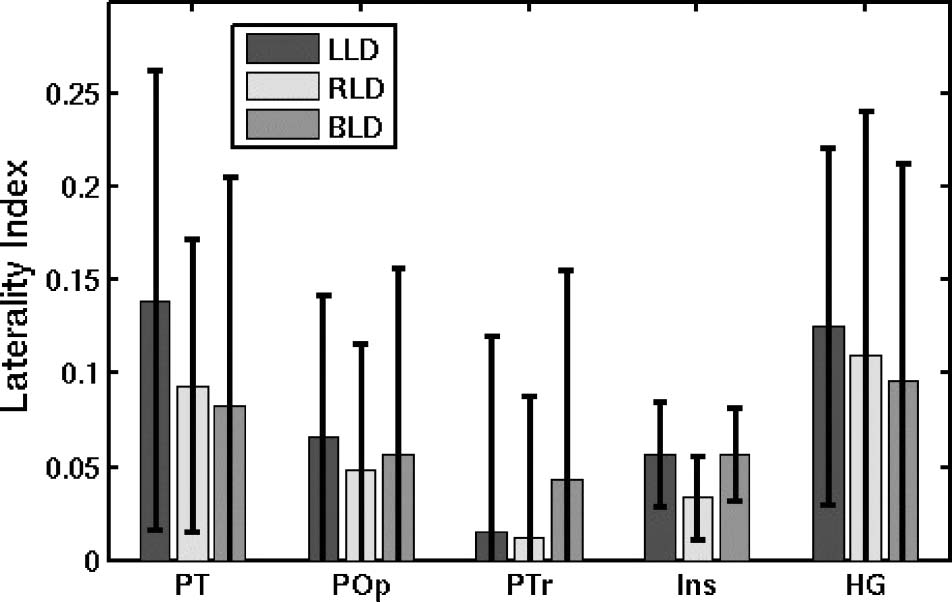
Figure 6 shows the average volume LI for the three groups.
The PT, POp, insula, and HG where all highly significantly
left-lateralized in both LLD and RLD ( p < .0001). Post hoc
analysis revealed that each ROI was significantly left
Figure 6. Mean volume LIs for anatomically defined ROIs (with
standard deviation bars). Ins: Insula.
lateralized in terms of surface area ( p < .0001), but not
thickness. The PTr was the only structure that was not
significantly left lateralized ( p = .31). All ROIs showed
numerically greater volume LIs for LLD over RLD; however,
the only structure to show a significant difference was
insula ( p = .0127, after correcting for five comparisons).
The PT significance was trending toward a difference ( p =
.13, uncorrected). The numerical laterality difference was
greater in PT than in insula, but the variability of insula
was much smaller leading to a greater effect size. Both
groups were left lateralized in insula, but LLD was more
so. A post hoc analysis revealed that LLD and RLD differed
significantly in insula area ( p = .0127), but not thickness
( p = .82), after correcting for five tests. The BLD group
showed roughly the same laterality pattern across the ROIs
as the LLD and RLD groups but was neither consistently
greater than nor less than LLD or RLD.
Exploratory Analysis
A whole-hemisphere map of the LLD-RLD difference is
shown in Figure 2. The left panel shows the results of
the LH-initialized template; the right panel shows the
results with the RH-initialized template. Figure 2 is a sig-
nificance map thresholded a vertex-wise uncorrected two-
sided threshold of p < .01 where red/yellow indicates
LLD > RLD and blue/cyan indicates RLD > LLD. Diese
statistics are shown on an “inflated” cortical surface of
the symmetric template where the light gray indicates a
gyrus and the dark gray indicates a sulcus. Within the
a priori regions (yellow boundaries), there is only one clus-
ter in STG that survives multiple comparison correction
(see also Figures 3A and 5B). No significant clusters were
found in the ALA. Outside these areas, there was a large
cluster in the ventral occipitotemporal (vOT) cortex that
survived whole hemisphere correction. See Table 2 for a list
of clusters, their p values, surface areas, and coordinates.
There are some other candidate clusters in Figure 2, Aber
they did not survive correction for multiple comparisons
Greve et al.
1483
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Tisch 2. Significant Clusters in the LLD–RLD Exploratory Analysis
Cluster
Initialization Hemisphere
Cluster p Value
Bereich (mm2)
STG
vOT
LH
RH
LH
RH
.0076
.0094
.0482
.0390
207
187
337
343
Centroid MNI305
Coordinates (xyz)
−58.2
58.6
−36.9
37.8
−39.9
−39.3
−49.6
−49.2
12.7
13.5
−17.8
−17.4
and so are not reported further. Both the maps in Figure 2
and the cluster summaries in Table 2 indicate the results
are robust to initialization hemisphere. For both the STG
and vOT clusters, the coordinates of the centroids between
LH and RH initialization are different by only about 1 mm
(after reversing the sign of the x coordinate).
Post hoc Tests and BLD Results
Post hoc tests were performed on the STG and vOT clus-
ters to determine whether the difference in groups was
being driven by laterality differences in one or both
groups and whether the underlying differences were be-
cause of volume, Bereich, or thickness differences. Abbildung 3B
shows a bar plot of the LIs for the STG cluster volume,
Bereich, and thickness broken down by group along with
p values corrected for eight post hoc tests. One can see
that there is a large difference in volume LI between the
groups; this is expected because this cluster was selected
for a volume LI difference (and why the LLD–RLD p value
for volume LI is not given). The LLD group has a positive
volume LI, and the RLD have a negative volume LI, beide
of which are significantly different than 0. This indicates
that both groups are asymmetric with opposite laterality.
The area LI is significantly different between the groups.
The LLD group has a significantly positive area LI; Die
RLD group has a negative area LI, but it was not signifi-
kippen. The LLD group shows no thickness laterality, Aber
the RLD group has a significantly thicker right cortex than
left in this cluster. There is a thickness LI difference be-
tween groups, but it does not survive correction for mul-
tiple post hoc tests. The results for the BLD group within
this cluster are also shown. On each measure, the BLD
group is between the LLD and RLD groups. We point
out that the BLD results are completely independent of
the participants used to construct the registration tem-
plate and the cluster.
A close-up of the vOT cluster is shown in Figure 4A
along with the post hoc tests in Figure 4B. The volume
is significantly lateralized in both groups in opposite di-
rections. There is a significant difference between groups
in area lateralization but not in thickness. The RLD group
is significantly rightward lateralized for both area and
Dicke. From this data, it appears that the difference
in GM volume LI in the vOT is being driven by area
and not thickness. The BLD group is between the LLD
and RLD in volume and surface area.
Confirmatory Cluster Analysis
The exploratory method developed in this manuscript is
novel, and so we would like to demonstrate that the STG
and vOT clusters are not merely an artifact of the non-
linear registration procedure or resampling of the indi-
vidual volume maps into template space. Im Idealfall, Die
STG and vOT clusters would be labeled independently
in each participant; these labels could then be used to
compute LLD and RLD statistics to verify results from
the exploratory analysis. The problem is to get clusters
in each participant that are independent of the registra-
tion and resampling procedures. We cannot simply map
the cluster from the template space back into the indi-
vidual space because that would be dependent upon
the registration. Stattdessen, we exploit the geometrical
relationship between the clusters and the labels of the
Destrieux atlas. Zum Beispiel, the STG cluster can be
approximated in the following way (siehe Abbildung 5). Erste,
the PT is divided into three sections along its long axis
(the green–brown–green labels in Figure 5A). Nächste, Die
lateral aspect of the STG (LASTG) is divided into five sec-
tionen (the pink-blue-pink labels in Figure 5A). When the
second section of the PT and the first section of the
LASTG are combined into a single ROI (orange outline
in Figure 5B), the STG cluster is covered fairly well.
The same geometric operations can be performed in
each individual brain to get an approximation of the
STG cluster without resorting to the registration used
to create the STG cluster. The results (Figur 7) have a
similar pattern to that of the original cluster analysis.
The volume LI for the LLD is still significantly greater than
that of the RLD ( P < .02), and the BLD is still between
the LLD and RLD. The difference is that all LIs are shifted
by about 0.13 to be more positive (e.g., RLD is now pos-
itive instead of negative). This shift is probably because of
the fact that the approximate STG cluster does not cover
the actual cluster perfectly. A similar analysis was applied
to the vOT cluster with consistent findings. Although the
confirmatory ROI is obviously informed by the exploratory
map-based analysis, the boundaries of the constructed
ROI in each participant are completely independent of
1484
Journal of Cognitive Neuroscience
Volume 25, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
o
9
c
4
n
2
_
5
a
/
_
j
0
o
0
c
4
n
0
5
_
a
p
_
d
0
0
b
4
y
0
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
sures for these participants. These include fMRI of an
LDT (Van der Haegen et al., 2012) and three behav-
ioral tasks: Dichotic Listening (DL, Van der Haegen,
Westerhausen, Hugdahl, & Brysbaert, 2013), VHF task
with pictures as stimuli ( VHFP), and VHF task with
words as stimuli ( VHFW; Van der Haegen et al., 2011).
We computed LIs for each of these behavioral and fMRI
measures. We also have the fMRI results for the subvocal
WG task; however, the correlation between the WG task
LI and the cluster LIs is circular because the WG task was
used to assign participants into LLD, RLD, and BLD. To
gain further insight into the functional correlates of the
anatomical asymmetries, we computed the correlation
coefficient between all pairs of LIs. The resulting matrix
is shown in Table 3. The behavioral tasks returned the
expected asymmetries: a right ear advantage in the DL
task for the LLD and a left ear advantage for the RLD,
and RVF advantages for LLD versus LVF advantages
for RLD. The only a priori defined ROI that correlated
with function was the insula, which correlated with ear
advantage in the DL task and fMRI laterality in the WG
task. The exploratory clusters showed higher correla-
tions with functional data: The STG cluster correlated
with LDT and VHFP. The vOT cluster correlated with
LDT and DL, but, surprisingly, not with the VHF tasks.
The correlations in Table 3 are based on all participants;
further analyses indicated that the conclusions remain
the same if the bilateral participants are omitted or when
Figure 7. Comparison of volume LI within the STG (exploratory)
cluster and within the (confirmatory) anatomically constructed ROI
(with standard deviation bars across subject). The bars for the
LLD and RLD are the same as the “Volume” bars in Figure 3.
the interhemispheric registration and so confirmatory
relative to this registration.
Correlations with fMRI and Behavioral Measures
In addition to the anatomical information, we also have
access to various functional language lateralization mea-
Table 3. Correlation Coefficients between Anatomical Asymmetries and Functional Data (Number of Participants in the Lower Left Half )
Anatomical Measures
fMRI
Behavioral
PT
65
65
65
65
65
65
65
52
42
58
58
POp
−.16
65
65
65
65
65
65
52
42
58
58
PT
POp
PTr
HG
Insula
STG
vOT
WG
LDT
DL
VHFP
VHFW
PTr
HG
Insula
STG
vOT
WG
−.12*
−.05
65
65
65
65
65
52
42
58
58
.11
.12
.10
65
65
65
65
52
42
58
58
−.06
.42**
.12 −.03
.15 −.11
−.10
.03
.15*
65
65
65
52
42
58
58
65
65
52
42
58
58
.10
.11
−.00
−.15
.27**
.41a
65
52
42
58
58
.17*
.16
−.06
.08
.38*
.46a
.47a
52
42
58
58
LDT
.05
.00
−.17
−.17
−.03
.26
.31*
.62**
35
48
48
DL
−.05
.16
.25
.07
.38*
.20
.49**
.58**
.23
38
38
VHFP
VHFW
.09
−.03
−.02
.08
.20
.11
.06
.53**
.23*
.38*
.60**
.07
.01
−.18
.05
.16
.27*
.11
.51**
.46**
.21
58
WG = LI fMRI BOLD signal in WG; LDT = LI fMRI BOLD signal in lexical decision; DL = LI accuracy dichotic listening; VHFP = LI naming times
pictures; VHFW = LI naming times words.
*p < .05 in this analysis or in the analysis of participants with data on all measures (n = 34).
**p < .05 in both analyses.
aA cell where the correlation is circular, for example, the STG and vOT clusters were defined based on fMRI activation in WG, so a high correlation is
expected.
Greve et al.
1485
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
o
9
c
4
n
2
_
5
a
/
_
j
0
o
0
c
4
n
0
5
_
a
p
_
d
0
0
b
4
y
0
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the analyses are limited to those participants for whom
we have data on all measures.
DISCUSSION
This study tested the surprising finding of Keller et al.
(2011) that only the volume of the insula differs between
healthy LLD and RLD participants (language dominance
based on asymmetry in the fMRI BOLD signals during a
silent WG task). We made use of the MRI scans collected
by Van der Haegen et al. (2011), augmented with 19 scans
collected later (footnote 2). In addition, we made use of
the FreeSurfer software, which allowed us to run explor-
atory analyses with a greater precision.
Our results replicate Keller et al. for all regions tested by
them. Only the size of the insula differed significantly be-
tween LLD and RLD participants. No significant differences
were found for Brocaʼs area or the PT, although the left-
ward asymmetry tended to be smaller for RLD participants
than for LLD participants, which is intuitively more accept-
able than the reverse trends reported by Keller et al. We
also failed to replicate the rightward insular asymmetry in
RLD participants, reported by Keller et al. As for the other
brain regions, we found evidence for a reduced left asym-
metry. In addition, we observed two new areas with reliable
differences between the LLD and RLD groups (showing
reversed asymmetries in the two groups). These were
STG and vOT, two regions involved in language perception
in the spoken and written modalities, respectively.
Our data, together with those of Keller et al. (2011),
point to a divergence between the anatomical and
functional asymmetries in participants with right speech
dominance. Although the participants had Brocaʼs area
clearly lateralized to the right and we know that most of
them also had spoken and written language perception
lateralized to the RH (Van der Haegen et al., 2012, 2013),
their overall volume asymmetry was leftward (Figure 6).
This casts doubts on the hypothesis that the anatomical
asymmetry of the human brain is the direct cause of the
functional asymmetries observed. This finding also seems
to put boundary conditions on the extent to which brain
use can modulate GM volume. After all, despite strong
functional asymmetries in language ( Van der Haegen
et al., 2011, 2012), tool use ( Vingerhoets et al., 2013),
and visuospatial processing (Cai et al., 2013), the anatomi-
cal differences between both groups were surprisingly
small in size and area.
In the remainder of the discussion, we give a more de-
tailed description of the findings in the different ROIs and
we end with some methodological observations, given
that this study also introduces various new ways of brain
analysis.
PT
The PT has been claimed to have a central role in spoken
language processing. Geschwind and Levitsky (1968),
found significant anatomical laterality in PT, and hypothe-
sized that this was related to language laterality. Several
studies to test this hypothesis followed, many of which
found that PT was lateralized to the left (Keller et al.,
2007, 2011; Dos Santos Sequeira et al., 2006; Eckert,
Leonard, Possing, & Binder, 2006), although not all
(Dorsaint-Pierre et al., 2006). Foundas, Leonard, Gilmore,
Fennell, and Heilman (1994) found PT asymmetry that
was reversed with RLD but only had one RLD participant.
Other studies have not been able to find a PT difference
between LLD and RLD (Keller et al., 2011; Dorsaint-Pierre
et al., 2006), although Tzourio et al. (1998) reported that
the area of left PT (rather than LI) was predictive of lan-
guage laterality. We also looked for unilateral differences
in PT surface area and found none. In fact, we found no
unilateral LLD–RLD differences on any of our measures.
As with other studies, we found a significant left later-
alization to PT in both LLD and RLD. As with Keller et al.
(2011) and Dorsaint-Pierre et al. (2006), we did not find
an LLD–RLD difference when analyzing the anatomically
defined PT. The asymmetry in volume was driven by
asymmetry in PT surface area and not in thickness. In the
exploratory analysis, however, a small area that partially
overlapped with both the PT and LASTG was found in
which volume LI was greater for LLD than RLD. A
post hoc analysis showed that the LLDs were leftward later-
alized (positive LI) whereas the RLDs were rightward later-
alized (negative LI). It also showed that LLD > RLD in
surface area LI but not in thickness. The STG is specifically
involved in processing the phonetic/phonological aspects
of spoken language (Turkeltaub & Coslett, 2010; Desai,
Liebenthal, Waldron, & Binder, 2008). It has also been
linked to auditory STM (Richardson et al., 2011). Interest-
ingly, this region does not seem to fit well within Hickok
and Poeppel (2007) model of speech processing, according
to which STG is involved in the spectrotemporal analysis of
auditory input and does so bilaterally. Andererseits,
the STG cluster is close to an area called SPT (sylvian parietal
temporal cortex). Hickok and Poeppel (2007) speculate that
area SPT is involved in translation between sensory speech
codes and the motor system (though this function extends
beyond speech tasks). Given that the functional asymmetry
was based on asymmetric fMRI results of a WG task, es ist
not impossible that the STG cluster overlaps with SPT.
At first glance, the exploratory analysis appears to con-
tradict the ROI results, which did not show a difference in
PT. Jedoch, the PT ROI was trending toward signifi-
Abbruch, and the STG cluster comprised only a small part
of PT. Analyzing the entire PT may wash out this effect.
An advantage of the exploratory technique is that it can
find clusters that do not fall into standard anatomical
definitions. This finding does add some support to the
Geschwind and Levitsky (1968) Hypothese.
The only other research to apply an exploratory
method to the study of LLD versus RLD was Dorsaint-
Pierre et al. (2006). They used VBM in a group of epilepsy
Patienten. Dorsaint-Pierre et al. (2006) did not find any
1486
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
LLD–RLD differences in the PT/STG area (although they
did find that simple laterality was significantly positive).
This could be for several reasons. Erste, they only had
11 RLD participants. Zweite, they used a sample of epi-
lepsy patients who may have reorganized language areas
(Brazdil, Zakopcan, Kuba, Fanfrdlova, & Rektor, 2003).
Endlich, it may be that VBM is less sensitive than surface-
based techniques to these types of effects.
Josse et al. (2009) performed a similar VBM analysis.
Jedoch, instead of classifying participants as LLD or
RLD, they performed a voxel-by-voxel regression analysis
between GM concentration asymmetry and fMRI activa-
tion asymmetry and found widespread correlation, In-
cluding in the PT/STG region. Although this supports
our results, there are several potential differences with
unsere Arbeit. Josse et al. (2009) report having 12 RLD par-
ticipants; Jedoch, by our definition of RLD (d.h., LI <
−.6), they would have had only four RLD. Therefore, it
is probably the case that our RLD participants are signifi-
cantly more right lateralized for language. In addition,
they found large correlations between GM density and
functional activation across the brain even for non-
language tasks. This suggests that the correlations may
have been an artifact possibly caused by partial voluming,
that is, if there is more GM in a voxel, then one would
expect more functional activation in that voxel.
POp and PTr
The POp and PTr comprise Brocaʼs area, which is related
to speech production.4 Studies by Keller et al. (2007,
2011) both found that POp was left lateralized but that
PTr was not. Keller et al. (2011) did not find an LLD–
RLD difference in either POp or PTr. Our anatomically
defined ROI results are in complete agreement as are
the exploratory results despite the fact that Keller et al.
(2007, 2011) employed manual labeling and used only
right-handed participants. We also found that the POp
laterality was driven by left–right differences in surface
area, not thickness. Dorsaint-Pierre et al. (2006), how-
ever, did find an LLD–RLD difference in GM concentra-
tion in Brocaʼs area using (exploratory) VBM. The use
of an epilepsy patient population may contribute to this
discrepancy. Dorsaint-Pierre et al. (2006) also comment
that their findings may be because of differences in cur-
vature of the cortex that can occur in certain types of
epilepsy and not because of change in GM volume. If
so, then the differences may not have shown up in this
surface-based because we only analyzed volume, area,
and thickness.
Insula
Keller et al., 2011). The numerical difference between LIs
for LLD and RLD was actually smaller for insula as com-
pared with PT. The significance came from smaller vari-
ability, which may be because of the fact that the insula
is a much larger structure and so its total volume will
not be very sensitive to small changes in its boundaries
or perhaps could be because of the fact that the circular
sulcus and the gyri of the insula are more easily identi-
fied. In Keller et al. (2011), the variance of insula was
also much less than for other structures, but they also
found a large difference in laterality. In this study, both
LLD and RLD had significant leftward asymmetries. Kel-
ler et al. (2011) showed a positive LI for LLD, but a ne-
gative LI for RLD. The reason for this is unclear but
perhaps related to the use of left-handers in this study
and right-handers in Keller et al. (2011). Interestingly,
there was not a cluster in insula in the exploratory ana-
lysis. Further investigation showed a consistent but
small LLD > RLD advantage across insula in the explora-
Tory-Analyse. It appears that this advantage is not strong
enough on a vertex-by-vertex basis to create a cluster,
but it is strong enough when averaged over the entire
insula. Keller et al. (2011) came to a similar conclusion
when they explored the spatial pattern of LLD/RLD dif-
ferences in insula.
Given our replication of Keller et al., we fully agree
with those authors that the role of the insula in language
processing needs further investigation, also given the
correlations with DL performance (Tisch 3). The insula
seems to be particularly involved in overt speech,
although there is also some evidence for a role in covert
Rede (Price, 2012), and insular lesions are related to
aphasia (Price, 2000; Donnan, Darby, & Saling, 1997).
Andererseits, the insula plays a role in a wide range
of intermodal functions (Flynn, Benson, & Ardila, 1999),
making its involvement in language processing less pro-
nounced than other peri-sylvian areas. Future research is
needed to investigate whether the central location within
the peri-sylvian network makes the insula one of the key
regions to determine language asymmetry.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
.
/
HG
HG, primary auditory cortex, has been studied in relation
to language laterality (Dorsaint-Pierre et al., 2006). Dort
was not a significant difference in HG anatomical asym-
metry between LLD and RLD in agreement with manual
labelings in Dorsaint-Pierre et al. (2006). We did find that
HG was significantly lateralized to the left, in agreement
with other studies (Dorsaint-Pierre et al., 2006; Penhune
et al., 1996, 2003; Rademacher et al., 1993).
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
There was a significant difference between LLD and RLD
in the volume asymmetry of insula (in agreement with
Keller et al., 2011). This difference was driven by area
asymmetries and not thickness (again in agreement with
vOT Cortex
The only other area to be significantly different be-
tween LLD and RLD was in the vOT with LLDs being more
Greve et al.
1487
left-lateralized and RLDs more right-lateralized in GM
Volumen. Dorsaint-Pierre et al. (2006) did not find an
LLD–RLD difference in vOT. To our knowledge, this is
the first report of anatomical asymmetry differences
between LLD and RLD in vOT. This area has been linked
to visual word recognition (Dehaene & Cohen, 2011;
Ben-Shachar, Dougherty, Deutsch, & Wandell, 2007;
Cohen et al., 2000), face recognition (Kanwisher, McDermott,
& Chun, 1997), and body and object recognition (Taylor &
Downing, 2011). The asymmetries observed are in line
with the functional lateralization of visual word recognition
(Van der Haegen et al., 2012; Seghier & Price, 2011; Cai
et al., 2008, 2010). Reduced GM volume in the left or bi-
lateral vOT regions has been found to be associated with
dyslexia (Raschle, Chang, & Gaab, 2011). The face recogni-
tion area (d.h., the fusiform face area) is also known to be
functionally lateralized but in the opposite direction rela-
tive to the visual word recognition area. Jedoch, we do
not see an area in Figure 4 where RLD > LLD inside of
vOT. A possible reason for this is that all the participants
in this study were left-handers and the FFA is less later-
alized in left-handers than right-handers (Willems, Peeling,
& Hagoort, 2010). Another reason might be that FFA and
the visual word recognition area are complementary (Plaut
& Behrmann, 2011; Dien, 2009) and cannot be separated
anatomically.
It does not come as a surprise that asymmetry in the
WG task predicts anatomical asymmetry in vOT. Beweis
suggests that phonological information contributes to
visual word recognition (Price & Devlin, 2011). Hemi-
sphere dominance for word reading tends to follow that
of speech production (Van der Haegen et al., 2012; Cai
et al., 2008, 2010), perhaps reflecting the need for fast
interactions between these areas. Auch, see below for cor-
relations between vOT and the LDT. In the history of the
human species, reading is a very recent skill with no real
evolutionary pressure for laterality yet. The fact that it is
lateralized is arguably because of the close interactions
with Brocaʼs area.
Bilateral Participants
Eleven participants did not show strong language lateral-
ität. In the anatomically defined ROI analysis, they showed
a similar laterality pattern as the LLD and RLD partici-
Hose, but there was no systematic relationship. Im
exploratory and confirmatory analysis of the STG cluster,
the LI for volume, Bereich, and thickness consistently fell
between LLD and RLD.
Correlations between fMRI and
Behavioral Measures
In line with the absence of a clear reversal of the overall
anatomical asymmetry in RLD participants (Figur 1),
most of the LIs in the anatomically defined ROIs were
uncorrelated with functional asymmetries. This was true
both for asymmetries based on fMRI BOLD signals ( WG
and LDT) and on behavioral data (DL, VHFP, and VHFW).
Only insula showed correlations with the functional
asymmetries (DL and WG). The STG cluster correlated
with VHFP but, überraschenderweise, not with DL. The anatomical
asymmetries in the vOT cluster correlated with DL and
LDT, in line with the proposal that this area plays a
central role in the integration of phonological and ortho-
graphic word information as mentioned above. The find-
ing that anatomical vOT asymmetries are correlated with
the ear advantage in DL is particularly noteworthy, wie es ist
in line with the hypothesis that the orthography of a
word helps the processing of spoken words in literate
individuals (Ziegler & Ferrand, 1988). It is surprising vOT
is not correlated with the VHF differences in word naming
and picture naming.
Statistical Model Selection
The statistical analysis requires a model of the relation-
ship between the functional LI (fLI) and structural LI
(sLI). One of two models is typically used: (1) a cate-
gorical model in which participants are divided into lateral-
ity classes; differences in class means are evidence of an
interaction between structure and function, oder (2) A
continuous model in which it is assumed that fLI and
sLI are linearly related; a nonzero slope is evidence of
an interaction. There are advantages and disadvantages
to both methods. Categorization models the fLI–sLI rela-
tionship as piece-wise constant. Some researchers (z.B.,
Keller et al., 2011) use two categories (LLD and RLD),
Andere (z.B., Dorsaint-Pierre et al., 2006) use three
(LLD, RLD, and BLD). We chose three for several reasons.
Erste, bilateral participants may represent a qualitatively
different language organization than the mirror reversal
hypothesized between LLD and RLD. Zum Beispiel, in einem
Wada test, bilaterality may manifest as both hemispheres
being impaired or neither being impaired (Dorsaint-Pierre
et al., 2006; Risse, Gates, & Fangman, 1997). Zweite,
having three classes makes the piece-wise constant model
more accurate in the case where there is continuous
Variation. We chose a relatively high threshold of 0.6 Zu
assure that our results were not influenced by bilateral
Teilnehmer (d.h., we would rather exclude a few true LLDs
or RLDs rather than risk the inclusion of BLDs).
A continuous model is attractive because language lat-
erality is never an “all-or-nothing” trait. The continuous
model only requires one regressor, saving several DOFs
relative to the categorical model—a very important point
when sample sizes are small. The ability to include bi-
lateral participants also increases the DOF, Aber, as men-
tioned above, it is not clear that the bilateral participants
belong with the others. The continuous model requires a
parametric function relating fLI and sLI (generally linear).
This is a very strong assumption given that the sLI and fLI
are computed from very different data and methods. In einem
1488
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
F
T
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
voxelwise structural analysis, this relationship could
change from region to region.
Daher, the categorical model makes fewer assumptions
than the continuous model but may have less power. Der
exact nature of the fLI–sLI relationship is complex and
will require much more data to sort out. For complete-
ness, we performed a follow-up test using a continuous
linear model. Although the exact p values changed, Dort
were no changes in the conclusions made from either the
ROI or the exploratory analysis (d.h., the STG and vOT
clusters and insula ROI were still significant and no new
areas became significant).
Methodological Considerations
This study exploited some unique methodologies. Erste,
we had a relatively large sample (n = 21) of healthy RLD
participants made possible by a low-cost and noninvasive
screening procedure. The sample was mainly selected
after screening 250 left-handers for behavioral signs of
RLD. Our sample is twice as large as comparable studies
(n = 10, Keller et al., 2011; n = 11, Dorsaint-Pierre et al.,
2006; n = 4, Eckert et al., 2006). One disadvantage of the
study is that the participants are only left-handers and
mostly women. This prevented us from exploring the
relationship of handedness and gender on the interaction
between language and anatomical laterality.
We employed automated surface-based methods to
compute anatomical measures. These methods include
3-D mesh models with subvoxel accuracy; surface-
based registration that aligns folding patterns across sub-
ject and hemisphere; and automated algorithms that
attempt to replicate the manual labeling of an anatomist.
The volumes of the anatomically defined ROIs were in
the range of published manual results but consistently
smaller. Zum Beispiel, we measured the left PT volume
(including PP) in LLD to be 2204 mm3 whereas other
Studien (Keller et al., 2007, 2011; Dos Santos Sequeira
et al., 2006) that employed manual labelings were in
the range of 3000–4000 mm3. A possible limitation of
the PT results is that the Destrieux definition of PT
included the PP, which is normally excluded. This may
have the effect of reducing asymmetry. In der Tat, the PT
LIs in this study (.09–.14) are much less than that of
Dos Santos Sequeira et al. (2006). Jedoch, they are
more than the PT LIs in Keller et al. (2007) (um .11)
and about the same as in Keller et al. (2011) (.09–.14).
The POp and PTr volumes in this study are about 66%
of that found in Keller et al. (2007). Although some of
this may have to do with the landmark definitions, it may
also be because of partial voluming. Voxels that are partially
in GM and WM will have an intensity that is darker than the
typical WM voxel, and a manual rater may be inclined to
label these as GM. The surface-based mesh model will
cut through the middle of such voxels and thereby give a
more accurate (and smaller) measure of GM volume.
Despite this difference, the ratio of LH to RH PT volume
in this study (1.3) is very close to that found in Dos Santos
Sequeira et al. (2006) (1.4), Keller et al. (2007) (1.2), Und
Keller et al. (2011) (1.17). For POp, the simple volume LI
was very similar for POp across the studies (0.12 for this
Studie, 0.11 for Keller et al., 2011, and about 0.13 for Keller
et al., 2007). There is a striking similarity between the ROI
results presented here and those of Keller et al. (2011)
despite appreciable differences in methodology. Obwohl
the ROI results are similar to previous work, we point out
that the exploratory results are novel.
The surface-based registration allowed an exploratory
analysis to search for areas of significant LLD/RLD differ-
ence that do not lie cleanly within the boundaries pre-
scribed by manual labels. Such a difference was found in
a cluster whose extent covered only part of the PT. Der
exploratory effect did not show up in the ROI approach
because this cluster did not fit neatly into an anatomically
defined region. The exploratory approach also allows
evaluating brain regions that are outside of traditional
language areas and so not targeted for tedious manual
labeling procedures. Such a region was found in vOT.
The interhemispheric registration procedure was veri-
fied in two ways. Erste, the entire study was replicated
using the RH and LH as initial targets, with very similar
results. Zweite, an ROI covering the exploratory STG
cluster was defined based on anatomical landmarks
(and so independent of the registration). This ROI
showed significant differences between LLD and RLD.
Although this by itself does not confirm a difference be-
tween LLD and RLD, it does show that the difference
found in the exploratory analysis is not a byproduct or
artifact of the registration method. The exploratory surface-
based analysis is different from VBM analysis in several
ways. It uses surface-based registration, which has been
shown to be superior to volume-based in many instances
(Tucholka, Fritsch, Polina, & Thirion, 2012; Zollei, Stevens,
Huber, Kakunoori, & Fischl, 2010; Postelnicu, Zollei, &
Fischl, 2009; Fischl et al., 2008). The surface-based method
allows the differences in GM volume to be decomposed
into effects of surface area and thickness. This is important
for shedding light on the origins of asymmetry. For exam-
Bitte, surface area and thickness are both highly heritable but
genetically independent (Winkler et al., 2010; Panizzon
et al., 2009). Separately measuring surface area and thick-
ness can also be important in understanding disorders. Für
Beispiel, dyslexia and reading skills have been found to be
related to surface area but not to thickness (Frye et al.,
2010). GM volume loss in aging is related to loss of surface
area not cortical thinning; Jedoch, cortical thinning
accompanies Alzheimer disease (Dickerson et al., 2009).
Andererseits, VBM may be sensitive to less specific
aspects of morphometry (Palaniyappan & Liddle, 2012).
Abschluss
The purpose of this study was to investigate the inter-
action between functional language laterality and anatomical
Greve et al.
1489
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
/
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
laterality. There were several novel aspects to the study
including the recruitment of a relatively large healthy sample
of RLD participants made possible by a previously devel-
oped low-cost and noninvasive screening procedure. Das
study also used automated surface-based analysis tech-
niques that provide several useful features: the ability to
automatically delineate standard language-related anatomi-
cal ROIs such as PT, POp, and PTr; the ability to compute
GM volume with subvoxel accuracy; the ability to decom-
pose differences in GM volume into differences in surface
area and thickness; and the ability to use surface-based
registration to align folding patterns across hemisphere as
well as participant for use in an exploratory analysis. Der
anatomical ROI-based results replicated those of a similar
study using manual labeling in healthy right-handers (Keller
et al., 2011) in that no LLD–RLD differences were found in
PT, POp, and PTr, but a strong difference was found in insula
GM volume. Interessant, the exploratory analysis did reveal
a small LLD > RLD cluster in STG that overlapped with PT.
This capability shows the utility of exploratory analysis for
finding structural differences that may cross the boundaries
of anatomically defined ROIs. The exploratory analysis also
indicated an LLD–RLD difference in vOT, an area associated
with visual word recognition. No other differences between
LLD and RLD were found in the rest of the brain, including in
Brocaʼs area. In all cases, differences in GM volume were
attributable to a difference in cortical surface area and not
cortical thickness. The sample used in this study was exclu-
sively left-handed and mostly female, making it impossible
to evaluate the effect of handedness and gender; Jedoch,
the use of a practical language dominance screening proce-
dure and automated surface-based analysis should make
more large-scale studies of language dominance possible.
Danksagungen
Support for this research was provided in part by the National
Institutes of Health (R01NS052585-03), the National Institute on
Altern (5R01AG029411, 5R01AG022381-08), the National Institute
of Neurological Disorders and Stroke (1R01NS069696-01A1), Die
National Center for Research Resources (P41-RR14075, R01
RR16594-01A1, the NCRR BIRN Morphometric Project BIRN002,
and Functional Imaging Biomedical Informatics Research Network
[FBIRN] U24 RR021382), and the National Institute for Biomedical
Imaging and Bioengineering (R01 EB001550, R01 EB006758, 1K25
EB013649-01). The scanning was made possible by an Odysseus
Grant paid by the Government of Flanders to M. B.
Reprint requests should be sent to Douglas N. Greve, Der
Martinos Center for Biomedical Imaging, Department of
Radiology, Massachusetts General Hospital, Room 2301, 149
13th Street, Charlestown, MA 02129, oder per E-Mail: greve@nmr.
mgh.harvard.edu.
Notes
It was also found that participants with right hemisphere
1.
speech lateralization had an atypical
left-ear advantage in a
dichotic listening task (Van der Haegen et al., 2013), vorschlagen
that the brain areas involved in spoken word recognition are
atypically lateralized as well, but no fMRI data have been col-
lected on this yet.
2. We tested 19 participants more after the Van der Haegen
et al. (2011) manuscript was accepted for publication. Most of
these additional participants were not previously screened with
the VHF tasks.
3. As can be seen in the Results section, this turned out to be
the case for about half of the participants.
4. Modern conceptions of Brocaʼs area are more complex
than simply a “motor speech” area, Zum Beispiel, Carreiras,
Pattamadilok, Meseguer, Barber, and Devlin (2012) have found
evidence that Brocaʼs area is involved in the understanding of
syntax.
VERWEISE
Amunts, K., Weiss, P. H., Mohlberg, H., Pieperhoff, P., Eickhoff,
S., Gurd, J. M., et al. (2004). Analysis of neural mechanisms
underlying verbal fluency in cytoarchitectonically defined
stereotaxic space-the roles of Brodmann areas 44 Und 45.
Neurobild, 22, 42–56.
Aschenbrenner, J., & Friston, K. J. (2000). Voxel-based morphometry-
The methods. Neurobild, 11, 805–821.
Ben-Shachar, M., Dougherty, R. F., Deutsch, G. K., & Wandell,
B. A. (2007). Differential sensitivity to words and shapes in
ventral occipito-temporal cortex. Hirnrinde, 17,
1604–1611.
Benson, D. F., & Geschwind, N. (1985). Aphasia and related
disorders: A clinical approach. In M. M. Mesulam (Ed.),
Principles of behavioral neurology (S. 193–228).
Philadelphia, PA: FA Davis.
Benson, R. R., FitzGerald, D. B., LeSueur, L. L., Kennedy, D. N.,
Kwong, K. K., Buchbinder, B. R., et al. (1999). Language
dominance determined by whole brain functional MRI in
patients with brain lesions. Neurologie, 52, 798–809.
Binder, J. R., Swanson, S. J., Hammeke, T. A., Morris, G. L.,
Mueller, W. M., Fischer, M., et al. (1996). Determination of
language dominance using functional MRI: A comparison
with the Wada test. Neurologie, 46, 978–984.
Brazdil, M., Zakopcan, J., Kuba, R., Fanfrdlova, Z., & Rektor, ICH.
(2003). Atypical hemispheric language dominance in left
temporal lobe epilepsy as a result of the reorganization of
language functions. Epilepsy & Behavior, 4, 414–419.
Cai, Q., Lavidor, M., Brysbaert, M., Paulignan, Y., & Nazir, T. A.
(2008). Cerebral lateralization of frontal lobe language
processes and lateralization of the posterior visual word
processing system. Zeitschrift für kognitive Neurowissenschaften, 20,
672–681.
Cai, Q., Paulignan, Y., Brysbaert, M., Ibarrola, D., & Nazir, T. A.
(2010). The left ventral occipito-temporal response to words
depends on language lateralization but not on visual
Vertrautheit. Hirnrinde, 20, 1153–1163.
Cai, Q., Van der Haegen, L., & Brysbaert, M. (2013).
Complementary hemispheric specialization for language
production and visuospatial attention. Verfahren der
Nationale Akademie der Wissenschaften, USA., 110, E322–E330.
Carreiras, M., Pattamadilok, C., Meseguer, E., Barber, H.,
& Devlin, J. T. (2012). Brocaʼs area plays a causal role in
morphosyntactic processing. Neuropsychologie, 50, 816–820.
Cohen, L., Dehaene, S., Naccache, L., Lehericy, S., Dehaene-
Lambertz, G., Henaff, M. A., et al. (2000). The visual word
form area: Spatial and temporal characterization of an initial
stage of reading in normal subjects and posterior split-brain
Patienten. Gehirn, 123, 291–307.
Dale, A. M., Fischl, B., & Sereno, M. ICH. (1999). Cortical surface-
based analysis I: Segmentation and surface reconstruction.
Neurobild, 9, 179–194.
1490
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
/
.
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Dehaene, S., & Cohen, L. (2011). The unique role of the visual
word form area in reading. Trends in den Kognitionswissenschaften, 15,
254–262.
Desai, R., Liebenthal, E., Waldron, E., & Binder, J. R. (2008).
Left posterior temporal regions are sensitive to auditory
categorization. Zeitschrift für kognitive Neurowissenschaften, 20,
1174–1188.
Desikan, R. S., Segonne, F., Fischl, B., Quinn, B. T., Dickerson,
B. C., Blacker, D., et al. (2006). An automated labeling
system for subdividing the human cerebral cortex on MRI
scans into gyral based regions of interest. Neurobild, 31,
968–980.
Destrieux, C., Fischl, B., Dale, A., & Halgren, E. (2010).
Automatic parcellation of human cortical gyri and sulci
using standard anatomical nomenclature. Neurobild, 53,
1-15.
Dickerson, B. C., Feczko, E., Augustinack, J. C., Pacheco, J.,
Morris, J. C., Fischl, B., et al. (2009). Differential effects of
aging and Alzheimerʼs disease on medial temporal lobe
cortical thickness and surface area. Neurobiologie des Alterns,
30, 432–440.
Dien, J. (2009). A tale of two recognition systems: Implications
of the fusiform face area and the visual word form area for
lateralized object recognition models. Neuropsychologie, 47,
1–16.
Donnan, G. A., Darby, D. G., & Saling, M. M. (1997).
Identification of brain region for coordinating speech
articulation. Lancet, 349, 221–222.
Dorsaint-Pierre, R., Penhune, V. B., Watkins, K. E., Neelin, P.,
Lerch, J. P., Bouffard, M., et al. (2006). Asymmetries of the
planum temporale and Heschlʼs gyrus: Relationship to
language lateralization. Gehirn, 129, 1164–1176.
Dos Santos Sequeira, S., Woerner, W., Walter, C., Kreuder, F.,
Lueken, U., Westerhausen, R., et al. (2006). Handedness,
dichotic-listening ear advantage, and gender effects on
planum temporale asymmetry-A volumetric investigation
using structural magnetic resonance imaging.
Neuropsychologie, 44, 622–636.
Eckert, M. A., Leonard, C. M., Possing, E. T., & Binder, J. R.
(2006). Uncoupled leftward asymmetries for planum
morphology and functional language processing. Gehirn
and Language, 98, 102–111.
Eckert, M. A., Lombardino, L. J., Walczak, A. R., Bonihla, L.,
Leonard, C. M., & Binder, J. R. (2008). Manual and automated
measures of superior temporal gyrus asymmetry: Concordant
structural predictors of verbal ability in children.
Neurobild, 41, 813–822.
Fischl, B., & Dale, A. M. (2000). Measuring the thickness of the
human cerebral cortex from magnetic resonance images.
Verfahren der Nationalen Akademie der Wissenschaften, 97,
11044–11049.
Fischl, B., Rajendran, N., Busa, E., Augustinack, J., Hinds, O.,
Yeo, B. T., et al. (2008). Cortical folding patterns and
predicting cytoarchitecture. Hirnrinde, 18, 1973–1980.
Fischl, B., Salat, D. H., Albert, M., Dieterich, M., Haselgrove, C.,
van der Kouwe, A., et al. (2002). Whole brain segmentation:
Automated labeling of neuroanatomical structures in the
human brain. Neuron, 33, 341–355.
Fischl, B., Sereno, M. ICH., & Dale, A. M. (1999). Cortical surface-
based analysis. II: Inflation, flattening, and a surface-based
coordinate system. Neurobild, 9, 195–207.
Fischl, B., Sereno, M. ICH., Tootell, R. B. H., & Dale, A. M. (1999).
High-resolution intersubject averaging and a coordinate
system for the cortical surface. Kartierung des menschlichen Gehirns, 8,
272–284.
Fischl, B., van der Kouwe, A., Destrieux, C., Halgren, E., Segonne,
F., Salat, D. H., et al. (2004). Automatically parcellating the
human cerebral cortex. Hirnrinde, 14, 11–22.
Flynn, F. G., Benson, D. F., & Ardila, A. (1999). Anatomy of the
insula—Functional and clinical correlates. Aphasiology, 13,
55–77.
Foundas, A. L., Leonard, C. M., Gilmore, R., Fennell, E., &
Heilman, K. M. (1994). Planum temporale asymmetry and
language dominance. Neuropsychologie, 32, 1225–1231.
Frye, R. E., Liederman, J., Malmberg, B., McLean, J., Strickland,
D., & Beauchamp, M. S. (2010). Surface area accounts for
the relation of gray matter volume to reading-related skills
and history of dyslexia. Hirnrinde, 20, 2625–2635.
Geschwind, N., & Levitsky, W. (1968). Human brain: Left-right
asymmetries in temporal speech region. Wissenschaft, 161,
186–187.
Hagler, D. J., Jr., Saygin, A. P., & Sereno, M. ICH. (2006).
Smoothing and cluster thresholding for cortical surface-based
group analysis of fMRI data. Neurobild, 33, 1093–1103.
Heim, S., Eickhoff, S. B., & Amunts, K. (2008). Specialisation in
Brocaʼs region for semantic, phonological, and syntactic
fluency? Neurobild, 40, 1362–1368.
Hickok, G., & Kacke, D. (2007). The cortical organization
of speech processing. Nature Reviews Neurowissenschaften, 8,
393–402.
Hunter, Z. R., & Brysbaert, M. (2008). Visual half-field
experiments are a good measure of cerebral language
dominance if used properly: Evidence from fMRI.
Neuropsychologie, 46, 316–325.
Josse, G., Kherif, F., Flandin, G., Seghier, M. L., & Price, C. J.
(2009). Predicting language lateralization from gray matter.
Zeitschrift für Neurowissenschaften, 29, 13516–13523.
Josse, G., Mazoyer, B., Crivello, F., & Tzourio-Mazoyer, N.
(2003). Left planum temporale: An anatomical marker of left
hemispheric specialization for language comprehension.
Gehirnforschung, Kognitive Gehirnforschung, 18, 1–14.
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). Der
fusiform face area: A module in human extrastriate cortex
specialized for face perception. Zeitschrift für Neurowissenschaften,
17, 4302–4311.
Keller, S. S., Highley, J. R., Garcia-Finana, M., Sluming, V.,
Rezaie, R., & Roberts, N. (2007). Sulcal variability, stereological
measurement and asymmetry of Brocaʼs area on MR images.
Journal of Anatomy, 211, 534–555.
Keller, S. S., Roberts, N., Garcia-Finana, M., Mohammadi, S.,
Ringelstein, E. B., Knecht, S., et al. (2011). Can the language-
dominant hemisphere be predicted by brain anatomy?
Zeitschrift für kognitive Neurowissenschaften, 23, 2013–2029.
Knecht, S., Drager, B., Deppe, M., Bobe, L., Lohmann, H.,
Floel, A., et al. (2000). Handedness and hemispheric
language dominance in healthy humans. Gehirn, 123,
2512–2518.
Kosslyn, S. M. (1987). Seeing and imagining in the cerebral
hemispheres: A computational approach. Psychological
Rezension, 94, 148–175.
Loring, D. W., Meador, K. J., Lee, G. P., Murro, A. M.,
Schmied, J. R., Flanigin, H. F., et al. (1990). Cerebral language
lateralization: Evidence from intracarotid amobarbital testing.
Neuropsychologie, 28, 831–838.
Luders, E., Gaser, C., Jancke, L., & Schlaug, G. (2004). A voxel-
based approach to gray matter asymmetries. Neurobild,
22, 656–664.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsychologie,
9, 97–113.
Palaniyappan, L., & Liddle, P. F. (2012). Differential effects of
surface area, gyrification and cortical thickness on voxel
based morphometric deficits in schizophrenia. Neurobild,
60, 693–699.
Panizzon, M. S., Fennema-Notestine, C., Eyler, L. T., Jernigan, T. L.,
Prom-Wormley, E., Neale, M., et al. (2009). Distinct genetic
Greve et al.
1491
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
influences on cortical surface area and cortical thickness.
Hirnrinde, 19, 2728–2735.
Penhune, V. B., Cismaru, R., Dorsaint-Pierre, R., Petitto, L. A., &
Zatorre, R. J. (2003). The morphometry of auditory cortex in
the congenitally deaf measured using MRI. Neurobild, 20,
1215–1225.
Penhune, V. B., Zatorre, R. J., MacDonald, J. D., & Evans, A. C.
(1996). Interhemispheric anatomical differences in human
primary auditory cortex: Probabilistic mapping and volume
measurement from magnetic resonance scans. Zerebral
Kortex, 6, 661–672.
Plaut, D. C., & Behrmann, M. (2011). Complementary neural
representations for faces and words: A computational
exploration. Cognitive Neuropsychology, 28, 251–275.
Postelnicu, G., Zollei, L., & Fischl, B. (2009). Combined
volumetric and surface registration. IEEE Transactions on
Medical Imaging, 28, 508–522.
Price, C. J. (2012). A review and synthesis of the first 20 Jahre
of PET and fMRI studies of heard speech, spoken language
and reading. Neurobild, 62, 816–847.
Price, C. J. (2000). The anatomy of language: Contributions
from functional neuroimaging. Journal of Anatomy, 197,
335–359.
Price, C. J., & Devlin, J. T. (2011). The interactive account of
ventral occipitotemporal contributions to reading. Trends in
Cognitive Sciences, 15, 246–253.
bodies, and objects. Zeitschrift für kognitive Neurowissenschaften, 23,
4122–4137.
Toga, A. W., & Thompson, P. M. (2003). Mapping brain
asymmetry. Nature Reviews Neurowissenschaften, 4, 37–48.
Tucholka, A., Fritsch, V., Polina, J. B., & Thirion, B. (2012).
An empirical comparison of surface-based and volume-based
group studies in neuroimaging. Neurobild, 63, 1443–1453.
Turkeltaub, P. E., & Coslett, H. B. (2010). Localization of
sublexical speech perception components. Brain and
Language, 114, 1-15.
Tzourio, N., Nkanga-Ngila, B., & Mazoyer, B. (1998). Left planum
temporale surface correlates with functional dominance
during story listening. NeuroReport, 9, 829–833.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Neurobild, 15, 273–289.
Van der Haegen, L., Cai, Q., & Brysbaert, M. (2012).
Colateralization of Brocaʼs area and the visual word form area
in left-handers: fMRI evidence. Brain and Language, 122,
171–178.
Van der Haegen, L., Cai, Q., Seurinck, R., & Brysbaert, M. (2011).
Further fMRI validation of the visual half field technique as an
indicator of language laterality: A large-group analysis.
Neuropsychologie, 49, 2879–2888.
Pujol, J., Deus, J., Losilla, J. M., & Capdevila, A. (1999). Zerebral
Van der Haegen, L., Westerhausen, R., Hugdahl, K., & Brysbaert,
lateralization of language in normal left-handed people
studied by functional MRI. Neurologie, 52, 1038–1043.
Rademacher, J., Caviness, V. S., Jr., Steinmetz, H., & Galaburda,
A. M. (1993). Topographical variation of the human primary
cortices: Implications for neuroimaging, brain mapping, Und
neurobiology. Hirnrinde, 3, 313–329.
Raschle, N. M., Chang, M., & Gaab, N. (2011). Structural brain
alterations associated with dyslexia predate reading onset.
Neurobild, 57, 742–749.
Richardson, F. M., Ramsden, S., Ellis, C., Burnett, S., Megnin, O.,
Catmur, C., et al. (2011). Auditory short-term memory
capacity correlates with gray matter density in the left
posterior STS in cognitively normal and dyslexic adults.
Zeitschrift für kognitive Neurowissenschaften, 23, 3746–3756.
Risse, G. L., Gates, J. R., & Fangman, M. C. (1997). A
reconsideration of bilateral language representation based on
the intracarotid amobarbital procedure. Brain and Cognition,
33, 118–132.
Seghier, M. L., & Price, C. J. (2011). Explaining left lateralization
for words in the ventral occipitotemporal cortex. Zeitschrift für
Neurowissenschaften, 31, 14745–14753.
Segonne, F., Dale, A. M., Busa, E., Glessner, M., Salat, D., Hahn,
H. K., et al. (2004). A hybrid approach to the skull stripping
problem in MRI. Neurobild, 22, 1060–1075.
Spreer, J., Arnold, S., Quiske, A., Wohlfarth, R., Ziyeh, S.,
Altenmuller, D., et al. (2002). Determination of hemisphere
dominance for language: Comparison of frontal and temporal
fMRI activation with intracarotid amytal testing.
Neuroradiology, 44, 467–474.
Springer, J. A., Binder, J. R., Hammeke, T. A., Swanson, S. J.,
Frost, J. A., Bellgowan, P. S., et al. (1999). Language
dominance in neurologically normal and epilepsy subjects: A
functional MRI study. Gehirn, 122, 2033–2046.
Taylor, J. C., & Downing, P. E. (2011). Division of labor between
lateral and ventral extrastriate representations of faces,
M. (2013). Speech dominance is a better predictor of
functional brain asymmetry than handedness: A combined
fMRI word generation and behavioral dichotic listening study.
Neuropsychologie, 51, 91–97.
Vingerhoets, G., Alderweireldt, A. S., Vandemaele, P., Cai, Q.,
Van der Haegen, L., Brysbaert, M., et al. (2013). Praxis and
language are linked: Evidence from co-lateralization in
individuals with atypical language dominance. Kortex, 49,
172–183.
Wada, J., & Rasmussen, T. (1960). Intracarotid-injection of
sodium amytal for the lateralization of cerebral speech
dominance, experimental and clinical observations. Zeitschrift
of Neurosurgery, 17, 266–282.
Watkins, K. E., Paus, T., Lerch, J. P., Seidenwald, A., Collins, D. L.,
Neelin, P., et al. (2001). Structural asymmetries in the human
Gehirn: A voxel-based statistical analysis of 142 MRI scans.
Hirnrinde, 11, 868–877.
Willems, R. M., Peeling, M. V., & Hagoort, P. (2010). Zerebral
lateralization of face-selective and body-selective visual areas
depends on handedness. Hirnrinde, 20, 1719–1725.
Winkler, A. M., Kochunov, P., Blangero, J., Almasy, L., Zilles, K.,
Fuchs, P. T., et al. (2010). Cortical thickness or gray matter
Volumen? The importance of selecting the phenotype for
imaging genetics studies. Neurobild, 53, 1135–1146.
Winkler, A. M., Sabuncu, M. R., Yeo, B. T., Fischl, B., Greve,
D. N., Kochunov, P., et al. (2012). Measuring and comparing
brain cortical surface area and other areal quantities.
Neurobild, 61, 1428–1443.
Ziegler, J., & Ferrand, L. (1988). Orthography shapes the
perception of speech: The consistency effect in auditory
word recognition. Psychonomic Bulletin & Rezension, 5,
683–689.
Zollei, L., Stevens, A., Huber, K., Kakunoori, S., & Fischl, B.
(2010). Improved tractography alignment using combined
volumetric and surface registration. Neurobild, 51, 206–213.
1492
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
4
9
7
/
7
1
1
4
9
7
4
7
5
/
5
1
6
7
0
7
Ö
9
C
4
N
2
_
5
A
/
_
J
0
Ö
0
C
4
N
0
5
_
A
P
_
D
0
0
B
4
j
0
G
5
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
T
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1