Cross-modal Emotional Attention: Emotional Voices
Modulate Early Stages of Visual Processing
Tobias Brosch, Didier Grandjean, David Sander, and Klaus R. Scherer
Abstracto
& Emotional attention, the boosting of the processing of
emotionally relevant stimuli, tiene, up to now, mainly been in-
vestigated within a sensory modality, por ejemplo, mediante el uso
emotional pictures to modulate visual attention. In real-life
entornos, sin embargo, humans typically encounter simulta-
neous input to several different senses, such as vision and
audition. As multiple signals entering different channels might
originate from a common, emotionally relevant source, el
prioritization of emotional stimuli should be able to oper-
ate across modalities. en este estudio, we explored cross-modal
emotional attention. Spatially localized utterances with emo-
tional and neutral prosody served as cues for a visually pre-
sented target in a cross-modal dot-probe task. Participantes
were faster to respond to targets that appeared at the spatial
location of emotional compared to neutral prosody. Event-
related brain potentials revealed emotional modulation of
early visual target processing at the level of the P1 component,
with neural sources in the striate visual cortex being more
active for targets that appeared at the spatial location of emo-
tional compared to neutral prosody. These effects were not
found using synthesized control sounds matched for mean
fundamental frequency and amplitude envelope. Estos resultados
show that emotional attention can operate across sensory
modalities by boosting early sensory stages of processing,
thus facilitating the multimodal assessment of emotionally rel-
evant stimuli in the environment. &
INTRODUCCIÓN
The human organism is constantly confronted with a
huge amount of stimulus input from the environment.
Due to limited capacity (Marois & Ivanoff, 2005), el
brain cannot exhaustively process all the input and has
to select some stimuli at the cost of others (Desimone
& Duncan, 1995). In addition to basic physical features
such as color or size (lobo & Horowitz, 2004), emo-
tional relevance is an important dimension which can
modulate this process. Emotional stimuli are privileged
in the competition for neural processing resources.
Brain activation elicited by emotional stimuli (como
pictures, palabras, or sounds) is higher than for neutral
estímulos, ref lecting a more robust and stable neural
representación ( Vuilleumier, 2005; Davidson, Maxwell,
& Shackman, 2004). A number of brain imaging stud-
ies have shown that detection and preferential process-
ing of emotional stimuli occurs even when they are not
initially in the focus of attention (Pourtois, Schwartz,
Seghier, Lazeyras, & Vuilleumier, 2006; Grandjean et al.,
2005; Vuilleumier, Armony, Conductor, & Dolan, 2001). El
amygdala, a neural structure in the medial-temporal
lobe with extensive connections to many other brain
regiones (El dulce, 2000), is crucially involved in the pref-
University of Geneva, Suiza
erential processing of emotional stimuli. Por ejemplo,
amygdala activity is correlated with enhanced responses
to emotional stimuli in the visual cortex (Morris et al.,
1998). Además, amygdala lesions can abolish the
enhanced activation for emotional compared to neutral
faces in the visual cortex ( Vuilleumier, Richardson,
Armony, Conductor, & Dolan, 2004). De este modo, it has been sug-
gested that increased perceptual processing of emo-
tional stimuli results from direct feedback signals from
the amygdala to cortical sensory pathways ( Vuilleumier,
2005).
The preferential treatment of emotional stimuli
es
reflected in participants’ behavior in several cognitive
paradigms, such as the visual search task (Brosch &
sharma, 2005; O¨ hman, Flykt, & Esteves, 2001), el
attentional blink paradigm (anderson, 2005), the atten-
tional cueing paradigm (Fox, ruso, & Dutton, 2002),
and the dot-probe task (Brosch, Sander, & Scherer,
2007; Lipp & Derakshan, 2005; Mogg & Bradley, 1999).
In the dot-probe task (ver figura 1), participants re-
spond to the location or identity of a target, cual
replaces one out of two simultaneously presented cues.
One of the cues is emotional, the other one is neu-
tral. Behavioral results in the dot-probe task show
facilitated processing when the target replaces the
emotional cue compared to the neutral cue, reflected
by faster response times toward the targets (Brosch
et al., 2007; Lipp & Derakshan, 2005). This is interpreted
D 2008 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 21:9, páginas. 1670–1679
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
1
yo
9
mi
–
1
pag
6
d
7
F
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
oh
7
C
0
norte
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
1
1
0
0
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
F
.
t
.
/
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Cifra 1. Experimental
secuencia.
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
1
yo
9
mi
–
1
pag
6
d
7
F
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
oh
7
C
0
norte
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
1
1
0
0
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
F
.
.
/
t
.
oh
norte
1
8
METRO
a
y
2
0
2
1
as the result of attentional capture by the emotional
stimulus, which then leads to increased processing of
the target.
Event-related potentials (ERPs) recorded during the
emotional dot-probe task reveal an augmentation of the
P1 component elicited by a target replacing the emotional
compared to the neutral cue (Brosch, Sander, Pourtois, &
Scherer, 2008; Pourtois, Grandjean, Sander, & Vuilleumier,
2004). Earlier ERP results have indicated that the P1 ex-
ogenous visual response is systematically enhanced in
amplitude in response to attended relative to unattended
spatial locations or stimuli (p.ej., Luck, Woodman, & Vogel,
2000). Amplitude modulations of the P1 as a function of
the deployment of visuospatial attention is thought to re-
flect a sensory gain control mechanism causing increased
perceptual processing in the visual cortex of attended
locations or stimuli (Hillyard, Vogel, & Luck, 1998). El
faster response times for targets replacing emotional cues
in the dot-probe paradigm are thus associated with mod-
ulations of early perceptual processing of the target (y
not due to postperceptual processes at the level of re-
sponse selection or action preparation).
Enhanced sensory representations of emotional stim-
uli have been found not only for the visual ( Vuilleumier
et al., 2001; Morris et al., 1998) but also for the auditory
domain. Several fMRI studies have shown that emotional
prosody increases activity in the associative auditory cor-
tex (superior temporal sulcus), more particularly in the
sensitive voice regions (Belin, Zatorre, Lafaille, Ahad, &
Pike, 2000). This effect was observed for positive and
negative emotions (Ethofer et al., 2006), and emerged
even when the focus of voluntary attention was directed
away from the emotional auditory stimuli using a dich-
otic listening task (Grandjean et al., 2005; Sander et al.,
2005). Además, stroke patients with left auditory
extinction showed a detection increase of emotional
compared to neutral prosody stimulation on the left
lado, showing that emotion is able to moderate an
auditory extinction phenomenon (Grandjean, Sander,
lucas, Scherer, & Vuilleumier, 2008), as previous studies
have already shown for the visual domain ( Vuilleumier
& Schwartz, 2001).
Until now, studies investigating emotional modula-
tion of spatial attention have mainly examined within-
modality effects, most frequently using pictures of
emotional stimuli to modulate visual attention. Cómo-
alguna vez, humans typically encounter simultaneous input to
several different senses, such as vision and audition.
Signals entering these different channels might originate
from a common source, requiring mechanisms for the
integration of information (including emotional
infor-
formación) conveyed by multiple sensory channels. To re-
ceive maximal benefit from multimodal input, the brain
must coordinate and integrate the input appropriately
so that signals from a relevant common source are pro-
cessed across the different input channels. This inte-
gration is a computational challenge, as the properties
of the information representation differ greatly between
the input channels (Conductor & Spence, 1998).
The questions to which extent attention operates in-
dependently within each sensory modality and by which
mechanisms attention is coordinated across modalities
Brosch et al.
1671
have been investigated using simple nonemotional stim-
uli such as flashes of light or bursts of noise (Eimer
& Conductor, 2001; Conductor & Spence, 1998). The paradigm
most frequently used for the investigation of cross-modal
attentional modulation is the spatial cueing paradigm
(posner, 1980). In this paradigm, participants indicate
whether a target appeared either in the left or the right
visual field. Before the target, a spatially nonpredictive
peripheral cue in another modality is presented (p.ej., un
auditory cue preceding a visual target). Although the cue
is not predictive of the location of the target, respuestas
to the targets are faster and/or more accurate when the
targets are presented on the same side as the cue
(McDonald & Ward, 2000; Spence & Conductor, 1997).
Like for its unimodal counterpart, ERP recordings have
been used to examine the neural correlates of the cross-
modal attentional modulation effect (Eimer & Conductor,
2001; McDonald & Ward, 2000). In an ERP study of ex-
ogenous attentional cueing using auditory cues and visual
objetivos, an attentional negativity (Nd) was elicited for vi-
sual ERPs recorded from lateral occipital sites (PO7/PO8)
entre 200 y 400 msec after stimulus onset for valid
compared to invalid trials (McDonald & Ward, 2000).
No cueing effects were observed for the P1 component.
This suggests that cross-modal effects of a nonemo-
tional auditory event on visual processes may be located
at a stage after the initial perceptual processing of visual
información.
Not much is known about the modulatory effect of
emotional stimuli on attention across modalities. Auto-
matic enhanced sensory responses of specific brain
areas to emotional events have been shown both for
visual (Vuilleumier et al., 2001) and auditory (Grandjean
et al., 2005; Sander et al., 2005) events. This probably
reflects a fundamental principle of human brain orga-
nization, namely to prioritize the processing of emo-
tionally relevant stimuli, even if they are outside the
focus of attention. Such a mechanism should be able
to operate across modalities, as multiple signals enter-
ing different channels might originate from a com-
mon, emotionally relevant source. Consistente con esto
vista, we recently showed that emotional prosody, el
changes in the tone of the voice that convey information
about a speaker’s emotional state (Scherer, Johnstone,
& Klasmeyer, 2003), can facilitate detection of a visual
objetivo (Brosch, Grandjean, Sander, & Scherer, 2008). En
this cross-modal emotional dot-probe paradigm (ver
MacLeod, Mathews, & Tata, 1986), participants indicated
the location of a visual target that was preceded by a
binaurally presented pair of auditory pseudowords, uno
of which was uttered with anger prosody (in one ear),
the other one with neutral prosody (in the other ear).
Although delivered through headphones, the emotional
and neutral auditory stimuli were spatialized to produce
the compelling illusion that they originated from a dis-
tinctive source localized either in the left or right peri-
personal space (see Methods for details). Response
times toward (nonemotional) visual targets were shorter
when they appeared in a position spatially congruent with
the perceived source of the emotional prosody (Brosch,
Grandjean, et al., 2008).
The aim of the present study was to investigate the
neural underpinnings of cross-modal modulation of vi-
sual attention by emotional prosody. Of special interest
was the question of whether cross-modal emotional at-
tention affects early sensory stages of processing—as
might be expected on the basis of investigations of emo-
tional attention within one modality (Brosch, Sander,
et al., 2008; Pourtois et al., 2004), or not—as might be
expected on the basis of investigations of nonemotional
cross-modal attention modulation (McDonald & Ward,
2000).
We recorded ERPs while participants performed the
cross-modal emotional dot-probe task (Brosch, Grandjean,
et al., 2008). Based upon earlier work investigating the
modulation of visual attention by visual emotional stim-
uli (Brosch, Sander, et al., 2008; Pourtois et al., 2004), nosotros
predicted that a cross-modal emotional modulation of
early sensory states would manifest as a modulation of
the amplitude of the P1 component in form of larger
amplitudes toward validly cued targets (ver figura 1)
than toward invalidly cued targets.
MÉTODOS
Participantes
Seventeen students of the University of Geneva par-
ticipated in the experiment. Data from two female
participants were excluded due to poor quality of the
physiological recording, leaving a final sample of 15 par-
ticipants (13 women, edad media = 21.4 años, DE = 3.3).
All participants were right-handed, had normal self-
reported audition and normal or corrected-to-normal
visión, and had no history of psychiatric or neurological
enfermedad.
Estímulos
The auditory stimuli consisted of meaningless but
word-like utterances (pseudowords ‘‘goster,’’ ‘‘niuvenci,''
‘‘figotleich’’) pronounced with either anger or neutral
prosody. Sixty different utterances by 10 different speak-
ers with a duration of 750 mseg (50% male speakers,
50% anger prosody) were extracted from a database of
pseudosentences that had been acquired and validated
in earlier work (Banse & Scherer, 1996). The anger stim-
uli were directly adopted from the database, the neutral
stimuli were selected from the ‘‘boredom’’ and ‘‘inter-
est’’ stimuli, selecting the most neutral on the basis of
a judgment study investigating the ‘‘neutrality’’ and ‘‘emo-
tionality’’ of these stimuli. Fifteen participants (9 women,
edad media = 25.3 años) judged the stimuli on two visual
analog rating scales (‘‘neutral’’ and ‘‘emotional’’). Basado
1672
Revista de neurociencia cognitiva
Volumen 21, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
1
yo
9
mi
–
1
pag
6
d
7
F
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
oh
7
C
0
norte
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
1
1
0
0
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
.
.
/
t
F
oh
norte
1
8
METRO
a
y
2
0
2
1
on those ratings, el 20 ‘‘interest’’ and 20 ‘‘boredom’’
‘‘emotional’’ ratings and maximal
stimuli with minimal
‘‘neutral’’ ratings were selected. Además, we per-
formed a judgment study on the excerpts selected for
the present experiment (anger, neutral) as well as emo-
tional prosody excerpts not used in the current study
(sadness, happiness, and fear). This was done to test the
recognizability of the different emotional stimuli and to
be sure that the neutral stimuli are perceived as ‘‘neutral’’
rather than ‘‘interest’’ or ‘‘boredom.’’
Sixteen participants (undergraduate students, 14 wom-
en) judged on visual analog scales (from ‘‘not at all’’ to
‘‘totally’’) to what extent the excerpts were pronounced
with anger, neutral, boredom, interés, despair, elation,
pride, disgust, desprecio, happiness, sadness, miedo, y
surprise emotional intonation. A test of repeated mea-
sures ANOVA using the within-subjects factors emotional
prosody and emotion scale revealed, as predicted, un
interaction effect [F(48, 912) = 75.78, pag < .001]. Anger
stimuli were mainly rated as expressing ‘‘anger’’ [con-
trast ‘‘anger’’ scale vs. other scales: F(1, 19) = 459.46,
p < .001] and neutral stimuli were mainly rated as ‘‘neu-
tral’’ [contrast ‘‘neutral’’ scale vs. other scales: F(1, 19) =
87.88, p < .001]. A contrast comparing the ‘‘neutral,’’
‘‘boredom,’’ and ‘‘interest’’ ratings for the neutral
stimuli showed that the neutral stimuli were rated sig-
nificantly higher on the ‘‘neutral’’ scale than on the
‘‘boredom’’ or ‘‘interest’’ scale [contrast neutral vs.
boring–interest: F(1, 19) = 52.94, p < .01]. All stimuli
were combined to 40 stereophonically presented paired
utterances containing one angry and one neutral ut-
terance. To avoid interactions of speaker sex and emo-
tionality in stimulus pairs, only utterance pairs from
same-sex speakers were combined. Each pair was matched
for mean acoustic energy.
The fundamental frequency F0 and the distribution
of energy in time play an important role in conveying
emotional information in voices (Grandjean, Ba¨nziger,
& Scherer, 2006; Banse & Scherer, 1996). In addition
to these low-level stimulus properties, emotional infor-
mation in prosody is conveyed by other, more complex
perceived acoustical characteristics corresponding to
objective acoustical parameters, such as spectral energy
distribution in time or the temporal dynamic of the
F0 (see e.g., Banse & Scherer, 1996). The complex in-
teractions of these different acoustical parameters over
time are crucial for emotional prosody perception. To
for the low-level physical properties of our
control
stimuli related to prosody, we included a control con-
dition by synthesizing control stimuli matched for the
mean fundamental frequency and the amplitude en-
velope of each vocal stimulus used in the experiment
using Praat. After controlling for the low-level stimulus
properties, any effect reflecting voice-specific processes
that is not driven by a particular range of frequency or
a specific amplitude contour should only be found for
the prosody cues, not for the control cues.
In order to give the subjective impression that the
sounds originate from a specific location in space, we
manipulated the interaural time difference (ITD) of the
sounds using a head-related transfer function (HRTF)
implemented in the plug-in Panorama used with Sound-
Forge (for more details about this procedure, see e.g.,
Spierer, Meuli, & Clarke, 2007). The audio pairs were
transformed via binaural synthesis to be equivalent to
sound sources at a distance of 110 cm and at an angle
of 248 to the left and to the right of the participants (see
Figure 1). We used spatially localized stimuli instead of
the simpler dichotic presentation mode, as it is a closer
approximation of real-life contexts in which concomi-
tant auditory and visual information can originate from
a common source localized in space. The HRTF method
enables us to investigate the relationship between emo-
tion and spatial attention processes based on realistic
spatial localization rather than investigating ear effects.
Previous studies with brain-damaged patients have
shown a double dissociation between auditory extinction
and ear extinction, highlighting the fact that these two
processes are very different in terms of the brain regions
involved (Spierer et al., 2007).
The experiment was controlled by E-Prime. The audi-
tory cues were presented using Sony MDR-EX71 head-
phones. The visual targets were presented using a Sony
VPL CX 10 projector.
Procedure
Figure 1 shows the experimental sequence. During the
whole experiment, a fixation cross was presented. Each
trial started with a random time interval between 500
and 1000 msec, after which the acoustic cue sound pair
was presented. One of the sounds in the pair had emo-
tional prosody, the other one neutral prosody.
The target, a neutral geometric figure (a triangle which
could either point upward or downward), was presented
with a variable cue–target stimulus onset asynchrony
(SOA) of 550, 600, 650, 700, or 750 msec after sound
onset. The target was presented for 100 msec on the left
or right side, at a distance of 45 cm from the fixation
cross. The participants were seated at 100 cm from the
projection screen. Thus, the angle between the target
and the fixation cross was 248, which is equivalent to
the synthesized location of the audio stimulus pairs. In a
valid trial, the target appeared on the side of the emo-
tional sound, whereas in an invalid trial, the target
appeared on the side of the neutral sound. Valid and
invalid trials were presented in randomized order with
an equal proportion of valid and invalid trials (50%).
Participants were instructed to press the ‘‘B’’ key of the
response keyboard using the index finger of their right
hand only when the orientation of the triangle corre-
sponded to their respective GO condition (triangle point-
ing upward or downward, counterbalanced across
participants). Participants had a maximum of 1500 msec
Brosch et al.
1673
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
9
e
-
1
p
6
d
7
f
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
o
7
c
0
n
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
1
0
0
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
f
.
.
/
o
n
1
8
M
a
y
2
0
2
1
to respond, after that time, the next trial started. The
experiment consisted of one practice block of 10 trials,
followed by four experimental blocks of 160 trials each
(total 640 trials). In two blocks, sounds with emotional
and neutral prosody were presented, and in two blocks,
the synthesized control sounds were presented. We de-
signed a small number of go trials which required a motor
response (10%) to study covert spatial orienting toward
emotional stimuli in a vast majority of trials where there
is no overt motor response (90% no-go trials), therefore
minimizing the contamination of motor preparation or
execution on EEG signal quality.
EEG Recordings
EEG was recorded with a sampling rate of 512 Hz
using the ActiveTwo system (BioSemi, Amsterdam,
Netherlands). Horizontal and vertical EOGs were re-
corded using four facial electrodes placed on the outer
canthi of the eyes and in the inferior and superior areas
of the left orbit. Scalp EEG was recorded from 64 Ag/
AgCl electrodes attached to an electrode cap and posi-
tioned according to the extended 10–20 EEG system.
The EEG electrodes were referenced off-line to average
reference. The data were filtered using a high pass of
0.53 Hz and a low pass of 30 Hz. Data were downsam-
pled to 256 Hz and segmented around target onsets in
epochs of 1000 msec (from (cid:1)200 msec to +800 msec).
A reduction of artifacts related to vertical eye move-
ments was implemented using the algorithm developed
by Gratton, Coles, and Donchin (1983). A baseline
correction was performed on the prestimulus interval
using the first 200 msec. EEG epochs exceeding 70 AV
were excluded from the analysis. The artifact-free epochs
were averaged separately for each electrode, condition,
and individual. Grand-average ERPs were finally gener-
ated by computing the mean ERPs across participants in
each condition.
Data Analysis
Behavioral Data
Response times for correct responses between 200 and
1000 msec were analyzed in a 2 (cid:2) 2 (cid:2) 2 repeated mea-
sures ANOVA with the factors voice condition (prosody/
synthesized control sounds), cue validity (valid/invalid)
and target position (left/right).
EEG Experiment
Based on our a priori hypotheses and on inspection of
the present ERP dataset, we analyzed the P1 component
(130–190 msec) time-locked to the onset of the target
in valid and invalid trials. Peak amplitudes and latencies
were measured at lateral occipital sites (PO7/O1 and PO8/
O2; see Figure 3). These sites were selected on the basis
of related effects in previous studies (Brosch, Sander,
et al., 2008; Pourtois et al., 2004; Martinez et al., 1999) and
on conspicuous topographic properties of the present
ERP dataset. The amplitudes and latencies of the P1 were
analyzed using 2 (cid:2) 2 (cid:2) 2 (cid:2) 2 (cid:2) 2 ANOVAs with the
repeated factors voice condition (prosody/synthesized
control sounds), cue validity (valid/invalid), target posi-
tion (left/right), hemisphere (left/right), and electrode
position (PO/O). To estimate the likely configuration
of intracranial neural sources underlying the observed
scalp topographic maps of interest, we used a distributed
inverse solution method on the basis of a Local Auto-
Regressive Average model of the unknown current den-
sity of the brain (LAURA; see Grave de Peralta Menendez,
Gonzalez Andino, Lantz, Michel, & Landis, 2001). The
method is derived from biophysical laws describing elec-
tric fields in the brain. It computes a three-dimensional
reconstruction of the generators of the brain’s electro-
magnetic activity measured at the scalp on the basis of
biophysically driven inverse solutions without a priori
assumptions on the number and position of the possi-
ble generators (see also Michel et al., 2004, for further
details).
RESULTS
Behavioral Data
Figure 2 shows the response times for valid and invalid
trials in the prosody condition and the control condition.
There was a trend toward a Voice condition (cid:2) Cue
validity interaction [F(1, 14) = 2.51, p = .14]. In the pros-
ody condition, participants responded faster toward valid
(549 msec) than toward invalid (565 msec) targets, as in-
dicated by a marginally significant t test [t(14) = 1.68, p =
.06, one-tailed], thus replicating our previous behavioral
findings (Brosch, Grandjean, et al., 2008). Note that in
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
9
e
-
1
p
6
d
7
f
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
o
7
c
0
n
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
1
0
0
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
/
.
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Response times (msec) for the prosody condition and
the control condition. In the prosody condition, participants
responded faster toward valid than invalid targets ( p = .06).
No such facilitation was observed for the control condition.
1674
Journal of Cognitive Neuroscience
Volume 21, Number 9
contrast to Brosch, Grandjean, et al. (2008), in the pres-
ent study, participants responded only on 10% of the
trials, as we wanted to analyze brain activity for the 90%
of trials without contamination by motor responses. In the
control condition, no differences were found in response
times between valid (570 msec) and invalid (572 msec)
trials [t(14) = 0.4, ns]. The interaction Voice condition (cid:2)
Target position revealed longer response times toward
targets presented to the left visual hemifield (580 msec)
compared to the right visual field (562 msec) in the con-
trol condition [F(1, 14) = 7.36, p = .02, partial h2 = .35].
ERP Analysis and Source Localization
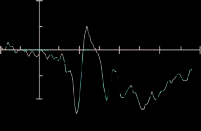
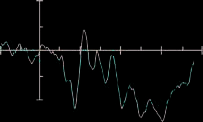
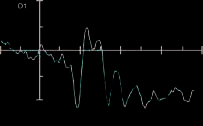
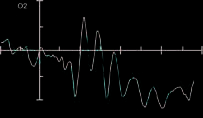
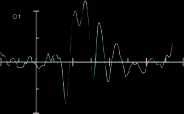
Figure 3 shows the ERPs time-locked to target onset for
targets presented to the left visual field and ERPs for the
valid and invalid conditions for the prosody condition at
electrodes PO7, PO8, O1, and O2.
P1 amplitude was larger in the prosody trials (3.0 AV)
than in the control trials (1.9 AV), as revealed by the
main effect of voice condition [F(1, 14) = 68.98, p < .001,
partial h2 = .83]. P1 for targets presented to the right
hemisphere peaked earlier (164 msec) than P1 for tar-
gets presented to the left hemisphere (171 msec), as
indicated by a main effect of target position [F(1, 14) =
15.14, p = .002, partial h2 = .52].
Most important for our hypotheses, the interaction
Voice condition (cid:2) Cue validity was statistically significant
[F(1, 14) = 5.78, p = .03, partial h2 = .29]. We thus
analyzed the data for the prosody condition and the
control condition separately with regards to the effects
of cue validity. In the prosody condition, amplitude of
the P1 was larger in valid (3.2 AV) than in invalid (2.8 AV)
trials as shown by a main effect of cue validity [F(1,
14) = 6.82, p = .021, partial h2 = .33]. This effect was
driven by targets presented to the left visual field (left
visual field invalid: 2.6 AV, left visual field valid: 3.3 AV,
right visual field invalid: 2.9 AV, right visual field valid:
3.0 AV), as indicated by the interaction Cue validity (cid:2)
Target position [F(1, 14) = 5.07, p = .041, partial h2 =
.27] and a follow-up t test comparing valid and invalid
targets presented to the left visual field [t(14) = 3.9,
Figure 3. Results from
the ERP analysis: (A) ERPs
time-locked to target onset
for targets presented to the
left visual field and ERPs for
the valid (red) and invalid
(black) conditions for
the prosody condition at
electrodes PO7, PO8, O1,
and O2. (B) ERPs at O1 and
O2 for the control condition.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
9
e
-
1
p
6
d
7
f
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
o
7
c
0
n
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
1
0
0
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
.
f
/
o
n
1
8
M
a
y
2
0
2
1
Brosch et al.
1675
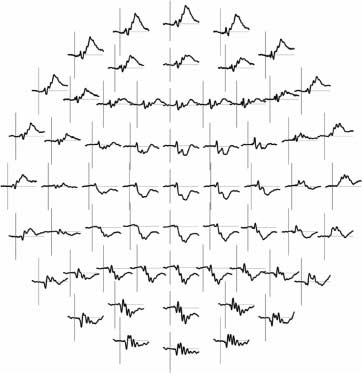
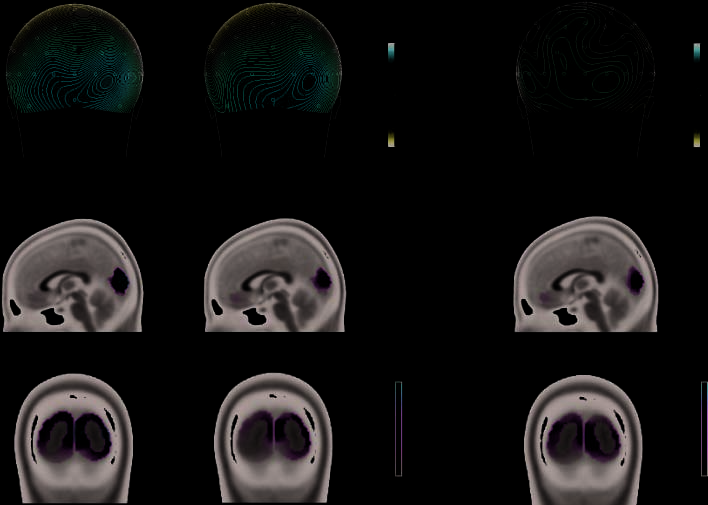
Figure 4. Top row:
Topographic maps for the
P1 in valid and invalid trials
and topographic difference
map. Middle and bottom
rows: Inverse solution based
on LAURA revealed the
intracranial generators of
the P1 in the striate and
extrastriate visual cortex.
Values of the inverse solution
for valid and invalid trials are
shown on a continuous scale
from 0 to 0.015 AA/mm3,
for the difference map on a
continuous scale from 0 to
0.01 AA/mm3. A region-of-
interest analysis taking into
account the inverse solution
points in the peak activation
in the visual cortex confirmed
stronger activation to valid
than to invalid targets.
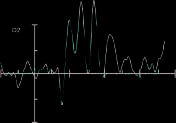
p = .001, one-tailed]. In the control condition, no effect
involving cue validity was significant (all p > .17, left visual
field invalid: 2.0 AV, left visual field valid: 2.1 AV, bien
visual field invalid: 1.8 AV, right visual field valid: 1.6 AV).
Finalmente, we applied an inverse solution on the basis of
LAURA to the peak of the P1 potential for valid and in-
valid trials in the prosody condition. Results confirmed
that the intracranial generators of the P1 were located in
the striate and extrastriate visual cortex (ver figura 4),
a pattern of brain regions which has been repeatedly
found when looking at the generators of this early vi-
sual response (Noesselt et al., 2002; Martinez et al., 1999).
A region-of-interest analysis, based on the inverse solu-
tion points in the peak activation in the visual cortex
(ver figura 4), confirmed stronger activation to valid
(0.015 AV) than to invalid (0.010 AV) objetivos [main effect
cue validity: F(1, 14) = 11.01, pag = .005, partial h2 = .44].
DISCUSIÓN
During this cross-modal emotional dot-probe task, nosotros
recorded scalp ERPs to investigate at what stage of stim-
ulus processing the deployment of visuospatial attention
toward simple nonemotional visual targets was affected
by spatially congruent or incongruent emotional infor-
mation conveyed in affective prosody. At the behavioral
nivel, participants were faster to respond to the orien-
tation of a visual target when it appeared at the spatial
location of a previously presented utterance with an-
ger prosody compared to neutral prosody. Este resultado es
consistent with our previous behavioral findings (Brosch,
Grandjean, et al., 2008), even though the effect in the
present study was only marginally significant ( pag = .06),
probably due to the lower number of GO trials requiring
a manual response. En tono rimbombante, this cross-modal emo-
tional effect was not present when using synthesized con-
trol stimuli matched for the mean fundamental frequency
and the amplitude envelope of each vocal stimulus used
in the experiment, ruling out the possibility that these
low-level acoustic parameters trigger the cross-modal
emotional effect.
Analysis of scalp ERPs revealed a selective modulation
of the P1 component toward visual targets preceded by
spatially congruent auditory cues conveying emotional
prosody, which was restricted to targets presented to
the left visual hemifield. P1 amplitude was higher when
the visual target appeared at the location of the source
of the anger compared to neutral prosody. This modu-
lation of the P1 as a function of the affective prosody
was not observed in the control condition. De este modo, this P1
effect consecutive to visual target processing most likely
depends upon the activation of voice-specific processes
(Grandjean et al., 2005; Belin et al., 2000) and cannot be
explained by the processing of a particular range of fre-
quency or a specific amplitude contour in the auditory
estímulos.
Here we show that the cross-modal modulation of
spatial attention triggered by emotional prosody affected
early sensory stages of visual processing. The observed
modulation by emotional prosody took place earlier than
the modulation observed with nonemotional auditory
1676
Revista de neurociencia cognitiva
Volumen 21, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
1
yo
9
mi
–
1
pag
6
d
7
F
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
oh
7
C
0
norte
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
1
1
0
0
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
/
.
.
t
F
oh
norte
1
8
METRO
a
y
2
0
2
1
cross-modal cues (McDonald & Ward, 2000), cual
emerged as an attentional negativity between 200 y
400 mseg. McDonald and Ward (2000) interpreted the
absence of a P1 modulation as suggesting that the cross-
modal effects of an auditory event on visual processes are
located after the initial sensory processing of visual in-
formación. In contrast to their finding, our results show
a modulation during initial stages of visual processing
caused here by emotional auditory cues. Two methodo-
logical differences between the study by McDonald and
Ward (2000) and our study should be discussed when
comparing the results. The former study used a modified
exogenous cueing paradigm, where only one auditory
cue was presented, whereas in our study, we presented
two cues simultaneously in a modified dot-probe para-
digm. Sin embargo, one would expect a more exhaustive
processing of the cue stimulus when it is presented with-
out direct competition for processing resources, not when
it has to compete with other stimuli. De este modo, it is unlikely
that this accounts for the differences in early perceptual
Procesando. A second methodological difference con-
cerns the SOA between the cue and the target: Whereas
McDonald and Ward (2000) used SOAs between 100
y 300 mseg, we used SOAs between 550 y 750 mseg.
Our choice of SOAs was motivated by the fact that pros-
ody is mainly due to temporal changes such as variations
in stress and pitch (muchacho, 1996), y por lo tanto, needs some
time to unfold.
Assuming that the different results are not due to
methodological differences between the studies, ellos
might reflect fundamental differences in the processing
of emotional and nonemotional stimuli. A system that
prioritizes orienting toward emotionally significant stim-
uli, operating across modalities, might produce a differ-
ent pattern of modulation and integration than a system
for the prioritization of perceptually salient stimuli. El
perception and evaluation of emotional stimuli involves
the activity of neural structures, especially the amygdala
( Vuilleumier, 2005; Sander, Grafman, & Zalla, 2003),
which are not involved in the cueing of attention to-
ward merely perceptively salient stimuli (Desimone &
Duncan, 1995). The amygdala plays a crucial role in
highlighting relevant events by providing both direct
and indirect top–down signals in sensory pathways
which modulate the representation of emotional events
( Vuilleumier, 2005). Affective prosody leads to increased
activation of the amygdala and the superior temporal
sulcus (Grandjean et al., 2005; Sander & Scheich, 2001).
Functional connections between the amygdala and the
visual cortex have been observed in animal tracer stud-
es (Freese & Amaral, 2005) and in humans using dif-
fusion tensor MRI (Catani, jones, Donato, & Ffytche,
2003). Además, increased activation of the visual cor-
tex when listening to emotional prosody (Sander et al.,
2005) or familiar voices (von Kriegstein, Kleinschmidt,
Sterzer, & Giraud, 2005) probably reflects a functional
coupling between auditory and visual cortices that can
facilitate the visual processing of targets ( Vuilleumier,
2005).
The behavioral effect as well as the modulation of
the P1 component observed in our study might reflect
a boosting of perceptual representation of the visual
stimulus in occipital brain areas, here triggered by a pre-
ceding affective voice. This conjecture is substantiated
by our source localization results, which clearly indicate
that the P1 modulation originated from generators lo-
calized in the visual cortex. Based on previous anatom-
ical evidence, we suggest that this enhanced occipital
activation for visual targets preceded by valid emotional
voice cues is probably driven by feedback connections
from the amygdala to the visual cortex, including the pri-
mary visual cortex (Freese & Amaral, 2005; Vuilleumier,
2005; Catani et al., 2003).
Emotional prosody is generally processed by both
hemispheres (Schirmer & Kotz, 2006; Van Lancker &
Sidtis, 1992). Some particularly relevant acoustical fea-
tures related to emotional prosody, sin embargo, seem to
involve the right hemisphere to a greater extent and
induce more stimulus-related processing in this hemi-
sphere (ross & Monnot, 2008), as shown by neuro-
imaging results ( Wildgruber, Ackermann, Kreifelts, &
Ethofer, 2006; Sander & Scheich, 2001) and behavioral
studies such as the dichotic listening task (Carmon &
Nachshon, 1973; Haggard & Parkinson, 1971). This later-
alization is in line with our findings, which indicated that
the modulation effect was mainly driven by targets pre-
sented to the left visual field, which are primarily pro-
cessed by the right hemisphere.
Further studies might investigate the effect of differ-
ent types of prosody (such as happy, surprised, or dis-
gusted) on attentional modulation. As no difference in
strength of amygdala activation is observed when com-
paring positive and negative prosody (Sander & Scheich,
2001), one would expect that our findings are not
restricted to anger prosody, but can be generalized to
different kinds of emotional prosody. We recently pre-
sented evidence for a similar generalization for the vi-
sual modality in form of rapid attentional modulation
toward several different kinds of emotionally relevant
estímulos (Brosch, Sander, et al., 2008).
To sum up, in this study we explored the effects of
cross-modal emotional attention. Both behavioral and
electrophysiological data converge on the central finding
that emotional attention can also operate across two
different sensory modalities by boosting early sensory
stages of processing.
Expresiones de gratitud
We thank Gilles Pourtois for valuable comments on a previous
draft of the article. This work was supported by the National
Centre of Competence in Research (NCCR) Affective Sciences,
financed by the Swiss National Science Foundation (No. 51NF40-
104897), and hosted by the University of Geneva.
Brosch et al.
1677
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
1
yo
9
mi
–
1
pag
6
d
7
F
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
oh
7
C
0
norte
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
1
1
0
0
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
t
.
F
.
/
oh
norte
1
8
METRO
a
y
2
0
2
1
Reprint requests should be sent to Tobias Brosch, Swiss Centre
for Affective Sciences, University of Geneva, 7, Rue des Battoirs,
1205 Geneva, Suiza, o por correo electrónico: Tobias.Brosch@unige.ch.
REFERENCIAS
anderson, A. k. (2005). Affective influences on the attentional
dynamics supporting awareness. Revista de experimentación
Psicología: General, 134, 258–281.
Banse, r., & Scherer, k. R. (1996). Acoustic profiles in vocal
emotion expression. Journal of Personality and Social
Psicología, 70, 614–636.
Belin, PAG., Zatorre, R. J., Lafaille, PAG., Ahad, PAG., & Pike, B.
(2000). Voice-selective areas in human auditory cortex.
Naturaleza, 403, 309–312.
Brosch, T., Grandjean, D., Sander, D., & Scherer, k. R. (2008).
Behold the voice of wrath: Cross-modal modulation of visual
attention by anger prosody. Cognición, 106, 1497–1503.
Brosch, T., Sander, D., Pourtois, GRAMO., & Scherer, k. R. (2008).
Beyond fear: Rapid spatial orienting towards positive
emotional stimuli. ciencia psicológica, 19, 362–370.
Brosch, T., Sander, D., & Scherer, k. R. (2007). That baby
caught my eye. . . Attention capture by infant faces.
Emoción, 7, 685–689.
Brosch, T., & sharma, D. (2005). The role of fear-relevant
stimuli in visual search: A comparison of phylogenetic
and ontogenetic stimuli. Emoción, 5, 360–364.
Carmon, A., & Nachshon, I. (1973). Ear asymmetry in
perception of emotional non-verbal stimuli. Acta
Psychologica, 37, 351–357.
Catani, METRO., jones, D. K., Donato, r., & Ffytche, D. h. (2003).
Occipito-temporal connections in the human brain.
Cerebro, 126, 2093–2107.
wrath: Brain responses to angry prosody in meaningless
speech. Neurociencia de la naturaleza, 8, 145–146.
graton, GRAMO., Coles, METRO. GRAMO., & Donchin, mi. (1983). A
new method for off-line removal of ocular artifact.
Electroencephalography and Clinical Neurophysiology,
55, 468–484.
Grave de Peralta Menendez, r., Gonzalez Andino, S., Lantz, GRAMO.,
Michel, C. METRO., & Landis, t. (2001). Noninvasive localization
of electromagnetic epileptic activity. I. Method descriptions
and simulations. Brain Topography, 14, 131–137.
Haggard, METRO. PAG., & Parkinson, A. METRO. (1971). Stimulus and
task factors as determinants of ear advantages. Quarterly
Revista de Psicología Experimental, 23, 168–177.
Hillyard, S. A., Vogel, mi. K., & Luck, S. j. (1998). Sensory
gain control (amplification) as a mechanism of selective
atención: Electrophysiological and neuroimaging
evidencia. Philosophical Transactions of the Royal
Society of London, Serie B, Ciencias Biologicas, 353,
1257–1270.
muchacho, D. R. (1996). Intonational phonology. Cambridge:
Prensa de la Universidad de Cambridge.
El dulce, j. mi. (2000). Emotion circuits in the brain. Annual
Review of Neuroscience, 23, 155–184.
Lipp, oh. v., & Derakshan, norte. (2005). Attentional bias to
pictures of fear-relevant animals in a dot probe task.
Emoción, 5, 365–369.
Luck, S. J., Woodman, GRAMO. F., & Vogel, mi. k. (2000).
Event-related potential studies of attention. Trends
in Cognitive Sciences, 4, 432–440.
MacLeod, C., Mathews, A., & Tata, PAG. (1986). Attentional bias
in emotional disorders. Journal of Abnormal Psychology,
95, 15–20.
Marois, r., & Ivanoff, j. (2005). Capacity limits of information
processing in the brain. Tendencias en Ciencias Cognitivas, 9,
296–305.
Davidson, R. J., Maxwell, j. S., & Shackman, A. j. (2004).
Martinez, A., Anllo-Vento, l., Sereno, METRO. I., Franco, l. r.,
The privileged status of emotion in the brain. Actas
of the National Academy of Sciences, EE.UU., 101,
11915–11916.
Desimone, r., & Duncan, j. (1995). Neural mechanisms of
selective visual attention. Revisión anual de neurociencia,
18, 193–222.
Conductor, J., & Spence, C. (1998). Crossmodal attention.
Opinión actual en neurobiología, 8, 245–253.
Eimer, METRO., & Conductor, j. (2001). Crossmodal links in
endogenous and exogenous spatial attention: Evidencia
from event-related brain potential studies. Neurociencia
and Biobehavioral Reviews, 25, 497–511.
Ethofer, T., Anders, S., Wiethoff, S., Erb, METRO., Herbert, C.,
Saur, r., et al. (2006). Effects of prosodic emotional
intensity on activation of associative auditory cortex.
NeuroReport, 17, 249–253.
Fox, MI., ruso, r., & Dutton, k. (2002). Attentional bias
for threat: Evidence for delayed disengagement from
emotional faces. Cognition and Emotion, 16, 355–379.
Freese, j. l., & Amaral, D. GRAMO. (2005). The organization of
projections from the amygdala to visual cortical areas TE
and V1 in the macaque monkey. Journal of Comparative
Neurología, 486, 295–317.
Grandjean, D., Ba¨nziger, T., & Scherer, k. R. (2006). Entonación
as an interface between language and affect. Progress in
Brain Research, 156, 235–247.
Grandjean, D., Sander, D., lucas, NORTE., Scherer, k. r., &
Vuilleumier, PAG. (2008). Effects of emotional prosody on
auditory extinction for voices in patients with spatial
neglect. Neuropsicología, 46, 487–496.
Grandjean, D., Sander, D., Pourtois, GRAMO., Schwartz, S.,
Seghier, METRO. l., Scherer, k. r., et al. (2005). The voices of
Buxton, R. B., Dubowitz, D. J., et al. (1999). Involvement
of striate and extrastriate visual cortical areas in spatial
atención. Neurociencia de la naturaleza, 2, 364–369.
McDonald, j. J., & Ward, l. METRO. (2000). Involuntary listening
aids seeing: Evidence from human electrophysiology.
ciencia psicológica, 11, 167–171.
Michel, C. METRO., Murray, METRO. METRO., Lantz, GRAMO., González, S., Spinelli, l.,
& Grave de Peralta, R. (2004). EEG source imaging.
Clinical Neurophysiology, 115, 2195–2222.
Mogg, K., & Bradley, B. PAG. (1999). Orienting of attention to
threatening facial expressions presented under conditions
of restricted awareness. Cognition and Emotion, 13,
713–740.
morris, j. S., Friston, k. J., Buchel, C., Frith, C. D., Joven,
A. w., Calder, A. J., et al. (1998). A neuromodulatory role
for the human amygdala in processing emotional facial
expresiones. Cerebro, 121, 47–57.
Noesselt, T., Hillyard, S. A., Wolforff, METRO. GRAMO., Schoenfeld, A.,
Hagner, T., Jancke, l., et al. (2002). Delayed striate
cortical activation during spatial attention. Neurona, 35,
575–587.
O¨ hman, A., Flykt, A., & Esteves, F. (2001). Emotion drives
atención: Detecting the snake in the grass. Diario
of Experimental Psychology: General, 130, 466–478.
posner, METRO. I. (1980). Orienting of attention. Quarterly
Revista de Psicología Experimental, 32, 3–25.
Pourtois, GRAMO., Grandjean, D., Sander, D., & Vuilleumier, PAG.
(2004). Electrophysiological correlates of rapid spatial
orienting towards fearful faces. Corteza cerebral, 14,
619–633.
Pourtois, GRAMO., Schwartz, S., Seghier, METRO. l., Lazeyras, F., &
Vuilleumier, PAG. (2006). Neural systems for orienting
1678
Revista de neurociencia cognitiva
Volumen 21, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
1
yo
9
mi
–
1
pag
6
d
7
F
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
oh
7
C
0
norte
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
1
1
0
0
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
.
t
.
/
F
oh
norte
1
8
METRO
a
y
2
0
2
1
attention to the location of threat signals: Un
event-related fMRI study. Neuroimagen, 31, 920–933.
ross, mi. D., & Monnot, METRO. (2008). Neurology of affective
prosody and its functional–anatomic organization in
hemisferio derecho. Brain and Language, 104, 51–74.
Sander, D., Grafman, J., & Zalla, t. (2003). The human
amygdala: An evolved system for relevance detection.
Reviews in the Neurosciences, 14, 303–316.
Sander, D., Grandjean, D., Pourtois, GRAMO., Schwartz, S., Seghier,
METRO. l., Scherer, k. r., et al. (2005). Emotion and attention
interactions in social cognition: Brain regions involved in
processing anger prosody. Neuroimagen, 28, 848–858.
Sander, K., & Scheich, h. (2001). Auditory perception of
laughing and crying activates human amygdala regardless
of attentional state. Cognitive Brain Research, 12,
181–198.
Scherer, k. r., Johnstone, T., & Klasmeyer, GRAMO. (2003). Vocal
expression of emotion. En R. j. Davidson, k. R. Scherer,
& h. h. Goldsmith (Editores.), Handbook of affective sciences
(páginas. 433–456). Oxford: prensa de la Universidad de Oxford.
Schirmer, A., & Kotz, S. A. (2006). Beyond the right
hemisferio: Brain mechanisms mediating vocal emotional
Procesando. Tendencias en Ciencias Cognitivas, 10, 24–30.
Spence, C., & Conductor, j. (1997). Audiovisual links in exogenous
covert spatial orienting. Percepción & Psicofísica, 59,
1–22.
Spierer, l., Meuli, r., & Clarke, S. (2007). Extinction of auditory
stimuli in hemineglect: Space versus ear. Neuropsicología,
45, 540–551.
Van Lancker, D., & Sidtis, j. j. (1992). The identification of
affective–prosodic stimuli by left- and right-hemisphere-
damaged subjects. Journal of Speech and Hearing
Investigación, 35, 963–970.
von Kriegstein, K., Kleinschmidt, A., Sterzer, PAG., & Giraud,
A. l. (2005). Interaction of face and voice areas during
speaker recognition. Revista de neurociencia cognitiva,
17, 367–376.
Vuilleumier, PAG. (2005). How brains beware: Neural mechanisms
of emotional attention. Tendencias en Ciencias Cognitivas, 9,
585–594.
Vuilleumier, PAG., Armony, j. l., Conductor, J., & Dolan, R. j.
(2001). Effects of attention and emotion on face
processing in the human brain: An event-related fMRI
estudiar. Neurona, 30, 829–841.
Vuilleumier, PAG., Richardson, METRO. PAG., Armony, j. l., Conductor, J.,
& Dolan, R. j. (2004). Distant influences of amygdala
lesion on visual cortical activation during emotional
face processing. Neurociencia de la naturaleza, 7, 1271–1278.
Vuilleumier, PAG., & Schwartz, S. (2001). Beware and be
aware: Capture of spatial attention by fear-related
stimuli in neglect. NeuroReport, 12, 1119–1122.
Wildgruber, D., Ackermann, h., Kreifelts, B., & Ethofer, t.
(2006). Cerebral processing of linguistic and emotional
prosody: fMRI studies. Progress in Brain Research, 156,
249–268.
lobo, j. METRO., & Horowitz, t. S. (2004). What attributes guide
the deployment of visual attention and how do they do
él? Naturaleza Reseñas Neurociencia, 5, 495–501.
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
1
yo
9
mi
–
1
pag
6
d
7
F
0
/
1
2
9
1
3
/
7
9
8
/
7
1
6
6
oh
7
C
0
norte
/
1
2
0
7
0
6
9
4
5
2
6
1
2
1
1
/
0
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
1
1
0
0
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
F
/
.
.
.
t
oh
norte
1
8
METRO
a
y
2
0
2
1
Brosch et al.
1679