Competition between Associations in Memory
Jeremy B. Caplan1,2 , Nora Hennies2, and Tobias Sommer2
Abstracto
■ If two associations share an item, one may be remembered at
the expense of the other (BC recalled but not AB). Aquí, we iden-
tify the neural processes by which this competition materializes
and is resolved. We analyzed fMRI signal while participants stud-
ied sets of pairs that reliably induced pair-to-pair associative inter-
ference, but which participants could not fully resolve. Precuneus
activity tracked retrieval of previous pairs during study of later
overlapping pairs. This retrieval apparently produced interfer-
ence by diverting study resources from the currently displayed
pair. Sin embargo, when activity in ventromedial prefrontal cortex,
as well as anterior subregions of the hippocampus, was present
while the earlier pair had been studied, interference was reversed,
and both pairs were likely to be recalled. Angular gyrus and mid-
frontal activity were related to interference resolution once the
participant had seen both pairs. Tomados juntos, asociaciones
compete via precuneus-mediated competitive retrieval, but ven-
tromedial prefrontal cortex may neutralize this by ensuring that
when the earlier association is remembered while studying the
later pair, memories of the two pairs can overcome interference
likely via activity in mid-frontal cortex and angular gyrus. ■
INTRODUCCIÓN
Knowledge often demands that we remember associa-
tions that share an item. Suppose you are learning about
animals. You first find out that chickadees eat seeds, un
association, AB, between seeds (A) and chickadees (B).
Más tarde, you find out that chickadees (B), Sucesivamente, are eaten
by hawks (C), the association BC (boldface is used here to
highlight the shared item). In a second example, you are
keeping track of which teams have played each other in a
children’s sports tournament, to be able to plan future
juegos. Team A (“Trojans”) played against Team B
(“United”) who played against Team C (“Strikers”) en un
later game. De nuevo, this entails remembering both the AB
and BC associations; there is no sense in which the later
association replaces the earlier association. Sin embargo, el
repeated item (B) in two pairs introduces associative inter-
ference. Without mechanisms to specifically address this
interference, mathematical models predict a somewhat
mutually exclusive relationship between the AB and BC
memories (p.ej., Caplan, Rehani, & Andrews, 2014). Eso
es, if AB is remembered, BC is less likely to be remembered
y viceversa. This formal argument seems different than
our daily experience. Claramente, we can remember new infor-
mation related to B without losing the previous memories.
Our overarching question is whether this competition
ever materializes in human memory tasks and is then
resolved, or possibly is never a challenge to begin with.
Note that in these examples, the order of the two associa-
tions relative to each other might or might not be known.
Aquí, we are focused on whether both associations can be
1Universidad de Alberta, Edmonton, Canada, 2University Medical
Center Hamburg-Eppendorf, Hamburg, Alemania
© 2022 Instituto de Tecnología de Massachusetts
remembered, or just one at the expense of the other,
whether or not their relative order is also known.
Most associative interference studies have been mod-
eled not on the AB/BC arrangement but on AB/AC learn-
En g, where the left-hand item is always the shared item
(note that AB/AC is also an associative interference para-
digm, but the shared item is always the cue, whereas in
AB/BC, the cue switches positions). Además, most of
the theory of associative interference has been developed
with two-list procedures, where List 1 contains unambigu-
ous pairs (all the AiBi plus control pairs) y lista 2 introducción-
duces interference (all the AiCi plus more control pairs).
One can look for evidence of competition by comparing
exactitud, on average, of memory for interference pairs
compared with control pairs with no repeated items. Cómo-
alguna vez, it has long been pointed out that there can be a gen-
eral effect, dónde, Por ejemplo, the interference pairs as a
set are remembered worse than the control pairs as a set
(p.ej., Kliegl & Bäuml, 2021; Martín, 1971b; Postman, Rígido,
& Fraser, 1968; Underwood & Schulz, 1960). With two
liza, this could be as simple as participants inhibiting all
the response items of interference pairs in List 2 (or simi-
larly in List 1), regardless of their specific pairings. Return-
ing to our situation, where the overlapping item switches
posición (AiBi and BiCi), if interference occurs directly
between pairs sharing an item, what is needed is a test
of whether a particular AiBi pair competes with a particular
BiCi pair—in other words, a correlation across overlapping
pairs indexed by i. If AiBi and BiCi compete, then if one
pair is remembered, it will often be at the expense of the
otro, producing a negative correlation between the pairs.
If there is no competition at the level of pairs, Podríamos
instead find zero correlation, indicating independence
Revista de neurociencia cognitiva 34:11, páginas. 2144–2167
https://doi.org/10.1162/jocn_a_01900
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
oh
C
norte
_
a
_
0
1
9
0
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(accuracy of BiCi is unrelated to accuracy of AiBi), o un
positive correlation, indicating facilitation (AiBi and BiCi
tend to both be remembered or both be forgotten).
Empirical results actually show correlations of zero, indi-
cating no competition but independence of the memo-
ries, or even positive correlations, indicating a facilitatory
relationship between the memories (Burton, Lek, &
Caplan, 2017; Tulving & watkins, 1974; Delprato, 1972;
Greeno, James, & DaPolito, 1971; Martín, 1971a, 1971b;
Wichawut & Martín, 1971). This could mean that contrary
to the intuition of many researchers, as well as memory
models with competitive retrieval, pairs sharing an item
never, En realidad, compete in memory. Alternativamente, alguno
characteristics of the tasks may have enabled participants
to overcome and sometimes even reverse competition
between overlapping memories by the time memory is
probado. In support of the latter, Caplan et al. (2014) eran
able to show unambiguous evidence for the presence of
competition between overlapping pairs. The key was to
construct lists of the form AB, BC, CD, DE, EF, FA (shuf-
fled). In such “double-function” lists (Primoff, 1938), cada
item is a left-hand item of one pair and a right-hand item of
another pair. When ambiguous pairs are segregated to dif-
ferent lists, as is frequently done with AB/AC paradigms,
participants can use list-membership to resolve interfer-
ence (Kliegl & Bäuml, 2021). Sin embargo, with the double-
function paradigm, which we adopt here (Cifra 1),
list-discrimination cannot be used because interfering
pairs are studied within a single list. Pairs were tested with
a two-response procedure.1 For example, given B as a cue,
participants attempt to recall A and C. In response to B as a
cue, accuracy for BC was negatively correlated with accu-
racy for AB (“same-probe” correlation, where accuracies
of AB and or BC were derived from the two responses
to a single (B) cue; ver Figura 1B, C). Follow-up data-
analyses showed that it was not just the two responses (A
and C) competing to be retrieved, but the memories for
the AB and BC where associations were stored with some-
what mutually competitive strengths (the distinct-probe
correlation described in the methods and elaborated in
Caplan et al., 2014).
If the range of interference resolution is book-ended by
this competition effect and classic independence with
AB/AC learning, this suggests that associative competition
es, en efecto, present initially (evidencia: negative correlation
with the one-list double-function task), but is often
resolved by the time researchers test memory (evidencia:
independence with the two-list AB/AC task). The single list
in the double-function paradigm prevents participants
from using list-membership to protect against associative
ambiguity. Además, the long chains of double-function
pairs might explain why this task produces pair-specific
competition whereas double-function lists with shorter,
three-item chains has produced pair-to-pair indepen-
dence or even net facilitation (Horner, Bisby, Arbusto, lin,
& Burgess, 2015; Horner & Burgess, 2013, 2014). Porque
interference was present but not complete, this paradigm
is well positioned to investigate neural processes underly-
ing both the materialization and resolution of competition
between associations.
Previous studies of neurocognitive mechanisms of asso-
ciative interference, mostly using AB/AC tasks, tener
reported interference on average but not tested for pair-
to-pair competition. Those that have tested for pair-level
effects have produced results consistent with the behav-
ioral literature, confirming near-zero or positive correla-
tions in pair-specific analyses (p.ej., in AB/AC lists, kühl,
Shah, DuBrow, & Wagner, 2010, and in short, three/four-
element double-function lists, Horner et al., 2015). Estos
studies have therefore focused on understanding how the
brain produces good memories of both pairs, but a clear
view of processes by which competition initially material-
izes has remained elusive. Aquí, we investigate the initial
piece of the story when competition first emerges, por
studying brain activity in a one-list double-function asso-
ciative interference paradigm that does show evidence
of competition at the level of pairs. Además, mientras
most neuroimaging studies of associative interference
have recorded only during retrieval and encoding of the
later-studied pairs, we also recorded and analyzed activity
during encoding of the earlier-studied pairs. Even in our
paradigma, interference is resolved for a substantial pro-
portion of pairs. This might occur during study of the
later pair. Sin embargo, a hypothesis that has not yet been
tested is that processes, already during encoding of the
earlier pair, might make it more likely that later inter-
ference can be resolved.
The Present Experiment
We scanned participants while they studied double-
tested with two-response cued recall
function lists,
(Cifra 1). We sought activity related to interference and
its resolution during study of the later pair, but also pro-
spectively, during study of the earlier pair linked by a
common item (AiBi and BiCi; due to randomization,
the earlier pair could be either AiBi or Bi Ci). probamos
the following nonmutually exclusive hypotheses, cual
build on and connect with prior studies that have traced
what are presumably the later interference-resolution
stages (reviewed in more detail in the Discussion sec-
ción). These analyses specifically take advantage of the
fact that we recorded brain activity during the earlier-
and later-studied pairs.
Regarding the source of interference, our first hypothe-
sis was that neurocognitive processes that lead to good
memory also lead the pair to compete, which falls out of
models that assume retrieval is competitive. De este modo, si
encoding strength is indicated by brain activity that shows
a subsequent-memory effect (greater activity during later-
remembered vs. later-forgotten pairs; kim, 2011), eso
same activity should also be associated with competition
between two pairs. En otras palabras, the neurocognitive
processes that lead to good memory also lead the pair to
Caplan, Hennies, y verano
2145
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
oh
C
norte
_
a
_
0
1
9
0
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
oh
C
norte
_
a
_
0
1
9
0
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
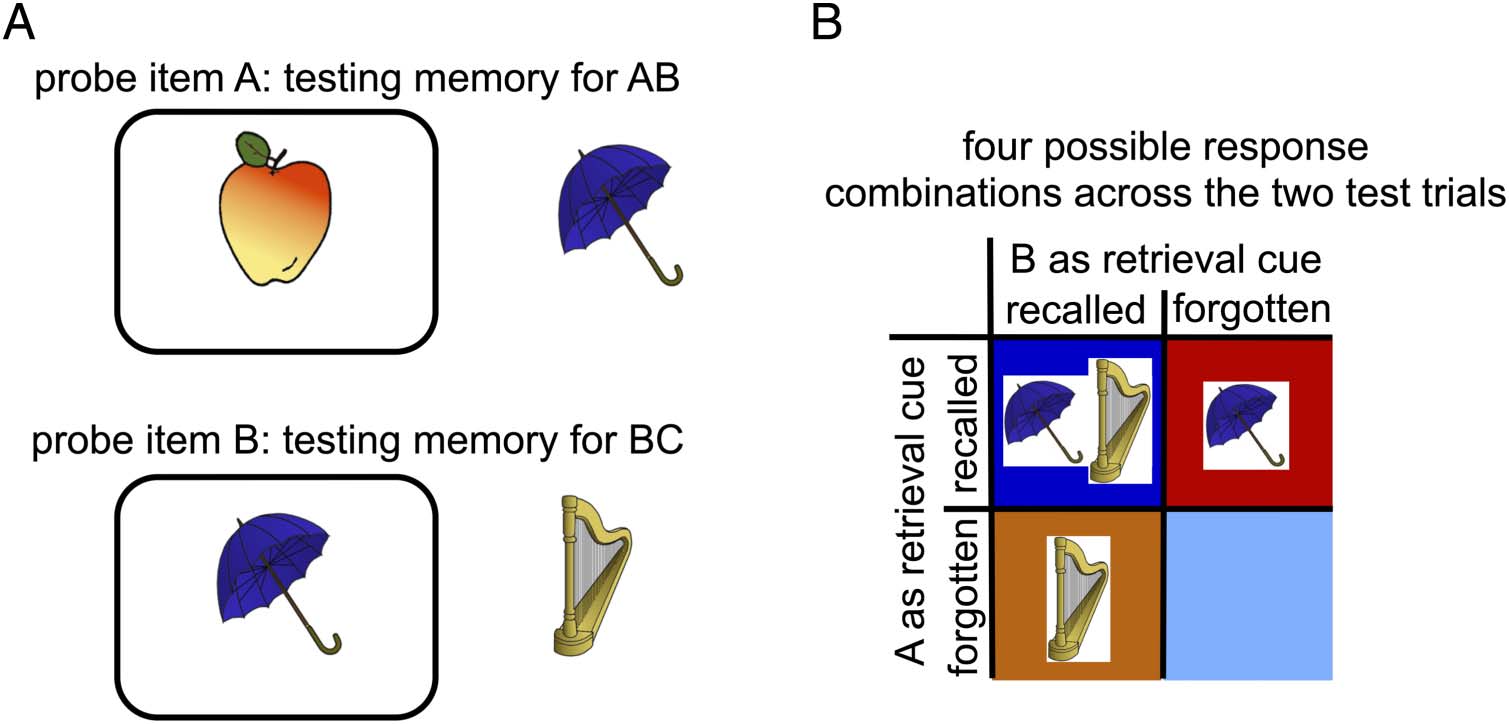
Cifra 1. The double-function list procedure. (A) In each cycle of the task, participants study a sequentially presented set of 12 pares, donde cada
item appears in the left position of one pair and the right position of another pair. After a distractor task (not depicted), each item appears one single
time as a cue, where participants attempt to vocally recall both associates. The procedure affords several ways of scoring accuracy, to assess the
relationship between two overlapping pairs in memory. (B) An example test trial, where A (the apple) is the cue item and the participant can (vocally)
respond with the umbrella, which would indicate memory of AB (tested in the forward direction) and/or with the vase, which would indicate memory
for LA (tested in the backward direction). (C) Four combinations of responses (accuracies) are possible and tallied as in the depicted 2 × 2
contingency table to compute the “same-probe” correlation. (“Forward” here indicates that scoring is done for pairs of pairs considering the forward-
probe direction for the current pair and consequently, the backward-probe direction for the competing pair.) When analyzing brain activity, nosotros
standardize such that the pair that is tested forward on a given test trial is the “current” pair and the pair that is tested backward on the same trial is
the competing” pair. De este modo, the brain-activity analyses are subdivided depending on whether the current pair was the earlier-studied pair or later-
studied pair, respectivamente. Note that the color coding in the contingency tables is maintained across the other figures.
compete. Próximo, consider that associative interference
studies have found evidence that participants remember
the previous pair when presented with a pair with a
repeated item (p.ej., Richter, Chanales, & kühl, 2016;
Horner et al., 2015; kühl, Rissman, Chun, & Wagner,
2011; Kuhl et al., 2010). Extending these findings to
explain pair-specific competition in our paradigm, nuestro
second hypothesis was that retrieval during study pro-
duces interference, consistent with behavioral effects
found by Caplan et al. (2014). De este modo, retrieval-related
activity during the later pair, such as is found in the precu-
neus in related paradigms (p.ej., Himmer, Schönauer,
Heib, Schabus, & Gais, 2019; Brodt et al., 2016; Wimber
et al., 2008; Phillips & Niki, 2002), may be associated with
mutually exclusive memory—one pair remembered at the
expense of the other. Además, activity during the earlier
pair that reflects this propensity to reactivate may then
produce proactive interference. The two hypotheses are
not mutually exclusive and might coexist. As elaborated
in detail in the Results section, the former would be sup-
ported if regions showing the simple subsequent-memory
effect contrast also appear in the interference contrast
during the earlier-studied pair. De lo contrario, the hypothesis
will not be supported, but also not strictly rejected, porque
there could be activity beyond the sensitivity of our mea-
sure. The latter will be supported if regions that appear in
the contrast aimed to isolate reactivation also appear in
the interference contrast during the later-studied pair.
Regarding the resolution of interference, our first
hypothesis was inspired a different body of work, associa-
tive inference. This paradigm has similar task design but
very different research goals. In studies of associative
inferencia, having studied AB and BC, participants must
infer the association AC. Given the major role ascribed
to the hippocampus in transitive and associative inference
(p.ej., Zeithamova & Bowman, 2020; Zeithamova,
2146
Revista de neurociencia cognitiva
Volumen 34, Número 11
Dominick, & Preston, 2012; Bunsey & Eichenbaum, 1996),
our first hypothesis was that activity in the hippocampus
overcomes competition between associations and can
possibly even reverse it. Our second hypothesis, cual
could coexist with the first, was that interference may be
resolved when the participant thinks about (es decir., retrieves)
the earlier-studied pair while viewing the later-studied
pair. Simply retrieving the earlier pair, as mentioned in
the previous paragraph, would be expected to exacerbate
a negative correlation between encoding strengths of the
two pairs, so for interference to be resolved, adicional
processes should be present. Previous studies have
implicated ventromedial prefrontal cortex (vmPFC) en
associative inference (p.ej., Zeithamova & Bowman, 2020;
Spalding et al., 2018; Zeithamova et al., 2012; Kumaran,
Summerfield, Hassabis, & Maguire, 2009). A third hypoth-
esis is possible and testable with data recorded during the
earlier-studied pairs. Eso es, there might be cognitive
processes during the earlier-studied pair that make its rep-
resentation in memory conducive to resolution with the
later-studied pair, reminiscent of prior-knowledge effects
(verano, Hennies, Luis, & Alink, 2022; verano, 2017).
These three hypothesis are not mutually exclusive and
might all coexist. They are each tested in their own right.
The first interference-resolution hypothesis would be sup-
ported if hippocampal regions were isolated in the resolu-
tion contrasts during the later-studied pair. El segundo
hypothesis would be supported if the same region or set
of regions were found to be significant in both the reacti-
vation and resolution contrasts during the later-studied
pair. The third hypothesis would be supported if a region
or regions showed robust effects in the resolution con-
trast during the earlier-studied pair.
Finalmente, once we obtained activity consistent with reacti-
vation of the earlier pair while studying the later pair, nosotros
sought convergent evidence that reactivation was, En realidad,
occurring, using representational similarity analysis (RSA;
Kriegeskorte, Mur, & Bandettini, 2008) and single-voxel
correlations across trials. We also interrogated the nature
of that reactivated activity, testing the hypothesis that later
reactivation of memory of a pair reactivates different,
higher-order, actividad (nonoverlapping areas; Favila, Sotavento,
& kühl, 2020) than the original on-line processing of the
estímulos.
MÉTODOS
Participantes
Thirty (20 women, 10 hombres, edad 28.8 ± 3.5 años; objetivo
sample size was set in advance based on related studies
with similar expected sensitivity; Caplan & Madan, 2016)
healthy participants were recruited from the university
comunidad. Participants had normal or corrected-to-
normal vision and reported no past or present psychiatric
or neurological disorders. The study was approved by the
local ethics committee, Board of Physicians, Hamburg,
Alemania. All participants gave written informed consent
and received monetary reimbursement (10 A/h).
Behavioral Methods
We first adapted the verbal paradigm used by Caplan et al.
(2014) to pictures, similar to our previous studies on emo-
tional associates (Fujiwara, Madan, Caplan, & verano,
2021; Caplan, verano, Madan, & Fujiwara, 2019; Madan,
Fujiwara, Caplana, & verano, 2017). The task is illustrated
En figura 1. To increase power for the fMRI analyses, nosotros
omitted the single-function (control) pairs that were in
the original design. Había 12 double-function pairs
per list, an interpair active-baseline task, only one test
per item, vocal responses instead of typed, and changes
to the timing. The experiment was implemented with
home-grown MATLAB code and the PsychToolbox
(Kleiner et al., 2007; Brainard, 1997; Pelli, 1997) y
CogToolbox (Fraundorf et al., 2014) libraries. The testing
session began with practice outside the scanner (no
analyzed), to familiarize the participant with the proce-
duro. The experimenter ensured that the participants
understood their tasks and were able to recognize the
practice-list stimuli. Most participants then did 18 carreras
(full procedure relevant to a given study set; one with only
17 runs and two with only 16 runs due to failure to start the
scanner, and one with 14 runs due to withdrawing early)
with scanning during the study phase only.
Materials
Stimuli were nameable, colored line-drawing object
images from Rossion and Pourtois (2004), with some stim-
uli removed by the authors when they were thought to be
difficult for German participants to identify. A stimulus was
never used on more than one list (including the practice
lista), and a fresh full random assignment of stimuli to lists
was done for each participant. In each study set, 12 objects
were randomly assigned to a set of 12 pares, con el
restriction that they comprised a ring structure (AB, BC,
CD, DE, EF, FG, GH, HI, IJ, JK, KL, LA) wherein every word
was the left-hand member of one pair and the right-hand
member of another pair (related to the stimulus structure
of Horner et al., 2015, closed-loop triads, but differing in
that here, the “loop” is longer and all items are of the same
material, objects). The classic finding of associative sym-
metry of cued recall of pairs explains some important
aspects of our task design. Eso es, in a standard list of non-
overlapping pairs, when the left-hand and right-hand
items are treated the same (Horowitz, Norman, & Day,
1966), forward (given A, recall B) and backward (given B,
recall A) cued-recall accuracies are equal (Asch, 1969).
Además, in past experiments, if each pair is tested twice,
cued recall of a given pair in the forward and backward
direction nearly always produces the same accuracy; en
otras palabras, there is a very high correlation between for-
ward and backward cued recall, computed across pairs
Caplan, Hennies, y verano
2147
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
oh
C
norte
_
a
_
0
1
9
0
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(Kahana, 2002; Rizzuto & Kahana, 2000, 2001). Both the
equivalence of mean accuracy and high correlation were
also confirmed with the double-function list-structure we
use here (Caplan et al., 2014).
Procedimiento
Pairs were presented sequentially in
Study phase.
random order. The items of each pair were displayed
simultaneously, with the two items separated by a space
in the center of the screen. Each pair was displayed for
3960 mseg (2 × repetition time [TR] = 1980 mseg),
followed by a 150-msec blank interpair interval. Following
the blank ISI, participants completed an active baseline
tarea (described below), lasting from 2- to 4-integer multi-
ples of 2/3 TR (step size 1.32 segundo; range: 2640–5280 msec)
to introduce randomly selected jitter into the timing. Nota
that the onset of the picture was not always at the same
time with respect to the scanner pulse.
Scanning halted during the test phase so that
Test phase.
vocal responses could be recorded. Each item served as a
cue exactly once, requesting up to two responses, pre-
sented in random order. Each test trial, preceded by a
block of the active baseline arrow task as in the study
phase, consisted of a cue word centered on the screen.
The phrase “Bild #1” (traducción: “Picture #1”) was dis-
played centered, underneath the cue object while the par-
ticipant was asked to vocally recall a word or phrase
describing one of the two images that were targets of
the cue object. The vocal response was recorded for
7850 msec into a sound file, but also scored in real time
by the experimenter and a research assistant on a scoring
sheet printed out in advance of the session. The recording
was voice-activated, and the onset time was logged as well.
A second response was collected the same way, with “Bild
#2” displayed. Following previous implementations of the
two-response procedure, participants were told they
could give the two responses in any order they chose
(Caplan et al., 2014; Barnes & Underwood, 1959). Accu-
racy was determined by matching the response with stim-
uli in the word pool. A response was considered correct if
it was one of the two responses given, regardless of the
other response and regardless of whether it was the first
or second response given to the cue.
We were able to confirm that our task produced asso-
ciative interference that could not be entirely resolved by
the time memory was tested, replicating the central find-
ings of Caplan et al. (2014) with the changes described
arriba (see Results section).
Active baseline: Arrow task. To suppress rehearsal and
reduce rest-related hippocampal activity (Rígido & Squire,
2001), participants viewed an arrow pointing left or right,
and responded with the button box with the button con-
gruent with the arrow direction. Each arrow-task trial
lasted a fixed duration (2/3 TR), and the number of trials
was selected to fill the interpair jitter interval.
Behavioral Data Analysis
Correlations between pairs of accuracy outcomes were
evaluated with Yule’s Q, equivalent to a gamma correla-
tion for bivariate data (Kahana, 2002), but can otherwise
be interpreted much like Pearson correlation; Q = 0 indi-
cates statistical independence, Q > 0, positive coupling
between the variables, and Q < 0, negative coupling, or
some level of mutual exclusion (one memory tends to
be recalled at the expense of the other). Statistics were
conducted on log-odds-transformed Q values (logits),
for which residuals are theoretically approximately
Gaussian, thus appropriate for parametric tests, and
resulting p values are the same as if one conducted a χ2
test on the same contingency table (Hayman & Tulving,
1989; Bishop, Fienberg, & Holland, 1975).
Yule’s Q is computed from 2 × 2 contingency table
composed of tallies. As illustrated in Figure 1C, accuracy
of one pair is in rows and accuracy of the other pair is in
columns. If we label the four cells such that Cell A counts
the number of trials for which both pairs are correct, Cell B
when the first pair is correct and the second is incorrect,
Cell C when the first pair is incorrect but the second is
correct, and Cell D when both pairs are incorrect, Q =
(ad − bc)/(ad + bc). The main relationship we are
interested in is Qsame−probe, where accuracy of two pairs
sharing an item is derived from the test trial where the
common item was the cue (for pairs AB and BC, we would
use accuracy from the trial for which B was the cue, and
both A and C were requested as responses). An example
where the earlier pair was evaluated in what would be
equivalent to a forward probe in a noninterference list of
pairs (given B, did the participant recall C?) and the later
pair was evaluated in a backward probe (given B, did
the participant recall A?) is illustrated in Figure 1C. If
associative interference is present, Qsame−probe would
be expected to be negative. If the interference between
the earlier and later pairs is reversed, we would expect
Qsame−probe to be positive. Following Caplan et al. (2014),
we also compute Qdistinct−probe, where accuracy of the
earlier pair is derived from a different test-cue trial than
accuracy of the later pair. This exploits the fact that each
item was given as a cue one time, with spaces for two valid
responses. Consequently, each pair is tested twice, once in
the forward direction, on the trial where its left-hand
item is the cue, and once in the backward direction,
on a different trial where its right-hand item is the cue
(Figure 2A). Thus, the distinct-probe correlation is
computed from a contingency table assembled from
the relationship between pairs sharing an item, where
accuracy of one pair was evaluated on a different test trial
than accuracy of the other pair (Figure 2B). For example,
we can assess memory for AB on the trial with A as the
cue (correct if B was one of the two responses given and
2148
Journal of Cognitive Neuroscience
Volume 34, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. The distinct-probe correlation. (A) As indicated by our terminology, the “distinct-probe” correlation is computed between two different
trials, each of which tests a different one of two pairs sharing an item. One example is depicted here. (B) The “distinct-probe” correlation is then
computed by the contingency table tallied across such pairings; one example is illustrated here. Note that the color coding in the contingency tables
is maintained across the other figures.
incorrect otherwise, corresponding to the forward
response (Figure 2A). This would be yoked to the assess-
ment of memory for BC on the trial with B as the cue
(correct if C was one of the two responses given and
incorrect otherwise; Figure 2A). In this example, both
pairs were tested in the forward direction (on different
trials). This pair of pairs would thus increment, in the
contingency table, Cell A if both were correct, Cell D if
both were incorrect, and Cells B and C if one were correct
and the other incorrect (Figure 2B). The other relevant
cases are where both pairs were tested in the backward
direction (on different trials). Thus, Qdistinct−probe also
measures competition between memories of two associ-
ations sharing an item, but it eliminates the contribution
of immediate competition between two candidate
responses to a single cue, because memory accuracy is
evaluated based on two different test-cue trials. Finally,
we compute a control correlation, Qcontrol, which is a
bootstrap, computed on a contingency table composed
of pairs from the same list that do not share an item
(e.g., AB and CD). This estimates the positive correlation
expected due to variability across lists (Caplan et al.,
2014; Hintzman, 1980). If response candidates compete
in response to a single cue, Qsame−probe would be more
negative than Qcontrol. If, in addition, memories of two
pairs sharing an item have been encoded in a compet-
itive relationship, as found by Caplan et al. (2014),
Qdistinct−probe will also be more negative than Qcontrol;
otherwise, Qdistinct−probe would be equivalent to Qcontrol.
fMRI Methods
Data Acquisition and Preprocessing
fMRI was performed on a 3 T system (Siemens Trio) with a
32-channel head coil. An echo planar imaging T2*-
sensitive sequence in 64 contiguous axial slices (2 × 2 ×
2 mm); TR, 1.98 sec; echo time, 26 msec; Multiband 2; par-
allel acquisition techniques factor 2; flip angle, 70°; matrix
64 × 64) was employed. High resolution (1 × 1 × 1 mm
voxel size) T1-weighted structural MRIs were acquired for
each subject using a 3D magnetization prepared rapid gra-
dient echo sequence. Functional imaging data were pro-
cessed using the SPM12 software (Wellcome Department
of Cognitive Neurology; https://www.fil.ion.ucl.ac.uk
/spm). Functional images were realigned and unwarped
to correct for susceptibility-by-movement artifacts. For
quality control, it was then checked whether individual
participants had excessively moved within run and the
normalization was checked via comparison of the tem-
plate and normalized T1 using the contour-function in
SPM. The anatomical images were coregistered to the
mean functional image of that participant. The anatomical
images were then segmented and transformed into stan-
dard stereotaxic Montreal Neurological Institute (MNI)
space using Diffeomorphic Anatomical Registration
Through Exponentiated Lie Algebra (DARTEL) as imple-
mented in SPM12 and the deformation field applied to
the functional images of the same participant. Functional
images were smoothed with FWHM of 6 mm.
Univariate fMRI Analyses
Individual subjects and group level data were analyses
using the general linear model as implemented in SPM12
in a mass univariate approach. Here, we describe the full
first-level model, with eight regressors of interest. In the
Results section, we describe, in turn, each second-level
model that is derived from those eight regressors.
First-level model. For the first-level model, we sorted
the encoding trials according to the subsequent perfor-
mance in the same probe forward test, that is, current
and competing pair remembered, only current but not
competing pair, not the current but only the competing,
Caplan, Hennies, and Sommer
2149
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
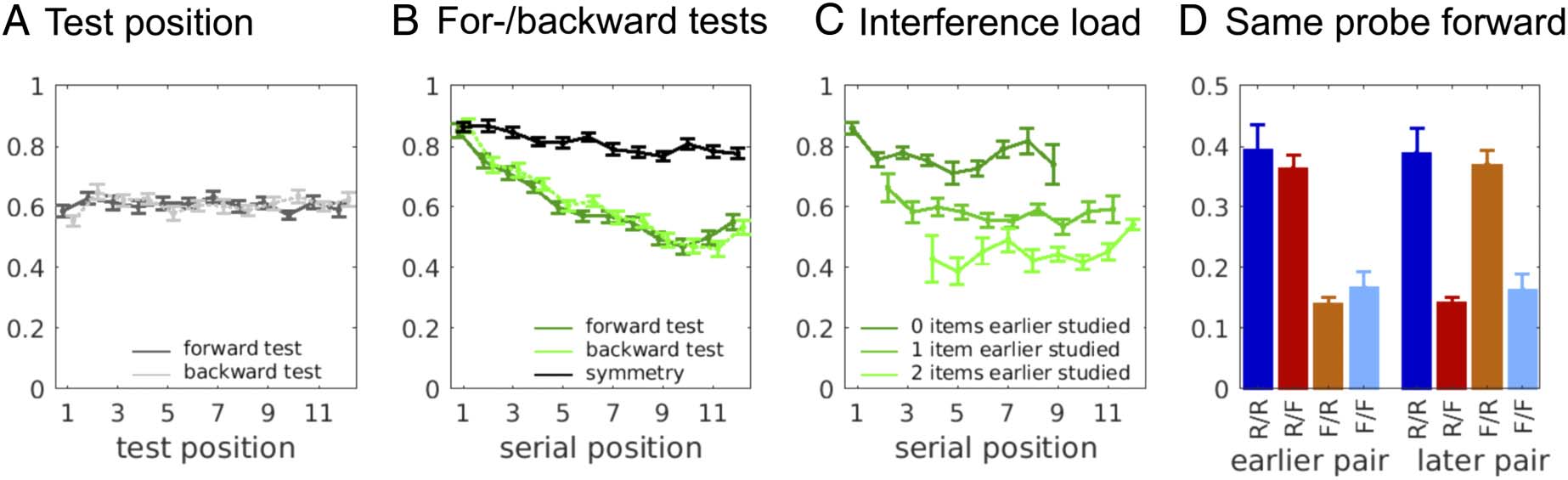
Figure 3. (A) Accuracy as a function of test position and probe direction, illustrating null effects of both factors. (B) Accuracy as a function of serial
position and probe direction, as well as “symmetry,” referring to the proportion of pairs for which either both directions were correct or both were
incorrect. This illustrates that recall was largely symmetric, and serial position is a major factor. (C) Accuracy as a function of number of items within
the pair previously studied (challenge due to interference) and serial position, showing that the steep serial-position effects in (B) are largely
explained by the repetition of items. (D) Accuracy broken down by cell of the contingency table from which Qsame−probe is computed. The high rates
in the R/ F and F/R outcome-conditions are responsible for the negative correlation. The predominance of R/F in the earlier pair and F/R in the later
pair shows that the majority of the competition is proactive interference; the earlier pair is remembered at the expense of the later pair. The colors of
the bars correspond to the cells in Figure 1C.
and neither of the pairs remembered (Figure 1C). In
other words, we sorted trials according to their Yule’s
Q cell in the same-probe forward/backward relationship
(Figure 1C). We focused on the forward/backward rela-
tionship (i.e., the cue that tests the current pair in the
forward direction and the competing pair in the back-
ward direction) because of the expected high associative
symmetry (Kahana, 2002), which was previously con-
firmed for this paradigm using verbal stimuli (Caplan
et al., 2014) and—foreshadowing our current results—
also observed in this experiment (Figure 3B). The symme-
try characteristic ensures that the results of one test direc-
tion generalize to the other. Although the same-probe and
distinct-probe correlations were significantly different
from one another (Figure 4), we focused on the same-
probe relationship for two reasons. First, the number of
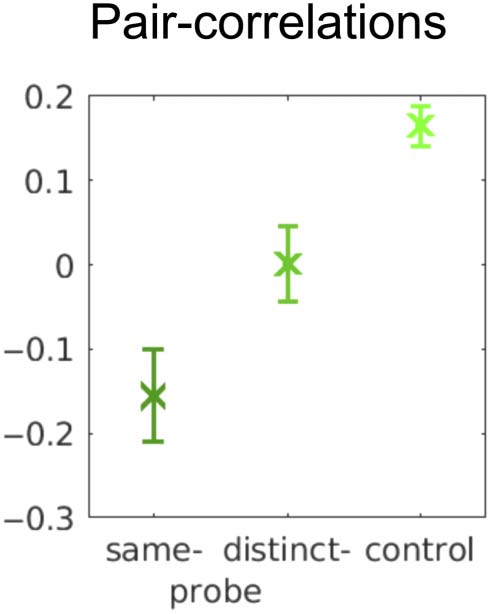
Figure 4. Correlations between pairs of pairs using response accuracy
from a single cued-recall probe (same-probe) or from distinct probe
trials (distinct-probe). Control is a bootstrap, controlling for
independence by estimating the correlation due to list-to-list variability
in accuracy. (See text for more detail.)
trials differentiating the same- and distinct-probe pairings
was too small to be able to reliably identify brain activity
that might be unique to the same-probe activity. Second,
when we reran the analyses based on the distinct-probe
relationship, activation maps were quite similar to those
based on the same-probe relationship. We present only
the same-probe results, as they should include both
competition both at the item level (competition between
the two response candidates to the cue item) and at the
association level (competition between memories of two
associations sharing an item).
Moreover, we dissociated “earlier-studied” pairs from
“later-studied” pairs. “Earlier-studied” pairs were stud-
ied before the competing pair had been studied (AB, in
the case of A as a probe in the example illustrated in
Figure 1C). “Later-studied” were studied after the compet-
ing pair had been studied (LA, in the case of A as a probe).2
Based on these considerations, the first-level model
included eight regressors (earlier- vs- later-studied pair ×
4 Yule’s Q cells) that were created by convolving the
onsets of the conditions with the canonical hemodynamic
response function. The Yule’s Q cells were the four cells of
the contingency table used to compute Q (see Figure 1C).
In addition, six movement regressors were added as nui-
sance variables. The encoding runs of each participants
were concatenated with appropriate adjustments to the
runs-specific constant, the autocorrelation structure, and
the high-pass filter.
Second-level model. On the second level, we contrasted
the parameter-estimates of the these regressors with par-
ticipant as a random factor to identify brain areas where
activity exhibited contrasts consistent with (re-)encod-
ing, interference, and resolution during processing of
earlier- and later-studied pairs. The nonsphericity correc-
tion for violation of the independent and identically
2150
Journal of Cognitive Neuroscience
Volume 34, Number 11
distributed assumption was applied. The particular con-
trasts applied will be detailed in the Results section (see
also Table 1). The fourth cell of the Yule’s Q contingency
table (both pairs incorrect) is ambiguous. Both pairs could
be forgotten because neither was studied well, or because
both were studied well but the two pairs competed such
that neither response could be produced in response to
the cue. For this reason, this fourth cell was usually left
Table 1. fMRI Results during Encoding of the Earlier-studied Pairs
Contrast
Area
x y z Coordinates
z Value
Encoding [1 1 −1 −1]
Anterior hippocampus
vmPFC
Fusiform/lingual/inferior
Occipital gyri
Temporal pole
Middle cingulate gyrus
Superior temporal gyrus
Interference [−1 2 −1 0]
Lingual gyrus
Posterior hippocampus
Inferior frontal gyrus
Posterior hippocampus
[−1 1 0 0]
Resolution [2 −1 −1 0]
vmPFC
Posterior hippocampus
Anterior hippocampus
Precuneus
Insula
Orbitofrontal cortex
Postcentral gyrus
Precentral gyrus
Cerebellum
vmPFC
Posterior hippocampus
Precuneus
[1 −1 0 0]
−20 −10 −18
22 −10 −18
0 36 −24
4 54 −6
22 −86 −10
−36 −74 −16
40 6 −42
−2 −10 38
−52 −20 −8
8 −92 −10
−20 −34 −6
24 −40 2
20 −32 −6
−54 12 6
24 −40 2
8 24 −12
−2 50 2
30 −26 −16
22 −12 −16
−20 −10 −20
8 −54 10
−40 2 12
−34 8 −10
−28 16 −24
−36 −28 54
46 −8 24
12 −56 −26
6 26 −8
−30 −24 −16
−2 −56 20
4.73
4.11
5.29
4.51
7.40
6.43
5.51
5.02
5.26
5.37
3.76
4.43
4.12
5.14
3.93
5.21
4.64
4.72
3.76
3.84
4.39
5.58
5.42
5.33
4.96
5.29
5.49
3.85a
3.50a
3.74a
x y z coordinates of the peaks of clusters in MNI space. Correction for multiple comparisons was done on the whole-brain level or within predefined
anatomical ROIs, specifically the vmPFC, bilateral anterior hippocampus, and precuneus. In the contrasts, the regressors are: (1) current (earlier-
studied) and competing (later-studied) pair remembered, (2) only current pair remembered, (3) only competing pair remembered, (4) neither pair
remembered. When two contrasts are listed for the same named contrast (here, “interference” and “resolution”), the contrasts should not be viewed
as independent, but, rather, the second as a follow-up refinement of the first, to test the robustness of the results.
a Trend toward significance p < .1.
Caplan, Hennies, and Sommer
2151
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
out of the contrasts, but beta values nonetheless plotted
alongside the beta values for the other cells when illustrat-
ing the results.
It is important to note that the presence of competition,
its neutralization, or reversal is evaluated by computing
correlations across pairs. We cannot infer whether compe-
tition between memory of any one AB pair was resolved
with memory of its corresponding BC pair. The way we
have structured our analyses should be viewed in terms
of working on the assumption that, for example, more
cases for which both pairs are remembered reflect resolu-
tion of interference than cases for which one, but not the
other, pair is remembered. No different than other
contrast-based analyses of neuroimaging data, the results
should be viewed with this limitation in mind. If two
conditions in a contrast were truly sampling from a single
distribution and differed only because of random noise,
the contrast would usually be nonsignificant.
Psycho-physiological interaction analyses. As a follow-
up analysis, we conducted four psycho-physiological inter-
actions (PPI; Friston et al., 1997) analyses using the results
of the second-level resolution contrast, namely, left and
right angular and mid frontal gyri (from Figure 6B) as seeds
(thresholded p < .05, corrected) and compared coupling
during resolution (Condition 1) with retro- and proactive
interference (Conditions 2 and 3). Parameter estimates of
the individual PPIs were tested on the second level using a
one-sample t test. Because we did not a priori select which
areas to use as seed regions, these PPIs should be viewed
as exploratory.
Trial-to-trial Variability: Multi-voxel and Single-voxel
Effects Related to Visual Perception and Reactivation
Whereas the analyses just described identify changes in
mean activity across conditions, the following set of
analyses identify activity that varies across trials within con-
dition. Our first question was whether we could find con-
vergent evidence in support of the idea that memory of
the earlier-studied pair is, in any concrete sense, retrieved
(reactivated) during the later-studied pair, as other neuro-
imaging studies have found. Then, we asked whether later
reactivation of memory of a pair reactivates the same or
different activity (nonoverlapping areas) as on-line pro-
cessing of the stimuli (Favila et al., 2020).
To get activity estimates for each individual trial as input
for these analyses, detailed in the following sections, for
each trial, we created an independent first-level model
with one regressor containing only the corresponding
trial, that is, its onset convolved with the canonical hemo-
dynamic response function, and one for all other trials in
that fMRI run (Mumford, Turner, Ashby, & Poldrack,
2012). Again, six movement regressors were added as nui-
sance variables; the correction for autocorrelation and a
high-pass filter were applied. The t maps testing the beta
of the trial of interest in each model against the implicit
baseline was used for the following RSA to reduce the
influence of noisy voxels (Dimsdale-Zucker & Ranganath,
2018). To maintain consistency with the other fMRI analyses,
we used smoothed single-trial data with the same 6-mm
FWHM kernel because it has been shown that that smooth-
ing does not decrease the sensitivity of RSA (Hendriks,
Daniels, Pegado, & Op de Beeck, 2017; Kriegeskorte,
Cusack, & Bandettini, 2010; Op de Beeck, 2010).
Representational similarity analyses. We used RSA to
evaluate voxel-pattern similarity between pairs of study
trials. In contrast to the previous contrasts and the follow-
ing single-voxel series of analyses, RSA analyses identify
activity that produces a “voxel-pattern,” that is, carrying
information in the relative weightings across voxels (e.g.,
Kriegeskorte et al., 2008; Haynes & Rees, 2005) that would
traditionally be considered close enough together com-
prise a single “region.”
The first RSA was conducted to identify areas involved in
on-line processing of the objects and the second to identify
areas involved in successful reactivation from memory of
earlier-studied pairs while studying the later pairs. In both
RSAs, we employed a whole-brain searchlight approach
(radius 5 voxels) and correlated (Pearson correlation across
voxels) the resulting vectors of trial-specific t values across
conditions of interest where only t values of trials within the
same run were correlated. These correlation coefficients
were averaged after Fisher z transform and were saved as
value for the center voxel of the current searchlight.
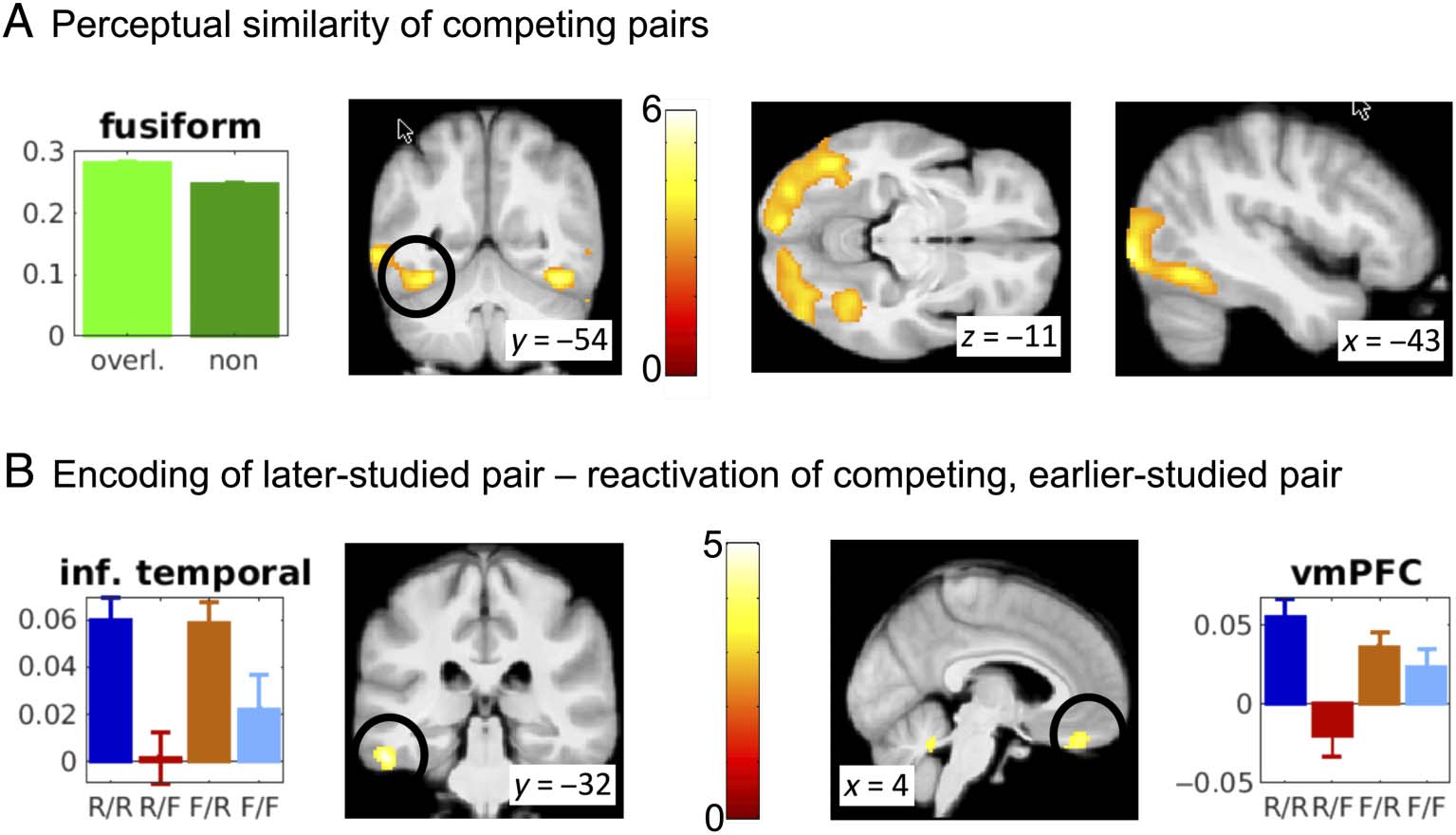
Perceptual processing. With the first RSA, we aimed to
identify areas involved in processing the objects based on
perceptual similarity of the two trials sharing an object.
Therefore, we correlated activity patterns of pairs with
one overlapping item (e.g., BC with AB and CD in
Figure 1A) and contrasted these against the correlation
of each pair with all other pairs in that run (e.g., BC with
DE, LA, EF, etc.; Figure 1A). The correlation coefficients
of both conditions were contrasted in a paired t test as
implemented in SPM12.
Successful reactivation. With the second RSA, we aimed
to identify areas involved in successful reactivation of the
competing pair. The rationale of this RSA was to identify
areas where the similarity of activity patterns during
encoding of later- and its competing earlier-studied pairs
was greater when reactivation was presumed to have taken
place versus no evidence of reactivation. These two pairs
(e.g., BC and AB in Figure 1A) share the overlapping item
(e.g., B), which will result in the same degree of similarity
in all of the 4 Yule’s Q cells. However, only when the
competing pair is reactivated during processing the later-
studied pair is there additional similarity expected in areas
that are involved in processing and reactivation of these
pairs (e.g., AB is reactivated during BC-studying resulting
in similarity of brain activity with AB). The resulting corre-
lation coefficients in each of the 4 Yule’s Q conditions
were contrasted on the second level using SPM12.
2152
Journal of Cognitive Neuroscience
Volume 34, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Pattern similarity and univariate activity differences.
Pattern similarity can be caused not only by distributed
patterns of activity but also by univariate activity differ-
ences between conditions because a stronger signal in a
condition could lead to stronger correlations of their trials
(Wagner, van Buuren, Bovy, & Fernández, 2016). To rule
out this confound, we averaged the activity in spheres
(radius 5 voxels) around the peaks identified by the RSAs.
First, we contrasted this mean activity between conditions
to test for differences in univariate activity that would not
have survived the correction for multiple comparisons
applied to the univariate analyses described above. Then,
we correlated the individual difference in mean activity
between conditions with the individual difference in
pattern similarity.
Single-voxel activity correlations across trials. Finally,
we conducted single-voxel analogues of the two RSAs. This
series of analyses tests for regional activity that might
reflect perceptual processing and reactivation regardless
of whether that activity produces a “voxel-pattern.” If a
voxel reflects activity that is reactivated (whether high or
low in value), even if there is no difference in mean activity
between successful and unsuccessful reactivations, its
activity may covary between pairs that are successfully
reactivated. For each voxel individually, we correlated
activity between the earlier and later pair sharing an
item, across such pairings. We contrasted those correla-
tions with correlations across pairings that did not share
an item to identify voxels related to perceptual process-
ing. We also contrasted correlations of voxel-activity
between pairs sharing an item for which the earlier pair
was later recalled versus not recalled, to assess possible
reactivation of single-voxel activity. The larger the corre-
lation, the more within-condition variability of activity
between pairs of trials. This across-pair, within-condition
correlation analysis was done on the single-voxel level but
also using a searchlight approach (radius 5 voxels) and
averaging the correlation coefficients within the spheres.
The resulting correlation maps were contrasted between
conditions on the group level as in the RSAs using
SPM12.
Statistical Significance
Results of all fMRI analyses were considered significant at
p < .05, FWE corrected for multiple comparisons across
the entire scan volume or within the a priori defined
anatomical ROIs. Based on the previous literature, ROIs
for the univariate analyses were the hippocampus, precu-
neus, and vmPFC, for the mulivariate analyses in addition
the inferior temporal and fusiform gyri. Bilateral hippo-
campus, bilateral precuneus, bilateral inferior temporal
gyrus (all three subsections combined), and bilateral
fusiform gyrus were computed from the Harvard-Oxford
cortical and subcortical structural atlases. A vmPFC ROI
was manually traced on the mean T1 image based
on previously published postmortem data (Mackey &
Petrides, 2014) using ITK-SNAP 3.6.0 ( Yushkevich et al.,
2006).
RESULTS
Behavior
We first report behavioral results showing that the prior
finding of pair-specific competition could be replicated
with a pure double-function design (and thus, twice as
many double-function pairs per list as in the mixed lists
used by Caplan et al., 2014), with particular attention to
whether associative interference is present, rather than
being largely resolved. We also examined whether there
was a predominance of proactive or retroactive interfer-
ence in the behavioral data. Accuracy was in the middle
of the allowable range (Figure 3), comfortably far from
ceiling and floor, conducive to examining modulation of
accuracy by competing pairs. There was little effect
on accuracy of test position (Figure 3A) or direction
(Figure 3A, B), replicating symmetry of mean cued-recall
accuracy (Asch & Ebenholtz, 1962; Kahana, 2002). Next,
consider that each item was used as a recall cue just once,
but two responses were collected. This means that for a
single pair, BC, the outcome of forward cued recall of BC
can be evaluated by checking whether the participant
produced C in response to B as the cue. Backward cued
recall is evaluated on a different test trial: Given C as the
cue, did the participant produce B as one of the
responses? Tallied in this way, the correlation of forward
and backward recall of individual pairs was high; Q = .87
(SEM interval: [−.017, +.015]), extending previously
observed associative symmetry (e.g., Sommer, Schoell,
& Büchel, 2008; Sommer, Rose, & Büchel, 2007; Caplan,
2005; Kahana, 2002; Rizzuto & Kahana, 2001) to pairs of
pictures. More importantly, this shows that associative
symmetry holds even in the presence of heightened com-
petition (Caplan et al., 2014; Rehani & Caplan, 2011).
Thus, we can safely collapse over test position and test
direction in the remaining analyses. There were large
effects of serial position on accuracy (Figure 3B). How-
ever, plotted differently, Figure 3C shows that the driving
factor was not serial position, but rather the amount of
interference present while the pair was studied. Pairs that
were presented before either constituent item had been
studied were most accurate, followed by pairs for which
one, but not the other item had been seen, and the low-
est accuracy for pairs for which both items had previously
appeared in other pairs. In other words, proactive inter-
ference is a major source of variability in accuracy in this
task. Figure 3D shows the breakdown of accuracy as a
function of cell within the contingency table from which
Qsame−probe is computed (Figure 1C), that is, between
the earlier-studied and later-studied of two pairs sharing
an item. The first and fourth conditions are cases where
both pairs are recalled or both pairs are not recalled,
Caplan, Hennies, and Sommer
2153
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
respectively. The middle conditions indicate competition
between memory for the two pairs. Inspection of those
middle bars shows that it was more common for the
earlier-studied pair to be remembered at the expense
of the later-studied pair, than the other way around. Con-
sistent with Figure 3C, proactive interference was more
common than retroactive interference.
Analyses of the interference-related Yule’s Q values
(Figure 4, explained in the Methods section and illus-
trated in Figures 1 and 2) confirmed the presence of
direct competition between pairs sharing an item. Namely,
both Qsame−probe and Qdistinct−probe were significantly
below Qcontrol, t(29) = −6.57, p < .0001, and t(29) =
−5.28, p < 0.0001, respectively, replicating Caplan et al.
(2014). Qsame−probe was also significantly more negative
than Qdistinct−probe, t(29) = −3.25, p = .0029, a novel
finding suggesting the presence of both simultaneous
competition at time of test between two candidate target
items, and a competitive relationship between memory
for the pairs, themselves.
The conceptual replication of the negative correlation
between overlapping pairs extends the boundary condi-
tions for this result. Modeled on the task used by Caplan
et al. (2014), our paradigm differed in several ways: 1) The
stimuli were drawings of objects instead of words. 2) Recall
was vocal rather than typed. 3) To maximize data-yield to
support the analyses of interest, we omitted single-function
pairs and doubled the number of double-function pairs.
Despite all these changes, recall of pairs sharing an item
was negatively correlated, indicating that as in Caplan et al.
(2014), pairs competed directly in memory. The correlation
was significantly negative (differing from behavioral findings
from associative interference paradigms as described in
the Introduction), but not as negative as possible (−1).
This satisfied the initial conditions we sought: Associative
interference was present, and partly, but not completely
resolved by the time memory was tested.
fMRI Results: Overview
As described in the Methods section, the first-level model
included eight regressors of interest (earlier- vs. later-
studied pair × 4 memory-outcome conditions corre-
sponding to the quadrants of the Yule’s Q table;
Figure 1C). Isolating activity during earlier-studied pairs
identifies activity during encoding that results in either
proactive interference (only the current but not the
later-studied pair will be remembered) or in resolution
of interference of the current pair with the later-studied
pair (both associations will be remembered). Activity dur-
ing encoding of later-studied pairs (similar to AC in classic
paradigms) can be related to either proactive interference
of the earlier studied pair, retroactive interference of the
current pair, or resolution with the earlier-studied pair. We
first report analyses of activity during the earlier-studied
pair and then during the later-studied pair.
The order of the regressors representing the 4 Yule’s Q
conditions was always (1) current and competing pair
remembered, (2) only current pair remembered, (3) only
competing pair remembered, (4) neither pair remem-
bered. In the text, because our main analyses are restricted
either to the earlier- or to the later-studied pair, we use
shorthand, noting only the first four (earlier-studied) or
last four (later-studied) regressors.
It should be borne in mind that the following contrasts
are not designed to be mutually exclusive, but rather, to
identify particular relationships of regional activity to the
task. As we will note, when a region appears in one con-
trast, whether it does or does not appear in another one
may further specify its putative role in the task.
fMRI Results: Activity during Encoding of Earlier-
studied Pairs
Subsequent memory of the earlier-studied pair. Before
delving into effects specifically related to interference
and its resolution, we conducted a simple subsequent-
memory effect analysis, to identify the “basic” encoding
regions. We identified areas showing a general subsequent
memory effect for the current (earlier-studied) pair irre-
spective of memory for the competing (later-studied) pair
by the contrast [1 1 −1 −1]. A set of regions comprising
bilateral anterior hippocampus, vmPFC, and visual areas
showed robust subsequent memory effect (SME) (Table 1,
“encoding”).
Activity related to proactive interference. To identify
areas where activity during encoding of the current pair
resulted later in interference with the competing pair, that
is, that showed greater activity when the current pair was
remembered but the competing pair was not remem-
bered, the contrast [−1 2 −1 0] was applied. The case of
both pairs forgotten was omitted from this contrast
because it is ambiguous whether such cases are due to a
failure of a resolution attempt or that one or both
associations were individually not remembered. Regions
within posterior hippocampus on both sides showed this
effect (Table 1, “interference,” and Figure 5A), suggesting
that particular hippocampal-dependent study processes
produce a memory that eventually will compete with
encoding of the later-studied pair. The inferior frontal
and lingual gyri also showed this effect.
Following up on these findings, the contrast [−1 1 0 0]
specifically tests for greater activity when only the current,
and not both pairs, will be remembered. Only a right pos-
terior hippocampal region showed this effect. We did not
observe any areas showing activity related to retroactive
interference from the later-studied competing pair (con-
trast [−1 −1 2 0] as well as [0 −1 1 0]), consistent with
the small corresponding behavioral effect (Figure 3D).
Activity related to proactive resolution. Next, we ask if
there is any activity present during processing the earlier-
studied pair that later results in resolution with the
2154
Journal of Cognitive Neuroscience
Volume 34, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
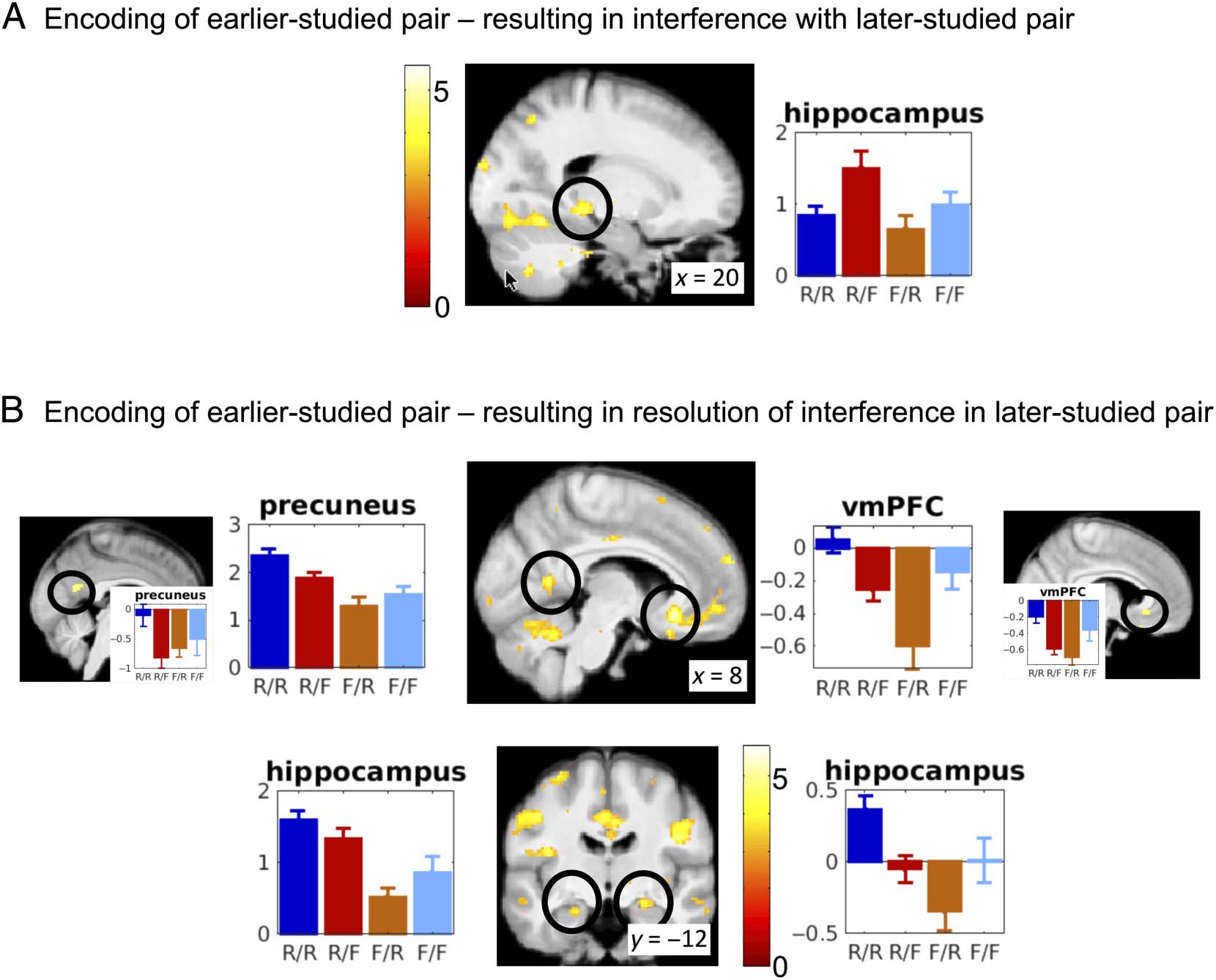
Figure 5. fMRI activity during encoding of earlier-studied pairs. (A) Among other areas (Table 1), activity in the bilateral posterior hippocampus
during processing of earlier-studied pairs resulted in interference with the later-studied competing pair as identified by the contrast [−1 2 −1 0]. (B)
Activity in the anterior hippocampus, vmPFC, and precuneus (and other areas) during processing of the earlier-studied pairs resulted in resolution
with the later encoded competing pair. The large plots of the parameters estimates of the four conditions reflect activity in the peak identified by the
contrast [2 −1 −1 0]. The colors of the bars corresponds to the cells in Figure 1C. The small plots of parameter estimates inserted in the precuneus
and vmPFC represent activity in subregions showing also specifically greater activity when both pairs were remembered compared with when only
the earlier-studied pair was remembered, that is, [1 −1 0 0]. Statistical maps are overlayed on the mean normalized structural image of the
participants. Visualization threshold p < .001. Activity on the y axis is in arbitrary units. Circles surround the local maxima from which activity is
plotted. In these contrasts, the current pair is the earlier-studied pair and the competing pair is the later-studied pair.
competing pair (Table 1, “resolution,” and Figure 5B),
testing our third hypothesis about the cause of resolution.
The contrast [2 −1 −1 0] isolates activity associated with
both pairs being subsequently remembered, versus only
one pair remembered but not the other. Again, hippocam-
pal subregions bilaterally, most extensively in the anterior
subdivision, showed greater activity during successful
encoding of the earlier-studied pair when the later-
studied competing pair was also successfully remem-
bered. Numerous other regions, most importantly the
vmPFC and ventral precuneus, showed a similar pattern
of activity.
Following up on these findings, the contrast [1 −1 0 0]
specifically tests for greater activity when both pairs were
remembered versus the current pair remembered but the
(later-studied) competing pair not. In other words, given
that the current pair was remembered, was the competing
pair also remembered or not? This contrast also isolates
resolution-related brain activity during study of the earlier
of the two pairs. Anterior hippocampus, vmPFC, and pre-
cuneus were also found in this contrast.
fMRI Results: Activity during Encoding of
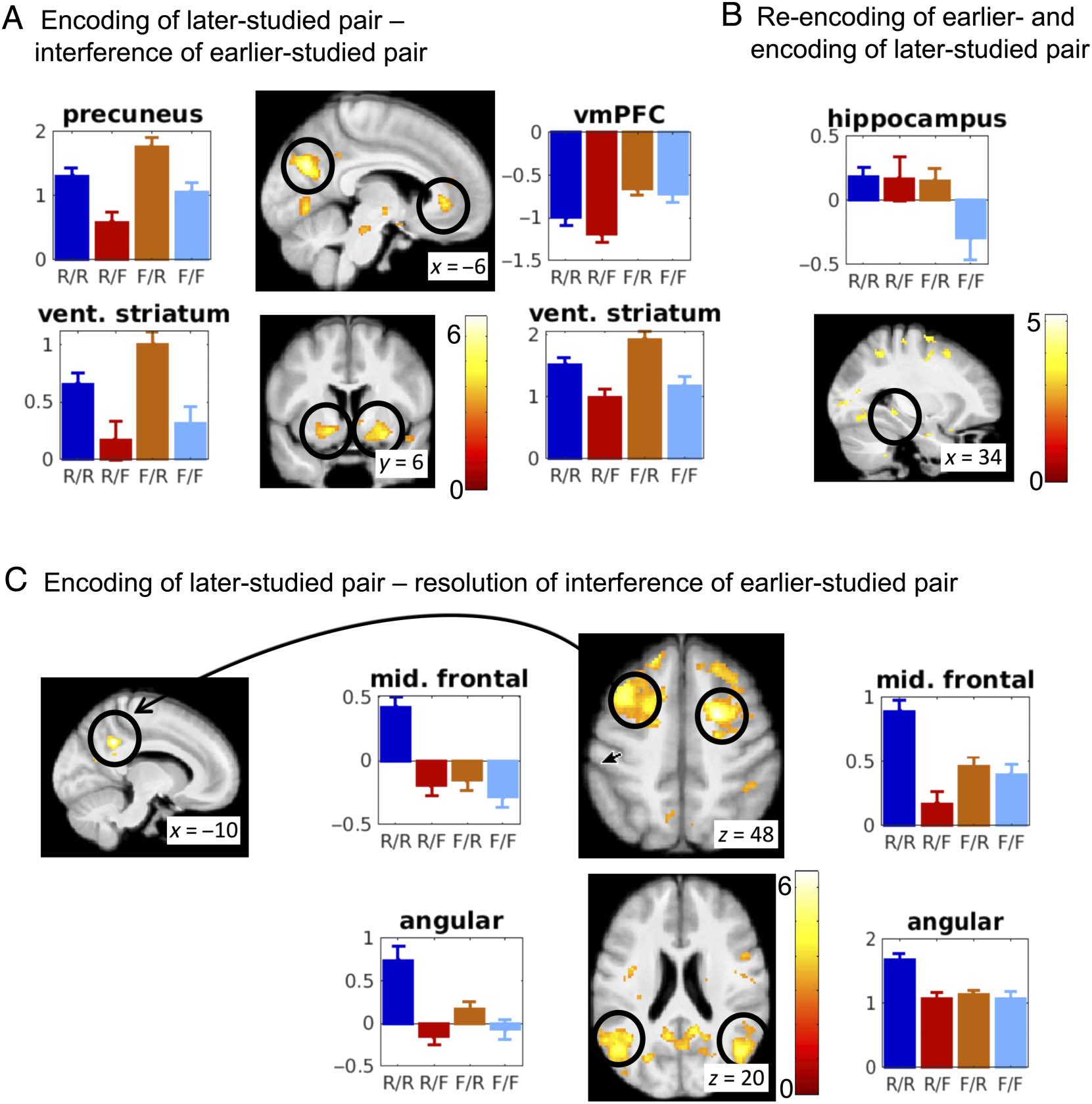
Later-studied Pairs
Paralleling the analyses of the earlier-studied pairs, we first
tested, for the later-studied pairs, which brain areas show a
simple SME (contrast [1 1 −1 −1]), which revealed a set of
regions comprising the supramarginal, middle temporal, and
fusiform gyri but not the hippocampus (Table 2, “encod-
ing”). However, this might be simply an effect of lower power
because there was less successful encoding of the later-
studied pairs, in particular whereas the proportions of
later-studied pairs in the first regressors (Yule’s Q cell 1)
was similar to the earlier-studied pairs (Figure 3D), there
were substantially fewer in the second regressor due to pro-
active interference. Moreover, this contrast confounds inter-
ference effects with subsequent-memory effects, as we shall
Caplan, Hennies, and Sommer
2155
Table 2. fMRI Results during Encoding of the Later-studied Pairs
Contrast
Area
x y z Coordinates
z Value
Encoding [1 1 −1 −1]
Supramarginal gyrus
Middle temporal gyrus
Temporal pole
Fusiform gyrus
Reactivation [1 −2 1 0]
vmPFC
Interference [−1 −1 2 0]
Precuneus
Angular gyrus
Thalamus
Middle frontal gyrus
vmPFC
Precuneus
Ventral striatum
[−1 0 1 0]
Precuneus
(Re-)encoding [1 1 1 −3]
Posterior hippocampus
Resolution [2 −1 −1 0]
Precuneus/posterior cingulate
Middle temporal gyrus
Middle frontal gyrus
Angular gyrus
Temporal pole
[1 −1 0 0]
Precuneus/posterior cingulate
Middle frontal gyrus
Angular gyrus
Temporal pole
36 −34 40
−58 −36 34
−56 −56 4
−38 6 −38
30 −62 −6
−2 32 2
8 52 −12
4 −62 26
−40 −72 34
−4 −12 −6
30 4 56
−6 36 −2
−8 −70 28
12 −66 30
−16 6 −10
24 10 −8
−8 −72 30
12 −68 30
−28 −38 −6
34 −30 −6
−50 −52 6
4 −62 26
−12 −48 26
22 4 48
−36 16 48
−46 −64 20
58 8 −24
4 −62 26
−12 −56 24
24 2 50
−14 22 56
−46 −64 20
40 −68 32
58 8 −24
5.29
5.77
5.13
5.04
4.89
4.47
4.30
6.43
5.03
5.53
5.05
4.50
6.46
5.15
4.65
5.01
3.82a
3.75a
3.74
3.92
5.04
4.59
5.64
6.19
5.29
5.73
5.09
6.28
5.18
5.74
5.13
5.16
4.93
5.12
x y z coordinates of the peaks of clusters in MNI space. Correction for multiple comparisons was done on the whole-brain level or within predefined
anatomical ROIs, specifically the vmPFC, bilateral hippocampus, and precuneus. In the contrasts, the regressors are: (1) current (later-studied) and
competing (earlier-studied) pair remembered, (2) only current pair remembered, (3) only competing pair remembered, (4) neither pair remem-
bered. When two contrasts are listed for the same named contrast (here, “interference” and “resolution”), the contrasts should not be viewed as
independent but, rather, the second as a follow-up refinement of the first, to test the robustness of the results.
a Trend toward significance p < .1.
2156
Journal of Cognitive Neuroscience
Volume 34, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
see below when we seek activity related to encoding or
“re-encoding of earlier and encoding of later-studied pair.”
Activity during the later-studied pair that might reflect
memory of the earlier-studied pair. Next, we looked
for regions that might reflect retrieval of the earlier-studied
pair during study of the later pair (although this should be
viewed not as conclusive; for convergent evidence, see the
follow-up correlational analyses below). Areas that showed
a reactivation-like pattern of activity were identified by the
contrast [1 −2 1 0], on the logic that if the earlier pair was
remembered while studying the later pair, it is more likely
to be recalled correctly during the subsequent memory test
than if the earlier pair were not remembered during the
later pair. Moreover, reactivation offers more encoding
time to the earlier pair, which should also increase the
probability that the earlier pair would later be recalled.
Regions showing this pattern included the vmPFC, precu-
neus, middle frontal, and angular gyri (Table 2, “reactiva-
tion”). Because these analyses measure the amount of
activity increase rather than the information-content of that
activity, this set of putative “reactivation” regions might
reflect neural processes that evoke reactivation rather than
the reactivated information, itself, which might be housed
elsewhere. The RSA analyses will show some convergent
support for this latter interpretation.
During study of a later pair, retrieval of the earlier pair
could result either in interference with the current encod-
ing or in resolution of interference, which we target in the
next contrasts. Regions that appear in the current “reacti-
vation” contrast and the “interference” contrast are candi-
date regions for introducing interference via reactivation.
Likewise, regions that appear in the current “reactivation”
contrast and the “resolution” contrast are candidate
regions for resolving interference by acting on memory
of the earlier pair.
A potential cause of proactive interference. We next
asked if there is any activity present during processing
the later-studied pair that reflects proactive interference
from the earlier-studied pair (Table 2, “interference,”
and Figure 6A). We applied the contrast [−1 −1 2 0] to
identify areas where only the competing but not the cur-
rent pair will be remembered, excluding the ambiguous
case where both pairs are forgotten. In addition to the
vmPFC and precuneus that also showed reactivation-like
activity patterns, the bilateral ventral striatum showed
greater activity when only the competing, earlier-studied
but not the current pair was successfully encoded. Very
similar precuneus regions also appeared in a follow-up
contrast, [−1 0 1 0], although not reaching significance
( p < .1), contrasting only the competing pair recalled
versus both pairs recalled.
We pause here to emphasize how remarkable it is that
precuneus, vmPFC, and ventral striatum regions showed
such robust activity differences that were primarily due to
memory for a pair studied at an entirely different time in
the study phase. This is compellingly consistent (although
not conclusive) with the idea that these regions retrieve
prior memories, but if there is too much reactivation, this
risks leaving little opportunity to resolve those retrieved
memories with the current pair. Convergent evidence for
the presence of reactivation is reported below, in the trial-
to-trial correlational analyses. We did not observe any region
where activity related to retroactive interference on the
competing, earlier-studied pair (contrast [−1 2 −1 0]).
Resolution of the earlier-studied pair with the current
later-studied pair. Next, we isolated areas where reacti-
vation of the competing, earlier-studied pair resulted not
in interference but possibly resolution with the currently
processed pair (Table 2, “resolution” and Figure 6C). As
with the earlier-studied pairs, the contrast [2 −1 −1 0]
was used, contrasting both pairs recalled versus only one
recalled. The precuneus/posterior cingulate as well as the
middle frontal and angular gyri were active during proba-
ble resolution of interference. These regions also exhib-
ited the simpler, follow-up contrast, [1 −1 0 0], both pairs
recalled versus the current pair forgotten but the compet-
ing pair recalled. Our first resolution hypothesis, that the
hippocampus produces resolved associations, was not
supported (although it is possible that a hippocampal
effect is present but underpowered). Rather, our second
(although not mutually exclusive) resolution hypothesis
was supported, namely, nonhippocampal regions more
plausibly produce resolved associations.
Of the four PPIs, using the left and right angular and
mid-frontal gyri as seed regions, only one reached signifi-
cance individually ( p = .014, Z = 4.30; but after a Bonfer-
roni correction, was only a trend). This was characterized
by the left mid-frontal gyrus seed region exhibiting stron-
ger coupling with the precuneus during integration than
interference ([−8 −50 36], arrow in Figure 6C).
Re-encoding of earlier and encoding of later-studied
pair. The suspected presence of reactivation raises the
possibility that the earlier, retrieved pair could be encoded
during study of the later pair, or both could be encoded at
that time. We wondered if activity in the hippocampus
might reflect this associative encoding, where sometimes
the current (later-studied) pair is encoded, other times the
competing pair (earlier-studied, retrieved during the cur-
rent trial) is instead being encoded, or both. We collapsed
together trials for which either the current or competing
pair was correct, and contrasted those with trials for which
both pairs were forgotten (Table 2, “(re-)encoding,” and
Figure 6C). In other words, the contrast [1 1 1 −3] expresses
the idea that activity in a particular brain region might reflect
the total amount of encoding occurring, whether it is
devoted exclusively to the current pair, exclusively to the
(reactivated) competing pair, or split somehow between
the two. A region within the posterior hippocampus on each
side had an activity profile very much like this, as well as a
region within the middle temporal gyrus. Thus, posterior
Caplan, Hennies, and Sommer
2157
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. fMRI activity during encoding of later-studied pairs. (A) Activity in the vmPFC, precuneus, and striatum during processing of later-studied
pairs resulted in interference by the earlier-studied competing pair as identified by the contrast [−1 2 −1 0] (Table 2). (B) The hippocampus was
involved when either the presumably reactivated competing pair, the current pair, or both were (re-)encoded. (C) Activity in the middle frontal and
angular gyri as well as the posterior cingulate showed greater activity if the competing earlier-studied pair was successfully resolved. The arrow in (C)
illustrates the higher coupling of the left mid-frontal gyrus with precuneus during interference resolution than interference (this exploratory PPI
reached a trend toward significance when corrected for the four possible seeds shown in (C). The large plots of the parameters estimates of the four
conditions reflect activity in the peaks. The colors of the bars correspond to the cells in Figure 1C. Statistical maps are overlayed on the mean
normalized structural image of the participants. Visualization threshold p < .001. Circles surround the local maxima from which activity is plotted. In
these contrasts, the current pair is the later-studied pair and the competing pair is the earlier-studied pair.
hippocampal and middle temporal activity during the
later-studied pair could reflect encoding of whatever is in
mind—the current or competing pair or potentially both.
It is interesting that anterior regions of the hippocampus
showed neither the “naïve” subsequent-memory effect,
nor this (re-)encoding effect during the later-studied pair.
Trial-to-trial Variability Analyses of fMRI: Convergent
Evidence of Reactivation
The set of regions that we identified with the “reactivation”
contrasts is consistent with the idea that reactivation
occurs, but the link to this interpretation is still quite
indirect. If reactivation is, in fact, taking place, then as in pre-
vious studies, we should be able to observe some similarity
in brain activity between the earlier- and later-studied pairs
when reactivation presumably succeeded. The regions
identified in the previous contrasts more likely reflect con-
trol processes that identify the repetition, initiate or
retrieve the information, rather than the information-
content of the reactivated memory, itself. In fact, it is also
possible that those “reactivation” regions enhance later
memory of the earlier-studied pair completely apart from
any putative reactivation. Here, we ask if we can obtain
more concrete evidence that the earlier pair is sometimes
remembered during study of the later pair. Namely, is
2158
Journal of Cognitive Neuroscience
Volume 34, Number 11
there similarity of brain activity between earlier- and
later-studied pairs that might reflect the memory that is
first constructed and then later remembered? (See, e.g.,
Koen & Rugg, 2016; Staresina, Alink, Kriegeskorte, &
Henson, 2013; Lee, Kravitz, & Baker, 2011, for this ap-
proach). In addition, coupled to this: What is, in fact,
reactivated? To tackle this question, we focused on a
hypothesis from Favila et al. (2020) that reactivated
information is different (nonoverlapping regions) than
on-line perceptual processing of the stimuli. In other
words, what the participant remembers is at a different
level of representation, presumably higher-order, than
visual-processing of the object-pairs.
We therefore conducted analyses seeking brain activity
that reflected the hypothetical reactivation, itself
(described in the Methods section). That is, we sought
activity patterns, as well as individual voxels, that were
common to the earlier and later pair sharing an item, when
the earlier pair was subsequently remembered (presum-
ing successful reactivation) versus not (less successful
reactivation). We compared this to a control analysis that
identified similarity of activity due to the common item
being visually processed during both pairs.
RSA. The maxima of the perceptual similarity RSA were
in the inferior occipital ([−42 −88 −2], Z = 5.82; [46
−80 −6], Z = 4.97) and fusiform gyri ([−34 −54 −16],
Z = 4.80; [40 −54 −16], Z = 5.11) where the fusiform
cluster were the most anterior (Figure 7A). This estab-
lishes the set of regions showing pattern similarity likely
due to visual processing of the item that was common
between the earlier- and later-studied pair.
Reactivation was tested as in the univariate analyses with
the contrast vector [1 −2 1 0]. This revealed a cluster in left
inferior temporal gyrus ([−52 −32 −22], Z = 4.42) and
vmPFC ([−4 40 −24], Z = 3.96; Figure 7B). Thus, the reac-
tivation regions were further in the ventral visual pathway
and did not include those whose voxel-patterns reflected
visual processing of the shared stimuli, similar to Favila
et al. (2020).
To test whether the reactivation RSA results were driven
by univariate differences effects, we first contrasted mean
activity in the spheres around the peak voxels identified by
the RSA between Conditions 1 and 3 with Condition 2.
Second, we correlated the individual differences between
these conditions with the corresponding differences in
similarity ( Wagner et al., 2016). (Note that this control
analysis of mean activity within spheres is not sensible
for the perceptual similarity RSA due to the fact
that all trials are in both conditions lead to equal mean
activity in both conditions.) In the inferior temporal
gyrus, mean activity showed a trend toward a significant
difference between conditions, t(30) = 1.55, p = .065,
and this difference did not correlate with the individual
difference in similarity between conditions (r = .094,
p = .612). In the vmPFC, a similar pattern emerged,
namely, a trend toward a significant difference between
conditions, t(30) = 1.43, p = .082, and no correlation of
the individual differences with similarity difference (r =
.274, p = .136).
Figure 7. RSA. The measure ( y axis and color scale) is similarity (Fisher z-transformed correlation coefficients). Note that the absolute values of the
correlation coefficients cannot be directly interpreted, for instance the nearly zero correlation in Condition 2 (R/F) in (B) does not necessarily mean
that the patterns do not show any similarity (Dimsdale-Zucker & Ranganath, 2018). (A) Similarity greater between pairs sharing an item than pairs
with no shared items. This presumably reflects similarity in visual-perceptual processing of the stimuli, caused by the common item. (B) For pairs
sharing an item, similarity was greater when the earlier pair was later remembered than when it was later forgotten, regardless of whether the later
pair was recalled. This is presumably caused by memory of the earlier pair while studying the later pair, which results in additional encoding of the
earlier pair. Note that the color coding in the bar graphs corresponds to that in the contingency tables in Figure 1C.
Caplan, Hennies, and Sommer
2159
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Single-voxel correlations in activity across trials. For
the perceptual similarity analysis, the searchlight approach
resulted in a very similar set of regions as the perceptual
similarity RSA, that is, bilateral early visual cortex (calcarine
[−14 −96 −8], Z = 5.52; inferior occipital gyrus [−50 −72
−2], Z = 5.72; [52 −72 −2], Z = 6.26) and fusiform gyrus
([−32 −48 −10], Z = 3.73; [24 −46 −12], Z = 4.56) and,
in addition, the precuneus ([−6 −54 10], Z = 4.09). With-
out the searchlight, the single-voxel analysis produced less
smooth results but were otherwise similar, also identifying
the early visual cortex ([−12 −94 −8], Z = 6.09; [20 −88
−8], Z = 5.87) and the fusiform gyrus ([−28 −44 −12],
Z = 4.10; [30 −40 −14], Z = 4.58).
For the reactivation analysis, the searchlight approach
converged with the RSA analysis, revealing clusters in
the left inferior temporal gyrus ([−54 −32 −24], Z =
3.35; and vmPFC ([−4 34 −26], Z = 4.34). Without the
searchlight, the single-voxel analysis also identified the
left inferior temporal gyrus ([−54 −38 −24], Z = 3.16)
and vmPFC ([0 34 −20], Z = 3.4). However, those
clusters were not significant after correcting for multiple
comparisons.
Thus, both distributed patterns (RSA analyses) and
regional activity (single-voxel correlations) reflect pair-
specific activity that is present during study of the earlier
pair and then reactivated during study of the corre-
sponding later pair. The regions showing these effects
are different and higher-order than those reflecting per-
ceptual processing of the displayed stimulus.
DISCUSSION
With the first behavioral paradigm in which participants
are challenged by pair-specific associative interference
but cannot fully resolve it, we identified neural processes
that explain how competition between associations mate-
rializes, and how it can be overcome. We review these
main findings and discuss how they dovetail with findings
from prior studies in which pair-specific interference has
been largely resolved.
The Origin of Associative Interference
A rare feature of our paradigm, offering access to brain
activity throughout the study phase, is that it allows us
to follow proactive interference-related activity from the
earlier-studied pair to the later-studied pair. Our first
hypothesis, that simple competitive retrieval based on
encoding strengths is the source of interference, was not
supported; brain regions that led to good memory for the
earlier-studied pair (“encoding” contrast in Table 1) did
not show interference effects (“interference” contrasts).
Consistent with the established importance of the hippo-
campus for associative memory (e.g., Caplan & Madan,
2016; Saksida & Bussey, 2010; Konkel & Cohen, 2009;
Eichenbaum, Yonelinas, & Ranganath, 2007; Mayes,
Montaldi, & Migo, 2007; Davachi, 2006; Rudy & O’Reilly,
2001; Cohen, Poldrack, & Eichenbaum, 1997; Nadel &
Moscovitch, 1997; Rudy & Sutherland, 1989; O’Keefe &
Nadel, 1978), hippocampal regions were associated with
subsequent memory of the earlier-studied pair. However,
different hippocampal subregions (more posterior) were
related to future associative interference.
Instead, our second hypothesis was supported; the pri-
mary source of interference appeared to be retrieval of the
earlier-studied pair while studying the later-studied pair.
This echoes neuroimaging findings on pairs with repeated
items. This includes associative interference paradigms for
which pair-specific interference had been largely success-
fully resolved (e.g., Richter et al., 2016; Horner et al., 2015;
Kuhl et al., 2011; Kuhl et al., 2010), as well as associative
inference paradigms, where the participant’s explicit
goal is to combine information from both component
pairs to answer inference questions (e.g., van Kesteren,
Rignanese, Gianferrara, Krabbendam, & Meeter, 2020;
Zeithamova & Bowman, 2020; Zeithamova et al., 2012;
Zeithamova & Preston, 2010) and confirms behavioral
evidence of such retrieval occurring in the one-list
double-function paradigm (Caplan et al., 2014). Specifi-
cally, high activity in posterior hippocampus (Figure 5A),
with insufficient activity in vmPFC, precuneus, and ante-
rior hippocampus (Figure 5B), lay the basis for interfer-
ence with the later-studied pair.
During later-pair processing, the “reactivation” and
“interference” contrasts share regions with nearby peaks
in a set of regions comprising vmPFC, precuneus, and
striatum (Table 2, Figure 6). Precuneus and striatum have
been implicated in memory retrieval (Clos, Schwarze,
Gluth, Bunzeck, & Sommer, 2015; Schwarze, Bingel,
Badre, & Sommer, 2013; Huijbers et al., 2012). While
studying the later-presented pair, right precuneus, along
with bilateral vmPFC, left angular gyrus and thalamus,
and right middle frontal gyrus, was associated with good
memory for the earlier-studied pair, regardless of memory
of the current pair. This is remarkable, because the con-
trast completely omits brain activity during initial study
of the earlier-studied pair.
Aligning these results with those from the interference
contrast (later-studied pair), a left-sided vmPFC region
and the right precuneus region recur in the interference
contrast with nearby peaks. Although speculative, this
pattern of findings reinforces our previous suggestion that
strong encoding of the earlier-studied pair in posterior
hippocampus together with weak encoding in the
vmPFC-precuneus-anterior hippocampus might result in
subsequent reactivation via the striatum and precuneus
that reduces encoding of the later, competing pair (cf.
Long & Kuhl, 2019).
Resolution of Interference
As with the origin of interference, our paradigm also
allows us to track processes supporting resolution of
interference across time, from the earlier- to the later-
2160
Journal of Cognitive Neuroscience
Volume 34, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
studied pair. Our first hypothesis, extrapolating from
transitive/associative inference (e.g., Horner et al., 2015;
Zeithamova et al., 2012; Heckers, Zalesak, Weiss, Ditman,
& Titone, 2004; Preston, Shrager, Dudukovic, & Gabrieli,
2004; Dusek & Eichenbaum, 1997; Bunsey & Eichenbaum,
1996), that the hippocampus supports successful resolu-
tion of the two pairs was unsupported. Perhaps the hippo-
campus was implicated in prior inference tasks for its
role in supporting memory of the component associations
(A → B and B → C), enabling a chained retrieval solution
to the inference, or supporting encoding of the inferred
A → C association during the B → C trial (Koster et al.,
2018), without necessarily resolving any competition.
Our second hypothesis was supported. Although reacti-
vation can produce interference, if there is not too much
reactivation, the participant might sometimes use extra-
hippocampal processes (Figure 6C) to resolve competi-
tion between the two co-activated associations, potentially
producing facilitation between the memory for the two
pairs (Burton et al., 2017; Wahlheim, Maddox, & Jacoby,
2014; Jacoby & Wahlheim, 2013; Wahlheim & Jacoby,
2013; Kuhl et al., 2010). Evidence for this can be seen in
regions that appeared in both the “reactivation” and “res-
olution” contrasts (Table 2), which include a region within
right precuneus, middle frontal, and angular gyri. Consis-
tent with this, it is interesting that the activity level when
both pairs were recalled (Figure 6A, R/R condition) was
not the highest, but actually lower than when only the ear-
lier (competing) pair was remembered and greater than
when only the later (current) pair was remembered. Thus,
if activity in the precuneus reflects the total amount of
retrieval of the earlier pair while studying the later pair,
this suggests that too much reactivation leaves too little
opportunity for mid-frontal and angular gyrus to resolve
interference. The angular gyrus has been linked to integra-
tion and retrieval of supramodal complex semantic knowl-
edge (Gilboa & Marlatte, 2017) and multimodal feature
integration during episodic retrieval (Bonnici, Richter,
Yazar, & Simons, 2021). The computations of the angular
gyrus, as part of a wider lateral parietal system, enable the
on-line dynamic buffering of multisensory spatiotempo-
rally extended representations (Humphreys, Ralph, &
Simons, 2021; Xie, Li, Xie, Xu, & Peng, 2019). The mid-
frontal gyrus has been implicated in attention (Bourgeois,
Sterpenich, Iannotti, & Vuilleumier, 2022), which may be
this region’s specific role in resolving interference here.
Finally, our third hypothesis, which we were for the first
time able to investigate, was supported. That is, activity
during the earlier-studied pair apparently set the initial
conditions for future resolution of interference. Among
the regions that exhibited this effect (“resolution” contrast
in Table 1) were vmPFC bilaterally, anterior hippocampus
bilaterally, and precuneus. Similar regions were identified
with the simple “encoding” contrast. This suggests that
(contrary to the first interference hypothesis discussed
in the previous section), a well-studied pair that is likely
to be remembered is also one that has a good chance of
being reconciled with an overlapping association in
the future. This hippocampal activity was during the
earlier-studied pair, so it does not reflect the resolution
process, itself, but rather the formation of a memory with
favorable properties for future resolution. The role of
vmPFC here is consistent with numerous prior studies
implicating vmPFC in encoding (Fujiwara et al., 2021)
and in forming integrated representations (Gilboa &
Marlatte, 2017). However again, vmPFC is not apparently
carrying out the resolution, itself, but laying down the con-
ditions for future resolution by other regions, namely,
middle frontal and angular gyri and posterior cingulate,
whereas the later competing pair is studied. Interestingly,
an interplay of the vmPFC and hippocampus with the pre-
cuneus during encoding has been previously associated
with incorporation of novel information into existing sche-
mata (Sommer et al., 2022; Sommer, 2017). Liu, Grady,
and Moscovitch (2017) found that an advantage for houses
associated with famous faces versus nonfamous faces was
attributable to activity in anterior hippocampus, vmPFC,
and precuneus activity—similar to what we observed.
The famous faces were presumed to provide richer repre-
sentations to which the houses could be bound. In our
task, this same set of regions may similarly provide addi-
tional details to the representation of the first pair, making
it easier for the later pair to be reconciled with the earlier
memory. This is in contrast to posterior hippocampus,
which may produce a memorable association that is less
amenable to resolution of competition with another pair.
Importantly, participants do not simply resolve interfer-
ence once it materializes, during the later-studied pair.
Rather, at least as important, the way in which the
earlier-studied pair is processed can be critical for subse-
quent successful resolution.
Cognitive Mechanisms of Resolution
of Interference
Having first identified the source of pair-specific interfer-
ence and then identified brain activity related to the neu-
tralization, or even reversal, of that interference, we now
consider the cognitive processes that the latter activity
might reflect.
First, we note that the term “integration” arises repeat-
edly in the associative interference neuroimaging literature,
but with several meanings, each of which might be related
to the neurocognitive processes we identified here. Integra-
tion can refer to the formation of a composite representa-
tion of two pairs in memory, such as encoding not just AB
and BC, but something like ABC. This is the idea behind
instructions to participants to form integrative imagery as
a way to resolve interference (e.g., Burton et al., 2017;
Anderson & Bell, 2001; Smith & Hunt, 2000; Anderson &
McCulloch, 1999). However, such integrated representa-
tions in memory are hard, if not impossible, to confirm.
Correct recall of both pairs (or even a positive correlation
across pairs of pairs; both pairs remembered or both
Caplan, Hennies, and Sommer
2161
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
forgotten; Burton et al., 2017) could conceivably result from
such an integrated representation, but this kind of result
could have other plausible causes. Strictly speaking, the
positive correlation only tells us that memory for AiBi and
BiCi have a source of shared variance.
Consider retrieval of the earlier pair while studying the
later pair. If the earlier pair is well encoded, it might be
retrieved with little effort and rapidly, thus displacing very
little encoding time from the later pair. In this way, a highly
recallable earlier pair might facilitate encoding and subse-
quent recall of the later pair without any direct integration.
Conversely, a poorly studied earlier pair, itself less likely to
be recalled correctly, may require more study time to be
recalled, thus also obstructing encoding of the later pair,
making it likely that both pairs will not later be recalled.
Our findings are somewhat in line with this; hippocampal
activity that produced a subsequent-memory effect
during the earlier pair was associated with good memory
for both pairs.
Second, drawing an analogy to associative inference,
resolution, and even reversal of interference in our task,
when it does occur, might be caused by participants
encoding the inferred association, AC, after retrieving
the earlier pair while studying the later pair. This could
positively couple the two pairs by adding a new retrieval
route. Suppose that given B as a probe, BC were not
remembered. If AB was remembered, A could then be
used as a retrieval cue for C, via the encoded inferred AC
association. Inferring the indirect association might be
one role of mid-frontal and angular gyrus activity during
the later pair. Then, posterior hippocampal activity, which
appears to be an agnostic (re-)encoder during the later
pair, could then store the indirect association if it were
successfully produced, or else the retrieved earlier associ-
ation or the current association.
A different notion is that ambiguity between similar stim-
uli may be addressed in part by changing the representa-
tions so that they are more orthogonal, termed “pattern
separation” (e.g., Poppenk, Evensmoen, Moscovitch, &
Nadel, 2013; Norman & O’Reilly, 2003; O’Reilly &
McClelland, 1994; Marr, 1971). Interestingly, Becker
(2017) found that the resolution of interference was solved
by her model by making representations more similar
rather than less similar (echoing the effects of strategy
found by Burton et al., 2017), but the more general idea
that representations may become more distinct (not nec-
essarily orthogonal) when both associations are brought to
mind together has been proposed by, for example, Kuhl
et al. (2010, 2011), and Chanales, Dudukovic, Richter,
and Kuhl (2019). Representational Hierarchical Theory,
which assumes no special role for the hippocampus in
memory, per se, implicates the hippocampus precisely in
offering the brain the ability to discriminate stimuli that
would be processed as highly similar by more upstream
regions (Cowell, Barense, & Sadil, 2019; Bartko, Cowell,
Winters, Bussey, & Saksida, 2010; Cowell, Bussey, &
Saksida, 2010; Saksida & Bussey, 2010; Bussey & Saksida,
2002). This could neutralize interference by reducing the
similarity-based ambiguity in encoded memories, but it
might also reverse interference if the formation of distinc-
tive representations were synergistic, likely to be successful
for both pairs or unsuccessful for both pairs. The latter
mechanism is, in fact, the antithesis of an integrated repre-
sentation. The mid-frontal and angular gyrus activity during
the later pair, related to success with both pairs, might con-
tribute to distinctive encoding of one or both pairs, that is,
however, unlikely to be like pattern separation.
If construction of distinctive representations is the main
mechanism of resolution of interference, our findings dur-
ing the earlier pair suggest that the distinctiveness process
can begin even before both pairs are known. The anterior
hippocampal and vmPFC activity during the earlier pair
might already achieve some distinctiveness, enabling the
participant to focus more on forming a distinctive repre-
sentation of the later pair once it is presented.
This repertoire of possible mechanisms of resolution of
interference suggests why our paradigm, in contrast to
other associative interference paradigms, leaves some
competition unresolved. Associative interference in other
paradigms is typically in triad form, such as AB/AC. Con-
structing an integrated representation, ABC could result
in AB and AC both being remembered (or both forgotten).
Encoding the inferred, BC association, could result in
good memory of both AB and AC. As already noted, assum-
ing AC was not memorable, if the participant can retrieve B
with A as a cue, the BC association offers a backup retrieval
path to potentially produce C as well. In the three-item
loops used by Horner et al. (2015), again, storing ABC or
storing all component associations could both produce
positive correlations in memory of AB, BC, and CA. With
our larger ring structure, those approaches may resolve
competition in one part of the ring, but at the same time
increase competition in another part of the ring. Suppose
the BCD is stored. That may positively correlate memory
tests of BC and CD with one another, but it introduces an
additional source of competition when testing the pairs AB
and DE. Thus, both “integrative” solutions may explain
why our participants can resolve some competition
between overlapping pairs, but this reasoning also shows
why it may be quite challenging to resolve all interference
after only a single exposure to a list.
Forming more distinctive representations is a process that
might have a benefit without such a cost. Increasing the
distinctiveness of BC from CD will also be likely to increase
the distinctiveness of BC from AB. In fact, as just suggested,
this might be the mechanism by which activity during the
earlier-studied pair increases the chance of resolution of
interference, even before the later-studied pair is known.
Convergent Evidence of Retrieval during Study and
the Nature of Retrieved Memories
A complementary set of analyses of trial-to-trial variability
produced more direct evidence for the presence of
2162
Journal of Cognitive Neuroscience
Volume 34, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
reactivation of the memory of the earlier-studied pair
while studying the later pair. These analyses also sug-
gested the relevant reactivated information was at a rela-
tively high level of representation, different from more
face-value visual processing of the stimuli. To understand
the logic here, assume that a region (either a set of individ-
ual voxels or a legitimate voxel-pattern) reflects high-order
features of memorial representations. Those features
might be high in value or low in value; hence, no differ-
ence is expected in average activity (and would be missed
by the simple contrasts). What matters is whether the
same value occurs both during the earlier pair and the later
pair, when the earlier pair is brought back to mind. For
example, suppose Region X reflects the amount of vivid-
ness of an image constructed to bind two objects together.
One stimulus pair, AB, might be high in vividness; evi-
dence of its reactivation would be high BOLD signal both
during presentation of AB and during subsequent presen-
tation of BC. A different pair, DE, might be remembered
with verbal rather than imagery mediation. Consequently,
BOLD signal in Region X would be low during presentation
of DE. If X reflects information that is reactivated, that low-
value BOLD level would be expected to reiterate itself dur-
ing study of a later pair, EF. Thus, the prediction is not that
Region X should (necessarily) exhibit greater activity when
reactivation is successful versus not, but that its value
should covary across trials (pairs) for which reactivation
was likely to have been successful. This correlation should
be greater than when computed across pairs for which
reactivation was probably not successful. The results
indeed supported similarity between the earlier-studied
and later-studied pairs when reactivation would presum-
ably have been more likely to have succeeded.
Moreover, the reactivation leading to subsequent mem-
ory of the earlier pair appeared at a high level of represen-
tation, further into the ventral visual pathway than on-line
perceptual processing (Favila et al., 2020). Thus, partici-
pants do not, apparently, reactivate in the sense of re-
imagining the two objects in the original pair with their
detailed visual features, but rather, remember a highly
processed, combined representation they had con-
structed of the two items. The control analyses confirmed
that those were pair-specific distributed activity patterns
(Koen & Rugg, 2016). Given the involvement of vmPFC,
in particular, trial-to-trial variability in this region may
reflect variability in producing the high-order representa-
tion of the association that is conducive to further elabo-
ration. Such elaboration might, for example, support
formation of an integrated representation or storage of
the inferred, indirect association as just discussed.
Alternatively, if retrieval of the earlier-studied pair is
decoupled from the low-level visual information in the
stimulus, but rather, at a high level of representation,
the more abstract representation might be more condu-
cive to being transformed into a representation with
more distinctiveness from other representations, or the
high-level nature of the retrieval might indicate that the
encoded representation of the earlier-studied pair may
have often already been modified to be more distinctive.
Limitations
To stick closely to the previous paradigm that produce
unambiguous evidence of pair-specific competition
(Caplan et al., 2014), we had to use verbal stimuli that
would be conducive to recall. Our findings might be
restricted to the verbal domain. To expand into nonverbal
memory domains, it will first be necessary to adapt and
validate the paradigm for forced-choice responses.
Conclusion
Our behavioral paradigm, with brain activity analyzed
during the entire study phase, revealed that associative
interference is not produced passively due to strength-
based competition between overlapping memories, but
rather due to proactive interference when the earlier-
studied pair diverts encoding resources away from the
later-studied pair, reflected in activity in the precuneus,
among other regions. However, if the retrieval-related pre-
cuneus activity is not too strong, numerous additional
regions, possibly coordinating with the precuneus, includ-
ing angular gyrus and mid-frontal gyrus (but perhaps not
hippocampus), can resolve interference. Finally, resolu-
tion of interference is enabled when the earlier-studied
pair is studied in a particular way, involving activity in
vmPFC and anterior hippocampus.
Reprint requests should be sent to Jeremy B. Caplan, Depart-
ment of Psychology, Biological Sciences Building P217, Univer-
sity of Alberta, Edmonton, AB, T6G 2E9, Canada, or via e-mail:
jcaplan@ualberta.ca.
Data Availability Statement
Data will be made available upon email request to the cor-
responding author.
Funding Information
Natural Sciences and Engineering Research Council of
Canada (https://dx.doi.org/10.13039/501100000038).
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Caplan, Hennies, and Sommer
2163
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance.
Notes
1. This was confusingly called “modified modified free recall”
in the past; see, for example, Barnes and Underwood (1959),
Burton et al. (2017), and Tulving and Watkins (1974).
2.
In the familiar AB/AC procedure, earlier- and later-studied
pairs would correspond to AB and AC, respectively, but in the
current paradigm, pairs could appear in any order (i.e., LA could
just as likely precede AB in our example).
REFERENCES
Anderson, M. C., & Bell, T. (2001). Forgetting our facts: The
role of inhibitory processes in the loss of propositional
knowledge. Journal of Experimental Psychology: General,
130, 544–570. https://doi.org/10.1037/0096-3445.130.3.544,
PubMed: 11561927
Anderson, M. C., & McCulloch, K. C. (1999). Integration as a
general boundary condition on retrieval-induced forgetting.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 25, 608–629. https://doi.org/10.1037/0278
-7393.25.3.608
Asch, S. E. (1969). A reformulation of the problem of
associations. American Psychologist, 24, 92–102. https://doi
.org/10.1037/h0027121
Asch, S. E., & Ebenholtz, S. M. (1962). The principle of
associative symmetry. Proceedings of the American
Philosophical Society, 106, 135–163.
Barnes, J. M., & Underwood, B. J. (1959). “Fate” of first-list
associations in transfer theory. Journal of Experimental
Psychology, 58, 97–105. https://doi.org/10.1037/h0047507,
PubMed: 13796886
Bartko, S. J., Cowell, R. A., Winters, B. D., Bussey, T. J., &
Saksida, L. M. (2010). Heightened susceptibility to
interference in an animal model of amnesia: Impairment in
encoding, storage, retrieval—Or all three? Neuropsychologia,
48, 2987–2997. https://doi.org/10.1016/j.neuropsychologia
.2010.06.007, PubMed: 20561536
Becker, S. (2017). Neurogenesis and pattern separation: Time
for a divorce. Wiley Interdisciplinary Reviews: Cognitive
Science, 8, e1427. https://doi.org/10.1002/wcs.1427, PubMed:
28026915
Bishop, Y. M. M., Fienberg, S. E., & Holland, P. W. (1975).
Discrete multivariate analysis: Theory and practice.
Cambridge, MA: MIT Press.
Bonnici, H. M., Richter, F. R., Yazar, Y., & Simons, J. S. (2021).
Multimodal feature integration in the angular gyrus during
episodic and semantic retrieval. Journal of Neuroscience, 36,
5462–5471. https://doi.org/10.1523/JNEUROSCI.4310-15
.2016, PubMed: 27194327
Bourgeois, A., Sterpenich, V., Iannotti, G. R., & Vuilleumier, P.
(2022). Reward-driven modulation of spatial attention in
the human frontal eye-field. Neuroimage, 247, 118846.
https://doi.org/10.1016/j.neuroimage.2021.118846, PubMed:
34942365
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436. https://doi.org/10.1163/156856897X00357,
PubMed: 9176952
Academy of Sciences, U.S.A., 113, 13251–13256. https://doi
.org/10.1073/pnas.1605719113, PubMed: 27803331
Bunsey, M., & Eichenbaum, H. B. (1996). Conservation of
hippocampal memory function in rats and humans. Nature,
379, 255–257. https://doi.org/10.1038/379255a0, PubMed:
8538790
Burton, R. L., Lek, I., & Caplan, J. B. (2017). Associative
independence revisited: Competition between conflicting
associations can be resolved or even reversed in one trial.
Quarterly Journal of Experimental Psychology, 70, 832–857.
https://doi.org/10.1080/17470218.2016.1171886, PubMed:
27112421
Bussey, T. J., & Saksida, L. M. (2002). The organization of visual
object representations: A connectionist model of effects
of lesions in perirhinal cortex. European Journal of
Neuroscience, 15, 355–364. https://doi.org/10.1046/j.0953
-816x.2001.01850.x, PubMed: 11849301
Caplan, J. B. (2005). Associative isolation: Unifying associative
and order paradigms. Journal of Mathematical Psychology,
49, 383–402. https://doi.org/10.1016/j.jmp.2005.06.004
Caplan, J. B., & Madan, C. R. (2016). Word-imageability
enhances association-memory by increasing hippocampal
engagement. Journal of Cognitive Neuroscience, 28,
1522–1538. https://doi.org/10.1162/jocn_a_00992, PubMed:
27315268
Caplan, J. B., Rehani, M., & Andrews, J. C. (2014). Associations
compete directly in memory. Quarterly Journal of
Experimental Psychology, 67, 955–978. https://doi.org/10
.1080/17470218.2013.838591, PubMed: 24131316
Caplan, J. B., Sommer, T., Madan, C. R., & Fujiwara, E. (2019).
Reduced associative memory for negative information:
Impact of confidence and interactive imagery during study.
Cognition and Emotion, 33, 1745–1753. https://doi.org/10
.1080/02699931.2019.1602028, PubMed: 30990113
Chanales, A. J. H., Dudukovic, N. M., Richter, F. R., & Kuhl, B. A.
(2019). Interference between overlapping memories is
predicted by neural states during learning. Nature
Communications, 10, 5363. https://doi.org/10.1038/s41467
-019-13377-x, PubMed: 31767880
Clos, M., Schwarze, U., Gluth, S., Bunzeck, N., & Sommer, T.
(2015). Goal- and retrieval-dependent activity in the striatum
during memory recognition. Neuropsychologia, 72, 1–11.
https://doi.org/10.1016/j.neuropsychologia.2015.04.011,
PubMed: 25868676
Cohen, N. J., Poldrack, R. A., & Eichenbaum, H. (1997). Memory
for items and memory for relations in the procedural/
declarative memory framework. Memory, 5, 131–178. https://
doi.org/10.1080/741941149, PubMed: 9156097
Cowell, R. A., Barense, M. D., & Sadil, P. S. (2019). A roadmap
for understanding memory: Decomposing cognitive
processes into operations and representations. eNeuro, 6,
1–19. https://doi.org/10.1523/ENEURO.0122-19.2019,
PubMed: 31189554
Cowell, R. A., Bussey, T. J., & Saksida, L. M. (2010). Functional
dissociations within the ventral object processing pathway:
Cognitive modules or a hierarchical continuum? Journal of
Cognitive Neuroscience, 22, 2460–2479. https://doi.org/10
.1162/jocn.2009.21373, PubMed: 19929757
Davachi, L. (2006). Item, context and relational episodic
encoding in humans. Current Opinion in Neurobiology,
16, 693–700. https://doi.org/10.1016/j.conb.2006.10.012,
PubMed: 17097284
Delprato, D. J. (1972). Pair-specific effects in retroactive
inhibition. Journal of Verbal Learning and Verbal Behavior,
11, 566–572. https://doi.org/10.1016/S0022-5371(72)80040-9
Brodt, S., Pöhlchen, D., Flanagin, V. L., Glasauer, S., Gais, S., &
Dimsdale-Zucker, H. R., & Ranganath, C. (2018).
Schönauer, M. (2016). Rapid and independent memory
formation in the parietal cortex. Proceedings of the National
Representational similarity analyses: A practical guide for
functional MRI applications. In D. Manahan-Vaughan (Ed.),
2164
Journal of Cognitive Neuroscience
Volume 34, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Handbook of behavioral neuroscience (pp. 509–525).
Elsevier. https://doi.org/10.1016/B978-0-12-812028-6.00027-6
Dusek, J. A., & Eichenbaum, H. (1997). The hippocampus and
memory for orderly stimulus relations. Proceedings of the
National Academy of Sciences, U.S.A., 94, 7109–7114. https://
doi.org/10.1073/pnas.94.13.7109, PubMed: 9192700
Eichenbaum, H., Yonelinas, A. R., & Ranganath, C. (2007).
The medial temporal lobe and recognition memory.
Annual Review of Neuroscience, 30, 123–152. https://doi.org
/10.1146/annurev.neuro.30.051606.094328, PubMed:
17417939
Favila, S. E., Lee, H., & Kuhl, B. A. (2020). Transforming the
concept of memory reactivation. Trends in Neurosciences,
43, 939–950. https://doi.org/10.1016/j.tins.2020.09.006,
PubMed: 33041061
Fraundorf, S. H., Diaz, M., Finley, J., Lewis, M. L., Tooley, K. M.,
Isaacs, A. M., et al. (2014). Cogtoolbox for MATLAB
[computer software]. https://www.scottfraundorf.com
/cogtoolbox.html.
Friston, K. J., Buechel, C., Fink, G. R., Morris, J., Rolls, E., &
Dolan, R. J. (1997). Psychophysiological and modulatory
interactions in neuroimaging. Neuroimage, 6, 218–229.
https://doi.org/10.1006/nimg.1997.0291, PubMed: 9344826
Fujiwara, E., Madan, C. R., Caplan, J. B., & Sommer, T. (2021).
Emotional arousal impairs association memory: Roles of
prefrontal cortex regions. Learning & Memory, 28, 76–81.
https://doi.org/10.1101/lm.052480.120, PubMed: 33593925
Gilboa, A., & Marlatte, H. (2017). Neurobiology of schemas and
schema-mediated memory. Trends in Cognitive Sciences, 21,
618–631. https://doi.org/10.1016/j.tics.2017.04.013, PubMed:
28551107
Greeno, J. G., James, C. T., & DaPolito, F. J. (1971). A cognitive
interpretation of negative transfer and forgetting of
paired associates. Journal of Verbal Learning and Verbal
Behavior, 10, 331–345. https://doi.org/10.1016/S0022-5371
(71)80032-4
Hayman, C. A. G., & Tulving, E. (1989). Contingent dissociation
between recognition and fragment completion: The method
of triangulation. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 15, 228–240. https://doi
.org/10.1037/0278-7393.15.2.228, PubMed: 2522512
Haynes, J.-D., & Rees, G. (2005). Predicting the orientation of
invisible stimuli from activity in human primary visual cortex.
Nature Neuroscience, 8, 686–691. https://doi.org/10.1038
/nn1445, PubMed: 15852013
Heckers, S., Zalesak, M., Weiss, A. P., Ditman, T., & Titone, D.
(2004). Hippocampal activation during transitive inference in
humans. Hippocampus, 14, 153–162. https://doi.org/10.1002
/hipo.10189, PubMed: 15098721
Hendriks, M. H. A., Daniels, N., Pegado, F., & Op de Beeck, H. P.
(2017). The effect of spatial smoothing on representational
similarity in a simple motor paradigm. Frontiers in Psychology,
8, 222. https://doi.org/10.3389/fneur.2017.00222, PubMed:
28611726
Himmer, L., Schönauer, M., Heib, D. P. J., Schabus, M., & Gais,
S. (2019). Rehearsal initiates systems memory consolidation,
sleep makes it last. Science Advances, 5, eaav1695. https://doi
.org/10.1126/sciadv.aav1695, PubMed: 31032406
Hintzman, D. L. (1980). Simpson’s paradox and the analysis of
memory retrieval. Psychological Review, 87, 398–410. https://
doi.org/10.1037/0033-295X.87.4.398
Horner, A. J., Bisby, J. A., Bush, D., Lin, W.-J., & Burgess, N.
(2015). Evidence for holistic episodic recollection via
hippocampal pattern completion. Nature Communications,
6, 7462. https://doi.org/10.1038/ncomms8462, PubMed:
26136141
Horner, A. J., & Burgess, N. (2013). The associative structure of
memory for multi-element events. Journal of Experimental
Psychology: General, 142, 1370–1383. https://doi.org/10.1037
/a0033626, PubMed: 23915127
Horner, A. J., & Burgess, N. (2014). Pattern completion in
multielement event engrams. Current Biology, 24, 988–992.
https://doi.org/10.1016/j.cub.2014.03.012, PubMed: 24746796
Horowitz, L. M., Norman, S. A., & Day, R. S. (1966). Availability
and associative symmetry. Psychological Review, 73, 1–15.
https://doi.org/10.1037/h0022661, PubMed: 5324566
Huijbers, W., Vannini, P., Sperling, R. A., Pennartz, C. M.,
Cabeza, R., & Daselaar, S. M. (2012). Memory-related
deactivations and activations in the posteromedial cortex.
Neuropsychologia, 50, 3764–3774. https://doi.org/10.1016/j
.neuropsychologia.2012.08.021, PubMed: 22982484
Humphreys, G. F., Ralph, M. A. L., & Simons, J. S. (2021). A
unifying account of angular gyrus contributions to episodic
and semantic cognition. Trends in Neurosciences, 44,
452–463. https://doi.org/10.1016/j.tins.2021.01.006, PubMed:
33612312
Jacoby, L. L., & Wahlheim, C. N. (2013). On the importance of
looking back: The role of recursive remindings in recency
judgments and cued recall. Memory & Cognition, 41, 625–637.
https://doi.org/10.3758/s13421-013-0298-5, PubMed: 23371792
Kahana, M. J. (2002). Associative symmetry and memory theory.
Memory & Cognition, 30, 823–840. https://doi.org/10.3758
/BF03195769, PubMed: 12450087
Kim, H. (2011). Neural activity that predicts subsequent
memory and forgetting: A meta-analysis of 74 fMRI studies.
Neuroimage, 54, 2446–2461. https://doi.org/10.1016/j
.neuroimage.2010.09.045, PubMed: 20869446
Kleiner, M., Brainard, D., Pelli, D., Ingling, A., Murray, R., &
Broussard, C. (2007). What’s new in Psychtoolbox-3?
Perception, 36, 1–16.
Kliegl, O., & Bäuml, K.-H. T. (2021). Buildup and release from
proactive interference—Cognitive and neural mechanisms.
Neuroscience and Biobehavioral Reviews, 120, 264–278.
https://doi.org/10.1016/j.neubiorev.2020.10.028, PubMed:
33221329
Koen, J. D., & Rugg, M. D. (2016). Memory reactivation predicts
resistance to retroactive interference: Evidence from
multivariate classification and pattern similarity analyses.
Journal of Neuroscience, 36, 4389–4399. https://doi.org/10
.1523/JNEUROSCI.4099-15.2016, PubMed: 27076433
Konkel, A., & Cohen, N. J. (2009). Relational memory and the
hippocampus: Representations and methods. Frontiers in
Neuroscience, 3, 166–174. https://doi.org/10.3389/neuro.01
.023.2009, PubMed: 20011138
Koster, R., Chadwick, M. J., Chen, Y., Berron, D., Banino, A.,
Düzel, E., et al. (2018). Big-loop recurrence within the
hippocampal system supports integration of information
across episodes. Neuron, 99, 1342–1354. https://doi.org/10
.1016/j.neuron.2018.08.009, PubMed: 30236285
Kriegeskorte, N., Cusack, R., & Bandettini, P. (2010). How
does an fMRI voxel sample the neuronal activity pattern:
Compact-kernel or complex spatiotemporal filter?
Neuroimage, 49, 1965–1976. https://doi.org/10.1016/j
.neuroimage.2009.09.059, PubMed: 19800408
Kriegeskorte, N., Mur, M., & Bandettini, P. (2008).
Representational similarity analysis—Connecting the
branches of systems neuroscience. Frontiers in Systems
Neuroscience, 2, 4. https://doi.org/10.3389/neuro.06.004
.2008, PubMed: 19104670
Kuhl, B. A., Rissman, J., Chun, M. M., & Wagner, A. D. (2011).
Fidelity of neural reactivation reveals competition between
memories. Proceedings of the National Academy of
Sciences, U.S.A., 108, 5903–5908. https://doi.org/10.1073/pnas
.1016939108, PubMed: 21436044
Kuhl, B. A., Shah, A. T., DuBrow, S., & Wagner, A. D.
(2010). Resistance to forgetting associated with
Caplan, Hennies, and Sommer
2165
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
hippocampus-mediated reactivation during new learning.
Nature Neuroscience, 13, 501–506. https://doi.org/10.1038
/nn.2498, PubMed: 20190745
Kumaran, D., Summerfield, J. J., Hassabis, D., & Maguire, E. A.
(2009). Tracking the emergence of conceptual knowledge
during human decision making. Neuron, 63, 889–901.
https://doi.org/10.1016/j.neuron.2009.07.030, PubMed:
19778516
Lee, S.-H., Kravitz, D. J., & Baker, C. I. (2011). Disentangling
visual imagery and perception of real-world objects.
Neuroimage, 59, 4064–4073. https://doi.org/10.1016/j
.neuroimage.2011.10.055, PubMed: 22040738
Liu, Z.-X., Grady, C., & Moscovitch, M. (2017). Effects of
prior-knowledge on brain activation and connectivity
during associative memory encoding. Cerebral Cortex, 27,
1991–2009. https://doi.org/10.1093/cercor/bhw047, PubMed:
26941384
Long, N. M., & Kuhl, B. A. (2019). Decoding the tradeoff
between encoding and retrieval to predict memory for
overlapping events. Neuroimage, 201, 116001. https://doi.org
/10.1016/j.neuroimage.2019.07.014, PubMed: 31299369
Mackey, S., & Petrides, M. (2014). Architecture and morphology
of the human ventromedial prefrontal cortex. European
Journal of Neuroscience, 40, 2777–2796. https://doi.org/10
.1111/ejn.12654, PubMed: 25123211
Madan, C. R., Fujiwara, E., Caplana, J. B., & Sommer, T. (2017).
Emotional arousal impairs association-memory: Roles of
amygdala and hippocampus. Neuroimage, 156, 14–28.
https://doi.org/10.1016/j.neuroimage.2017.04.065, PubMed:
28483720
Marr, D. (1971). Simple memory: A theory for archicortex.
Philosophical Transactions of the Royal Society of London,
Series B: Biological Sciences, 262, 23–81. https://doi.org/10
.1098/rstb.1971.0078, PubMed: 4399412
Martin, E. (1971a). Stimulus component independence.
Journal of Verbal Learning and Verbal Behavior, 10,
715–721. https://doi.org/10.1016/S0022-5371(71)80080-4
Martin, E. (1971b). Verbal learning theory and independent
retrieval phenomena. Psychological Review, 78, 314–332.
https://doi.org/10.1037/h0031030
Mayes, A. R., Montaldi, D., & Migo, E. (2007). Associative
memory and the medial temporal lobes. Trends in Cognitive
Sciences, 11, 126–135. https://doi.org/10.1016/j.tics.2006.12
.003, PubMed: 17270487
Mumford, J. A., Turner, B. O., Ashby, F. G., & Poldrack, R. A.
(2012). Deconvolving BOLD activation in event-related
designs for multivoxel pattern classification analyses.
Neuroimage, 59, 2636–2643. https://doi.org/10.1016/j
.neuroimage.2011.08.076, PubMed: 21924359
Nadel, L., & Moscovitch, M. (1997). Memory consolidation,
retrograde amnesia and the hippocampal complex. Current
Opinion in Neurobiology, 7, 217–227. https://doi.org/10.1016
/S0959-4388(97)80010-4, PubMed: 9142752
Norman, K. A., & O’Reilly, R. C. (2003). Modeling hippocampal
and neocortical contributions to recognition memory: A
complementary-learning-systems approach. Psychological
Review, 110, 611–646. https://doi.org/10.1037/0033-295X.110
.4.611, PubMed: 14599236
O’Keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive
map. Oxford University Press.
Op de Beeck, H. P. (2010). Against hyperacuity in brain reading:
Spatial smoothing does not hurt multivariate fMRI analyses?
Neuroimage, 49, 1943–1948. https://doi.org/10.1016/j
.neuroimage.2009.02.047, PubMed: 19285144
O’Reilly, R. C., & McClelland, J. L. (1994). Hippocampal
conjunctive encoding, storage and recall: Avoiding a tradeoff.
Hippocampus, 4, 661–682. https://doi.org/10.1002/hipo
.450040605, PubMed: 7704110
Pelli, D. G. (1997). The VideoToolbox software for visual
psychophysics: Transforming numbers into movies. Spatial
Vision, 10, 437–442. https://doi.org/10.1163
/156856897X00366, PubMed: 9176953
Phillips, S., & Niki, K. (2002). Separating relational from item
load effects in paired recognition: Temporoparietal and
middle frontal gyral activity with increased associates, but
not items during encoding and retention. Neuroimage, 17,
1031–1055. https://doi.org/10.1006/nimg.2002.1190, PubMed:
12377177
Poppenk, J., Evensmoen, H. R., Moscovitch, M., & Nadel, L.
(2013). Long-axis specialization of the human hippocampus.
Trends in Cognitive Sciences, 17, 230–240. https://doi.org/10
.1016/j.tics.2013.03.005, PubMed: 23597720
Postman, L., Stark, K., & Fraser, J. (1968). Temporal changes
in interference. Journal of Verbal Learning and Verbal
Behavior, 7, 672–694. https://doi.org/10.1016/S0022-5371(68)
80124-0
Preston, A. R., Shrager, Y., Dudukovic, N. M., & Gabrieli, J. D. E.
(2004). Hippocampal contribution to the novel use of
relational information in declarative memory. Hippocampus,
14, 148–152. https://doi.org/10.1002/hipo.20009, PubMed:
15098720
Primoff, E. (1938). Backward and forward associations as an
organizing act in serial and in paired associate learning.
Journal of Psychology, 5, 375–395. https://doi.org/10.1080
/00223980.1938.9917578
Rehani, M., & Caplan, J. B. (2011). Interference and the
representation of order within associations. Quarterly
Journal of Experimental Psychology, 64, 1409–1429. https://
doi.org/10.1080/17470218.2010.549945, PubMed: 21598202
Richter, F. R., Chanales, A. J. H., & Kuhl, B. A. (2016). Predicting
the integration of overlapping memories by decoding
mnemonic processing states during learning. Neuroimage,
124, 323–335. https://doi.org/10.1016/j.neuroimage.2015.08
.051, PubMed: 26327243
Rizzuto, D. S., & Kahana, M. J. (2000). Associative symmetry vs.
independent associations. Neurocomputing, 32–33, 973–978.
https://doi.org/10.1016/S0925-2312(00)00268-X
Rizzuto, D. S., & Kahana, M. J. (2001). An autoassociative neural
network model of paired-associate learning. Neural
Computation, 13, 2075–2092. https://doi.org/10.1162
/089976601750399317, PubMed: 11516358
Rossion, B., & Pourtois, G. (2004). Revisiting Snodgrass and
Vanderwart’s object pictorial set: The role of surface detail in
basic-level object recognition. Perception, 33, 217–236.
https://doi.org/10.1068/p5117, PubMed: 15109163
Rudy, J. W., & O’Reilly, R. C. (2001). Conjunctive representations,
the hippocampus, and contextual fear conditioning.
Cognitive, Affective, & Behavioral Neuroscience, 1, 66–82.
https://doi.org/10.3758/CABN.1.1.66, PubMed: 12467104
Rudy, J. W., & Sutherland, R. J. (1989). The hippocampal
formation is necessary for rats to learn and remember
configurational discriminations. Behavioural Brain
Research, 34, 97–109. https://doi.org/10.1016/S0166-4328(89)
80093-2, PubMed: 2765175
Saksida, L. M., & Bussey, T. J. (2010). The representational–
hierarchical view of amnesia: Translation from animal to
human. Neuropsychologia, 48, 2370–2384. https://doi.org/10
.1016/j.neuropsychologia.2010.02.026, PubMed: 20206190
Schwarze, U., Bingel, U., Badre, D., & Sommer, T. (2013).
Ventral striatal activity correlates with memory confidence
for old- and new-responses in a difficult recognition test.
PLoS One, 8, e54324. https://doi.org/10.1371/journal.pone
.0054324, PubMed: 23472064
Smith, R. E., & Hunt, R. R. (2000). The effects of distinctiveness
require reinstatement of organization: The importance of
intentional memory instructions. Journal of Memory and
2166
Journal of Cognitive Neuroscience
Volume 34, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Language, 43, 431–446. https://doi.org/10.1006/jmla.2000
.2707
Sommer, T. (2017). The emergence of knowledge and how it
supports the memory for novel related information. Cerebral
Cortex, 27, 1906–1921. https://doi.org/10.1093/cercor
/bhw031, PubMed: 26908636
Sommer, T., Hennies, N., Lewis, P. A., & Alink, A. (2022). The
assimilation of novel information into schemata and its
efficient consolidation. Journal of Neuroscience, 42,
5916–5929. https://doi.org/10.1523/JNEUROSCI.2373-21
.2022, PubMed: 35710624
Sommer, T., Rose, M., & Büchel, C. (2007). Associative
symmetry versus independent associations in the memory
for object–location associations. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 33, 90–106.
https://doi.org/10.1037/0278-7393.33.1.90, PubMed: 17201555
Sommer, T., Schoell, E., & Büchel, C. (2008). Associative
symmetry of the memory for object–location associations
as revealed by the testing effect. Acta Psychologica, 128,
238–248. https://doi.org/10.1016/j.actpsy.2008.01.003,
PubMed: 18289507
Spalding, K. N., Schlichting, M. L., Zeithamova, D., Preston, A. R.,
Duff, M. C., Tranel, D., et al. (2018). Ventromedial prefrontal
cortex is necessary for normal associative inference and
memory integration. Journal of Neuroscience, 38, 3767–3775.
https://doi.org/10.1523/JNEUROSCI.2501-17.2018, PubMed:
29555854
Staresina, B. P., Alink, A., Kriegeskorte, N., & Henson, R. N.
(2013). Awake reactivation predicts memory in humans.
Proceedings of the National Academy of Sciences, U.S.A.,
110, 21159–21164. https://doi.org/10.1073/pnas.1311989110,
PubMed: 24324174
Stark, C. E. L., & Squire, L. R. (2001). When zero is not zero:
The problem of ambiguous baseline conditions in fMRI.
Proceedings of the National Academy of Sciences, U.S.A.,
98, 12760–12766. https://doi.org/10.1073/pnas.221462998,
PubMed: 11592989
Tulving, E., & Watkins, M. J. (1974). On negative transfer:
Effects of testing one list on the recall of another. Journal of
Verbal Learning and Verbal Behavior, 13, 181–193. https://
doi.org/10.1016/S0022-5371(74)80043-5
Underwood, B. J., & Schulz, R. W. (1960). Response dominance
and rate of learning paired associates. Journal of General
Psychology, 62, 153–158. https://doi.org/10.1080/00221309
.1960.9920406, PubMed: 13840364
van Kesteren, M. T. R., Rignanese, P., Gianferrara, P. G.,
Krabbendam, L., & Meeter, M. (2020). Congruency and
reactivation aid memory integration through reinstatement
of prior knowledge. Scientific Reports, 10, 4776. https://doi
.org/10.1038/s41598-020-61737-1, PubMed: 32179822
Wagner, I. C., van Buuren, M., Bovy, L., & Fernández, G. (2016).
Parallel engagement of regions associated with encoding and
later retrieval forms durable memories. Journal of
Neuroscience, 36, 7985–7995. https://doi.org/10.1523
/JNEUROSCI.0830-16.2016, PubMed: 27466342
Wahlheim, C. N., & Jacoby, L. L. (2013). Remembering change:
The critical role of recursive remindings in proactive effects
of memory. Memory & Cognition, 41, 1–15. https://doi.org/10
.3758/s13421-012-0246-9, PubMed: 22918874
Wahlheim, C. N., Maddox, G. B., & Jacoby, L. L. (2014). The role
of reminding in the effects of spaced repetitions on cued
recall: Sufficient but not necessary. Memory & Cognition, 40,
94–105. https://doi.org/10.1037/a0034055, PubMed: 23937236
Wichawut, C., & Martin, E. (1971). Independence of A-B and
A-C associations in retroaction. Journal of Verbal Learning
and Verbal Behavior, 10, 316–321. https://doi.org/10.1016
/S0022-5371(71)80061-0
Wimber, M., Bäuml, K.-H., Bergström, Z., Markopoulos, G.,
Heinze, H.-J., & Richardson-Klavehn, A. (2008). Neural
markers of inhibition in human memory retrieval. Journal
of Neuroscience, 28, 13419–13427. https://doi.org/10.1523
/JNEUROSCI.1916-08.2008, PubMed: 19074015
Xie, Y. J., Li, Y. Y., Xie, B., Xu, Y. Y., & Peng, L. (2019). The
neural basis of complex audiovisual objects maintenances in
working memory. Neuropsychologia, 133, 107189. https://doi
.org/10.1016/j.neuropsychologia.2019.107189, PubMed:
31513808
Yushkevich, P. A., Piven, J., Hazlett, H. C., Smith, R. G., Ho, S.,
Gee, J. C., et al. (2006). User-guided 3D active contour
segmentation of anatomical structures: Significantly improved
efficiency and reliability. Neuroimage, 31, 1116–1128. https://
doi.org/10.1016/j.neuroimage.2006.01.015, PubMed: 16545965
Zeithamova, D., & Bowman, C. R. (2020). Generalization and
the hippocampus: More than one story? Neurobiology of
Learning and Memory, 175, 107317. https://doi.org/10.1016/j
.nlm.2020.107317, PubMed: 33007461
Zeithamova, D., Dominick, A. L., & Preston, A. R. (2012).
Hippocampal and ventral medial prefrontal activation during
retrieval-mediated learning supports novel inference.
Neuron, 75, 168–179. https://doi.org/10.1016/j.neuron.2012
.05.010, PubMed: 22794270
Zeithamova, D., & Preston, A. R. (2010). Flexible memories:
Differential roles for medial temporal lobe and prefrontal
cortex in cross-episode binding. Journal of Neuroscience, 30,
14676–14684. https://doi.org/10.1523/JNEUROSCI.3250-10
.2010, PubMed: 21048124
Caplan, Hennies, and Sommer
2167
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
1
2
1
4
4
2
0
4
8
4
8
1
/
/
j
o
c
n
_
a
_
0
1
9
0
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3