Coming Unbound: Disrupting Automatic Integration
of Synesthetic Color and Graphemes by Transcranial
Magnetic Stimulation of the Right Parietal Lobe
Michael Esterman1,2, Timothy Verstynen1, Richard B.. Ivry1,
and Lynn C. Robertson1,2
Abstracto
& En algunos individuos, a visually presented letter or number
automatically evokes the perception of a specific color, an ex-
perience known as color–grapheme synesthesia. It has been
suggested that parietal binding mechanisms play a role in the
fenómeno. We used a noninvasive stimulation technique,
transcranial magnetic stimulation (TMS), to determine whether
the posterior parietal lobe is critical for the integration of color
and shape in color–grapheme synesthesia, as it appears to be
for normal color–shape binding. Using a color-naming task with
colored letters that were either congruent or incongruent with
the synesthetic photism, we demonstrate that inhibition of
the right posterior parietal lobe with repetitive TMS transiently
attenuates synesthetic binding. These findings suggest that syn-
esthesia (the induction of color from shape) relies on similar
mechanisms as found in normal perception (where the percep-
tion of color is induced by wavelength). &
INTRODUCCIÓN
In color–grapheme synesthesia, specific graphemes
are automatically seen in specific colors. Por ejemplo,
the letter ‘‘A’’ is always perceived in a particular shade
of red. This unusual variety of color–form binding
has been well characterized behaviorally. Sin embargo, el
neural correlates of synesthesia are not well under-
permaneció (Rich & Mattingley, 2002). Although functional
magnetic resonance imaging (resonancia magnética funcional) has shown that
increased activity in extrastriate cortex is associated
with the perception of synesthetic photisms (Sperling,
Prvulovic, Linden, Cantante, & Stirn, 2006; Hubbard, Arman,
Ramachandran, & Boynton, 2005), other brain imaging
studies of color–grapheme synesthesia have shown neu-
ral correlates outside of the ventral occipital cortex in
the posterior parietal cortex (PPC). Rich et al. (2003)
found activation in the right PPC (at the junction of
the occipital cortex) during synesthetic color–grapheme
percepción, an area implicated in normal binding of
color and shape (Donner et al., 2002). Others have
found activation in the intraparietal sulcus (IPS) associ-
ated with color–grapheme synesthesia, both in the left
hemisferio (Weiss, Zilles, & Fink, 2005) and bilaterally
(Elías, Saucier, Hardie, & Sarty, 2003).
The PPC has already been associated with color–form
binding in normal perception using tasks that involve con-
1Universidad de California, berkeley, 2Veterans Administration
Centro Médico, Martinez, California
junction visual search (Donner et al., 2002; Ashbridge,
Walsh, & Cowey, 1997). Además, patients with bilateral
PPC lesions have difficulty in correctly conjoining shape
and color. Por ejemplo, R.M., a patient with Balint’s syn-
drome, makes frequent illusory conjunctions in free view-
ing conditions (Friedman-Hill, Robertson, & Treisman,
1995). Juntos, these findings suggest that although syn-
esthesia is an atypical integration of color and shape, él
may rely on similar mechanisms to normal perceptual fea-
ture binding (Cohen-Kadosh & Henik, 2006; Sagiv, Heer,
& Robertson, 2006; Robertson, 2003; Grossenbacher &
Lovelace, 2001).
Further evidence that parietal mechanisms may be
involved in synesthesia come from behavioral studies
demonstrating that binding of the synesthetic photism
requires awareness of the inducing grapheme and is
modulated by whether the grapheme is within the
spotlight of attention (Sagiv et al., 2006; Palmeri, Blake,
Marois, Flanery, & Whetsell, 2002; Mattingley, Rich,
Yelland, & Bradshaw, 2001). The idea that synesthesia
arises from operations of an attentional binding mech-
anism is challenged by findings indicating preattentive
activation of synesthetic percepts (Smilek, dixon, &
Merikle, 2004; Ramachandran & Hubbard, 2001). Estos
contradictory results may be explained in part by indi-
vidual differences between synesthetes (Hubbard &
Ramachandran, 2005).
We sought to test the hypothesis that parietal binding
mechanisms that are necessary for normal perception
also play a role in color–grapheme synesthesia. To this
Revista de neurociencia cognitiva 18:9, páginas. 1570–1576
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
9
–
1
pag
5
d
7
F
0
/
1
1
8
9
/
3
5
9
9
/
7
1
8
5
7
oh
0
C
/
norte
1
2
7
0
5
0
6
6
3
1
5
8
1
/
9
j
1
oh
5
C
7
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
9
t
.
oh
1
norte
5
0
7
8
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
F
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
end, we used a color-naming task (Mattingley et al.,
2001; dixon, Smilek, Cudahy, & Merikle, 2000). On
each trial, a colored letter was presented in one of
three colors and the participant pressed a key to indi-
cate the color. The color was either congruent or
incongruent with the synesthetic photism (Figura 1B).
Synesthetic facilitation occurs when responses to let-
ters presented in their synesthetic color (congruente
condición) are faster than baseline responses (neutral
characters that do not evoke a synesthetic photism).
Synesthetic interference occurs when responses to let-
ters presented in colors that are incongruent with
their synesthetic photism (incongruent condition) son
slower than baseline. This task was performed follow-
ing transcranial magnetic stimulation (TMS) del
left or right parietal cortex. This method allowed us to
investigate whether transient inhibition of the parietal
cortex influenced synesthetic induction of color by
forma. Específicamente, would parietal stimulation reduce
any photism-induced facilitation or interference? Semejante
changes would be consistent with this region contribut-
ing to binding of color and shape in synesthesia.
We targeted the angular gyrus at the junction of the
posterior IPS and transverse occipital sulcus (IPS/TOS),
a region associated with color–form binding in normal
percepción (Donner et al., 2002). Además, activación
in the right IPS/TOS has been observed during synes-
thetic color–grapheme perception (Rich et al., 2003),
suggesting a correspondence between normal and syn-
esthetic binding of color and form. In separate sessions,
stimulation was applied over this parietal region of the
right and left hemisphere. In another session, repetitive
TMS (rTMS) was applied over the primary visual cortex
(V1) to test for the effects of generalized brain stimula-
tion on responses in this task.
MÉTODOS
Participantes
All procedures for this experiment were approved by the
local ethical review board at University of California at
berkeley. We tested two color–grapheme synesthetes
recruited from the local population at the university:
C.P. (27 years old) and E.F. (22 years old), both right-
handed women. Both report having a unique set of
alphanumeric–color associations (ver Figura 1A) that are
stable over time. Both reported their colors were pro-
jected in the external world and appeared as a ‘‘property’’
of the inducing character, which was also in the ‘‘mind’s
eye.’’ Both synesthetes would be classified as ‘‘projectors’’
(Sagiv et al., 2006; dixon, Smilek, & Merikle, 2004). Tiene
been shown in a prior behavioral study that attention
modulates the extent of C.P.’s synesthetic experience
(Sagiv et al., 2006). Además, her experiences were pre-
viously associated with modest activation of lower visual
areas,
leading to her classification as a ‘‘higher synes-
thete’’ (see participant C.H.P. in Hubbard et al., 2005).
We did not test participants with normal perception
in this study because they have no synesthetic photism
to influence color naming (see behavioral procedures
described below) and training controls with color–
grapheme associations would not replicate the experi-
ence of synesthesia.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
9
–
1
pag
5
d
7
F
0
/
1
1
8
9
/
3
5
9
9
/
7
1
8
5
7
oh
0
C
/
norte
1
2
7
0
5
0
6
6
3
1
5
8
1
/
9
j
1
oh
5
C
7
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
9
t
.
oh
1
norte
5
0
7
8
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
F
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Esterman et al.
1571
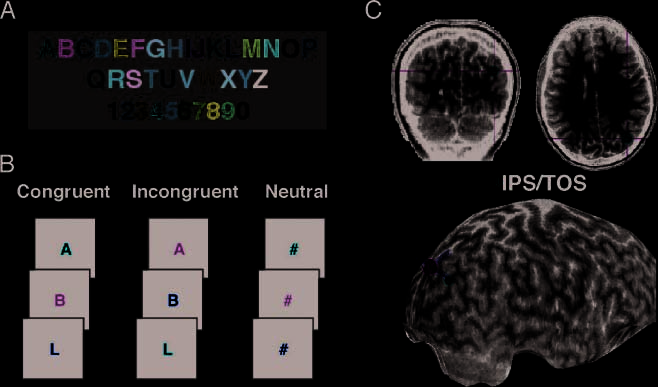
Cifra 1. Behavioral and TMS
procedure for participant E.F.
(A) Synesthetic alphabet. (B)
Individualized experimental
condiciones. The task was to
name the stimulus color. En
the congruent condition, el
stimulus color matched the
synesthetic color. En el
incongruent condition, el
stimulus color did not match
the synesthetic color. Neutral
characters did not evoke a
synesthetic color. (C) Target
stimulation site (right IPS/
TOS) shown on coronal
and axial slices, plus a 3-D
reconstruction of the
anatomical image for E.F.
Target location is shown
in red, center of magnetic
coil is shown as green
spheres in the 3-D image,
and estimated pulse and
orthogonal trajectories
are shown as yellow lines.
Behavioral Procedure
Before testing, we estimated the specific RGB screen
values for each letter of each participant’s synesthetic
alphabet. They were seated comfortably (cid:1)30 cm in front
of a 19-in. CRT monitor (70 Hz refresh rate). An alpha-
numeric symbol was presented on the screen and the
participant was instructed to adjust the RGB values until
the color on the screen matched their synesthetic
photism. For testing, we chose those letters that evoked
the most red, verde, and yellow color associations,
respectivamente. Each shade of red, verde, and yellow was
chosen to closely match the synesthetic photism.
To quantify each participant’s synesthesia we used a
color-naming task. A series of colored single letters (rojo,
verde, or yellow; see above) were presented, each for
1000 mseg, in a color that was either congruent (estafa-
gruent trials) or incongruent (incongruent trials) con
the synesthetic photism, or a symbol (‘‘#’’) was pre-
sented that did not evoke a synesthetic photism (neutral
ensayos; see Figure 1B for examples). Participants pressed a
button with their right hand indicating the true color of
the letter as quickly as possible while trying to ignore the
synesthetic photism. Reaction time to manually respond
to the screen color was recorded as the difference
between the onset of the letter and the subsequent
keypress. There was no deadline to respond. Following
a response, there was a 1000 msec intertrial interval.
Stimuli were presented with PsychLab software (Teren
Gum, Bostón, Virginia), and recorded with a CMU button box
(three buttons pressed with three different fingers).
Each block consisted of 120 ensayos (48 congruente, 48
incongruente, y 24 neutral, representing all combina-
tions of colors and characters). After each stimulation
epoch (sham rTMS or real rTMS; see below), participar-
pants were tested on two blocks of trials: an early block
(1–5 min poststimulation) and a late block (6–10 min
poststimulation). Order of sham and real TMS was
counterbalanced for each participant. Each sham and
real TMS epoch was performed twice in each session,
alternating between stimulation types. Separate sessions
were done for each target region (right parietal,
izquierda
parietal, V1). For each session, 960 trials were collected
per participant.
Transcranial Magnetic Stimulation
We first obtained high-resolution anatomical MRIs for
cada participante. The images for C.P. were acquired in a
previous study using a MPRAGE sequence on a 1.5T
Siemens Vision scanner at the University of California,
San Diego (1 (cid:2) 1 (cid:2) 1 isotropic voxels, 180 slices). El
images for E.F. were acquired using a MPFLASH protocol
on a Varian INNOVA 4T system at the University of
California, berkeley (2 (cid:2) 2 (cid:2) 2 isotropic voxels, 128
slices). We identified the target regions for TMS stimu-
lation from each participant’s MRI (left and right IPS/
TOS and V1). The voxel location was marked on a skull-
stripped reconstructed image.
The scalp location was determined by using a stereo-
taxic localization system (Brainsight, Rogue-Research
Cª, Montréal, Canada). Coil position over the target
regions was monitored online during the stimulation
epochs. Además, trajectory estimates of the TMS
pulse were estimated intermittently throughout record-
En g (green dots and yellow lines in Figure 1C). A pesar de
we chose our region of interest (ROI) based on each
individual’s anatomy, we estimated the Talairach coor-
dinates of the right IPS/TOS site as (30, (cid:3)74, 32), close
to the area associated with feature binding (22, (cid:3)71, 27)
in Donner et al. (2002). Talairach space is primarily driv-
en by the need to standardize across individuals and
assumes a standard sulcal and gyral geometry. By nature,
it is insensitive to individual differences in brain struc-
tura. The locations in the present study were identified
by using the pattern of gross anatomical landmarks of
the IPS, angular gyrus, and TOS. De este modo, this coordinate
is only an estimate of our stimulation location.
rTMS was performed by using an iron-cored figure-8
coil (NeoTonus Inc., Marietta, Georgia; see Epstein & Davey,
2002). Before each session, the participant’s active mo-
tor threshold was determined as the point at which four
to six visible twitches of the thumb were detected fol-
mugiendo 10 pulses over the motor cortex while the thumb
and index finger were held together in a pinchlike pos-
tura. C.P.’s threshold was between 35% y 38% maxi-
mum stimulator output, whereas E.F.’s threshold was
consistent at 45%. Stimulation was then set to 115% de
motor threshold for the remainder of the experiment.
Each stimulation epoch consisted of 480 consecutive
pulses that were delivered at a rate of 1 Hz (8 mín.). Este
low-frequency design causes a transient inhibition of the
underlying cortex, with the duration of the effect rough-
ly equal to the duration of the stimulation at 1 Hz
(Pascual-Leone et al., 1998). De este modo, maximal behavioral
effects of TMS were expected during the early block
(minutes 1–5 post stimulation) diminishing during the
late block (minutes 6–10 post stimulation). During rTMS
bloques, the coil was oriented to deliver stimulation
directly to the target cortical location. For sham control
rTMS blocks, the coil was oriented 908 away from the
scalp so that no pulses perturbed underlying neural
tejido. The subjects were naive to whether they were
receiving sham or real stimulation.
Análisis de los datos
Given the small sample size and the propensity for
large between-subjects variability with synesthesia, nosotros
adopted a strong within-subject method to analyze the
stimulation effects. The TMS effects were statistically
analyzed by using randomization methods on a single-
subject basis (p.ej., bootstrapping; see Manly, 1997). Todo
analyses excluded any reaction times greater than four
1572
Revista de neurociencia cognitiva
Volumen 18, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
9
–
1
pag
5
d
7
F
0
/
1
1
8
9
/
3
5
9
9
/
7
1
8
5
7
oh
0
C
/
norte
1
2
7
0
5
0
6
6
3
1
5
8
1
/
9
j
1
oh
5
C
7
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
9
t
.
oh
1
norte
5
0
7
8
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
standard deviations from the mean (less than 1% of trials
for each participant). We first recorded the interference
effect for each of the six conditions (3 regiones (cid:2) 2 rTMS/
sham epochs) by subtracting reaction times to incon-
gruent trials from neutral trials. A ‘‘true’’ TMS effect
puntaje (Xt) was determined by subtracting the interfer-
ence effect observed following sham stimulation from
the effect found after real rTMS. A score of zero indicates
no change in interference following rTMS, a positive
score indicates increased interference following rTMS,
and a negative score represents a decreased interference
effect after rTMS. We then randomly recategorized
reaction times between neutral and incongruent trials
and recalculated a new interference effect. This was done
separately for rTMS and sham trials, following which a
new TMS effect score was calculated. This scrambling,
recategorization, and subtraction process was repeated
para 10,000 iterations to produce a hypothetical probabil-
ity distribution of interference effects that would occur
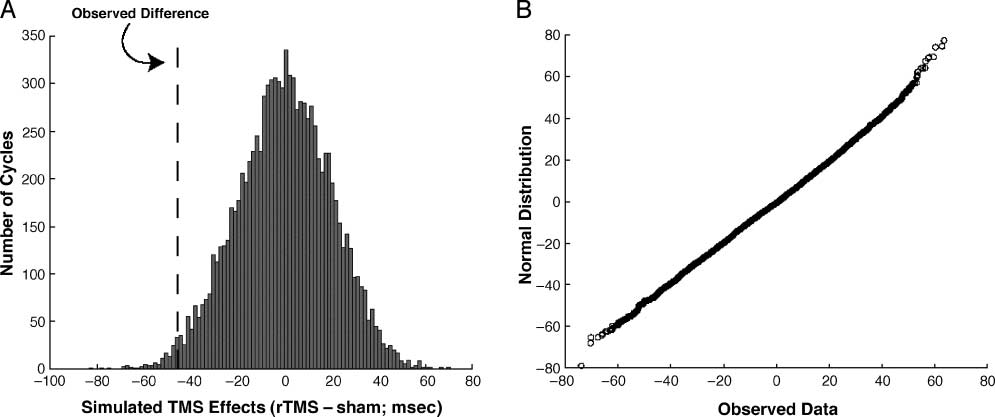
simply by chance. Figure 2A shows an example of a
random distribution of
interference effects for C.P.
following right-hemisphere stimulation. The dashed line
represents her true TMS effect. These distributions
closely resembled normal distributions, as verified using
probability plots (Figura 2B; Chambers, Cleveland,
Kleiner, & Tukey, 1983).
The probability of getting the true observed effects by
chance ( pag) can then be directly calculated as:
p ¼ (cid:1)ðXr > XtÞ
where Xr
randomized null distribution.
is the array of values pulled from the
RESULTS AND DISCUSSION
Following sham rTMS, both participants showed syn-
esthetic interference in all six blocks (E.F.: mean =
51 mseg, range 32–90 msec; C.P.: mean = 31 mseg;
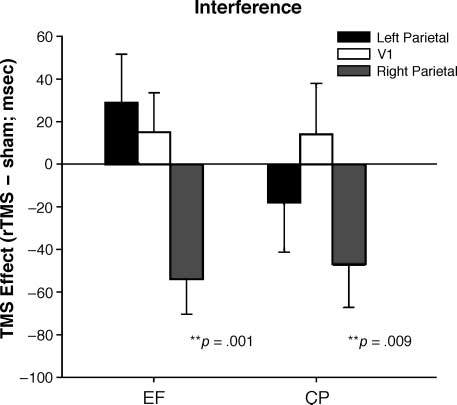
range, 19–48 msec). A diferencia de, rTMS of the right pa-
rietal ROI significantly attenuated interference for both
participants during the early test block (por 54 msec for
E.F. y 48 msec for C.P., pag < .01; see Figure 3). For
E.F., interference returned to baseline levels in the late
block (61 msec); however, C.P.’s scores were still atten-
uated (10 msec), suggesting the effects of rTMS had
not completely worn off. C.P.’s responses were generally
faster overall following real rTMS,
likely due to gen-
eralized arousal. More importantly, the attenuation of
interference seen after right IPS/TOS stimulation is the
result of a disproportionate decrease in reaction times
on incongruent trials (see Table 1).
In contrast to right-hemisphere stimulation, rTMS
over the left PPC had no effect on interference (see
Figure 3; C.P.: p = .22, E.F.: p = .09). Thus, the
disruption of synesthesia following right parietal rTMS
does not generalize to the left-hemisphere homologue.
This is consistent with neurophysiological evidence that
the right parietal cortex plays a more critical role in
normal color–form binding than does the left parietal
cortex (Ashbridge et al., 1997).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
8
e
9
-
1
p
5
d
7
f
0
/
1
1
8
9
/
3
5
9
9
/
7
1
8
5
7
o
0
c
/
n
1
2
7
0
5
0
6
6
3
1
5
8
1
/
9
j
1
o
5
c
7
n
0
.
p
2
d
0
0
b
6
y
.
g
1
u
8
e
.
s
9
t
.
o
1
n
5
0
7
8
0
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Figure 2. (A) A histogram of simulated TMS effects representing a hypothetical null distribution for C.P. after right PPC stimulation. The
observed reaction times were randomly recategorized (with replacement) and new interference effects were calculated from these permutated
data sets. This process was repeated 10,000 times to produce a distribution of values that represent what would be observed purely by chance.
The dashed line represents the true observed TMS effect immediately following stimulation. (B) A normal probability plot, comparing the
data presented in (A) with values from a normal distribution with mean of (cid:3)0.04 msec and standard deviation of 20 msec. The approximately
straight line indicates that the hypothetical null distribution ref lect a normal gaussian process.
Esterman et al.
1573
disruption of synesthetic color–form integration, thus
reducing the conflict when the synesthetic photism
did not correspond to the real color of the letter.
These findings are consistent with evidence implicating
the IPS/TOS in feature binding in normal perception
(Donner et al., 2002; Friedman-Hill et al., 1995) and
suggests that this region contributes to feature binding
even when the perceived color is evoked by shape
rather than wavelength (Robertson, 2003).
The present study supports the theory that feedback
from a multimodal association region, like the parietal
cortex, contributes to the perception of a synesthetic
photism (Cohen-Kadosh & Henik, 2006; Sagiv et al.,
2006; Robertson, 2003; Grossenbacher & Lovelace,
2001). Although this does not exclude the possibility
that direct connections between fusiform regions also
play a role in synesthesia (Ramachandran & Hubbard,
2001), it suggests that cross wiring between these re-
gions is not always sufficient to bind the synesthetic
percept to the inducing character.
An alternative explanation for our findings is that
parietal TMS attenuates competition between perceptu-
ally related codes, similar to competition produced be-
tween the perceptually and conceptually related codes
in the traditional Stroop task where the color of the
word may be congruent or incongruent with the word
itself. According to this view, parietal TMS would inter-
fere with competition between the color word and color
(traditional Stroop) as well as competition between two
colors, one induced by wavelength and the other by the
letter (synesthesia), rather than interfere with feature
binding per se. Although imaging studies have asso-
ciated parietal activity with Stroop tasks (MacLeod &
MacDonald, 2000), it is unlikely that the parietal lobe
plays a critical role in the competitive process underlying
the traditional Stroop effect for several reasons. First,
bilateral stimulation of the posterior parietal lobes does
not influence the magnitude of Stroop interference or
facilitation (Hayward, Goodwin, & Harmer, 2004).
Second, neuropsychological studies of patients with
parietal lesions have demonstrated normal Stroop inter-
ference compared to controls. Interestingly, this effect
has been reported in patients with either left, right, or
bilateral infarcts (Vivas, Humphreys, & Fuentes, 2003;
Robertson, Treisman, Friedman-Hill, & Grabowecky,
1997; Berti, Frassinetti, & Umilta, 1994), and these same
patients may exhibit profound deficits in color–form
binding (see Robertson, 2003). In contrast, alterations
in Stroop performance have been associated with dam-
age to the prefrontal cortex (Kato, 2001; Stuss, Floden,
Alexander, Levine, & Katz, 2001).
Third, the three studies that have implicated the
parietal lobe in color–grapheme synesthesia have not
involved Stroop tasks, indicating that the parietal lobe’s
role in synesthesia is not dependent on response in-
terference (Weiss et al., 2005; Elias et al., 2003; Rich
et al., 2003). Together, these different lines of research
Figure 3. Effects of TMS on synesthetic interference. Graph
displays effects of rTMS on synesthetic interference (incongruent
minus neutral) for both participants immediately following
stimulation (early block; see text). Both participants demonstrated
a strong attenuation of interference following right parietal
compared to sham rTMS, but no change following stimulation
of the left parietal region or V1 (see Methods for discussion of
statistical procedure and Table 1 for reaction times and standard
deviations in all conditions).
rTMS over V1 also had no effect on interference (see
Figure 3; C.P.: p = .28, E.F.: p = .21). Thus, the
disruption of synesthesia following right parietal rTMS
is not due to generalized arousal following stimulation
and suggests that V1 does not play a critical role in
color–form binding.
Unlike interference, facilitation was not reliably ob-
served in our participants. Naming colors that were
synesthetically congruent produced consistently faster
responses for participant E.F only (36 msec). C.P. did
not show consistent facilitation in the sham blocks
(mean = 14 msec; range, –26 to 44 msec). Facilitation
is not reliably observed in traditional Stroop tasks in
normal observers (Tzelgov, Henik, & Berger, 1992;
MacLeod, 1991) as well as tasks similar to ours when
tested in other synesthetes (Sagiv et al., 2006; Mattingley
et al., 2001; Dixon et al., 2000). It has been proposed
that facilitation, when observed in traditional Stroop
tasks, may be due to inadvertent reading, which would
explain our weak facilitation effect, given that ‘‘reading’’
the letter would not lead to correct responses in our
paradigm (MacLeod, 1991). Our results are consistent
with studies of normal perceivers showing that facilita-
tion and interference rely on different mechanisms
(Tzelgov et al., 1992).
In sum, immediately following rTMS of the right PPC,
both participants showed a significant attenuation of
interference normally induced by their synesthesia. We
propose that right parietal rTMS produced a transient
1574
Journal of Cognitive Neuroscience
Volume 18, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
8
e
9
-
1
p
5
d
7
f
0
/
1
1
8
9
/
3
5
9
9
/
7
1
8
5
7
o
0
c
/
n
1
2
7
0
5
0
6
6
3
1
5
8
1
/
9
j
1
o
5
c
7
n
0
.
p
2
d
0
0
b
6
y
.
g
1
u
8
e
.
s
9
t
.
o
1
n
5
0
7
8
0
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Table 1. Single-subject Reaction Times
Participant E.F.
Participant C.P.
Neutral
Congruent
Incongruent
Neutral
Congruent
Incongruent
Right parietal
Early phase
Sham
rTMS
Late phase
Sham
rTMS
Left parietal
Early phase
Sham
rTMS
Late phase
Sham
rTMS
V1
Early phase
Sham
rTMS
Late phase
Sham
rTMS
476 ± 103
481 ± 88
483 ± 86
509 ± 117
424 ± 77
423 ± 58
453 ± 97
441 ± 80
492 ± 120
469 ± 84
434 ± 81
449 ± 93
496 ± 90
471 ± 95
462 ± 94
462 ± 108
452 ± 76
440 ± 99
423 ± 75
443 ± 98
419 ± 70
412 ± 62
413 ± 64
418 ± 74
546 ± 114
497 ± 96
531 ± 92
570 ± 131
523 ± 116
530 ± 141
529 ± 119
532 ± 133
485 ± 91
488 ± 107
514 ± 113
501 ± 84
615 ± 161
550 ± 145
574 ± 158
511 ± 118
548 ± 108
591 ± 166
575 ± 169
546 ± 145
503 ± 99
503 ± 104
499 ± 118
471 ± 96
533 ± 113
495 ± 114
521 ± 111
490 ± 103
591 ± 121
511 ± 106
547 ± 141
486 ± 107
568 ± 117
524 ± 127
556 ± 168
511 ± 132
647 ± 186
535 ± 94
597 ± 185
601 ± 184
544 ± 152
523 ± 134
557 ± 132
536 ± 145
611 ± 156
544 ± 144
587 ± 150
529 ± 152
Shown are mean reaction times and standard deviations across each condition (neutral, congruent, or incongruent), phase (early, late), participant
(E.F., C.P.) and each stimulation site (right parietal, left parietal, V1).
strongly suggest that our effect is not due to response
interference in general.
Behavioral and physiological evidence suggests that
synesthesia is a heterogeneous phenomenon (Dixon &
Smilek, 2005; Hubbard et al., 2005). These individual
differences may be related to the degree of parietal
involvement. One behavioral factor that may contribute
to this heterogeneity is the difference between projec-
tors, those who see the synesthetic photism in the
world, and associators, those who see the photism in
the mind’s eye (Dixon et al., 2004). Although this factor
has not been explored with respect to the degree of
involvement, one hypothesis to consider in
parietal
future research is that synesthetes who show greater
parietal involvement may tend to be ‘‘projectors,’’ par-
ticularly when the synesthetic percept is tightly bound
spatially to the inducing grapheme. Physiologically, the
degree of parietal contribution among synesthetes may
also be greater in individuals such as C.P., in whom the
extent of early visual activation has been shown to be
lesser (see Hubbard et al., 2005). Parietal participation in
the synesthetic experience may correspond to the ex-
tent that attention is required to elicit the photism. The
variability in the expression of color–grapheme synes-
thesia makes the examination of individual differences
vital to our understanding of this fascinating phenome-
non (Dixon & Smilek, 2005; Hubbard et al., 2005; Dixon
et al., 2004). Our results have illuminated one of poten-
tially several mechanisms responsible for synesthesia
and are consistent with theories of how normal binding
occurs within the human brain.
Future studies must examine the role of parietal bind-
ing mechanisms in other synesthetes, as well as investi-
gate other parietal and temporal lobe regions that have
been implicated in spatial attention and binding, such as
the superior parietal cortex and the superior temporal
Esterman et al.
1575
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
8
e
9
-
1
p
5
d
7
f
0
/
1
1
8
9
/
3
5
9
9
/
7
1
8
5
7
o
0
c
/
n
1
2
7
0
5
0
6
6
3
1
5
8
1
/
9
j
1
o
5
c
7
n
0
.
p
2
d
0
0
b
6
y
.
g
1
u
8
e
.
s
9
t
.
o
1
n
5
0
7
8
0
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
sulcus, using both TMS and fMRI. In addition, single-pulse
TMS will be able to elaborate on the temporal dynamics
of the right PPC’s involvement in synesthesia.
Acknowledgments
We thank Noam Sagiv for discussions and motivation that led
to this project, Noam Sobel for comments on an early version
of this manuscript, and Edward Hubbard for his constructive
comments during the review process.
Reprint requests should be sent to Michael Esterman, Depart-
ment of Psychology and Neuroscience Institute, University of Cali-
fornia, Berkeley, CA, USA, or via e-mail: esterman@berkeley.edu.
REFERENCES
Ashbridge, E., Walsh, V., & Cowey, A. (1997). Temporal
aspects of visual search studied by transcranial magnetic
stimulation. Neuropsychologia, 35, 1121–1131.
Berti, A., Frassinetti, F., & Umilta, C. (1994). Nonconscious
MacLeod, C. M. (1991). Half a century of research on the
Stroop effect: An integrative review. Psychological
Bulletin, 109, 163–203.
MacLeod, C. M., & MacDonald, P. A. (2000). Interdimensional
interference in the Stroop effect: Uncovering the
cognitive and neural anatomy of attention. Trends in
Cognitive Sciences, 4, 383–391.
Manly, B. (1997). Randomization, bootstrap and Monte
Carlo methods in biology (2nd ed.). New York: Chapman
and Hall/CRC.
Mattingley, J. B., Rich, A. N., Yelland, G., & Bradshaw, J. L.
(2001). Unconscious priming eliminates automatic binding
of colour and alphanumeric form in synaesthesia. Nature,
410, 580–582.
Palmeri, T. J., Blake, R., Marois, R., Flanery, M. A., & Whetsell,
W. (2002). The perceptual reality of synesthetic colors.
Proceedings of the National Academy of Sciences, U.S.A.,
99, 4127–4131.
Pascual-Leone, A., Tormos, J. M., Keenan, J., Tarazona, F.,
Canete, C., & Catala, M. D. (1998). Study and modulation
of human cortical excitability with transcranial magnetic
stimulation. Journal of Clinical Neurophysiology, 15,
333–343.
reading? Evidence from neglect dyslexia. Cortex, 30, 181–197.
Ramachandran, V. S., & Hubbard, E. M. (2001). Psychophysical
Chambers, J., Cleveland, W., Kleiner, B., & Tukey, P. (1983).
Graphical methods for data analysis (Wadsworth).
Boston: Duxbury Press.
Cohen-Kadosh, R., & Henik, A. (2006). Color congruity
effect: Where do colors and numbers interact in
synesthesia? Cortex, 42, 259–263.
Dixon, M. J., & Smilek, D. (2005). The importance of
individual differences in grapheme–color synesthesia.
Neuron, 45, 821–823.
Dixon, M. J., Smilek, D., Cudahy, C., & Merikle, P. M.
(2000). Five plus two equals yellow. Nature, 406, 365.
Dixon, M. J., Smilek, D., & Merikle, P. M. (2004). Not
all synaesthetes are created equal: Projector versus
associator synaesthetes. Cognitive, Affective, and
Behavioral Neuroscience, 4, 335–343.
Donner, T. H., Kettermann, A., Diesch, E., Ostendorf, F.,
Villringer, A., & Brandt, S. A. (2002). Visual feature and
conjunction searches of equal difficulty engage only partially
overlapping frontoparietal networks. Neuroimage, 15, 16–25.
Elias, L. J., Saucier, D. M., Hardie, C., & Sarty, G. E. (2003).
Dissociating semantic and perceptual components of
synaesthesia: Behavioral and functional neuroanatomical
investigations. Cognitive Brain Research, 16, 232–237.
Epstein, C. M., & Davey, K. R. (2002). Iron-core coils for
transcranial magnetic stimulation. Journal of Clinical
Neurophysiology, 19, 376–381.
Friedman-Hill, S., Robertson, L., & Treisman, A. (1995). Parietal
contributions to visual feature binding: Evidence from a
patient with bilateral lesions. Science, 269, 853–855.
Grossenbacher, P. G., & Lovelace, C. T. (2001). Mechanisms
of synesthesia: Cognitive and physiological constraints.
Trends in Cognitive Sciences, 5, 36–41.
Hayward, G., Goodwin, G. M., & Harmer, C. J. (2004). The
role of the anterior congulate cortex in the counting
Stroop task. Experimental Brain Research, 154, 355–358.
Hubbard, E. M., Arman, A. C., Ramachandran, V. S., &
Boynton, G. M. (2005). Individual differences among
grapheme–color synesthetes: Brain–behavior correlations.
Neuron, 45, 975–985.
Hubbard, E. M., & Ramachandran, V. S. (2005). Neurocognitive
mechanisms of synesthesia. Neuron, 48, 509–520.
Kato, M. (2001). Prefrontal lobes and the attentional control:
A neuropsychological study using modified Stroop test.
Rinsho Shinkeigaku, 41, 1134–1136.
investigations into the neural basis of synaesthesia.
Proceedings of the Royal Society of London, Series B,
Biological Sciences, 268, 979–983.
Rich, A. N., & Mattingley, J. B. (2002). Anomalous perception
in synaesthesia: A cognitive neuroscience perspective.
Nature Reviews, 3, 43–52.
Rich, A. N., Puce, A., Syngeniotis, A., Williams, M. A.,
Howard, M. A., McGlone, F., & Mattingley, J. B. (2003).
Colour my brain: A functional neuroimaging study of
color–graphemic synaesthesia. Paper presented at the
annual meeting of the Cognitive Neuroscience Society,
New York, NY.
Robertson, L., Treisman, A., Friedman-Hill, S., & Grabowecky,
M. (1997). The interaction of spatial and object pathways:
Evidence from Balint’s syndrome. Journal of Cognitive
Neuroscience, 9, 295–317.
Robertson, L. C. (2003). Binding, spatial attention and
perceptual awareness. Nature Reviews Neuroscience, 4,
93–102.
Sagiv, N., Heer, J., & Robertson, L. C. (2006). Does binding
of synesthetic color to the evoking grapheme require
attention? Cortex, 42, 232–242.
Smilek, D., Dixon, M. J., & Merikle, P. M. (2004). Binding of
graphemes and synesthetic colors in color–graphemic
synesthesia. In N. Sagiv & L. Robertson (Eds.), Synesthesia:
Perspectives from cognitive neuroscience. New York:
Oxford University Press.
Sperling, J. M., Prvulovic, D., Linden, D. E. J., Singer, W., &
Stirn, A. (2006). Neuronal correlates of colour–graphemic
synaesthesia: A fMRI study. Cortex, 42, 295–303.
Stuss, D. T., Floden, D., Alexander, M. P., Levine, B., &
Katz, D. (2001). Stroop performance in focal lesion
patients: Dissociation of processes and frontal lobe
lesion location. Neuropsychologia, 39, 771–786.
Tzelgov, J., Henik, A., & Berger, J. (1992). Controlling Stroop
effects by manipulating expectations for color words.
Memory & Cognition, 20, 727–735.
Vivas, A. B., Humphreys, G. W., & Fuentes, L. J. (2003).
Inhibitory processing following damage to the parietal
lobe. Neuropsychologia, 41, 1531–1540.
Weiss, P. H., Zilles, K., & Fink, G. R. (2005). When visual
perception causes feeling: Enhanced cross-modal
processing in grapheme–color synesthesia. Neuroimage,
28, 859–868.
1576
Journal of Cognitive Neuroscience
Volume 18, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
8
e
9
-
1
p
5
d
7
f
0
/
1
1
8
9
/
3
5
9
9
/
7
1
8
5
7
o
0
c
/
n
1
2
7
0
5
0
6
6
3
1
5
8
1
/
9
j
1
o
5
c
7
n
0
.
p
2
d
0
0
b
6
y
.
g
1
u
8
e
.
s
9
t
.
o
1
n
5
0
7
8
0
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1