Brain-to-Brain Synchrony and Learning Outcomes Vary by
Student–Teacher Dynamics: Evidence from a Real-world
Classroom Electroencephalography Study
Dana Bevilacqua1*, Ido Davidesco1*, Lu Wan2, Kim Chaloner3, Jess Rowland1,4,
Mingzhou Ding2, David Poeppel1,5, and Suzanne Dikker1,6
Abstracto
■ How does the human brain support real-world learning?
We used wireless electroencephalography to collect neuro-
physiological data from a group of 12 senior high school stu-
dents and their teacher during regular biology lessons. Six
scheduled classes over the course of the semester were orga-
nized such that class materials were presented using different
teaching styles (videos and lectures), and students completed
a multiple-choice quiz after each class to measure their reten-
tion of that lesson’s content. Both students’ brain-to-brain syn-
chrony and their content retention were higher for videos
than lectures across the six classes. Brain-to-brain synchrony
between the teacher and students varied as a function of stu-
dent engagement as well as teacher likeability: Students who
reported greater social closeness to the teacher showed
higher brain-to-brain synchrony with the teacher, pero esto
was only the case for lectures—that is, when the teacher is
an integral part of the content presentation. Además, estu-
dents’ retention of the class content correlated with student–
teacher closeness, but not with brain-to-brain synchrony. Estos
findings expand on existing social neuroscience research by
showing that social factors such as perceived closeness are re-
flected in brain-to-brain synchrony in real-world group settings
and can predict cognitive outcomes such as students’ academic
actuación. ■
INTRODUCCIÓN
Methodological advances in neuroscience research have
enabled novel approaches to investigating how the brain
supports dynamic real-world social interactions. por ejemplo-
amplio, researchers have begun to study the neural basis
of social interactions by comparing the brain responses of
multiple individuals during a variety of seminaturalistic tasks
(para una revisión, see Hasson & Frith, 2016; Babiloni & Astolfi,
2014; Scholkmann, Holper, Lobo, & Lobo, 2013; Hasson,
Ghazanfar, Galantucci, Garrod, & Keyser, 2012). Investigación
involving turn-taking in gestural (Schippers, Roebroeck,
Renken, Nanetti, & Keyser, 2010) as well as verbal
(Dikker, Silbert, Hasson, & Zevin, 2014; Esteban, Silbert,
& Hasson, 2010) communication have demonstrated a re-
lationship between brain synchrony and comprehension
as well as the predictability of another person’s commu-
nicative act. Further work has shown that complex audio-
visual stimuli (p.ej., natural movies) elicit similar brain
This paper appeared as part of a Special Focus deriving from
a symposium at the 2017 annual meeting of the Cognitive
Neuroscience Society, entitled, “Real World Neuroscience.”
1New York University, 2University of Florida, 3Grace Church
School, Nueva York, Nueva York, 4School of Visual Arts, Nueva York, Nueva York,
5Max Planck Institute for Empirical Aesthetics, Frankfurt am
Main, Alemania, 6Universidad de Utrecht
*These authors contributed equally.
activity among viewers and emotional responses and,
crucialmente, vary as a function of participants’ attentional
engagement (Ki, Kelly, & Parra, 2016; Chang et al.,
2015; Nummenmaa et al., 2012; Jääskeläinen et al.,
2008; Hasson, Nir, Exacción, Fuhrmann, & Malach, 2004).
Although these experiments explore the similarities
and differences in neural activity across participants as
they engage in similar or pseudointeractive tasks, ellos
do not capture the dynamic nature of real-world settings.
Methodological constraints limit the ways in which
researchers have been able to explore the brain basis of
social interactions as they occur in real world. A pesar de
providing promising results, these studies are still largely
confined to the laboratory, mostly limited to dyads, y
typically use neuroimaging technology with low temporal
resolution (p.ej., functional near-infrared spectroscopy).
We know that the direct study of face-to-face exchanges
is critical to fully understand social interactions, yet there
is a gap in the research exploring the underlying neural
mechanisms of joint behavior as it naturally unfolds
(Dumas, 2011). To be able to investigate how the brain
supports interactions that resemble the complexity of the
interactions we encounter in everyday life, hyperscanning
research will have to accommodate more ecologically
valid situations (Babiloni & Astolfi, 2014; Schilbach
et al., 2013; Dumas, 2011). En el estudio actual, nosotros
investigated the neuroscience of real-world classroom
© 2018 Instituto de Tecnología de Massachusetts. Published under a
Creative Commons Attribution 4.0 Internacional (CC POR 4.0) licencia.
Revista de neurociencia cognitiva 31:3, páginas. 401–411
doi:10.1162/jocn_a_01274
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
oh
C
norte
_
a
_
0
1
2
7
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
learning using mobile electroencephalography (EEG)
headsets to simultaneously record participants in support
of previous experimentation by Dikker et al. (2017).
Cada vez más, research shows that, during joint actions,
people become “coupled” at motor, perceptual, and cog-
nitive levels in both planned and improvised coordina-
ción (Knoblich, Butterfill, & Sebanz, 2011). Participantes
during synchronized motor activity modify their own
actions in response to their partners (Dumas, Nadel,
Soussignan, Martinerie, & Garnero, 2010). Hyperscan-
ning neuroscience research has shown not only a rela-
tionship between synchrony at the motoric and neural
niveles (Dumas et al., 2010) but also that face-to-face inter-
actions moderate the relationship between social factors
and brain-to-brain synchrony (Dikker et al., 2017; Jiang
et al., 2015; Hari, Himberg, Nummenmaa, Hämäläinen, &
Parkkonen, 2013; Scholkmann et al., 2013; Jiang, dai, Peng,
Liu, & Lu, 2012; Dumas et al., 2010). Específicamente, joint action
tasks demonstrate that synchronous motor activity within
interactive partners leads to increased feelings of affil-
iation and social cohesion (Valdesolo, Ouyang, & DeSteno,
2010; Hove & Risen, 2009; Bernieri, 1988), particularmente en
cooperative versus competitive contexts, and that this is
reflected at the neural level (cheng, li, & Hu, 2015; Cual,
Bryant, & Reiss, 2012; Yun, Watanabe, & Shimojo, 2012).
The classroom setting is an exemplary environment to
systematically investigate group interactions—between
students and students with their teacher—under semi-
controlled conditions, while measuring behavioral and
cognitive outcomes (p.ej., academic performance and stu-
dent engagement; Scholkmann et al., 2013; Watanabe,
2013). The dynamic interaction between a teacher and
a group of students is fundamental to classroom learning
and has been shown to affect both student engagement
and academic achievement ( Watanabe, 2013; abrazos,
Wu, Kwok, Villarreal, & Johnson, 2012; Walton & cohen,
2011; Martillo & Planta, 2001; Bernieri, 1988). Teaching
and learning can be viewed as a joint action between
the teacher and the students such that features of the
interactive partner and the event are treated as stimuli
in a reciprocal exchange (Sensevy, Gruson, & Forest,
2015). Research into student–teacher relationship ex-
changes in the classroom suggests that exploring under-
lying neural activity may support understanding and
predicting educational outcomes from the perspective
of the teacher and the student (Holper et al., 2013). Re-
cently, researchers have used portable EEG equipment
in the classroom to record nine students simultaneously
during natural movie viewing and reproduced findings
from similar, laboratory-based, experimental designs
with commercial-grade equipment, demonstrating the
potential for real-world measurement of students’ atten-
tional engagement (Poulsen, Kamronn, Dmochowski,
Parra, & Hansen, 2017).
In further recent classroom-based experimentation,
which forms the foundation for the current work, autores
report that brain-to-brain synchrony (quantified as total
interdependence [TI] or interbrain coherence; Wen,
Mo, & Ding, 2012) between students during class activi-
ties was correlated with student engagement and class-
room social dynamics (Dikker et al., 2017). Students’
synchrony to the group was higher in their preferred
teaching style (p.ej., video over lecture) and related to
greater student focus, group affinity, and empathy
(Dikker et al., 2017). Además, findings in group social
dynamics speak directly to the presence of others as a
moderator of student synchrony during class. Para examen-
por ejemplo, higher student ratings of their teacher correlated
with a smaller difference between video (donde el
teacher played no role) and lecture conditions (dónde
the teacher was central), and students who engaged in
prelesson face-to-face baseline recordings showed the
highest pairwise synchrony during class with their mutual
gaze partner compared with other random students in
el grupo (Dikker et al., 2017). Juntos, their results
suggest that brain-to-brain synchrony is driven by a com-
bination of (i) stimulus properties, (ii) individuo difiere-
ences, y (iii) social dynamics.
The Current Study
In the context of classroom learning, attention is known to
play a critical role in learning and maintaining information
(reyes, Brackett, Rivers, Blanco, & Salovey, 2012), y
student attention is a challenge even for the most experi-
enced teachers (Evertson & Weinstein, 2013). If brain-to-
brain synchrony indeed increases as a function of shared
atención (to the teacher, the lesson content, colegas), como
suggested by the research summarized above (Dikker
et al., 2017; Ki et al., 2016), and attention increases reten-
ción (cohen & Parra, 2016), we can then ask whether a
student’s neural synchrony to the rest of the group or with
the teacher predict their retention of the content.
To capture the unique underlying neural activity of the
social and behavioral factors in the class, we simulta-
neously recorded students and their teacher during their
usual high school biology lessons, which included both
video and lecture components, and tested students’ re-
tention postlesson. These teaching styles generated data
that were relatively free of motion artifacts, a consider-
able concern in real-world EEG research (see Dikker
et al., 2017, supplementary materials for an extensive dis-
cussion and evidence showing that motion artifacts do
not explain brain-to-brain synchrony). We aimed to
address two research questions, pertaining to the rela-
tionship between brain-to-brain synchrony (TI; Wen
et al., 2012) on the one hand and classroom learning
and student–teacher relationships on the other.
2.
1. Does brain-to-brain synchrony between a student and
their peers predict their retention of the class content?
Is there a relationship between student–teacher
brain-to-brain synchrony, classroom learning, y
student–teacher relationships, respectivamente?
402
Revista de neurociencia cognitiva
Volumen 31, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
oh
C
norte
_
a
_
0
1
2
7
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
In Dikker et al. (2017), both student ratings (p.ej., en-
gagement) and brain-to-brain synchrony between stu-
dents were higher when students viewed lesson-related
videos compared with lectures, which allows us to ask
if such a parametric difference also exists for content
retention (Research Question 1). Además, como el
teacher plays a pivotal role during lectures but not during
videos, we ask whether the student–teacher relationship
matters more when the teacher is present (Investigación
Question 2). To address these questions, we employed
a similar setup as the classroom EEG findings from
Dikker et al. but included two metrics in addition to
student-to-group synchrony: (1) student performance
y (2) student–teacher brain-to-brain synchrony.
MÉTODOS
Overall Procedure
This experiment took place between February 2016 y
Puede 2016. We collaborated with a New York City high
school biology class composed of juniors and seniors.
Students received an introduction to neuroscience and
the experimental background in the fall semester preced-
ing the experiment and then a refresher in the spring
semester right before the classroom EEG sessions
(Figura 1A). Students were also introduced and trained
to use the wireless EEG equipment to assist in presession
setup and postsession breakdown procedures (ver
vimeo.com/212150060 for an impression of the class-
room EEG setup in a different school). Pre-experimental
questionnaires were electronically mailed to students and
the teacher for completion before recording sessions.
After all recording sessions, we returned for a nonrecord-
ing session to administer the same preexperiment elec-
tronic questionnaires.
During six classroom sessions, 80 min each, we visited
the class to record neurophysiological activity as the
students and their teacher engaged in semiregular class-
room activities (Figure 1B and C). Class content followed
the teacher’s preplanned biology curriculum. For every
session, to preserve time, participants and experimenters
worked together to set up the mobile EEG headsets and
test connectivity across all channels. Students also filled
out presession questionnaires during this time (ver
abajo). EEG was recorded from students and their
teacher simultaneously for all conditions.
For each session, students and their teacher’s EEG ac-
tivity was recorded during a preexperimental baseline in
three conditions (p.ej., facing the wall, facing a partner,
and facing the group; 2 min each). Después, all partici-
pants were simultaneously EEG-recorded during the
lección, which was composed of two lecture blocks and
two video blocks (interleaved) of approximately 5 mín.
cada, con 20 min per lesson. The final EEG recording
was a repeated baseline condition also for 2 min each
through altering the sequence of conditions. Después
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
oh
C
norte
_
a
_
0
1
2
7
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
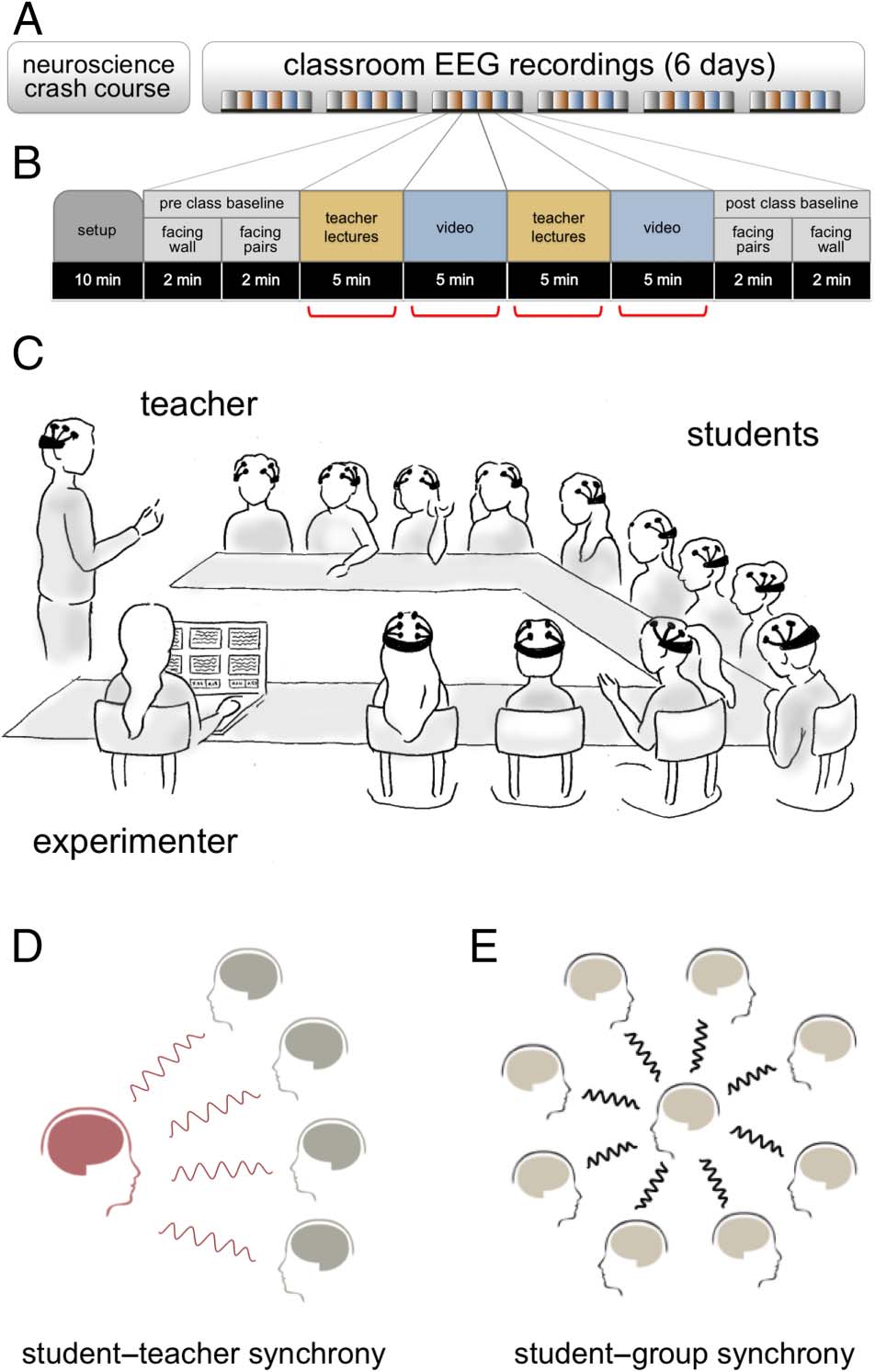
Cifra 1. Experimental setup, procedimiento, and synchrony. (A)
Experimentation was preceded by a neuroscience crash course to
introduce students to the foundations for the current study. EEG
recordings occurred over six sessions. (B) Each session was 80 mín.
and composed of setup, pretesting and posttesting baseline EEG
recordings, and four blocks of experimental conditions (two lectures
interleaved with two videos). After EEG recordings, students were
tested for lesson content retention with quizzes (five questions
per lesson block) and finished with a postsession self-report
cuestionario. (C) Illustration reproduced from Dikker et al. (2017)
of the general classroom recording setup for each session. Synchrony
between student-to-teacher (D) and student-to-group (mi) can be
explored using TI in relation to performance and social factors.
removing EEG equipment, students completed a post-
lesson multiple-choice quiz as a measure of students’
lesson content retention and a self-report questionnaire
for students’ engagement, focus, and likability of the
teaching style (p.ej., lecture vs. video; see details below).
Participantes
Participants of this study were 12 healthy high school
estudiantes (seven girls and five boys, aged 16–18 years),
randomly chosen from the entire class of 19 (one student
de 19 declined participation altogether and was assigned
the role of experimental assistant). Consent forms were
Bevilacqua et al.
403
distributed to all students and the teacher (incluido
parental consent for students under the age of 18 años)
before the beginning of data collection. The institutional
review board of New York University approved all exper-
imental procedures for this study. One student (a girl)
was not included in the statistical analysis reported below
because of limited TI values (p.ej., only student-to-group
TI for three lecture blocks in total for all EEG recording
sessions, and no student-to-teacher TI, was available for
computation).
Teaching Styles
Classroom activities included two “teaching styles”—the
teacher’s lectures and lesson-related educational videos.
Before each recording session, we designed lesson con-
tent with the teacher based on her normal curriculum for
the class and semester so as to maintain continuity and
normalcy in relation to the overall class structure. Ser-
fore each session, the researcher collaborated with the
teacher to design lectures and choose relevant educa-
tional videos suited to the level of the class, estructurando
content to fit the session time constraints (∼5 min per
condición). In each session, the teacher lectured for
aproximadamente 5 min and then presented the students
with a 5-min instructional video, and after a short break,
this sequence was repeated in the same order. Data anal-
yses were performed for each student on each day for
each teaching style. De este modo, there was one TI (see below)
value per student per day per teaching style (Lectures 1
y 2 were averaged together and compared with the
average of Videos 1 y 2).
Student Retention
Students completed a 20-question multiple-choice
knowledge quiz immediately after each recording session
(six in total) to test retention of core concepts from the
day’s lesson. The knowledge quiz included five questions
for each lecture or video unit (de este modo 20 questions in total).
The teacher and the researchers codesigned quizzes be-
fore recording sessions to ensure that quiz content was
adequately paired to the students’ general comprehen-
sion level. Scores were computed as an average per
teaching style per session to assess information reten-
tion for lectures compared with videos. Outlier (<0.2)
and at-ceiling performance (1) was removed in the
statistical analyses reported below.
Questionnaires
Two types of questionnaires were administered to stu-
dents: (i) before and after all recording sessions and (ii)
before and after each recording session. The prestudy
and poststudy questionnaires included demographic
information (gender and age), class and content likability,
and closeness ratings toward the teacher and each stu-
dent in the class. Before and after the recording session,
students filled out brief self-report items including their
engagement and general likability of the day’s lesson and
experimental experience. Students were asked directly
how much they enjoyed and felt engaged in both teach-
ing styles separately for comparison. All self-report ques-
tionnaires were on a scale of 1–7 and were max–min
normalized in all figures for presentation purposes.
Data Collection, Preprocessing, and Analysis
EEG Data Collection
Recordings were collected over six class sessions
throughout the spring semester (February 2016 through
May 2016). Students were briefed on basic EEG technol-
ogy and uses and were aware of movement, speech, and
eye blink artifacts. Further instructions to minimize
movement and speech during recording segments were
given before each lesson, and students were instructed to
reserve questions and discussion for after the recording
session was over. Thus, minimal to no conversational ex-
change occurred between students and their teacher dur-
ing the EEG recordings. EEG activity was recorded
simultaneously from 12 students and their teacher using
Emotiv wireless EMOTIV EPOC EEG headsets (14 chan-
nels; sampling rate = 128 Hz, online notch filter; mastoid
reference locations; Debener, Minow, Emkes, Gandras, &
de Vos, 2012). Custom software built using the Open-
Frameworks software package (www.openframeworks.
com) was used to record EEG data from all 13 partici-
pants simultaneously onto a single computer (MacBook
Pro). Individual laptops (a combination of students’ per-
sonal computers and those provided by the school) were
set up at the beginning of each session to test each par-
ticipant’s headset connectivity and electrode impedance
before data collection. For a detailed discussion of the
software and experimental setup and evaluation of the
EEG data quality, see Dikker et al. (2017; supplemental
materials).
Analysis: Quantifying Brain-to-Brain Synchrony
EEG Preprocessing
The raw EEG data for students and the teacher for each
class and each teaching style (videos and lectures) were
filtered and preprocessed using EEGLAB (Delorme &
Makeig, 2004). The signals were band-pass filtered be-
tween 0.5 and 35 Hz and divided into 1-sec epochs for
artifact rejection and EEG analysis. Artifacts in the data
were both automatically and manually excluded. We first
set a rejection threshold in EEGLAB of ±100 μV for all
channels and then visually inspected each 1-sec epoch
to further exclude eye, muscle, and speech-related arti-
facts. This resulted in an average rejection rate (across
students and days) of 59% of EEG-recorded epochs dur-
ing lecture lessons and 54% of EEG-recorded epochs
404
Journal of Cognitive Neuroscience
Volume 31, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
o
c
n
_
a
_
0
1
2
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
during video lessons. Subsequently, channels with aver-
age amplitude diverging from the mean channel ampli-
tude by 4 SDs were excluded from analysis.
Computing Brain-to-Brain Synchrony: TI
Brain-to-brain synchrony was computed using the method
of TI (Dikker et al., 2017; Wen et al., 2012). Spectral coher-
ence was computed based on the Welch method to limit
bias in coherence estimation (Dikker et al., 2017; Burgess,
2013). For every student–student and student–teacher
pair during each 1-sec epoch recorded per teaching style,
TI was computed for a pair of simultaneously acquired
time series (e.g., (x1,y1), (x2,y2),… (x3,y3)) as:
TIx;y¼ − 1
2π
(cid:2)
λð Þ 1 − C2
xy
ln 1 − C2
xy
− 1
2π
Z π
Z π
dλ
λð Þ
ln
−π
−π
(cid:2)
(cid:3)
where Cxy(λ) is the coherence between the two signals,
x and y, at frequency f = λ/2π.
In this study, six preselected channels, which were
most often free of noise across students in Dikker et al.
(2017), were used for computing TI. These included two
occipital channels (O1, O2), two frontal channels (F3,
F4), and two parietal channels (P7, P8)—unless elec-
trodes have been rejected based on artifacts. TI was
assessed according to the methodology of Dikker et al.
First, we computed the magnitude-squared coherence
using the Welch method for the six preselected one-on-
one paired combinations of electrodes from two partici-
pants. This coherence was calculated for the frequency
range between 1 and 20 Hz by tapering nonoverlapping
1-sec epochs with a Hanning window (zero-padded to
4 sec; Mäki & Ilmoniemi, 2010; Lalor et al., 2005) and per-
forming the Fourier transform with a 0.25-Hz frequency
resolution. A minimum of 30 artifact-free mutual epochs
for pairs was required to be included in the analysis for
the corresponding teaching style per day. For each pair
of participants, TI values across all paired electrodes were
averaged. Then, student-to-group TI was calculated by
averaging all possible pairwise combinations between
one participant and the rest of the group. Student-to-
teacher TI was a pairwise value between the student
and their teacher. Student-to-group TI and student-to-
teacher TI were then entered in the multilevel models
for statistical analysis (Figure 1D and E) to evaluate
students’ synchrony with the group and their teacher in
relation to the performance and social factors.
Analysis Strategy
To investigate the relationship between student-to-
group TI, student-to-teacher TI, quiz scores, and ques-
tionnaire metrics across days, we created multilevel
models (Goldstein, 1995), with days nested within stu-
dents. Multilevel models were implemented in the SAS
PROC MIXED procedure (random effects were modeled
wherever possible; Singer, 1998). Table 1 summarizes the
repeated-measures analyses that were conducted and
their corresponding research questions.
RESULTS
Student-to-Group Synchrony and
Memory Retention
We first tested whether brain-to-brain synchrony be-
tween a student and their peers predicted content reten-
tion. Overall, students retained content presented in
videos better than content from the lectures (video:
Table 1. Repeated-Measures Multilevel Models and Corresponding Research Question (Related to Figures 2 and 3)
Analysis
Research Question
(1) Repeated-measures multilevel regression analysis assessing
Do quiz scores vary by teaching style and as a function of
effects of Teaching Style × Quiz Scores on student-to-group TI
student-to-group TI?
(2) Repeated-measures multilevel regression analysis assessing
Does student-to-teacher TI vary by teaching style and as a
effects of Teaching Style × Engagement on student-to-teacher TI
function of engagement?
(3) Repeated-measures multilevel regression analysis assessing
effects of teacher-modulated student-to-group TI on student-
to-teacher TI and any possible interaction by teaching style
Does any effect of the teacher in student-to-group TI covary
with student-to-teacher TI? Is this the case only for the
(teacher-relevant) lecture teaching style?
(4) Repeated-measures multilevel regression analysis assessing
effects of student-to-teacher closeness on student-to-teacher
TI and any possible interaction by teaching style
Does student-to-teacher closeness predict student-to-teacher TI?
Is this the case only for the (teacher-relevant) lecture teaching
style?
(5) Repeated-measures multilevel regression analysis assessing
effects of student-to-teacher closeness on quiz scores and any
possible interaction by teaching style
Does student-to-teacher closeness predict quiz scores? Is
this the case only for the (teacher-relevant) lecture
teaching style?
(6) Repeated-measures multilevel regression analysis assessing
Does student-to-teacher TI predict quiz scores? Is this
effects of student-to-teacher TI on quiz scores and any
possible interaction by teaching style
the case only for the (teacher-relevant) lecture teaching
style?
Bevilacqua et al.
405
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
o
c
n
_
a
_
0
1
2
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
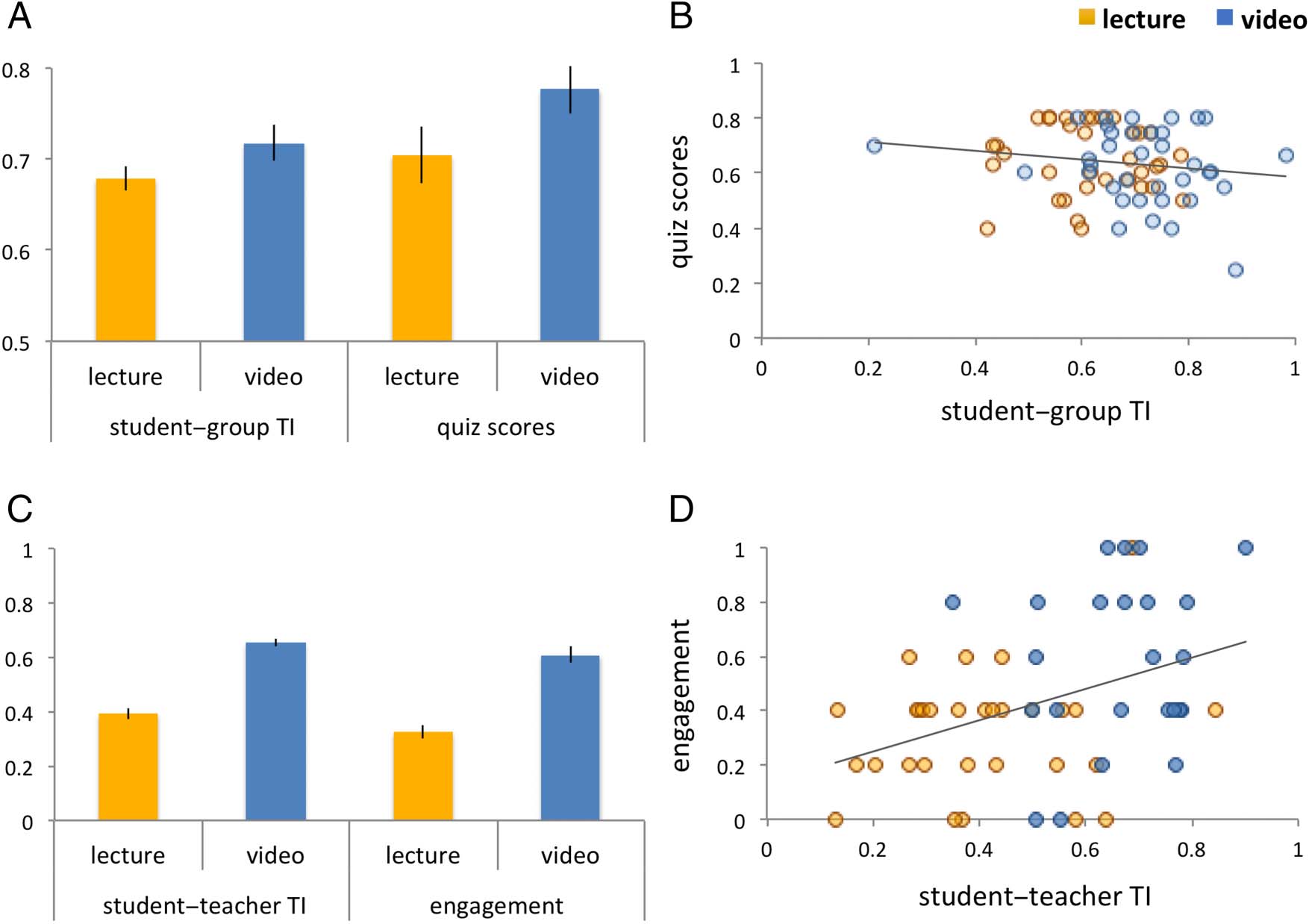
0.78 ± 0.02; lecture: 0.70 ± 0.03; Teaching Style × Quiz
Scores: F(1, 10) = 6.40, p = .029; Figure 2A, right). How-
ever, contrary to our prediction based on previous find-
ings (Cohen & Parra, 2016), there was no significant
relationship between student-to-group brain synchrony
and lesson content retention (Student-to-Group TI ×
Quiz Scores: F(1, 70) = 1.16, p = .2847; Figure 2B;
Table 1, Cell 1).
Brain-to-Brain Synchrony, Student–Teacher
Dynamics, and Learning
Similar to our previous findings (Dikker et al., 2017), stu-
dents reported higher daily engagement for videos com-
pared with lectures (video: 5.05 ± 0.21; lecture: 3.63 ±
0.18; Daily Engagement × Teaching Style: F(1, 9) =
14.67, p = .004). In addition, student-to-teacher TI sug-
gests variations according to teaching style where brain
synchrony with the teacher was significantly higher for
videos compared with lectures (video: 0.65 ± 0.02; lec-
ture: 0.39 ± 0.03; Student-to-Teacher TI × Teaching
Style: F(1, 10) = 35.33, p = .0001; Figure 2C, left), and
there was a strong interaction between the two variables
(Student-to-Teacher TI × Daily Engagement: F(1, 43) =
10.33, p =.003; Figure 2D; Table 1, Cell 2). Interestingly,
daily student engagement was not correlated with
student-to-group brain synchrony (not shown), replicat-
ing similar findings from Dikker et al. (2017), who found
that postsemester engagement ratings, but not daily
engagement ratings, correlated with student-to-group
TI. This suggests that students’ relationship to their
teacher may be a stronger predictor of class engagement
than a student’s peer.
We next explored whether “teacher-relevant” factors
varied as a function of teaching style. As pointed out
above, a major distinction between video and lecture
teaching styles is that the teacher plays a pivotal role dur-
ing lectures, whereas this is not the case for videos.
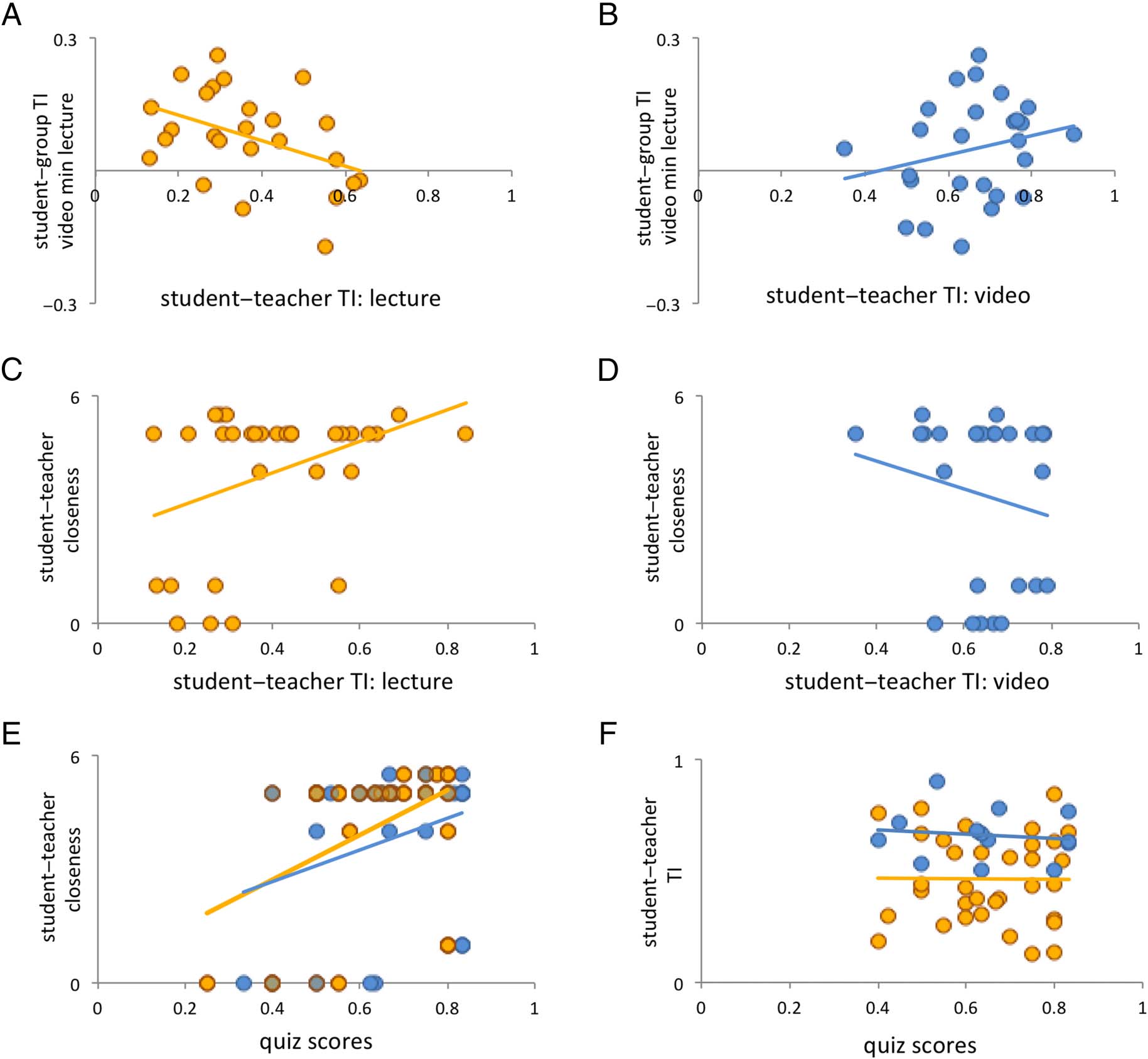
Dikker et al. (2017) found that a student’s teacher like-
ability rating was significantly correlated with the differ-
ence between a student’s student-to-group TI during
video as opposed to lecture content (i.e., student-to-group
TI during videos was used as a “baseline” condition): The
greater the teacher likeability, the smaller the difference
between conditions (recall that video sessions overall
show larger TI values, because of a combination of stimu-
lus properties and engagement factors; see Dikker et al.,
2017). Here, we extend this finding to student-to-teacher
TI: As shown in Figure 3A and B, the correlation between
student-to-teacher TI during lectures, on the one hand,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
o
c
n
_
a
_
0
1
2
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Brain-to-brain synchrony, student retention, and individual differences. (A) Average student-to-group TI (left) and average quiz scores
(right) for two teaching styles (videos vs. lectures). Students’ brain synchrony with the group did not correlate with performance on a knowledge
quiz (B). (C) Student-to-teacher TI (left) and students’ day-to-day self-reported engagement (right). Students’ daily engagement predicted student-to-
teacher TI (D).
406
Journal of Cognitive Neuroscience
Volume 31, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
o
c
n
_
a
_
0
1
2
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Social dynamics predict student–teacher synchrony and retention. (A) The difference in student-to-group TI (video TI minus lecture TI)
predicted greater synchrony with the teacher during lectures, but not during videos (B). (C) Student–teacher closeness predicted student-to-
teacher synchrony during lectures, but not during videos (D). Student–teacher closeness predicted better quiz scores (E), but student-to-teacher
TI did not (F).
and the video–lecture difference between student-to-
group TI, on the other, suggests significant variations
by teaching style (Student-to-Teacher TI × Student-to-
Group TIvideo−lecture: F(1, 28) = 6.48, p = .0167; Table 1,
Cell 3). There was a negative correlation between the two
variables during lectures (r = −.426, p < .05), but not
during videos (r = .23, p = .258). These exploratory
analyses suggest that students who showed greater
brain synchrony with their teacher during the lecture
conditions also showed less of a relative reduction in
their synchrony with the group for lectures compared
with videos.
We observed a similar interaction for the correlation
between student–teacher closeness and student-to-
teacher TI (r = .382, p < .05): Student–teacher closeness
ratings only predicted student-to-teacher TI during lec-
tures, not videos (student–teacher closeness: 3.94 ±
0.26; Student-to-Teacher Closeness × Teaching Style:
F(1, 39) = 3.98, p = .05; Figure 3C and D; Table 1, Cell 4).
Interestingly, there was a significant correlation between
student–teacher closeness and quiz scores (r = .352, p =
.003; Figure 3E; Table 1, Cell 5). However, like student-
to-group TI, student-to-teacher TI did not predict quiz
scores for either condition (Student-to-Teacher TI ×
Quiz Scores: F(1, 35) = 0.05, p = .818; Figure 3F; Table 1,
Cell 6).
DISCUSSION
In an effort to understand the neural basis of student–
teacher interactions and explore the relationship be-
tween brain-to-brain synchrony and classroom learning,
we recorded simultaneous EEG from a group of high
school students and their teacher during their normally
Bevilacqua et al.
407
scheduled biology classes. EEG data were analyzed in
conjunction with a set of behavioral measures, including
quiz scores, self-reported engagement, and student–
teacher closeness.
We show that quiz scores were higher for videos than
lectures, as was brain synchrony. However, although pri-
or work has already demonstrated that brain-to-brain syn-
chrony relates to successful communication (Dikker
et al., 2014; Stephens et al., 2010), memory formation
(Hasson, Furman, Clark, Dudai, & Davachi, 2008), and
information retention (Cohen & Parra, 2016), there was
no relationship between student retention and brain-to-
brain synchrony: Neither student-to-group synchrony nor
student–teacher synchrony predicted quiz scores. It is
worth emphasizing that we did see a similar overall
pattern where quiz scores, student-to-group synchrony,
student–teacher synchrony, and engagement were all
higher for videos as compared with lectures, and we know
from past research that social factors such as student–
teacher closeness and student engagement are related
to student learning (Holper et al., 2013; Hughes et al.,
2012).
There are multiple possible reasons as to why we failed
to replicate previous findings between neural activity and
students’ performance (Cohen & Parra, 2016). First, mea-
suring neurophysiological activity in the real world comes
with its own unique limitations, for example, the trade-off
between preserving naturalistic exchanges to the greatest
degree and minimizing artifacts in data. Classroom ex-
changes are often dynamic and expressive, and although
we instructed participants to minimize movement and
speech during recording segments (recall that only the
teacher spoke during lectures), a certain amount of natural
gesturing was inherent to the design and environment
compared with laboratory-based research. Second, and
most important, prior laboratory-based research that has
linked neural activity and learning used a larger sample
of participants and longer quizzes (Cohen & Parra,
2016). Here, we were constrained by class duration and
class size.
Furthermore, we estimated the overall relationship be-
tween retention and synchrony during different teaching
activities. This means the synchrony values included
moments in which learned items were presented as well
as moments with concepts that were later forgotten. This
may have been too coarse as an approach for measuring
students’ learning: Most past research has compared syn-
chrony during the presentation of remembered versus for-
gotten items (Battro et al., 2013; Holper et al., 2013). Thus,
to accurately quantify the relationship between brain syn-
chrony and learning, it might be necessary to reconstruct
when the content featured on the quizzes was presented
during class and relate student retention of that informa-
tion to group synchrony during those specific “learning”
moments (along the lines of, e.g., Kang & Wheatley,
2017; Wagner, Kelley, Haxby, & Heatherton, 2016).
Unfortunately, this information was unavailable in the
current study as lectures were not scripted verbatim or
videotaped, and the onset of the instructional videos was
not synchronized with the EEG recordings. Finally, the
EEG equipment we used is less precise than laboratory-
grade EEG equipment used in classic experimentation,
maybe resulting in failing to capture subtle effects. As
technology advances in wireless EEG recording options
(e.g., affordable headsets with more electrodes), isolat-
ing neurophysiological activity in relation to specific
stimuli events will be discernible and should be explored
more fully in student-to-group and student-to-teacher
interactions.
Our second research goal concerned the relationship
between student–teacher brain-to-brain synchrony, class-
room learning, and student–teacher social closeness. We
observed that student–teacher synchrony was predicted
by teacher closeness during lectures, but not videos.
One way to interpret this result is that the teacher is a
greater “attractor” of synchrony during lectures than
videos, independent of students’ preferences for videos
over lectures. More simply put: The teacher is the “stim-
ulus” during lectures, but not during videos.
The finding that brain synchrony reflects student–
teacher closeness relates to a growing body of literature
about how social networks are represented in the brain
(Curley & Ochsner, 2017; Parkinson, Kleinbaum, &
Wheatley, 2017; Zerubavel, Bearman, Weber, & Ochsner,
2015). It was recently demonstrated that similarity in
fMRI responses to video stimuli across individuals varies
with distance in a social network, with close friends exhi-
biting the highest degree of neural similarity (Parkinson,
Kleinbaum, & Wheatley, 2018). Another study found that
information about social network position was spontane-
ously activated when participants viewed familiar individ-
uals (Parkinson et al., 2017). Similarly, in our study,
student–teacher closeness was predicted by brain syn-
chrony during lectures, when the students (presumably)
were not actively thinking about their relationship with
the teacher.
The student–teacher results also support an indirect
relationship between synchrony and performance.
Although student–teacher synchrony did not directly relate
to students’ quiz scores, student–teacher closeness did, sup-
porting previous findings (Dikker et al., 2017; Watanabe,
2013). Given the relationship between student–teacher
closeness and quiz scores, future research may further
elucidate whether students better retain information from
lectures than videos over time, as prior research tested
information retention after a period of 3 weeks from pre-
sentation (Cohen & Parra, 2016). In addition, student-to-
teacher synchrony, but not student-to-group synchrony,
was predicted by students’ daily engagement ratings, sug-
gesting that students’ relationship to their teacher, rather
than to their peers, may be a stronger predictor of engage-
ment. This also suggests a crucial link between attention,
identifying the relevant stimulus features for interpreta-
tion, and retention of information related to the target
408
Journal of Cognitive Neuroscience
Volume 31, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
o
c
n
_
a
_
0
1
2
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
stimuli—specifically, as attending to different features of
stimuli can alter interpretation and is reflected in changes
in neural activity (Cooper, Hasson, & Small, 2011).
It is important to clarify that significant correlations in
brain-to-brain synchrony research do not indicate that
brains are “physically linked” (Babiloni & Astolfi, 2014).
Rather, brain-to-brain synchrony is a neural marker,
across all participants, that is a quantifiable reflection of
underlying cognitive psychological processes. One pro-
posal, with growing support (Dikker et al., 2017; Poulsen
et al., 2017; Ki et al., 2016), is that brain-to-brain syn-
chrony increases as shared or joint attention modulates
entrainment by “tuning” neural oscillations to the tempo-
ral structure of our surroundings. Temporally aligned en-
trainment to the oscillatory features of external stimuli
(e.g., teacher’s voice) is thought to support information
extraction from the stimulus, such as in parsing continu-
ous speech into syllables (Giraud & Poeppel, 2012) and
attentional selection of relevant information (Lakatos,
Karmos, Mehta, Ulbert, & Schroeder, 2008). Thus,
stimulus-evoked responses drive the relationship be-
tween similar brain activity in groups and naturalistic
stimuli, and multiple perception-related processes, such
as attentional engagement as well as structural features of
the stimuli, modulate this relationship (Ki et al., 2016;
Poulsen et al., 2017).
It is widely shown that stimulus entrainment heavily
depends on attention (Fiebelkorn, Saalmann, Kastner,
2013; Zion Golumbic et al., 2013; Lakatos et al., 2008).
For example, several recent studies demonstrated that,
in the “cocktail party effect,” when confronted with two
speakers and paying attention only to one of them, oscil-
lations in high-order auditory areas track only the
attended speaker’s voice (Zion Golumbic et al., 2013;
Mesgarani & Chang, 2012). In social interactions, joint at-
tention and mutual gaze drive the defining characteristics
of the exchange: initiator and responder roles, shared
intention and motivation, and the interactive context
(Koike et al., 2016). Stimulus properties (e.g., teaching
style or richness of the audiovisual environment; Hasson
et al., 2004), individual differences (e.g., focus, engage-
ment, personality traits; Nummenmaa et al., 2012), and
social dynamics (e.g., social closeness and social inter-
action; Koike et al., 2016) each mediate attention and
brain-to-brain synchrony.
In our analyses of student-to-group and student-to-
teacher synchrony, we begin to see how neural syn-
chrony reflects the complex interaction between attention
and social dynamics. During the lectures, student–teacher
closeness varied with student–teacher synchrony. Still,
videos overall generated stronger student–teacher syn-
chrony than lectures overall. This may be due to low-level
differences between the two types of stimuli (e.g., stron-
ger audiovisual cues in the videos), in line with prior find-
ings suggesting that correlated neurophysiological activity
is partially driven by low-level visual features (Poulsen
et al., 2017). Together, these findings are readily explained
within a stimulus entrainment account: When the teacher
is the stimulus, student–teacher synchrony increases as a
function of increased attention-modulated stimulus
entrainment (indirectly measured via student–teacher
closeness in our study). Independently, videos are a stron-
ger “entrainer stimulus” than lectures because of their rich
(and spatially constrained) audiovisual content, resulting
in an increase of brain-to-brain synchrony (Ki et al.,
2016). As discussed, a classroom has complex sensory fea-
tures and dynamics, such as educational videos, group pro-
jects and discussion, and interactions with the teacher. This
increases variability of how students may receive and retain
information. Although the student–teacher relationship
mimics more classically explored leader–follower dynamics
(Jiang et al., 2015), little research has directly investigated
the specific nuanced features of this complex social ex-
change to provide insight into the neural underpinnings
of attentional engagement in the real world (Ki et al., 2016).
The interaction between students and their teacher is
implicit and explicit, is social, flows bidirectionally and
continually, and is influenced by behavioral contagion as
individuals automatically imitate each other ( Watanabe,
2013)—all with the added component of performance-
based evaluations and assessments. In this study, we rep-
licated previous findings (Dikker et al., 2017) showing
that students reported higher daily engagement for video
lessons compared with lectures and built upon these
findings by showing that students also performed better
in quizzes measuring content retention in the lesson type
they preferred (i.e., videos). In summary, in addition to
the nature of the stimulus (here, lectures vs. videos),
social dynamics, specifically student–teacher social close-
ness, appear to drive brain-to-brain synchrony. Our find-
ings on brain-to-brain synchrony in a group setting marry
two lines of prior research, namely, studies investigating
neural entrainment to engaging stimuli (e.g., Poulsen
et al., 2017) and studies linking social connectedness to
brain responses (Parkinson et al., 2017, 2018).
Further investigating these complex dynamics as they
occur naturally—such as those between students, peers,
and their teacher in relation to class content—can reveal
more about the nuanced interplay of the various factors
that affect learning in the real world.
Acknowledgments
This research was supported by NSF INSPIRE Track 1 Award
1344285, NSF ECR Award 1661016, and Netherlands Organi-
zation for Scientific Research Award 275-89-018. We thank
the school staff at Grace Church School and especially the
AP Biology students for all of their time, support, and enthu-
siasm; Hasibe Melda Kahraman and Diego Reinero for their
research assistance; and J. Van Bavel for analysis consultation.
We thank Matthias Oostrik for the software design.
Reprint requests should be sent to Dana Bevilacqua or Suzanne
Dikker, New York University, Department of Psychology,
6 Washington Place, Suite 275–276, New York, NY 10003,
or via e-mail: dana.bev@nyu.edu, sdikker@gmail.com.
Bevilacqua et al.
409
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
o
c
n
_
a
_
0
1
2
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Babiloni, F., & Astolfi, L. (2014). Social neuroscience and
hyperscanning techniques: Past, present and future.
Neuroscience & Biobehavioral Reviews, 44, 76–93.
Battro, A. M., Calero, C. I., Goldin, A. P., Holper, L., Pezzatti, L.,
Shalóm, D. E., et al. (2013). The cognitive neuroscience of
the teacher–student interaction. Mind, Brain, and
Education, 7, 177–181.
Bernieri, F. J. (1988). Coordinated movement and rapport in
teacher–student interactions. Journal of Nonverbal
Behavior, 12, 120–138.
Burgess, A. P. (2013). On the interpretation of synchronization
in EEG hyperscanning studies: A cautionary note. Frontiers
in Human Neuroscience, 7, 881.
Chang, W., Jääskeläinen, I. P., Belliveau, J. W., Huang, S., Hung,
A., Rossi, S., et al. (2015). Combined MEG and EEG show
reliable patterns of electromagnetic brain activity during
natural viewing. Neuroimage, 114, 49–56.
Cheng, X., Li, X., & Hu, Y. (2015). Synchronous brain activity
during cooperative exchange depends on gender of partner:
A fNIRS-based hyperscanning study. Human Brain Mapping,
36, 2039–2048.
Cohen, S. S., & Parra, L. C. (2016). Memorable audiovisual
narratives synchronize sensory and supramodal neural
responses. eNeuro, 3.
Cooper, E. A., Hasson, U., & Small, S. L. (2011). Interpretation-
mediated changes in neural activity during language
comprehension. Neuroimage, 55, 1314–1323.
Cui, X., Bryant, D. M., & Reiss, A. L. (2012). NIRS-based
hyperscanning reveals increased interpersonal coherence in
superior frontal cortex during cooperation. Neuroimage, 59,
2430–2437.
Curley, J. P., & Ochsner, K. N. (2017). Neuroscience: Social
networks in the brain. Nature Human Behaviour, 1, 0104.
Debener, S., Minow, F., Emkes, R., Gandras, K., & de Vos, M.
(2012). How about taking a low-cost, small, and wireless EEG
for a walk? Psychophysiology, 49, 1617–1621.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Dikker, S., Silbert, L. J., Hasson, U., & Zevin, J. D. (2014). On
the same wavelength: Predictable language enhances
speaker–listener brain-to-brain synchrony in posterior
superior temporal gyrus. Journal of Neuroscience, 34,
6267–6272.
Dikker, S., Wan, L., Davidesco, I., Kaggen, L., Oostrik, M.,
McClintock, J., et al. (2017). Brain-to-brain synchrony tracks
real-world dynamic group interactions in the classroom.
Current Biology, 27, 1375–1380.
Dumas, G. (2011). Towards a two-body neuroscience.
Communicative & Integrative Biology, 4, 349–352.
Dumas, G., Nadel, J., Soussignan, R., Martinerie, J., & Garnero,
L. (2010). Inter-brain synchronization during social
interaction. PLoS One, 5, e12166.
Evertson, C. M., & Weinstein, C. S. (2013). Handbook of
classroom management: Research, practice, and
contemporary issues. New York: Routledge.
Fiebelkorn, I. C., Saalmann, Y. B., & Kastner, S. (2013).
Rhythmic sampling within and between objects despite
sustained attention at a cued location. Current Biology, 23,
2553–2558.
Giraud, A., & Poeppel, D. (2012). Cortical oscillations and
speech processing: Emerging computational principles and
operations. Nature Neuroscience, 15, 511–517.
Goldstein, H. (1995). Multilevel statistical models. London:
Edward Arnold.
Hamre, B. K., & Pianta, R. C. (2001). Early teacher–child
relationships and the trajectory of children’s school
outcomes through eighth grade. Child Development, 72,
625–638.
Hari, R., Himberg, T., Nummenmaa, L., Hämäläinen, M., &
Parkkonen, L. (2013). Synchrony of brains and bodies during
implicit interpersonal interaction. Trends in Cognitive
Sciences, 17, 105–106.
Hasson, U., & Frith, C. D. (2016). Mirroring and beyond:
Coupled dynamics as a generalized framework for modelling
social interactions. Philosophical Transactions of the Royal
Society of London, Series B, Biological Sciences, 371,
20150366.
Hasson, U., Furman, O., Clark, D., Dudai, Y., & Davachi, L.
(2008). Enhanced intersubject correlations during movie
viewing correlate with successful episodic encoding. Neuron,
57, 452–462.
Hasson, U., Ghazanfar, A. A., Galantucci, B., Garrod, S., &
Keysers, C. (2012). Brain-to-brain coupling: A mechanism for
creating and sharing a social world. Trends in Cognitive
Sciences, 16, 114–121.
Hasson, U., Nir, Y., Levy, I., Fuhrmann, G., & Malach, R. (2004).
Intersubject synchronization of cortical activity during natural
vision. Science, 303, 1634–1640.
Holper, L., Goldin, A. P., Shalóm, D. E., Battro, A. M., Wolf, M.,
& Sigman, M. (2013). The teaching and the learning brain: A
cortical hemodynamic marker of teacher–student interactions
in the Socratic dialog. Science Direct, 59, 1–10.
Hove, M., & Risen, J. (2009). It’s all in the timing: Interpersonal
synchrony increases affiliation. Social Cognition, 27, 949–960.
Hughes, J. N., Wu, J., Kwok, O., Villarreal, V., & Johnson, A. Y.
(2012). Indirect effects of child reports of teacher–student
relationship on achievement. Journal of Educational
Psychology, 104, 350–365.
Jääskeläinen, I. P., Koskentalo, K., Balk, M. H., Autti, T.,
Kauramäki, J., Pomren, C., et al. (2008). Inter-subject
synchronization of prefrontal cortex hemodynamic activity
during natural viewing. Open Neuroimaging Journal, 2,
14–19.
Jiang, J., Chen, C., Dai, B., Shi, G., Ding, G., Liu, L., et al. (2015).
Leader emergence through interpersonal neural
synchronization. Proceedings of the National Academy of
Sciences, U.S.A., 112, 4274–4279.
Jiang, J., Dai, B., Peng, D., Liu, L., & Lu, C. (2012). Neural
synchronization during face-to-face communication. Journal
of Neuroscience, 32, 16064–16069.
Kang, O., & Wheatley, T. (2017). Pupil dilation patterns
spontaneously synchronize across individuals during shared
attention. Journal of Experimental Psychology: General,
146, 569–576.
Ki, J. J., Kelly, S. P., & Parra, L. C. (2016). Attention strongly
modulates reliability of neural responses to naturalistic
narrative stimuli. Journal of Neuroscience, 36, 3092.
Knoblich, G., Butterfill, S., & Sebanz, N. (2011). Psychological
research on joint action: Theory and data. In B. Ross (Ed.),
The psychology of learning and motivation (pp. 59–101).
Cambridge, MA: Academic Press.
Koike, T., Tanabe, H. C., Okazaki, S., Nakagawa, E., Sasaki, A. T.,
Shimada, K., et al. (2016). Neural substrates of shared
attention as social memory: A hyperscanning functional
magnetic resonance imaging study. Neuroimage, 125,
401–412.
Lakatos, P., Karmos, G., Mehta, A. D., Ulbert, I., & Schroeder,
C. E. (2008). Entrainment of neuronal oscillations as a
mechanism of attentional selection. Science, 320, 110–113.
Lalor, E. C., Kelly, S. P., Finucane, C., Burke, R., Smith, R., Reilly,
R. B., et al. (2005). Steady-state VEP-based brain–computer
interface control in an immersive 3D gaming environment.
410
Journal of Cognitive Neuroscience
Volume 31, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
o
c
n
_
a
_
0
1
2
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EURASIP Journal on Applied Signal Processing, 2005,
3156–3164.
Mäki, H., & Ilmoniemi, R. J. (2010). EEG oscillations and
magnetically evoked motor potentials reflect motor system
excitability in overlapping neuronal populations. Clinical
Neurophysiology, 121, 492–501.
Mesgarani, N., & Chang, E. F. (2012). Selective cortical
representation of attended speaker in multi-talker speech
perception. Nature, 485, 233–236.
Nummenmaa, L., Glerean, E., Viinikainen, M., Jääskeläinen, I. P.,
Hari, R., & Sams, M. (2012). Emotions promote social
interaction by synchronizing brain activity across individuals.
Proceedings of the National Academy of Sciences, U.S.A.,
109, 9599–9604.
Parkinson, C., Kleinbaum, A., & Wheatley, T. (2017).
Spontaneous neural encoding of social network position.
Nature Human Behaviour, 1, 1–7.
Parkinson, C., Kleinbaum, A., & Wheatley, T. (2018). Similar neural
responses predict friendship. Nature Communications, 9, 332.
Poulsen, A. T., Kamronn, S., Dmochowski, J., Parra, L. C., &
Hansen, L. K. (2017). EEG in the classroom: Synchronised
neural recordings during video presentation. Scientific
Reports, 7, 43916.
Reyes, M. R., Brackett, M. A., Rivers, S. E., White, M., & Salovey,
P. (2012). Classroom emotional climate, student
engagement, and academic achievement. Journal of
Educational Psychology, 104, 700–712.
Schilbach, L., Timmermans, B., Reddy, V., Costall, A., Bente, G.,
Schlicht, T., et al. (2013). Toward a second-person
neuroscience. Behavioral and Brain Sciences, 36, 393–414.
Schippers, M. B., Roebroeck, A., Renken, R., Nanetti, L., &
Keysers, C. (2010). Mapping the information flow from one
brain to another during gestural communication.
Proceedings of the National Academy of Sciences, U.S.A.,
107, 9388–9393.
Scholkmann, F., Holper, L., Wolf, U., & Wolf, M. (2013). A new
methodical approach in neuroscience: Assessing inter-
personal brain coupling using functional near-infrared
imaging (fNIRI) hyperscanning. Frontiers in Human
Neuroscience, 7, 813.
Sensevy, G., Gruson, B., & Forest, D. (2015). On the nature of
the semiotic structure of the didactic action: The joint action
theory in didactics within a comparative approach.
Interchange, 46, 387–412.
Singer, J. D. (1998). Using SAS PROC MIXED to fit multilevel
models, hierarchical models, and individual growth models.
Journal of Educational and Behavioral Statistics, 23,
323–355.
Stephens, G. J., Silbert, L. J., & Hasson, U. (2010). Speaker–
listener neural coupling underlies successful communication.
Proceedings of the National Academy of Sciences, U.S.A.,
107, 14425–14430.
Valdesolo, P., Ouyang, J., & DeSteno, D. (2010). The
rhythm of joint action: Synchrony promotes cooperative
ability. Journal of Experimental Social Psychology, 46,
693–695.
Wagner, D. D., Kelley, W. M., Haxby, J. V., & Heatherton, T. F.
(2016). The dorsal medial prefrontal cortex responds
preferentially to social interactions during natural viewing.
Journal of Neuroscience, 36, 6917.
Walton, G. M., & Cohen, G. L. (2011). A brief social-belonging
intervention improves academic and health outcomes of
minority students. Science, 331, 1447.
Watanabe, K. (2013). Teaching as a dynamic phenomenon with
interpersonal interactions. Mind, Brain, and Education, 7,
91–100.
Wen, X., Mo, J., & Ding, M. (2012). Exploring resting-state
functional connectivity with total interdependence.
Neuroimage, 60, 1587–1595.
Yun, K., Watanabe, K., & Shimojo, S. (2012). Interpersonal body
and neural synchronization as a marker of implicit social
interaction. Scientific Reports, 2, 959.
Zerubavel, N., Bearman, P. S., Weber, J., & Ochsner, K. N.
(2015). Neural mechanisms tracking popularity in real-world
social networks. Proceedings of the National Academy of
Sciences, U.S.A., 112, 15072–15077.
Zion Golumbic, E., Ding, N., Bickel, S., Lakatos, P., Schevon, C.,
McKhann, G., et al. (2013). Mechanisms underlying selective
neuronal tracking of attended speech at a “cocktail party”.
Neuron, 77, 980–991.
Bevilacqua et al.
411
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
3
4
0
1
1
9
5
6
4
0
8
/
j
o
c
n
_
a
_
0
1
2
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3