Bilateral Gamma/Delta Transcranial Alternating
Current Stimulation Affects Interhemispheric
Speech Sound Integration
Basil C. Preisig1,2,3, Matthias J. Sjerps1,2, Alexis Hervais-Adelman3,
Anne Kösem4, Peter Hagoort1,2, and Lars Riecke5
D
oh
w
norte
yo
oh
a
d
mi
d
Abstracto
■ Perceiving speech requires the integration of different
speech cues, eso es, formants. When the speech signal is split
so that different cues are presented to the right and left ear
(dichotic listening), comprehension requires the integration
of binaural information. Based on prior electrophysiological
evidencia, we hypothesized that the integration of dichotically
presented speech cues is enabled by interhemispheric phase
synchronization between primary and secondary auditory cor-
tex in the gamma frequency band. We tested this hypothesis
by applying transcranial alternating current stimulation (TACS)
bilaterally above the superior temporal lobe to induce or
disrupt interhemispheric gamma-phase coupling. A diferencia de
to initial predictions, we found that gamma TACS applied in-
phase above the two hemispheres (interhemispheric lag 0° )
perturbs interhemispheric integration of speech cues, possibly
because the applied stimulation perturbs an inherent phase lag
between the left and right auditory cortex. We also observed
this disruptive effect when applying antiphasic delta TACS (en-
terhemispheric lag 180°). We conclude that interhemispheric
phase coupling plays a functional role in interhemispheric
speech integration. The direction of this effect may depend
on the stimulation frequency. ■
INTRODUCCIÓN
Our ability to discriminate and assign meaning to speech
sounds relies on the identification and integration of
spectrotemporal cues carried by the acoustic speech
señal. A common approach to study the mechanisms
underlying speech cue integration is to present distinct
cues to different ears (dichotic listening) and to investi-
gate the conditions under which they give rise to the sub-
jective experience of an integrated unified speech sound
(Preisig & Sjerps, 2019; Mathiak, Hertrich, Lutzenberger,
& Ackermann, 2001; Liberman & Mattingly, 1989; Rand,
1974). Although the auditory nerve projects from each
ear to both cerebral hemispheres, processing of acoustic
input is initially dominant in the neural pathway, incluir-
ing the auditory cortex that is “contralateral” to the ear of
presentación (Pollmann, Maertens, por cramon, Lepsien,
& Hugdahl, 2002; Sparks & Geschwind, 1968; Kimura,
1967); for reviews, ver (Hugdahl & Westerhausen, 2016;
Westerhausen & Hugdahl, 2008). Por lo tanto, the unifica-
tion of the binaurally presented speech cues requires in-
terhemispheric integration, eso es, the grouping and
1Donders Institute for Brain, Cognition and Behaviour, Nimega,
Los países bajos, 2Max Planck Institute for Psycholinguistics,
Nimega, Los países bajos, 3University of Zurich, 4Lyon
Neuroscience Research Center (CRNL), Lyon, Francia, 5Maastricht
Universidad
© 2019 Instituto de Tecnología de Massachusetts
fusion of cues that are initially processed by different cere-
bral hemispheres. Además, processing of speech and
idioma, Por ejemplo, phoneme recognition, is dominant
in the left hemisphere (Mesgarani, Cheung, Johnson, &
Chang, 2014; Giraud & Poeppel, 2012; Chang et al., 2010;
Obleser, Zimmermann, Van Meter, & Rauschecker, 2007;
Jäncke, 2002; Zatorre & Belin, 2001). De este modo, the integration
of binaurally presented speech cues may require interhemi-
spheric transfer of information from the right to the left
auditory cortex via the corpus callosum, as described in
the so-called callosal relay model (Steinmann et al., 2014,
2018; Bayazıt, Oniz, Hahn, Güntürkün, & Ozgören, 2009;
Westerhausen, Grüner, Specht, & Hugdahl, 2009; Jäncke,
2002).
The interhemispheric transfer and integration of sen-
sory information has been suggested to be facilitated
through phase synchronization between neural oscilla-
tions in the two hemispheres (Fell & Axmacher, 2011;
Fries, 2005). Por eso, interhemispheric phase synchroni-
zation may play a crucial role for the integration of dichotic
speech cues. In support of this idea, Steinmann et al.
(2014) have shown modulation of interhemispheric gamma
(30–100 Hz) phase synchronization during dichotic speech
escuchando. Más precisamente, increased gamma functional con-
nectivity was observed in a condition requiring transfer of
speech cues for phoneme recognition, and this connectivity
was directed from the right to the left secondary auditory
Revista de neurociencia cognitiva 32:7, páginas. 1242–1250
https://doi.org/10.1162/jocn_a_01498
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
r
t
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
C
2
yo
7
mi
1
–
2
pag
4
d
2
F
2
/
0
3
1
2
3
/
5
7
4
/
6
1
2
oh
4
C
2
norte
_
/
a
1
_
8
0
6
1
1
4
6
9
1
8
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
8
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
/
t
F
.
oh
norte
0
5
METRO
a
y
2
0
2
1
corteza (Steinmann et al., 2018). De este modo, interhemispheric
phase synchronization in the gamma band in the posterior
superior temporal cortex plays a role in the interhemi-
spheric integration of speech.
It is still unclear whether this synchronization contrib-
utes functionally to speech integration or merely results
from it (Zaehle, Lenz, Ohl, & Herrmann, 2010). Más-
encima, it is unclear whether its role is limited to oscillations
in the gamma range. Besides their role in interhemi-
spheric integration of speech, cortical oscillations in the
lower gamma range (25–40 Hz) are also important
para
the processing of phonetic information such as formant
transitions or voicing (Rufener, Oechslin, Zaehle, &
Meyer, 2016; Giraud & Poeppel, 2012; Shamir, Ghitza,
Epstein, & Kopell, 2009; Poeppel, 2003). Slower oscilla-
tions in the delta and theta (∼1–8 Hz) band overlap with
intelligibility-relevant temporal fluctuations in the acous-
tic speech signal and may contribute functionally to the
processing of syllabic information during diotic speech
percepción (Riecke, Formisano, Sorger, Başkent, &
Gaudrain, 2018; Zoefel, Archer-Boyd, & davis, 2018;
Keitel, Ince, Bruto, & Kayser, 2017; Rimmele, Zion
Golumbic, Schröger, & Poeppel, 2015; Gross et al., 2013;
luo & Poeppel, 2007; for a comprehensive review, ver
Kösem & Wassenhove, 2017). De este modo, these slow oscillations
may contribute to diotic speech perception, but there
exists no evidence to suggest that they play a role in
interhemispheric integration.
en este estudio, we investigated the mechanisms under-
lying dichotic speech cue integration. We tested the hy-
pothesis that interhemispheric phase synchronization
plays a functional role in interhemispheric speech inte-
gration. We experimentally manipulated interhemispheric
phase synchronization by applying transcranial alternating
current stimulation (TACS) simultaneously above the audi-
tory cortex in the lateral superior temporal lobe of each
hemisferio. To functionally couple the two regions, nosotros
fixed the phase of TACS across the two stimulation sites
(in-phase condition). En cambio, to functionally decouple
the two regions, we reversed the phase of TACS at one
site (anti-phase condition; Preisig, Sjerps, Kösem, &
Riecke, 2019; Saturnino, Madsen, Siebner, & Thielscher,
2017). This approach has already been successfully
applied to modulate bistable perception in the visual do-
principal (Helfrich et al., 2014). To test for a specific role of
gamma oscillations for interhemispheric speech sound
integración, we applied TACS at 40 Hz (gamma condi-
ción). Además, we included 3.125-Hz TACS (delta
condición) and sham stimulation as control conditions
to enable establishing frequency specificity of the puta-
tive effect of gamma TACS on interhemispheric speech
integración.
Interhemispheric speech integration was assessed
using a dichotic listening task. An ambiguous speech
sound (“base,” perceptually intermediate between the
syllables /ga/ and /da/) was presented to the participants’
right ear and a disambiguating acoustic cue (“chirp,"
which was either a low or high third formant, F3) to their
left ear. Interhemispheric integration of the base and the
chirp is reflected by an increased number of /ga/ reports
in the low-F3 condition and an increased number of /da/
reports in the high-F3 condition (Preisig & Sjerps, 2019).
We predicted that interhemispheric phase synchroni-
zación (in-phase condition) would significantly increase
interhemispheric speech integration, as reflected by an
increased number of /ga/ reports in the low-F3 condition
and an increased number of /da/ reports in the high-F3
condición, compared with interhemispheric phase de-
synchronization (anti-phase condition). Además,
we predicted that functional coupling of bilateral auditory
cortices in the gamma, but not delta frequency band,
would strengthen interhemispheric speech integration,
compared with sham stimulation.
MÉTODOS
Participantes
Thirty-six right-handed volunteers (m = 22.56 años, DE =
2.93; 14 hombres) participated in the study. All participants
had normal or corrected-to-normal visual acuity. The par-
ticipants reported no history of neurological, psychiatric,
or hearing disorders, and all had normal hearing (hearing
thresholds of less than 25 dB HL at 250, 500, 750, 1000,
1500, 3000, y 4000 Hz, tested on both ears separately
using pure tone audiometry) and no threshold difference
between the left and the right ear larger than 5 dB for any
of the tested frequencies. All participants gave written in-
formed consent before the experiment. Ethical approval to
conduct this study was provided by the local ethics com-
mittee (CMO region Arnhem-Nijmegen). This study was
conducted in accordance with the principles of the latest
version of the Declaration of Helsinki.
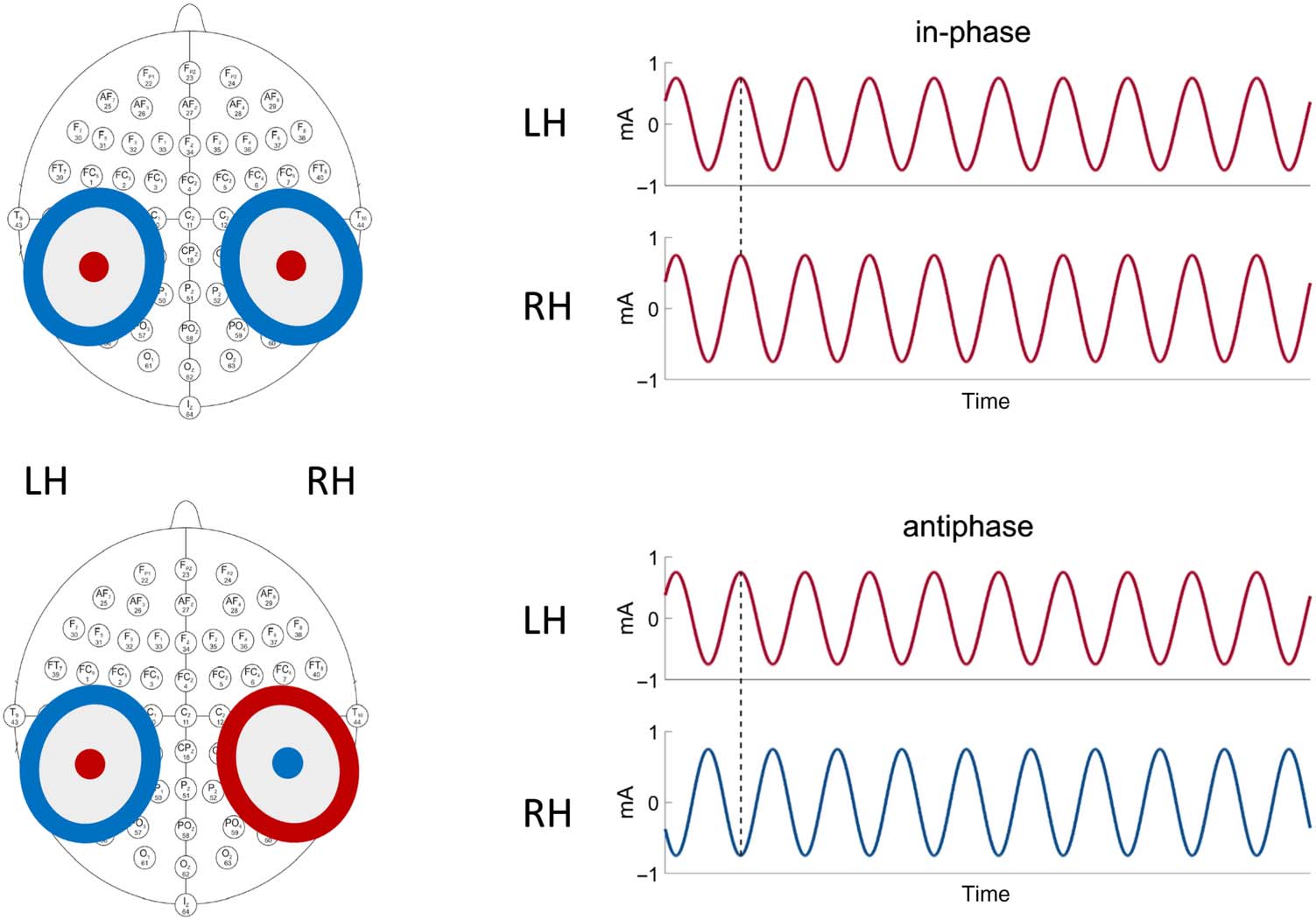
Electric Stimulation
Electric currents were applied through two high-density elec-
trode configurations, each consisting of concentric rubber
electrodes: a central circular electrode (radius = 1.25 cm)
and a surrounding ring electrode (inner radius = 3.5 cm,
outer radius = 4.8 cm). Each electrode configuration was
connected to a separate battery-driven transcranial current
stimulator (Neuroconn, Ilmenau, Alemania), similar to pre-
vious two-channel approaches (Ten Oever et al., 2016;
Riecke, Formisano, Herrmann, & Sack, 2015). The elec-
trode configurations were centered according to the inter-
national 10–20 system over CP5 (above the left cerebral
hemisferio) and CP6 (above the right cerebral hemi-
sphere; ver figura 1). These scalp locations were chosen
to produce relatively strong currents in the target regions
over the auditory speech areas (es decir., left and right lateral
superior temporal lobe), as suggested by prior electric field
simulations on a standard head model using the simnibs
toolbox (Thielscher, Antunes, & Saturnino, 2015).
Preisig et al.
1243
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
r
t
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
C
2
yo
7
mi
1
–
2
pag
4
d
2
F
2
/
0
3
1
2
3
/
5
7
4
/
6
1
2
oh
4
C
2
norte
_
/
a
1
_
8
0
6
1
1
4
6
9
1
8
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
8
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
t
/
.
F
oh
norte
0
5
METRO
a
y
2
0
2
1
Cifra 1. Dual-site high-density
TACS setup. Left: The electrode
configurations were centered
according to the international
10–20 system over CP5 (arriba
the left cerebral hemisphere)
and CP6 (above the right
cerebral hemisphere). Right:
The interhemispheric phase
synchrony was manipulated
using in-phase TACS (0° phase
lag between stimulation sites,
dotted line) and anti-phase
(180° phase lag, dotted line)
TACS. The colors represent the
polarity (positive = red;
negative = blue) of the current
for the time stamp highlighted
by the dotted line. RH = right
hemisferio; LH = left
hemisferio.
TACS was applied at a frequency in the low gamma
banda (40 Hz) or in the delta band (3.125 Hz), the latter
matched the timescale of the syllabic envelope, eso es,
the duration of the syllable matched the length of a half
cycle of the delta TACS. Before starting the actual exper-
mento, we ensured that all participants tolerated the
TACS well. TACS intensity was adjusted individually to
the point for which the participant reported feeling com-
fortable or uncertain about the presence of the current
(1.4 ± 0.1 mA peak-to-peak, mean ± SD across par-
ticipants). Impedance was kept below 10 kΩ. el promedio
current density was 0.2 mA/cm2 at the center electrode
y 0.06 mA/cm2 at the concentric ring electrode. Stim-
ulation was ramped over the first and the last 10 sec of
each experimental block using raised cosine ramps.
The timing of the electric and auditory stimuli was con-
trolled using a multichannel D/A converter (National
Instrumentos, sampling rate: 11 kHz) and Datastreamer
software (Ten Oever et al., 2016). Visual stimulation and
response recording were controlled using Presentation
software ( Versión 18.0, Neurobehavioral Systems, Cª,
berkeley, California).
Behavioral Pretest
Interhemispheric speech sound integration was assessed
by simultaneously presenting an ambiguous base and a
disambiguating chirp (F3) to the right and left ear,
respectivamente. The chirp supported either a /da/ (high F3
∼ 2.9 kHz) or a /ga/ (low F3 ∼ 2.5 kHz) interpretation of
the ambiguous base. Because perceptual category bound-
aries may vary across individuals, a pretest was used to
define participant-specific ambiguous base stimuli for the
main experiment. The pretest included the presentation
of nine stimuli of the /da/–/ga/ continuum, cada 16 veces,
in random order. To make this pretest most similar to the
main experiment, syllables of the /da/–/ga/ continuum
were presented to the right ear, and a single F3 chirp
(identical to the F3 component in the ambiguous base
stimulus) to the left ear. Subjective category boundaries
were estimated by assessing individual psychometric
curves and identifying the point at which participants re-
ported perceiving the stimulus as /da/ or /ga/ in ∼50% of
the trials. The stimulus associated with this individual
category boundary was then used as the base stimulus
for the subsequent main experiment (Preisig & Sjerps,
2019). Further detail concerning stimulus creation is re-
ported in a previous publication using the same materials
(Preisig & Sjerps, 2019).
Experimental Design and Task
The experiment included four stimulation conditions and
sham stimulation. Electric stimulation was applied at one
of the two frequencies, 40 Hz and 3.125 Hz. Each of these
frequency conditions was presented in two interhemi-
spheric phase synchronization conditions: (A) “In-phase
stimulation” was applied with a phase lag of 0° between
the central electrodes placed over the left and the right
auditory speech areas (es decir., bilateral superior temporal
lobe) to induce interhemispheric synchronization. (B)
“Anti-phase stimulation” was applied with a relative phase
lag of 180° to induce interhemispheric desynchronization
(Preisig et al., 2019; Saturnino et al., 2017; ver figura 1).
1244
Revista de neurociencia cognitiva
Volumen 32, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
r
t
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
3
C
2
yo
7
mi
1
–
2
pag
4
d
2
F
2
/
0
3
1
2
3
/
5
7
4
/
6
1
2
oh
4
C
2
norte
_
/
a
1
_
8
0
6
1
1
4
6
9
1
8
8
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
1
oh
4
norte
9
0
8
8
.
pag
S
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
/
.
t
F
oh
norte
0
5
METRO
a
y
2
0
2
1
During “sham stimulation” (placebo), the onset ramp was
followed immediately by an offset ramp, eso es, no elec-
tric stimulation was applied during the actual experiment.
The ramp was repeated at the end of the block.
The experiment consisted of 10 experimental blocks.
Each block consisted of 48 trials containing the ambigu-
ous base stimulus (for which the F3 frequency was set at
the participant-specific subjective category boundary
value that was obtained in the pretest) y 12 trials con-
taining unambiguous base stimuli (which contained an
F3 component that supported a clear /da/ interpretation
[∼2.9 kHz] or a /ga/ interpretation [∼2.5 kHz]) presentado
to the right ear. The ambiguous base stimulus was paired
with a disambiguating F3 chirp presented to the left ear
(24 trials with the high F3 chirp and 24 trials with the low
F3 chirp). In the unambiguous stimuli, a chirp with the
same F3 frequency as the base was presented to the left
ear. Unambiguous stimuli did not require interhemispheric
integration for disambiguation because participants could
readily identify these stimuli based on monaural input
solo, eso es, the unambiguous base stimulus presented
to the right ear. The first half of the experiment included
five blocks of trials: gamma in-phase TACS, gamma anti-
phase TACS, delta in-phase TACS, delta anti-phase TACS,
and sham. The order of the first five experimental blocks
was reversed in the second half of the experiment. The or-
der of all blocks was pseudorandomized, such that blocks
of the same TACS frequency followed upon each other and
counterbalanced across participants. This pseudorandomi-
zation scheme was used to account for potential cross-
frequency carryover effects (zorros, Bruto, & Thut, 2015).
After each block, participants were asked to rate the
subjective strength of any sensations induced by the
stimulation on a visual analogue scale from 0 cm (No
subjective sensations) a 10 cm (strong subjective sen-
sations). Although sensation ratings were relatively low
in all conditions, TACS blocks (m = 2.64, DE = 1.38)
were rated significantly higher than sham blocks (m =
1.77, DE = 1.46), t(25) = 3.11, pag < .01. However, even
though participants rated TACS and sham blocks dif-
ferently (Turi et al., 2019; Zoefel, Allard, Anil, & Davis,
2020), this unlikely influenced our main results, as we
found no association between sensation ratings and be-
havioral performance, Pearson’s R(128) = −0.14, p =
.10, across stimulation conditions.
The auditory stimuli were presented with an ISI of on
average 3.5 sec. The exact ISI was set so that the syllable
onset occurred at one of six predefined, equidistant
TACS phases (TACS/syllable onset lag: 30°, 90°, 150°,
210°, 270°, 330° ). This allowed compensating for indi-
vidual differences in the optimal relative TACS syllable
timing (Zoefel, Davis, Valente, & Riecke, 2019; Riecke
et al., 2018; Riecke, Formisano, et al., 2015; Riecke, Sack,
& Schroeder, 2015), with the aim to improve the detect-
ability of putative stimulation effects in the group-level
analysis. In this study, we did not observe any effect of
TACS/syllable onset lag (Figure 2). Thus, the behavioral
data were pooled across the six TACS/syllable onset lags
for each stimulation condition. Every stimulus was pre-
ceded by a fixation cross presented 600 msec before
auditory stimulus onset. At 1450 msec after the fixation
cross onset, the response options /ga/ and /da/ were pre-
sented (one above and one below the fixation cross,
falling within a visual angle of 9.43° ). The participants
indicated their response by pressing the corresponding
response button with their left index finger.1 Participants
were instructed to perform as accurately and as fast as
possible. The position (up vs. down) of the response
options was counterbalanced across participants.
Data Analysis
In a first step, we assessed the reliability of the categorical
judgments of individual participants on unambiguous
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
2
p
4
d
2
f
2
/
0
3
1
2
3
/
5
7
4
/
6
1
2
o
4
c
2
n
_
/
a
1
_
8
0
6
1
1
4
6
9
1
8
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
8
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
f
.
t
o
n
0
5
M
a
y
2
0
2
1
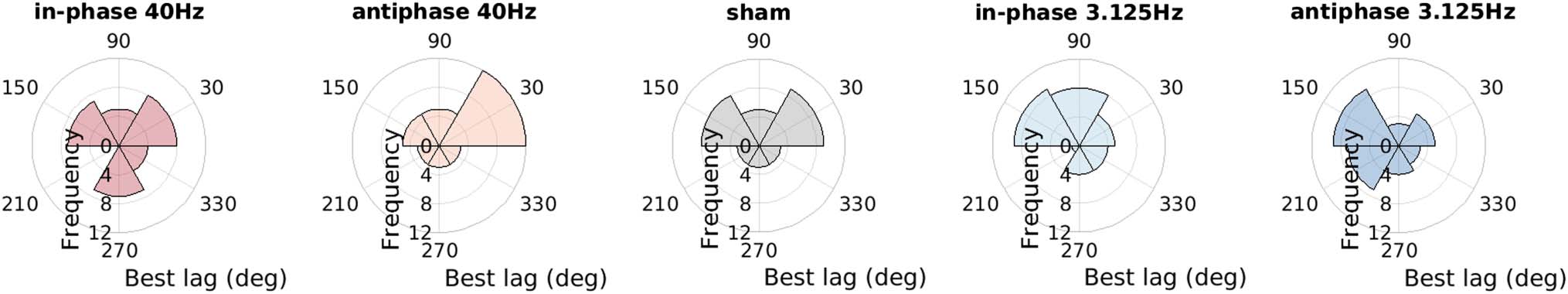
Figure 2. The phase angle histograms show the distribution of the participants’ best TACS/syllable onset lag in the stimulation condition labeled
above each histogram (in-phase 40 Hz, anti-phase 40 Hz; sham, in-phase 3.125 Hz, anti-phase 3.125 Hz). The optimal timing of TACS and the
syllable presentation, that is, average best lag was 87° ± 12° (mean ± SEM ) across participants. The distribution of the participants’ best lag pooled
across TACS stimulation condition (in-phase 40 Hz, anti-phase 40 Hz; sham, in-phase 3.125 Hz, anti-phase 3.125 Hz) did not deviate significantly
from uniformity (Rayleigh test for nonuniformity of circular data, z = 1.733, p = .177), suggesting that the best lag varied substantially across
participants. Moreover, the best lags observed in the different stimulation conditions were observed to be uncorrelated ( ps >.08), Indicando que
participants’ best lag varied across stimulation conditions. To compensate for these individual differences in optimal relative TACS syllable timing,
the best lag was aligned across participants, and the remaining phase bins were phase wrapped, separately for each participant and stimulation
condición (for details, see Experimental Design and Task section). Initial analysis of the aligned data for an effect of TACS/syllable onset lag revealed
no significant result; por lo tanto, the six TACS/syllable onset lags were pooled in all subsequent analyses.
Preisig et al.
1245
endpoint trials (base and chirp stimuli with the same F3
endpoint, supporting the interpretation of /ga/ or /da/)
collected during the sham blocks. Por eso, for each partic-
ipant, we tested with a chi-square test whether the pro-
portion of /ga/ responses differed between /ga/ and /da/
endpoint stimuli. Based on this criterion, the data of four
participants were excluded from further analyses because
their classification accuracy did not significantly exceed
chance level. One additional participant was excluded
because of a technical error during the experiment.
De este modo, the final data set included data from 31 Participantes
(m = 22.63 años, DE = 3.20, 12 hombres).
Two dependent variables were analyzed: the categori-
cal response on each individual trial (0 = /da/; 1 = /ga/)
and the proportion of responses consistent with the pre-
sented F3 chirp (es decir., those in which interhemispheric in-
tegration occurred), per condition. These variables were
computed based on participants’ responses to the stimuli
requiring interhemispheric integration, eso es, the stim-
uli composed of an ambiguous base and a disambi-
guating F3 chirp. For each stimulation condition, el
proportion of integrated trials was calculated per TACS/
syllable onset lag, which were concatenated to build a
behavioral time series. To compensate for individual dif-
ferences (Cifra 2), el maximo (best lag) of the time
series was subsequently aligned across individuals.
Because we did not observe any effect of TACS/syllable
onset lag, the behavioral time series were pooled across
the six TACS/syllable onset lags for each stimulation con-
condición. Statistical analyses were conducted in R ( Versión
3.3.3) using parametric tests (normality assumption was
fulfilled, the dependent variable in each of our conditions
was normally distributed, Shapiro–Wilk test of normality,
ps > .19): Linear mixed-effect models were used to ana-
lyze categorical responses, and repeated-measures
ANOVAs were used to test for a stimulation effect, enterrar-
hemispheric phase effect, frequency effect, and interac-
ciones. Post hoc comparisons were conducted using
paired t tests and false discovery rate (FDR) corrections
for multiple comparisons were applied (Benjamini &
Hochberg, 1995).
RESULTADOS
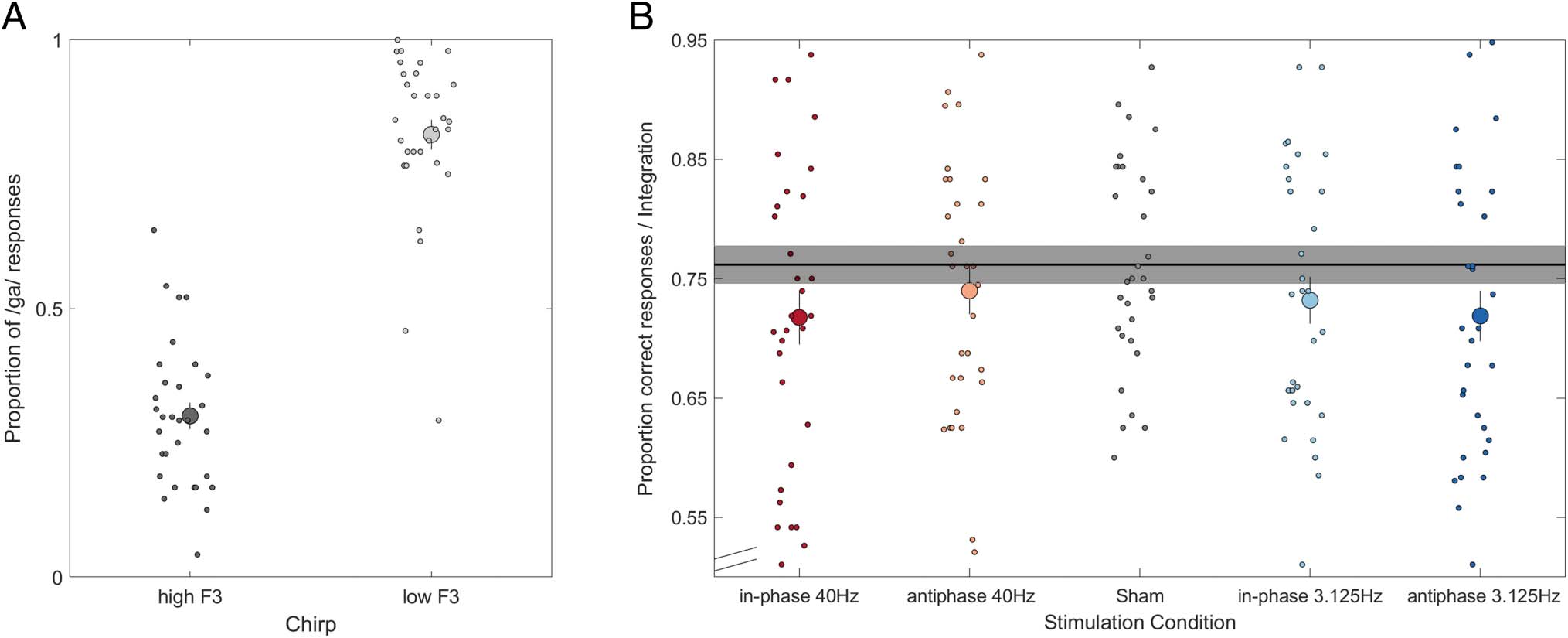
The average classification accuracy (%) including unambig-
uous stimuli (extreme points from the /ga/–/da/ continuum)
during sham blocks was high (m = 90.59, DE = 8.74). Para
trials that required interhemispheric integration, par-
ticipants integrated the information from the F3 chirp on
average on 73.4 ± 9.7% (mean ± SEM) of the trials that
included an ambiguous base stimulus. Primero, probamos
whether participants’ responses to ambiguous base stimuli
were influenced by the frequency of the disambiguating F3
chirp presented to the contralateral ear. For this analysis,
we only included sham blocks. We observed that partici-
pants gave on average 34.12 ± 10.08% (mean ± SEM )
/ga/ responses to ambiguous bases combined with the high
(∼2.9 kHz) F3 chirp and 80.92 ± 9.14% (mean ± SEM) /ga/
responses to ambiguous bases combined with the low
(∼2.5 kHz) F3 chirp. To confirm that the chirp F3 frequency
influenced participants’ response (0 = /da/; 1 = /ga/ re-
respuesta), a logistic linear mixed-effect model with the fixed
factor “chirp type” (niveles: high F3 = −1; low F3 = 1), y
by-participant random intercepts and slopes were fitted to
los datos. The analysis revealed a main effect of chirp type
(B = 2.733, z = 13.052, pag < .001). This result indicates that
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
2
p
4
d
2
f
2
/
0
3
1
2
3
/
5
7
4
/
6
1
2
o
4
c
2
n
_
/
a
1
_
8
0
6
1
1
4
6
9
1
8
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
8
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
f
t
o
n
0
5
M
a
y
2
0
2
1
Figure 3. (A) The proportion of /ga/ responses (mean ± SEM ) as a function of chirp type (high F3, low F3) in the sham condition. (B) Participants’
average performance (mean ± SEM across participants) is shown for each stimulation condition relative to sham (gray reference line, shaded
area represents SEM across participants). Dots represent the data points of single participants.
1246
Journal of Cognitive Neuroscience
Volume 32, Number 7
interhemispheric speech integration occurred (i.e., the
participants integrated the chirp and the contralateral
ambiguous base) during the sham blocks as we expected
(Figure 3A).
Figure 3B shows participants’ average performance for
each stimulation condition and the sham condition. In the
stimulation conditions, overall performance ranged on
average between 71.75% and 73.98%, whereas in the sham
condition, it was significantly better (76.16%; average dif-
ference: 3.46%), t(30) = 2.89, p = .007, d = 0.37. To test
whether the strength of this general stimulation effect
depended on the frequency or phase synchrony of the
TACS, a two-way repeated-measures ANOVA, including
the within-subject factors Stimulation Frequency (40 Hz,
3.125 Hz) and Interhemispheric Phase Synchronization
(in-phase, anti-phase) was conducted for the dependent
variable difference in interhemispheric integration of the
chirp as compared with sham stimulation. Delta values be-
tween the performance in each stimulation condition and
the sham condition were included in the analysis. Contrary
to our predictions, this analysis revealed no significant
interaction Stimulation Frequency × Interhemispheric
Phase Synchronization, F(1, 30) = 2.59, p = .12, ηp
2 =
.01, or main effects of Stimulation Frequency, F(1, 30) =
0.12, p = .73, ηp
2 = .0004, or Interhemispheric Phase
Synchronization, F(1, 30) = 0.20, p = .66, ηp
2 = .0008.
The lack of a main effect of Interhemispheric Phase
Synchronization implies no significant difference between
in-phase versus anti-phase stimulation.
To identify the specific TACS conditions under which the
stimulation effect occurred, a one-way repeated-measures
ANOVA, including the within-subject factors Stimulation
Condition (sham, in-phase 40 Hz, anti-phase 40 Hz, in-phase
3.125 Hz, anti-phase 3.125 Hz) was conducted for the de-
pendent variable the proportion of integrated trials. This
analysis revealed a significant main effect of Stimulation
Condition, F(4, 120) = 2.99, p = .02, ηp
2 = .02. Pairwise
comparisons revealed significantly reduced performance in
the in-phase 40-Hz condition, t(30) = −2.78, p = .049, FDR-
corrected, d = −0.37, and anti-phase 3.125-Hz condition,
t(30) = −2.76, p = .049, FDR-corrected, d = −0.40, com-
pared with sham stimulation, but not in the anti-phase
40-Hz condition, t(30) = −1.44, p = .32, FDR-corrected,
d = −0.22, or in-phase 3.125-Hz condition, t(30) =
−2.16, p = .13, FDR-corrected, d = −0.29. These results
indicate that the bihemispheric TACS modulated inter-
hemispheric speech integration.
DISCUSSION
In this study, we tested the hypothesis that interhemi-
spheric phase synchronization facilitates interhemispheric
speech integration. To test this, we applied TACS simulta-
neously above listeners’ left and right auditory speech
areas (either in-phase or anti-phase) to synchronize or de-
synchronize the two areas and measured the effect on
interhemispheric speech integration. Based on previous
evidence from electrophysiological studies (Steinmann
et al., 2014, 2018), interhemispheric integration of speech
might be causally related to phase synchronization of bilat-
eral auditory speech areas in the gamma frequency band.
No such effect has been reported for the delta frequency
band. Thus, we predicted that functional coupling of bilat-
eral auditory speech areas in the gamma, but probably not
in delta frequency band, would strengthen interhemi-
spheric speech integration, compared with functionally
decoupling them.
Our results show a reduction of interhemispheric in-
tegration under gamma TACS compared with sham stim-
ulation. This reduction was significant when gamma TACS
was applied in-phase above the two cerebral hemispheres.
We also observed a significant reduction when anti-phase
delta TACS was applied. We found no significant difference
between in-phase compared with anti-phase conditions for
either gamma or delta TACS. Although we found a general
reduction of performance during TACS versus sham stimu-
lation, we observed no main effect or interaction in an over-
all ANOVA comparing these reductions across the different
TACS conditions. However, the observed pattern of signif-
icant (in-phase gamma TACS, anti-phase delta TACS) and
nonsignificant (anti-phase gamma TACS, in-phase delta
TACS) changes in speech perception relative to sham stim-
ulation strongly suggests that TACS modulated interhemi-
spheric speech cue integration.
Contrary to our prediction, in-phase, not anti-phase,
gamma TACS perturbed interhemispheric speech cue in-
tegration. This finding implies that full interhemispheric
phase synchronization (0° difference) at 40 Hz is not ben-
eficial for interhemispheric speech cue integration. This
observation could be related to interindividual differences
in interhemispheric auditory transfer times (Henshall
et al., 2012). Strongest interhemispheric integration may
occur when gamma phase in the two hemispheres differs
in a manner commensurate with individual auditory trans-
fer times. This notion is supported by findings showing
that the auditory event-related N100 to dichotically pre-
sented syllables occurs at a different latency over the right
versus the left auditory cortex (Eichele, Nordby, Rimol, &
Hugdahl, 2005). The reported lag is on average 15 msec,
which closely matches with the half cycle duration of our
gamma TACS (12.5 msec). In line with this, a recent study
found that anti-phase TACS applied at 40 Hz does not
affect response laterality during dichotic listening (Meier
et al., 2019). Critically, the authors could show in a
follow-up analysis that, only in participants with intrinsic
gamma phase asymmetries closer to 0°, anti-phase gamma
TACS led to a reduction of interhemispheric integration,
that is, a shift in response laterality to right ear. These re-
sults corresponds well with our finding that anti-phase
gamma TACS, which imposes an interhemispheric lag
of 12.5 msec, may not perturb speech cue integration.
Our current experimental design does not allow further
testing this idea; this may be done in future studies that
Preisig et al.
1247
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
2
p
4
d
2
f
2
/
0
3
1
2
3
/
5
7
4
/
6
1
2
o
4
c
2
n
_
/
a
1
_
8
0
6
1
1
4
6
9
1
8
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
8
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
parametrically manipulate interhemispheric phase asyn-
chrony in multiple steps across the gamma cycle.
Other studies have reported that bilateral 40-Hz TACS
perturbs phonemic processing, decreasing discrimina-
bility of syllables with different VOTs in young adults
(Rufener, Zaehle, Oechslin, & Meyer, 2016) but increas-
ing it in older adults (Rufener, Oechslin, et al., 2016) and
in dyslexic individuals (Rufener, Krauel, Meyer, Heinze,
& Zaehle, 2019). Therefore, we cannot rule out that our
gamma TACS also affected local phoneme processing.
We positioned our electrodes so as to stimulate espe-
cially cortical speech areas in the lateral superior tempo-
ral lobe; therefore, we believe that the observed effect
originates from these areas. We cannot exclude that
other regions were stimulated by spreading current and
also contributed to the effect as our design did not in-
clude control regions. In addition to that, gamma TACS
might have affected deployment of attentional resources,
considering that unilateral 40-Hz TACS may affect perfor-
mance on dichotic working memory tasks ( Wöstmann,
Vosskuhl, Obleser, & Herrmann, 2018).
Surprisingly, our results suggest that not only gamma-
phase coupling but also delta-phase coupling plays a role
for interhemispheric speech cue integration. Our observa-
tion that anti-phase delta TACS perturbed behavioral per-
formance suggests that this type of stimulation disrupts
cross-lateral transfer of speech cues as well. Previous stud-
ies using dichotic stimulus presentation did not report
phase coupling in this frequency band (Steinmann et al.,
2014, 2018). Therefore, we speculate that anti-phase TACS
may have caused a difference in neural excitability be-
tween hemispheres during the processing of the binaural
input: When the current was positive over one site, it was
negative over the contralateral site, and vice versa. This
may have been particularly relevant for the delta TACS
condition, in which the applied current matched the
syllabic envelope. Increased neural excitability in one
hemisphere and decreased excitability in the other may
have resulted in an interhemispheric difference in the ef-
fectiveness with which the dichotic syllabic components
(chirp or ambiguous base) were processed. Indeed, trans-
cranial direct current stimulation has been shown to have
polarity-specific effects on temporal and spectral pro-
cessing of auditory input (Heimrath, Kuehne, Heinze, &
Zaehle, 2014; Schaal, Williamson, & Banissy, 2013; Zaehle,
Beretta, Jäncke, Herrmann, & Sandmann, 2011; Vines,
Schnider, & Schlaug, 2006).
An important additional consideration is that anti-
phase delta TACS may disrupt interhemispheric cross-
frequency dynamics between delta and gamma oscillations
during speech perception (Giraud & Poeppel, 2012).
Coupling of these frequency bands could be of particular
relevance for interhemispheric integration, because re-
gions in the left and right auditory cortex may be differ-
ently tuned with respect to these frequency bands, with
a relative leftward dominance of low-gamma neural oscil-
lations and/or rightward dominance of slow frequency
oscillations (Flinker, Doyle, Mehta, Devinsky, & Poeppel,
2019; Bouton et al., 2018; Giraud & Poeppel, 2012;
Saoud et al., 2012; Poeppel, 2003). In addition to this,
there is support that right hemispheric auditory process-
ing may be tuned for spectral information (Preisig &
Sjerps, 2019; Bouton et al., 2018) and left hemispheric
auditory processing may be tuned for temporal informa-
tion (Flinker et al., 2019; Saoud et al., 2012)—a theoretical
framework originally formulated in the asymmetric sam-
pling theory (Poeppel, 2003; for a similar framework,
see Zatorre & Belin, 2001). In a previous study, we found
that the laterality of initial chirp sound processing, that is,
the ear of presentation, did not influence participants’
perceptual decisions (Preisig & Sjerps, 2019). However,
stimulus laterality influenced the processing speed of
integration. Thus, we cannot rule out that the ear of
presentation contributes to the observed TACS effect.
Our current experimental design does not allow further
testing this idea; this may be done in future studies apply-
ing interhemispheric cross-frequency delta–gamma TACS
stimulation presenting the chirp to the left and the right
ear, respectively.
In summary, our results indicate that both gamma and
delta TACS affect interhemispheric speech integration,
but in different ways. The induced perturbations imply
that interhemispheric phase coupling plays a functional
role in interhemispheric speech integration.
Acknowledgments
This work was supported by the Swiss National Science Foun-
dation (P2BEP3_168728 /PP00P1_163726) and the Janggen-Pöhn
Stiftung. The authors would like to thank Brigit Knudsen, Iris
Schmits, and Sarah Kemp for their assistance.
Reprint requests should be sent to Basil C. Preisig, Donders
Institute for Brain Cognition and Behaviour, Radboud University,
P.O. Box 9101, Nijmegen, Gelderland 6500 HB, The Netherlands,
or via e-mail: basilpreisig@gmx.ch.
Note
1. To activate the right motor cortex, in line with a related on-
going neuroimaging study examining speech processing in the
left cerebral hemisphere.
REFERENCES
Bayazıt, O., Oniz, A., Hahn, C., Güntürkün, O., & Ozgören,
M. (2009). Dichotic listening revisited: Trial-by-trial ERP
analyses reveal intra- and interhemispheric differences.
Neuropsychologia, 47, 536–545.
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false
discovery rate: A practical and powerful approach to multiple
testing. Journal of the Royal Statistical Society: Series B:
Methodological, 57, 289–300.
Bouton, S., Chambon, V., Tyrand, R., Guggisberg, A. G., Seeck,
M., Karkar, S., et al. (2018). Focal versus distributed temporal
cortex activity for speech sound category assignment.
Proceedings of the National Academy of Sciences, U.S.A.,
115, E1299–E1308.
1248
Journal of Cognitive Neuroscience
Volume 32, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
2
p
4
d
2
f
2
/
0
3
1
2
3
/
5
7
4
/
6
1
2
o
4
c
2
n
_
/
a
1
_
8
0
6
1
1
4
6
9
1
8
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
8
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
t
f
o
n
0
5
M
a
y
2
0
2
1
Chang, E. F., Rieger, J. W., Johnson, K., Berger, M. S., Barbaro,
Mesgarani, N., Cheung, C., Johnson, K., & Chang, E. F. (2014).
N. M., & Knight, R. T. (2010). Categorical speech representation
in human superior temporal gyrus. Nature Neuroscience, 13,
1428–1432.
Phonetic feature encoding in human superior temporal
gyrus. Science, 343, 1006–1010.
Obleser, J., Zimmermann, J., Van Meter, J., & Rauschecker, J. P.
Eichele, T., Nordby, H., Rimol, L. M., & Hugdahl, K. (2005).
Asymmetry of evoked potential latency to speech sounds
predicts the ear advantage in dichotic listening. Cognitive
Brain Research, 24, 405–412.
Fell, J., & Axmacher, N. (2011). The role of phase
synchronization in memory processes. Nature Reviews
Neuroscience, 12, 105–118.
Flinker, A., Doyle, W. K., Mehta, A. D., Devinsky, O., & Poeppel,
D. (2019). Spectrotemporal modulation provides a unifying
framework for auditory cortical asymmetries. Nature Human
Behaviour, 3, 393–403.
Fries, P. (2005). A mechanism for cognitive dynamics: Neuronal
communication through neuronal coherence. Trends in
Cognitive Sciences, 9, 474–480.
Giraud, A. L., & Poeppel, D. (2012). Cortical oscillations and
speech processing: Emerging computational principles and
operations. Nature Neuroscience, 15, 511–517.
Gross, J., Hoogenboom, N., Thut, G., Schyns, P., Panzeri, S.,
Belin, P., et al. (2013). Speech rhythms and multiplexed
oscillatory sensory coding in the human brain. PLoS Biology,
11, e1001752.
Heimrath, K., Kuehne, M., Heinze, H. J., & Zaehle, T. (2014).
Transcranial direct current stimulation (tDCS) traces the
predominance of the left auditory cortex for processing of
rapidly changing acoustic information. Neuroscience, 261,
68–73.
Helfrich, R. F., Knepper, H., Nolte, G., Strüber, D., Rach, S.,
Herrmann, C. S., et al. (2014). Selective modulation of
interhemispheric functional connectivity by HD-tACS shapes
perception. PLOS Biology, 12, e1002031.
Henshall, K. R., Sergejew, A. A., McKay, C. M., Rance, G., Shea,
T. L., Hayden, M. J., et al. (2012). Interhemispheric transfer
time in patients with auditory hallucinations: An auditory
event-related potential study. International Journal of
Psychophysiology, 84, 130–139.
Hugdahl, K., & Westerhausen, R. (2016). Speech processing
asymmetry revealed by dichotic listening and functional brain
imaging. Neuropsychologia, 93, 466–481.
Jäncke, L. (2002). Does “callosal relay” explain ear advantage
in dichotic monitoring? Laterality: Asymmetries of Body,
Brain and Cognition, 7, 309–320.
Keitel, A., Ince, R. A. A., Gross, J., & Kayser, C. (2017).
Auditory cortical delta-entrainment interacts with
oscillatory power in multiple fronto-parietal networks.
Neuroimage, 147, 32–42.
Kimura, D. (1967). Functional asymmetry of the brain in
dichotic listening. Cortex, 3, 163–178.
Kösem, A., & Wassenhove, V. v. (2017). Distinct contributions
of low- and high-frequency neural oscillations to speech
comprehension. Language, Cognition and Neuroscience,
32, 536–544.
Liberman, A. M., & Mattingly, I. G. (1989). A specialization for
speech perception. Science, 243, 489–494.
Luo, H., & Poeppel, D. (2007). Phase patterns of neuronal
responses reliably discriminate speech in human auditory
cortex. Neuron, 54, 1001–1010.
Mathiak, K., Hertrich, I., Lutzenberger, W., & Ackermann, H.
(2001). Neural correlates of duplex perception: A whole-
head magnetencephalography study. NeuroReport, 12,
501–506.
Meier, J., Nolte, G., Schneider, T. R., Engel, A. K., Leicht, G., &
Mulert, C. (2019). Intrinsic 40Hz-phase asymmetries predict
tACS effects during conscious auditory perception. PLoS One,
14, e0213996.
(2007). Multiple stages of auditory speech perception
reflected in event-related fMRI. Cerebral Cortex, 17,
2251–2257.
Poeppel, D. (2003). The analysis of speech in different temporal
integration windows: Cerebral lateralization as ‘asymmetric
sampling in time’. Speech Communication, 41, 245–255.
Pollmann, S., Maertens, M., von Cramon, D. Y., Lepsien, J., &
Hugdahl, K. (2002). Dichotic listening in patients with
splenial and nonsplenial callosal lesions. Neuropsychology,
16, 56–64.
Preisig, B. C., & Sjerps, M. J. (2019). Hemispheric specializations
affect interhemispheric speech sound integration during
duplex perception. Journal of the Acoustical Society of
America, 145, EL190–EL196.
Preisig, B. C., Sjerps, M. J., Kösem, A., & Riecke, L. (2019). Dual-
site high-density 4 Hz transcranial alternating current
stimulation applied over auditory and motor cortical speech
areas does not influence auditory-motor mapping. Brain
Stimulation, 12, 775–777.
Rand, T. C. (1974). Dichotic release from masking for speech.
Journal of the Acoustical Society of America, 55,
678–680.
Riecke, L., Formisano, E., Herrmann, C. S., & Sack, A. T. (2015).
4-Hz transcranial alternating current stimulation phase
modulates hearing. Brain Stimulation, 8, 777–783.
Riecke, L., Formisano, E., Sorger, B., Başkent, D., & Gaudrain, E.
(2018). Neural entrainment to speech modulates speech
intelligibility. Current Biology, 28, 161–169.
Riecke, L., Sack, A. T., & Schroeder, C. E. (2015). Endogenous
delta/theta sound-brain phase entrainment accelerates the
buildup of auditory streaming. Current Biology, 25,
3196–3201.
Rimmele, J. M., Zion Golumbic, E., Schröger, E., & Poeppel, D.
(2015). The effects of selective attention and speech
acoustics on neural speech-tracking in a multi-talker scene.
Cortex, 68, 144–154.
Rufener, K. S., Krauel, K., Meyer, M., Heinze, H.-J., & Zaehle, T.
(2019). Transcranial electrical stimulation improves phoneme
processing in developmental dyslexia. Brain Stimulation,
12, 930–937.
Rufener, K. S., Oechslin, M. S., Zaehle, T., & Meyer, M. (2016).
Transcranial Alternating Current Stimulation (tACS)
differentially modulates speech perception in young and
older adults. Brain Stimulation, 9, 560–565.
Rufener, K. S., Zaehle, T., Oechslin, M. S., & Meyer, M. (2016).
40 Hz-Transcranial alternating current stimulation (tACS)
selectively modulates speech perception. International
Journal of Psychophysiology, 101, 18–24.
Saoud, H., Josse, G., Bertasi, E., Truy, E., Chait, M., & Giraud,
A. L. (2012). Brain-speech alignment enhances auditory
cortical responses and speech perception. Journal of
Neuroscience, 32, 275–281.
Saturnino, G. B., Madsen, K. H., Siebner, H. R., & Thielscher, A.
(2017). How to target inter-regional phase synchronization
with dual-site transcranial alternating current stimulation.
Neuroimage, 163, 68–80.
Schaal, N. K., Williamson, V. J., & Banissy, M. J. (2013).
Anodal transcranial direct current stimulation over the
supramarginal gyrus facilitates pitch memory. European
Journal of Neuroscience, 38, 3513–3518.
Shamir, M., Ghitza, O., Epstein, S., & Kopell, N. (2009).
Representation of time-varying stimuli by a network
exhibiting oscillations on a faster time scale. PLoS
Computational Biology, 5, e1000370.
Preisig et al.
1249
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
2
p
4
d
2
f
2
/
0
3
1
2
3
/
5
7
4
/
6
1
2
o
4
c
2
n
_
/
a
1
_
8
0
6
1
1
4
6
9
1
8
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
8
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
/
t
o
n
0
5
M
a
y
2
0
2
1
Sparks, R., & Geschwind, N. (1968). Dichotic listening in man
after section of neocortical commissures. Cortex, 4, 3–16.
Steinmann, S., Leicht, G., Ertl, M., Andreou, C., Polomac, N.,
Westerhausen, R., et al. (2014). Conscious auditory
perception related to long-range synchrony of gamma
oscillations. Neuroimage, 100, 435–443.
Steinmann, S., Meier, J., Nolte, G., Engel, A. K., Leicht, G., &
Mulert, C. (2018). The callosal relay model of
interhemispheric communication: New evidence from
effective connectivity analysis. Brain Topography, 31,
218–226.
Ten Oever, S., de Graaf, T. A., Bonnemayer, C., Ronner, J., Sack,
A. T., & Riecke, L. (2016). Stimulus presentation at specific
neuronal oscillatory phases experimentally controlled with
tACS: Implementation and applications. Frontiers in Cellular
Neuroscience, 10, 240.
Thielscher, A., Antunes, A., & Saturnino, G. B. (2015). Field
modeling for transcranial magnetic stimulation: A useful tool
to understand the physiological effects of TMS? In Proceeding
of the Engineering in Medicine and Biology Society (EMBC),
2015 37th Annual International Conference of the IEEE, pp.
222–225.
Turi, Z., Csifcsák, G., Boayue, N. M., Aslaksen, P., Antal, A.,
Paulus, W., et al. (2019). Blinding is compromised for
transcranial direct current stimulation at 1 mA for 20 min in
young healthy adults. European Journal of Neuroscience,
50, 3261–3268.
Vines, B. W., Schnider, N. M., & Schlaug, G. (2006). Testing for
causality with transcranial direct current stimulation: Pitch
memory and the left supramarginal gyrus. NeuroReport, 17,
1047–1050.
Vossen, A., Gross, J., & Thut, G. (2015). Alpha power increase
after transcranial alternating current stimulation at alpha
frequency (α-tACS) reflects plastic changes rather than
entrainment. Brain Stimulation, 8, 499–508.
Westerhausen, R., Grüner, R., Specht, K., & Hugdahl, K. (2009).
Functional relevance of interindividual differences in
temporal lobe callosal pathways: A DTI tractography study.
Cerebral Cortex, 19, 1322–1329.
Westerhausen, R., & Hugdahl, K. (2008). The corpus callosum
in dichotic listening studies of hemispheric asymmetry: A
review of clinical and experimental evidence. Neuroscience &
Biobehavioral Reviews, 32, 1044–1054.
Wöstmann, M., Vosskuhl, J., Obleser, J., & Herrmann, C. S.
(2018). Opposite effects of lateralised transcranial alpha
versus gamma stimulation on auditory spatial attention.
Brain Stimulation, 11, 752–758.
Zaehle, T., Beretta, M., Jäncke, L., Herrmann, C. S., &
Sandmann, P. (2011). Excitability changes induced in the
human auditory cortex by transcranial direct current
stimulation: Direct electrophysiological evidence.
Experimental Brain Research, 215, 135–140.
Zaehle, T., Lenz, D., Ohl, F. W., & Herrmann, C. S. (2010).
Resonance phenomena in the human auditory cortex:
Individual resonance frequencies of the cerebral cortex
determine electrophysiological responses. Experimental
Brain Research, 203, 629–635.
Zatorre, R. J., & Belin, P. (2001). Spectral and temporal
processing in human auditory cortex. Cerebral Cortex, 11,
946–953.
Zoefel, B., Allard, I., Anil, M., & Davis, M. H. (2020). Perception
of rhythmic speech is modulated by focal bilateral
transcranial alternating current stimulation. Journal of
Cognitive Neuroscience, 32, 226–240.
Zoefel, B., Archer-Boyd, A., & Davis, M. H. (2018). Phase
entrainment of brain oscillations causally modulates neural
responses to intelligible speech. Current Biology, 28, 401–408.
Zoefel, B., Davis, M. H., Valente, G., & Riecke, L. (2019). How to
test for phasic modulation of neural and behavioural
responses. Neuroimage, 202, 116175.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
2
p
4
d
2
f
2
/
0
3
1
2
3
/
5
7
4
/
6
1
2
o
4
c
2
n
_
/
a
1
_
8
0
6
1
1
4
6
9
1
8
8
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
9
0
8
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
.
t
o
n
0
5
M
a
y
2
0
2
1
1250
Journal of Cognitive Neuroscience
Volume 32, Number 7