Are Color Experiences the Same across the Visual Field?
Ariel Zeleznikow-Johnston1
, Yasunori Aizawa2,3
Makiko Yamada2, and Naotsugu Tsuchiya1,4,5
,
Abstracto
■ It seems obvious to laypeople that neurotypical humans
experience color equivalently across their entire visual field.
To some neuroscientists, psychologists, and philosophers,
aunque, this claim has been met with skepticism, as neurophys-
iological evidence indicates the mechanisms that support color
perception degrade with eccentricity. Sin embargo, the argument
that this entails altered color experience in peripheral vision
is not universally accepted. Aquí, we address whether color
experience is essentially equivalent between central and periph-
eral vision. To assess this, we will obtain similarity relationships
between color experiences across the visual field using both
online and laboratory-based far-field displays, while removing
the confounds of saccades, memory, and expectation about
color experiences. Our experiment was designed to provide
clear evidence that would favor either unchanged or altered
color experience relationships in the periphery. Nuestros resultados
are consistent with lay people’s phenomenological reports:
Color experiences, as probed by similarity relationships in
central vision and the far field (60°), are equivalent when elic-
ited by large stimuli. These findings challenge the widespread
view in philosophy and cognitive science that peripheral color
experiences are illusory, and are discussed in the context of
their related neurophysiological, psychophysical, and philo-
sophical literature. ■
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INTRODUCCIÓN
It seems obvious to laypeople that neurotypical humans
experience color across their entire visual field. Any dis-
tinct point in the visual field has some color associated
with it, without a sense that the possible colors one could
experience differs at different locations. To neuroscien-
tists though, this claim is sometimes met with skepticism
because of differences in perceptual performance across
the visual field. Performance on tasks using small fixed-
size stimuli decreases with eccentricity (Strasburger,
Rentschler, & Juttner, 2011), and the required color con-
trast for differences to be detected increases (Hansen,
Pracejus, & Gegenfurtner, 2009). There are associated
neurophysiological changes with increasing retinal
eccentricity, such as declines in the density of color-
sensitive photoreceptors, cones (Curcio, Lento, Kalina,
& Hendrickson, 1990). This behavioral and physiological
evidence is consistent with the claim that abilities to per-
ceive color differ across the visual field. Sin embargo, alguno
researchers go even further, believing that color percep-
tion is not just different but actually degraded in periph-
eral vision compared with foveal vision, and that this
entails a degraded experience of color in the periphery.
1Monash University, Melbourne, Victoria, Australia, 2National
Institutes for Quantum and Radiological Science and Technol-
ogia, Chiba, Japón, 3Tohoku University, Sendai, Japón, 4National
Institute of Information and Communications Technology
(NICT), Suita, Japón, 5Advanced Telecommunications Research
Institute International, Kioto, Japón
© 2023 Instituto de Tecnología de Massachusetts
Comments such as “perceptual experience lacks a sur-
prising amount of color” (cohen & Rubenstein, 2020),
“introspection [about peripheral color] cannot be truly
reliable” (Giron, Lau, & Knotts, 2018), “our intuitive sense
of a rich, colorful visual world is largely incorrect” (cohen,
Botch, & Robertson, 2020), and “it seems that our color
vision goes right way out to the edge of our vision. Él
doesn’t.” (Carroll & Dennett, 2020) are common. Estos
authors believe that the unreliable nature of introspection
into perception should lead us to be skeptical of reports
about conscious experience of peripheral color. We seek
to test the validity of these color-sceptic claims by deter-
mining whether color experiences are equivalent across
the visual field.
The most dramatic support for the skeptical view comes
from a recent study showing that people can be entirely
unaware of gradual, yet complete, removal of peripheral
visual color inputs under seminaturalistic viewing condi-
ciones (Cohen et al., 2020). Using a virtual-reality setup,
the authors demonstrated that the majority of people do
not notice when color is removed from eccentricities
greater than 20 degrees of visual angle (DVA). Más
support comes from studies that asked participants to
assign a color to a given stimulus presented for short dura-
ciones (380–1000 msec) at varying eccentricities. Using the
same stimuli presented centrally or peripherally, informes
of a roughly 50% decrease in perceived saturation at
alrededor 40 DVA are observed, as well as shifts in the
assigned hue and increased contrast required for detec-
ción (Hansen et al., 2009; McKeefry, Murray, & Parry,
Revista de neurociencia cognitiva 35:4, páginas. 509–542
https://doi.org/10.1162/jocn_a_01962
2007; Ayama, Sakurai, Carlander, Derefeldt, & Eriksson,
2004; Sakurai, Ayama, & Kumagai, 2003), although periph-
erally presented blue stimuli can increase in perceived
saturation (Vanston & Crognale, 2018). This behavioral work
is supported by neurophysiological studies showing that
increasing retinal eccentricity is associated with decreasing
density of cones (Curcio et al., 1990) and retinal ganglion
cell density (Curcio & allen, 1990) as well as a decline in
cortical resource allocation (Daniel & Whitteridge, 1961).
The sceptic’s case is clear: The evidence indicates that
peripheral color experiences are degraded or absent,
and the naive introspection is unreliable.
Todavía, support for the naive view of color experience
across the visual field can be found in much of the same
literature that sceptics cite. The same studies showing
desaturation and hue-shifts with constant-size stimuli
shown centrally and peripherally also show that if the
stimuli are magnified in size as they increase in eccentricity,
participants assign stimuli a consistent color (Hansen et al.,
2009; Ayama et al., 2004; Sakurai et al., 2003; Abramov,
gordon, & chan, 1991; gordon & Abramov, 1977). Allá
is also corresponding neurophysiological support for color
experience across the visual field. Although cone densities
decline dramatically with eccentricity, decreasing the
resolution of incoming perceptual information, the retinal
ganglion cells that pool photoreceptor inputs compensate
for this with increasingly large receptive fields, ensuring
color information is still available for downstream process-
En g (Curcio & allen, 1990). Similarmente, the axonal projection
zones of these retinal ganglion cells into the lateral genic-
ulate nucleus and then to the primary visual cortex become
larger with increasing eccentricity (Lennie, 1998). Estos
neurophysiological findings do suggest that peripheral
color perception is different from central perception in
terms of acuity and contrast sensitivity, yet while also
making plausible the naive case for color experiences
across the visual field (Haun, 2021).
Resolving the conflict between these two positions
requires a definition of what it would mean for color expe-
riences to “feel the same” across the visual field. The major-
ity of traditional color psychophysics research does not
address this, instead focusing on related questions. Uno
such example is “under what conditions can the same
physical stimulus induce different color experiences at
different locations?" (Figura 1A). Although important, él
provides data only on the stimulus–experience relation-
barco, rather than the experience–experience relationship,
which we are after here. In addition to the material
described above, there are data to suggest that the same
stimulus can elicit a different stimulus–experience relation-
ship even at different retinal locations of the same eccen-
tricity (Afraz, Pashkam, & Cavanagh, 2010).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
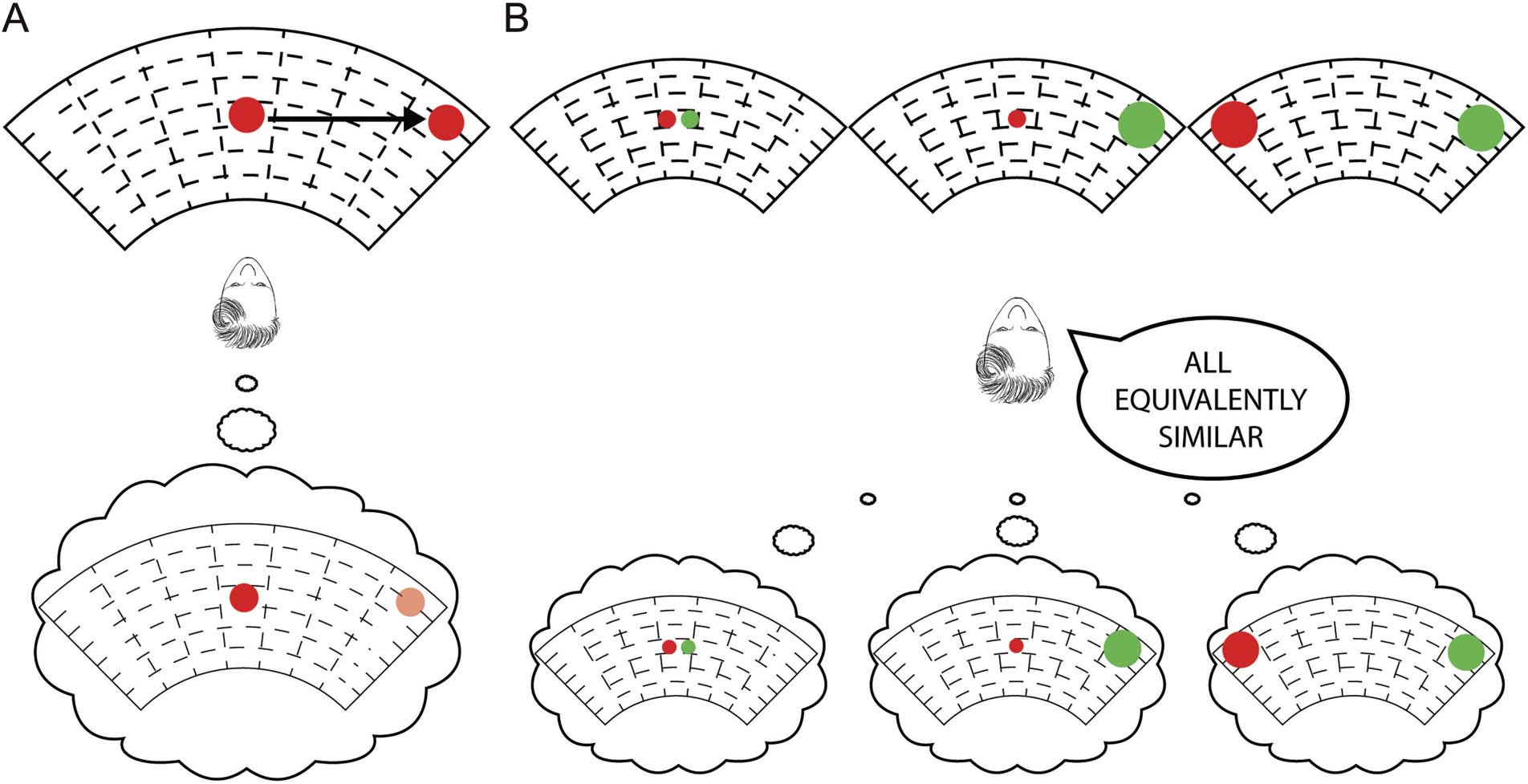
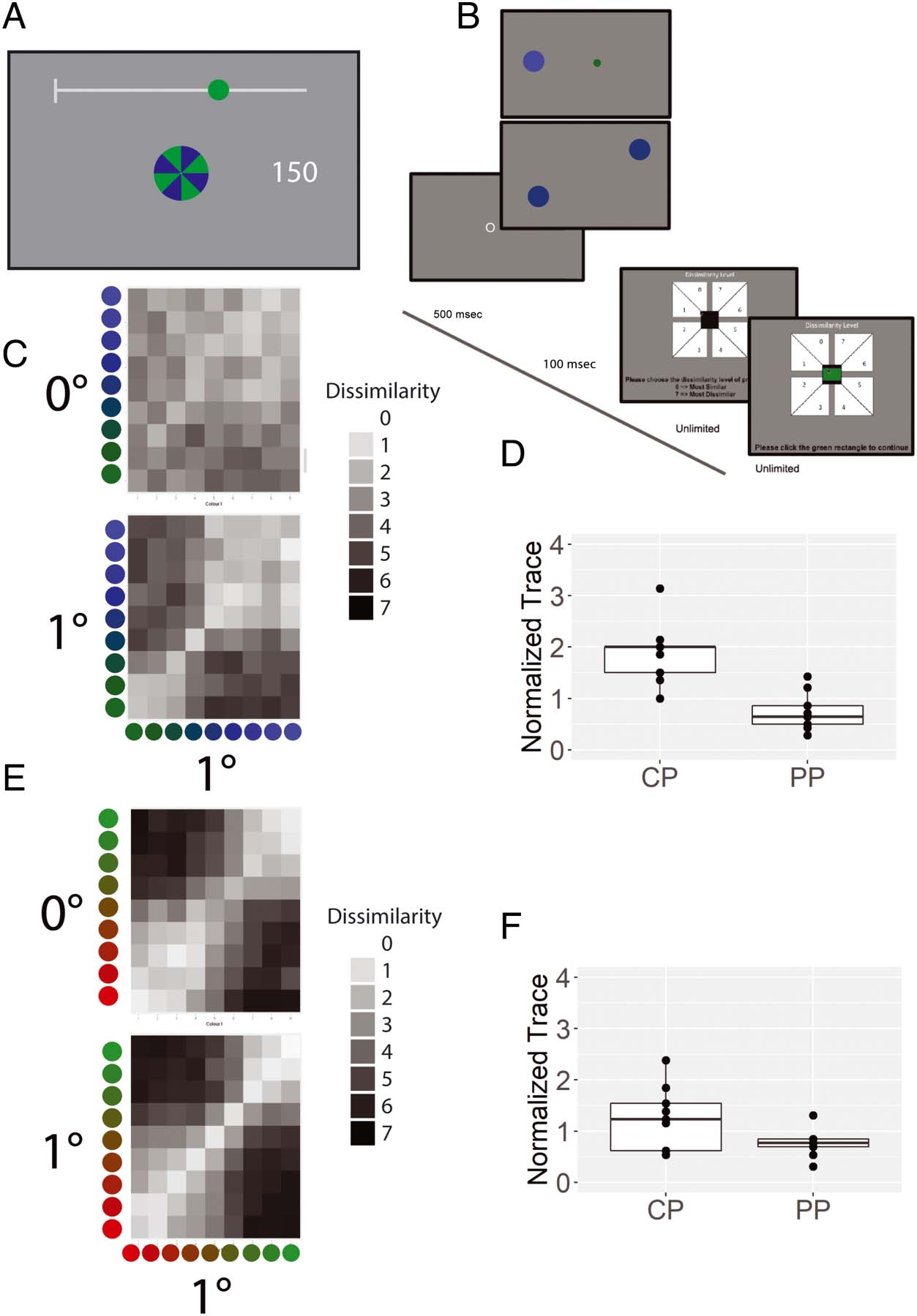
Cifra 1. Traditional psychophysics and a novel equivalence test of color experience equivalence. There has long been controversy as to whether
color experiences are “the same” across the visual field. The controversy originates from two issues: (1) how to measure color experience and
(2) how to define “the same.” In this article, we offer novel solutions to both of these issues. (A) A majority of traditional psychophysics has been
concerned with whether “physically” identical stimuli generate the “same” color experience when presented at different retinal locations. Estos
studies often used isolated instances of a single stimulus presented at a time, to which participants responded with “same/different responses.”
The results from this line of research tell us that small peripheral stimuli appear desaturated under certain conditions, demonstrating some
“physically same” stimuli can be perceived as “different.” (B) A diferencia de, our novel approach is concerned with whether “the structure of color
experiences” differs at different visual field locations. To characterize the structure, we focus on the similarity relationships between a set of color
experiences at different locations, with one possible pair out of the set shown here.
510
Revista de neurociencia cognitiva
Volumen 35, Número 4
A diferencia de, we are seeking to answer “whether color
experience relationships are the same or not at differ-
ent locations in the visual field” (Figura 1B). We have pre-
viously suggested that characterization of an experience in
relation to other possible experiences a participant could
be having is sufficient to assess the notion of subjective
equivalence that is meant by “the experiences felt the
same” (Fink, Kob, & Lyre, 2021; Tsuchiya & Saigo, 2021).
To operationalize the concept of “sameness” and charac-
terize experience relationships, we will make use of “sub-
jective equivalence tests.” One possible equivalence test is
the systematic pairwise comparison of the similarity
between experiences at different visual locations. If com-
paring Experiences A or B to every other possible pairing
of experiences results in sets of similarity judgments that
are statistically indistinguishable, then A and B can be con-
sidered equivalent. This equivalence test will allow us to
capture and compare aspects of experiences at different
visual locations in a systematic manner. Conceptualizing
experiences in this manner is becoming increasingly
prominent in consciousness science (Fink et al., 2021).
Además, this relational approach is also related to rep-
resentational similarity analysis of neural representations,
a prominent method for linking brain activity to behavior
(Kriegeskorte & Kievit, 2013). Aquí, we focus on the sim-
ilarity relationships among color experiences in central
and peripheral vision and test whether they are equiv-
alente, so as to assess whether the experiences themselves
differ across the visual field.
Partial inspiration for this approach comes from similar-
ity experiments that have been used previously to build
geometric models of color experience. The earliest publi-
cation was performed by Helm (1964), who asked partici-
pants to place colored chips at distances from other chips
that were proportional to their subjective similarity. Unfor-
tunately for the question at hand, no attempt was made to
fix the retinal stimulus size or duration of presentation to
preclude the possible eye movements. These consider-
ations have also been neglected in more recent studies
(Bonnardel et al., 2016; Quemaduras & Shepp, 1988). Work using
another paradigm to obtain subjective similarity between
colores, asking participants to provide a numerical dissimi-
larity rating between two color patches, has also ignored
the eccentricity of presentation. These studies have typi-
cally been performed with large stimuli (2–6 DVA), usando
both limited (p.ej., 500 mseg) and unlimited duration pre-
sentaciones (Bosten, robinson, Jordán, & Mollon, 2005;
Izmailov & Sokolov, 1991). Although this research pro-
vides support for a 3-D geometric model (p.ej., HSV,
CIELAB) of color experience in central vision, it cannot
answer whether this model is valid for experiences across
the visual field.
Además, the previously described work examining
color perception in the periphery (Ayama et al., 2004;
Sakurai et al., 2003; gordon & Abramov, 1977) cannot
be used to assess whether the similarity relationships
between color experiences changes across the visual field.
This is because, in these studies, participants were forced
to report their peripheral color experiences according
to the 3-D model developed for relationships between
central color experiences, without verifying that this
model was appropriate for the periphery. There are two
main shortcomings of this approach. The first is that par-
ticipants were constrained to respond by clicking on
points in a 3-D space. This may well have influenced par-
ticipant responses, as the predefined space of possible
responses already assumes the relationships between
the colors. This constraint matters, as it has been shown
that responses are different if participants are instructed
to provide responses through the framework of color
opponency compared with if they are uninstructed (Ennis
& Zaidi, 2019). Segundo, it was not reported whether par-
ticipants fully endorsed the responses that they provided
or whether the responses provided merely had the lowest
possible error given the reporting method’s constraints.
The relevance of this question is demonstrated by analo-
gous experiments examining stimuli similarity under
different illumination conditions. En estos, Participantes
are simultaneously able to report which achromatic color
stimuli match “best” while denying that any possible
stimuli adjustments can make the experiences equivalent
when presented in different illumination contexts
(Logvinenko & Maloney, 2006). This could not have been
detected if the methods of the previous peripheral color
experiments described above had been used. Addressing
these issues requires comparing the similarity of color
experiences at different eccentricities without prespeci-
fying the color relationships.
We believe that a combination of three methods can be
used to overcome the limitations of these prior studies. El
first is to determine whether dissimilarity judgments for
color experiences at different visual locations are
correlacionado, which would indicate their structural equiva-
lence. The second is to directly ask participants whether
color experiences generated by identical stimuli presented
at different locations are maximally similar (zero dissimilar-
idad), which more directly establishes their equivalence. El
third is to assess whether non-identical color pairs have the
same similarity relationships at different locations, cual
establishes their experiential equivalence. If all of these
are answered in the affirmative, it would establish the
equivalence of the experiences at different locations.
This present study seeks to use these three methods to
determine whether color experiences are equivalent
across the visual field. We will do so by obtaining and
analyzing the relationships between color experiences at
varying eccentricities. Específicamente, we will collect similarity
ratings by pairwise comparison of experiences generated
by stimuli presented either both in the central visual field
(CC), one central and one peripheral (CP), or both periph-
eral (PÁGINAS). To reduce the concern of possible eye move-
ments as well as cognitive effects such as expectation,
we will present two color patches for brief durations
(250 mseg) at unpredictable locations. Each participant
Zeleznikow-Johnston et al.
511
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
will rate the similarity between every combination of
stimuli and eccentricity to allow for full determination of
the color experience relationships with respect to the
chosen set of stimuli. Además, to relate our findings
on color experience relationships to that of the previous
work on perceptual changes with eccentricity, Lo haremos
determine whether similarity judgments are affected by
the size of the stimuli used to generate color experiences.
En particular, we will examine whether keeping the size of
the stimuli in the periphery results in these stimuli
inducing altered color experience structures with respect
to central vision. To buttress our novel methodology,
we performed three control experiments described
in Appendix D. They establish the sensitivity and re-
liability of our methods (p.ej., robustness against display
inhomogeneity). They also demonstrate that our methods
can demonstrate the expected phenomenological
consequences of a lack of S-cones at the central fovea
(Magnussen, Spillmann, Stürzel, & Werner, 2004).
If the structure of dissimilarity relationships between
color experiences has any degree of equivalence across
the visual field, then participants should provide corre-
lated dissimilarity judgments for sets of color stimuli
shown at different eccentricities. Para cada participante,
we will obtain the correlation between their judgments
for each of the eccentricity conditions, eso es, CC to CP,
CC to PP, and CP to PP. We hypothesize that the mean
across-participants correlation will be positive for each of
these comparisons (H1A–C).
Correlation can establish one notion of structural equiv-
alence, but given it is insensitive to uniform changes in
judgments, it alone is insufficient to establish experiential
equivalence across the visual field. Additional tests involve
directly examining the magnitudes of the dissimilarity
respuestas. Por ejemplo, do identical color stimuli elicit
completely similar experiences when presented at differ-
ent visual field locations? If color experiences either do not
existir, or are substantially degraded or altered in the
periphery, then participants should provide the same dis-
tribution of responses for pairs of both physically identical
and non-identical color pairs when one or both of the
patches are presented at the periphery. Alternativamente, si
color experiences exist in the periphery and physically
identical stimuli can elicit the same experience both cen-
trally and peripherally, it should be possible to distinguish
between the distribution of similarity responses given for
identical versus non-identical stimuli pairs shown periph-
erally. Para cada participante, we will obtain the mean
dissimilarity response for both physically identical and
non-identical stimuli shown in each of the CC, CP, y
PP conditions. The difference in the mean ratings for
identical versus non-identical patches will be used to
assess subjective equivalence as per the following hypoth-
eses: Participants presented with peripherally magnified
stimuli will provide a mean dissimilarity difference rating
greater than zero (H2A–C) or alternatively close to zero,
indicating no difference (H0).
Además, we suspect that small stimuli shown in
the periphery without peripheral magnification may
not induce color experiences that are equivalent to
when the same stimuli are presented centrally. Para probar esto,
we will use a 2 × 2 design altering stimulus size and
peripheral magnification and observing the resulting
mean ratings. We hypothesize that there will be an
interaction effect on the normalized traces provided in
the PP conditions (H2D).
Not only should identical stimuli elicit the same dissim-
ilarity responses if color experience structures are equiva-
lent across the visual field, but so too should any pair of
color experiences, identical or non-identical, elicit the
same dissimilarity response regardless of eccentricity.
We will analyze whether this is indeed the case by examin-
ing whether participants make the same dissimilarity
response for a color stimulus pair across the CC, CP, y
PP trial conditions. We will quantify this by examining the
between-conditions response variance for each color pair.
If participants provide the same response for a given color
pair under all three conditions, the variance in dissimilarity
across those trials is zero. We will calculate variance in dis-
similarity response between the three trial conditions
across all color pairings for each participant and use the
per-participant mean variance as the outcome measure.
A diferencia de, if the similarity between color pairs is inconsis-
tent between CC, CP, and PP trials, then the variance in
dissimilarity across the three trials would be large. En esto
caso, shuffling the correspondence between color-pair
labels and a participant’s similarity judgments for their
CP and PP trials would have no effect on the mean
variance, as their judgments would already be inconsis-
tent across eccentricity conditions. We will examine if
participants presented with peripherally magnified stim-
uli will provide consistent judgments across eccentricity
conditions by assessing whether the difference in the
mean variance of their judgments is greater than zero
(H3), or whether they are inconsistent with no difference
in mean variance between the original and scrambled
judgments (H0).
All specific hypotheses are listed in the Hypothesis
Summary Table (Table A1).
We have initially validated our methodology through
online versions of both our preregistered experiment and
replications of previously reported findings (Apéndice D).
After performing screen size and viewing distance
calibrations, participants performed successive compari-
sons between color experiences generated by stimuli
shown either centrally (1 DVA) or peripherally (10 DVA).
Although the peripheral eccentricity was limited with
respect to the maximal extent of human peripheral vision,
this is still well outside the fovea, which comprises only a
couple of degrees either side of the center of vision. Para
our preregistered experiments, we will expand on this
pilot work by performing the same experiments but at a
peripheral eccentricity of 60 DVA in a laboratory setting.
All other aims and hypotheses remain the same.
512
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
MÉTODOS
En esta sección, we provide the methods of the preregis-
tered in-laboratory experiment. For our online pilot exper-
elementos, see Appendix D.
Ética
All participants provided written informed consent before
participating in the laboratory study, which has been
approved by the Ethics Committee of the National
Institute of Quantum Science and Technology. Allá
are no conflicts of interest to declare. Los participantes fueron
compensated for their time at a rate of ¥5000 JPY/day.
Diseño
Participantes
Participants were obtained through responses to an e-mail
list of registered participants held by the institute. Participe-
pants provided written consent before the commence-
ment of the experiments.
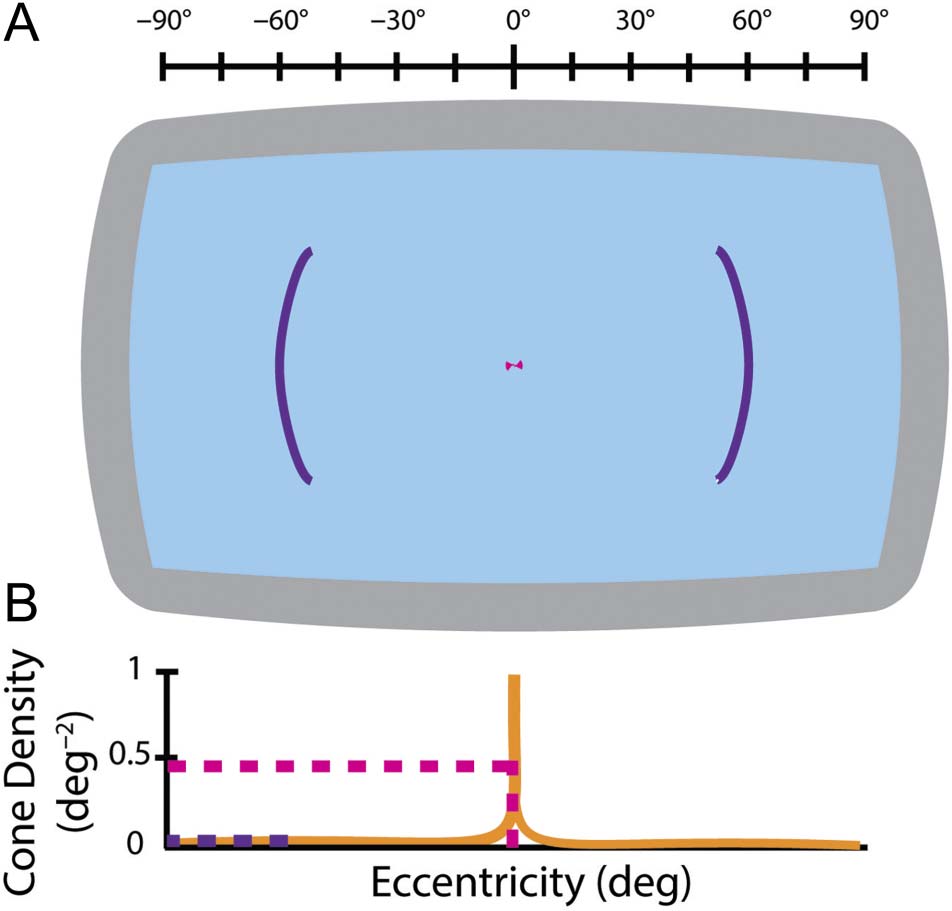
Display Apparatus
Stimuli were presented on a Panoworks display (Orihal-
con Technologies). Participants were seated 1359 mm
away from the center of the screen, providing a screen
tamaño de 180 × 109.4 DVA (Figure A1). Stimuli were pre-
sented via Inquisit 6 (Millisecond) display software.
Estímulos
All stimuli were presented on a gray background corre-
sponding to [0, 0, 0.5] in HSV color space. Usamos un
stimulus set of nine fully saturated and luminant colors
with maximal hue spacing (see Table B1 for full specifica-
ción). All stimuli were presented as solid-colored circles
that vary in position and size based on task conditions.
All specifications of position are made with reference to
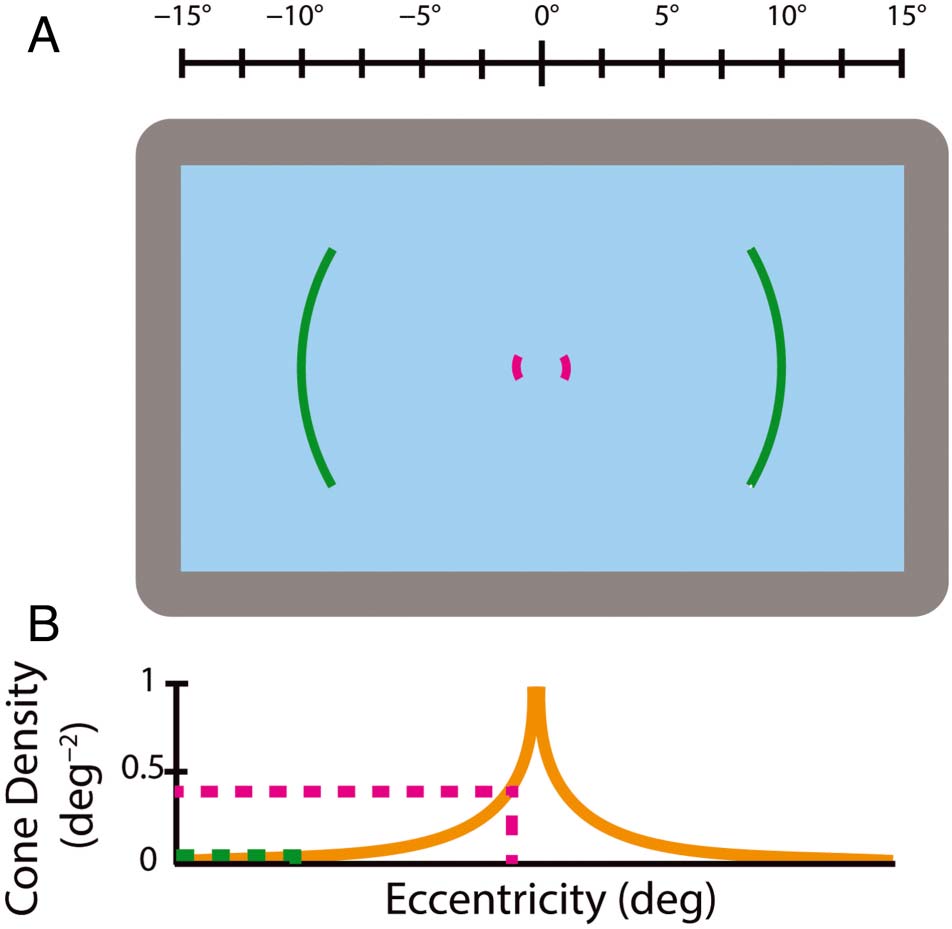
the center of the screen (Figure 2A and B).
Centrally presented color stimuli were presented with
their center at 1 DVA to avoid overlap. As the fovea extends
to approximately 2 DVA and the macula to approximately
9 DVA, these stimuli are well within central vision. Periph-
eral stimuli were centered at 60 DVA, well outside of foveal
visión (Figura 2B).
In experimental conditions with peripherally magnified
estímulos, peripheral stimuli were scaled relative to central
stimuli according to the average size of the V2 receptive
field increase relative to central vision (Hombre libre &
Simoncelli, 2011). Por ejemplo, this accounts for a 4.5×
increase in radius when presenting a stimulus at 10 DVA
versus 1 DVA, or a 27× increase at 60 DVA. The diameter
of the central stimuli was 0.1 DVA for the small stimulus
y 2 DVA for the large stimulus. On any given trial, el
stimuli are randomly selected to be centered anywhere
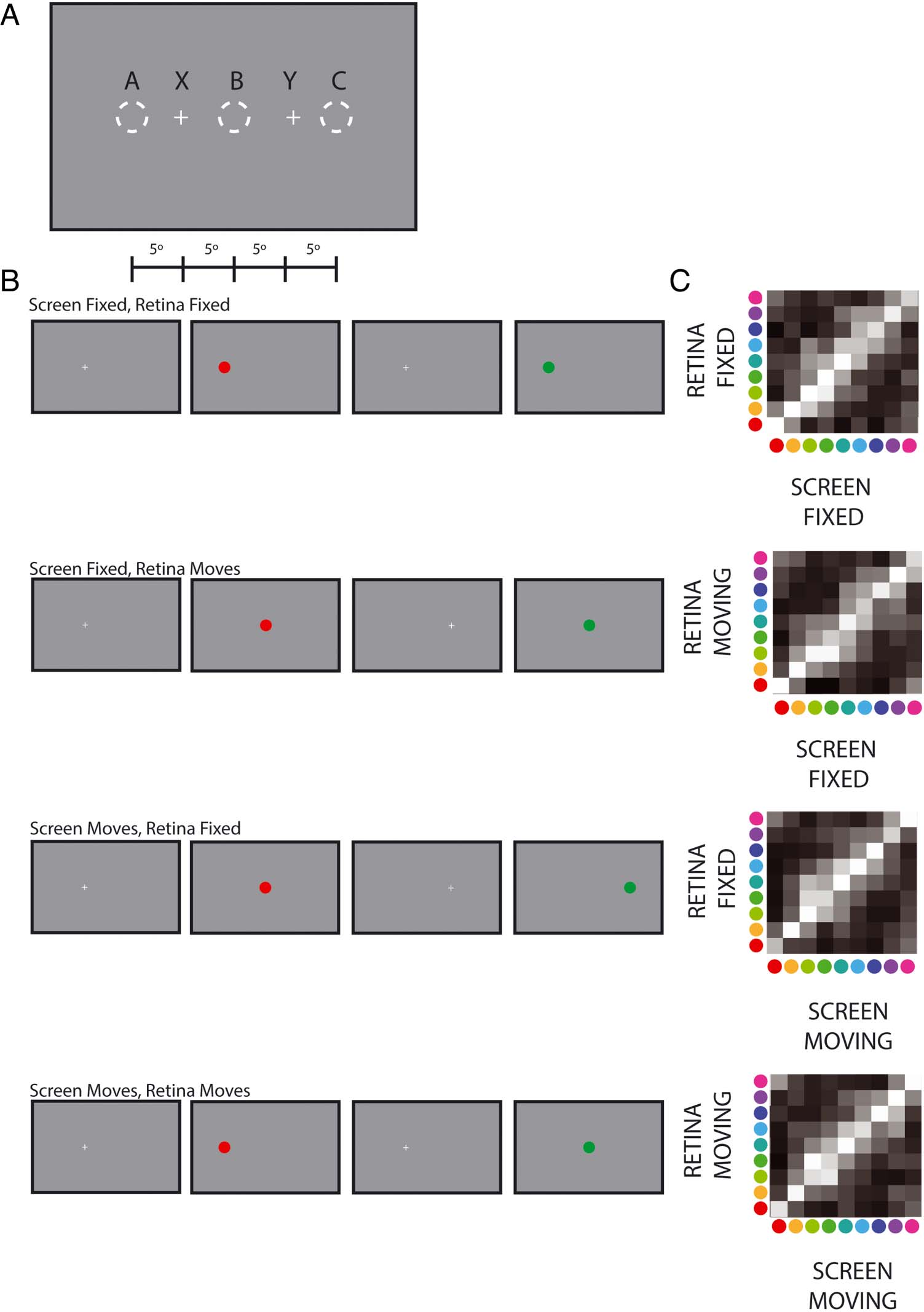
Cifra 2. (A) Schematic of the achievable eccentricities of stimuli using
our Panoworks display. Central and peripheral stimuli are presented at
random locations centered on their appropriate eccentricity, cual es
shown schematically by the pink (1 DVA) and violet (60 DVA) líneas,
respectivamente. See Figure A1 for more details. (B) Cone photoreceptor
densities at central and peripheral eccentricities relative to the retinal
maximum for the range of the Panoworks display ( watson, 2014;
Curcio et al., 1990). Eccentricities for (A) y (B) are shown by the scale
arriba (A).
on the circle that corresponds to their appropriate eccen-
tricity so long as the stimulus is not occluded by the edge
of the screen (shown as colored areas in Figure 2A).
Stimuli were always presented opposite each other (180°
apart in polar angle).
Procedimiento
Instructions. After consent and screen size calibrations,
participants were shown a sequence of task instructions
(for the online pilot version, the demo is available here:
https://mili2nd.co/76ub). Inicialmente, they are presented with
an animation of what the stimulus display will look like and
instructed about what they will need to fixate on in the
center of the screen. On the next page, they are informed
that they will need to rate the similarity of the two stimuli
presented on each trial and asked to ignore any differ-
ences in size and location between the stimuli. They were
told they need to provide a judgment between 0 (mayoría
similar/least different) y 7 (least similar/most different)
by clicking the appropriate integer, where the numbers
are displayed in a circular fashion around the center of
the screen after stimuli presentation (Cifra 3). Participe-
pants were also encouraged to use the full range of possi-
ble values when providing responses. On this same
pantalla, they are also presented with an animation of
responses being provided on the response screen. Después
they have made their choice, they click on the center of
Zeleznikow-Johnston et al.
513
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
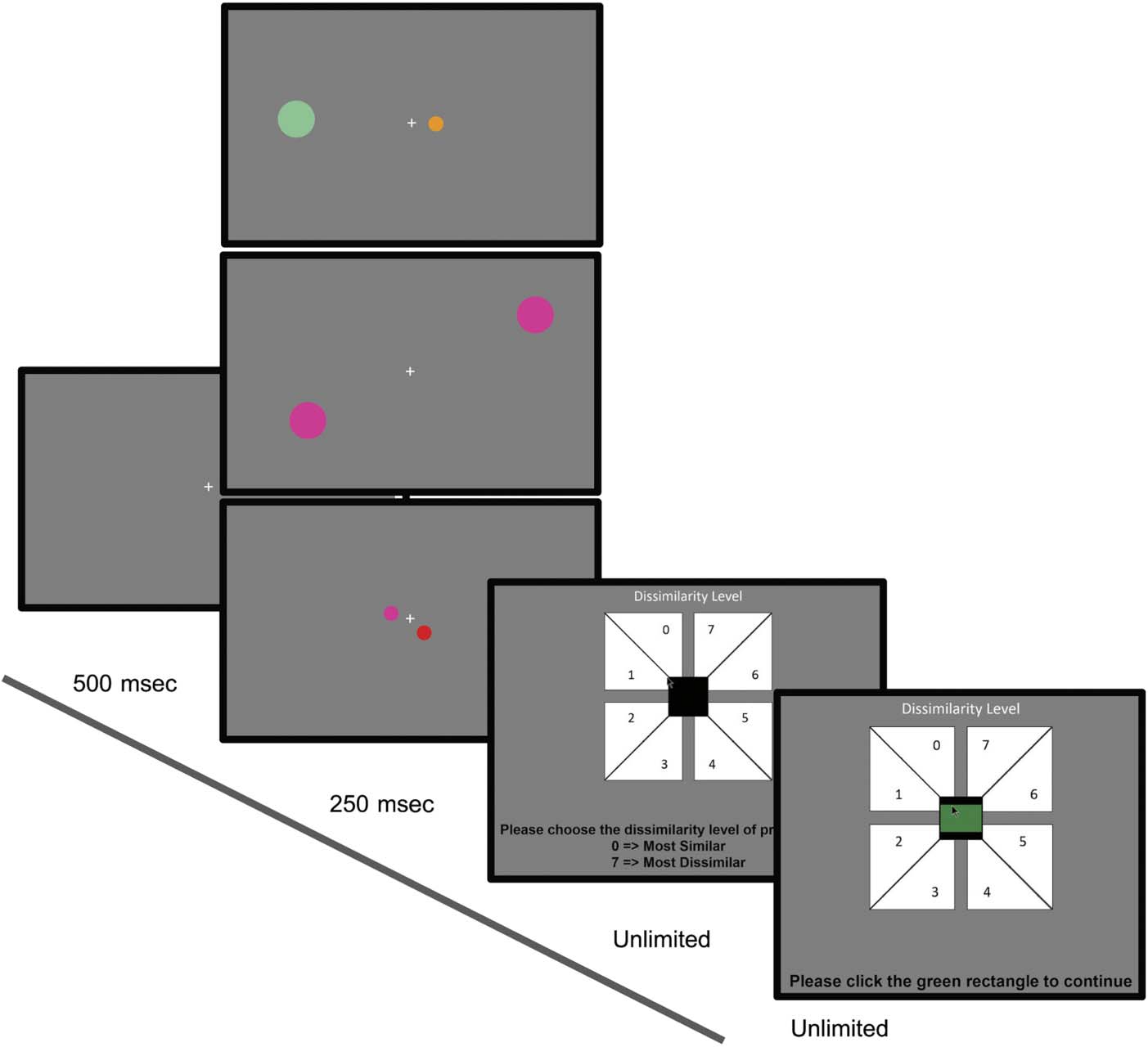
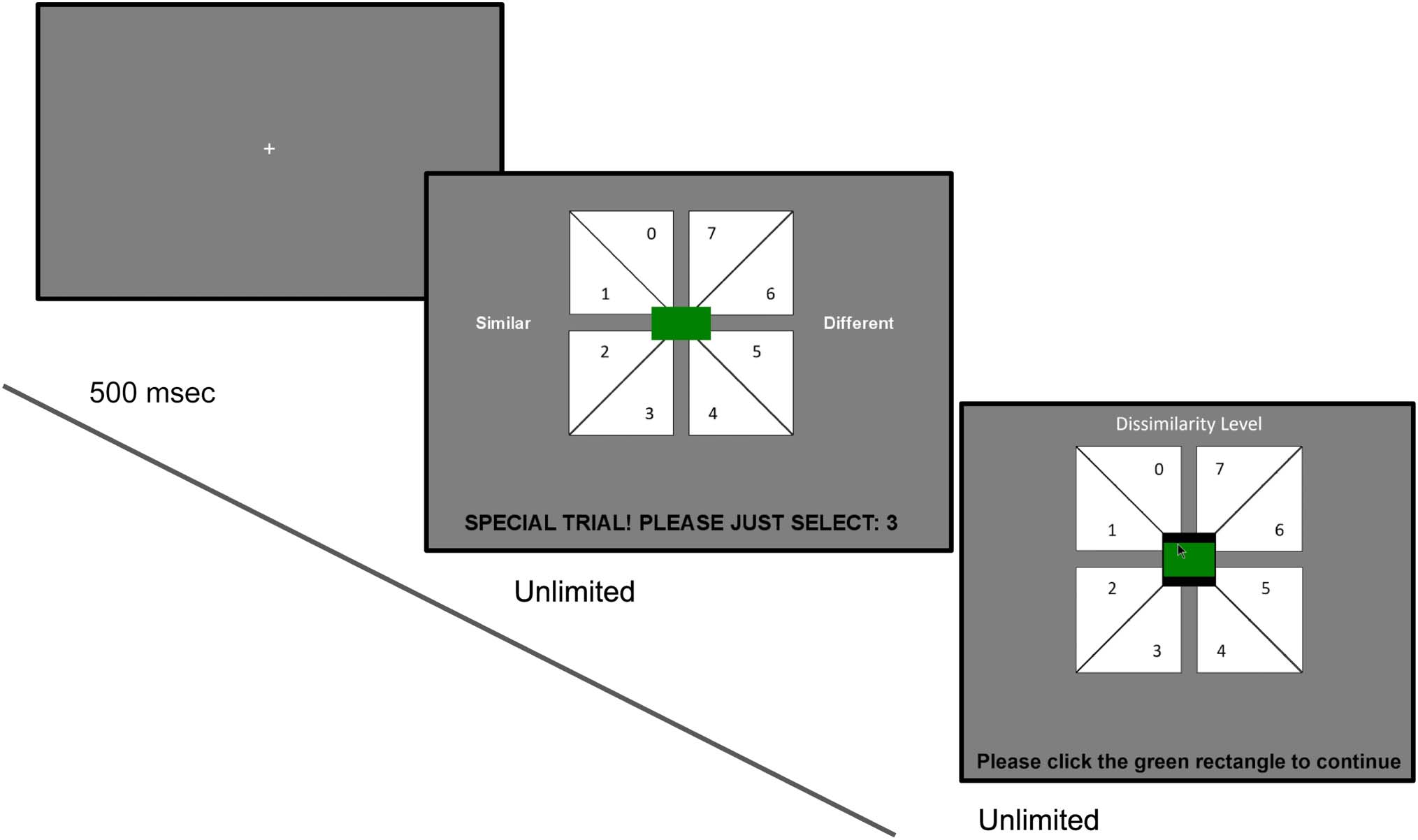
Cifra 3. Schematic of the
main trial procedure. Cada
trial begins with a 500-msec
fixation cross. This is followed
by stimuli presentation for
250 msec in either the CC
(abajo), PÁGINAS (medio), or CP
(arriba) configuración. Participantes
are then asked to rate the
dissimilarity of the two stimuli
on the response screen by
clicking on one of eight values.
After responding, Participantes
are asked to click on a green
rectangle in the center of the
screen to encourage fixation
back to the center.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
a rectangle in the center of the screen to direct their
fixation and the mouse pointer back to the center for
the next trial.
On the next screen, they are informed that some trials
will be catch trials, where no stimuli will be displayed and,
en cambio, participants are to select a specified value. Último, a
provide context for similarity rating judgments, participar-
pants are also shown a 3 × 3 grid of colored circles
corresponding to all the color stimuli they will see
throughout the experiment (Figure B1). Después,
they perform nine practice trials to familiarize them selves
with the experimental procedure. Para estas pruebas, ellos
were provided with feedback on what selection they
hecho, consisting of both the value they selected and the
text “Very Similar,” “Similar,” “Different,” or “Very Differ-
ent” for selections of 0–1, 2–3, 4–5, 6–7, respectivamente. En
the cessation of these practice trials, they were asked to
press the SPACE button to proceed to the main trial set.
En cada prueba, participants first fixate on a
Main task.
central fixation cross for 500 mseg. After this, hay
three possible trial types. In main trials, two stimuli will
be displayed, either in a center–center (CC), center–
periphery (CP), or periphery–periphery (PÁGINAS) configura-
ción (Cifra 3). The stimuli are presented for 250 mseg,
after which a response screen is displayed. By clicking
the appropriate location, participants are asked to
report the perceived similarity between the stimuli
in terms of their color. In catch trials, where no color
stimuli are displayed, participants are presented with a
response screen and are directed to select a specified
valor (Figure B2). To initiate the next trial, ellos son
directed to click a rectangle in the center of the screen.
Within each experimental condition, cada participante
completed 81 CC, 162 CP, y 81 PP main trials and 10
catch trials, equaling 334 trials in total per experimental
condición. Trials are randomly ordered within sequence,
and a different sequence will be presented to each partic-
ipant. Each participant completed each of the four
experimental conditions: large stimulus, periférico
aumento (MAG BIG); large stimulus, no peripheral
aumento (FIX BIG); small stimulus, peripheral magni-
ficción (MAG SMALL); and small stimulus, no peripheral
aumento (FIX SMALL). The sequence in which
participants perform the experimental conditions was
randomized across the different participants.
Sampling Plan
Participant Exclusion
Participant quality control occured both before and after
the main experiment. Before the experiment, Participantes
were excluded if they self-report visual acuity or color
vision impairments, including participants who normally
use glasses. After collecting the data and based on our
online pilot experiments, participants who score < 77%
on catch trials or who do not complete all main trials were
514
Journal of Cognitive Neuroscience
Volume 35, Number 4
excluded (Figure C1; Appendix D). All other participants
were included in the data analysis.
Bayes Factor Design Analysis
We use the Bayes Factor (BF) Design Analysis approach
to determine the number of participants required to
provide compelling evidence for or against our hypoth-
eses (Table A1, Summary Table). We use a sequential
design with maximum participant approach to recruit
participants until either: the BF provides strong evidence
for the null hypothesis (BF < 0.1) or the alternative
hypothesis (BF > 10); or a total of 50 Participantes (con-
out counting those that are excluded by the above cri-
teria) has been reached (Schönbrodt & carpinteros,
2018). Más específicamente, the BF for each and all of
Hypotheses 1A, 1B, 1C, 1D, and 2A must each exceed
one of the thresholds for recruitment to be halted before
alcanzando 50 Participantes. We also collect a minimum
of five participants who pass the exclusion criteria.
We compute BFs using the bayestestR package of the
R Environment forStatistical Computing (Makowski,
Ben-Shachar, & Lüdecke, 2019). BFs are calculated as
appropriate for each hypothesis (see Analysis Plan section
para más detalles).
Analysis Plan
Planned Analyses
Abajo, we use the results of the online pilot experiments
to explain our analyses strategies and detailed hypotheses
(Figures 4–7, Apéndice D).
Descriptive statistics. For the registered in-laboratory
experimento, we visually demonstrate the group average
dissimilarity matrix as in Figure 4. We also perform metric
multidimensional scaling (MDS) on the dissimilarity data
to assess whether the data recapitulate the previously
reported color hue rings. Último, for the planned experi-
mentos, we provide the within-subject response correlation
across the first and second pass per condition. We did not
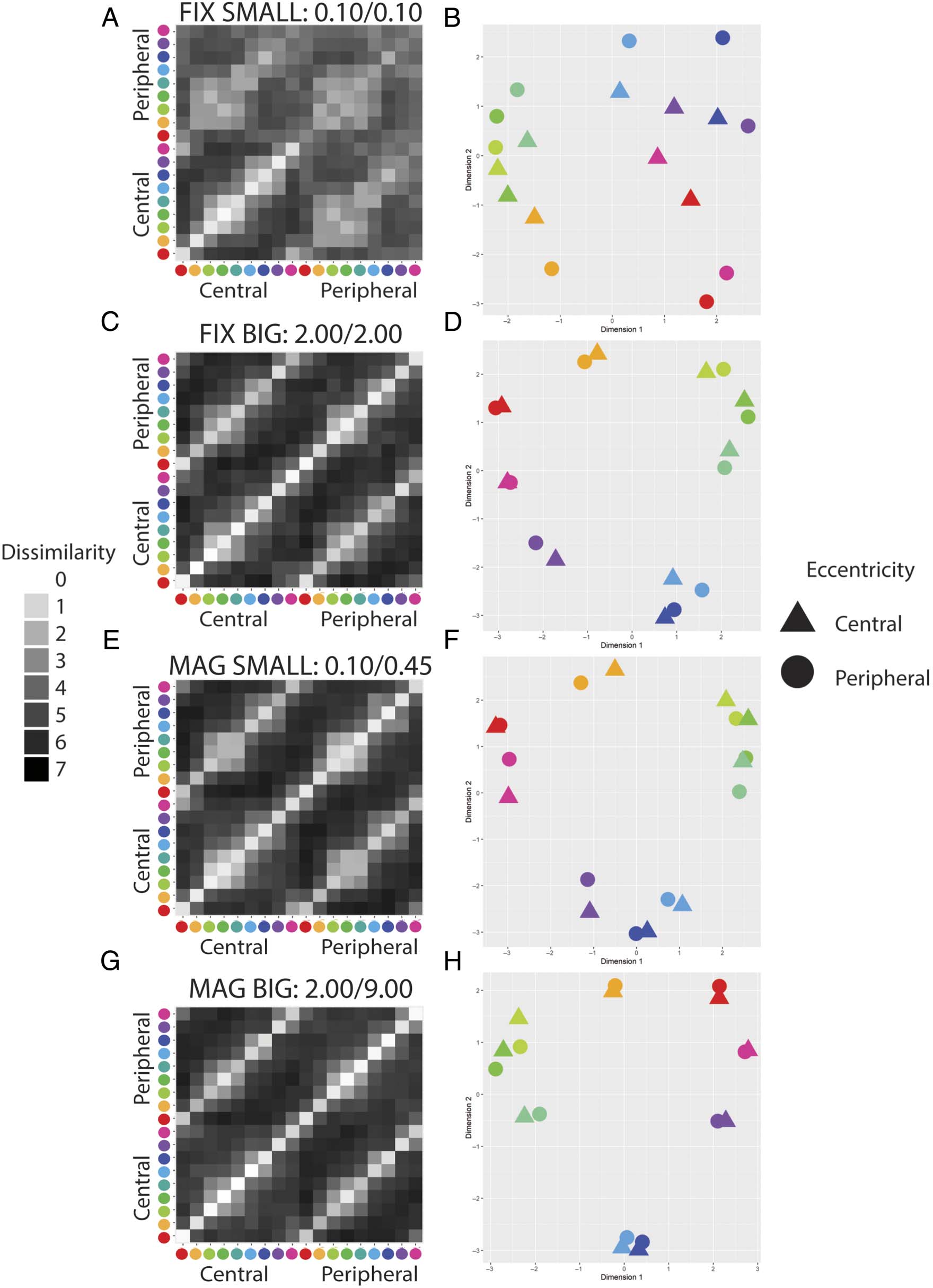
collect this for the pilot study. In our pilot data, all groups
except the participants shown small, unmagnified stimuli
(Figure 4A and B) provide clearly structured responses
across the full range of dissimilarities.
We have also collected n = 1 control participant data
for one of the stimulus and magnification groups using
the Panoworks display, confirming the feasibility of our
estudiar.
Are the overall structures of similarity reports equiva-
If color
lent for different eccentricities? (Hipótesis 1).
experiences are equivalent across the visual field, entonces
dissimilarity reports for color pairs experiences at one
eccentricity should be correlated with those at another
eccentricity (Figure 5A). Para cada participante, we obtain
the correlation between their judgments for each of the
eccentricity conditions, eso es, CC to CP, CC to PP, y
CP to PP (Figura 5B).
For the registered laboratory experiment, we hypothe-
size that participants presented with peripherally
magnified stimuli (en 60 DVA) will provide judg-
ments for each eccentricity condition that are posi-
tively correlated with each other (H1A: CC to CP,
H1B: CC to PP, H1C: CC to CP). We assess this by
first Fisher-Z transforming the correlations and then per-
forming a one-sample Bayesian t test with the default
Jeffreys prior for the variance and Cauchy prior for the
effect size for each hypothesis. We consider a BF ≥ 3,
indicating that the distribution of correlations is nonzero,
to support our hypotheses. The online pilot data (periph-
ery at 10 DVA) we have already collected support this
hipótesis (Figure 5C, Table C1).
Are physically equivalent stimuli subjectively equivalent
If color expe-
at different eccentricities? (Hipótesis 2).
riences are equivalent across the visual field, then an
experience of a color should be reportable as identical to
the same color presented elsewhere. Assuming this is the
caso, participants may report dissimilarity values for the
comparison of physically identical stimuli that are close
to zero and significantly smaller than for non-identical
stimuli pairs. We assess whether dissimilarity values for
identical pairs are significantly smaller than for non-
identical pairs. Para cada participante, we obtain the mean
dissimilarity response to physically identical stimuli,
termed a “normalized trace,” in each of the CC, CP, y
PP conditions (Figure 6A and B), as well as their mean
rating for non-identical stimuli (Figure 6C). The per-
participant difference between their mean response to
non-identical pairs and the normalized trace is used to
assess subjective equivalence, termed a “normalized
trace difference” (Figure 6D).
For the registered laboratory experiment, we hypothe-
size that participants presented with peripherally
magnified stimuli (en 60 DVA) will provide a normal-
ized trace difference significantly greater than zero
for each of the CC (H2A), CP (H2B), and PP (H2C)
condiciones. We assess this by fitting the normalized
trace difference of these participants using a Bayesian
beta regression with no regressors. A beta regression is
appropriate as the normalized traces are bounded by the
minimum and maximum possible dissimilarity responses
(Cribari-Neto & Zeileis, 2010). We perform a transforma-
tion to ensure boundary offset (Smithson & Verkuilen,
2006). The data are fitted using the R package Rstanarm
(Goodrich, Gabry, Alí, & Brilleman, 2020). We use the
default prior that the data mean (model intercept) will
be centered around zero, which corresponds to a prior
belief that the participants provide the same responses
to non-identical and identical color pairs. reportamos el
intercept estimate and its Bayesian 95% highest-posterior-
density (HPD) interval, as well as the BF10 for the intercept
Zeleznikow-Johnston et al.
515
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
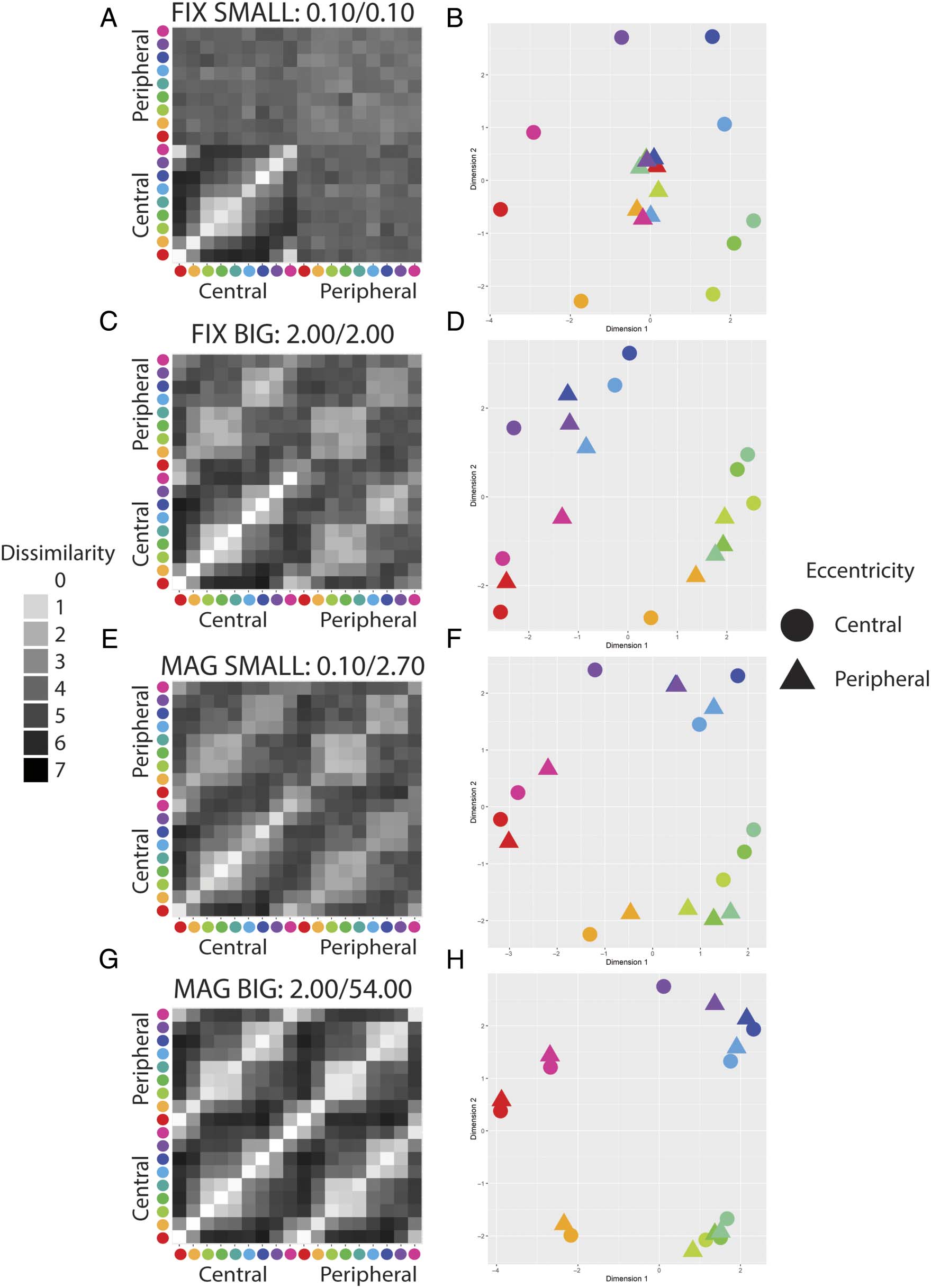
Cifra 4. Results of the pilot online experiment (n = 13–17). (A, C, mi, GRAMO) Group mean dissimilarity matrices for each experimental condition. El
data have been symmetrized across the main diagonal. (B, D, F, h) The corresponding MDS plot for each condition. Circles and triangles indicate
centrally and peripherally presented stimuli, respectivamente. Peripheral stimuli are centered at 10 DVA. FIX = peripheral stimuli of fixed-size with
respect to central stimuli; MAG = peripherally magnified (×4.5) estímulos; SMALL = central stimulus diameter of 0.1 DVA; BIG = central stimulus
diameter of 2.0 DVA.
516
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
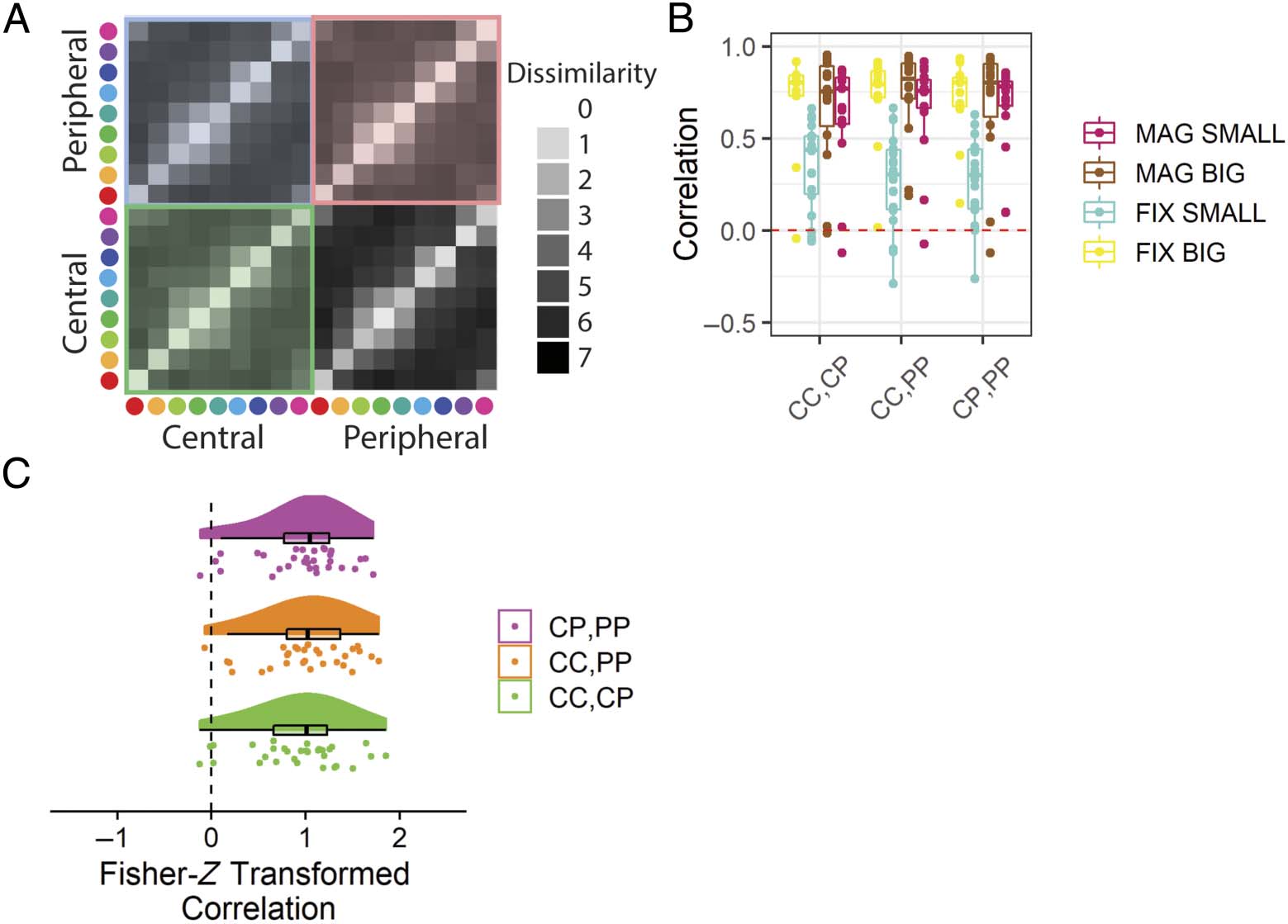
Cifra 5. Results from the pilot online experiment that address whether similarity reports for the same stimuli shown at different eccentricities
are correlated. (A) A schematic illustration with the set of dissimilarity values reported with the values being compared marked with colored shading.
(B) The within-subject correlations between judgments given in different eccentricities. The red dashed line indicates zero correlation. (C) El
Fisher-Z transformed within-subject correlations between judgments across eccentricity for participants shown peripherally magnified stimuli.
Peripheral stimuli are centered at 10 DVA. Boxplots centerline is median, box shows 25th–75th percentile, and whiskers show the largest value within
150% of the interquartile range. CC = central–central; CP = central–peripheral; PP = peripheral–peripheral; FIX = peripheral stimuli of fixed-size
with respect to central stimuli; MAG = peripherally magnified (×4.5) estímulos; SMALL = central stimulus diameter of 0.1 DVA; BIG = central stimulus
diameter of 2.0 DVA.
using the Savage-Dickey density ratio ( carpinteros,
Lodewyckx, Kuriyal, & Grasman, 2010). We consider a
BF ≥ 3, indicating that the model intercept is less than
cero, to support our hypothesis. The online pilot data
(periphery at 10 DVA) we have already collected support
this hypothesis (Figure 6E, Table C2).
We suspect that small stimuli shown in the periph-
ery may not induce color experiences that are
equivalent to when the same stimuli are presented
centrally. Además, we expect that this can be
corrected by magnifying the size of stimuli in the
periphery (H2D). We assess this by fitting all the
participant normalized trace data with a Bayesian beta
regression with stimulus size, peripheral magnification,
and Size × Magnification interaction as regressors. Nosotros
use the default prior, which corresponds to a prior belief
that the coefficient for all of the regressors is zero. Nosotros
consider a BF ≥ 3 for the interaction term to support
our hypothesis. The online data we have already collected
are inconclusive for this hypothesis (Figure 6F; Table C3).
Are color-pair comparisons equivalent across the visual
If color experience structures are
campo? (Hipótesis 3).
equivalent across the visual field, any pair of color experi-
ences should elicit the same dissimilarity response
regardless of eccentricity. We analyze whether this is
indeed the case by examining whether participants make
the same dissimilarity response for a color stimulus pair
in the CC, CP, and PP trial conditions. If participants pro-
vide the same responses for all three conditions, el vari-
ance in dissimilarity across those trials is zero (Figure 7A
y B). A diferencia de, if the response to a color pair in a par-
ticular trial type is uninformative with respect to the other
trial types (es decir., the correspondence in dissimilarity values
for a particular color pair under CC, CP, and PP conditions
is no greater than between color pair dissimilarity values
randomly sampled from a participant’s judgments), entonces
the variance in dissimilarity across trials in the three
conditions would be substantial (Figure 7B). We assess
whether participants provide consistent responses across
eccentricity conditions by comparing the mean variance of
their actual color pair judgments across CC, CP, and PP
conditions to the case where the corresponding CP and
PP judgments are randomly scrambled (Figure 7B;
Figure C3). If the difference in mean variance between
participants’ original and scrambled data is greater
than zero, then participants are providing consistent
responses across eccentricity conditions. De este modo, Lo haremos
obtain the difference in mean variance for original and
CP/PP-scrambled dissimilarity responses across the three
Zeleznikow-Johnston et al.
517
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
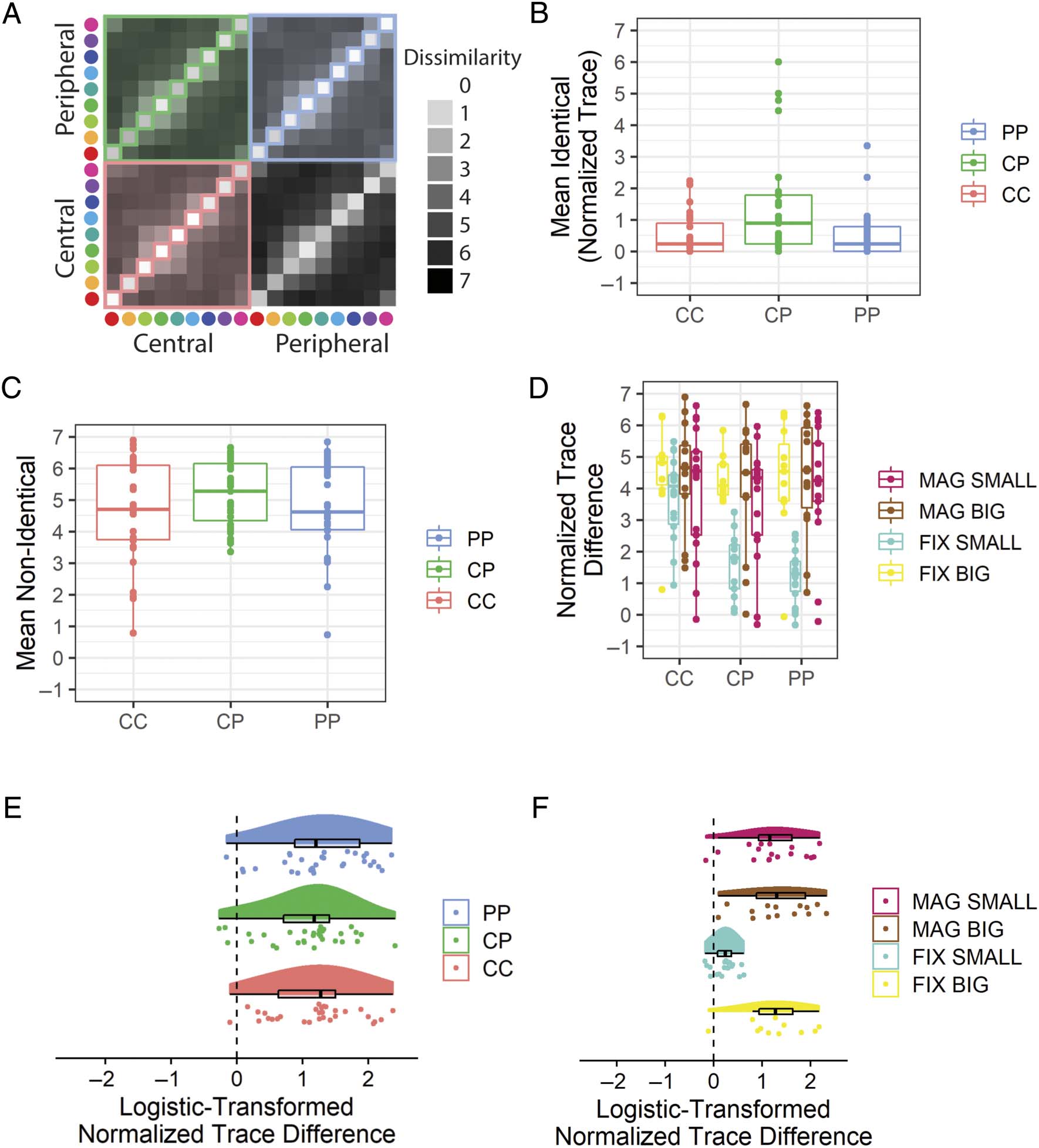
Cifra 6. Results from the pilot online experiment that address whether identical color stimuli are reported as the same at different viewing
ubicación. (A) A schematic illustration with the set of dissimilarity values reported for identical color stimuli in each condition marked by the diagonal
squares, whereas non-identical color stimuli are marked by colored shading. (B) The mean rating given to identical color pair stimuli in each
eccentricity condition, for participants shown peripherally magnified stimuli. (C) As per (B), but for non-identical color pairs. (D) la diferencia
between the mean dissimilarity value for non-identical and identical color patch pairs, called a “normalized trace difference.” The normalized trace
difference is plotted for each condition for each participant. (mi) The normalized trace difference values for each condition from participants shown
peripherally magnified stimuli. The values are scaled from 0 a 1 based on the maximum and minimum normalized trace differences possible and
then logistically transformed. (F) The transformed normalized trace differences for the PP condition for all participants. Peripheral stimuli are
centered at 10 DVA. Boxplots centerline is median, box shows 25th–75th percentile, and whiskers show the largest value within 150% del
interquartile range. CC = central–central; CP = central–peripheral; PP = peripheral–peripheral; FIX = peripheral stimuli of fixed-size with respect
to central stimuli; MAG = peripherally magnified (×4.5) estímulos; SMALL = central stimulus diameter of 0.1 DVA; BIG = central stimulus diameter
de 2.0 DVA.
trial conditions and all color pairings for each participant
(Figure 7C and D).
We hypothesize that participants presented with
peripherally magnified stimuli will provide a
difference in mean variance for color pairs shown
in different eccentricities significantly greater than
that expected from inconsistent color pair judg-
ment responses (H3). We assess this by fitting the
518
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
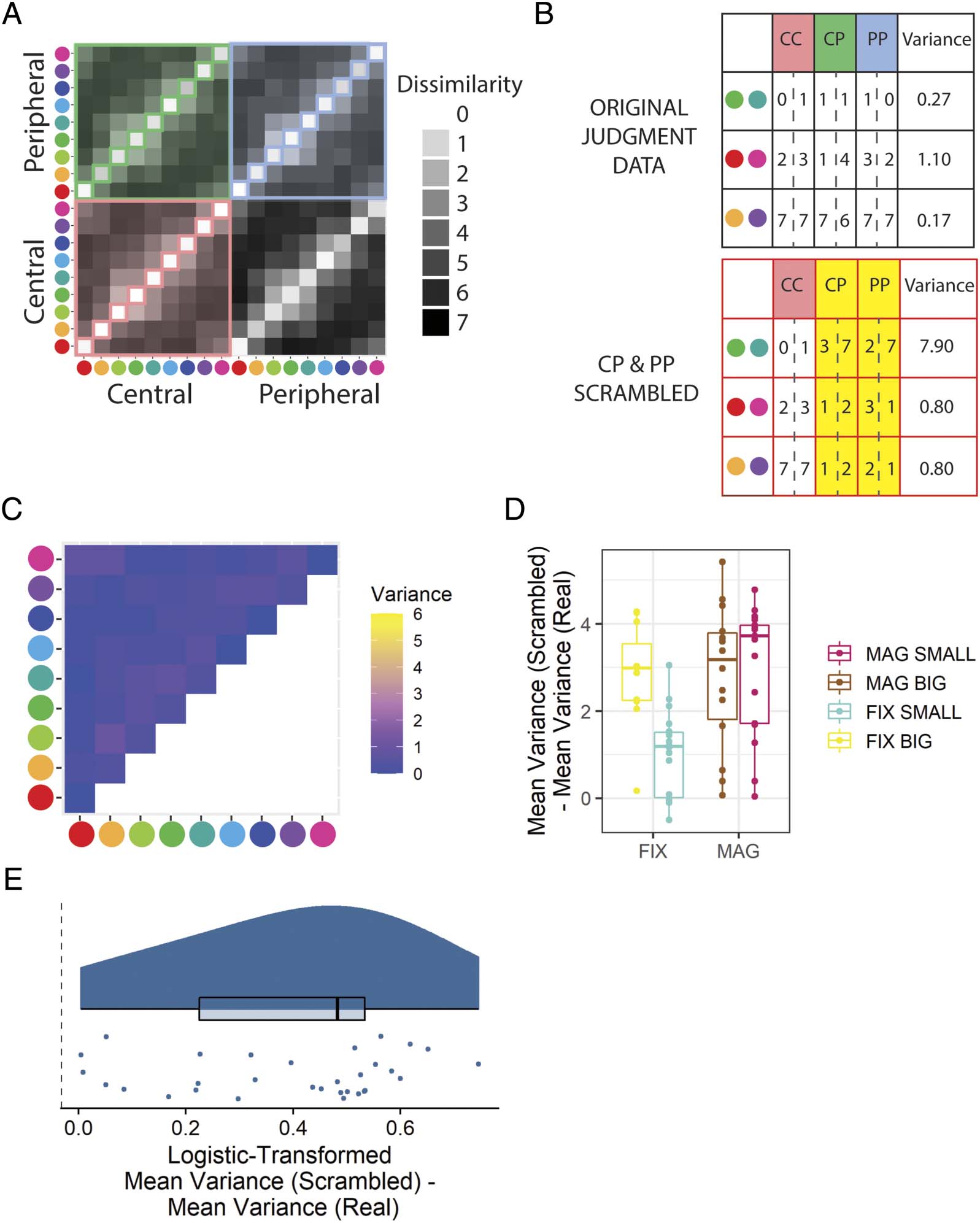
Cifra 7. Results from the online experiment that address whether arbitrary pairs of color stimuli are reported to have the same dissimilarity at different
viewing locations (n = 13–17). (A) The set of dissimilarity values collected from participants for each possible color pair in each condition is marked
by the colored shading. Participants provided two judgments for each non-identical color pair in each condition. (B) A demonstration of how variance
in dissimilarity values is calculated for a given color pair shown in different conditions. Two judgments are made for each color pair in each condition,
meaning the variance is calculated across six values. en el análisis, this variance is compared with the mean variance that would be seen if random color
pairs were sampled as opposed to the same color pair under different eccentricity conditions. See also Figure C3. (C) The mean variance across participants
shown peripherally magnified stimuli for each color pair. (D) The difference in mean color-pair variance for scrambled and real ratings provided by
cada participante. (mi) The difference in mean color-pair variance between scrambled and real data for each participant shown peripherally magnified
estímulos. The values are scaled from 0 a 1 based on the maximum and minimum mean variance difference possible and then logistically transformed.
Peripheral stimuli are centered at 10 DVA. Boxplots centerline is median, box shows 25th–75th percentile, and whiskers show the largest value within 150%
of the interquartile range. CC = central–central; CP = central–peripheral; PP = peripheral–peripheral; FIX = peripheral stimuli of fixed-size with respect to
central stimuli; MAG = peripherally magnified (×4.5) estímulos; SMALL = central stimulus diameter of 0.1 DVA; BIG = central stimulus diameter of 2.0 DVA.
mean variance difference from these participants using a
Bayesian beta regression with no regressors and the
default prior that the data mean (model intercept) will
correspond to 0. We consider a BF ≥ 3, Indicando que
la media (model intercept) is greater than that expected
from inconsistent responding, to support our hypothe-
hermana. The online data we have already collected support
this hypothesis (Figure 7E; Table C4).
Zeleznikow-Johnston et al.
519
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 8. Visualization of the participant dissimilarity judgments for the in-laboratory experiment (norte = 14). (A, C, mi, GRAMO) Group mean dissimilarity
matrices for each experimental condition. The data have been symmetrized across the main diagonal. (B, D, F, h) The corresponding MDS plot
for each condition. Circles and triangles indicate centrally and peripherally presented stimuli, respectivamente. Peripheral stimuli are centered at 60 DVA.
FIX = peripheral stimuli of fixed-size with respect to central stimuli; MAG = peripherally magnified (×27) estímulos;. SMALL = central stimulus
diameter of 0.1 DVA; BIG = central stimulus diameter of 2.0 DVA.
520
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
Mesa 1. A Summary of the Stimulus Eccentricity and Size Parameters across the Online and In-laboratory Experiments
Central Stimulus
Eccentricity (DVA)
Central Stimulus
Diameter (DVA)
Peripheral Stimulus
Eccentricity (DVA)
Peripheral Stimulus
Diameter (DVA)
1
1
1
1
1
1
1
1
0.10
2.00
0.10
2.00
0.10
2.00
0.10
2.00
10
10
10
10
60
60
60
60
0.10
2.00
0.45
9.00
0.10
2.00
2.70
54.00
Nombre
En línea
FIX SMALL
FIX BIG
MAG SMALL
MAG BIG
In-laboratory
FIX SMALL
FIX BIG
MAG SMALL
MAG BIG
RESULTADOS
Participantes
For the in-laboratory experiment, we recruited 14 saludable
volunteers (8 mujer, edad media = 36.4, DE = 13.8) con
normal or corrected-to-normal vision (color vision not
formally assessed) before reaching our stopping criteria.
No participants were rejected because of meeting exclu-
sion criteria.
Estadísticas descriptivas
Informal visual inspection of both the group mean par-
ticipant dissimilarity matrix and a 2-D metric MDS repre-
sentation of that data for each experimental condition
suggests that whether color experiences are equivalent
across the visual field depends on the size of the stimuli
that elicit them (Cifra 8, Mesa 1). Photographs of the
monitor display for these conditions can be seen in
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
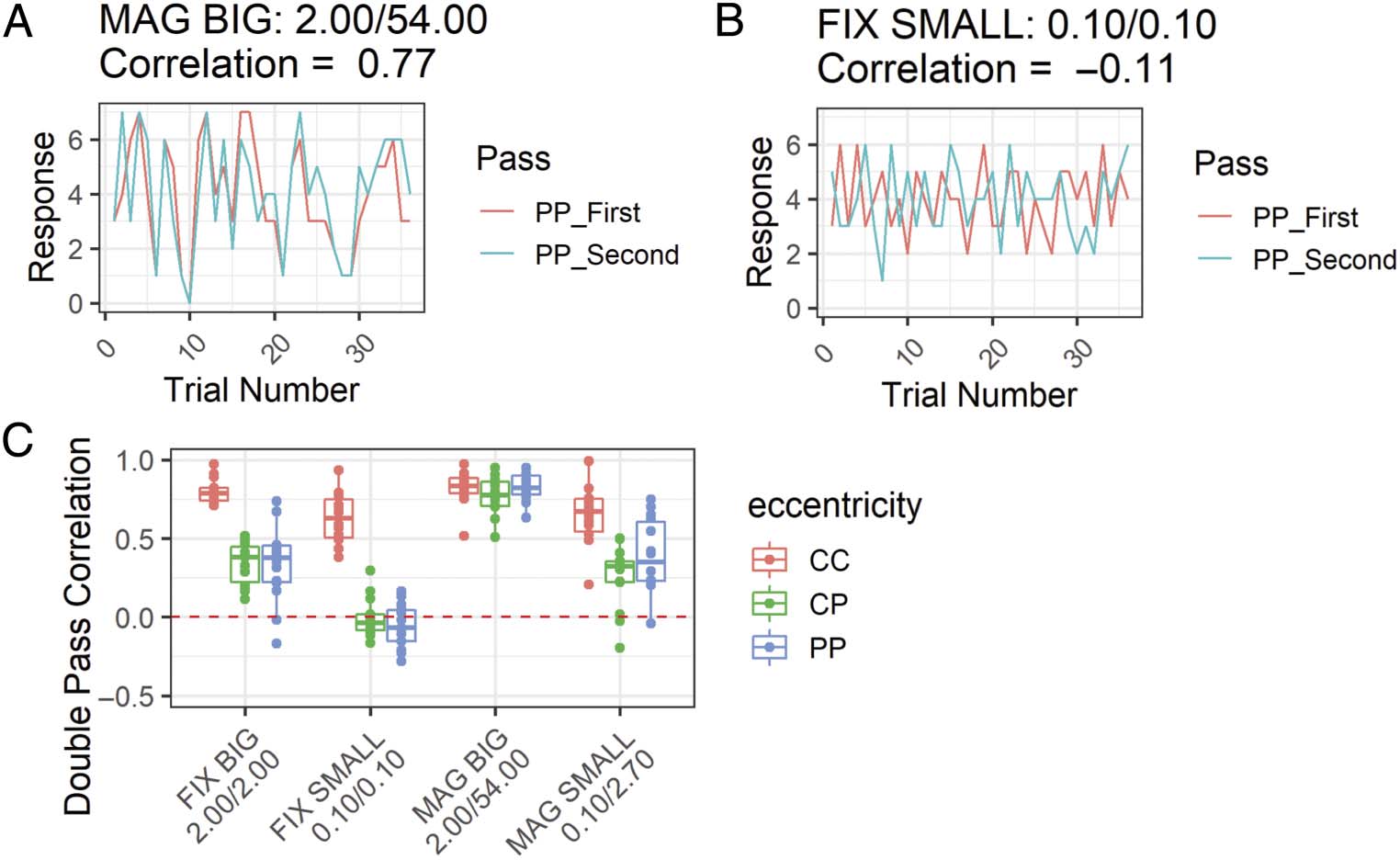
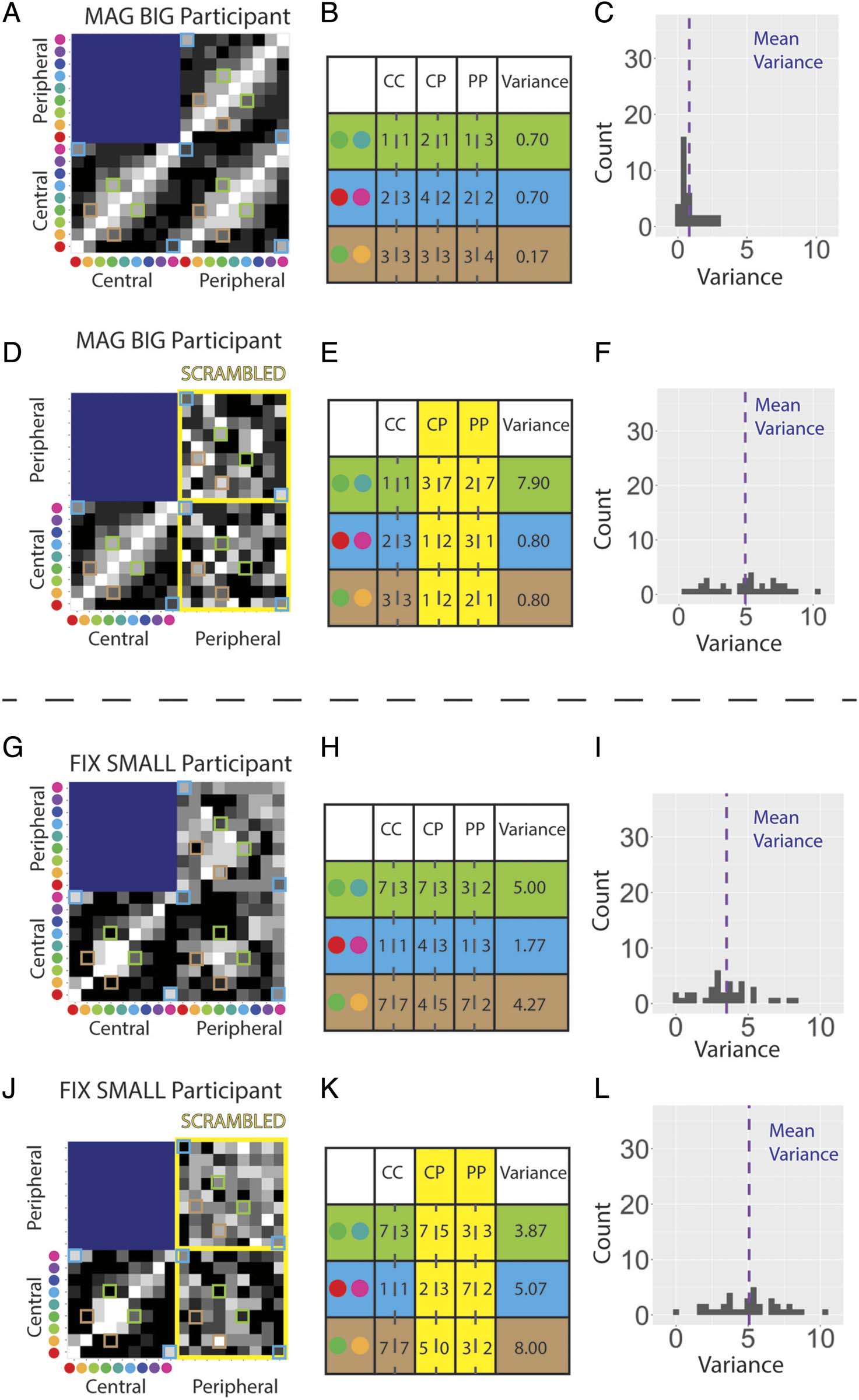
Cifra 9. Results from the double-pass analysis for judgment consistency. (A) The first and second responses for each color pair in the PP presentations
for Participant 1 in the MAG BIG condition. The high correlation between the two passess indicates consistent responding. (B) As per (A) si no fuera por
Participant 1 during the FIX SMALL condition. Low correlation indicates inconsistent responding, perhaps because of failure to perceive the stimuli.
(C) The double pass correlation across all participants for each condition and presentation type. FIX = peripheral stimuli of fixed size with respect to
central stimuli. Peripheral stimuli are centered at 60 DVA. MAG = peripherally magnified (×27) estímulos; SMALL = central stimulus diameter of 0.1 DVA;
BIG = central stimulus diameter of 2.0 DVA; CC = central–central; CP = central–peripheral; PP = peripheral–peripheral comparison.
Zeleznikow-Johnston et al.
521
Figure A2. Raw responses for the individual participants
can be seen in Figure A3.
Each participant performed each color pair judgment
twice for each eccentricity condition within each experi-
mental condition, with the “first pass” and “second pass”
together comprising a “double pass paradigm.” Deter-
mining the correlation between a participant’s responses
for their first and second pass allows us to judge how
consistent they are in their responses given identical
stimulus conditions. When participants can see the stim-
uli in a consistent manner, the correlation should be
alto (Figure 9A). Note that high correlations are possible
even with differences in the quality of color experiences
at the fovea and periphery (p.ej., if peripheral stimuli
look desaturated in a consistent manner, it should still
result in high correlations; Decock & Douven, 2013).
When participants cannot see the stimuli per se, or at
least not their colored aspect, their similarity responses
may become more random, resulting in lower correla-
tions across passes (Figure 9B). Resultantly, examination
of the double pass correlations across all experimental
conditions for all participants gives some suggestion of
their ability to perceive the stimuli (Figure 9C). En el
MAG BIG condition, participants give highly correlated
judgments regardless of eccentricity, suggesting consis-
tent perception of the stimuli. A diferencia de, partícipe
judgments are uncorrelated for the CP and PP trials dur-
ing the FIX SMALL condition, indicating a potential failure
to perceive these small, unmagnified stimuli (cualquiera
entirely or at least their color aspect). This matches the
informal reports some participants spontaneously pro-
vided to the experimenter during the FIX SMALL condi-
tion claiming they could not see the color or location of
the stimulus. Whereas CC judgments remain highly cor-
related in both the FIX BIG and MAG SMALL conditions,
participant responses for CP and PP trials have a relatively
reduced but still positive correlation.
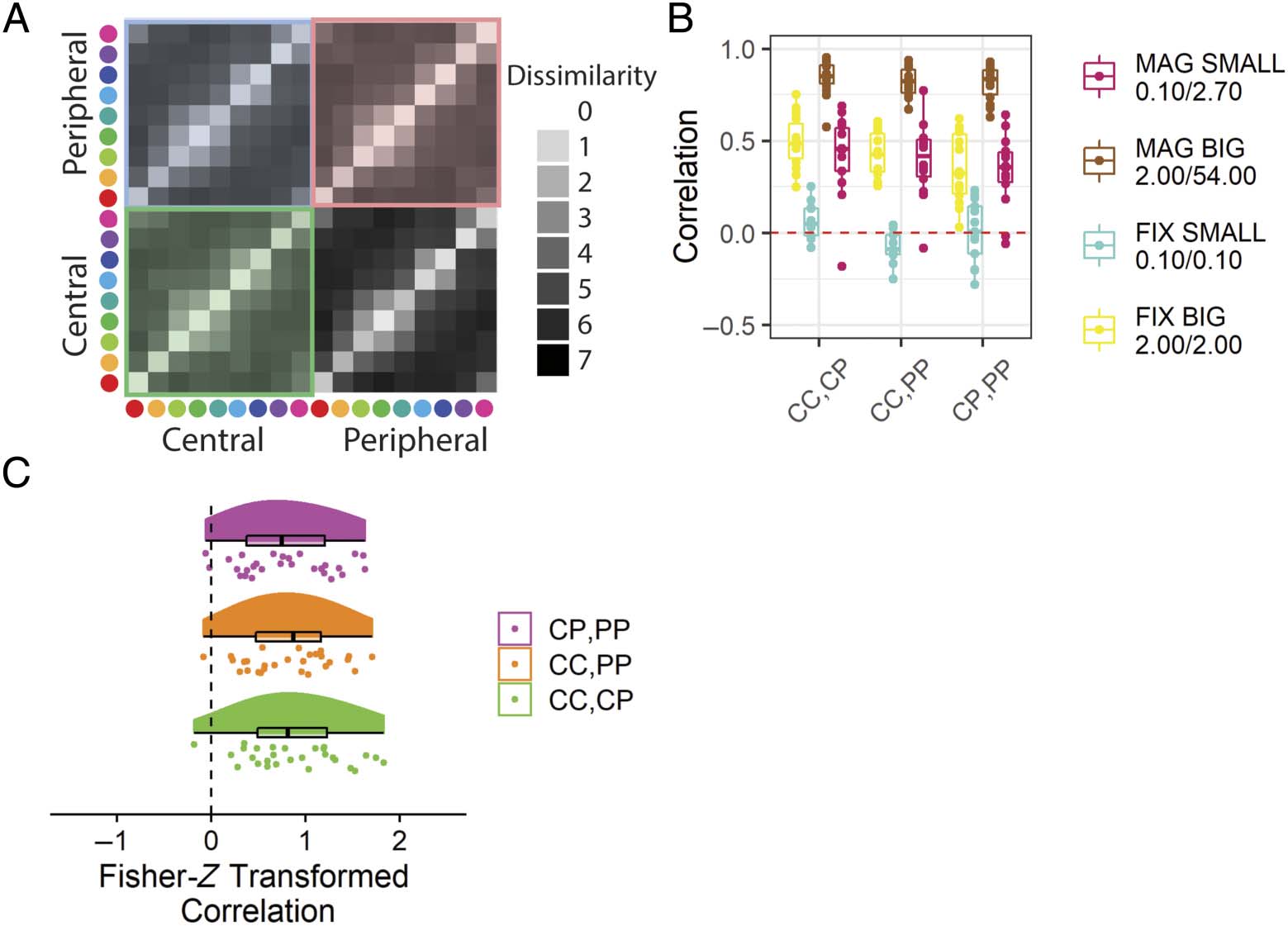
Similarity Structure Correlations (Hipótesis 1)
We hypothesized that participants presented with periph-
erally magnified stimuli would provide dissimilarity judg-
ments at each eccentricity condition that are positively
correlated with each other (H1A: CC to CP, H1B: CC to
PÁGINAS, H1C: CC to CP). All of these hypotheses were sup-
ported (Figure 10C, Mesa 2).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 10. Results from the in-laboratory experiment that address whether similarity reports for the same stimuli shown at different eccentricities
are correlated. (A) A schematic illustration with the set of dissimilarity values reported with the values being compared marked with colored shading.
(B) The within-subject correlations between judgments given in different eccentricities. The red dashed line indicates zero correlation. (C) El
Fisher-Z transformed within-subject correlations between judgments across eccentricity for participants shown peripherally magnified stimuli.
Peripheral stimuli are centered at 60 DVA. Boxplots centerline is median, box shows 25th–75th percentile, and whiskers show the largest value within
150% of the interquartile range. CC = central–central; CP = central–peripheral; PP = peripheral–peripheral; FIX = peripheral stimuli of fixed-size
with respect to central stimuli; MAG = peripherally magnified (×27) estímulos; SMALL = central stimulus diameter of 0.1 DVA; BIG = central stimulus
diameter of 2.0 DVA.
522
Revista de neurociencia cognitiva
Volumen 35, Número 4
Mesa 2. A Summary of the Experimental Results
Similarity Structure Correlations
Hipótesis
Comparison
Mean Correlation
H1A
H1B
H1C
CC, CP
CC, PÁGINAS
CP, PÁGINAS
0.71
0.68
0.65
Hipótesis
Condition
H2A
H2B
H2C
CC
CP
PÁGINAS
Parameter
Interceptar
Size
Magnification
H2D
Size × Magnification
Hipótesis
H3
Experiential Equivalence of Identical Stimuli
Significar
3.19
2.21
2.49
Estimate
−0,10
0.5
0.0
0.0
Significar
2.13
95% HPD
2.48 a 3.80
1.45 a 2.91
1.76 a 3.19
95% HPD
−0.36 to 0.13
0.12 a 0.83
0.01 a 0.04
−0.02 to 0.02
Overall Judgment Consistency
95% HPD
0.97 a 3.23
BF10
> 109
> 107
> 107
BF10
> 105
> 102
> 104
BF10
0.05
BF10
11.17
The corresponding hypotheses are listed in the text and summarized in Table A1.
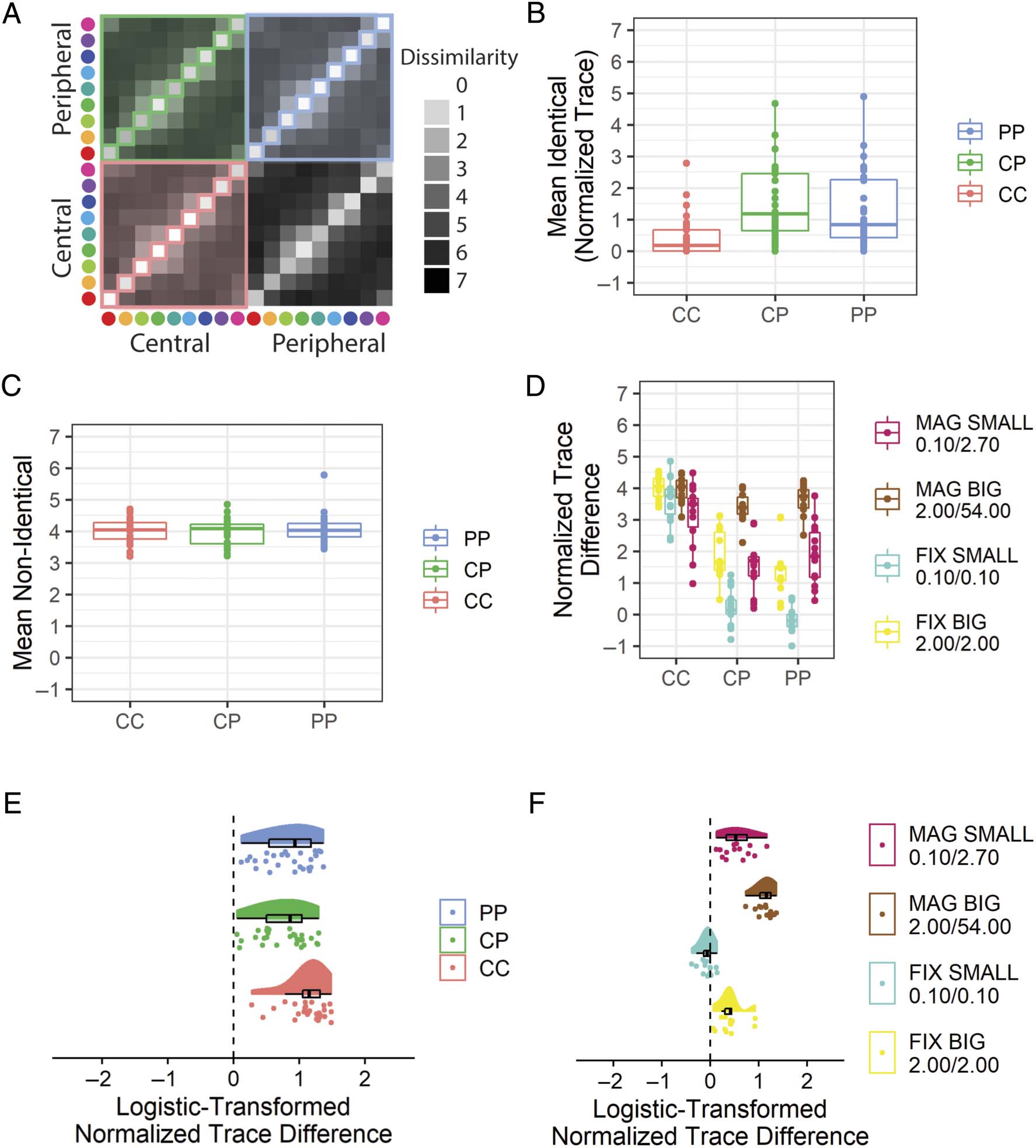
Experiential Equivalence of Identical Stimuli
(Hipótesis 2)
We hypothesized that participants presented with periph-
erally magnified stimuli would provide a normalized
trace difference significantly greater than zero for each
of the CC (H2A), CP (H2B), and PP (H2C) condiciones.
All of these hypotheses were supported (Figure 11E,
Mesa 2).
Además, we hypothesized that there would be an
interaction between peripheral magnification and stimu-
lus size on participant dissimilarity judgments. Este
hypothesis was not supported (Figure 11F; Mesa 2).
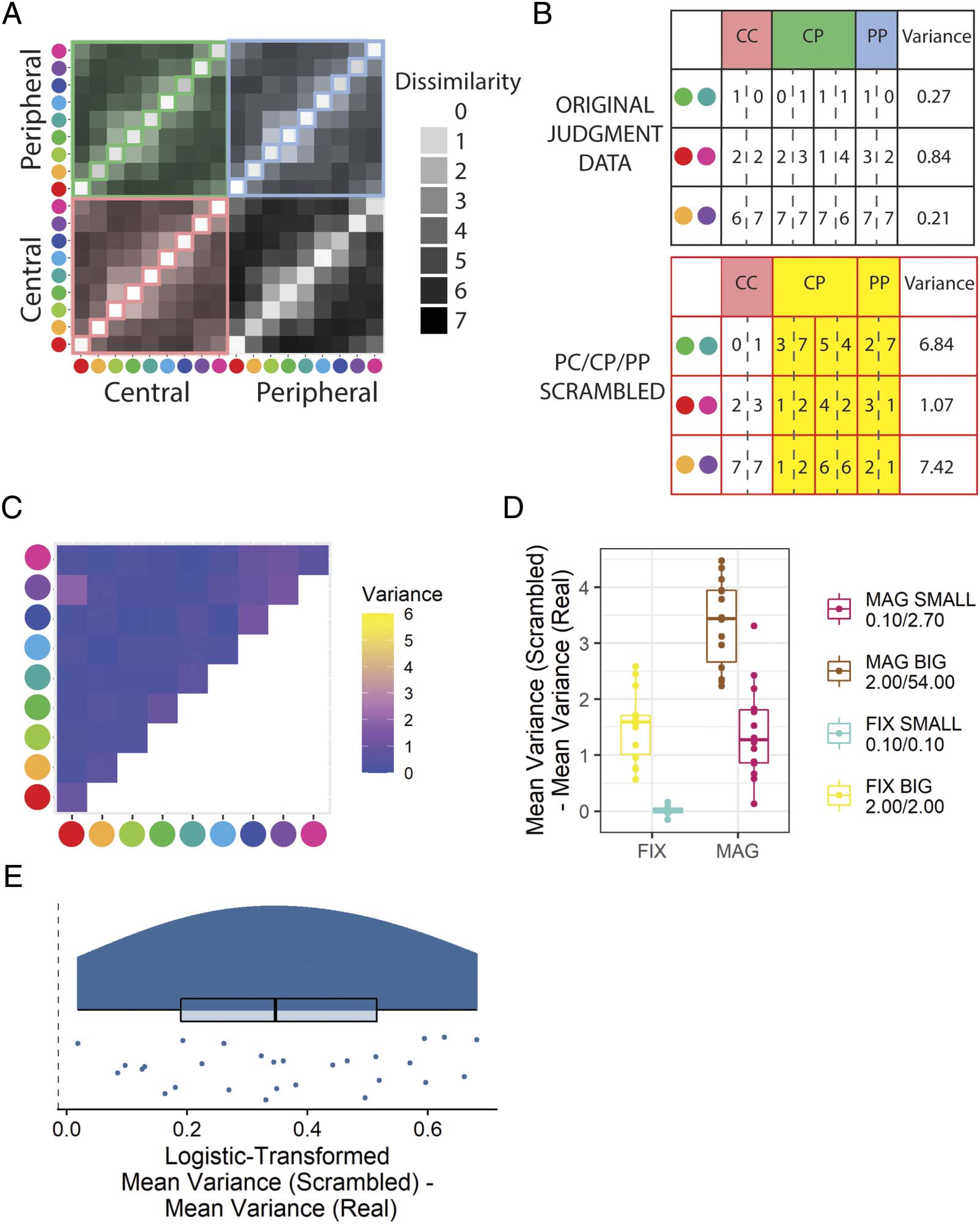
Overall Judgment Consistency (Hipótesis 3)
We hypothesized that participants presented with periph-
erally magnified stimuli would provide a difference in
mean variance for color pairs shown in different eccentric-
ities significantly greater than that expected from inconsis-
tent color pair judgment responses (H3). This hypothesis
was supported (Figure 12E; Mesa 1).
DISCUSIÓN
We set out to examine whether color experience relation-
ships are the same across the visual field, in contrast to pre-
vious psychophysics studies examining whether the same
stimulus always evokes the same experience irrespective
of retinal location. Our first hypothesis was that color judg-
ments for color-pair experiences at different locations
would be correlated, indicating structural equivalence.
We found that this was indeed the case (H1A–C). Our sec-
ond hypothesis was that (peripherally magnified) identical
stimuli would evoke maximally similar (zero dissimilarity)
experiencias, supporting structural equivalence that goes
beyond mere correlation. This too was supported by the
datos (H2A–C). Our third hypothesis was that non-identical
color stimuli pairs would elicit the same similarity
responses irrespective of presentation location, establish-
ing their experiential equivalence at different locations.
This was also affirmed (H3). Además, participants could
provide these responses in a consistent manner, como
demonstrated by the double-pass analysis. Juntos, el
collected data unambiguously demonstrate the structural
Zeleznikow-Johnston et al.
523
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 11. Results from the in-laboratory experiment that address whether identical color stimuli are reported as the same at different viewing
ubicación. (A) A schematic illustration with the set of dissimilarity values reported for identical color stimuli in each condition marked by the diagonal
squares, whereas non-identical color stimuli are marked by colored shading. (B) The mean rating given to identical color pair stimuli in each
eccentricity condition, for participants shown peripherally magnified stimuli. (C) As per (B), but for non-identical color pairs. (D) la diferencia
between the mean dissimilarity value for non-identical and identical color patch pairs, called a “normalized trace difference.” The normalized trace
difference is plotted for each condition for each participant. (mi) The normalized trace difference values for each condition from participants shown
peripherally magnified stimuli. The values are scaled from 0 a 1 based on the maximum and minimum normalized trace differences possible
and then logistically transformed. (F) The transformed normalized trace differences for the PP condition for all participants. Peripheral stimuli are
centered at 60 DVA. Boxplots centerline is median, box shows 25th–75th percentile, and whiskers show the largest value within 150% del
interquartile range. CC = central–central; CP = central–peripheral; PP = peripheral–peripheral; FIX = peripheral stimuli of fixed-size with respect
to central stimuli; MAG = peripherally magnified (×27) estímulos; SMALL = central stimulus diameter of 0.1 DVA; BIG = central stimulus diameter
de 2.0 DVA.
equivalence of color experiences across the visual field
when elicited by peripherally magnified stimuli.
Our data-supported hypotheses in turn support the
naive view that neurotypical humans perceive and
experience color across their entire visual field. Peripheral
color perception, eso es, the ability for large peripheral
stimuli to be consistently detected, appraised and com-
pared, is demonstrated explicitly by the formal hypotheses
524
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 12. Results from the in-laboratory experiment that address whether arbitrary pairs of color stimuli are reported to have the same dissimilarity
at different viewing locations. (A) The set of dissimilarity values collected from participants for each possible color pair in each condition is marked
by the colored shading. Participants provided two judgments for each non-identical color pair in each condition. (B) A demonstration of how
variance in dissimilarity values is calculated for a given color pair shown in different conditions. Two judgments are made for each color pair in
the CC and PP conditions and four for each color pair in the CP condition, meaning the variance is calculated across eight values. en el análisis,
this variance is compared with the mean variance that would be seen if random color pairs were sampled as opposed to the same color pair
under different eccentricity conditions. See also Figure C3. (C) The mean variance across participants shown peripherally magnified stimuli for
each color pair. (D) The difference in mean color-pair variance for scrambled and real ratings provided by each participant. (mi) la diferencia
in mean color-pair variance between scrambled and real data for each participant shown peripherally magnified stimuli. The values are scaled
de 0 a 1 based on the maximum and minimum mean variance difference possible and then logistically transformed. Boxplots centerline is median,
box shows 25th–75th percentile, and whiskers show the largest value within 150% of the interquartile range. Peripheral stimuli are centered
en 60 DVA. CC = central–central; CP = central–peripheral; PP = peripheral–peripheral; FIX = peripheral stimuli of fixed-size with respect to
central stimuli; MAG = peripherally magnified (×27) estímulos; SMALL = central stimulus diameter of 0.1 DVA; BIG = central stimulus diameter
de 2.0 DVA.
Zeleznikow-Johnston et al.
525
along with the double-pass correlations. Además,
peripheral color experiences, eso es, “what-it-feels-like to
experience colors in the periphery,” were demonstrated
to be structurally equivalent to those in central vision.
Generalizing these findings outside the laboratory would
sugerir, as naively expected, that we really can perceive
and experience a canopy of leaves as green, the clouds
as gray, and the sky as blue in our peripheral vision.
Sin embargo, we did not hypothesize, and our data do not
sugerir, that we perceive small stimuli equivalently when
presented at different visual field locations. Our sole inva-
lidated hypothesis was that there would be an interaction
effect between stimulus size and peripheral magnification
on dissimilarity responses for identical versus non-
identical stimuli shown in the periphery. We formulated
this hypothesis as we believed that only small, unmagni-
fied, peripherally presented stimuli would fail to produce
differing judgments for identical versus non-identical stim-
uli. We expected that either increasing stimulus size (de
0.10- to 2.00-DVA diameter) or peripherally magnifying
the stimulus (de 0.10- to 2.70-DVA diameter) would fully
attenuate this effect and produce equivalent judgments to
when the stimuli were presented centrally. Instead of an
interaction though, these judgments instead appeared to
be affected additively by stimulus size and peripheral
aumento. This suggests that for small stimuli to elicit
fully equivalent color experiences at 60 DVA to those seen
in the fovea, they need to be peripherally magnified by a
greater factor than the V2 receptive field scaling that we
chose. V4 scaling may potentially be a more appropriate
elección (Hombre libre & Simoncelli, 2011).
There are at least two distinct phenomenological
situations that may be occurring in the small, unmagnified
stimuli paradigm. One possibility is that these stimuli were
perceived and elicited color experiences, but with a
genuine change in the experienced color and their corre-
sponding similarities. Por ejemplo, previous studies have
suggested that peripheral stimuli appear less saturated
(Sakurai et al., 2003; gordon & Abramov, 1977), a pesar de
see Rajananda, Peters, Lau, and Odegaard (2017). Cómo-
alguna vez, our double-pass analysis (Cifra 9) revealed that
repeated presentation of small 0.10-DVA stimuli pairs
en 60 DVA produced entirely uncorrelated responses,
whereas ∼2.00-DVA stimuli responses were positively cor-
related yet reduced compared with larger stimuli. Más-
más, participants’ responses to small stimuli pairs were
inconsistent compared with larger stimuli, en lugar de
equal variance but different value. This pattern of results
seems hard to reconcile with an explanation proposing
participants perceived peripheral stimuli as uniformly
and reliably less saturated in this experiment. The more
likely scenario is that these small stimuli were simply
harder to consciously detect than their larger counterparts,
resulting in a failure to elicit reliable similarity responses.
Given our current findings of color perception in
peripheral vision and related earlier work in the literature,
why then do some claim that “the feeling that our entire
‘subjective visual world’ is richly colored…must be an
illusion” (Chater, 2018)? We believe it stems from a
particular interpretation of the known findings on how
perceptual abilities differ across the visual field. Acuity
for detecting stimuli of a fixed size drops with eccentricity
(Strasburger et al., 2011), and this effect is more pro-
nounced for chromatic than achromatic stimuli (anderson,
Mullen, & Hesse, 1991; Mullen, 1991). Contrast-sensitivity
for high spatial frequencies declines exponentially with
eccentricity ( Cual, chi, & Makous, 1995), whereas in
general, increased contrast is required for chromatic stim-
ulus detection (Hansen et al., 2009; McKeefry et al., 2007;
Ayama et al., 2004; Sakurai et al., 2003). Último, although it
is unclear if it is affected by the chromaticity of the stimuli,
susceptibility to crowding is possibly the clearest behav-
ioral deficit of peripheral vision (Rosenholtz, 2016). Nosotros
note though that changes in behavioral performance do
not always unidirectionally decrease with eccentricity; para
ejemplo, the central fovea is tritanopic because of a lack
of S-cones (williams, MacLeod, & Hayhoe, 1981; Ver también
Apéndice D, Control Experiment 1), and textural segrega-
tion performance peaks outside of the fovea (Yeshurun &
Carrasco, 1998, 2000).
Presumiblemente, it is these psychophysical facts that
motivate the claims that peripheral color perceptual
experience is illusory. Before critiquing these, it is worth
noting that there are at least two possible interpretations
of these statements, hinging on what is meant by the term
“illusory”: (1) Perceptual performance for detection, dis-
crimination, and characterization of colored stimuli
in the periphery is dissociated from expected perfor-
mance, eso es, we are metacognitively overconfident
about our peripheral color perception performance; o
(2) our knowledge of our subjective experience of periph-
eral color at each moment is fragile and easily revised
or distorted by expectation and memory. This happens
to such an extent that the subjective reports of naive par-
ticipants may completely dissociate from their actual
color experience. We are unsure of whether the skeptics
mean to defend either or both of these claims, but as we
believe both are inaccurate, we shall address each in turn.
On the first interpretation: Variation in perceptual
abilities across the visual field does not imply an outright
lack of perception per se. The lack of high-acuity chro-
matic perception in the periphery does not suggest a
complete absence of chromatic perception, no more than
an inability to detect microscopic stimuli presented
foveally implies an absence of foveal vision (Anstis,
1998). Similarmente, formal modeling of variation in chromatic
perception across the visual field suggests that natural
stimuli should still appear colorful at peripheral eccentric-
ities (Haun, 2021). Many previous studies have shown that
chromatically presented stimuli can be detected when
presented peripherally, and even identified with foveal
stimuli if appropriately magnified (Hansen et al., 2009;
Abramov et al., 1991). Our current study supports this pre-
vious work and adds to it the finding that the perception of
526
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
the relational properties of these stimuli is equivalent
when they are peripherally magnified.
Regarding the second: When considering the subjective
experience of color as opposed to just stimulus detection,
it is not clear that any experiment based simply on behav-
ioral performance can be used to argue for an absence of
peripheral color experience. Por ejemplo, color constancy
and metamers demonstrate the potential dissociation
between stimulus properties and subjective experience
(Gegenfurtner, Bloj, & Toscani, 2015; Adelson, 2000;
Nimeroff & Yurow, 1965). Además, people sometimes
report color experiences in the complete absence of
estímulos, such as in afterimages (van Boxtel, Tsuchiya, &
Koch, 2010), the McCollough effect (Humphrey &
Goodale, 1998), or even while dreaming (Kahn, Dement,
Pescador, & Barmack, 1962). De este modo, claims of an absence of
color experience in peripheral vision based on failures of
stimuli detection could just as easily be used to deny the
presence of color experiences, despite subjective reports
to the contrary, in all these other circumstances.
Given all this, what then explains the seeming discrep-
ancy between our findings and the failure of participants to
notice the complete absence of peripheral color stimuli in
the virtual reality experiments of Cohen and colleagues
(2020)? We believe the answer is a combination of how
our differing paradigms exploited attention and expecta-
ción. Our experimental setup involved participants
observing briefly presented colored stimuli in random
locations against an otherwise featureless gray back-
ground, with participants discouraged from making sac-
Cades. This paradigm presumably enhances the salience
of the peripheral stimuli through their sudden onset and
lack of predictable location, engaging exogenous atten-
ción. A diferencia de, Cohen’s virtual reality display involved
participants observing colorful natural scene imagery, con
participants free to saccade, for multiple seconds before
gradual peripheral desaturation. Notablemente, Participantes
often failed to notice peripheral desaturation even when
they were aware of its upcoming possibility, in a manner
reminiscent of gradual change blindness (simons,
Franconeri, & Reimer, 2000). We suspect this is likely
because of memory of the scene colors along with a strong
prior perceptual expectation that naturalistic scenes have
constant physical color. One possible result of these fac-
tors is that participant expectation of constant peripheral
color results in subjectively experienced color even when
stimulus color is removed, eso es, “filling in” (Balas &
Sinha, 2007; Komatsu, 2006). Another possibility is that
participants consciously experience the desaturation
but fail to report it because of a failure to attend to and
subsequently remember their peripheral experiences,
eso es, inattentional blindness or amnesia (Chen & Wyble,
2016; Mack, 2003; lobo, 1999). Either way, although we
agree with Cohen’s assertion that the participants can have
surprisingly nonveridical color expectation, memory, y
reports under these conditions, we disagree that this
necessarily implies “the immediate impression of a rich,
colorful experience that encompasses their entire visual
world…is surprisingly inaccurate.” (Cohen et al., 2020).
Sin embargo, both virtual reality headsets and our own
Panoworks display provide a means to perform far-field
psychophysics with significant advantages over historical
aproches. Both approaches allow for easy manipulation
of stimulus duration, tamaño, and type in combination with
head fixation and eye tracking. One interesting question
that remains using both techniques is what is the source
of individual differences in subjective reports across the
different stimulus parameter conditions (Figure A3).
Potential candidates include differences in: cortical magni-
ficción (Hombre libre & Simoncelli, 2011); color sensitivity in
the periphery, perhaps because of interindividual cone
receptor density variations (Afraz et al., 2010), or attention.
Último, we wish to conclude with the connections between
our experimental approach and recent developments in
the philosophy of consciousness. A number of philoso-
phers have suggested the conceptual possibility of charac-
terizing phenomenal consciousness through obtaining the
structural relationships between experiences (Sotavento, 2021;
Rosenthal, 2015; Chalmers, 1996; Nagel, 1974), con algunos
going so far as to say these relations provide a complete
description of any experience (Lyre, 2022; Fink et al.,
2021). A similar claim of the feasibility of using a relational
approach to characterize qualia has been made from a
more formal category theoretical perspective (Tsuchiya &
Saigo, 2021; Tsuchiya, Taguchi, & Saigo, 2016). Estos
theoretical research programs go hand in hand with the
popular neuroscientific method of representational similar-
ity analysis (Shinkuma, Nishida, Kado, maeda, & Nishimoto,
2019; Kriegeskorte & Kievit, 2013), among other efforts to
pursue neuroscience of consciousness research related to
qualia spaces (Lau, Michel, El dulce, & flamenco, 2022;
Tallon-Baudry, 2022). Our experimental approach provides
a method for exploring these ideas empirically in a way that
has actual consequences for these relational theories.
Future studies that would also have interesting philosoph-
ical implications while making use of this approach could
extend this work into other modalities than vision, carácter-
terize nonneurotypical populations such as those with
color blindness, or collect subjective relationship reports
in combination with neural recordings.
Zeleznikow-Johnston et al.
527
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
5
2
8
j
oh
tu
r
norte
a
yo
oh
F
C
oh
gramo
norte
i
t
i
v
mi
norte
mi
tu
r
oh
s
C
i
mi
norte
C
mi
V
oh
yo
tu
metro
mi
3
5
,
norte
tu
metro
b
mi
r
4
APPENDIX A
Table A1. Hypothesis Summary Table
Question
1. Are the overall
structures of color
dissimilarity reports at
different eccentricities
lo mismo?
Hipótesis
Sampling Plan
Analysis Plan
H1A) Participants presented with
peripherally magnified physically
equivalent stimuli will provide
CC & PP responses that are
positively correlated.
Recruitment will stop when any of the
following are met:
(cid:129) The BF for the hypothesis exceeds 10
(cid:129) The BF for hypothesis is below 0.1
(cid:129) 50 participants have been recruited.
Participants will be excluded if: they self-
report visual impairments (incluido
glasses); fail to complete all trials; o
score below 77% on catch trials. Todo otro
participants will be included.
H1B) As above, but for CC & CP
As above
H1C) As above, but for CP & PÁGINAS
As above
1. Are physically
equivalent stimuli
presented at differing
eccentricities
subjectively equivalent?
H2A) Participants presented with
peripherally magnified physically
equivalent stimuli will provide
dissimilarity responses closer to
zero than expected from random
responding in the CC condition.
As above
We will calculate the Pearson correlation
of CC and CP judgments for each
partícipe.
We will first Fisher-Z transform these
correlations and then test whether the
mean of their distribution is nonzero
by performing a Bayesian one-sample
t test using the default Jeffreys prior for
the variance and Cauchy prior for the
effect size.
As above, but for CC & CP
As above, but for CP & PÁGINAS
The difference in dissimilarity responses
to identically vs. non-identically colored
stimuli will be converted to a normalized
trace difference for each participant in the
peripherally magnified groups. Lo haremos
fit this data from the CC condition to a
Bayesian beta regression model with no
regressors to calculate the intercept.
We will use the default prior of a mean
de 0 y una desviación estándar de 2.5,
which corresponds to a prior belief that
participants will respond equivalently
for identical and non-identical stimuli.
Interpretation Given
to Different Outcomes
(cid:129) The results are
consistent with the
hypothesis if the
BF for the model
intercept is ≥ 3.
(cid:129) The results are
consistent with the
null hypothesis if the
BF for the model
intercept is ≤ 1/3.
(cid:129) The results will
be taken to be
inconclusive if
1/3 ≤ BF ≤ 3
As above
As above
As above
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
As above
As above
H2B) As above, but for the
As above
As above, but for the CP condition
As above
CP condition
H2C) As above, but for the
As above
As above, but for the PP condition
As above
PP condition
2. Does failure to
peripherally magnify
small stimuli prevent
color equivalence in
the periphery?
As above
H2D) Participants presented
with small, unmagnified stimuli
in the periphery that are
physically equivalent will provide
dissimilarity responses larger
than those of participants shown
larger or peripherally magnified
estímulos, es decir., that there will be an
interaction effect between size
and magnification.
3. Are color-pairs
subjectively equivalent
regardless of
presentación
eccentricity?
As above
H3) Participants will provide the
same dissimilarity responses to
the same pair of colored stimuli
under CC, CP, or PP conditions,
and hence the mean variance in
responses across the conditions
across all color pairs will be
closer to zero than expected
from random responding.
z
mi
yo
mi
z
norte
i
k
oh
w
–
j
oh
h
norte
s
t
oh
norte
mi
t
a
yo
.
5
2
9
As above
As above
The difference in dissimilarity responses
to identically vs. non-identically colored
stimuli will be converted to a normalized
trace difference for each participant.
We will fit the subset of this data
corresponding to PP judgments with a
Bayesian beta regression model with size,
magnification and Size × Magnification as
regressors to calculate the coefficient for
the Size × Magnification regressor. Nosotros
will use the default prior that corresponds
to a prior belief that the coefficient for all
of the regressors is zero.
For each participant in the peripherally
magnified groups, variance in the
dissimilarity responses to pairs of
colored stimuli across the CC, CP, y
PP conditions will be calculated, y el
mean of this variance across all color
pairs obtained. We will also calculate a
participant’s mean variance in the case
where the color pair labels for the
judgments are scrambled for the CP and
PP eccentricities. We will then take the
difference between the scrambled and
original mean variance case for each
participant as a measure of their judgment
consistency across eccentricities.
We will fit this mean variance difference
data to a Bayesian beta regression model
with no regressors to calculate the
intercept. We will use the default prior
of a mean of 0 and a standard deviation
de 2.5, which corresponds to the prior
belief that randomizing the structure of
responses in different eccentricities will
not affect the consistency of participant
judgments across eccentricities.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Figure A1. Panoworks display and setup. Horizontal streaking is an artifact
of the camera and not perceived by participants.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Figure A2. Panoworks display and setup for each condition. (A) FIX SMALL during a PP trial presentation. Stimuli have a 0.1 DVA and are presented
en 60 DVA. The stimuli are too small to be easily seen in this image. (B) FIX BIG during a PP trial presentation. Stimuli are again presented centered at
60 DVA but with a diameter of 2 DVA. (C) MAG SMALL during a CP trial. The central stimulus is presented centered at 1 DVA with a diameter of 0.1
DVA and is too small to see on this image. The peripheral stimulus is centered at 60 DVA with a 2.7-DVA diameter. (D) MAG BIG during a PP trial.
Both stimuli are presented centered at 60 DVA with a 54-DVA diameter.
530
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Figure A3. Individual raw dissimilarity values from the in-laboratory experiment placed into matrices for each participant. Each row is an individual
partícipe, and each column corresponds to a stimulus parameter condition. The plots are asymmetric as participants were presented with each
(non-identical) color pair twice and each response is plotted. The numbers above each plot show the double-pass correlation for each eccentricity
condition for each stimulus parameter condition per participant.
Zeleznikow-Johnston et al.
531
APPENDIX B
Figure B1. A sample color grid presented at the start of the experiment
to familiarize participants with the colors they will be comparing. El
colors were presented in random locations in the grid for each
partícipe.
Table B1. Stimuli Used throughout the Experiments
Hex Code
#7f7f7f
#ff0000
#ffaa00
#aaff00
#00ff00
#00ffa9
#00a9ff
#0000ff
#aa00ff
#ff00aa
HSV
[0, 0, 0.5]
[0, 1, 1]
[40, 1, 1]
[80, 1, 1]
[120, 1, 1]
[160, 1, 1]
[200, 1, 1]
[240, 1, 1]
[280, 1, 1]
[320, 1, 1]
Nombre
Fondo
Color 1
Color 2
Color 3
Color 4
Color 5
Color 6
Color 7
Color 8
Color 9
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Figure B2. Task procedure for catch trials. Ten catch trials are randomly inserted into the main trial sequence. After the 500-msec fixation, en lugar de
stimuli display, participants are instead shown a response screen where they are directed to select a particular value chosen at random and varying
between catch trials. Participants are then directed to click on the green rectangle at the center of the screen to proceed to the next trial.
532
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Figure B3. (A) Schematic for the pilot online experiments. The panel shows the achievable
eccentricities of stimuli with typical computer monitors. Central and peripheral stimuli are
presented at random locations ±30° of horizontal and centered on the pink (10 DVA) y
verde (1 DVA) líneas, respectivamente. (B) Cone photoreceptor densities at central and peripheral
eccentricities relative to the retinal maximum ( watson, 2014; Curcio et al., 1990). Eccentricities
para (A) y (B) are shown by the scale above (A).
APPENDIX C
Figure C1. Pilot online results. Catch trial accuracy (chance performance 12.5%) plotted
against the mean RT on main trials for a participant. The horizontal red line indicates the catch
score cutoff of > 0.77. Only participants above the cutoff were included in the pilot analysis.
Zeleznikow-Johnston et al.
533
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Figure C2. Individual raw dissimilarity values from the online pilot experiment placed into matrices for each participant in the (A) pequeño, fixed-size
estímulos; (B) grande, fixed-size stimuli; (C) pequeño, peripherally magnified stimuli; y (D) grande, peripherally magnified stimuli groups. Participants who
appear to have misunderstood the response instructions and inverted their dissimilarity responses are marked in red. Participants who otherwise
have CC normalized traces over 3.5 are marked in blue. Participants marked in red or blue had their responses inverted for the pilot analysis.
Participants who otherwise are suspected of misunderstanding the instructions and/or task noncompliance are marked in green.
534
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Figure C3. A demonstration of how mean variance is calculated for each participant. (A) The raw dissimilarity values for a representative participant
from the MAG BIG experimental condition. (B) An illustration of how the variance in dissimilarity responses is calculated for three different color
pares. The background color of each row marks the six dissimilarity values marked by the corresponding colored boxes in (A). (C) A histogram of the
variance for all color pairs for the participant. (D) The same participant as in (A), but with their similarity judgments for the CP and PP conditions
randomly scrambled. (mi) The same variance calculations as per (B) but with the scrambled values for CP and PP. (F) The variance in similarity
judgments for all color pairs in the given CP and PP shuffling. The mean variance of the distribution is considerably higher than in (C), indicating the
original similarity judgments were consistent across eccentricity conditions. (G–L) as per (A–F), but for a participant in the FIX SMALL group. Nota
that the mean variance is similar in both the original (I) and scrambled (l) casos, indicating that the participant’s similarity judgments were
inconsistent across eccentricity conditions.
Zeleznikow-Johnston et al.
535
Table C1. Normalized Trace Difference per Condition from the
Online Pilot Experiment (norte = 31) Using Peripherally Magnified
Stimuli Shown at 10 DVA
experiment and provided electronic consent to partici-
pate. Participants were compensated for their time at a
rate of $6/hour.
Condition
CC, CP
CC, PÁGINAS
CP, PÁGINAS
BF10
> 1014
> 1013
> 1013
Table C2. Normalized Trace Difference per Condition from the
Online Pilot Experiment (norte = 31) Using Peripherally Magnified
Stimuli Shown at 10 DVA
Condition
Significar
95% HPD
CC
CP
PÁGINAS
3.31
2.93
3.40
2.49 a 3.99
2.10 a 3.65
2.60 a 4.07
BF10
> 103
> 103
> 103
Table C3. Normalized Trace Difference Data from All Participant
Groups in the Online Pilot Experiment (norte = 31) PP Condition
Fitted to a Bayesian Beta Regression
b
95% HPD
BF10
Interceptar
Size
Magnification
−0.1
−0.47 to 0.27
1.2
0.3
0.57 a 1.86
0.15 a 0.39
Size × Magnification
−0.2
−0.44 to −0.05
1.49
Table C4. Mean Variance from the Online Pilot Experiment
(norte = 31) Using Peripherally Magnified Stimuli Shown at 10 DVA
Significar
2.65
95% HPD
1.27 a 3.96
BF10
20.44
APPENDIX D
Abajo, we describe methods related to the online pilot
experimentos.
Pilot Experiment: Online Version of the
Main Experiment
Ética
Experimental procedures were approved by the Monash
University Human Research Ethics Committee (Proyecto
ID: 17674). Participants were provided electronically with
written consent forms before the commencement of the
Diseño
Participants were recruited remotely
Participantes.
through CloudResearch (Litman, robinson, & Abberbock,
2017), a virtual wrapper for Amazon’s Mechanical Turk
(MTRT) platform. Participants accessed the experiment
through the MTurk worker area and provided data using
their own personal computers. Only English native
speakers from Australia, Canada, India, and the United
States of America were recruited. Participants were famil-
iar with the online MTurk platform as they were only
invited to participate if they had performed at least 1000
unrelated tasks online with at least 97% aprobación. Participe-
pants were only recruited from those with a Windows
operating system.
Display apparatus. Because of the nature of online
experimentation, participants used their own computer
screen to perform the experiment. Screen properties neces-
sary for appropriate stimulus display were obtained via
calibration steps at the beginning of the task, descrito
below in Procedure section. Stimuli were presented via
Inquisit 6 (milisegundo) display software. Centrally pre-
sented stimuli were presented at 1 DVA as per the main
métodos. Peripherally presented stimuli in the online
experiment were centered at 10 DVA, outside of the
macula’s visual field and thus in the near periphery
(Figure B3).
Consent. Once participants agreed to participate in our
study in the TurkPrime interface, they were provided with
a link that directs them to our study hosted by Millisecond.
The first page of the experiment was a consent form that
they could electronically sign by pressing the spacebar.
Participants were informed that the data collection pro-
cess was anonymous and that they could quit the experi-
ment at any time.
Screen size calibrations. Following the consent page,
participants were directed through a series of steps to
obtain both their physical screen size and viewing distance
from the screen and were instructed that they must per-
form these properly to complete the task. La siguiente
protocol has been adapted from a previous study validat-
ing this method (li, Joo, Yeatman, & Reinecke, 2020).
Participants were asked to place a card (p.ej., credit card,
student card, or any other card that is the same dimension
as a credit card: 8.6 cm × 5.4 centímetro) against the screen and
adjust a rectangle on their screen to match the size of the
physical card. As these cards come in a standard size, este
allowed us to infer the screen size from the number of
pixels in the displayed rectangle. Participants were then
directed to fixate on a point on the right-hand side of their
536
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
screen with their left eye while closing their right eye. Par-
ticipants were then shown a small circle that moves from
right to left and asked to press the ENTER key when the
circle disappears from view, which occurs upon entering
the participant’s blind spot. As the blind spot is located
about 13.5° from the center of vision, this allowed for trig-
onometric calculation of the viewing distance if the screen
size is known. If the calibration values suggest the partici-
pant would be unable to complete the experiment
because of inadequate screen size, participants are
rejected from the experiment. For a demonstration of
the calibration procedure, see the online version of the
tarea: https://mili2nd.co/frub.
Tarea. Online participants performed trials with only
one stimulus size and peripheral magnification setting.
Each participant completed 81 CC, 81 CP, y 81 PP main
trials and 10 catch trials, equaling 253 ensayos. Todo otro
methodological details are as per the main text.
Sampling Plan
Participant quality control
Participant exclusion.
occurred both before and after the main experiment.
Before the main experiment and after the calibration
procedimiento, we stopped the experiment if participants’
estimated screen size did not allow us to present the
peripheral stimuli without being cut off by the edge of
the screen. Además, we excluded participants whose
estimated screen diameter is less than 26 cm or viewing
distance is less than 30 centímetro, as these values would suggest
the task is being performed on a phone or tablet.
Following the main experiment, we calculated the mean
and standard deviation of the participant catch scores. Nosotros
excluded participants who scored less than 1 SD below
the median (77%) on catch trials, as this is suggestive of
inattentive behavior (Figure C1). Participants who did
not complete all of the main experimental trials were also
excluded. All other participants were included in the data
análisis.
Analysis Plan
Preprocesamiento. Upon visual inspection of the online
datos, it appeared that some participants provided dissimi-
larity responses that were inverted relative to what would
be expected even in central vision presentation condi-
ciones, presumably because of misunderstanding the
response method. Como resultado, for the pilot data, nosotros
inverted the dissimilarity responses of participants with
a CC normalized trace > 3.5. We will not perform this
for the laboratory-based experiment should it occur then.
En cambio, we have optimized the task instructions and
added practice trial feedback to make participant misun-
derstandings less likely. All other analyses are the same
as per the main text.
Control Experiment 1: Similarity Judgments in the
Central Fovea
As a check of the efficacy and robustness of our methods,
we performed a control experiment to assess the impact of
the absence of blue-sensitive S-cones in the central retina
on color similarity judgments. The central fovea is similar
to the condition of tritanopia, which results in impair-
ments in blue-green color discrimination ( Williams et al.,
1981). We attempted to replicate these previous findings
using our similarity judgment technique.
Online participants performed a modified version of
our color similarity judgment task, where stimuli varied
either from blue to green (prueba) or blue to red (control).
Before commencing the task, participants performed a
heterochromatic flicker adjustment task to equilibrate
the luminance values for their specific monitor ( Wagner
& Boynton, 1972; Figure D1A). This was intended to
increase the likelihood that stimuli varied subjectively in
chroma rather than in luminance. Participants also per-
formed the screen size calibrations as per Experiment 1.
Following the calibrations, participants performed a mod-
ified version of our original task (Figure D1B). Estímulos
were presented under two conditions: 0°/1° or 1°/1°. En
the 0° position, stimuli with a diameter of 0.1° were pre-
sented centrally at 0°, which is free of S-cones (Magnussen
et al., 2004). In the 1° position, stimuli with a diameter of
0.45° are presented in a region of the fovea with S-cones.
The fixation cross was changed to a fixation circle to avoid
overlap with the 0° stimuli. The stimuli presentation time
was changed from 250 msec to 100 msec to help prevent
microsaccades. All other aspects, including the response
protocolo, remained the same as the previously reported
experimentos. Primero, we the authors performed the task,
which was also replicated by n = 14 online participants
with similar results.
Figure D1C (arriba) shows the dissimilarity rating matrix as
expected from tritanopic, blue-green blind vision. All pairs
of isoluminant blue-green patches between 0° and 1°
patches look similar (es decir., the mean rating of ∼2 out of
7), with an indistinct relationship structure. A diferencia de,
participants gave a much clearer and structured dissimilar-
ity rating matrix as expected from normal vision when they
compared two small color patches both at 1° (Figure D1C,
abajo). Participant dissimilarity responses to physically
identical pairs (the mean of the diagonal, es decir., normalized
trace) were much smaller when both stimuli were dis-
played on retinal areas with S-cones present (Figure D1;
pag = .014). A control experiment with isoluminant red-
green patches (Figure D1E and F) confirmed that this pat-
tern was not observed ( pag = .985).
Control Experiment 2: Examining the Possibility of
Screen Chromatic Heterogeneity
We examined the effects of possible variation in the chro-
matic properties on a participant’s monitor. para hacerlo, nosotros
Zeleznikow-Johnston et al.
537
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Figure D1. Color similarity
judgments in the central fovea.
(A) Participant-specific color
equiluminance values were
obtained via the minimal-flicker
técnica. Los participantes fueron
shown a stimulus flickering
between two solid colors (cualquiera
blue/green or red/green) en
the center of the screen and
asked to adjust the brightness
of one of the colors using a
slider until the flickering was
minimized and the colors
appeared equiluminant. (B) El
main similarity judgment task
followed the luminance and
screen size calibrations. El
calibrations allowed participants
to be shown a set of stimuli
that were isoluminant on their
monitors at the center of the
pantalla. Stimuli presentation was
preceded by a fixation circle
(500 mseg) with an interior
larger than the central stimulus.
(C) Group-mean dissimilarity
judgment matrices for
experiences generated by
equiluminant stimuli varying
from green to blue in 0° versus
1° comparison (arriba) and 1°
versus 1° (abajo) condiciones.
Note that stimuli were always
displayed within the fovea.
(D) The normalized traces from
the dissimilarity matrices for
participants show stimuli that
varied from blue to green.
Participants rated the physically
identical blue–green pairs as
more different (es decir., clasificación
of ∼2) in the 0° versus 1°
comparison condition but
more similar between the 1°
and 1° comparison condition
(es decir., ∼0.7). (E and F) Lo mismo que (C) y (D) but for red to green. Red–green pairs looked similar at both the 1° versus 1° (es decir., ∼0.7) y
0° versus 1° (es decir., ∼1.2). Boxplots centerline is median, box shows 25th–75th percentile, and whiskers show the largest value within 150% de
the interquartile range. n = 13–14.
modified Experiment 1, systematically varying the retinal
or screen location of a stimulus (Figure D2). Online partic-
ipants performed color similarity judgments for stimuli
presented under 2 (screen location: fixed or not) × 2 (ret-
inal location: fixed or not; Figure D2A and B). Each partic-
ipant provided a full set of judgments for all color pairs
under each of the four conditions, with the trials for differ-
ent conditions randomly interspersed. Three different
stimulus presentation points were selected for the exper-
mento, each 5° apart. Each trial had four phases. Primero, a fix-
ation cross was presented at the left side of the screen for
500 mseg, followed by a stimulus for 250 msec in one
of two possible locations. After this, a fixation cross
(500 mseg) was presented in one of two locations,
followed by an additional stimulus (250 mseg) in one of
two possible locations depending on the second fixation
punto. This design meant that stimuli always had a 50%
probability of presentation to the left or right of a fixation
cross after presentation.
The same pattern of similarity relationships are
observed regardless of condition (Figure D2C). Esta sugerencia-
gests that chromatic inhomogeneity of commercial dis-
plays does not affect the pattern of similarity judgments
provided. We note also that as our methods are based
on analyzing the structure of sets of similarity judgments
(p.ej., rather than binary classification of stimuli), ellos son
robust to minor changes in stimulus properties that lead
to only minor changes in similarity structure.
538
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Figure D2. Examining the possibility of screen chromatic heterogeneity. Online participants performed color similarity judgments for stimuli
presented under four different conditions. Each participant provided a full set of judgments for all color pairs under all four conditions, with the trials
for different conditions randomly interspersed. (A) Two different fixation points and three different stimulus presentation points were selected for
the experiment, each 5° apart. (B) Illustration of the four possible conditions for stimuli presentation: screen location fixed, retinal location fixed,
neither, o ambos. (C) The mean dissimilarity judgments for each color pair across participants. Each participant performed all four conditions. El
same pattern of similarity relationships are observed regardless of condition. norte = 7.
Zeleznikow-Johnston et al.
539
Expresiones de gratitud
A. Z.-J./ N. t. are supported by Australian Research Council
(DP180100396), National Health and Medical Research Council
(APP1183280), and the Foundational Questions Institute and
Fetzer Franklin Fund, a donor advised fund of Silicon Valley
Community Foundation (FQXi-RFP-CPW-2017). norte. T./ M. Y.
are supported by Grant-in-Aid for Transformative Research
Areas (B; norte. T.: 20H05710, METRO. y.: 20H05711) from Japan Society
for the Promotion of Science. METRO. Y. is supported by Moonshot
R&D Grant ( JPMJMS2295-01) from Japan Science and Technol-
ogy Agency and KAKENHI (22H01108, 22 K18265) from Japan
Society for the Promotion of Science. The funders have/had
no role in study design, Recogida y análisis de datos, decisión
a publicar, o preparación del manuscrito. We would addi-
tionally like to thank Ruitong Fan for helping develop the initial
pilot experiment.
Reprint requests should be sent to Ariel Zeleznikow-Johnston,
Turner Institute for Brain and Mental Health & School of Psy-
chological Sciences, Faculty of Medicine, Nursing, and Health
Ciencias, Monash University, 770 Blackburn Rd., Melbourne,
Victoria, Australia, 3800, o por correo electrónico: ariel.zeleznikow
-johnston@monash.edu, or Makiko Yamada, Institute for
Quantum Medical Science, National Institutes for Quantum and
Radiological Science and Technology, 4-9-1 Anagawa, Inage-ku,
Chiba, 263-8555, Japón, o por correo electrónico: yamada.makiko@qst.go.jp.
Declaración de disponibilidad de datos
We published a preregistered research plan on the Open
Science Framework before data collection (https://osf.io
/rg7kf/). Our experimental code, raw data, and analysis
code are all available on the Open Science Framework
(https://osf.io/5pfrg/).
Contribuciones de autor
Ariel Zeleznikow-Johnston: Conceptualización; Data cura-
ción; Análisis formal; Investigación; Metodología; Proyecto
administración; Software; Visualización; Writing—Original
borrador; Writing—Review & edición. Yasunori Aizawa: Datos
curation; Investigación; Administración de proyecto; Writing—
Revisar & edición. Makiko Yamada: Adquisición de financiación;
Administración de proyecto; Supervisión; Writing—Review &
edición. Naotsugu Tsuchiya: Conceptualización; Formal
análisis; Adquisición de financiación; Metodología; Proyecto
administración; Supervisión; Writing—Review & edición.
Diversidad en las prácticas de citas
Análisis retrospectivo de las citas en cada artículo publicado.-
publicado en esta revista de 2010 a 2021 revela una persistente
patrón de desequilibrio de género: Aunque las proporciones de
equipos de autoría (categorizado por iden de género estimado-
tificación del primer autor/último autor) publicando en la revista-
final de neurociencia cognitiva ( JoCN) durante este período
fueron M(un)/m = .407, W.(Omán)/m = .32, M/M = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/M = .109, and W/ W = .085 (Postle y
fulvio, JoCN, 34:1, páginas. 1–3). Como consecuencia, JoCN anima-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance.
REFERENCIAS
Abramov, I., gordon, J., & chan, h. (1991). Color appearance in
the peripheral retina: Effects of stimulus size. Diario de la
Optical Society of America A, 8, 404–414. https://doi.org/10
.1364/JOSAA.8.000404, PubMed: 2007915
Adelson, mi. h. (2000). Lightness perception and lightness illusions.
En m. Gazzaniga (Ed.), The new cognitive neurosciences
(2y ed., páginas. 339–351). Cambridge, MAMÁ: CON prensa.
Afraz, A., Pashkam, METRO. v., & Cavanagh, PAG. (2010). Espacial
heterogeneity in the perception of face and form attributes.
Biología actual, 20, 2112–2116. https://doi.org/10.1016/j.cub
.2010.11.017, PubMed: 21109440
anderson, S. J., Mullen, k. T., & Hesse, R. F. (1991). Humano
peripheral spatial resolution for achromatic and chromatic
estímulos: Limits imposed by optical and retinal factors. Diario
of Physiology, 442, 47–64. https://doi.org/10.1113/jphysiol
.1991.sp018781, PubMed: 1798037
Anstis, S. (1998). Picturing peripheral acuity. Percepción,
27, 817–825. https://doi.org/10.1068/p270817, PubMed:
10209644
Ayama, METRO., Sakurai, METRO., Carlander, o., Derefeldt, GRAMO., & Eriksson,
l. (2004). Color appearance in peripheral vision. En B. mi.
Rogowitz & t. norte. Pappas (Editores.), Proceedings of the Human
Vision and Electronic Imaging IX (pag. 260). https://doi.org/10
.1117/12.522240
Balas, B., & Sinha, PAG. (2007). “Filling-in” colour in natural
escenas. Cognición visual, 15, 765–778. https://doi.org/10
.1080/13506280701295453
Bonnardel, v., Beniwal, S., Dubey, NORTE., Pande, METRO., Ajo, K.,
& Bimler, D. (2016). Perceptual color spacing derived from
maximum likelihood multidimensional scaling. Diario de la
Optical Society of America A, 33, A30–A36. https://doi.org/10
.1364/JOSAA.33.000A30, PubMed: 26974936
Bosten, j. METRO., robinson, j. D., Jordán, GRAMO., & Mollon, j. D. (2005).
Multidimensional scaling reveals a color dimension unique to
‘color-deficient’ observers. Biología actual, 15, R950–R952.
https://doi.org/10.1016/j.cub.2005.11.031, PubMed: 16332521
Quemaduras, B., & Shepp, B. mi. (1988). Dimensional interactions and
the structure of psychological space: The representation of
hue, saturation, and brightness. Percepción & Psicofísica,
43, 494–507. https://doi.org/10.3758/BF03207885, PubMed:
3380640
Carroll, S., & Dennett, D. C. (2020). Daniel Dennett on minds,
patrones, and the scientific image – Sean Carroll (No. 78).
https://www.preposterousuniverse.com/podcast/2020/01/06
/78-daniel-dennett-on-minds-patterns-and-the-scientific
-image/
Chalmers, D. j. (1996). The conscious mind: In search of a
fundamental theory. prensa de la Universidad de Oxford.
Chater, norte. (2018). Mind is flat: The remarkable shallowness of
the improvising brain. Prensa de la Universidad de Yale. https://doi.org
/10.12987/9780300240610
Chen, h., & Wyble, B. (2016). Attribute amnesia reflects a
lack of memory consolidation for attended information.
Revista de Psicología Experimental: Percepcion humana
y rendimiento, 42, 225–234. https://doi.org/10.1037
/xhp0000133, PubMed: 26348066
cohen, METRO. A., Botch, t. l., & Robertson, C. mi. (2020). El
limits of color awareness during active, real-world vision.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
540
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
117, 13821–13827. https://doi.org/10.1073/pnas.1922294117,
PubMed: 32513698
cohen, METRO. A., & Rubenstein, j. (2020). How much color do we
see in the blink of an eye? Cognición, 200, 104268. https://doi
.org/10.1016/j.cognition.2020.104268, PubMed: 32473406
Cribari-Neto, F., & Zeileis, A. (2010). Beta regression in R.
Journal of Statistical Software, 34, 1–24. https://doi.org/10
.18637/jss.v034.i02
Curcio, C. A., & allen, k. A. (1990). Topography of ganglion
cells in human retina. Journal of Comparative Neurology,
300, 5–25. https://doi.org/10.1002/cne.903000103, PubMed:
2229487
Curcio, C. A., Lento, k. r., Kalina, R. MI., & Hendrickson, A. mi.
(1990). Human photoreceptor topography. Diario de
Comparative Neurology, 292, 497–523. https://doi.org/10
.1002/cne.902920402, PubMed: 2324310
Daniel, PAG. METRO., & Whitteridge, D. (1961). The representation of
the visual field on the cerebral cortex in monkeys. Diario de
Physiology, 159, 203–221. https://doi.org/10.1113/jphysiol
.1961.sp006803, PubMed: 13883391
Decock, l., & Douven, I. (2013). Qualia compression.
Philosophy and Phenomenological Research, 87, 129–150.
https://doi.org/10.1111/j.1933-1592.2011.00545.x
Ennis, R. J., & Zaidi, q. (2019). Geometrical structure of
perceptual color space: Mental representations and
adaptation invariance. Revista de visión, 19, 1. https://doi.org
/10.1167/19.12.1, PubMed: 31573606
Fink, S. B., Kob, l., & Lyre, h. (2021). A structural constraint
on neural correlates of consciousness. Philosophy and
the Mind Sciences, 2, 7. https://doi.org/10.33735/phimisci
.2021.79
Hombre libre, J., & Simoncelli, mi. PAG. (2011). Metamers of the ventral
stream. Neurociencia de la naturaleza, 14, 1195–1201. https://doi.org
/10.1038/nn.2889, PubMed: 21841776
Gegenfurtner, k. r., Bloj, METRO., & Toscani, METRO. (2015). The many
colours of ‘the dress.’ Current Biology, 25, R543–R544.
https://doi.org/10.1016/j.cub.2015.04.043, PubMed: 25981790
Giron, C., Lau, h., & Knotts, j. D. (2018). Are open interviews
superior to button presses? A commentary on Haun et al.
(2017). Symposium on the Brains Blog [Blog post]. https://
philosophyofbrains.com/2018/04/13/symposium-on-haun
-tononi-koch-and-tsuchiya-are-we-underestimating-the
-richness-of-visual-experience.aspx
Goodrich, B., Gabry, J., Alí, I., & Brilleman, S. (2020). Rstanarm:
Bayesian applied regression modeling via Stan. RAPACIO R
versión 2.21.1. https://mc-stan.org/rstanarm
gordon, J., & Abramov, I. (1977). Color vision in the peripheral
retina. II. Hue and saturation. Revista de la Sociedad Óptica
de América, 67, 202–207. https://doi.org/10.1364/JOSA.67
.000202, PubMed: 839300
Hansen, T., Pracejus, l., & Gegenfurtner, k. R. (2009). Color
perception in the intermediate periphery of the visual field.
Revista de visión, 9, 26. https://doi.org/10.1167/9.4.26,
PubMed: 19757935
Haun, A. METRO. (2021). What is visible across the visual field?
Neurociencia de la conciencia, 2021, niab006. https://doi
.org/10.1093/nc/niab006, PubMed: 34084558
Helm, C. mi. (1964). Multidimensional ratio scaling analysis of
perceived color relations. Journal of the Optical Society of
America, 54, 256–262. https://doi.org/10.1364/JOSA.54
.000256, PubMed: 14123941
Humphrey, GRAMO. K., & Goodale, METRO. A. (1998). Sondeo
unconscious visual processing with the McCollough effect.
Conciencia y cognición, 7, 494–519. https://doi.org/10
.1006/ccog.1998.0369, PubMed: 9787058
Izmailov, C. A., & Sokolov, mi. norte. (1991). Spherical model of
color and brightness discrimination. ciencia psicológica,
2, 249–260. https://doi.org/10.1111/j.1467-9280.1991
.tb00143.x
Kahn, MI., Dement, w., Pescador, C., & Barmack, j. mi. (1962).
Incidence of color in immediately recalled dreams. Ciencia,
137, 1054–1055. https://doi.org/10.1126/science.137.3535
.1054, PubMed: 14453103
Komatsu, h. (2006). The neural mechanisms of perceptual
filling-in. Naturaleza Reseñas Neurociencia, 7, 220–231. https://
doi.org/10.1038/nrn1869, PubMed: 16495943
Kriegeskorte, NORTE., & Kievit, R. A. (2013). Representational
geometría: Integrating cognition, cálculo, and the brain.
Tendencias en Ciencias Cognitivas, 17, 401–412. https://doi.org/10
.1016/j.tics.2013.06.007, PubMed: 23876494
Lau, h., Michel, METRO., El dulce, j. MI., & flamenco, S. METRO. (2022). El
mnemonic basis of subjective experience. Reseñas de naturaleza
Psicología, 1, 479–488. https://doi.org/10.1038/s44159-022
-00068-6
Sotavento, A. Y. (2021). Modeling mental qualities. Philosophical Review,
130, 263–298. https://doi.org/10.1215/00318108-8809919
Lennie, PAG. (1998). Single units and visual cortical organization.
Percepción, 27, 889–935. https://doi.org/10.1068/p270889,
PubMed: 10209632
li, P., Joo, S. J., Yeatman, j. D., & Reinecke, k. (2020).
Controlling for participants’ viewing distance in large-scale,
psychophysical online experiments using a Virtual Chinrest.
Informes Científicos, 10, 904. https://doi.org/10.1038/s41598-019
-57204-1, PubMed: 31969579
Litman, l., robinson, J., & Abberbock, t. (2017). TurkPrime
.com: A versatile crowdsourcing data acquisition platform for
the behavioral sciences. Métodos de investigación del comportamiento, 49,
433–442. https://doi.org/10.3758/s13428-016-0727-z, PubMed:
27071389
Logvinenko, A. D., & Maloney, l. t. (2006). The proximity
structure of achromatic surface colors and the impossibility of
asymmetric lightness matching. Percepción & Psicofísica,
68, 76–83. https://doi.org/10.3758/BF03193657, PubMed:
16617831
Lyre, h. (2022). Neurophenomenal structuralism. A philosophical
agenda for a structuralist neuroscience of consciousness.
Neurociencia de la conciencia, 2022, niac012. https://doi
.org/10.1093/nc/niac012, PubMed: 36004320
Mack, A. (2003). Inattentional blindness: Looking without
seeing. Current Directions in Psychological Science, 12,
180–184. https://doi.org/10.1111/1467-8721.01256
Magnussen, S., Spillmann, l., Stürzel, F., & Werner, j. S. (2004).
Unveiling the foveal blue scotoma through an afterimage.
Vision Research, 44, 377–383. https://doi.org/10.1016/j.visres
.2003.09.023, PubMed: 14659964
Makowski, D., Ben-Shachar, METRO. S., & Lüdecke, D. (2019).
BayestestR: Describing effects and their uncertainty,
existence and significance within the Bayesian framework.
Journal of Open Source Software, 4, 1541. https://doi.org/10
.21105/joss.01541
McKeefry, D. J., Murray, I. J., & Parry, norte. R. A. (2007). Perceived
shifts in saturation and hue of chromatic stimuli in the near
peripheral retina. Journal of the Optical Society of America
A, 24, 3168–3179. https://doi.org/10.1364/JOSAA.24.003168,
PubMed: 17912307
Mullen, k. t. (1991). Colour vision as a post-receptoral
specialization of the central visual field. Vision Research, 31,
119–130. https://doi.org/10.1016/0042-6989(91)90079-k,
PubMed: 2006545
Nagel, t. (1974). What is it like to be a bat? Filosófico
Revisar, 83, 435–450. https://doi.org/10.2307/2183914
Nimeroff, I., & Yurow, j. A. (1965). Degree of metamerism.
Journal of the Optical Society of America, 55, 185–190.
https://doi.org/10.1364/JOSA.55.000185
Zeleznikow-Johnston et al.
541
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Rajananda, S., Peters, METRO. A. K., Lau, h., & Odegaard, B. (2017).
Subjective inflation of color saturation in the periphery under
temporal overload. bioRxiv. https://doi.org/10.1101/227074
Rosenholtz, R. (2016). Capabilities and limitations of peripheral
visión. Annual Review of Vision Science, 2, 437–457. https://
doi.org/10.1146/annurev-vision-082114-035733, PubMed:
28532349
Rosenthal, D. (2015). Quality spaces and sensory modalities.
En P. Coates & S. Coleman (Editores.), Phenomenal qualities
(páginas. 33–65). prensa de la Universidad de Oxford. https://doi.org/10.1093
/acprof:oso/9780198712718.003.0002
Sakurai, METRO., Ayama, METRO., & Kumagai, t. (2003). Color appearance
in the entire visual field: Color zone map based on the
unique hue component. Journal of the Optical Society of
America A, 20, 1997–2009. https://doi.org/10.1364/JOSAA.20
.001997, PubMed: 14620327
Schönbrodt, F. D., & carpinteros, E.-J. (2018). Bayes factor
design analysis: Planning for compelling evidence. psiconomico
Boletín & Revisar, 25, 128–142. https://doi.org/10.3758
/s13423-017-1230-y, PubMed: 28251595
Shinkuma, r., Nishida, S., Kado, METRO., maeda, NORTE., & Nishimoto, S.
(2019). Relational network of people constructed on the basis
of similarity of brain activities. IEEE Access, 7, 110258–110266.
https://doi.org/10.1109/ACCESS.2019.2933990
simons, D. J., Franconeri, S. l., & Reimer, R. l. (2000). Cambiar
blindness in the absence of a visual disruption. Percepción,
29, 1143–1154. https://doi.org/10.1068/p3104, PubMed:
11220207
Smithson, METRO., & Verkuilen, j. (2006). A better lemon squeezer?
Maximum-likelihood regression with beta-distributed dependent
variables. Psychological Methods, 11, 54–71. https://doi.org
/10.1037/1082-989X.11.1.54, PubMed: 16594767
Strasburger, h., Rentschler, I., & Juttner, METRO. (2011). Peripheral
vision and pattern recognition: Una revisión. Diario de
Vision, 11, 13. https://doi.org/10.1167/11.5.13, PubMed:
22207654
Tallon-Baudry, C. (2022). The topological space of subjective
experiencia. Tendencias en Ciencias Cognitivas, 26, 1068–1069.
https://doi.org/10.1016/j.tics.2022.09.002, PubMed: 36243671
Tsuchiya, NORTE., & Saigo, h. (2021). A relational approach to
consciousness: Categories of level and contents of
consciousness. Neurociencia de la conciencia, 2021,
niab034. https://doi.org/10.1093/nc/niab034, PubMed:
34659799
Tsuchiya, NORTE., Taguchi, S., & Saigo, h. (2016). Using category
theory to assess the relationship between consciousness and
integrated information theory. Neuroscience Research, 107,
1–7. https://doi.org/10.1016/j.neures.2015.12.007, PubMed:
26748074
van Boxtel, j. j. A., Tsuchiya, NORTE., & Koch, C. (2010). Opposing
effects of attention and consciousness on afterimages.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
107, 8883–8888. https://doi.org/10.1073/pnas.0913292107,
PubMed: 20424112
Vanston, j. MI., & Crognale, METRO. A. (2018). Effects of eccentricity
on color contrast. Journal of the Optical Society of America
A, 35, B122–B129. https://doi.org/10.1364/JOSAA.35.00B122,
PubMed: 29603965
carpinteros, E.-J., Lodewyckx, T., Kuriyal, h., & Grasman, R.
(2010). Bayesian hypothesis testing for psychologists: A
tutorial on the savage–dickey method. Psicología cognitiva,
60, 158–189. https://doi.org/10.1016/j.cogpsych.2009.12.001,
PubMed: 20064637
Wagner, GRAMO., & Boynton, R. METRO. (1972). Comparison of four
methods of heterochromatic photometry. Diario de la
Optical Society of America, 62, 1508–1515. https://doi.org/10
.1364/JOSA.62.001508, PubMed: 4643012
watson, A. B. (2014). A formula for human retinal ganglion cell
receptive field density as a function of visual field location.
Revista de visión, 14, 15. https://doi.org/10.1167/14.7.15,
PubMed: 24982468
williams, D. r., MacLeod, D. I. A., & Hayhoe, METRO. METRO. (1981).
Foveal tritanopia. Vision Research, 21, 1341–1356. https://doi
.org/10.1016/0042-6989(81)90241-8, PubMed: 6976039
lobo, j. METRO. (1999). Inattentional amnesia. In Fleeting
recuerdos: Cognition of brief visual stimuli (páginas. 71–94).
Cambridge, MAMÁ: CON prensa.
Cual, J., chi, X., & Makous, W.. (1995). Zero frequency masking
and a model of contrast sensitivity. Vision Research, 35,
1965–1978. https://doi.org/10.1016/0042-6989(94)00285-t,
PubMed: 7660602
Yeshurun, y., & Carrasco, METRO. (1998). Attention improves or
impairs visual performance by enhancing spatial resolution.
Naturaleza, 396, 72–75. https://doi.org/10.1038/23936, PubMed:
9817201
Yeshurun, y., & Carrasco, METRO. (2000). The locus of attentional
effects in texture segmentation. Neurociencia de la naturaleza, 3,
622–627. https://doi.org/10.1038/75804, PubMed: 10816320
542
Revista de neurociencia cognitiva
Volumen 35, Número 4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
4
5
0
9
2
0
7
3
6
5
6
/
j
oh
C
norte
_
a
_
0
1
9
6
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3