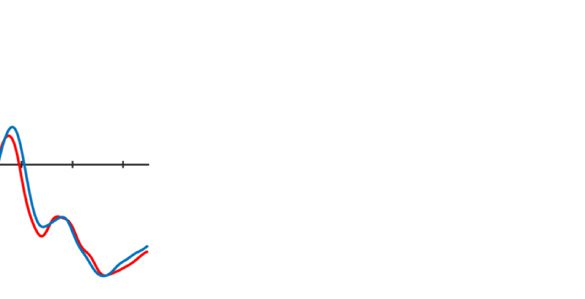Anticipatory Distractor Suppression Elicited
by Statistical Regularities in Visual Search
Benchi Wang, Joram van Driel, Eduard Ort, and Jan Theeuwes
Abstracto
■ Salient yet irrelevant objects often capture our attention and in-
terfere with our daily tasks. Distraction by salient objects can be re-
duced by suppressing the location where they are likely to appear.
The question we addressed here was whether suppression of fre-
quent distractor locations is already implemented beforehand, in an-
ticipation of the stimulus. Using EEG, we recorded cortical activity of
human participants searching for a target while ignoring a salient dis-
tractor. The distractor was presented more often at one location
than at any other location. We found reduced capture for distractors
at frequent locations, indicating that participants learned to avoid
distraction. críticamente, we found evidence for “proactive suppression”
as already “prior to display onset,” there was enhanced power in
parieto-occipital alpha oscillations contralateral to the frequent dis-
tractor location—a signal known to occur in anticipation of irrelevant
información. Locked to display onset, ERP analysis showed a distrac-
tor suppression-related distractor positivity (PD) component for this
ubicación. En tono rimbombante, this PD was found regardless of whether dis-
tracting information was presented at the frequent location. In addi-
ción, there was an early PD component representing an early
attentional index of the frequent distractor location. Nuestros resultados
show anticipatory (proactive) suppression of frequent distractor
locations in visual search already starting prior to display onset. ■
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
oh
C
norte
_
a
_
0
1
4
3
3
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INTRODUCCIÓN
In everyday life, we try to attend to events that are rele-
vant to us and ignore information that could distract us.
Por ejemplo, when driving along a busy road, we devote
attention to relevant objects such as road signs and other
cars while ignoring salient irrelevant objects such as bill-
boards. The extent to which we are able to avoid distrac-
tion from salient events has been a central question for
many years.
Traditionally, attentional selection is assumed to be the
result of the interaction between the goals of the observer
(current selection goals) and the physical properties of the
visual environment (salience of the objects; Theeuwes,
2010; Itti & Koch, 2001; Egeth & Yantis, 1997). Recientemente, él
was pointed out that, in many cases, selection is neither the
result of goals of the observer nor the result of stimulus-
driven (es decir., bottom–up saliency) factores (Failing &
Theeuwes, 2018; Theeuwes, 2018; Awh, Belopolsky, &
Theeuwes, 2012). A third category labeled as “selection his-
tory” referred to those instances in which selection is the
result of previous attentional deployments (Theeuwes,
2019). These previous deployments can result in lingering
and enduring selection biases that are neither related to
the current goals nor to the saliency of objects.
A well-known example of how selection history affects
attentional selection is research known as “contextual
cueing,” demonstrating that search for a target is
Vrije Universiteit Amsterdam
© 2019 Instituto de Tecnología de Massachusetts
facilitated when it appears in a visual layout that was pre-
viously searched relative to visual layouts that were never
seen before ( Jiang & Chun, 2001; Chun & Jiang, 1998,
1999). These studies have highlighted that people pick
up on the statistical regularities present in the display,
such that they selectively attend those locations that
are relevant for the task. More recently, it was also dem-
onstrated that people did not only activate locations that
are relevant but also selectively inhibit locations of
salient distractors that distract them (Failing, Wang, &
Theeuwes, 2019; Wang & Theeuwes, 2018a, 2018b,
2018C). In these studies, Wang and Theeuwes used the
well-established additional singleton task (Theeuwes,
1992) in which observers search for a unique shape sin-
gleton while a unique and salient color singleton is simul-
taneously present. Because observers cannot search for
one specific target feature, large capture effects are typi-
cally observed in this paradigm (see Wang & Theeuwes,
2018C). Wang and Theeuwes also manipulated the distri-
butional properties of the location of salient distractor. Él
was shown that when a salient distractor appeared more
often in one location than in all other locations, its dis-
tracting effect (the extent to which it captured attention)
was attenuated, suggesting that this frequent distractor
location competed less for attention than all other loca-
ciones. They also showed that when the target happened
to be presented at that location, its selection was ham-
pered (see Ferrante et al., 2018, for similar results).
Además, there was a spatial gradient of suppression
around this location.
Revista de neurociencia cognitiva 31:10, páginas. 1535–1548
doi:10.1162/jocn_a_01433
The finding that we are able to inhibit frequent dis-
tractor locations that are likely to contain salient dis-
tractor has important implications for theories of visual
selection and other related fields. ERP studies have
shown that ignoring of a salient distractor is typically accom-
panied by a distractor positivity (PD) component in the ERP
waveform (Feldmann-Wüstefeld, Uengoer, & Schubö, 2015;
Burra & Kerzel, 2014; Gaspar & McDonald, 2014; Sawaki &
Luck, 2010), indicating spatial suppression of its location
during search (Hickey, Di Lollo, & McDonald, 2008).
Sin embargo, it remains unclear whether such suppression
of frequent distractor locations is already implemented
beforehand, in anticipation of the stimulus.
En el estudio actual, using EEG, we recorded cortical
activity of human participants who were searching for a
objetivo, while ignoring a salient distractor. The salient
distractor was presented more often in one location
(high-probability location) than in all other locations
(low-probability location). Consistent with a suppression
account and our previous behavioral studies ( Wang &
Theeuwes, 2018a, 2018b, 2018C), we expected to observe
the classic distractor-suppression-related PD component
in the ERP signal when a distractor was presented at
the high-probability location. Sin embargo, if spatial suppres-
sion to this location is applied well before display onset,
we would also expect to observe increased power in
parieto-occipital alpha oscillations contralateral to the lo-
cation that was more likely to contain the distractor rela-
tive to the ipsilateral location ( Worden, Foxe, Wang, &
Simpson, 2000; see Klimesch, 1999, para una revisión). Nosotros también
conducted a baseline experiment in which all locations
were equally likely to contain a distractor (es decir., sin
any statistical regularities), and here, we expected to see
no EEG components related to spatial suppression.
MÉTODOS
The study was approved by the ethical review committee
of the Faculty of Behavioral and Movement Sciences of the
Vrije Universiteit Amsterdam.
Participantes
Separate groups with 18 participants each (13 women
y 5 men in Experiment 1 with a mean age of 20.9
años; 17 women and 1 man in Experiment 2 con un
mean age of 19.7 años) took part in the study. All par-
ticipants provided written informed consent prior to the
experiment and were reimbursed for their participation
either with money or course credit. They reported
normal color vision and normal or corrected-to-normal
visual acuity.
Apparatus and Stimuli
Stimulus presentation and response registration were
controlled by custom scripts written in Python run on
an HP Compaq 8000 Elite computer with a 22-in. liquid
crystal display color monitor (Samsung Syncmaster 2233,
1680 × 1050 pixels at 120 Hz). Participants were seated
in a sound-attenuated and dimly lit laboratory, at a viewing
distance of 75 cm. They were instructed to sit as relaxed as
possible to minimize muscle activity and other electro-
physiological noise to appear in the EEG signal.
The primary experimental display contained a visual
search array consisting of eight discrete stimuli with dif-
ferent shapes (one circle vs. seven unfilled diamonds, o
viceversa), with each shape containing a vertical or hor-
izontal gray line (0.3° × 1.5°) inside. These stimuli were
presented on an imaginary circle with a radius of 4°, cen-
tered at the fixation (a white cross measuring 1° × 1°),
against a black background (6 cd/m2). The circle’s radius
was 1°, the unfilled diamond was subtended 2° × 2°, y
each shape had a red or green outline (the color was cho-
sen from CIE Lab color space, centered at L = 70, a = 5,
b = 0, with a radius of 60, with an almost equal lumi-
nance range: 36–44 cd/m2).
Procedure and Design
A fixation cross appeared and remained visible through-
out the trial. Después 1350 mseg, the search array was pre-
sented for 2450 msec or until the participant responded.
Participants had to search for a circle (objetivo) entre
seven diamonds (distractors) or vice versa and indicate
whether the line segment inside the target was vertical
or horizontal by pressing the up or left key on the key-
board as fast as possible. Because the target singleton
could either be a diamond between circles or a circle
between diamonds, observers most likely adopted a
“singleton detection mode,” which makes it impossible
to selectively attend one specific feature (see Bacon &
Egeth, 1994). The intertrial interval was between 0 y
350 msec at random.
Experimento 1: With High-Probability Location
The target was presented in each trial, and it was equally
likely to be a circle or a diamond. A uniquely colored dis-
tractor singleton appeared in 66% of the trials, con el
same shape as other distractors but a different color (rojo
or green with an equal probability). All conditions were
randomized within blocks. Those display elements were
presented at eight locations from the imaginary ring with
a 4° radius. The target and distractor singleton were only
presented along the vertical or horizontal axis. El
remaining four locations served as neutral display ele-
mentos. Para cada participante, one of the horizontal loca-
tions contained a distractor singleton in 55% of trials
(high-probability location); for three other locations (en
the cardinal axis), the probability of containing a distractor
singleton was 15% cada (low-probability location). El
high-probability location was counterbalanced across
Participantes. In the distractor singleton present condition,
1536
Revista de neurociencia cognitiva
Volumen 31, Número 10
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
oh
C
norte
_
a
_
0
1
4
3
3
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
the target appeared equally often at each possible loca-
ción, except for the high-probability location.1 In the no-
distractor condition, the target appeared equally often at
each possible location.
Participants completed 40 practice trials and 13 bloques,
each containing 120 ensayos. Warning messages were pre-
sented if they did not respond or pressed the wrong
key. Depending on the singleton distractors and target
ubicación, a total of five conditions were analyzed:
(1) the distractor singleton was presented at the high-
probability location while the target was presented verti-
cally; (2) the target was presented at high-probability
location while the distractor singleton was either absent
or presented vertically; (3) the distractor singleton was
either absent or presented vertically, while the target
was presented vertically; (4) the distractor singleton
was presented along the horizontal axis at the lateral
low-probability location, while the target was presented
vertically; y (5) the distractor singleton was either
absent or presented vertically, while the target was
presented along the horizontal axis at the lateral low-
probability location.
Experimento 2: Without High-Probability Location
Experimento 2 served as a baseline experiment to compare it
with the results acquired in Experiment 1. In Experiment 2,
we used the same design as in Experiment 1 except that all
four potential target and singleton distractor locations were
equiprobable. Participants completed 40 practice trials and
nine blocks, each containing 120 ensayos. As there was no
high-probability location in Experiment 2, only three condi-
tions were analyzed: (1) the distractor singleton was pre-
sented laterally, while the target was presented vertically;
(2) the target was presented laterally when the distractor
singleton was absent or presented vertically; y (3)
the distractor singleton was either absent or presented
vertically, while the target was also presented vertically.
Tenga en cuenta que, in both experiments, both target and distractor
singleton were salient objects.
Data Recording and Preprocessing
EEG data were collected using a BioSemi ActiveTwo sys-
tema (www.biosemi.com) de 64 electrodes, placed ac-
cording to the extended 10–20 system and digitized at
512 Hz. Además, to record eye movements, the ver-
tical EOG was recorded from electrodes located 2 cm
above and below the right eye, and the horizontal EOG
was recorded from electrodes 1 cm lateral to the external
canthi.
EEG data were re-referenced to the mean of the left
and right earlobes and high-pass filtered using a cutoff
de 1.5 Hz (for independent component analysis [ICA]
solo) y 0.1 Hz (for final analyses). High-pass filtering
was done with default settings of the pop_basicfilter
function of ERPLAB (Lopez-Calderon & Luck, 2014),
using a Butterworth IIR filter. Continuous EEG was first
epoched from –3000 to 3500 msec relative to search dis-
play onset (to avoid edge artifacts resulting from wavelet
convolution; see below). Trial rejection and ICA were
done on smaller epochs (–1500 to 1500 mseg) to avoid
overlapping time points across epochs. para mejorar
ICA, epochs were baseline-normalized using the whole
epoch as baseline (Groppe, Makeig, & Kutas, 2009).
Malfunctioning electrodes were visually detected and
temporarily removed from the data. An adapted version
of an automatic trial rejection procedure, as imple-
mented in the Fieldtrip toolbox (Oostenveld, Fries,
Maris, & Schoffelen, 2011), was used to detect EMG arti-
hechos. Específicamente, we used a 110–140 Hz bandpass filter
to capture muscle activity and allowed for variable z-score
cutoffs per participant based on the within-subject vari-
ance of z scores. After trial rejection, ICA, as implemented
in the EEGLAB toolbox (Delorme & Makeig, 2004), era
performed on the clean electrodes only. Together with
the vertical EOG and horizontal EOG signals, we visually
inspected and removed ICA components that captured
eye blinks, eye movement, or other artifacts that were
clearly not brain-driven activity. Afterwards, we interpo-
lated the malfunctioning electrodes identified earlier
using spherical spline interpolation as implemented in
EEGLAB’s eeg_interp.m function. Trials with an incorrect
(or no) response were also excluded from further analy-
ses. All steps together left 87.4% (±6.0% across partici-
pants) trials in Experiment 1 y 73.5% (±3.4% across
Participantes) trials in Experiment 2.
Time–Frequency Analysis
Time–Frequency Decomposition: Laplacian
Before time–frequency decomposition, we estimated the
surface Laplacian of the EEG data (Perrin, Pernier,
Bertrand, & Echallier, 1989), which is equivalent to the
current source density approach. This method acts as a
spatial high-pass filter by accentuating local effects while
filtering out distant effects due to volume conduction;
de este modo, it sharpens the EEG topography (Kayser & Tenke,
2015; cohen, 2014). For estimating the surface Laplacian,
we used a 10th-order Legendre polynomial, and λ was
set at 10−5.
Morlet Wavelet Convolution
The epoched EEG time series were decomposed into
their time–frequency representations using custom-
written MATLAB scripts (github.com/joramvd/tfdecomp).
The time series were convolved with a set of Morlet wave-
lets with frequencies ranging from 1 a 40 Hz in 25 loga-
rithmically spaced steps. The complex wavelets were
created by multiplying sine waves (ei2πft, where i is the
complex operator, f is frequency, and t is time) con un
−t^2/2s^2, where s is the width of the Gaussian).
Gaussian (mi
Wang et al.
1537
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
oh
C
norte
_
a
_
0
1
4
3
3
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The width of the Gaussian was set as s = δ/2πf, where δ
represents the number of cycles of each wavelet, logarith-
mically spaced between 3 y 12 to have a good trade-off
between temporal and frequency precision. Convolution
was applied in the frequency domain: The fast Fourier
transform was applied to both the EEG data and the
Morlet wavelets, and these were multiplied in the fre-
quency domain, after which the inverse fast Fourier trans-
form was applied. The frequency-specific power at each
time point was defined as the squared magnitude of the
complex signal resulting from the convolution (real(Zt)2 +
imagen(Zt)2). Raw power was transformed into one com-
mon scale across frequencies using a within-subject per-
mutation testing procedure (see below).
Frequency Band and Time Window of Interest
Based on previous studies, we selected PO3/4, PO7/8, y
O1/2 for the analysis (de Vries, van Driel, & Olivers, 2017;
van Driel, Gunseli, Meeter, & Olivers, 2017). We were spe-
cifically interested in the lower (7.5–10 Hz) and upper
(10–12.5 Hz) alpha frequency ranges (see Klimesch,
1996, 1999, 2012, for reviews) to track how attention is
guided by implicit regularities before the search display
onset, in a time window comprising the intertrial interval
(–1500 to 0 mseg). Además, because participants could
only anticipate the high-probability distractor location and
not the target location nor the actual distractor location,
we combined all five conditions together in both experi-
ments for the prestimulus alpha-band analysis.
Z Transformation of Lateralized Power
Raw power can only take positive values and has a 1/f
power scaling, which requires a transformation such as
dB conversion or percent signal change (cohen, 2014).
Such relative metrics, Sucesivamente, require a baseline time
range, for which typically a prestimulus interval is chosen.
Sin embargo, for this analysis, we were specifically interested
in a between-group comparison of condition average
lateralized (contra- vs. ipsilateral to anticipated distractor
ubicación) alpha power during this prestimulus interval.
We thus Z-transformed time–frequency power through
a within-subject statistical procedure. For each contra-
and ipsilateral channel pair (thus excluding midline elec-
trodes) and each time–frequency point, we first computed
the trial average difference scores of contra- minus ipsi-
lateral power. Próximo, at each of 1000 permutations, nosotros
swapped for a random subset of trials the order of this
comparación (es decir., ipsilateral minus contralateral) and com-
puted the trial average across all trials. This gave a distri-
bution of “lateralized” power values under the null
hypothesis of no difference between contra- and ipsilateral
canales. The observed lateralized power values were
then statistically transformed with respect to the mean
and standard deviation of this distribution, by computing
their Z values. This procedure yields a metric that shows
interpretable dynamics over time as well as frequency.
Because there was no anticipated distractor location in
the baseline experiment (Experimento 2), we labeled one
lateral side as “contralateral” and the other side as “ipsilat-
eral” consistently for all participants and accordingly com-
puted the same Z transformation for time–frequency
power as was done for the real testing (Experimento 1) con
predictable lateralized distractors.
Estadísticas
A nivel de grupo, time–frequency maps of power of the
selected electrode groups were first statistically tested
against baseline within each experimental group, usando
a permutation-based approach with cluster size correc-
ción. Sin embargo, with permutation testing, the more points
are added to the analysis, the less sensitive it becomes. Si
it is known in advance that a specific frequency band is
likely to exhibit the effect of interest, then one should in-
corporate this prior knowledge and limit the statistical
test to this certain band (Oostenveld et al., 2011; Maris
& Oostenveld, 2007). Our a priori hypothesis pertained
specifically to preonset alpha band enhancement. De este modo,
in addition to a group-specific descriptive test over all
time–frequency points, we further narrowed our hypoth-
esis test of a group difference to the alpha-band only.
We ran two types of cluster-corrected permutation
pruebas: (1) Within each group, we used one-sample t tests
across time and frequency; at each test, the sign of the
power values was shuffled across participants in 2000
permutations to yield a null distribution against which
observed clusters of summed t values were thresholded
using p < .05; (2) Between groups, we used indepen-
dent-samples t tests, comparing the two groups across
time and frequency, and on alpha-band average power
over time. Here, we shuffled participants across experi-
ment in 2000 permutations for the null distribution.
ERP Analysis
The ERP analysis was done on a prespecified time window
from −100 to 900 msec relative to display onset. To
obtain ERPs, single-trial EEG data were first baseline-
corrected using a prestimulus baseline period from
−100 to 0 msec, after which data were trial-averaged.
Experiment 1: With High-Probability Location
For each participant, mean contralateral and ipsilateral activ-
ity over conditions relative to the high-probability location
was calculated for electrodes PO7/8. To determine the time
window used for further statistical analyses of different ERP
components, the lateralized ERP (contralateral minus ipsi-
lateral to the high-probability location) was calculated,
showing lateralized activity due to the high-probability
distractor location. The time window was also selected re-
ferred to previous studies (Feldmann-Wüstefeld & Vogel,
1538
Journal of Cognitive Neuroscience
Volume 31, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
2019; Hickey et al., 2008; Hickey, McDonald, & Theeuwes,
2006; Eimer, 1996). Thus, the early PD epoch was deter-
mined as ±20 msec from the most positive peak be-
tween 50 and 150 msec. The PD epoch was determined
as ±20 msec from the most positive peak between 150 and
250 msec. When the target was presented at the lateral
low-probability location, the target N2pc epoch was deter-
mined as ±20 msec from the most positive peak be-
tween 150 and 250 msec (note that the positive peak
was used, because the lateralized ERP was defined
with respect to the high-probability distractor location).
When the target was presented at the high-probability
location, the target–N2pc epoch was determined as the
most negative peak between 250 and 350 msec, because
we assumed a delayed target selection due to the sup-
pression on the high-probability location. The mean am-
plitude for these time windows was calculated separately
for laterality (contra vs. ipsi), each condition, and each
participant.
Experiment 2: Without High-Probability Location
For each participant, mean contralateral and ipsilateral activ-
ity over conditions relative to the target or distractor single-
ton was calculated for electrodes PO7/8. To determine the
time window used for further statistical analyses of different
ERP components, both the lateralized ERP (contralateral
minus ipsilateral) for trials with lateral target and for trials
with lateral distractor singleton were collapsed, showing
lateralized activity due to targets or distractor singletons.
The target–N2pc epoch was determined as ±15 msec from
the most negative peak between 200 and 300 msec in the
lateral targets waveform. The mean amplitude for these time
windows was calculated separately for laterality (contra vs.
ipsi), each condition, and each participant.
RESULTS
Behavior
Consistent with Wang and Theeuwes (2018a, 2018b),
distractors at the high-probability location interfered less
(828 msec) than distractors at the low-probability loca-
tion (865 msec), t(17) = 3.42, p = .003. Compared with
the no-distractor condition (777 msec), there were signif-
icant attentional capture effects for a distractor present at
the high-probability location, t(17) = 4.26, p = .001, and
the low-probability location, t(17) = 8.61, p < .001. Also,
consistent with earlier studies ( Wang & Theeuwes,
2018a, 2018b, 2018c), target selection was less efficient
when the target appeared at the high-probability distrac-
tor location (808 msec) relative to the low-probability
distractor location (767 msec), t(17) = 3.73, p = .002.
To show that these effects were due to the predictability
of the high-probability location, we also ran a baseline exper-
iment (Experiment 2) in which all locations were equally
likely to contain a salient distractor singleton. Consistent
with earlier studies (Theeuwes, 1991, 1992), this showed a
classic attentional capture effect, with slower RTs when the
distractor singleton was present (850 msec) compared with
when it was absent (773 msec), t(17) = 10.59, p < .001. See
Appendix for more details.
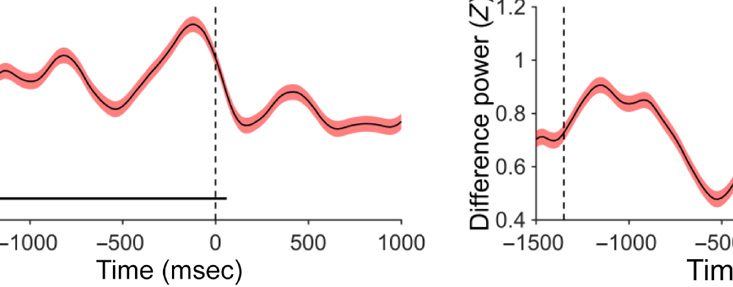
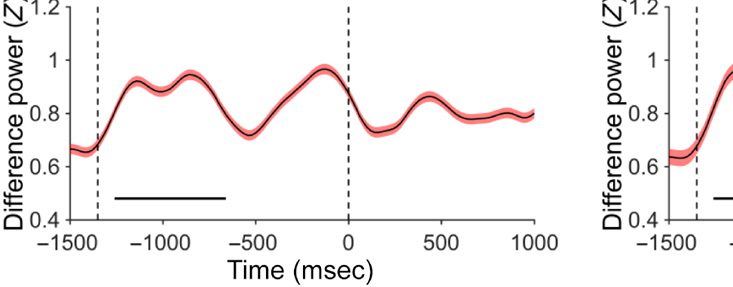
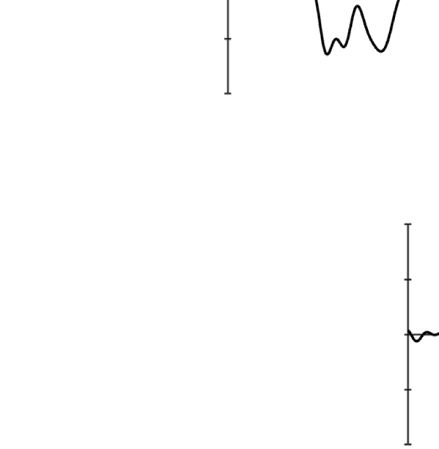
Prestimulus: Local Alpha Power
As the measure of anticipation, we chose lateralized
alpha power (following Klimesch, 1996). We analyzed
the oscillation in the alpha band (7.5–12.5 Hz) for elec-
trodes PO3/4, PO7/8, and O1/2 (see Methods section
for oscillation band and electrode selection) and found
a lateralized effect consisting of a stronger enhancement
in power for contra- versus ipsilateral relative to the high-
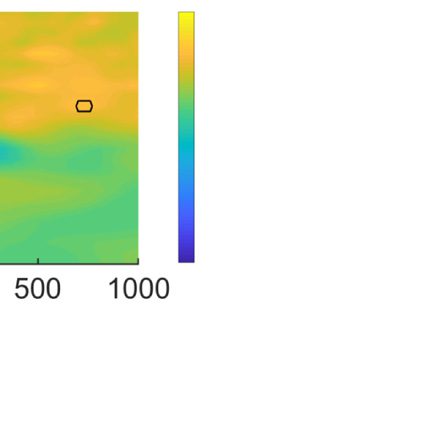
probability location in Experiment 1 (see Figure 1A).
Importantly, there was no lateralized enhancement in
Experiment 2 (see Figure 1B). A permutation test across
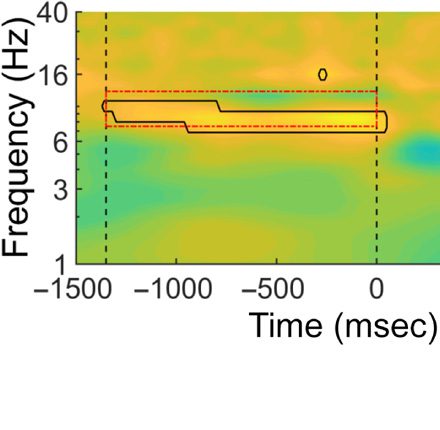
time and frequency comparing Experiments 1 and 2
confirmed this finding, showing that the significant differ-
ence ( p < .05, uncorrected) between groups was mainly
seen in the alpha band (see Figure 1C, upper panels for
an illustration). Importantly, a cluster-based permutation
test on the alpha-band comparing Experiments 1 and 2
revealed a reliable difference between the two groups
in the −1220 and −660 msec interval prior to display
onset ( p < .05; see Figure 1C, lower left panel for an
illustration). Also, even without using the cluster-based
permutation test, when comparing the average increased
alpha power between the two groups, there was a reli-
able difference before display onset, t = 2.37, p = .03.
We interpret these results as evidence for anticipatory
suppression of the high-probability location.
Previous research has shown that lower alpha band
(7.5–10 Hz) is more related to short-lasting expectation
and attentional processing, whereas the upper alpha
band (10–12.5 Hz) is more related to semantic memory
processing (see Klimesch, 1996, 1999, 2012, for reviews).
Given these findings, we divided the alpha band in two,
in order to explore whether the anticipatory suppression
effect related to the high-probability location would in-
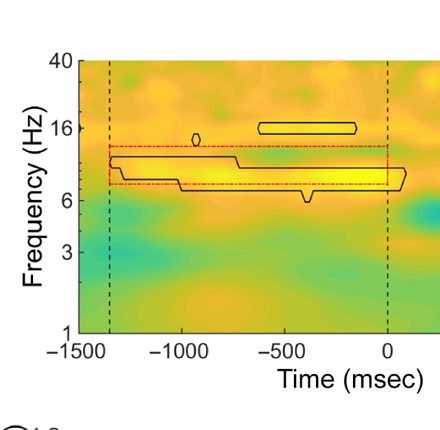
deed only occur in the lower alpha band. For the lower
alpha band, a cluster-based permutation test between ex-
periments showed that the power enhancement was sig-
nificant from −1260 to 60 msec relative to display onset
( p < .05; see Figure 1C, middle panel). No effects were
observed for the upper alpha band (see Figure 1C, right
panel).
In short, current results suggest that observers start sup-
pressing the high-probability location about ∼1200 msec
before display onset. Moreover, this prestimulus sup-
pression (i.e., implicit expectation that a distractor would
appear on the high-probability location) is mainly reflected
in the lower alpha band, which is consistent with previous
studies (see Klimesch, 2012, for a review).
Wang et al.
1539
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
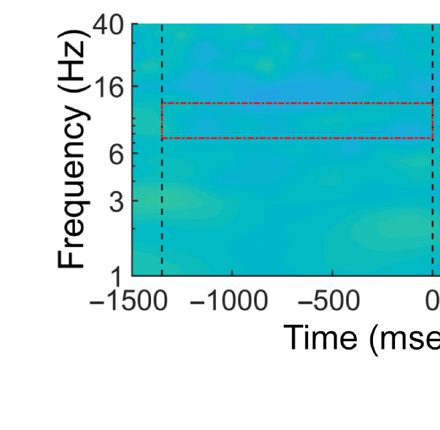
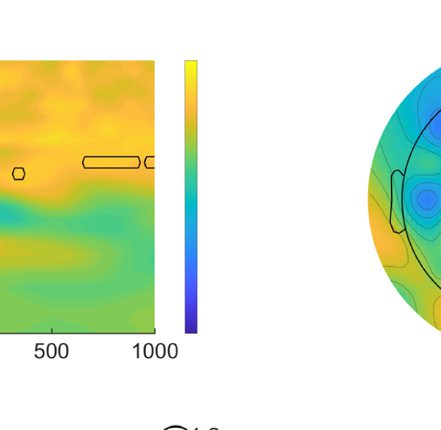
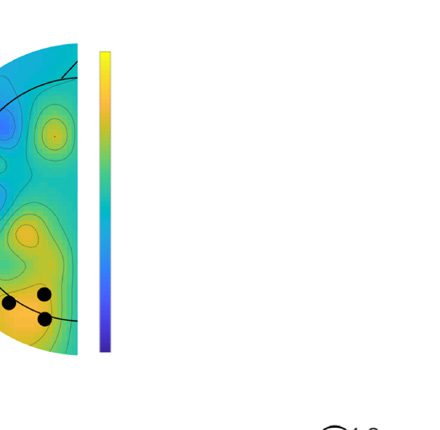
Figure 1. (A) Lateralized posterior alpha power in Experiment 1 with high-probability location, (B) lateralized posterior alpha power in Experiment 2 as a
baseline, and (C) their comparison results. A and B show time–frequency plots of contralateral minus ipsilateral power at the average of O1/2, PO3/4,
and PO7/8 in different experiments (red dashed outline indicates the alpha band oscillation between stable fixation and search display onset, and black
solid outline indicates [uncorrected] significant power difference between contralateral and ipsilateral in grand average at p < .05). In C, upper left
panel shows time–frequency plots of Experiment 1 minus Experiment 2 (baseline) power at the average of O1/2, PO3/4, and PO7/8; upper right panel
shows half topographies, grand-averaged scalp distributions of alpha power at ipsilateral electrodes subtracted from those at contralateral electrodes,
collapsed across hemispheres, and power is averaged over the alpha band oscillation between stable fixation and search display onset; lower panels present
time series of contralateral minus ipsilateral alpha power difference between Experiments 1 and 2 for overall alpha band, lower alpha band, and upper
alpha band (black horizontal line represents time points with a significant difference between experiments after cluster-size thresholding at p < .05).
The red shadow area indicates means ± SEM of difference values.
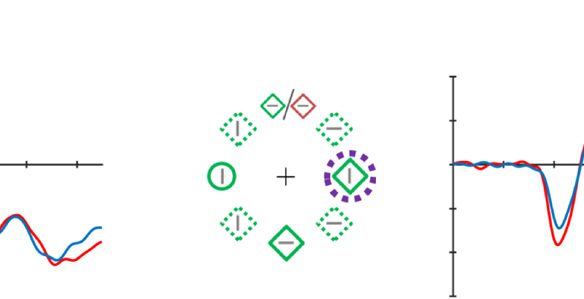
Poststimulus: PD
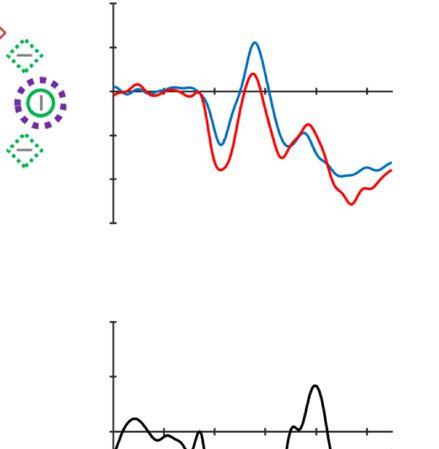
To provide converging evidence that the high-probability
location is suppressed, we first examined whether a PD
was elicited when the distractor singleton was presented
at the high- versus the low-probability location (see
Hickey et al., 2008). Consistent with the behavioral data
showing reduced capture, we observed a PD when the
distractor was presented in the high-probability location
and not when it was presented in the low-probability
location. A two-way ANOVA with within-subject factors
Laterality (contra- vs. ipsilateral to the distractor single-
ton) and Distractor location (high vs. low probability)
was conducted on the average ERP amplitude within the
PD time window (186–226 msec; see Methods section for
choice of time window). The results showed a main effect
for Laterality, F(1, 17) = 9.51, p = .007, ηp
2 = .36, but not for
Distractor location, F(1, 17) = 2.92, p = .105, ηp
2 = .15.
However, importantly, the interaction was reliable, F(1,
17) = 4.98, p = .039, ηp
2 = .23. Planned comparisons trac-
ing the source of the interaction indeed showed a reliable
PD only for high-probability locations, t(17) = 3.15, p =
.006 (see Figure 2, upper middle and lower left
panels).2
One possible reason for not observing a PD for distrac-
tors presented at the low-probability location might be
that there are opposing lateralized suppression effects
working at the same time (i.e., the spatial suppression of
the high-probability location and the feature suppression
of the distractor singleton at the opposing low-probability
location). If the PD reflecting the suppression on high-
probability location is larger than the PD reflecting the sup-
pression on the distractor at the low-probability location,
then it is possible that this larger PD overshadows the
smaller distractor-generated PD. Note, however, it is also
possible that the distractor at the low-probability location
1540
Journal of Cognitive Neuroscience
Volume 31, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
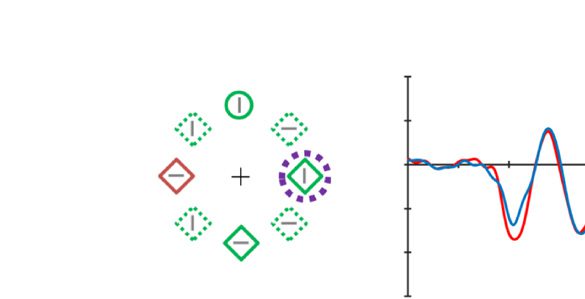
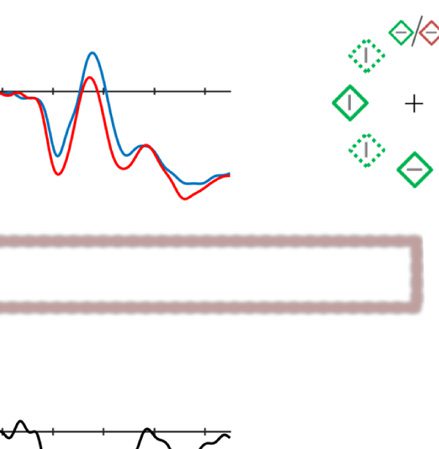
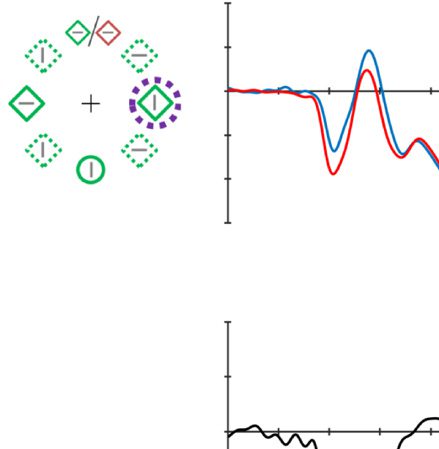
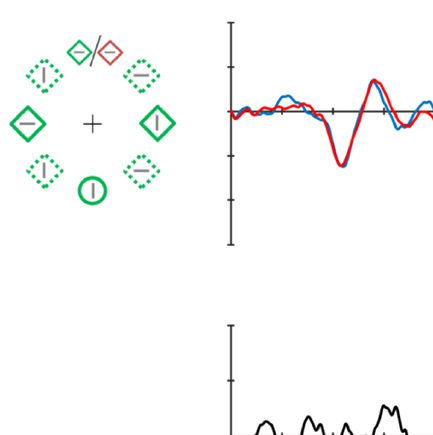
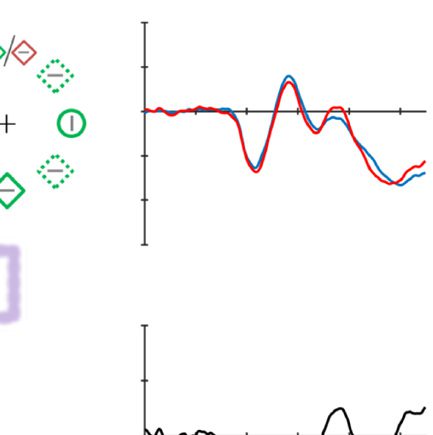
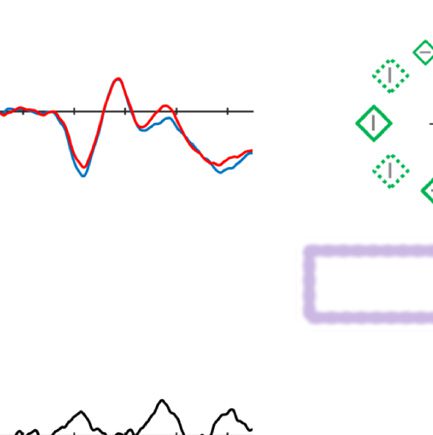
Figure 2. Examples of the search displays in various conditions of Experiment 1. The dashed circles indicate the high-probability location. The
display elements with solid outline indicate possible target and distractor singleton locations; the display elements with dashed outline indicate
locations that never contained a target or distractor singleton. Each subpanel shows, per condition, a grand-averaged ERP recorded at PO7/PO8
and the lateralized difference waves (contra- minus ipsilateral). Green, red, and blue dashed squares indicate the time window that was used to
calculate mean amplitude of early PD, PD, and N2pc, respectively. Note that, in the lower right panel, the ERP waves were plotted contralateral to the
high-probability location, but the target was presented at the low-probability location. Thus, the N2pc was reversed here, suggesting a selection
of the target. All waveforms were low-pass filtered at 30 Hz (half power cutoff, 24 dB) for visualization.
does not generate a PD at all, as the distractor at this loca-
tion generates a strong capture effect.
Moreover, if the suppression is both anticipatory- and
location-based and not elicited by the presence of spe-
cific distractor information, we would expect a PD for
the high-probability location even if no distractor single-
ton is presented there. We analyzed this component
separately for when the target appeared at this location
(upper right panel of Figure 2), for when the distractor
singleton appeared at this location (upper middle panel
of Figure 2), and for when a neutral display element was
presented at this location (no target nor distractor single-
ton, upper left panel of Figure 2). A two-way ANOVA with
within-subject factors Laterality (contra- vs. ipsilateral to
the high-probability location) and Type of display ele-
ment (distractor, target, or neutral display element) was
conducted on average ERP amplitude within PD time win-
dow. The results showed a main effect for Laterality, F(1,
17) = 7.96, p = .012, ηp
2 = .32, but not for Type of display
element, F(2, 34) = 2.15, p = .13, ηp
2 = .11, and no inter-
action was observed, F(2, 34) = 1.93, p = .161, ηp
2 = .1.
The activity was more positive in the contralateral side for
each condition, all ts ≥ 2.46, all ps ≤ .025, indicating that a
PD was found regardless of which display element was pre-
sented there. This suggests that the suppression we ob-
served is completely spatial in origin. Note, however, as
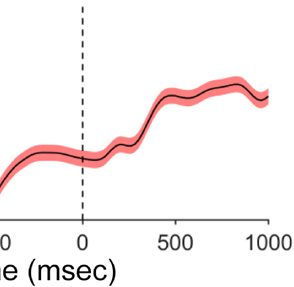
can be seen in Figure 3, the mean PD amplitude generated
when a distractor was present at the high-probability loca-
tion (1.58 μV) was slightly larger than the amplitude when
a neutral display element presented at high-probability lo-
cation (1.09 μV), t = 2.14, p = .047. This suggests that the
presence of distracting information at this high-probability
location generates additional suppression above and be-
yond suppression that is already present when no dis-
tracting information is present. There was no reliable
difference when the target (1.43 μV) and the neutral dis-
play element were presented at high-probability location,
t = 1.2, p = .248.
Wang et al.
1541
presented at the low-probability location (with the average
peak latency of 205 msec) were significantly different,
t(17) = 7.89, p < .001.
It is clear that the N2pc for targets presented at the
high-probability location is weaker than in the low-proba-
bility location. If we use the same time period (±20 msec)
for calculating the N2pc, as was used for targets presented
at the low-probability location, then there N2pc is no
longer reliable. This is probably due to the suppression
(indexed by the PD) exerted at the high-probability loca-
tion, which makes the N2pc less robust. Therefore, we
used the peak point to calculate the mean N2pc ampli-
tude for targets presented at the high-probability location.
When we use this method, a reliable yet less robust N2pc
was observed, as we have outlined above.
Poststimulus: Early PD
Consistent with the notion of anticipatory suppression of
the high-probability location, we found evidence for an
early PD representing an early modulation of the high-
probability location in Experiment 1. A two-way ANOVA
with within-subject factors Laterality (contra- vs. ipsilateral
to the high-probability location) and Type of display ele-
ment (distractor, target, or neutral display element) was
conducted on average ERP amplitude within the early
PD time window (74–114 msec; see Methods section for
choice of time window). The results indicated a main
significant effect for Laterality only, F(1, 17) = 4.94, p =
.04, ηp
2 = .23, and no interaction across conditions (see
Figure 2), suggesting that the early PD was present for
the high-probability location regardless of whether a
target, a distractor, or a neutral display element was
presented.
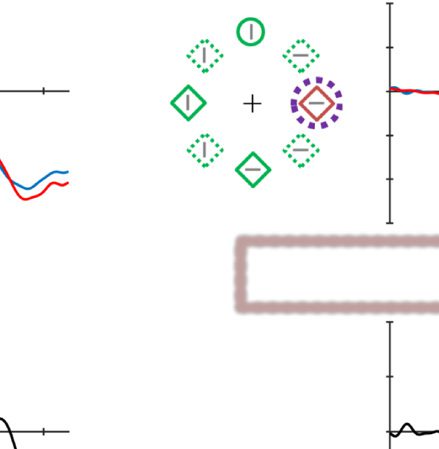
ERP Results in Experiment 2 (Baseline)
In the baseline experiment, the target and distractor single-
ton were equally likely presented at each location. Based
on previous studies (Hickey et al., 2006), we expected an
N2pc component for both the target and distractor single-
ton. A two-way ANOVA with within-subject factors Laterality
(contra- vs. ipsilateral to the singleton) and Lateralized item
(target vs. distractor) was conducted. The results showed a
significant main effect for Laterality, F(1, 17) = 5.01, p =
.039, ηp
2 = .23 (see Figure 4), but no interaction suggesting
that both target and distractor singletons generated an
N2pc (Hickey et al., 2006). There was indeed a significant
N2pc for targets, t(17) = 2.17, p = .045, and a trend in the
same direction for distractors, t(17) = 1.75, p = .099. The
same time window was chosen to analyze when a neutral
display element was presented laterally (i.e., like what we
did in Experiment 1). The results indicate that there was no
difference between contralateral and ipsilateral sides, t < 1.
It should be noted that, unlike in Experiment 1 (which had
the high-probability location), in the current baseline
experiment, no PD was observed in any condition.
Figure 3. Upper panel shows mean PD amplitude for different items
presented at high-probability location. Lower panel shows mean N2pc
amplitude for target presented at high- and low-probability locations.
Error bars denote within-subject 95% confidence intervals.
Poststimulus: Target-elicited N2pc
To determine whether there was attentional selection of
the target and/or the distractor when presented at the
low-probability location, we analyzed the N2pc toward
the target or distractor singleton ( lower panels of
Figure 2). A two-way ANOVA with within-subject factors
Laterality (contra- vs. ipsilateral to the low-probability lo-
cation) and Type of display element (distractor vs. target)
was conducted on mean ERP amplitude within the N2pc
time window (186–226 msec; see Methods section for
choice of time window). Only the interaction was reli-
able, F(1, 17) = 7.5, p = .014, ηp
2 = .31. Subsequent
planned comparisons tracing the source of the inter-
action showed a reliable N2pc for targets only, t(17) =
2.35, p = .031 (see Figure 2, lower panels).
Furthermore, we also observed what seems to be a
delayed N2pc (peaked at 294 msec) when the target
was presented at high-probability location, t(17) =
1.88, p = .039 (one-tailed). This amplitude of the delayed
N2pc for the target at the high-probability location did
not differ from the amplitude of the N2pc for the target
at the low-probability location, t < 1 (see Figure 3). We
used a jackknife procedure (Kiesel, Miller, Jolicœur, &
Brisson, 2008) to statistically test the difference between
the N2pc latencies for targets presented at the high-
versus low-probability location and obtained a reliable
estimate of the numerical difference. The peak latencies
between the delayed N2pc (with the average peak latency
of 294 msec) for target presented at the high-probability
location and the N2pc observed when the target was
1542
Journal of Cognitive Neuroscience
Volume 31, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
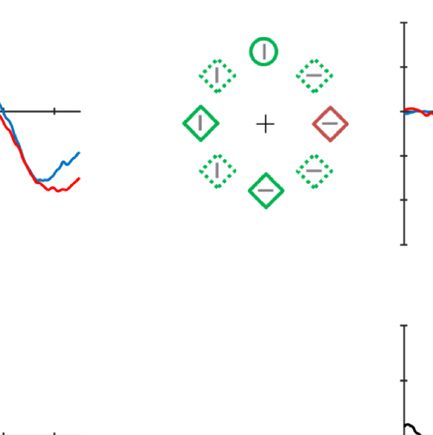
Figure 4. Example of the search displays in various conditions of Experiment 2. The dashed circles indicate the high-probability location. The
display elements with solid outline indicate possible target and distractor singleton locations; the display elements with dashed outline indicate
locations that never contained a target or distractor singleton. Grand-averaged ERPs recorded at PO7/PO8 and their difference waves (contra-
minus ipsilateral) for different conditions were presented in different subpanels. Blue dashed squares indicate the time window that was used to
calculate mean amplitude of N2pc. All waveforms were low-pass filtered at 30 Hz (half power cutoff, 24 dB) using digital filtering.
DISCUSSION
Our data show that the reduction of attentional capture
by a salient singleton is accompanied by prestimulus an-
ticipatory suppression. Because participants have learned
that the salient distractor is more likely to appear at one
specific location, this location is suppressed as there
is increased alpha power contralateral to this location
relative to the ipsilateral location. Notably, alpha-band
oscillations have been associated with neural inhibition
( Jensen & Mazaheri, 2010), serving as a general gating
mechanism. Previous studies have reported increased
alpha power at electrodes over ipsilateral visual cortex,
representing the location where participants expected
distractors to appear (Kelly, Lalor, Reilly, & Foxe,
2006). This was interpreted as evidence of active atten-
tional suppression. It has been claimed that prestimulus
alpha operates as a gating mechanism, allowing the sup-
pression of distracting information (Moore & Armstrong,
2003).
When the salient distractor was presented at the high-
probability location, we found a PD, suggesting spatial
suppression. Critically, following this PD, the attentional
capture by salient distractor was attenuated, as no N2pc
was observed toward that location. In the baseline exper-
iment, when all locations were presented equally likely,
there was no PD, and consequently, attention was cap-
tured by the salient distractor as evidenced by an N2pc,
consistent with Hickey et al. (2006). Notably a PD was
also found when the target was presented at the high-
probability location in Experiment 1, resulting in a delayed
selection (i.e., delayed N2pc) of the target consistent with
the behavioral data showing delayed RT when the target is
presented at that location.
Previous studies have shown that attentional suppres-
sion of distractors is typically accompanied by a PD wave-
form (Feldmann-Wüstefeld & Vogel, 2019; Cosman,
Lowe, Zinke, Woodman, & Schall, 2018; Gaspelin &
Luck, 2018a; Gaspar, Christie, Prime, Jolicœur, &
McDonald, 2016; Gaspar & McDonald, 2014; Feldmann-
Wüstefeld, Wykowska, & Schubö, 2013; Sawaki & Luck,
2013; Hickey et al., 2008). Observing a PD is attributed
to the processing of distracting information and is as-
sumed to reflect active suppression of this distractor infor-
mation (van Moorselaar & Slagter, 2019; Gaspelin & Luck,
2018b; Burra & Kerzel, 2014; Gaspar & McDonald, 2014;
Sawaki & Luck, 2013). For example, in a visual search
study, Gaspelin and Luck (2018a) showed that, when the
salient distractor was presented, it generated a PD, which
was correlated with behavior that showed suppression.
Gaspar et al. (2016) showed that a salient distractor
(e.g., a red item between green background items) gener-
ated a PD whereas a relatively low salient distractor (e.g.,
an orange item between red background items) did not
result in a PD. Overall these findings indicate that a PD is
only observed if a salient distractor is present and if its
salience signal needs to be actively suppressed. The find-
ings of the current study are dramatically different in this
respect. Consistent with previous finding, we do find a
PD when a salient distractor was present at the high-
probability location (Figure 2, upper middle panel); yet,
unlike previous studies, we also find a PD when neutral
(background) elements were present (Figure 2, upper left
panel) and even when the target was present at the high-
probability location (Figure 2, upper right panel). In the
latter case, the suppression signified by the PD resulted
in a delayed N2pc and a delayed behavioral response to
the target. It is possible that the suppression that we
Wang et al.
1543
report here is different from the suppression that is ob-
served when a salient distractor is present.
The type of suppression reported here can be labeled
as “proactive suppression,” in the sense that specific lo-
cations within the spatial priority map are suppressed
(Ferrante et al., 2018; Wang & Theeuwes, 2018a, 2018b,
2018c). This suppression is spatial and feature-blind, and
happens regardless of what information is present at that
location (whether it is a target, a salient distractor, or a
neutral element). Consequently, also the neural correlates
of such suppression (contralateral alpha enhancement,
early PD, and PD) are observed in all these conditions.
Critically, and consistent with the notion of proactive sup-
pression, the enhanced alpha power contralateral to the
high-probability location is observed prior to display onset.
This indicates that the suppression is indeed proactive in
the sense that the high-probability location is already
suppressed before display onset. As argued, increased
alpha-band oscillations are assumed to represent neural
inhibition ( Jensen & Mazaheri, 2010), serving as a general
attentional gating mechanism.
Also, there is another suppression that is considered to be
reactive in the sense that attention is first captured by the sa-
lient distractor (even for the briefest moment) and then is
immediately suppressed (Won, Kosoyan, & Geng, 2019).
Depending on the task design, it is possible that the
disengagement of attention can be so fast that there are
virtually very little (or no) effects of the presence of the salient
distractor (i.e., basically no attentional capture). It is similar to
Moher and Egeth’s (2012) conception of what has been
labeled as the “search and destroy” hypothesis. Moher and
Egeth showed that participants that were instructed to
inhibit a particular feature could only do so after attending
to the location of the to-be-ignored feature.
It should be noted that the PD that we observed when a
distractor singleton was present at the high-probability
location was slightly larger than the PD that we found for
neutral elements presented at the high-probability loca-
tion. This may suggest that the presence of distracting in-
formation generates additional (retroactive) suppression
above and beyond the proactive suppression that was
induced through statistical learning.
We also observed an early PD component (80–120 msec
poststimulus) toward the high-probability location in all
conditions regardless of whether a target, a salient dis-
tractor, or a neutral element was presented at the high-
probability location. An early PD (also termed “positivity
posterior contralateral” or “lateralized P1”) often co-occurs
with the later PD in displays with lateralized stimuli.
Initially, Luck and colleagues (Luck & Kappenman, 2012;
Luck, Heinze, Mangun, & Hillyard, 1990) found that the
early PD (named “lateralized P1” in their study) was en-
hanced when the target appeared at the attended hemi-
field, suggesting that it may represent facilitation of early
sensory processing. This is consistent with some studies
suggesting that the early PD is related to initial processing
of stimuli and/or is assumed to represent some low-level
sensory imbalance between the two hemispheres. (Jannati,
Gaspar, & McDonald, 2013; Fortier-Gauthier, Moffat,
Dell’Acqua, McDonald, & Jolicœur, 2012). However, in the
current study, the early PD was also found when nonsalient
neutral display elements were presented at the high-
probability location, indicating that the early PD does
not represent early sensory processing of salience nor
hemispheric imbalance but instead represents a signature
of early suppression ( Weaver, van Zoest, & Hickey, 2017).
The current study shows that participants learn to sup-
press the location where the salient distractor is likely to
appear. The pattern of results shows that relative to the
high-probability location, there is prestimulus, contralat-
eral alpha enhancement, an early PD, and the PD. The
neural correlates were found regardless of whether a sa-
lient distractor, a salient target, or a neutral element were
presented at this location. These findings suggest that
the location that is likely to contain a salient distractor
is proactively suppressed through statistical learning.
APPENDIX
Behavior Results in Experiment 1
Trials on which the RTs were faster than 200 msec and slower
than 2000 msec (2.17%) were excluded from analyses.
Attentional Capture Effect
Mean RTs and mean error rates are presented in Figure A1A.
With Distractor condition (high-probability location, low-
probability location, and no distractor) as a factor, a
repeated-measures ANOVA on mean RTs showed a main
effect, F(2, 34) = 32.09, p < .001, partial η2 = .65.
Subsequent planned comparisons showed that there
were significant attentional capture effects for a distractor
presented at the high-probability location, t(17) = 4.26,
p = .001, and the low-probability location, t(17) = 8.61,
p < .001. Crucially, however, there was also a reliable dif-
ference between the high- and low-probability locations,
t(17) = 3.42, p = .003, suggesting that attentional cap-
ture effect was reduced for trials in which the distractor
appeared at a high-probability location.
The results on error rates mimicked those for RTs. With
Distractor condition (high-probability location, low-
probability location, and no-distractor) as a factor, a repeated-
measures ANOVA showed a main effect, F(2, 34) =
21.41, p < .001, partial η2 = .56. Subsequent planned
comparisons showed that significant attentional capture
effects were observed when a distractor singleton was pre-
sented at the high-probability location, t(17) = 3.82, p =
.001, and the low-probability location, t(17) = 5.74, p <
.001. Importantly, however, there was also a reliable dif-
ference between the high- and low-probability locations,
t(17) = 3.35, p = .004. Taken together, these results
suggest that attentional capture effect was reduced for
trials in which the distractor singleton appeared in a
1544
Journal of Cognitive Neuroscience
Volume 31, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure A1. The mean RTs (left)
and the mean error rates (right)
in (A) singleton distractor
present condition and (B)
singleton distractor absent
(no-distractor) condition. Error
bars denote within-subject
95% confidence intervals.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
high-probability relative to the low-probability location, a
straightforward replication of Wang and Theeuwes
(2018a, 2018b).
Target at the High-probability Distractor Location
To determine whether not only attentional capture but
also the efficiency of selecting the target was affected,
we examined RTs in the no-distractor condition. We ex-
pected that, even when the distractor was absent, observers
would respond more slowly toward a target when it was
presented at the high-probability distractor location relative
to the low-probability location. This is exactly what we
found (see the left panel of Figure A1B): Participants were
slower to respond when the target was presented at the
high-probability distractor location rather than at the
Figure A2. The spatial
distribution of attentional
capture effect by the means
of RTs (left) and error rates
(right). Here, Dist-0 refers to
the high-probability distractor
location, Dist-1 refers to the
low-probability distractor
location with one unit (90° polar
angle) away from the high-
probability distractor location,
and so on. Error bars denote
within-subject 95% confidence
intervals.
Wang et al.
1545
low-probability distractor location, t(17) = 3.73, p = .002,
and the response was less accurate when the target was
presented at the high-probability distractor location,
t(17) = 2.15, p = .046.
The Spatial Distribution of the Suppression Effect
We examined RTs and error rates of the target when the
distractor singleton appeared at different locations in re-
lation to the distance from the high-probability location.
If the effect has a spatial extent, the RT and the error rate
are expected to scale with distance from this area. Thus,
the distractor location was divided into three distances
(dist-0, dist-1, dist-2)3 from the high-probability location.
The mean RTs and mean error rates for these conditions
are presented in Figure A2A.
Repeated-measures ANOVAs on mean RTs and mean
error rates both showed significant main effects for
Distance, F(2, 34) = 11.57, p < .001, partial η2 = .41
and F(2, 34) = 7.15, p = .003, partial η2 = .3, respec-
tively. Moreover, we fitted the data with a linear function
and used their slopes to determine whether mean RTs
and mean error rates changed with distance. The slopes
were significantly larger than zero for mean RTs (15.4 msec
per display element), t(17) = 4.43, p < .001, and mean
error rates (0.8% per display element), t(17) = 3.73, p =
.002, suggesting a spatial gradient (the distractor effect
grew when the distractor was presented farther away
from the high-probability distractor location).
In the no-distractor condition, there was also a signifi-
cant main effect for Distance, F(2, 34) = 7.64, p = .002,
partial η2 = .31. The gradient effect was now reversed:
With increased distance, the selection of the target be-
came progressively more efficient, signified by a slope
of −11.15 msec per display element that differed from
zero, t(17) = 3.31, p = .004. This latter effect suggests
spatial suppression that is not dependent on whether
or not a distractor is present.
Behavior Results in Experiment 2
Trials on which the RTs were faster than 200 msec and
slower than 2000 msec (2.24%) were excluded from anal-
yses. Mean RTs and mean error rates are presented in
Figure A3. Paired-samples t test between different distrac-
tor conditions (with vs. without distractor singleton)
showed a significant capture effect, reflected by mean
RTs, t(17) = 10.59, p < .001, and mean error rates,
t(17) = 4.16, p = .001.
Figure A3. The mean RTs (left)
and the mean error rates (right)
in different distractor condition.
Error bars denote within-
subjects 95% confidence
intervals.
1546
Journal of Cognitive Neuroscience
Volume 31, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Acknowledgments
This research was supported by a China Scholarship Council
(CSC) scholarship (201508330313) to Benchi Wang.
Reprint requests should be sent to Benchi Wang, Department
of Experimental and Applied Psychology, Vrije Universiteit
Amsterdam, Van der Boechorststraat 1, 1081 BT Amsterdam,
The Netherlands, or via e-mail: wangbenchi.swift@gmail.com.
Notes
1. Because the target never appears at the high-probability lo-
cation in the distractor present condition, it is possible that the
effects reported are due to the target probability and not to the
distractor probability. However, in a recent study, we showed
that the probability of the target does not matter: the suppres-
sion effect observed here and in previous studies (Wang &
Theeuwes, 2018a, 2018b, 2018c) was solely due to the distrac-
tor being presented more often in this one location (Failing
et al., 2019).
2. When manually removing all the eye movement trials in
Experiment 1, the results stay the same.
3. The dist-0 location refers to the high-probability distractor
location; the dist-1 location means that the distractor was pre-
sented next to the high-probability distractor location (90° polar
angle); the dist-2 location refers to 180° from the high-probability
distractor location.
REFERENCES
Awh, E., Belopolsky, A. V., & Theeuwes, J. (2012). Top–down
versus bottom–up attentional control: A failed theoretical
dichotomy. Trends in Cognitive Sciences, 16, 437–443.
Failing, M., & Theeuwes, J. (2018). Selection history: How
reward modulates selectivity of visual attention.
Psychonomic Bulletin & Review, 25, 514–538.
Failing, M., Wang, B., & Theeuwes, J. (2019). Spatial suppression
due to statistical regularities is driven by distractor
suppression not by target activation. Attention, Perception,
& Psychophysics, 81, 1405–1414.
Feldmann-Wüstefeld, T., Uengoer, M., & Schubö, A. (2015).
You see what you have learned. Evidence for an interrelation
of associative learning and visual selective attention.
Psychophysiology, 52, 1483–1497.
Feldmann-Wüstefeld, T., & Vogel, E. K. (2019). Neural evidence
for the contribution of active suppression during working
memory filtering. Cerebral Cortex, 29, 529–543.
Feldmann-Wüstefeld, T., Wykowska, A., & Schubö, A. (2013).
Context heterogeneity has a sustained impact on attention
deployment: Behavioral and electrophysiological evidence.
Psychophysiology, 50(8), 722–733.
Ferrante, O., Patacca, A., Di Caro, V., Della Libera, C.,
Santandrea, E., & Chelazzi, L. (2018). Altering spatial
priority maps via statistical learning of target selection and
distractor filtering. Cortex, 102, 67–95.
Fortier-Gauthier, U., Moffat, N., Dell’Acqua, R., McDonald, J. J.,
& Jolicœur, P. (2012). Contralateral cortical organisation of
information in visual short-term memory: Evidence from
lateralized brain activity during retrieval. Neuropsychologia,
50, 1748–1758.
Gaspar, J. M., Christie, G. J., Prime, D. J., Jolicœur, P., &
McDonald, J. J. (2016). Inability to suppress salient
distractors predicts low visual working memory capacity.
Proceedings of the National Academy of Sciences, U.S.A.,
113, 3693–3698.
Gaspar, J. M., & McDonald, J. J. (2014). Suppression of salient
objects prevents distraction in visual search. Journal of
Neuroscience, 34, 5658–5666.
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven
Gaspelin, N., & Luck, S. J. (2018a). Combined
attentional capture. Perception & Psychophysics, 55,
485–496.
Burra, N., & Kerzel, D. (2014). The distractor positivity (Pd) signals
lowering of attentional priority: Evidence from event-related
potentials and individual differences. Psychophysiology, 51,
685–696.
electrophysiological and behavioral evidence for the
suppression of salient distractors. Journal of Cognitive
Neuroscience, 30, 1265–1280.
Gaspelin, N., & Luck, S. J. (2018b). The role of inhibition in
avoiding distraction by salient stimuli. Trends in Cognitive
Sciences, 22, 79–92.
Chun, M. M., & Jiang, Y. (1998). Contextual cueing: Implicit
Groppe, D. M., Makeig, S., & Kutas, M. (2009). Identifying
learning and memory of visual context guides spatial
attention. Cognitive Psychology, 36, 28–71.
Chun, M. M., & Jiang, Y. (1999). Top–down attentional
guidance based on implicit learning of visual covariation.
Psychological Science, 10, 360–365.
Cohen, M. X. (2014). Analyzing neural time series data:
Theory and practice. Cambridge, MA: MIT Press.
Cosman, J. D., Lowe, K. A., Zinke, W., Woodman, G. F., &
Schall, J. D. (2018). Prefrontal control of visual distraction.
Current Biology, 28, 414.e3–420.e3.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
de Vries, I. E., van Driel, J., & Olivers, C. N. (2017). Posterior
α EEG dynamics dissociate current from future goals in working
memory-guided visual search. Journal of Neuroscience, 37,
1591–1603.
Egeth, H. E., & Yantis, S. (1997). Visual attention: Control,
representation, and time course. Annual Review of
Psychology, 48, 269–297.
Eimer, M. (1996). The N2pc component as an indicator of
attentional selectivity. Electroencephalography and Clinical
Neurophysiology, 99, 225–234.
reliable independent components via split-half comparisons.
Neuroimage, 45, 1199–1211.
Hickey, C., Di Lollo, V., & McDonald, J. J. (2008).
Electrophysiological indices of target and distractor
processing in visual search. Journal of Cognitive
Neuroscience, 21, 760–775.
Hickey, C., McDonald, J. J., & Theeuwes, J. (2006).
Electrophysiological evidence of the capture of visual
attention. Journal of Cognitive Neuroscience, 18,
604–613.
Itti, L., & Koch, C. (2001). Computational modelling of visual
attention. Nature Reviews Neuroscience, 2, 194–203.
Jannati, A., Gaspar, J. M., & McDonald, J. J. (2013). Tracking
target and distractor processing in fixed-feature visual search:
Evidence from human electrophysiology. Journal of
Experimental Psychology: Human Perception and
Performance, 39, 1713–1730.
Jensen, O., & Mazaheri, A. (2010). Shaping functional
architecture by oscillatory alpha activity: Gating by inhibition.
Frontiers in Human Neuroscience, 4, 186.
Jiang, Y., & Chun, M. M. (2001). Selective attention modulates
implicit learning. The Quarterly Journal of Experimental
Psychology: Human Experimental Psychology, 54,
1105–1124.
Wang et al.
1547
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Kayser, J., & Tenke, C. E. (2015). Issues and considerations for
using the scalp surface Laplacian in EEG/ERP research: A
tutorial review. International Journal of Psychophysiology,
97, 189–209.
Kelly, S. P., Lalor, E. C., Reilly, R. B., & Foxe, J. J. (2006).
Increases in alpha oscillatory power reflect an active
retinotopic mechanism for distracter suppression during
sustained visuospatial attention. Journal of Neurophysiology,
95, 3844–3851.
Kiesel, A., Miller, J., Jolicœur, P., & Brisson, B. (2008).
Measurement of ERP latency differences: A comparison of
single-participant and jackknife-based scoring methods.
Psychophysiology, 45, 250–274.
Klimesch, W. (1996). Memory processes, brain oscillations
and EEG synchronization. International Journal of
Psychophysiology, 24, 61–100.
Klimesch, W. (1999). EEG alpha and theta oscillations reflect
cognitive and memory performance: A review and analysis.
Brain Research Reviews, 29, 169–195.
Klimesch, W. (2012). Alpha-band oscillations, attention, and
controlled access to stored information. Trends in Cognitive
Sciences, 16, 606–617.
Lopez-Calderon, J., & Luck, S. J. (2014). ERPLAB: An open-
source toolbox for the analysis of event-related potentials.
Frontiers in Human Neuroscience, 8, 213.
Luck, S. J., Heinze, H. J., Mangun, G. R., & Hillyard, S. A. (1990).
Visual event-related potentials index focused attention
within bilateral stimulus arrays. II. Functional dissociation
of P1 and N1 components. Electroencephalography and
Clinical Neurophysiology, 75, 528–542.
Luck, S. J., & Kappenman, E. S. (2012). ERP components and
selective attention. In E. S. Kappenman & S. J. Luck (Eds.),
The Oxford handbook of event-related potential
components (pp. 295–327). Oxford, United Kingdom: Oxford
University Press.
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190.
Moher, J., & Egeth, H. E. (2012). The ignoring paradox: Cueing
distractor features leads first to selection, then to inhibition
of to-be-ignored items. Attention, Perception, & Psychophysics,
74, 1590–1605.
Moore, T., & Armstrong, K. M. (2003). Selective gating of visual
signals by microstimulation of frontal cortex. Nature, 421,
370–373.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J. M. (2011).
FieldTrip: Open source software for advanced analysis of
MEG, EEG, and Invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011, 9.
Perrin, F., Pernier, J., Bertrand, O., & Echallier, J. F. (1989).
Spherical splines for scalp potential and current density
mapping. Electroencephalography and Clinical
Neurophysiology, 72, 184–187.
Sawaki, R., & Luck, S. J. (2010). Capture versus suppression
of attention by salient singletons: Electrophysiological
evidence for an automatic attend-to-me signal. Attention,
Perception, & Psychophysics, 72, 1455–1470.
Sawaki, R., & Luck, S. J. (2013). Active suppression after
involuntary capture of attention. Psychonomic Bulletin &
Review, 20, 296–301.
Theeuwes, J. (1991). Cross-dimensional perceptual selectivity.
Perception & Psychophysics, 50, 184–193.
Theeuwes, J. (1992). Perceptual selectivity for color and form.
Perception & Psychophysics, 51, 599–606.
Theeuwes, J. (2010). Top–down and bottom–up control of
visual selection. Acta Psychologica, 135, 77–99.
Theeuwes, J. (2018). Visual selection: Usually fast and
automatic; seldom slow and volitional. Journal of Cognition,
1, 21.
Theeuwes, J. (2019). Goal-driven, stimulus-driven, and history-
driven selection. Current Opinion in Psychology, 29, 97–101.
van Driel, J., Gunseli, E., Meeter, M., & Olivers, C. N. L. (2017).
Local and interregional alpha EEG dynamics dissociate
between memory for search and memory for recognition.
Neuroimage, 149, 114–128.
van Moorselaar, D., & Slagter, H. A. (2019). Learning what is
irrelevant or relevant: Expectations facilitate distractor
inhibition and target facilitation through distinct neural
mechanisms. Journal of Neuroscience. https://doi.org/
10.1523/JNEUROSCI.0593-19.2019
Wang, B., & Theeuwes, J. (2018a). Statistical regularities modulate
attentional capture. Journal of Experimental Psychology:
Human Perception and Performance, 44, 13–17.
Wang, B., & Theeuwes, J. (2018b). How to inhibit a distractor
location? Statistical learning versus active, top–down
suppression. Attention, Perception, & Psychophysics, 80,
860–870.
Wang, B., & Theeuwes, J. (2018c). Statistical regularities
modulate attentional capture independent of search strategy.
Attention, Perception, & Psychophysics, 80, 1763–1774.
Weaver, M. D., van Zoest, W., & Hickey, C. (2017). A
temporal dependency account of attentional inhibition in
oculomotor control. Neuroimage, 147, 880–894.
Won, B.-Y., Kosoyan, M., & Geng, J. J. (2019). Evidence for
second-order singleton suppression based on probabilistic
expectations. Journal of Experimental Psychology: Human
Perception and Performance, 45, 125–138.
Worden, M. S., Foxe, J. J., Wang, N., & Simpson, G. V. (2000).
Anticipatory biasing of visuospatial attention indexed by
retinotopically specific alpha-band electroencephalography
increases over occipital cortex. Journal of Neuroscience,
20, RC63.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
0
1
5
3
5
1
8
6
0
8
1
1
/
/
j
o
c
n
_
a
_
0
1
4
3
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
1548
Journal of Cognitive Neuroscience
Volume 31, Number 10