An fMRI Investigation of Attributing Negative Social
Treatment to Racial Discrimination
Carrie L. Masten, Eva H. Telzer, and Naomi I. Eisenberger
Abstracto
■ We used fMRI to examine the neural responses that occur
during experiences of perceived racial discrimination. Anterior
neuroimaging studies have focused exclusively on the processes
underlying racial bias from the perpetratorʼs perspective and have
yet to examine the processes that occur when individuals are be-
ing discriminated against. To extend this work, we examined the
neural correlates associated with attributing negative social treat-
ment to racial discrimination to explore the cognitive and affective
processes that occur as discrimination is being experienced. Hacer
este, we scanned Black participants while they were ostensibly ex-
cluded by Whites and then measured distress levels and race-
based attributions for exclusion. In response to being socially ex-
cluded by Whites, Black participants who appeared to be more
distressed showed greater social pain-related neural activity and
reduced emotion regulatory neural activity. Además, those
who attributed exclusion to racial discrimination displayed less so-
cial pain-related and more emotion regulatory neural activity. El
potential negative impact that frequent negative social treatment
and discrimination-related distress regulation might have on indi-
vidualsʼ long-term mental and physical health is discussed. ■
INTRODUCCIÓN
Sixty percent of Black Americans perceive racial discrimina-
tion on a daily basis (Sellers & Shelton, 2003). Psychologists
have long noted that this repeated exposure to discrimina-
tion is psychologically harmful for the target and may con-
tribute to well-established health disparities between Black
and White individuals (Mays, Cochran, & Barnes, 2007). Sur-
prisingly, sin embargo, most research has focused on Whitesʼ
negative attitudes toward Blacks (Shelton, 2000), en cambio
of Blacksʼ experiences as the target of racial bias. Además,
psychological research that has investigated the experience
of discrimination from the targetʼs perspective has been
limited by its reliance on self-report or behavioral indices
of felt discrimination that are typically measured after the
act of racial discrimination has been perceived and pro-
cessed to some extent. Functional magnetic resonance im-
aging (resonancia magnética funcional) may be an important complement to these
self-report and behavioral assessments of discrimination
experiences because fMRI enables the measurement of
neural responses to these experiences as they unfold.
Examining on-line neural responses during discrimina-
tion may be particularly helpful given the inconsistencies
in behavioral research regarding which psychological pro-
cesses are involved in experiencing discrimination. En otra
palabras, although being the target of discrimination is asso-
ciated with reduced well-being (Major, Quinton, & McCoy,
2002) and long-term negative consequences for physical
and mental health (p.ej., Landrine & Klonoff, 2000; Krieger
& Sídney, 1996; see Mays et al., 2007 para una revisión), it is not
Universidad de California, Los Angeles
yet clear how attributing negative social treatment to dis-
crimination affects individuals in the moment. Para examen-
por ejemplo, some research suggests that attributing negative social
treatment to discrimination is distressing because discrimi-
nation is due to an internal characteristic of the target (dis-
crimination is due to “my race”), which cannot be changed
and might therefore lead to future instances of discrimina-
ción (Wirth & williams, 2009; Schmitt & Branscombe, 2002;
Branscombe, 1998). Sin embargo, other research has sug-
gested that attributing negative social treatment to discrim-
ination may be distress-reducing and provide a buffer if the
prejudice is viewed as a characteristic of the perpetrator
(discrimination is due to “others being prejudiced”), y
the likelihood of self-blame is reduced (Major, Kaiser, &
McCoy, 2003; Crocker & Major, 1989). It should be noted
that both of these accounts presume that receiving nega-
tive social treatment is distressing. What is in question in
these two lines of research is whether attributing this nega-
tive treatment to discrimination increases or decreases this
distress. To extend behavioral research and provide new
insight about the processes that occur as individuals at-
tribute negative social treatment to discrimination, we used
fMRI to examine the neural correlates associated with the
distress of negative social treatment as well as how making
discriminatory attributions (attributing negative treatment
to racial discrimination) alters Black participantsʼ immedi-
ate affective responses to this treatment.
Although neuroimaging research has not examined vic-
timsʼ experiences with discrimination, previous work has
established a network of neural regions supporting the
experience and regulation of responses to social threats
that provides a framework from which to examine racial
© 2011 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 23:5, páginas. 1042–1051
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
3
yo
5
mi
–
1
pag
0
d
4
F
2
/
1
2
9
3
4
/
1
5
0
/
4
1
2
0
oh
4
C
2
norte
/
1
2
0
7
1
7
0
5
2
2
8
1
0
5
2
/
0
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
1
tu
0
mi
.
s
t
2
oh
1
norte
5
2
0
0
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
t
/
F
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
discriminación. Research has implicated dorsal anterior
cingulate cortex (dACC) and anterior insula in the “pain-
ful” or distressing aspects of social rejection (DeWall et al.,
in press; Masten et al., 2009; Way, taylor, & Eisenberger,
2009; Eisenberger, Lieberman, & williams, 2003). Investigación
has also suggested that several regions of lateral prefrontal
corteza (PFC; ventrolateral, dorsolateral) may play a role
in regulating negative affective responses to social or painful
threats, as indexed by correlated reductions in dACC and an-
terior insular activity (Lieberman et al., 2004; Wager et al.,
2004; Eisenberger et al., 2003; Petrovic & Ingvar, 2002). simí-
mucho, regions of medial PFC and rostral ACC (rACC) may play
a regulatory role as well, as evidenced by their contribution
to pain regulation (Petrovic & Ingvar, 2002), extinction of
learned fears (Phelps, delgado, Nearing, & El dulce, 2004),
and positive interpretations of potentially negative stimuli
(kim, Somerville, Johnstone, Alexander, & Whalen, 2003).
Based on this neural framework, we predicted that the
distress associated with negative social treatment would
be associated with greater activity in regions associated with
social distress (dACC, anterior insula), and reduced activity
in regions involved in emotion regulation (PFC, rACC).
Además, we also examined how attributing this negative
social treatment to discrimination would relate to these
neural responses. Based on previous work, to the extent
that attributing negative social treatment to discrimination
is personally threatening and distressing in the moment,
making discriminatory attributions should be associated
with more activity in neural regions underlying distress
(dACC, anterior insula) and less activity in regions that regu-
late these negative experiences (PFC, rACC). Alternativamente,
to the extent that attributing negative social treatment to
discrimination provides a way of coping with negative treat-
ment and reduces distress in the moment, making discrimi-
natory attributions should be associated with less neural
evidence of distress and more evidence of regulation.
MÉTODOS
Participantes
Eighteen Black individuals (9 women), ranging in age from
19 a 28 años (m = 21.4), participated in the study. Participe-
pants consisted of UCLA students and staff, as well as mem-
bers of the surrounding community who were unaffiliated
with the University. Participants were recruited through
fliers distributed around the UCLA campus, announcements
made in undergraduate classes, postings on student list-
serves, and on-line advertisements in the community. Carrera
was never mentioned in any of the recruitment materials.
All participants completed written consent in accordance
with UCLAʼs Institutional Review Board.
Procedures
To examine how making discriminatory attributions for
negative social treatment would impact affective and neural
responses to negative social treatment, participants were
scanned as they were ostensibly excluded by two White
individuals and could thus believe that they were the target
of racial discrimination. In order to make race salient and
increase the probability that racial discrimination might be
considered a viable attribution for being excluded, participar-
pants were introduced to two White confederates (1 masculino,
1 femenino) prior to scanning. Además, interviewers and
scanning personnel were White. The participant and con-
federates completed consents together and were told that
they would be playing a game together during their scans.
They then introduced themselves by stating their name,
occupation or year and major in college, and something in-
teresting about themselves. Aside from this visible priming
of race, race was not discussed during any part of the experi-
mental session.
fMRI Task
To simulate a real, interactive experience of social exclusion,
participants completed an experimental paradigm called
“Cyberball” (Williams et al., 2002; williams, Cheung, & Choi,
2000), in which they were told that they would play a com-
puterized ball-tossing game via the Internet with two other
participants in other scanners. En realidad, participants played
with a computer. Throughout the game of Cyberball, el
ball was thrown back and forth among the three players, con
the participant choosing the recipient of their own throws,
and the throws of the other two “players” determined by
the preset program. Participants could see the images rep-
resenting the other two players on a computer screen,
as well as their own “hand” that they controlled using a
button-box. During the fMRI scan, participants played two
rounds of Cyberball, one in which they were included
equally for the whole game and one in which they were ex-
cluded after initially being included for 10 throws. This para-
digm has been used previously in several behavioral and
neuroimaging studies to successfully simulate an experience
of social exclusion and produce feelings of distress related
to this exclusion (Masten et al., 2009; Eisenberger, Gable,
& Lieberman, 2007; Van Beest & williams, 2006; Zadro,
williams, & Richardson, 2004; Eisenberger et al., 2003).
Following the completion of several postscan measures
administered at the end of the fMRI session, Participantes
were given a full debriefing explaining the deception in-
volved in the Cyberball game, including the intentional
priming of race, and were thoroughly questioned about
their feelings regarding this deception.
Postscan Measures
Following the scan, participants completed self-report mea-
sures of social distress and discriminatory attributions (de-
scribed below). Además, because self-report bias was of
particular concern, given the sensitive nature of discussing
oneʼs feelings about being discriminated against, nosotros también
examined a nonverbal behavioral measure of distress. Cada
participant completed a videotaped interview, in which they
Masten, Telzer, and Eisenberger
1043
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
3
yo
5
mi
–
1
pag
0
d
4
F
2
/
1
2
9
3
4
/
1
5
0
/
4
1
2
0
oh
4
C
2
norte
/
1
2
0
7
1
7
0
5
2
2
8
1
0
5
2
/
0
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
1
tu
0
mi
.
s
t
2
oh
1
norte
5
2
0
0
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
.
.
F
t
/
oh
norte
1
8
METRO
a
y
2
0
2
1
were asked to comment on their thoughts/feelings at the
time of the exclusion episode. These interviews were later
rated by observers for negative affect (“observer-rated dis-
tress”) as described below.
Self-reported Distress
To measure self-reported distress, participants completed
the Need–Threat Scale (Williams et al., 2000, 2002) seguir-
ing completion of the functional scan, and they were ex-
plicitly told to answer the questions based on the second
round of the game (during which the exclusion occurred).
This measure assesses 12 subjectively experienced con-
sequences of being excluded during the game (p.ej., “I felt
liked,” “I felt rejected”), using a 5-point scale ranging from
1 (not at all) a 5 (very much). Items were reverse-coded
appropriately and averaged to create a measure of self-
reported distress that was highly reliable (un = .86).
Observer-rated Distress
Because of the sensitive nature of this interracial situation
and the fact that self-reports of social distress could be tem-
pered or altered due to self-presentational concerns (Henry
& Hardin, 2006; Dovidio et al., 1997), we also included
a nonverbal, behavioral measure of participantsʼ distress
niveles. Immediately after the scanning session, each par-
ticipant was videotaped while completing an interview in
which they described their thoughts and feelings during
the game. They were asked to discuss the feelings that they
had when they were being excluded as well as the reasons
for why they might have been excluded; they were not
prompted to discuss race. Más tarde, 21 individuals who were
blind to the studyʼs purpose observed the videos and rated
each clip using a 5-point scale (1 = not at all to 5 = very
mucho) to indicate how upset, sad, rejected, angry, and de-
feated each participant seemed (p.ej., “How upset did the
subject seem?"). Ratings for these five emotions were aver-
aged and standardized to create a measure of observer-rated
distress (ICC = .91). Previous work has shown that non-
verbal measures of behavioral responses, similar to this
uno, are a better indicator of affect and less influenced by
self-presentational biases than self-reports (Kawakami, Phills,
Steele, & dovidio, 2007; Dovidio et al., 1997; Word, Zanna,
& Cooper, 1974).
Self-reported Discriminatory Attributions
After completing ratings of self-reported distress, participar-
pants indicated the extent to which they thought they
were excluded (during the second round of the Cyberball
juego) because of their race by answering three questions:
(1) “How much do you believe the others responded to
you during the game because of your race?," (2) “How
much did you feel you were discriminated against by the
other players during the game?,” and (3) “How much do
you believe the other players in the game were racist?,"
using a 5-point scale (1 = not at all to 5 = very much)
(Goodwin, williams, & Carter-Sowell, 2010). Responses
to these three questions were averaged to create a com-
posite score of self-reported discriminatory attributions
with good reliability (un = .79). These questions were im-
bedded in a larger questionnaire that included distracter
items in an effort to reduce suspicion regarding the pur-
pose of the study.
Neuroimaging Procedures
fMRI Data Acquisition
Imaging data were collected using a Siemens Allegra 3-Tesla
MRI scanner. An initial 2-D spin-echo image (TR = 4000 mseg,
TE = 40 mseg, matrix size 256 × 256, 4 mm thick, 1 mm
gap) in the sagittal plane was acquired to enable prescrip-
tion of slices obtained in structural and functional scans. En
addition, a high-resolution structural T2*-weighted echo-
planar imaging volume (TR = 4000 mseg, TE = 54 mseg,
matrix size 128 × 128, FOV = 200 mm, 36 slices, 1.56-mm in-
plane resolution, 3 mm thick) was acquired coplanar with the
functional scans. Each of the two rounds of Cyberball was
completed during a functional scan lasting 2 mín. 48 segundo
(echo-planar T2*-weighted gradient-echo, TR = 2000 mseg,
TE = 25 mseg, flip angle = 90°, 64 × 64 matrix, FOV =
200 mm; 36 slices, 3 mm thick, 3 mm3 voxel size, skip 1 mm).
fMRI Data Analysis
All neuroimaging data were preprocessed and analyzed
using Statistical Parametric Mapping (SPM5; Wellcome De-
partment of Cognitive Neurology, Institute of Neurology,
Londres, Reino Unido). Images for each participant were realigned
to correct for head motion (no participantʼs motion ex-
ceeded 1.5 mm), normalized into a standard stereotactic
space as defined by the Montreal Neurological Institute
and the International Consortium for Brain Mapping, y
smoothed using an 8-mm Gaussian kernel, full width at half
maximum, to increase the signal-to-noise ratio.
Cyberball was modeled as a block design. Each round
of Cyberball was modeled as a run with periods of inclu-
sion and exclusion modeled as blocks within the runs.
The order of blocks was not counterbalanced in an effort
to maintain ecological validity and avoid residual brain ac-
tivation from the exclusion round being confounded with
the activation recorded during inclusion. Because the para-
digm is self-advancing for each participant, block lengths
varied slightly across individuals; sin embargo, final rest peri-
ods allowed for this variation within a functional scan last-
ing a set amount of time. After modeling the Cyberball
paradigma, linear contrasts were calculated for each planned
condition comparison for each participant. These individ-
ual contrast images were then used in whole-brain, grupo-
level random-effects analyses across all participants.
Group-level analyses were thresholded at p < .005 for magnitude with a minimum cluster size threshold of 10 voxels (Lieberman & Cunningham, 2009), in order to 1044 Journal of Cognitive Neuroscience Volume 23, Number 5 D o w n l o a d e d l l / / / / j t t f / i t . : / / f r o m D h o t w t n p o : a / d / e m d i f t r o p m r c h . s p i l d v i e r e r c c t . h m a i r e . d u c o o m c / n j a o r c t i n c / e a - p r d t i 2 c 3 l 5 e - 1 p 0 d 4 f 2 / 1 2 9 3 4 / 1 5 0 / 4 1 2 0 o 4 c 2 n / 1 2 0 7 1 7 0 5 2 2 8 1 0 5 2 / 0 j o p c d n . b y 2 0 g 1 u 0 e . s t 2 o 1 n 5 2 0 0 7 . S p e d p f e m b y b e g r u 2 0 e 2 s 3 t / j / t . . . f o n 1 8 M a y 2 0 2 1 examine all regions for which a priori hypotheses were made, specifically those involved in the experience of social threat as well as the regulation of these negative affective responses (e.g., dACC, anterior insula, PFC). We used a more conservative threshold of p < .001 for magnitude with a minimum cluster size threshold of 20 voxels to examine all other areas of the brain (Lieberman & Cunningham, 2009). All coordinates are reported in Montreal Neurological Institute format. First, we examined neural activity during exclusion com- pared to inclusion. Next, to test the primary research questions, we conducted whole-brain regression analyses to examine how each of the following variables correlated with neural activity during exclusion compared to inclu- sion: (a) participantsʼ self-reported distress scores; (b) observer-rated distress scores; and (c) the degree to which each participant believed that he/she was excluded be- cause of his or her race. RESULTS Behavioral Results Analysis of postscan measures of distress and perceived dis- crimination demonstrated substantial variability in each of these measures. Self-reported distress among participants ranged from 2.08 to 4.50 on a scale from 1 (not at all) to 5 (very much), with an average rating of 2.99 (SD = .68), whereas observer-rated distress ranged from 1.37 to 3.29 on the same scale, with an average rating of 2.11 (SD = .55). Self-reported and observer-rated distress were posi- tively correlated (r = .50, p < .05). In terms of participantsʼ perceived discrimination, self-reports of discriminatory at- tributions ranged from 1 to 4.67 (on a scale from 1 to 5), with a mean rating of 2.13 (SD = 1.06); 15 of the 18 (83%) participants reported that they had been excluded because of their race to some degree (ratings above 1 for at least one of the three items). In addition, although participantsʼ self-reported discriminatory attributions correlated posi- tively with their self-reported distress (r = .51, p < .05), these attributions did not significantly relate to observer- rated distress (r = .32, ns).1 Neuroimaging Results Neural Activity during Exclusion versus Inclusion First, we examined the main effect of experiencing exclu- sion compared to inclusion. A whole-brain analysis revealed several regions that were significantly more active during exclusion versus inclusion, including both limbic regions previously linked with pain-related processing (e.g., anterior insula) and regions previously linked with emotion reg- ulation (e.g., VLPFC, rACC), consistent with previous stud- ies examining social exclusion (Eisenberger, Gable, et al., 2007; Eisenberger, Taylor, Gable, Hilmert, & Lieberman, 2007; Eisenberger et al., 2003; see Table 1 for a complete list of activations). Associations between Distress and Neural Activity during Exclusion versus Inclusion To examine how distress was related to neural activity, we correlated neural activity during exclusion versus in- clusion with: (a) participantsʼ self-reported distress result- ing from exclusion and (b) observer-rated distress resulting from exclusion. Whole-brain regression analyses (during exclusion vs. inclusion) revealed that self-reported dis- tress did not correlate positively with the limbic regions that have previously been seen in studies examining the experience of social threats (e.g., dACC, anterior insula), but did correlate negatively with PFC activity (Table 2A). Thus, this self-report measure may have assessed some- thing other than social distress as it is typically mea- sured, which is not surprising in this sensitive, interracial situation. Correlations with observer-rated distress, however, re- vealed a neural network consistent with previous studies examining social distress and the regulation of this distress (see Table 2B). Participants with higher observer-rated dis- tress displayed greater activity in dACC (Figure 1A) and anterior insula (Figure 1B), reduced activity in ventrolateral PFC ( VLPFC) and anterolateral PFC (ALPFC), regions in- volved in emotion regulation, and reduced activity in dorso- medial PFC (DMPFC; Table 1B), a region which has been linked with perspective-taking and mentalizing about others (Mitchell, Macrae, & Banaji, 2006), as well as emotion regu- lation (Ochsner, Bunge, Gross, & Gabrieli, 2002). In other words, individuals who were rated by outside observers as appearing more distressed during their postexclusion inter- views showed greater neural activity in regions associated with social distress and reduced activity in regions associ- ated with emotion regulation while they were being socially excluded.2 As such, this nonverbal behavioral measure of distress may provide an index that is more in line with what has been observed in previous studies of rejection-related distress. Associations between Perceived Discrimination and Neural Activity during Exclusion versus Inclusion Next, to examine how attributing social exclusion to dis- crimination related to neural activity, we correlated neural activity during exclusion versus inclusion with the extent to which participants reported that they were excluded because of their race (self-reported discriminatory attribu- tions). Interestingly, neural responses indicated that making discriminatory attributions for social exclusion was asso- ciated with less activity in neural regions associated with distress and threat perception and greater activity in neural regions associated with emotion regulation (see Table 3). Thus, in contrast to the relationship between observer-rated distress and neural activity, the more participants felt that they were excluded because of their race, the less activity they displayed in areas responsive to social threats, includ- ing dACC (Figure 2A), and the more activity they displayed Masten, Telzer, and Eisenberger 1045 D o w n l o a d e d l l / / / / j f / t t i t . : / / f r o m D h o t w t n p o : a / d / e m d i f t r o p m r c h . s p i l d v i e r e r c c t . h m a i r e . d u c o o m c / n j a o r c t i n c / e a - p r d t i 2 c 3 l 5 e - 1 p 0 d 4 f 2 / 1 2 9 3 4 / 1 5 0 / 4 1 2 0 o 4 c 2 n / 1 2 0 7 1 7 0 5 2 2 8 1 0 5 2 / 0 j o p c d n . b y 2 0 g 1 u 0 e . s t 2 o 1 n 5 2 0 0 7 . S p e d p f e m b y b e g r u 2 0 e 2 s 3 t / j f / . t . . o n 1 8 M a y 2 0 2 1 Table 1. Regions Activated during Exclusion versus Inclusion Anatomical Region BA Exclusion > Inclusion
Anterior insula
Anterior insula
rACC/ VMPFC
VLPFC
LPFC
LPFC
DMPFC
Premotor cortex†
MTG†
ITG†
STG†
Occipital cortex†
Occipital cortex†
Occipital cortex†
Cerebellum†
11
45
44/45
44/45
8
6
21
20
22
19
19
19
R
R
R
R
R
l
R
R
l
R
l
l
R
l
R
X
46
42
4
58
44
−50
0
52
−50
48
−58
−44
38
−28
34
y
z
t
24
22
44
30
12
12
42
4
−18
−22
−54
−80
−78
−84
−70
−10
0
−10
6
24
24
44
50
−8
−18
16
28
44
36
−14
4.47
4.13
4.59
3.75
5.58
3.92
5.07
4.65
6.86
5.53
5.13
5.97
5.81
4.58
5.65
k
212
212
58
445
445
138
182
115
30
35
84
38
79
39
25
pag
<.0005
<.0005
<.0005
<.001
<.0001
<.001
<.0001
<.0005
<.0001
<.0001
<.0001
<.0001
<.0001
<.0005
<.0001
Regions marked with a cross (†) are those that were not the primary focus of this investigation but that were still significant at p < .001, 20 voxels (k-values
listed for these were taken from the thresholding map at p < .001). All other regions (e.g., limbic, prefrontal) are listed at p < .005, 10 voxels. BA refers to
putative Brodmannʼs area; L and R refer to left and right hemispheres; x, y, and z refer to MNI coordinates; t refers to the t score at those coordinates (local
maxima); k refers to the number of voxels in each significant cluster. rACC = rostral anterior cingulate cortex; VMPFC = ventromedial prefrontal cortex;
VLPFC = ventrolateral prefrontal cortex; LPFC = lateral prefrontal cortex; DMPFC = dorsomedial prefrontal cortex; MTG = middle temporal gyrus; ITG =
inferior temporal gyrus; STG = superior temporal gyrus.
in regions associated with regulation of threat responses,
specifically rACC (Figure 2B; see Table 3).
DISCUSSION
This study is the first to examine the neural correlates of
racial bias from the victimʼs point of view. As such, two sets
of findings emerged as particularly novel and informative
for our understanding of experiences with racial discrimi-
nation: (1) those who appeared to be more distressed by
the exclusion episode showed greater social pain-related
neural activity and reduced emotion regulatory neural
activity; and (2) attributing this negative social treatment
to discrimination was associated with the reverse pattern-
reduced social pain-related neural activity and greater reg-
ulatory activity.
First, although Black participantsʼ self-reported distress
was not associated with greater activity in regions previously
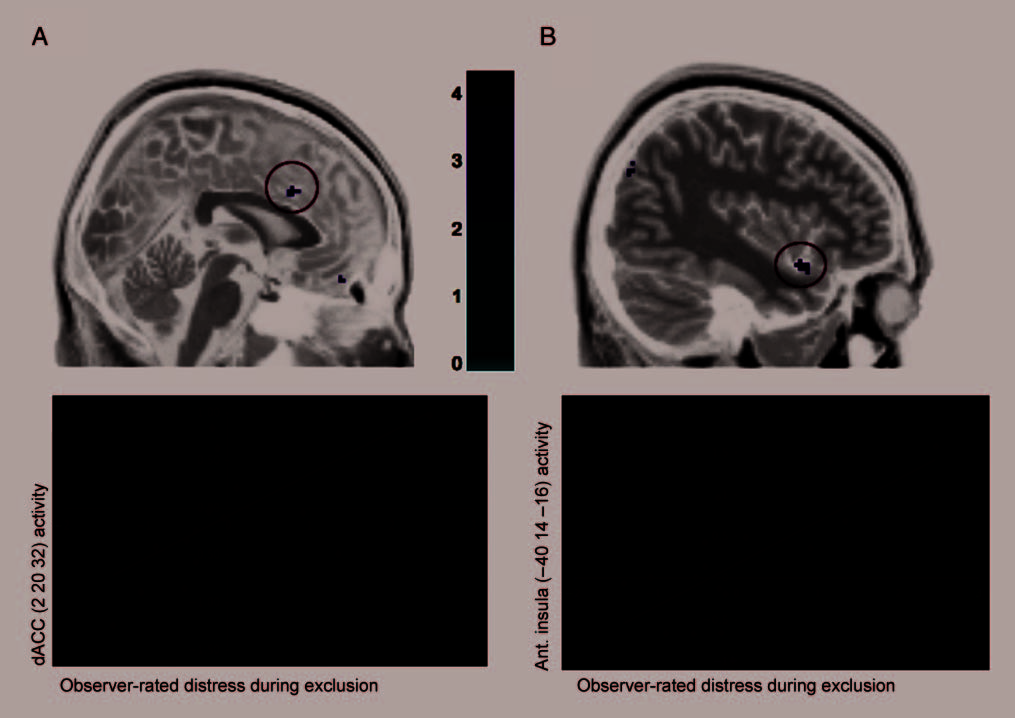
linked to social distress, observer-rated distress was asso-
ciated with greater activity in these neural regions. This is
consistent with previous research indicating that nonverbal
measures of behavioral responses provide a better index
of affective experience than self-reports (Kawakami et al.,
2007; Dovidio et al., 1997; Word et al., 1974) and suggests
that these nonverbal behavioral measures may provide a
better index of neural responding as well. These findings
also highlight the distress associated with negative social
treatment in general and suggest that Black individuals,
who may face negative social treatment more frequently,
may unfortunately experience this distress more often.
Given that experiences of negative social treatment have
been linked to increased physiological stress responding
(Eisenberger, Taylor, et al., 2007; Dickerson & Kemeny,
2004) and negative mental health outcomes (Slavich,
Thornton, Torres, Monroe, & Gotlib, 2009), it is likely that
repeated experiences with these negative events take a
cumulative toll.
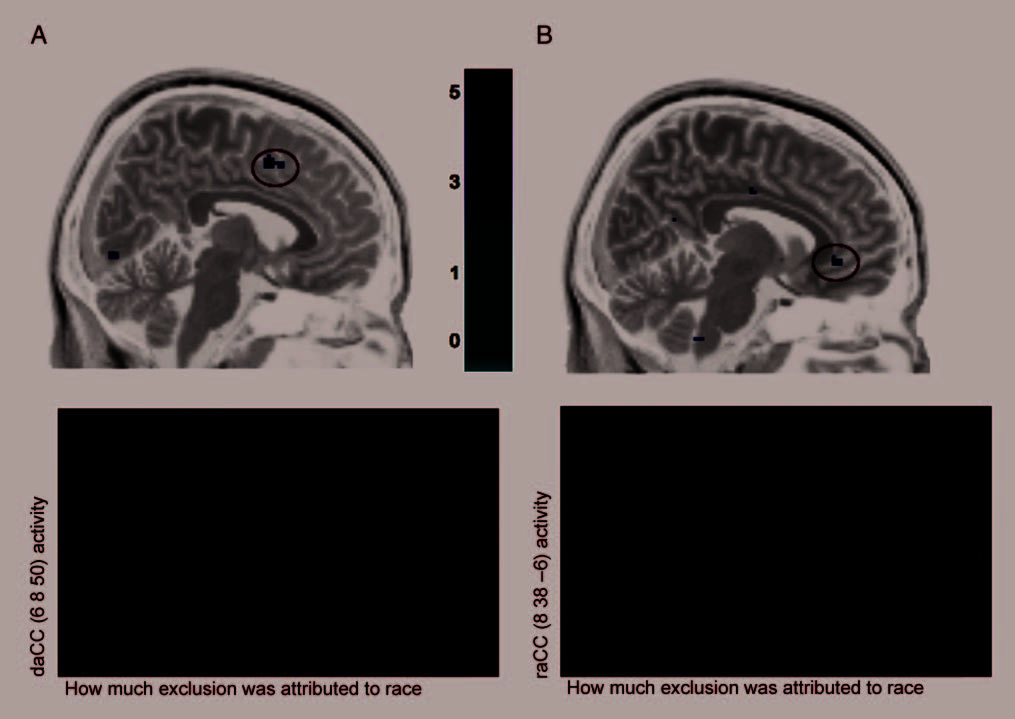
Second, our neural findings demonstrated that Black
participants who attributed exclusion to racial discrimina-
tion showed less activity in distress-related regions and
more activity in regions previously linked with emotion/
pain regulation. This provides support for the notion that
making discriminatory attributions may buffer against neg-
ative emotions as the perceived discrimination is taking
place. These findings extend previous behavioral work sug-
gesting that when individuals make discriminatory attribu-
tions in situations of negative social treatment, blame for
the negative treatment is attributed externally (discrimi-
nation is due to “others being prejudiced”), thus reducing
negative affect and self-blame (Major et al., 2003; Crocker
& Major, 1989). Thus, although other research has sug-
gested that making discriminatory attributions may increase
1046
Journal of Cognitive Neuroscience
Volume 23, Number 5
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
5
e
-
1
p
0
d
4
f
2
/
1
2
9
3
4
/
1
5
0
/
4
1
2
0
o
4
c
2
n
/
1
2
0
7
1
7
0
5
2
2
8
1
0
5
2
/
0
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
2
0
0
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
.
f
t
.
o
n
1
8
M
a
y
2
0
2
1
Table 2. Correlations with Social Distress
Anatomical Region
BA
x
y
z
t
r
k
p
(A)
Positive Associations with Self-reported Distress
subACC
25
L
−12
Negative Associations with Self-reported Distress
DMPFC
Premotor cortex†
8
6
L
L
−8
−24
(B)
Positive Associations with Observer-rated Distress
dACC
Anterior insula
VMPFC
DMPFC
32
11
8
R
L
R
L
Negative Associations with Observer-rated Distress
VLPFC
ALPFC
DMPFC
Somatosens. cortex†
Cerebellum†
IPL†
Precuneus†
Somatosens. cortex†
45
10
8/9
1/2/3
40
4
1/2/3
R
L
L
R
L
L
R
L
2
−40
4
−6
48
−20
−12
34
−22
−38
10
−22
16
42
4
20
14
48
54
32
44
42
−34
−50
−52
−36
−34
−8
44
54
32
−16
−18
48
8
4
42
74
−46
40
70
58
3.34
4.77
3.47
3.94
3.24
4.58
3.38
3.84
3.90
6.95
6.49
6.42
6.28
6.06
3.41
.65
10
<.005
−.64
−.75
16
111
<.005
<.0005
D
o
w
n
l
o
a
d
e
d
.66
.70
.63
.70
−.65
−.69
−.70
−.87
−.85
−.85
−.84
−.83
19
17
20
23
15
31
61
50
33
44
132
94
<.005
<.001
<.005
<.001
<.005
<.001
<.001
<.0001
<.0001
<.0001
<.0001
<.0001
Regions activated during exclusion versus inclusion that correlated significantly with: (A) self-reported distress and (B) observer-rated distress result-
ing from exclusion.
Regions marked with a cross (†) are those that were not the primary focus of this investigation but that were still significant at p < .001, 20 voxels (k-values
listed for these were taken from the thresholding map at p < .001). All other regions (e.g., limbic, prefrontal) are listed at p < .005, 10 voxels. BA refers to
putative Brodmannʼs area; L and R refer to left and right hemispheres; x, y, and z refer to MNI coordinates; t refers to the t score at those coordinates (local
maxima); r refers to the correlation coefficient representing the strength of the association between each regressor and activity in the specified region;
k refers to the number of voxels in each significant cluster. subACC = subgenual anterior cingulate cortex; DMPFC = dorsomedial prefrontal cortex;
dACC = dorsal anterior cingulate cortex; VMPFC = ventromedial prefrontal cortex; VLPFC = ventrolateral prefrontal cortex; ALPFC = anterolateral pre-
frontal cortex; somatosens. cortex = somatosensory cortex; IPL = inferior parietal lobule.
negative affect in the face of negative treatment (Wirth &
Williams, 2009; Schmitt & Branscombe, 2002; Branscombe,
1998), our examination of the underlying neural processes
associated with perceived discrimination suggests that mak-
ing race-related discriminatory attributions may facilitate
regulation of negative affective responses as negative treat-
ment is occurring, and provide a temporary coping mecha-
nism in such situations.
Although making discriminatory attributions may, in
some cases, be distress-reducing in the short term, these
findings should not be taken to imply that being the target
of discrimination has no negative consequences. Many be-
havioral studies have suggested otherwise, and additional
studies using novel levels of analysis will continue to eluci-
date these consequences. For example, as mentioned pre-
viously, facing repeated negative social treatment as the
target of discrimination may increase stress-related physio-
logical responding or lead to long-term mental and physical
health problems (Mays et al., 2007). In addition, the fre-
quent need to regulate oneʼs response to negative social
treatment (either through external attributions for negative
treatment or other means) may interfere with oneʼs ability
to cope with other stressors. For example, research on ego
depletion has shown that exerting self-control or emotion
regulatory processes can impair oneʼs ability to engage these
processes subsequently (Baumeister, Vohs, & Tice, 2007).
Masten, Telzer, and Eisenberger
1047
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
5
e
-
1
p
0
d
4
f
2
/
1
2
9
3
4
/
1
5
0
/
4
1
2
0
o
4
c
2
n
/
1
2
0
7
1
7
0
5
2
2
8
1
0
5
2
/
0
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
2
0
0
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
.
f
t
/
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
5
e
-
1
p
0
d
4
f
2
/
1
2
9
3
4
/
1
5
0
/
4
1
2
0
o
4
c
2
n
/
1
2
0
7
1
7
0
5
2
2
8
1
0
5
2
/
0
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
2
0
0
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
t
.
/
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 1. Activity during exclusion versus inclusion that correlated positively with observersʼ ratings of participantsʼ distress in the (A) dACC
and (B) anterior insula.
Table 3. Correlations with Discriminatory Attributions
Anatomical Region
BA
x
y
z
t
Positive Associations with Attributing Exclusion to Race
rACC
Lat. temp. cortex†
Brainstem†
32
21
R
L
R
8
−46
2
Negative Associations with Attributing Exclusion to Race
dACC
Motor cortex†
Motor cortex†
6/32
4
4
R
L
R
6
−46
50
38
−10
−38
8
−6
−12
−6
−22
−48
50
56
38
3.36
5.64
4.88
4.01
4.27
5.58
r
.64
.82
.77
−.71
−.73
−.81
k
11
47
24
45
62
534
p
<.005
<.0001
<.0001
<.001
<.0005
<.0001
Regions activated during exclusion vs. inclusion that correlated significantly with participantsʼ self-reported ratings of discriminatory attributions.
Regions marked with a cross (†) are those that were not the primary focus of this investigation but that were still significant at p < .001, 20 voxels
(k-values listed for these were taken from the thresholding map at p < .001). All other regions (e.g., limbic, prefrontal) are listed at p < .005,
10 voxels. BA refers to putative Brodmannʼs Area; L and R refer to left and right hemispheres; x, y, and z refer to MNI coordinates; t refers to
the t score at those coordinates (local maxima); r refers to the correlation coefficient representing the strength of the association between each
regressor and activity in the specified region; k refers to the number of voxels in each significant cluster. rACC = rostral anterior cingulate cortex; lat.
temp. cortex = lateral temporal cortex; dACC = dorsal anterior cingulate cortex.
1048
Journal of Cognitive Neuroscience
Volume 23, Number 5
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
5
e
-
1
p
0
d
4
f
2
/
1
2
9
3
4
/
1
5
0
/
4
1
2
0
o
4
c
2
n
/
1
2
0
7
1
7
0
5
2
2
8
1
0
5
2
/
0
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
2
0
0
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
.
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Correlations between neural activity and self-reported discriminatory attributions. (A) dACC activity during exclusion versus inclusion that
correlated negatively with the degree to which participants believed their exclusion was related to their race. (B) rACC activity during exclusion
versus inclusion that correlated positively with the degree to which participants believed their exclusion was related to their race.
One implication of this is that having to cope with or regulate
the stress of discrimination may affect oneʼs ability to cope
with or regulate subsequent stressors (Baumeister, Faber,
& Wallace, 1999). Thus, it is possible that, for individuals
who experience discrimination frequently, making discrimi-
natory attributions may help reduce distress at the time of
the experience, but may also leave these individuals ego-
depleted—decreasing their ability to cope with subsequent
stressors and thus increasing psychological and physical
stress over time. Although we have shown that making dis-
criminatory attributions during the unfolding of a negative
social event is associated with less distress-related neural
activity and heightened activity in regulatory regions, the
cumulative toll that making these attributions and regulat-
ing distress has over time remains unclear and will be an
important topic of future research.
It is not clear in this study why discriminatory attribu-
tions correlated negatively with distress-related neural
activity but positively with self-reported social distress.
One possibility is that the self-report measure of social
distress and the distress-related neural activity reflect some-
what different affective consequences of perceiving dis-
crimination. For example, Major et al. (2003) have shown
that making discriminatory attributions for negative social
treatment may attenuate self-directed emotions (e.g., de-
pressed, worthless), but not necessarily other-directed
emotions (e.g., angry, mad). Thus, it is possible that the
self-report measure and the neural measure were assessing
slightly different features of affective experience. Another
possibility is that making discriminatory attributions may
have different effects on immediate affective experience
(which might be better captured by the neural responses)
versus reflective affective experience (which might be bet-
ter captured by the self-report measures) (Redelmeier &
Kahneman, 1996). Given that the self-report measure of
social distress did not demonstrate typical patterns of corre-
lations with neural activity, it is not clear what this measure
was assessing in the current study. Future research will be
needed to more fully examine this as well as to delineate
the effects of making discriminatory attributions on the
neural correlates of self- versus other-directed emotions
and immediate versus reflective affective responses follow-
ing negative social treatment.
In addition, future studies should also examine other
contextual factors that might have influenced the findings
reported here. For example, one limitation of the current
Masten, Telzer, and Eisenberger
1049
study is that the researchers and interviewers who asked
participants about their exclusion experience were White.
Although this likely boosted the implicit priming of race
(i.e., all the individuals interacting with the participant
prior to the scan were White), it is also possible that the
participant acted differently during their interview than
they would have if a Black experimenter conducted the
interview—particularly given that race was a frequent topic
of discussion raised by the participants. It would be useful
for future work to specifically examine the impact of inter-
acting with either racial ingroup or outgroup members
after a discrimination experience. An additional limitation
of the current study is that we did not have a videotaped
interview before participants were scanned, and thus, we
do not have a baseline for participantsʼ affect prior to the
experimental manipulation. This would be a useful ad-
dition in future studies. Lastly, in interpreting our neuro-
imaging results, some inferences were based on previous
research linking specific regions and behavioral functions.
The ability to judge with certainty the meaning of activation
in a particular region is limited, given the multiple func-
tions that a region may be involved in (Poldrack, 2006),
and future work directly testing these inferences will be
useful in providing additional support for our proposed
interpretations.
Finally, it is worth noting that although we did find greater
activity in the anterior insula in the direct comparison of
exclusion versus inclusion, we did not find greater activity
in dACC as previous research typically has, reflecting the dis-
tress that participants feel as they are being excluded (e.g.,
DeWall et al., in press; Way et al., 2009; Eisenberger, Gable,
et al., 2007; Eisenberger, Taylor, et al., 2007; Eisenberger
et al., 2003). Given that one of our hypotheses was that at-
tributing negative social treatment to race might reduce the
negative consequences of social exclusion, it is not too sur-
prising that the previously observed main effect of dACC
activity during social exclusion compared to inclusion did
not replicate. In fact, if it is the case that making discrimina-
tory attributions is pain-reducing, and the experimental
situation (being excluded by outgroup members) elicited
a high frequency of discriminatory attributions, it would fol-
low that certain types of pain-related neural activity might
not be observed during social exclusion.
To conclude, the findings reported here provide new in-
sight into the underlying processes that occur when individ-
uals perceive racial discrimination. Although the long-term
negative consequences of discrimination are undeniable,
these findings suggest one possible mechanism whereby
individuals who deal with negative social treatment on a
regular basis are able to cope with these experiences as
they occur. As such, this study is the first to provide neural
evidence that discriminatory attributions may help reduce
immediate negative affect among stigmatized groups in
situations of negative treatment and provides new insight
into the experiences of those targeted by racial bias. Fu-
ture work will be needed to examine the long-term conse-
quences of these processes.
Acknowledgments
We thank the staff of the UCLA Brain Mapping Center for their
assistance. This project was supported by a grant awarded to
N. Eisenberger from the UCLA Export Center on Research, Educa-
tion, Training and Strategic Communication on Minority Health
Disparities (Grant no. 5 P60 MD000508-04). Preparation of this
manuscript was supported by a National Science Foundation
Graduate Research Fellowship, an Elizabeth Munsterberg Koppitz
Fellowship, and a Ruth L. Kirschstein National Research Service
Award to C. Masten and a National Science Foundation Graduate
Research Fellowship to E. Telzer.
Reprint requests should be sent to Naomi I. Eisenberger, 1285
Franz Hall, Box 951563, Los Angeles, CA 90095-1563, or via e-mail:
neisenbe@ucla.edu.
Notes
1. One participant was found through visual inspection to be
a multivariate outlier across all behavioral (self-report) correla-
tional analyses. Further analysis of the standardized residuals and
Cookʼs Distance for each of these behavioral analyses confirmed
that this participantʼs data was disproportionately influential on
the regression line for the majority of these analyses, and this
participant was excluded as a result. Given that this participant
was not an outlier on any single scale, this participant was included
in all neuroimaging analyses, which included only one behavioral
variable in any given regression analysis. It is worth noting, how-
ever, that none of the neuroimaging findings showed any
meaningful differences when this participant was excluded from
analyses.
2. To ensure that the observed correlations with observer-rated
distress were not simply due to individual differences in the
tendency to experience negative affect, we examined correlations
with neuroticism. Participants completed the neuroticism sub-
scale of the Eysenck Personality Questionnaire (EPQ; Eysenck &
Eysenck, 1975). There was no relationship between neuroticism
scores and observer-rated distress (r = −.003, ns), and control-
ling for neuroticism did not change the associations between
observer-rated distress and neural activity during observed ex-
clusion versus inclusion.
REFERENCES
Baumeister, R. F., Faber, J. E., & Wallace, H. M. (1999).
Coping and ego depletion. In C. R. Snyder (Ed.), Coping:
The psychology of what works (pp. 50–60). New York:
Oxford University Press.
Baumeister, R. F., Vohs, K. D., & Tice, D. M. (2007). The
strength model of self-control. Current Directions in
Psychological Science, 16, 351–355.
Branscombe, N. R. (1998). Thinking about oneʼs gender
groupʼs privileges or disadvantages: Consequences for
well-being in women and men. British Journal of Social
Psychology, 37, 167–184.
Crocker, J., & Major, B. (1989). Social stigma and self-esteem:
The self-protective properties of stigma. Psychological
Review, 96, 608–630.
DeWall, C. N., MacDonald, G., Webster, G. D., Masten, C. L.,
Baumeister, R. F., Powell, C., et al. (in press). Tylenol
reduces social pain: Behavioral and neural evidence.
Psychological Science.
Dickerson, S. S., & Kemeny, M. E. (2004). Acute stressors
and cortisol responses: A theoretical integration and
synthesis of laboratory research. Psychological Bulletin,
130, 355–391.
1050
Journal of Cognitive Neuroscience
Volume 23, Number 5
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
5
e
-
1
p
0
d
4
f
2
/
1
2
9
3
4
/
1
5
0
/
4
1
2
0
o
4
c
2
n
/
1
2
0
7
1
7
0
5
2
2
8
1
0
5
2
/
0
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
2
0
0
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
f
/
.
o
n
1
8
M
a
y
2
0
2
1
Dovidio, J. F., Gaertner, S. L., Validzic, A., Matoka, A.,
Johnson, B., & Frazier, S. (1997). Extending the benefits
of recategorization: Evaluations, self-disclosure, and
helping. Journal of Experimental Social Psychology, 33,
401–420.
Eisenberger, N. I., Gable, S. L., & Lieberman, M. D. (2007).
fMRI responses relate to differences in real-world social
experience. Emotion, 7, 745–754.
Eisenberger, N. I., Taylor, S. E., Gable, S. L., Hilmert, C. J.,
& Lieberman, M. D. (2007). Neural pathways link social
support with attenuated neuroendocrine stress responses.
Neuroimage, 35, 1601–1612.
Eisenberger, N. I., Lieberman, M. D., & Williams, K. D. (2003).
Does rejection hurt? An fMRI study of social exclusion.
Science, 302, 290–292.
Eysenck, S. B. G., & Eysenck, H. J. (1975). Manual of the
Eysenck Personality Questionnaire. London: Hodder &
Stoughton.
Goodwin, S. A., Williams, K. D., & Carter-Sowell, A. R. (2010).
The psychological sting of stigma: The costs of attributing
ostracism to racism. Journal of Experimental Social
Psychology, 46, 612–618.
Henry, P. J., & Hardin, C. D. (2006). The contact hypothesis
revisited: Status bias in the reduction of implicit prejudice
in the United States and Lebanon. Psychological Science,
17, 862–868.
Kawakami, K., Phills, C. E., Steele, J. R., & Dovidio, J. F. (2007).
(Close) distance makes the heart grow fonder: Improving
implicit racial attitudes and interracial interactions through
approach behaviors. Journal of Personality and Social
Psychology, 92, 957–971.
Kim, M. J., Somerville, L. H., Johnstone, T., Alexander, A. L.,
& Whalen, P. L. (2003). Inverse amygdala and medial
prefrontal cortex responses to surprised faces.
NeuroReport, 14, 2317–2322.
Krieger, N., & Sydney, S. (1996). Racial discrimination and
blood pressure. The CARDIA study of young black and
white women and men. American Journal of Public
Health, 6, 1370–1378.
Landrine, H., & Klonoff, E. A. (2000). Racial discrimination
and cigarette smoking among blacks: Findings from two
studies. Ethnicity and Disease, 10, 195–202.
Lieberman, M. D., & Cunningham, W. A. (2009). Type I and
Type II error concerns in fMRI research: Re-balancing the
scale. Social Cognitive and Affective Neuroscience, 4,
423–428.
Lieberman, M. D., Jarcho, J. M., Berman, S., Naliboff, B. D.,
Suyenobu, B. Y., Mandelkern, M., et al. (2004). The neural
correlates of placebo effects: A disruption account.
Neuroimage, 22, 447–455.
Major, B., Kaiser, C. R., & McCoy, S. K. (2003). Itʼs not my
fault: When and why attributions to prejudice protect
self-esteem. Personality and Social Psychology Bulletin,
29, 772–781.
Major, B., Quinton, W. J., & McCoy, S. K. (2002). Antecedents
and consequences of attributions to discrimination:
Theoretical and empirical advances. In M. P. Zanna (Ed.),
Advances in experimental social psychology ( Vol. 34,
pp. 251–330). San Diego, CA: Academic Press.
Masten, C. L., Eisenberger, N. I., Borofsky, L. A., Pfeifer, J. H.,
McNealy, K., Mazziotta, J., et al. (2009). Neural correlates
of social exclusion during adolescence: Understanding the
distress of peer rejection. Social Cognitive and Affective
Neuroscience, 4, 143–157.
Mays, V. M., Cochran, S. D., & Barnes, N. W. (2007). Race,
race-based discrimination, and health outcomes among
African American. Annual Reviews of Psychology, 58,
201–225.
Mitchell, J. P., Macrae, C. N., & Banaji, M. R. (2006). Dissociable
medial prefrontal contributions to judgments of similar
and dissimilar others. Neuron, 50, 655–663.
Ochsner, K. N., Bunge, S. A., Gross, J. J., & Gabrieli, J. D.
(2002). Rethinking feelings: An fMRI study of the cognitive
regulation of emotion. Journal of Cognitive Neuroscience,
14, 1215–1229.
Petrovic, P., & Ingvar, M. (2002). Imaging cognitive modulation
of pain processing. Pain, 95, 1–5.
Phelps, E. A., Delgado, M. R., Nearing, K. I., & LeDoux, J. E.
(2004). Extinction learning in humans: Role of the
amygdala and vmPFC. Neuron, 43, 897–905.
Poldrack, R. A. (2006). Can cognitive processes be inferred from
neuroimaging data? Trends in Cognitive Sciences, 10, 59–63.
Redelmeier, D. A., & Kahneman, D. (1996). Patients memories
of painful medial treatments: Real-time and retrospective
evaluations of two minimally invasive procedures. Pain,
66, 3–8.
Schmitt, M. T., & Branscombe, N. R. (2002). The causal loci
of attributions to prejudice. Personality and Social
Psychology Bulletin, 28, 620–628.
Sellers, R. M., & Shelton, J. N. (2003). The role of racial identity
in perceived racial discrimination. Journal of Personality
and Social Psychology, 84, 1079–1092.
Shelton, J. N. (2000). A reconceptualization of how we study
issues of racial prejudice. Personality and Social Psychology
Review, 4, 374–390.
Slavich, G. M., Thornton, T., Torres, L. D., Monroe, S. M., &
Gotlib, I. H. (2009). Targeted rejection predicts hastened
onset of major depression. Journal of Social and Clinical
Psychology, 28, 223–243.
Van Beest, I., & Williams, K. D. (2006). When inclusion
costs and ostracism pays, ostracism still hurts. Journal
of Personality and Social Psychology, 91, 918–928.
Wager, T. D., Rilling, J. K., Smith, E. E., Sokolik, A., Casey, K. L.,
Davidson, R. J., et al. (2004). Placebo-induced changes in
fMRI in the anticipation and experience of pain. Science,
20, 1162–1167.
Way, B. M., Taylor, S. E., & Eisenberger, N. I. (2009). Variation
in the mu-opioid receptor gene (OPRM1) is associated
with dispositional and neural sensitivity to social rejection.
Proceedings of the National Academy of Sciences, U.S.A.,
106, 15079–15084.
Williams, K. D., Cheung, C. K., & Choi, W. (2000). Cyberostracism:
Effects of being ignored over the Internet. Journal of
Personality and Social Psychology, 79, 748–762.
Williams, K. D., Govan, C. L., Croker, V., Tynan, D.,
Cruickshank, M., & Lam, A. (2002). Investigations into
differences between social- and cyberostracism. Group
Dynamics: Theory Research, and Practice. Special Issue:
Groups and Internet, 6, 65–77.
Wirth, J. H., & Williams, K. D. (2009). “They donʼt like our
kind”: Consequences of being ostracized while possessing
a group membership. Group Processes & Intergroup
Relations, 12, 111–127.
Word, C. O., Zanna, M. P., & Cooper, J. (1974). The
nonverbal mediation of self-fulfilling prophecies in
interracial interaction. Journal of Experimental Social
Psychology, 10, 109–120.
Zadro, L., Williams, K. D., & Richardson, R. (2004). How low
can you go? Ostracism by a computer is sufficient to lower
self-reported levels of belonging, control, self-esteem, and
meaningful existence. Journal of Experimental Social
Psychology, 40, 560–567.
Masten, Telzer, and Eisenberger
1051
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
5
e
-
1
p
0
d
4
f
2
/
1
2
9
3
4
/
1
5
0
/
4
1
2
0
o
4
c
2
n
/
1
2
0
7
1
7
0
5
2
2
8
1
0
5
2
/
0
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
2
0
0
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
t
.
.
/
o
n
1
8
M
a
y
2
0
2
1