Action Intention-based and Stimulus Regularity-based
Predictions: Same or Different?
Betina Korka1, Erich Schröger1, and Andreas Widmann1,2
Abstracto
■ We act on the environment to produce desired effects, pero
we also adapt to the environmental demands by learning what
to expect next, based on experience: How do action-based pre-
dictions and sensory predictions relate to each other? We explore
this by implementing a self-generation oddball paradigm, dónde
participants performed random sequences of left and right but-
ton presses to produce frequent standard and rare deviant tones.
By manipulating the action–tone association as well as the like-
lihood of a button press over the other one, we compare ERP
effects evoked by the intention to produce a specific tone, tono
regularity, and both intention and regularity. We show that the
N1b and Tb components of the N1 response are modulated by
violations of tone regularity only. Sin embargo, violations of action
intention as well as of regularity elicit MMN responses, cual
occur similarly in all three conditions. Regardless of whether
the predictions at sensory levels were based on either intention,
regularity, o ambos, the tone deviance was further and equally
well detected at hierarchically higher processing level, as re-
flected in similar P3a effects between conditions. We did not ob-
serve additive prediction errors when intention and regularity
were violated concurrently, suggesting the two integrate despite
presumably having independent generators. Even though they
are often discussed as individual prediction sources in the litera-
tura, this study represents to our knowledge the first to directly
compare them. Finalmente, these results show how, in the context of
acción, our brain can easily switch between top–down intention-
based expectations and bottom–up regularity cues to efficiently
predict future events. ■
INTRODUCCIÓN
Predicting forthcoming sensory input allows us to act effi-
ciently in the environment. According to the predictive
coding theory, the human brain is a probability calculator,
constantly preoccupied with predicting future events
(matar & Pouget, 2004). ERPs can be interpreted as a mea-
sure of prediction error, where attenuated sensory ERPs
indicate smaller prediction errors and thus better pre-
dictions (Friston, 2005). In the auditory domain, several
types of prediction signatures, along with their paradigms
of investigation, are discussed in the literature (Schröger,
Marzecová, & SanMiguel, 2015; Bendixen, SanMiguel, &
Schröger, 2012; abrazos, Desantis, & Waszak, 2012). Two
prominent lines focus on action-based predictions investi-
gated in self-generation paradigms and sensory predic-
tions investigated in variants of the oddball paradigm.
We act to produce desired outcomes in the environ-
mento; according to the ideomotor theory, performing
an action results in an association between the action it-
self and its sensory consequences, and once the associa-
tion has been learned, action selection is determined
postdictively based on its corresponding perceptual conse-
quences (Elsner & Hommel, 2001; Príncipe, 1997). De este modo, nuestro
1University of Leipzig, 2Leibniz Institute for Neurobiology,
Magdeburg, Alemania
© 2019 Instituto de Tecnología de Massachusetts
own actions represent top–down information sources used
to generate predictions. In this context, self-generated
tones (most often via button presses) are commonly
found to elicit attenuated N1 and often P2 ERP responses
in comparison to externally generated but otherwise
identical tones (Horváth, 2015). The frontocentral N1b
and the temporal Tb peak of the T-complex represent
N1 subcomponents that have associated with sensory-
specific predictions, in contrast to the “unspecific” N1
observable with large ISIs (SanMiguel, Todd, & Schröger,
2013; Hari, Kaila, Katila, Tuomisto, & Varpula, 1982). Incluso
though the N1–P2 are often discussed together in self-
generation studies, it has been suggested that the P2
reflects different processes compared with the N1 (Crowley
& Colrain, 2004), which are rather related to processing of
complex tone features (Shahin, Roberts, Pantev, Trainor,
& ross, 2005).
It has been proposed that the intention for action
(rather than the action itself ) is the crucial prediction
input—specifically, Timm and colleagues showed that only
voluntary button presses, in contrast to involuntary ones
induced by TMS, lead N1–P2 attenuation, in comparison
to externally generated tones (Timm, SanMiguel, Keil,
Schröger, & Schönwiesner, 2014). Sin embargo, the self-
external comparison is problematic because it confounds
several processes (for a detailed description, see Hughes
et al., 2012). Hughes and colleagues addressed this problem
Revista de neurociencia cognitiva 31:12, páginas. 1917–1932
https://doi.org/10.1162/jocn_a_01456
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
oh
C
norte
_
a
_
0
1
4
5
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
and further compared self-generated tones, which were
either congruent or incongruent with hand-specific learned
associations and showed that the congruent relative to the
incongruent tones, indeed lead to N1 attenuation (abrazos,
Desantis, & Waszak, 2013).
Not only do we act to change the environment, but we
also adapt to it by learning the relationship between cer-
tain events—we thus know what outcome to expect next
based on probabilities. In this context, tone regularity
represents a source of bottom–up predictive information,
automatically extracted from preceding sensory input. Él
was indeed shown that if the precise identity of the self-
generated tones is not stable between trials, the prediction
effect is reduced or even abolished (SanMiguel, Saupe, &
Schröger, 2013; SanMiguel, Widmann, Bendixen, Trujillo-
Baretto, & Schröger, 2013; Bäß, Jacobsen, & Schröger,
2008). Although less often discussed in the context of ac-
ción, variants of the oddball paradigm investigating sensory
predictions as differences between regularity-violating and
regularity-conforming tones are vast and well established
(for a comprehensive review, see Näätänen, Paavilainen,
Rinne, & Alho, 2007). The main component of interest
in oddball paradigms is the MMN, which represents the
difference between the rare deviant and frequent standard
tones, peaking in between 100 y 250 msec after the
occurrence of the deviancy (see Garrido, Kilner, Esteban,
& Friston, 2009, for a predictive coding interpretation of
the MMN).
Although “motor and sensory predictions may con-
stitute different sources for a single mechanism” (Lange,
2013), they are barely ever integrated into a common per-
perspectiva. This study takes a step forward by considering
intention- and regularity-based predictions as distinct pre-
diction sources for action-related predictions, in the con-
text of self-generated sounds. Específicamente, we first wanted
to determine effects of the violation of the predictions for
a particular sound that were either based on the action
intention (participant intentionally generated a particular
sound as effect of a particular action) or on the presentation
regularity (one of the two sounds was presented more
frequently than the other, without a reliable action–effect
coupling). Segundo, we wanted to see whether bottom–up
regularity-based and top–down intention-based predictions
have additive effects in case of concurrent violations. A
this end, we used a self-generation paradigm where par-
ticipants pressed buttons to produce frequent standard
and infrequent deviant tones, while we manipulated the
action–tone association as well as the likelihood of a but-
ton press over the other one.
The N1 and MMN components have been considered
as the same brain response ( Jääskeläinen et al., 2004),
but also fundamentally different (Näätänen, Jacobsen, &
Winkler, 2005). Además, the incongruency response, encima-
lapping in latency and morphology to the MMN, is yet
another prediction error elicited by incongruent audio-
visual pairs (Pieszek, Widmann, Gruber, & Schröger, 2013;
Widmann, Kujala, Tervaniemi, Kujala, & Schröger, 2004). A
clear distinction between these responses is thus difficult.
Aquí, we use temporal PCA for the ERP analysis, cual, en
contrast to visual inspection, reliably identifies the constitu-
ent wave components, given the complex nature of an ERP
ola (Dien, 2012). We shall therefore focus the analysis on
the obligatory components (negative and positive, como
identified in the data), rather than on the difference wave.
Finalmente, according to the stages of auditory distraction, si
the prediction errors reflected at the N1 and MMN levels
reach a strong enough threshold, a second processing
stage involving an involuntary attention switch occurs, cual
is reflected in the P3a component (Horváth, Winkler, &
Bendixen, 2008; Waszak & Herwig, 2007; Escera, Alho,
Winkler, & Näätänen, 1998; Schröger, 1997). This effect pre-
sumably represents the ERP signature of an orienting re-
sponse following motivationally significant stimuli, como
expectancy-violating deviant tones (Nieuwenhuis, De Geus,
& Aston-Jones, 2011). We test the hypothesis that tone
regularity as well as action intention lead to sensory
prediction effects, as reflected by significant N1/MMN dif-
ferences between self-generated deviants and standards.
If the two prediction sources are additive, this should lead
to larger deviant–standard differences in case of con-
current violations. Note that by looking at the differences
between standard (predicted) and deviant (unpredicted)
tones, we do not measure prediction directly, but we
probe the existence of predictions via the effects of pre-
diction violations. Sin embargo, for reasons of simplifying,
we should regard the effects obtained by prediction
violation as a measure of prediction. Además, we ex-
pected to find P3a enhancement for the deviant as com-
pared with standard tones, provided the prediction errors
elicited at the earlier processing level reach a strong
enough threshold.
MÉTODOS
Participantes
Data were collected from 24 Participantes (10 hombres, significar
age = 23.5 años, age range = 18–32 years), all of whom
gave written informed consent for the study partic-
ipation. All participants reported normal hearing and
normal-to-corrected vision, and none of them had any
history of neurological conditions. All participants were
right-handed, except one left-handed man. None was tak-
ing any prescribed drugs. The ethics committee of the
University of Leipzig, in agreement with the Declaration
of Helsinki, approved the study procedure (code of approval:
465/17-ek). Participants received compensation of either
A8/hr or course credits.
Stimuli and Apparatus
For the whole experiment duration, participants were
seated in a comfortable office chair in an electrically
shielded, double-walled sound booth (Industrial Acoustics
1918
Revista de neurociencia cognitiva
Volumen 31, Número 12
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
oh
C
norte
_
a
_
0
1
4
5
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Compañía). Stimuli were complex sine wave sounds with
a fundamental frequency of 352 Hz (the low tone) y
440 Hz (the high tone), including the second and third
harmonic attenuated by −3 and −6 dB, respectivamente, con
the duration of 100 mseg, including 5-msec rise-and-fall
veces, and presented binaurally over a pair of headphones
(Sennheiser HD 25) at an intensity level of 76 dB SPL. El
two keys participants pressed had dimensions of 6 × 6 cm
and were placed on a desk, in front of them. Visual feed-
back indicated how many times the left versus right keys
need to be pressed, as well as the time interval between
two consecutive key presses (presented in white numbers
on a black screen). This was provided on a 19-in. CRT
monitor (G90fB, ViewSonic, resolution 1024 × 768 píxeles,
refresh rate of 100 Hz), which was placed at a comfortable
watching distance in front of the participant (∼60 cm).
Stimuli were created and presented via the Psychtool-
box 3 (Kleiner et al., 2007), in combination with GNU
Octave Version 4.0.0 (Eaton, bateman, Hauberg, &
Wehbring, 2016), running on Linux OS.
Tarea
Participants pressed the left and right keys using their
left and right index fingers to generate tones, according
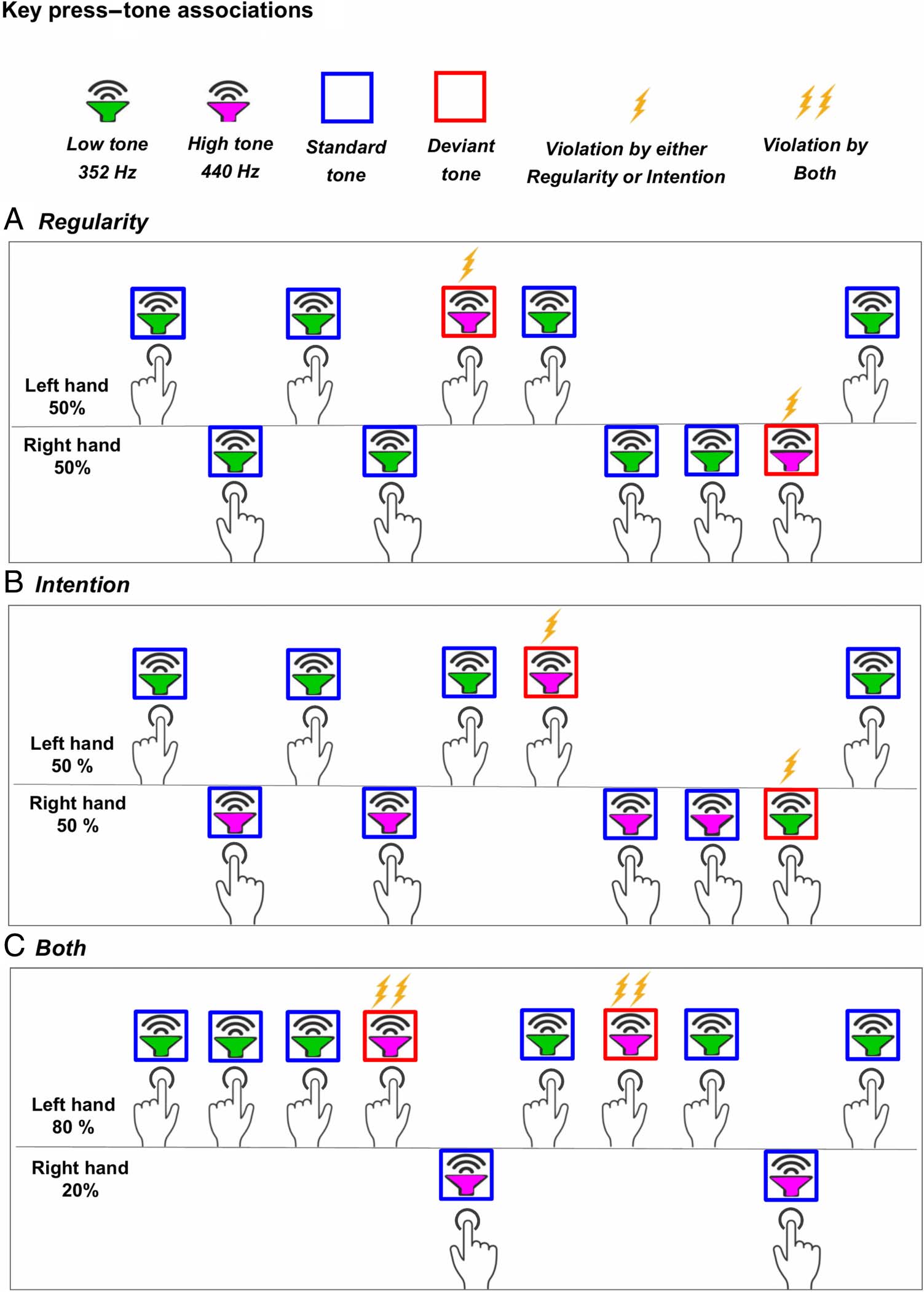
to the condition-specific instructions. Cifra 1 displays a
possible condition-specific representation of key press–tone
asociaciones. For the Regularity condition (Figura 1A),
participants were instructed to press both keys with 50–
50% chances to generate a low tone (which was presented
en 80% of the cases). Rarely, a high tone was presented
en cambio (20% of the cases). For the Intention condition
(Figura 1B), participants’ instructions were to press the
left key in 50% of the cases to generate a low tone and
the right key in 50% of the cases to generate a high tone
(presented with 80% probabilities), while on few occa-
siones, the left key generated a high tone and the right
key generated a low tone instead (20% of the cases).
For the Both condition (Figura 1C), participants were
instructed to press the left key frequently, en 80% del
casos, and the right key rarely, en 20% of the cases, mientras
the key–tone associations were the same as in the
Intention condition. For all conditions, participants were
made aware that, sometimes, another tone than the
expected one will be presented and told to ignore it if that
was the case and proceed normally to the next button
prensa, as we wanted to avoid the possibility that par-
ticipants stop throughout the block and report that some-
thing unexpected happened. Note that the key–tone
associations as well as the frequently pressed key in
Both condition have been counterbalanced and the
above-described mappings reflect only one possibility.
In all three conditions, participants’ task was to press a
key every approximately 1200 msec while, primero, following
the condition-specific instructions (press 50–50% or
80–20%) y, segundo, avoid producing fixed left/right
patterns of key presses (es decir., press the keys in a “random”
secuencia).
Experimental Procedure
One session consisted of 10 experimental blocks. The du-
ration of one block was about 3 mín., and participants
could take self-paced breaks in between. Three shorter
practice blocks were completed at the beginning of every
condición (blocks corresponding to the same condition
were run one after another). The condition order as well
as the key–tone associations and frequent versus rare key
presses were counterbalanced between participants.1
Two constraints were followed: primero, Regularity and the
frequently pressed key in Both generated the same stan-
dard tone, y segundo, the same key–tone association
was kept between Intention and Both. We thus insured
that there were no conflicting associations between con-
ditions. One complete experimental session lasted for
acerca de 45 mín..
The tone onset immediately followed the key press
(with a delay of ∼5 msec, due to technical limitations).
Trial duration was about 1200 mseg, and participants
were instructed to fixate on a fixation cross for the whole
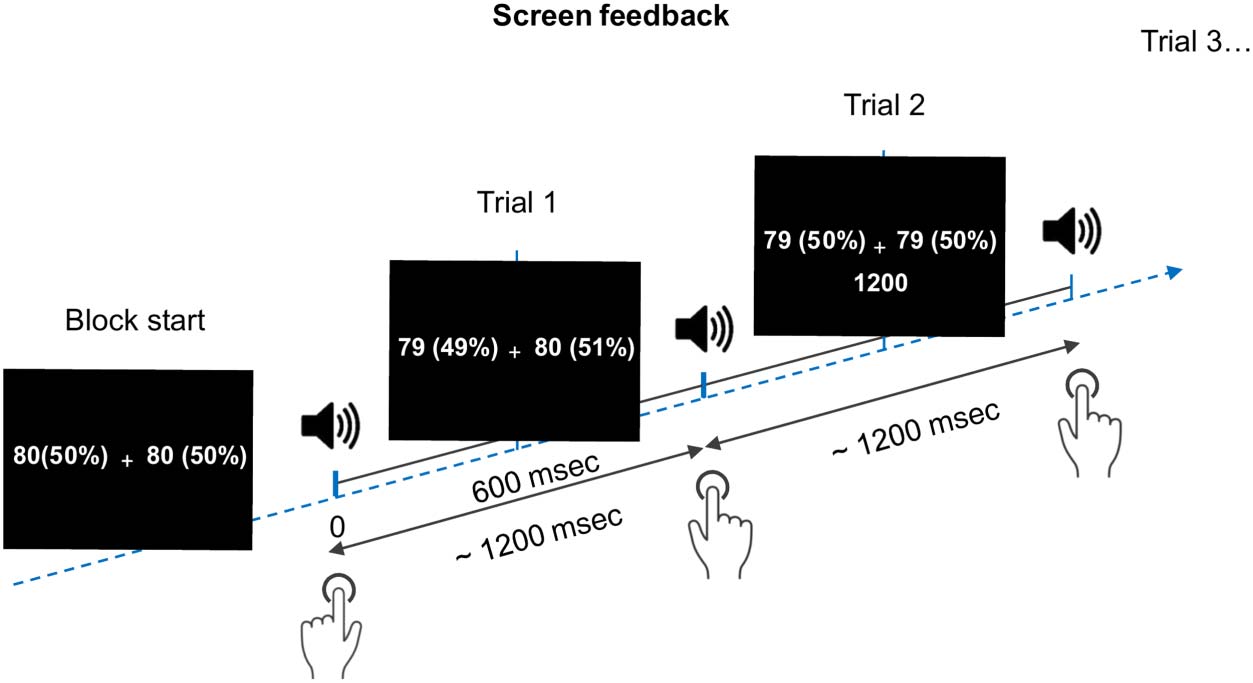
block duration. Cifra 2 illustrates a possible sequence of
trials at the start of a block, including screen feedback.
The screen feedback was designed to help participants
press the keys with equal (in Regularity and Intention)
or unequal (Ambos) chances and to press a key about every
1200 mseg. Each trial began with an indication of how
many times the left versus right keys need to be pressed,
displayed at the left and right sides of the fixation cross.
This was indicated in numbers as well as in percentages
(es decir., participants could see, Por ejemplo, that the left key
needs to be pressed 80 veces, que representa 50% de
the total number of key presses left for that block). El
numbers on the left referred to the left hand and vice
versa. Underneath the fixation cross and starting from
the second trial, the timing between two consecutive
key presses was displayed in milliseconds. Timing errors
were defined as intervals shorter or longer by more than
400 msec than the indicated time (1200 mseg). If a timing
error occurred, a corresponding error message (“Too
short/Too long”) was displayed on the screen, en cambio
of the timing between the key presses—the tone was
not presented in trials containing such errors, following
which participants proceeded normally to the next but-
ton press. Note that because a tone was not presented,
timing errors did not affect the total number of collected
trials for the standard or deviant tones. Although the fixation
cross was presented for the whole duration of one block,
the screen feedback was updating every trial 600 mseg
after tone onset.
To have an indication of task compliance, we recorded
timing errors (es decir., ±400 relative to 1200 mseg) and fixed
left–right sequences. The left–right sequences were ana-
lyzed online using the Walsh–Hadamard randomness test
Korka, Schröger, and Widmann
1919
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
oh
C
norte
_
a
_
0
1
4
5
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. An example of
condition-specific key press–
tone associations. En el
Regularity condition (arriba),
participants pressed the two
buttons with 50–50% chances to
generate a standard low tone
(marked in green) con 80%
probabilidad (marked in blue
square) and a deviant high tone
(marked in pink) con 20%
probabilidad (marked in red
square). In the Intention
condición (middle), Participantes
pressed the left button in 50%
of the cases to generate a
standard low tone and a deviant
high tone and conversely the
right button in 50% of the cases
to generate a standard high
tone and a deviant low tone. En
the Both condition, the key
press–tone associations were
the same as in Intention,
whereas the left key was
pressed frequently (en 80% de
the cases) and the right key
rarely (en 20% of the cases). Uno
lightning symbol marks tones
that violate either regularity
(arriba) or intention (middle).
Two lightning symbols mark
tones that violate both
regularity and intention
(abajo).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
oh
C
norte
_
a
_
0
1
4
5
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(Oprina, Popescu, Simion, & Simion, 2009), programmed
in Octave within the experiment. The test uses a sequence
of binary input (aquí, codes for left vs. right key presses)
and detects randomness failure based on autocorrelation.
If excessive fixed patterns were detected, participants were
verbally warned at the end of the block. Note that measur-
ing “pure” randomness was beyond our scope, because it is
controversial whether humans can produce completely ran-
dom sequences of actions, one major difficulty being the
very definition of mathematical randomness (Wagenaar,
1972). This was rather implemented to make sure partici-
pants press the two keys equally often (or one key four
times more often than the other one), without repeating
a certain sequence excessively. The timing errors were
analyzed offline as percentage errors from the total
number of trials.
The Regularity condition contained 384 standard
tones (80%) y 96 deviant tones (20%)—these were
grouped in three blocks, each containing 160 ensayos, de
cual 80 corresponded to the left and 80 to the right
key. In Intention, 192 high and 192 low tones inversely
associated with the left and right keys were generated as
1920
Revista de neurociencia cognitiva
Volumen 31, Número 12
Cifra 2. A possible sequence
of trials. Every key press
generates a tone (presentado
with a delay of ∼5 msec),
followed by the screen feedback
updating every 600 mseg después
tone onset. Participants fix their
gaze on the fixation cross and
press a key of their choice every
acerca de 1200 mseg. The screen
feedback indicates how many
times the left versus right
buttons need to be pressed in
numbers and in percentages,
and beginning with the second
trial, the time interval between
two consecutive key presses.
Regularity and Intention blocks
start with 80 trials for each of
the left and right keys (50–50
chances). Both blocks start with
120 trials for the frequently
pressed key and 30 trials for the
rarely pressed key (80–20 chances).
standards (80% probabilidad), y 48 low and 48 alto
tones inversely associated with the left and right keys
were generated as deviants (20% probabilidad). As in
Regularity, these were presented in three blocks, cada
de 160 ensayos, of which 80 corresponded to the left and
80 to the right key. In Both, the frequently pressed key
(80% of the cases) generated 384 standard tones (80%
probabilidad) y el 96 deviant tones (20% probabilidad).
The rarely pressed key (20% of the cases) generated 96
standard tones (80% probabilidad) y 24 deviant tones
(20% probabilidad). These were presented in four con-
secutive blocks, each containing 150 ensayos, of which
120 trials corresponded to the frequent and 30 trials cor-
responded to the rare key. The standard–deviant se-
quence of tones within a block (and within the same
key for Intention and Both) were randomized, con
the constraint that the first two tones were always
standards. Note that for the Both condition, nosotros sólo
analyzed the trials corresponding to the frequently
pressed key. De este modo, we recorded an equal number of
trials for all three conditions: 384 standards versus 96
deviants.
EEG Data Recording
EEG data were continuously recorded at a sampling rate of
500 Hz with a system equipped with 64 Ag–AgCl active elec-
trodes, using a BrainAmp amplifier and the Vision Recorder
software (Cerebro Productos GmbH, Munich, Alemania). Fifty-
eight electrodes were mounted in an elastic cap (actiCAP)
following the extended international 10–20 system (Chatrian,
Lettich, & nelson, 1985). Two additional electrodes were
placed on the mastoids. One electrode placed on the
tip of the nose served as online reference, a ground elec-
trode was placed on the forehead, and three elec-
trodes were used to record EOG activity, two of which
were placed on the left and right outer canthi and one
below the left eye.
EEG Preprocessing
The preprocessing was carried out in three steps using
the EEGLAB MATLAB-based software (Delorme &
Makeig, 2004). Primero, data were filtered using a 0.1-Hz
high-pass and 45-Hz low-pass windowed sinc finite
impulse response filter (Hamming window, filter order
8250 [high pass] y 166 [low pass]), in accordance with
the recommendations of Widmann, Schröger, and Maess
(2015). De término medio, 1.29 channels containing extreme
amplitudes were removed using a deviation criterion that
“calculates the robust z score of the robust standard
deviation for each channel” (Bigdely-Shamlo, Mullen,
Kothe, Su, & robbins, 2015). Data were then epoched
around the tone presentation (−200 to 600 mseg).
Epochs with amplitudes exceeding a 600-μV Delta thresh-
old were removed. Segundo, an independent component
analysis was computed on the raw data, which were first
filtered with a 1-Hz high-pass and 45-Hz low-pass filter,
epoched (−200 to 600 msec relative to tone presentation)
and cleaned by removing the same bad channels and epochs
detected at the first step. The obtained weights were stored
and transferred to the data sets obtained at the first step.
Tercero, the removal of components containing eye-related
(parpadea, lateral eye movements) and muscle artifacts
was done by visual inspection and based on measures
computed with FASTER (Nolan, Whelan, & Reilly, 2010),
ADJUST (Mognon, Jovicich, & Bruzzone, 2011), y
SASICA (Chaumon, obispo, & Busch, 2015).2 The missing
channels were interpolated using the built-in EEGLAB
spherical interpolation function, and data were baseline
Korka, Schröger, and Widmann
1921
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
oh
C
norte
_
a
_
0
1
4
5
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
corrected using the 200-msec prestimulus interval. Epochs
with amplitudes still exceeding a 200-μV Delta threshold
after the independent component analysis cleaning were
removed—epochs removed at both the first and third
steps represented less than 1% from the total number of
ensayos. Finalmente, condition-specific averages were calculated.
PCA Analysis
A temporal PCA was performed using the ERP PCA toolkit
MATLAB-based toolbox (Dien, 2010). We computed the
PCA on the individual averages of all conditions using
Promax rotation (k = 3) with a covariance relationship
matrix and Kaiser weighting. Horn’s parallel test was used
to determine the number of components to be retained.
Statistical Analysis
Each component of interest identified by temporal PCA
was separately tested using first frequentist and second
Bayesian repeated-measures ANOVA (rANOVA); the anal-
yses were conducted using IBM SPSS Statistics 25 y
JASP 0.9.1.0, respectivamente. Note that for the Intention
condición, standards from the left and right hands were
pooled together for the analyses (irrespective of whether
they were low- or high-pitched)—similarly, the deviants
from both hands were analyzed together. For the Both
condición, only tones generated by the frequently
pressed key were analyzed. A 3 × 2 design with factors
Condition (Regularity vs. Intention vs. Ambos) × Stimulus
(standard vs. deviant) was used for the frequentist anal-
ysis. Statistical significance was defined at the .05 alfa
nivel, and results are reported including the eta-square
effect sizes (η2). Follow-up t tests were computed for sta-
tistically significant interactions. The complementary 3 ×
2 Bayesian analysis was calculated to test all alternative
modelos, including main effects and interactions against
the null model, which included only the random factor,
eso es, participants’ variation. The Bayes factor (BF10)
was calculated using 10.000 sample repetitions; the null
hypothesis corresponded to a standardized effect size δ =
0, and the alternative hypothesis was defined as a Cauchy
prior distribution centered around 0 (Rojo, Morey,
Speckman, & Province, 2012). Bayesian t tests followed
up on the effects of the models including interactions,
provided these supported the alternative hypothesis best.
Por último, BFInclusion calculated across matched models (es decir.,
models that include vs. do not include the effect) pro-
vided a measure of change odds from prior to posterior
distributions. These were only calculated if more than
one model supported the alternative hypothesis to have
clear evidence whether the main effects or the interaction
explain the data best. In accord with existing recom-
mendations on how to interpret the Bayes factor (Sotavento &
carpinteros, 2014; Jeffreys, 1961), values ≤0.3 were tak-
en as evidence in favor of the null hypothesis, values ≥3 as
evidence in favor of the alternative hypothesis, mientras
values close to 1 were considered poor evidence.
Finalmente, we checked by means of t tests the Regularity +
Intention versus Both additivity model (p.ej., see Pieszek
et al., 2013, for a similar procedure). For this purpose, el
component scores representing the differences between de-
viants and standards (deviant − standard) were calculated for
every condition, the difference scores for Regularity and
Intention being subsequently added together. Note that the
additivity model was only tested if sensory prediction error
efectos (significant differences between deviants and stan-
dards) were found in all three conditions and only for those
sensory components in which such effects were found.
RESULTADOS
Timing Errors
The key press-to-key press time intervals longer or shorter
than 1200 msec by more than 400 msec were recorded as
timing errors. We calculated these as error percentages (%
ERR) relative to the total number of trials contained in
every block—blocks corresponding to the same condition
were subsequently averaged. Participants made on average
2.76 %ERR in Regularity (DE = 3.51, range = 0–12.29),
3.29 %ERR in Intention (DE = 3.40, range = 0–11.88),
y 2.38 %ERR in Both (DE = 3.28, range = 0–10)—these
indicate they followed the instructions and pressed the
keys at the suggested pace. A one-way ANOVA including
the three conditions (Regularity, Intention, Ambos) revealed
no significant Condition differences, F(2, 46) = 2.78, pag =
.114, η2 =.090. Committed errors do not indicate fewer
“correct” trials for the analysis of ERPs, because for the tri-
als containing timing errors, no tone was generated.
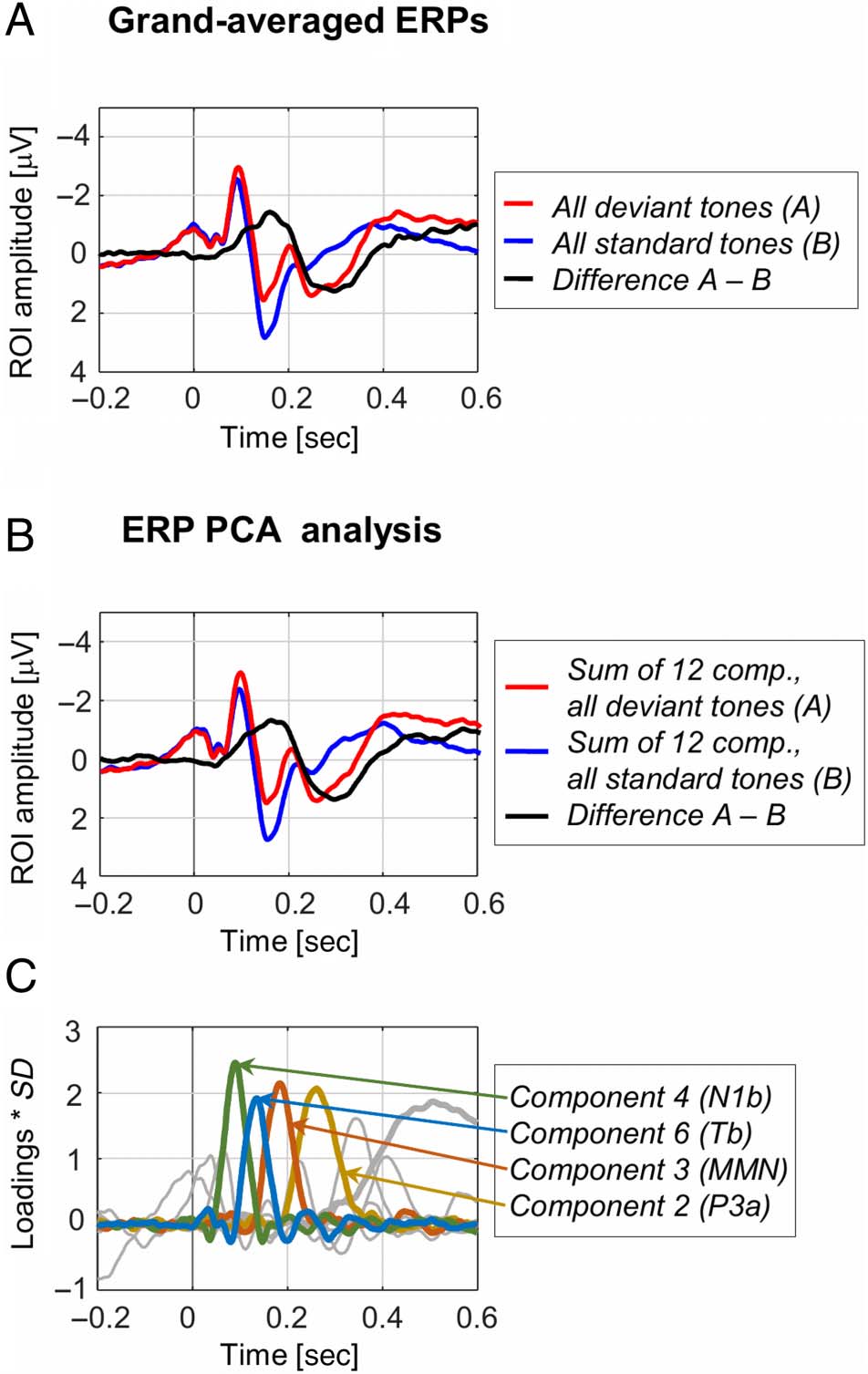
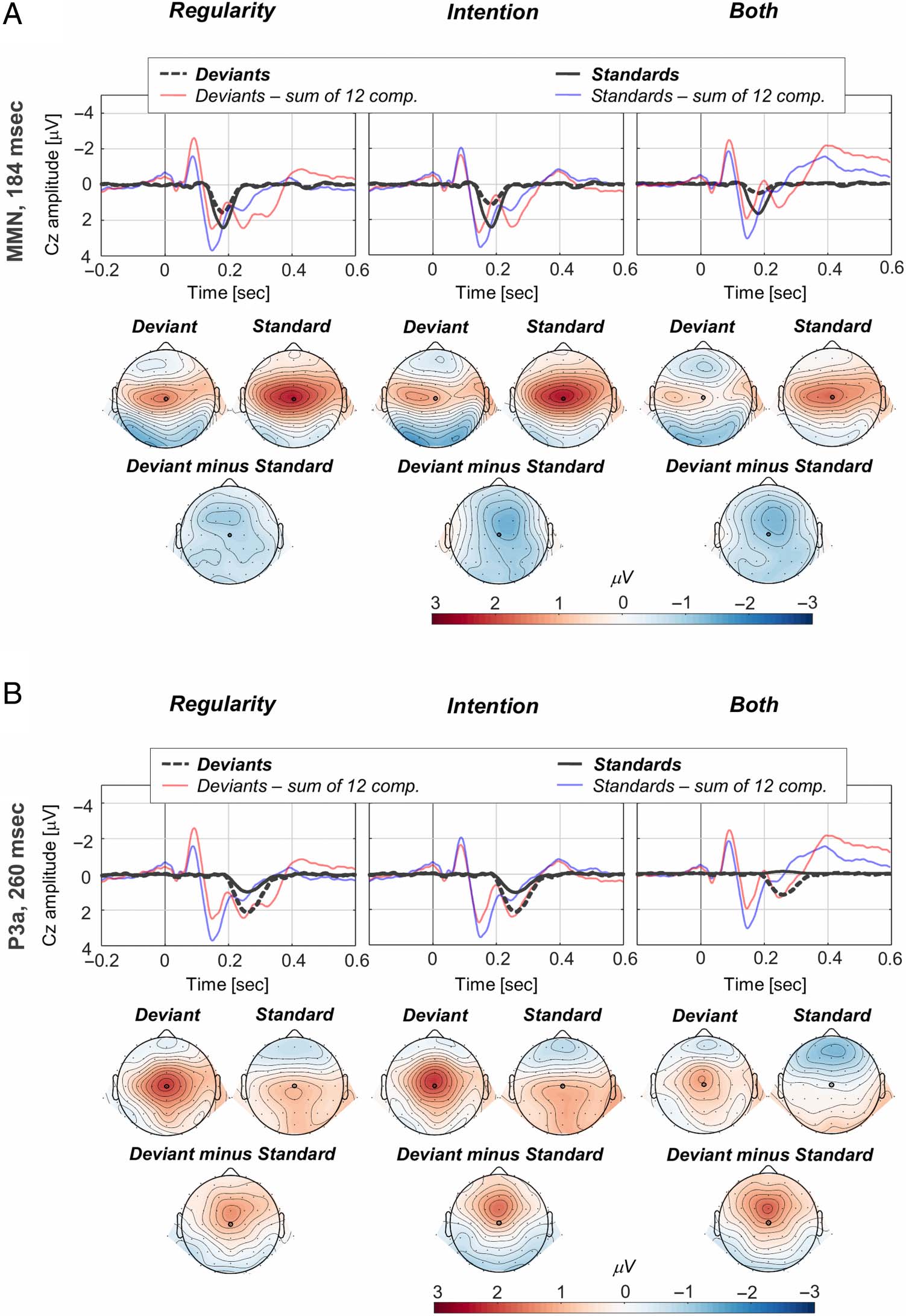
ERP PCA Results
Cifra 3 displays the grand-averaged ERPs, junto con el
PCA results. According to Horn’s parallel test, 12 compo-
nents were extracted explaining over 95% of the total
epoch variability (Figura 3C). The stimulus-specific waves
represented by the sum of the 12 retained components
firmly correspond to the stimulus-specific grand-averaged
ERPs—see Figure 3A–B for a visual comparison at the
level of an ROI representing an average of Fz, FCz, y
Cz, electrodes that typically are of interest in auditory pro-
cesando (p.ej., see results in Timm et al., 2014; abrazos
et al., 2013; SanMiguel, Saupe, et al., 2013; Horváth
et al., 2008; Näätänen et al., 2005). Of the retained 12,
we focused our attention on four, Components 2, 3, 4,
y 6, presumably representing the P3a, MMN, N1b, y
Tb peaks, respectivamente. The selection of the four compo-
nents of interest was based on latency and topographical
información. Note that they are ordered not by chronolog-
ical peak latency but by the explained variance. Eso es,
Component 2 corresponding to the P3a peak at 260 mseg
explains ∼15.2% of the epoch variability. Component 3
1922
Revista de neurociencia cognitiva
Volumen 31, Número 12
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
oh
C
norte
_
a
_
0
1
4
5
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Näätänen & Picton, 1987). Component 1 (not analyzed here
but marked in the thicker gray line in Figure 3C) explains
about half of the whole epoch variability (51.3%) and peaks
en 506 msec—this presumably represents the reorienting
negativity or N3 peak (Kotchoubey & Pavlov, 2019). El
time-invariant component scores represent the contribu-
tion of each component of interest to the ERP wave—these
have been subjected to statistical analyses. The time-variant
loadings of the components reflect their contribution to the
voltage maps at each point in time.
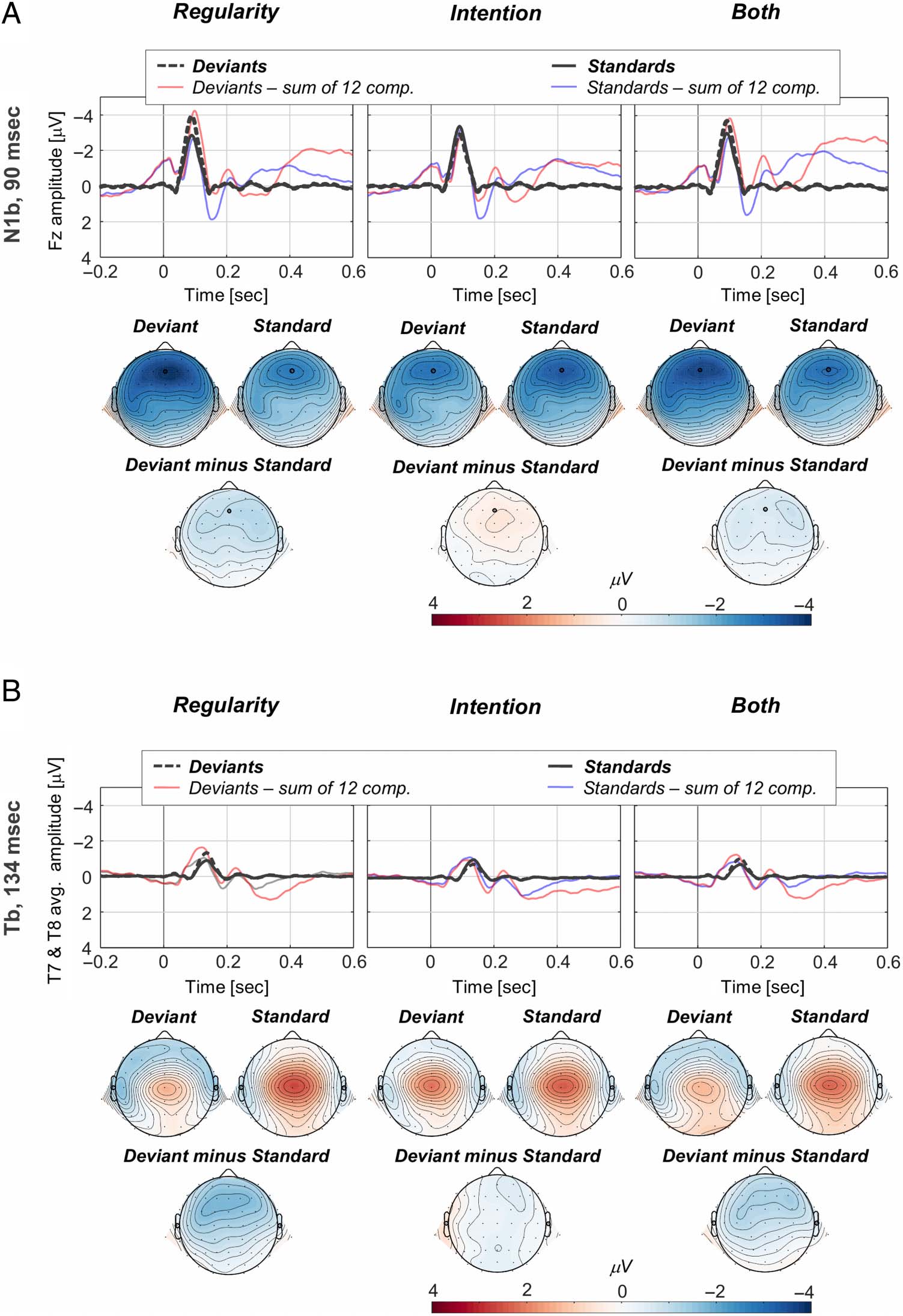
For each component of interest, we analyzed component
scores at the electrodes showing the largest score activations.
Correspondingly, the N1 scores were analyzed at electrode
Fz. The Tb has a bilateral distribution peaking around the
T7 and T8 temporal electrodes—we thus analyzed the aver-
age of the component scores corresponding to the two elec-
trodes. Finalmente, the scores corresponding to MMN and P3a
components were analyzed at electrode Cz. Figures 4 y 5
display for each component the condition-specific activations
for the standard and deviant tones, along with the corre-
sponding topographical maps (N1b, Figura 4A; Tb,
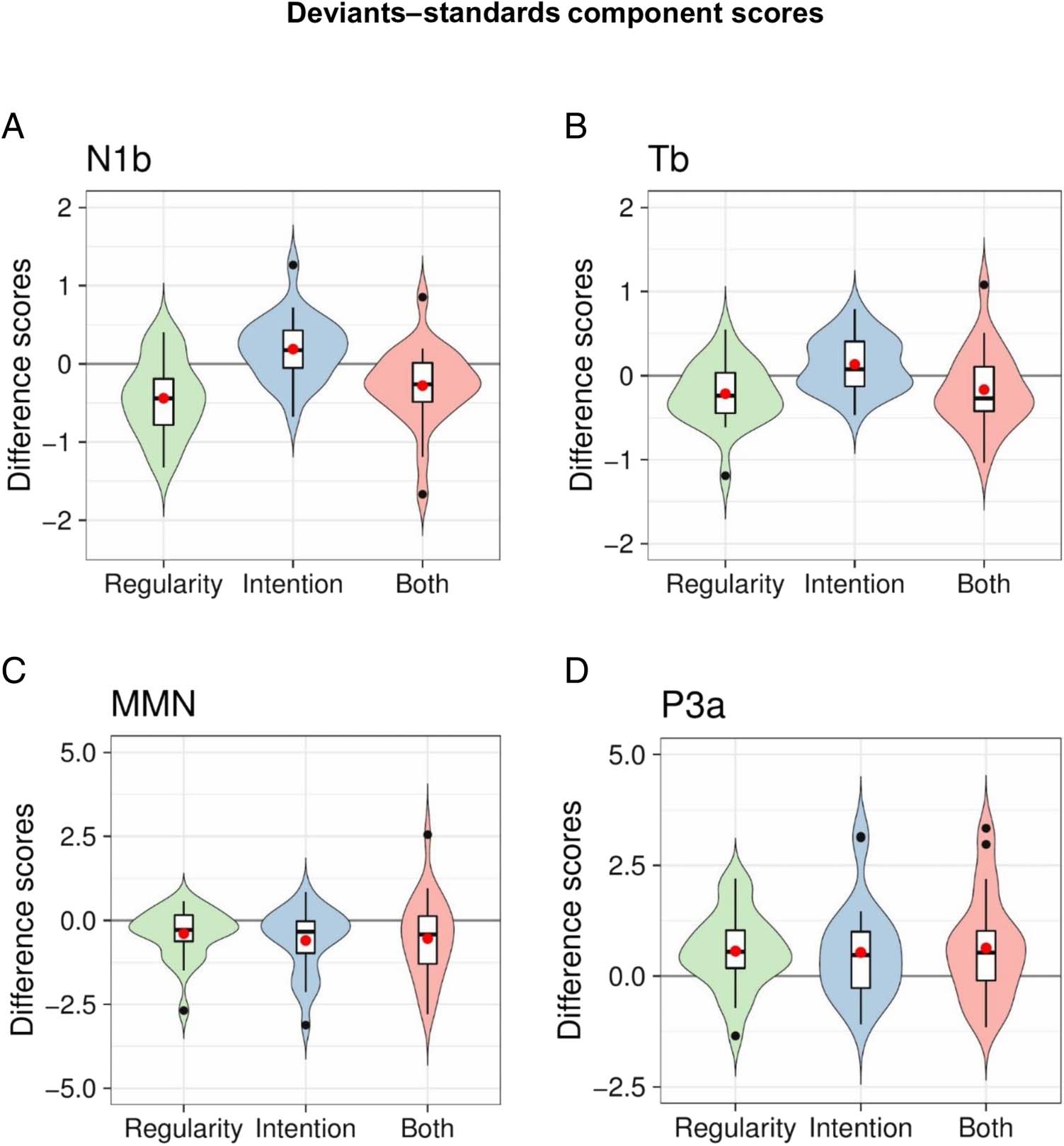
Figura 4B; MMN, Figure 5A; P3a, Figure 5B). Violin plots in
Cifra 6 display the condition-specific effects (es decir., com-
ponent scores plotted as deviant–standard differences)
for each of the four components (N1b, Figure 6A; Tb,
Figure 6B; MMN, Figure 6C; P3a, Figure 6D). We further
report component-specific statistical results. Main ef-
fects and interactions obtained in the frequentist versus
Bayesian analyses are summarized in Table 1.
N1b Enhancement for Deviants in Regularity and Both,
but Not in Intention
The frequentist rANOVA revealed a main effect of Stimulus,
F(1, 23) = 5.92, pag = .023, η2 = .205, and an interaction of
Condition × Stimulus, F(2, 46) = 18.86, pag < .001, η2 =
.451. Follow-up t tests indicate the N1b component scores
are significantly enhanced for the deviant tones in Regu-
larity, t(23) = 4.39, p < .001, and Both, t(23) = 2.76, p =
.011, but not in Intention, where, in contrast, the scores for
the standard tones are enhanced, t(23) = −2.26, p = .034.
The Bayesian rANOVA favored the full model, including
the main effects and the interaction term (Condition +
Stimulus + Condition × Stimulus, BF10 = 3.61 ± 1.96%;
see Table 1), whereas the models containing main effects
of Condition and Stimulus only provided anecdotal evi-
dence. Follow-up Bayesian t tests mirrored the frequentist
results by providing strong evidence for the alternative hy-
pothesis in Regularity (BF10 = 138.77 ± <0.001%), moder-
ate in Both (BF10 = 4.47 ± < 0.001%), and only anecdotal
evidence in Intention (BF10 = 1.78 ± 0.005%).
Tb Enhancement for Deviants in Regularity, but Not in
Intention and Both
The frequentist rANOVA revealed a significant main effect
of Stimulus, F(1, 23) = 5.44, p = .029, η2 = .191, and an
Korka, Schröger, and Widmann
1923
Figure 3. ERP PCA results. (A) Grand-averaged ERPs are presented
for the deviant (red) and standard (blue) tones, along with the
difference wave (black), averaged across all three conditions, for a ROI
composed of the electrodes Fz, FCz, and Cz. (B) Following the PCA
analysis, 12 components explaining over 95% of the epoch variability
were retained—the waves representing the sum of these 12
components for the deviant (red) and standard (blue) tones, as well as
the difference wave (black), firmly correspond to the original grand-
averaged ERPs. (C) The 12 retained components are presented
individually. Of these, Components 2, 3, 4, and 6 corresponding to the
P3, MMN, N1b, and Tb peaks, respectively, were further analyzed—
these are marked in color. Component 1, marked in the thicker gray
line, is related to the reorienting negativity or N3.
corresponding to the MMN response reflecting an increase
in negativity for the deviant compared with standard tones
peaking at 184 msec (P2 range) explains ∼9.7% of the
epoch variability. Component 4 representing the sensory-
specific N1b peak at 90 msec and Component 6 repre-
senting the Tb peak (corresponding to the T-complex)
at 134 msec explain ∼4.6% and ∼2.6% of the epoch var-
iability, respectively. Note that the identified time courses
and topographies (i.e., early latency and frontocentral
distribution for N1b, later latency, and temporal distribu-
tion for Tb) correspond to previous studies reporting N1-
constituent components (SanMiguel, Todd, et al., 2013;
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. N1b and Tb PCA
results. (A) The N1b component
peaks at 90 msec and is largest
at the Fz electrode. (B) The Tb
component peaks at 134 msec
and is largest over temporal T7
and T8 electrodes, the displayed
waves representing a mean of
the two. For all three
conditions, the component-
specific waves for the standards
(dark full lines) and deviants
(dark dashed lines) are
displayed along with the
“reconstruction waves,”
representing the sum of the 12
retained components for the
deviants (faded red lines) and
standards (faded blue lines).
The topographical maps have
been calculated based on
spherical spline interpolation
and illustrate the deviants and
standards evoked responses, as
well as the deviants–standards
difference maps. The electrodes
marked on the topographical
maps (N1b→Fz, Tb→T7, and
T8) represent the ones included
in the analysis.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
interaction of Condition × Stimulus, F(2, 46) = 3.33, p =
.044, η2 = .127. Follow-up t tests indicate the Tb compo-
nent scores are significantly enhanced for the deviant
tones in Regularity, t(23) = 2.92, p = .008, whereas in
Both, a nonsignificant trend was observed, t(23) =
1.77, p = .089. In Intention, the difference between the
standard and deviant tones was not significant, t(23) =
0.25, p = .798. The Bayesian rANOVA favored the model
containing the main effects (Condition + Stimulus, BF10 =
3.22 ± 1.36%; see Table 1), whereas all other models,
including the one with the interaction term, only provided
anecdotal evidence. The Bayesian analysis therefore did
not bring conclusive evidence in favor or against the
alternative hypothesis containing the interaction term
(BF10 = 0.96 ± 3.45%).
MMN in Regularity, Intention, and Both
The frequentist rANOVA revealed a main effect of Stimu-
lus, F(1, 23) = 11.24, p = .003, η2 = .328, with smaller
1924
Journal of Cognitive Neuroscience
Volume 31, Number 12
Figure 5. MMN and P3a PCA
results. The MMN and P3a
components are largest at
electrode Cz. MMN peaks at
184 msec (A), and the P3a peaks
at 260 msec (B). For all three
conditions, the component-
specific waves for the standards
(dark full lines) and deviants
(dark dashed lines) are
displayed along with the
“reconstruction waves”
representing the sum of the 12
retained components for the
deviants (faded red lines) and
standards (faded blue lines).
The topographical maps have
been calculated based on
spherical spline interpolation
and illustrate the deviants and
standards evoked responses, as
well as the deviants–standards
difference maps. The electrode
marked on the topographical
maps (Cz) represents the one
included in the analysis.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
positive scores for the deviant (M = 0.50) as compared
with the standard (M = 1.01) tones. A trend approaching
significance was observed as a main effect of Condition,
F(2, 46) = 3.10, p = .054, η2 = .119, but no significant
interaction term was observed, F(2, 46) = 0.43, p = .651,
η2 = .018; the presence of each of the three MMNs was
confirmed by paired t tests: Intention, t(23) = 3.06, p =
.006; Regularity, t(23) = 2.63, p = .015; Both, t(23) =
2.19, p = .038). Thus, the MMN is present across all three
conditions, but not significantly different between condi-
tions. The Bayesian rANOVA favored the model containing
the main effects of condition and stimulus (Condition +
Stimulus, BF10 = 316.99 ± 1.24%; see Table 1), whereas
the models containing the main effect of stimulus and
the full model also brought evidence in favor of the alter-
native hypothesis. However, averaged across the matched
models, the BFInclusion suggests only the main effect of
Stimulus should be retained (BFInclusion = 246.06), whereas
the main effects of Condition and the interaction only pro-
vided anecdotal evidence for the alternative hypothesis or
Korka, Schröger, and Widmann
1925
Figure 6. Condition-specific
effects: deviants–standards.
Violin plots display the
condition-specific deviant–
standard component scores for
N1b (A), Tb (B), MMN (C), and
P3a (D) components. The
estimated density distributions
(displayed in green for
Regularity, blue for Intention,
and red for Both) are shown
along with boxplots indicating
the medians, interquartile
ranges, and confidence
intervals, whereas the means
are displayed in red dots. The
black dots represent individual
data points falling outside the
confidence intervals. Note that
the scores are component-
specific and the scales between
A and D do not correspond to
each other.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
moderate evidence for the null hypothesis, respectively
(Condition: BFInclusion = 1.59; Condition × Stimulus:
BFInclusion = 0.14). Therefore, mirroring the frequentist re-
sults, this indicates that there are no condition differences
for the MMN component.
P3a Enhancement for Deviants in Regularity, Intention,
and Both
The frequentist rANOVA revealed a significant main effect
of Stimulus, F(1, 23) = 12.14, p = .002 η2 = .346, with
larger positive scores for the deviant (M = 0.87) as com-
pared with standard (M = 0.30) tones, and a significant
main effect of Condition, F(2, 46) = 6.64, p = .003, η2 =
.224, with larger positive values for Intention (M = 0.76),
followed by Regularity (M = 0.74) and Both (M = 0.26).
No significant interaction term was observed, F(2, 46) =
0.09, p = .908, η2 = .004, suggesting the P3a enhancement
effect is present across all three conditions, but not signifi-
cantly different between conditions. The Bayesian rANOVA
favored the model containing the main effects of Condition
and Stimulus (Condition + Stimulus, BF10 = 18231.89 ±
0.95%; see Table 1), whereas all other models containing
the main effect of Condition and the main effect of Stimu-
lus, as well as the full model, also brought evidence in favor
of the alternative hypothesis. However, averaged across the
matched models, the BFInclusion suggests the models con-
taining the main effects of Condition (BFInclusion = 23.17)
and stimulus (BFInclusion = 1719.02) should be retained,
whereas the interaction provided moderate evidence for
the null hypothesis (BFInclusion = 0.12). Again, mirroring
the frequentist results, this suggests that there are no
condition differences for the P3a component.
No Additivity of Regularity and Intention Effects
We calculated the Regularity + Intention versus Both ad-
ditivity model for the MMN component, where significant
prediction errors were observed in all three conditions.
The Regularity + Intention difference scores do not
equal the Both difference scores (i.e., are significantly dif-
ferent, t(23) = −2.08, p = .048), with the former being
1926
Journal of Cognitive Neuroscience
Volume 31, Number 12
BF10
± %ERR
Table 1. Results of Statistical Analyses
Frequentist Effects
Bayesian Models
Comp.
N1b
Cond
Stim
F
0.40
5.92
p
.669
.023
η 2
.017
.205
Cond
Stim
Cond + Stim
Cond × Stim
18.86
<.001
.451
Cond + Stim + Cond × Stim
Tb
Cond
Stim
2.38
5.44
.104
.029
.094
.191
Cond
Stim
Cond × Stim
3.33
.044
.127
Cond + Stim + Cond × Stim
Cond + Stim
MMN
Cond
Stim
3.10
11.24
.054
.003
.119
.328
Cond
Stim
Cond × Stim
0.43
.651
.018
Cond + Stim + Cond × Stim
Cond + Stim
P3a
Cond
Stim
6.64
12.14
.003
.002
.224
.346
Cond
Stim
Cond + Stim
0.11
1.79
0.21
3.61
1.54
1.81
3.22
0.96
1.09
199.07
316.99
46.24
10.06
786.12
18231.89
0.70
0.85
2.09
1.96
0.87
1.21
1.36
3.45
0.65
1.05
1.24
2.74
0.50
0.79
0.95
1.69
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
Cond × Stim
0.09
.908
.004
Cond + Stim + Cond × Stim
2285.46
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
For all analyzed components, the frequentist main effects and interactions with their corresponding p values and effect sizes are displayed (left), along with
the Bayes factors and corresponding errors for the full models including main effects and interactions (right). The significant main effects and interactions
from the frequentist analyses as well as the models with the best explanatory power from the Bayesian analysis are highlighted in bold.
more negative (i.e., larger effects, M = −0.99) than the
latter (M = −0.54). Thus, an additivity model does not
suit these data.
DISCUSSION
Action predictions based on action intention and sensory
predictions based on tone regularity are often described as
similar mechanisms, but a direct comparison is yet miss-
ing. We addressed this issue by using a “self-generation
oddball paradigm,” where participants produced standard
and deviant tones (high or low pitched) by performing
“random” sequences of left and right button presses. By
manipulating the action–tone association as well as the
likelihood of performing one action over the other, we
contrast predictions based on tone regularity versus inten-
tion to produce a specific tone versus both intention and
regularity. Our results indicate that the N1b and Tb com-
ponents of the N1 response are modulated by regularity
violations, but not by intention violations. Intention and
regularity violations are reflected in the MMN response,
which importantly is elicited with and without high global
probability of the standard tone. Even though regularity and
intention might represent independent generative sources,
their resulting prediction errors seem to integrate, rather
than add up—that is, we did not observe stronger effects
(indicating additivity) when the two were present together.
Finally, similar P3a effects for all conditions suggest that, re-
gardless of whether the sensory predictions are imple-
mented based on either tone regularity, motor intention,
or both, this does not influence the deviance detection
mechanism further implemented at the next processing
step. As follows, we discuss these findings in more detail.
Bottom–Up Regularity-based Prediction Errors Are
Reflected in the N1 and MMN Components
The use of temporal PCA has led to the distinct identifi-
cation of the N1b and Tb N1-constituent components, as
well as of the MMN component. We found N1b enhance-
ment effects for the deviant relative to the standard tones
in the Regularity and Both conditions, but not in the
Intention condition—this was supported by frequentist
Korka, Schröger, and Widmann
1927
as well as Bayesian analyses. Additionally, we found Tb
enhancement for the deviant relative to the standard
tones in the Regularity condition only. For the MMN com-
ponent, the typical enhancement for the standard relative
to the deviant tones was observed (peaking at P2 latency
range), leading to negativity responses for the deviant–
standard evoked tones. This effect was present across
the Intention, Regularity, as well as Both conditions, with
no differences between the three, as indicated by fre-
quentist as well as Bayesian analyses.
The N1b and Tb effects presumably reflect stimulus-
specific adaptation of the neuronal responses (Grill-
Spector, Henson, & Martin, 2006). The effect sizes and
Bayes factors suggest the N1 magnitude decreases as a
function of global tone probability, with strong effects
in the Regularity condition, followed by a decrease in
Both and finally no effects in Intention, where global reg-
ularity (standard-to-deviant probability, regardless of ac-
tion) is absent. That is, in the Regularity condition, the
standard tone was overall presented in 80% of the cases,
because the two buttons were equally pressed and gen-
erated the same frequent (and infrequent) tone. In the
Both condition, the two buttons were pressed with
80% versus 20% chances and were inversely associated
with the two tones. Subsequently, here, the frequently
presented tone was the standard generated by the fre-
quently pressed key, with a global regularity of 68% (re-
sulting from frequent key → standard 64% vs. deviant
16%; infrequent key → standard 16% vs. deviant 4%; stan-
dard 64% + deviant 4% [same tone between the two
keys] = 68%). Finally, in the Intention condition, the
mapping of standards and deviants was inversely associ-
ated with the left and right keys, which were pressed
equally frequent, meaning that the two tones were over-
all presented with equal chances.
The distinction typically made between the N1-consituent
components (SanMiguel, Todd, et al., 2013; Näätänen &
Picton, 1987; McCallum & Curry, 1980; Wolpaw & Penry,
1975) indicates that these N1b and Tb results can be inter-
preted as a consequence of “true” sensory predictions, in
contrast to mere orienting responses captured by the “un-
specific” N1, elicited with long ISIs (SanMiguel, Todd, et al.,
2013). Indeed, unlike most self-generation studies, we used
a short tone-to-tone interval; we made sure the timing be-
tween two consecutive button presses was stable around
1200 msec across all trials (in comparison to a range in be-
tween 2000 and 6000 msec, in typical self-generation stud-
ies). Participants proved to be able to keep the correct
pace for all blocks and conditions, as indicated by the
few timing errors (less than 3.5% on average), when they
produced intervals longer or shorter than 1200 msec by
more than 400 msec.
Next, our data suggest that the N1 component is
followed by MMN responses in the Regularity and Both
conditions. The N1–MMN succession is a typical result
associated with regularity-based prediction errors, where
while N1 reflects stimulus adaptation, the MMN
represents a memory- or prediction-driven comparison
of the expected versus received input (Garrido et al.,
2009; Näätänen et al., 2005). This pattern of results is also
compatible with the proposed stages of auditory distrac-
tion (Horváth et al., 2008), where, at an initial sensory
processing step, the N1 represents first-order and the
MMN second-order change detectors. Thus, in the
Regularity and Both conditions, first- and second-order
prediction errors are implemented in a bottom–up man-
ner via global tone regularity.
Top–Down Intention-based Prediction Errors Are
Reflected in the MMN Component
As already mentioned, the MMN is present in the
Regularity and Both conditions, where the global likeli-
hood of the standard tones was higher than the one of
the deviants, but also in the Intention condition, where
the two tones were presented with equal chances.
Because this effect is robust with and, importantly, with-
out global tone regularity, we propose that it represents
an intention-based prediction error (elicited in the
Intention condition). Consequently, we show that the
intention-based MMN is implemented in a top–down
manner, when controlled for neural adaptation reflected
in the early N1 response ( Jacobsen & Schröger, 2001).
One important distinction between adaptation at low
levels due to regular input and top–down effects is that
the first is an automatic and necessary side effect of
bottom–up sensory processing, whereas the second in-
volves high-level expectations, which are fed back down
the cortical hierarchy to achieve effects at sensory levels
(Lee & Mumford, 2003). It has indeed been shown that
top–down expectations regarding the quality (high or
low) of individual tones within a sequence modulate
the sensory ERP components starting from 100 msec
( Widmann et al., 2004). Altogether, these results indicate
that intention and regularity, in line with earlier (but
untested) suggestions, are distinct “sources for a single
mechanism” (Lange, 2013).
Referring to the intention-based expectations on devi-
ance processing, Waszak and Herwig asked participants
to generate standard versus deviant tones by voluntary
left and right key presses, similarly to here. Unlike here,
in their design, both key presses generated the same
standard and deviant tones in a test phase, whereas in
a previous acquisition phase, the left versus right actions
have been associated with either the standard or the de-
viant tone, with 100% certainty. Thus, in the test phase,
based on the intention to press a key over the other one,
either the standard or the deviant tone were to be ex-
pected, but based on tone regularity, the same standard
tone was frequently presented, regardless of the chosen
action. They have found P3a effects between the pre-
dicted and unpredicted deviants (i.e., the deviants asso-
ciated with the same button press as in the acquisition
phase vs. the deviants associated with the button press that
1928
Journal of Cognitive Neuroscience
Volume 31, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
has in the acquisition phase been associated with the other
tone) and conclude that this P3a effect is due to previously
formed intention-based expectations (Waszak & Herwig,
2007). Our results go a step further and show that, when
controlled for stimulus regularity, the intention-based ex-
pectations modulate deviance processing even earlier at
sensory levels reflected in the MMN response.
Congruent with the present MMN intention result, a
recent study by LeBars and colleagues reported that ef-
fects around 200 msec (at the level of what is described
as the N2b component) do indeed depend on whether
the participants are able to choose or not which button
to press. Specifically, similar to here, they had partici-
pants generate low and high standards and deviants,
which were inversely associated with left and right button
presses, and showed that only when the choice of which
key to press was determined by participants’ intention, in
contrast to externally cued, mismatch answers were elic-
ited (Le Bars, Darriba, & Waszak, 2019).
Further Deviance Detection Is Reflected in the P3a
Response for Bottom–Up and Top–
Down Predictions
Following up on the stages of auditory distraction, if the
deviation detection reflected in the N1 and MMN compo-
nents at the sensory processing step exceeds a certain
threshold, a second processing step reflected in the P3a
response follows (Horváth et al., 2008). We found P3a ef-
fects (larger positive amplitudes for the deviant, compared
with the standard tones) in all three conditions, with no
differences between conditions—these effects were once
again supported by frequentist as well as Bayesian analy-
ses. First, this indicates that the deviance has been strongly
perceived in all three conditions pointing to the success of
the experimental manipulation. Second, the P3a results
point out that, regardless of whether the predictions at
the initial sensory processing levels have been violated
based on either Regularity, Intention, or Both, this does
not seem to influence the following processing level
where the change detection mechanism reflects an invol-
untary attentional switch toward motivational (i.e., expec-
tancy violating) stimuli (Nieuwenhuis et al., 2011).
In the light of Waszak and Hervig’s P3a interpretation
(Waszak & Herwig, 2007), we cannot rule out the possibil-
ity that, in fact, the P3a effects reported here might also
reflect intention-based signatures across conditions. That
is, given that participants always chose when and which
key to press to produce a tone (regardless of whether that
tone was hand specific in the Intention and Both condi-
tions or associated with both hands in the Regularity
condition), the intention per se to perform an action to
produce an effect must have been a factor in all three con-
ditions similarly. Therefore, to better understand the P3a
effect in this context, it would be necessary to reduce the
intention-related processes in the case of regularity-based
predictions, for example, by cuing the left and right button
presses, similar to Le Bars et al. (2019). However, Le Bars
et al. (2019) do not show to what extent cuing the actions,
in contrast to intentional action, affects the magnitude of
the P3a effect. It thus remains for future research to es-
tablish the precise functional interpretation of the P3a
component following sensory predictions.
Concurrent Violations of Intention and Regularity
Do Not Lead to Stronger Prediction Errors
The interaction of top–down and bottom–up information
is widely discussed in the literature. It is generally
assumed that, in the context of hierarchical processing
of predictive information, bottom–up information and
top–down expectations are constantly contrasted and in-
tegrated in cortical feedforward/feedback loops (Lee &
Mumford, 2003). We did not find stronger prediction
errors elicited by concurrent violations of regularity and
intention—that is, the effects we report for the sensory
N1 and MMN components are not larger in the Both con-
dition, which would suggest additivity of Regularity and
Intention. This conclusion is supported by frequentist
and Bayesian analyses, as well as by testing the additivity
model (i.e., Regularity + Intention vs. Both). These data
therefore suggest that convergent predictions by regular-
ity and intention are integrated at lower levels of process-
ing (as opposed to being represented independently and
eliciting separate prediction error responses), despite the
two presumably having independent generative models.
A relevant study by Pieszek and colleagues compared
bottom–up regularity predictions with top–down predic-
tions based on audio-visual pairing. Specifically, within a
trial, the two types of information could either be contra-
dictory (i.e., one confirming vs. the other violating the
tone expectation), or both confirming, or concurrently
violating the expectation. Similar to this study, they
showed that bottom–up and top–down violations individ-
ually lead to prediction errors, as expressed by MMN and
incongruency responses, respectively. However, unlike
here, they report an additive bottom–up + top–down
model, where the difference wave corresponding to the
concurrent violations roughly matches the sum of the
two difference waves corresponding to the independent
predictions (Pieszek et al., 2013).
On the one hand, according to the predictive coding
theory, the prediction mechanism generates predictions
regarding both the context of the incoming stimulation,
as well as about the expected precision (Feldman &
Friston, 2010)—these effects can, in turn, be mediated
by attention (Schröger et al., 2015). From this perspective,
it would make sense that more precision (i.e., bottom–up
+ top–down) would lead to stronger prediction errors,
provided the expectations are violated—like in Pieszek
et al. (2013), for example. On the other hand, it is also like-
ly that the mechanism works based on an “efficiency rule,”
where, in the absence of special attentional resources,
once a reliable source of information is available for the
Korka, Schröger, and Widmann
1929
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
prediction (extracted from either tone regularity or action
intention), additional sources become redundant.
Regarding the first hypothesis according to which more
precision should enhance the prediction error, it is possible
that extra attentional resources would be required to en-
hance the prediction errors by additional information (as
opposed to additional information being redundant). It is
generally believed that attention and prediction work to-
gether to enhance precision (Schröger et al., 2015), and this
effect could also be transferable to concurrent bottom–up
and top–down predictions. To conclude, it remains for
future research to establish whether including an attention
manipulation in studying the nonconflicting predictions
based on regularity and intention changes the prediction
precision and leads to higher prediction errors for con-
current (both regularity and intention) in comparison to
single violations (either regularity or intention).
The second hypothesis becomes likely if we consider
that, in our design, throughout blocks corresponding to
the same condition (and within individual trials), the
bottom–up versus top–down predictions did not contra-
dict each other, in contrast to the study by Pieszek et al.
(2013). Specifically, in their design, within the same trial,
individual predictions could be violated based on tone
regularity, but confirmed based on the visual–auditory
pairing or vice versa. This situation is confusing, in com-
parison to when both information types unanimously
confirm or violate expectations; thus, it is unsurprising
that concurrently violating both predictions (i.e., high
certainty) leads to a stronger error than when violating
one but confirming the other (i.e., “confusion”). In this
study, regularity and intention seem to have provided
enough precision when presented individually, such that
presenting them together does not add certainty, but re-
dundancy. Note that another major difference between
the study of Pieszek et al. (2013) and this study consists
in the very nature of the top–down expectations (visual
vs. intention based), which might be implemented dif-
ferently, producing effects at different latencies. Indeed,
although in the study of Pieszek et al. (2013) the bottom–
up versus top–down effects are being additive for the
mean amplitudes around 105–130 msec, in this study
we show that several components in the 100–200 msec
latency ranges respond differently to bottom–up regular-
ity versus top–down intention manipulations.
No N1 Effects for Top–Down
Intention-based Modulations?
We shall finally point out that the lack of N1 effects in the
Intention condition is surprising, if we consider that pre-
vious studies bring forward the central contribution of
motor intention to explain the N1 results typically found
with self-generation paradigms (Timm et al., 2014;
Hughes et al., 2013). Because we can clearly exclude
the possibility that deviance has simply not been
detected in the Intention condition, based on the MMN
and P3a effects, different explanations can be considered.
First, as already mentioned, it has been proposed that the
N1 effects observed with self-generation studies represent
“unspecific” responses following tones presented at rather
long intervals (SanMiguel, Todd, et al., 2013) that do also
not necessarily have to be contingent to the button presses
(Horváth, 2013; Horváth, Maess, Baess, & Tóth, 2012).
Additionally, even though regularity is not often considered
as an explaining mechanism for the N1 effects reported with
typical self-generation paradigms, it might play a role.
Specifically, in contrast to self-generated tones where clear
temporal regularity is established via self-pacing, clear tem-
poral relations between consecutive tones cannot be as eas-
ily determined for externally generated tones (see Hughes
et al., 2012, 2013, for similar arguments). In this study, the
button presses determine sounds in all conditions at the
same self-pacing rate—this means there are no temporal
regularity differences between conditions and thus no
effects are elicited at this level. Altogether, this leaves the
later components around 200 msec as better candidates
for the interpretation based around motor intention, idea
previously suggested (SanMiguel, Todd, et al., 2013) and re-
cently confirmed by LeBars and colleagues who showed that
the N2b, but not the N1, is specific to intention-driven in com-
parison to stimulus-driven action (Le Bars et al., 2019).
Alternatively, a more trivial but also likely explanation is that
the action–tone associations in the Intention condition were
simply not strong enough to implement predictions at N1
sensory levels. In a comparable study, Hughes and colleagues
asked participants to produce action-specific tones by
inversely associating left versus right key presses with high
versus low pitch tones (Hughes et al., 2013). In the predict-
able condition, these associations generated their corre-
sponding tones with 100% probability, whereas in the
unpredictable condition, two additional buttons for each
hand generated high and low tones with equal chances.
Comparing predictable versus unpredictable tones across
both hands, they did not find an N1 effect—we thus replicate
these findings. However, Hughes et al. (2013) found an N1
effect by looking at hand-specific prediction-congruent
versus -incongruent tones. This result suggests that
intention-based predictions at N1 levels can occur if a strong
association is built-up between a hand-specific action and its
corresponding tone. Correspondingly, it is possible that if
the associations in our Intention condition were stronger,
we would observe prediction errors at the N1 level. This hy-
pothesis should be further tested—this is particularly worth
investigating, because, if stronger action–tone associations in
the Intention condition would lead to N1 effects (similar to
the Regularity condition), this could in turn lead to additivity
of bottom–up and top–down effects at N1 sensory levels.
Summary and Conclusion
This study represents, to our best knowledge, the first
direct comparison of intention and regularity effects, in
1930
Journal of Cognitive Neuroscience
Volume 31, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
the context of action-related predictions. We show that the
N1 component is modulated by tone regularity violations,
but not by intention violations—one possible explanation
is that it reflects a necessary consequence of neural adapta-
tion. However, the MMN component is modulated both by
regularity and intention violations. Because this effect is ro-
bust in all three conditions regardless of the global tone
regularity (i.e., when controlled for neural adaptation), we
argue that top–down predictions based on action intention
are reflected at this sensory processing level. We did not
observe stronger prediction errors when regularity and in-
tention were concurrently violated (i.e., the two did not
have additive effects). This suggests that, even though
the two presumably have independent generators, con-
verging predictions integrate (in contrast to prediction er-
ror responses adding up). Similar P3a effects across
conditions point out that the deviance is further pro-
cessed, regardless of whether the effects at the earlier sen-
sory levels were based on regularity, intention, or both.
Acknowledgments
We thank all participants who took part in this study. The data
collection and analysis were conducted at the Cognitive and
Biological Psychology Laboratory, University of Leipzig. This
project was funded by the German Research Foundation
(www.dfg.de) with project number GZ: SCHR 375/25-1. We
would also like to thank Taavi Wenk for his help with the data
collection as well as our lab colleagues for their useful com-
ments regarding this study design.
Reprint requests should be sent to Betina Korka, Cognitive and
Biological Psychology, Institute of Psychology, University of
Leipzig, Neumarkt 9-19, D-04109 Leipzig, Germany, or via
e-mail: betina-christiana.korka@uni-leipzig.de.
Notes
1. For the key–tone associations, in the Regularity condition, the
first half of the participants generated a low tone and the second
half generated a high tone as standards. In the Intention condi-
tion, half of the participants who generated the low standard tone
in the Regularity condition generated a low standard tone with the
left key and a high standard tone with the right key, and vice versa
for the other half. The same associations were implemented for
the second half of the participants who generated the high stan-
dard tone in the Regularity condition, thus far obtaining four
quarters of possible associations (1: Regularity low–Intention
left low–Intention right high; 2: Regularity low–Intention left
high–Intention right low; 3: Regularity high–Intention left low–
Intention right high; 4: Regularity high–Intention left high–
Intention right low). In the Both condition, half of the participants
(Quarters 1 and 4) pressed the left key frequently, and half of
them pressed the right key frequently (Quarters 2 and 3). The
key–tone associations were the same as in the Intention condi-
tion. Note that the standard tone for the frequently pressed key
in the Both condition was the same as in the Regularity condition.
For the condition order, permutations of Regularity, Intention,
and Both conditions result in six different combinations, each of
which was assigned to a participant within one quarter (6 condi-
tion orders × 4 quarters = 24 participants).
2. The measures computed by FASTER include the median
slope of time course, slope of the power spectrum, spatial
kurtosis, Hurst exponent, and correlation with eye channels.
ADJUST computes the spatial average difference, spatial eye dif-
ference, generic discontinuity spatial feature, maximum epoch
variance, and temporal kurtosis. Finally, the measures com-
puted by SASICA are low autocorrelation of time course, focal
channel topography, focal trial activity, correlation with vertical
EOG, and correlation with horizontal EOG. On average, 13.25
(range: 11–14) components per participant were removed.
REFERENCES
Bäß, P., Jacobsen, T., & Schröger, E. (2008). Suppression of the
auditory N1 event-related potential component with
unpredictable self-initiated tones: Evidence for internal
forward models with dynamic stimulation. International
Journal of Psychophysiology, 70, 137–143.
Bendixen, A., SanMiguel, I., & Schröger, E. (2012). Early
electrophysiological indicators for predictive processing in
audition: A review. International Journal of
Psychophysiology, 83, 120–131.
Bigdely-Shamlo, N., Mullen, T., Kothe, C., Su, K. M., &
Robbins, K. A. (2015). The PREP pipeline: Standardized
preprocessing for large-scale EEG analysis. Frontiers in
Neuroinformatics, 9, 16.
Chatrian, G. E., Lettich, E., & Nelson, P. L. (1985). Ten percent
electrode system for topographic studies of spontaneous
and evoked EEG activities. American Journal of EEG
Technology, 25, 83–92.
Chaumon, M., Bishop, D. V., & Busch, N. A. (2015). A practical
guide to the selection of independent components of the
electroencephalogram for artifact correction. Journal of
Neuroscience Methods, 250, 47–63.
Crowley, K. E., & Colrain, I. M. (2004). A review of the
evidence for P2 being an independent component process:
Age, sleep and modality. Clinical Neurophysiology, 115,
732–744.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trail EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Dien, J. (2010). The ERP PCA toolkit: An open source program
for advanced statistical analysis of event-related potential
data. Journal of Neuroscience Methods, 187, 138–145.
Dien, J. (2012). Applying principal components analysis to
event-related potentials: A tutorial. Developmental
Neuropsychology, 37, 497–517.
Eaton, J. W., Bateman, D., Hauberg, S., & Wehbring, R. (2016).
GNU Octave ( Version 4.2.0.).
Elsner, B., & Hommel, B. (2001). Effect anticipation and action
control. Journal of Experimental Psychology: Human
Perception and Performance, 27, 229–240.
Escera, C., Alho, K., Winkler, I., & Näätänen, R. (1998). Neural
mechanisms of involuntary attention to acoustic novelty and
change. Journal of Cognitive Neuroscience, 10, 590–604.
Feldman, H., & Friston, K. J. (2010). Attention, uncertainty, and
free-energy. Frontiers in Human Neuroscience, 4, 215.
Friston, K. (2005). A theory of cortical responses. Philosophical
Transactions of the Royal Society of London, Series B:
Biological Sciences, 360, 815–836.
Garrido, M. I., Kilner, J. M., Stephan, K. E., & Friston, K. J.
(2009). The mismatch negativity: A review of underlying
mechanisms. Clinical Neurophysiology, 120, 453–463.
Grill-Spector, K., Henson, R., & Martin, A. (2006). Repetition
and the brain: Neural models of stimulus-specific effects.
Trends in Cognitive Sciences, 10, 14–23.
Hari, R., Kaila, K., Katila, T., Tuomisto, T., & Varpula, T. (1982).
Interstimulus interval dependence of the auditory vertex
Korka, Schröger, and Widmann
1931
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
response and its magnetic counterpart: Implications for their
neural generation. Electroencephalography and Clinical
Neurophysiology, 54, 561–569.
Horváth, J. (2013). Action-sound coincidence-related
attenuation of auditory ERPs is not modulated by affordance
compatibility. Biological Psychology, 93, 81–87.
Horváth, J. (2015). Action-related auditory ERP attenuation:
Paradigms and hypotheses. Brain Research, 1626, 54–65.
Horváth, J., Maess, B., Baess, P., & Tóth, A. (2012). Action–
sound coincidences suppress evoked responses of the
human auditory cortex in EEG and MEG. Journal of
Cognitive Neuroscience, 24, 1919–1931.
Horváth, J., Winkler, I., & Bendixen, A. (2008). Do N1/MMN, P3a,
and RON form a strongly coupled chain reflecting the three stages
of auditory distraction? Biological Psychology, 79, 139–147.
Hughes, G., Desantis, A., & Waszak, F. (2012). Mechanisms of
intentional binding and sensory attenuation: The role of
temporal prediction, temporal control, identity prediction,
and motor prediction. Psychological Bulletin, 139, 133–151.
Hughes, G., Desantis, A., & Waszak, F. (2013). Attenuation of
auditory N1 results from identity-specific action–effect
prediction. European Journal of Neuroscience, 37, 1152–1158.
Jääskeläinen, I. P., Ahveninen, J., Bonmassar, G., Dale, A. M.,
Ilmoniemi, R. J., Levänen, S., et al. (2004). Human posterior
auditory cortex gates novel sounds to consciousness. Proceedings
of the National Academy of Sciences, U.S.A., 101, 6809–6814.
Jacobsen, T., & Schröger, E. (2001). Is there pre-attentive memory-
based comparison of pitch? Psychophysiology, 38, 723–727.
Jeffreys, H. (1961). Theory of probability. Oxford: Oxford
University Press.
Kleiner, M., Brainard, D., Pelli, D., Ingling, A., Murray, R., &
Broussard, C. (2007). What’s new in Psychtoolbox-3.
Perception, 36, 1–16.
Knill, D. C., & Pouget, A. (2004). The Bayesian brain: The role of
uncertainty in neural coding and computation. Trends in
Neurosciences, 27, 712–719.
Kotchoubey, B., & Pavlov, Y. G. (2019). A Signature of passivity?
An explorative study of the N3 event-related potential
component in passive oddball tasks. Frontiers in
Neuroscience, 13, 365.
Lange, K. (2013). The ups and downs of temporal orienting: A
review of auditory temporal orienting studies and a model
associating the heterogeneous findings on the auditory N1
with opposite effects of attention and prediction. Frontiers in
Human Neuroscience, 7, 263.
Le Bars, S., Darriba, Á., & Waszak, F. (2019). Event-related brain
potentials to self-triggered tones: Impact of action type and
impulsivity traits. Neuropsychologia, 125, 14–22.
Lee, T. S., & Mumford, D. (2003). Hierarchical Bayesian
inference in the visual cortex. Journal of the Optical Society
of America A: Optics, Image, Science, and Vision, 20,
1434–1448.
Lee, M. D., & Wagenmakers, E.-J. (2014). Bayesian cognitive
modeling: A practical course. Cambridge: Cambridge
University Press.
McCallum, W. C., & Curry, S. H. (1980). The form and
distribution of auditory evoked potentials and CNVs when
stimuli and responses are lateralized. Progress in Brain
Research, 48, 229–240.
Mognon, A., Jovicich, J., & Bruzzone, L. (2011). ADJUST:
An automatic EEG artifact detector based on the joint use
of spatial and temporal features. Psychophysiology, 48,
229–240.
Näätänen, R., Jacobsen, T., & Winkler, I. (2005). Memory-based
or afferent processes in mismatch negativity (MMN): A
review of the evidence. Psychophysiology, 42, 25–32.
Näätänen, R., Paavilainen, P., Rinne, T., & Alho, K. (2007). The
mismatch negativity (MMN) in basic research of central
auditory processing: A review. Clinical Neurophysiology,
118, 2544–2590.
Näätänen, R., & Picton, T. (1987). The N1 wave of the human
electric and magnetic response to sound: A review and an
analysis of the component structure. Psychophysiology, 24,
375–425.
Nieuwenhuis, S., De Geus, E. J., & Aston-Jones, G. (2011). The
anatomical and functional relationship between the P3 and
autonomic components of the orienting response.
Psychophysiology, 48, 162–175.
Nolan, H., Whelan, R., & Reilly, R. B. (2010). FASTER: Fully
automated statistical thresholding for EEG artifact rejection.
Journal of Neuroscience Methods, 192, 152–162.
Oprina, A., Popescu, A., Simion, E., & Simion, G. (2009). Walsh–
Hadamard randomness test and new methods of test results
integration. Bulletin of the Transilvania University of
Brasov. Mathematics, Informatics, Physics. Series III, 2,
93–105.
Pieszek, M., Widmann, A., Gruber, T., & Schröger, E. (2013).
The human brain maintains contradictory and redundant
auditory sensory predictions. PLoS One, 8, e53634.
Prinz, W. (1997). Perception and action planning. European
Journal of Perception and Action Planning, 9, 129–154.
Rouder, J. N., Morey, R. D., Speckman, P. L., & Province, J. M.
(2012). Default Bayes factors for ANOVA designs. Journal of
Mathematical Psychology, 56, 356–374.
SanMiguel, I., Saupe, K., & Schröger, E. (2013). I know what is
missing here: Electrophysiological prediction error signals
elicited by omissions of predicted “what” but not “when”.
Frontiers in Human Neuroscience, 7, 407.
SanMiguel, I., Todd, J., & Schröger, E. (2013). Sensory
suppression effects to self-initiated sounds reflect the
attenuation of the unspecific N1 component of the auditory
ERP. Psychophysiology, 50, 334–343.
SanMiguel, I., Widmann, A., Bendixen, A., Trujillo-Barreto, N., &
Schröger, E. (2013). Hearing silences: Human auditory
processing relies on preactivation of sound-specific brain
activity patterns. Journal of Neuroscience, 33, 8633–8639.
Schröger, E. (1997). On the detection of auditory deviations: A
pre-attentive activation model. Psychophysiology, 34,
245–257.
Schröger, E., Marzecová, A., & SanMiguel, I. (2015). Attention
and prediction in human audition: A lesson from cognitive
psychophysiology. European Journal of Neuroscience, 41,
641–664.
Shahin, A., Roberts, L. E., Pantev, C., Trainor, L. J., & Ross, B.
(2005). Modulation of P2 auditory-evoked responses by the
spectral complexity of musical sounds. NeuroReport, 16,
1781–1785.
Timm, J., SanMiguel, I., Keil, J., Schröger, E., & Schönwiesner,
M. (2014). Motor intention determines sensory attenuation
of brain responses to self-initiated sounds. Journal of
Cognitive Neuroscience, 26, 1481–1489.
Wagenaar, W. A. (1972). Generation of random sequences by
human subjects: A critical survey of literature. Psychological
Bulletin, 77, 65–72.
Waszak, F., & Herwig, A. (2007). Effect anticipation modulates
deviance processing in the brain. Brain Research, 1183,
74–82.
Widmann, A., Kujala, T., Tervaniemi, M., Kujala, A., & Schröger,
E. (2004). From symbols to sounds: Visual symbolic
information activates sound. Psychophysiology, 41, 709–715.
Widmann, A., Schröger, E., & Maess, B. (2015). Digital filter
design for electrophysiological data—A practical approach.
Journal of Neuroscience Methods, 250, 34–46.
Wolpaw, J. R., & Penry, J. K. (1975). A temporal component of
the auditory evoked response. Electroencephalography and
Clinical Neurophysiology, 39, 609–620.
1932
Journal of Cognitive Neuroscience
Volume 31, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
1
2
1
9
1
7
1
8
6
0
9
6
4
/
/
j
o
c
n
_
a
_
0
1
4
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3