Evolution of Autopoiesis and
Multicellularity in the Game of Life
Peter D. Turney*
Ronin Institute
peter.turney@ronininstitute.org
Abstract Recently we introduced a model of symbiosis, Model-S,
based on the evolution of seed patterns in Conwayʼs Game of Life. En
el modelo, the fitness of a seed pattern is measured by one-on-one
competitions in the Immigration Game, a two-player variation of the
Game of Life. Our previous article showed that Model-S can serve as
a highly abstract, simplified model of biological life: (1) The initial
seed pattern is analogous to a genome. (2) The changes as the game
runs are analogous to the development of the phenome. (3)
Tournament selection in Model-S is analogous to natural selection in
biology. (4) The Immigration Game in Model-S is analogous to
competition in biology. (5) The first three layers in Model-S are
analogous to biological reproduction. (6) The fusion of seed patterns
in Model-S is analogous to symbiosis. The current article takes this
analogy two steps further: (7) Autopoietic structures in the Game of
Life (still lifes, oscillators, and spaceships—collectively known as ashes) son
analogous to cells in biology. (8) The seed patterns in the Game of
Life give rise to multiple, diverse, cooperating autopoietic structures,
analogous to multicellular biological life. We use the apgsearch software
(Ash Pattern Generator Search), developed by Adam Goucher for the
study of ashes, to analyze autopoiesis and multicellularity in Model-S.
We find that the fitness of evolved seed patterns in Model-S is
highly correlated with the diversity and quantity of multicellular
autopoietic structures.
Palabras clave
Evolución, autopoiesis, multicellularity,
cellular automata, diversity, symbiosis
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
1 Introducción
In previous work (Turney, 2020b), we introduced Model-S, a computational model of the evolution
of symbiosis. In Model-S, seed patterns compete for survival in one-on-one competitions in the
Immigration Game (IG), invented by Don Woods in 1971 (LifeWiki, 2021mi; Wainwright, 1971).
The Immigration Game is a two-player variation on the solitaire Game of Life (GoL), invented
by John Conway in 1970 (jardinero, 1970; Poundstone, 2013). The winner of IG is the seed pattern
that grows the most. Model-S adds an external evolutionary algorithm to IG. The algorithm evolves
a population of seed patterns, selecting seeds according to their ability to win IG.
Model-S is constructed with four layers, each subsequent layer building on the previous layers.
The purpose of having four layers is to measure each additional layerʼs contribution to the fitness of
la población. This provides some insight into the strengths and weaknesses of each layer. Cifra 1
* Autor correspondiente.
© 2021 Instituto de Tecnología de Massachusetts.
Artificial Life 27: 26–43 (2021) https://doi.org/10.1162/artl_a_00334
Publicado bajo una atribución Creative Commons
4.0 Internacional (CC POR 4.0) licencia.
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. This flowchart outlines the process for selecting an individualʼs genome from the population and creating a new
genome. This process is a subroutine in a loop that produces a series of new individuals. For each individual that is added
to the population, another is removed; hence this is a steady-state model with a constant population size. The decision to
use a given layer is determined by the parameters of Model-S.
presents a flow chart that sketches the basic structure of Model-S. For a more detailed flow chart,
see our previous article (Turney, 2020b).
Layer 1 implements a simple form of asexual reproduction, with a fixed genome size (eso es, a
fixed binary matrix size). A member of the population is selected for reproduction using tournament
selección. The chosen seed pattern is mutated by randomly flipping some of the bits in the binary
matrix, and it then competes in a series of one-on-one Immigration Games with the other members
of the population. Its fitness is the average fraction of games it wins. Layer 2 implements a slightly
more sophisticated asexual reproduction. In addition to mutation by flipping bits (in Layer 1), el
binary matrix is allowed to grow or shrink by adding or subtracting a row or column to or from
the matrix. Layer 3 selects two seeds from the population and then combines them with genetic cross-
encima. The resulting child is then passed on to Layer 2 for mutation. Layer 4 adds symbiosis to Model-S.
Two seeds are selected from the population and fused together, side-by-side, creating a new symbiotic
genome. This new fused seed is treated as a whole; eso es, selection shifts from the level of the two parts
Artificial Life Volume 27, Número 1
27
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
to the level of the whole. The main result of our past work is evidence for the hypothesis that symbiosis
promotes fitness improvements in IG (Turney, 2020b).
In the course of our past research with Model-S, it became apparent that autopoiesis plays an
important role in determining the fitness of seed patterns. The term autopoiesis was introduced by
Humberto Maturana and Francisco Varela (Maturana & Varela, 1973). Randall Beer (2014, pag. 185)
states that “… an autopoietic system consists of a network of processes that produce components
whose interactions serve to generate and maintain the very network of processes that produced them.”
Beer has shown, in a series of articles (Beer, 2004, 2014, 2015), that GoL is an excellent platform
for modeling autopoiesis. The classes of structures in GoL that are most interesting to Beer regard-
ing autopoiesis are still lifes, oscillators, and spaceships (Beer, 2015). These three classes of patterns
are structures that naturally emerge in the game and capture the attention of GoL players. A popular
activity in the GoL community is to create a random soup and run it to quiescence (LifeWiki,
2021gramo). A soup is a random initial seed pattern. When the soup reaches quiescence, what remains
is called ash. Ashes are defined as still lifes, oscillators, and spaceships (LifeWiki, 2021h).
Cifra 2 presents a sample of the output of Model-S. The left column in Figure 2 shows four
initial seed patterns, randomly sampled from the final generations of four different runs of Model-S.
The right column shows the final ash that results from each seed. The first row presents a run of
Model-S with Layer 1 solo; the other layers are turned off. Each following row shows the effect of
adding another layer to the model.
Mesa 1 summarizes the types and quantities of ash for the four different runs of Model-S in
Cifra 2. Mesa 1 suggests that symbiosis (Layer 4) in Model-S yields large quantities of ash, com-
pared with the other layers. En la sección 4, we will present statistical analysis that supports the hypoth-
esis that symbiosis yields highly fit seed patterns by evolving seeds that yield diverse types and
large quantities of ash.
Model-S is a highly abstract, simplified model of biological life. Mesa 2 outlines a mapping be-
tween elements of biological life and elements of Model-S. Our previous article on Model-S (Turney,
2020b) discussed the analogies presented here in the first six rows in Table 2: (1) The initial seed
pattern in a cellular automaton is like a genome. It is a static encoding of the information that de-
termines how the game will unfold, according to the rules of the game. (2) Running IG is like the
growth of the phenome over time. Two seed patterns grow and compete with each other for space.
(3) The tournament selection algorithm in Model-S is like natural selection. The winner of a tour-
nament is selected for reproduction. (4) IG is used to measure the fitness of a player, based on the
number of living cells in the player. IG is a model of the fitness of organisms competing for repro-
ducción. (5) Layers 1, 2, y 3 in Model-S implement three different forms of reproduction: Layer 1
reproduces a selected genome with random binary mutations, Layer 2 reproduces a selected genome
with changes to the size of the genome, and Layer 3 combines two genomes with genetic crossover.
(6) The fusion of seed patterns in Layer 4 in Model-S is like symbiosis. En particular, fusion can be
viewed as a form of endosymbiosis with vertical transmission of genes. The current article focuses
on the last two rows in Table 2: (7) Ashes (still lifes, oscillators, and spaceships) are like living cells,
y (8) multiple ashes that grow from a single seed pattern are like multiple cells in a living organism.
Before we continue, letʼs pause to discuss a possible source of confusion. The word cell might
refer to a square in the grid of GoL or it might refer to an autopoietic structure, such as ash in GoL
or an actual biological cell. When there might be confusion, we will use the terms grid-cell for a square
in the grid and ash-cell for an autopoietic structure.
The main contributions of this article are (1) evidence that the fitness of a seed pattern depends
on the number of ashes that it is able to generate (the productivity of the seed pattern, en la sección 4.1),
(2) evidence that the fitness of a seed pattern depends on the variety of ashes that it is able to
generate (the diversity of types of ashes, en la sección 4.2), y (3) evidence that, although the fitness
of a seed pattern is correlated with its size, size alone is not sufficient for fitness: Productivity and
diversity require complex evolved structures (en la sección 4.3).
These three lines of evidence support the analogies presented in the last two rows of Table 2. (1)
A seed pattern that is highly productive is likely to win in competition against a seed pattern that is
28
Artificial Life Volume 27, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 2. These images show examples of initial evolved seed patterns for each layer on the left and the corresponding
final patterns on the right. The final patterns continue to change, but they are composed entirely of ashes. Gliders are
omitted in Layers 3 y 4 because they have moved far from the core ashes.
less productive. Asimismo, in biology, an organism with many cells may have an advantage when
competing against an organism with few cells. Model-S measures fitness using IG, where the winner
is the pattern with the most living grid-cells, but productivity is based on the number of ash-cells. Él
does not necessarily follow that productivity (number of ash-cells) is correlated with fitness (number
of grid-cells); por lo tanto, an experiment is required to determine the relation between productivity
and fitness (mira la sección 4.1). (2) A seed pattern that produces a diversity of types of ashes is likely to
have an advantage over a seed pattern that has little variety, because each type of ash may require a
Artificial Life Volume 27, Número 1
29
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
Mesa 1. This table summarizes the statistics for the final ash patterns shown in Figure 2.
Layer 1 ashes
Layer 2 ashes
Layer 3 ashes
Layer 4 ashes
Tipo
block
blinker
pond
Count
Tipo
Count
Tipo
Count
Tipo
Count
6
5
2
block
beehive
blinker
glider
barco
boat
7
3
1
1
1
1
block
blinker
boat
glider
beehive
barco
loaf
long boat
16
blinker
8
5
4
4
2
2
1
block
beehive
glider
boat
barco
pond
tub
loaf
pulsar
36
25
15
13
6
4
3
2
1
1
Num objects
Num types
13
3
Num objects
Num types
14
6
Num objects
Num types
42
8
Num objects
106
Num types
10
Nota. For more information about the types of ashes listed in the table, see Adam Goucherʼs Catagolue (LifeWiki,
2021F). Num = number of.
Mesa 2. This table shows correspondences between characteristics of biological life and Model-S. In a highly abstract and
simplified way, Model-S exhibits several of the main concepts of biology.
Biological Life
Model-S
Shared Characteristics
genome, DNA, chromosomes
initial seed pattern, initial soup
static information, código
phenome, organism,
desarrollo
automaton
playing the game, running the cellular
dynamic development,
growth
selección
natural selection
tournament selection
competition
the Immigration Game
fitness measure
reproduction with mutation,
Layers 1, 2, y 3 of Model-S
variación, sexo
reproduction with
heritable variation
mutualism, cooperation
Layer 4, fusion of seed patterns
symbiosis
cells
still lifes, oscillators, spaceships, ashes
autopoiesis
multicellular organisms
multiple ashes grown from the same
multicellularity
initial seed pattern
Artificial Life Volume 27, Número 1
1
2
3
4
5
6
7
8
30
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
different competitive strategy to defeat it. In biology, an organism with several types of cells (como
skin cells, muscle cells, and brain cells) may have an advantage when competing against an organism
with only a few types of cells (mira la sección 4.2). (3) Winning IG requires complex evolved seed pat-
charranes, just as survival of biological organisms requires complex evolved genomes (mira la sección 4.3).
Although we are using Model-S as a specific example in this article, the phenomena we observe
(increasing instances of autopoiesis and multicellularity) are a consequence of trying to improve the
odds of winning the Immigration Game; the phenomena are not specific to Model-S. It seems nec-
essary that any effort to create soups (seed patterns) that are increasingly better at winning Immi-
gration Games will result in soups that yield increasing productivity and diversity of ashes. From this
perspectiva, the current article tells us more about the nature of the Immigration Game and the
Game of Life than it tells us about Model-S.
We review related work in Section 2. En la sección 3, we present background information about
Model-S (Sección 3.1) and autopoiesis (Sección 3.2). This background supports the core of the paper
en la sección 4, the experiments with fitness and productivity (Sección 4.1), fitness and diversity
(Sección 4.2), the relations among productivity, diversity, and size (Sección 4.3), and the lessons
learned from the experiments (Sección 4.4). Sección 5 discusses future work and limitations of the
análisis. The article concludes in Section 6.
2 Trabajo relacionado
Conwayʼs Game of Life was introduced to the world by Martin Gardner (jardinero, 1970). His in-
troductory article mentioned still lifes, oscillators, and spaceships, the three classes of objects that are
now covered by the term ash (LifeWiki, 2021h). The term primordial soup was used by Wainwright in
1971 to describe a random initial state in GoL (jardinero, 1971). The GoL community generates
random soups and runs them to quiescence in order to find interesting ashes (Goucher, 2021b;
LifeWiki, 2021d).
Maturana and Varela introduced autopoiesis to the world in 1973 (Maturana & Varela, 1973).
McMullin (2004) reports that Maturana and Varela had coined the word autopoiesis in 1971 y ellos
were planning at that time to make a cellular automaton computer simulation of autopoiesis, inspired
by the work of von Neumann (1966) and Conway (jardinero, 1970). Their cellular automaton model
was published in 1974 (McMullin, 2004; Varela et al., 1974). It used a simulated chemistry in a two-
dimensional grid space. A cell in the space could be empty or it could contain one of three types of
particles. The rules of the artificial chemistry were designed to allow particles to connect and form a
kind of cell membrane. The membrane was capable of self-maintenance and self-construction.
Beer (2004, 2014, 2015) has made a strong case for GoL as a model of autopoiesis. At first, él
focused on gliders as instances of autopoiesis (Beer, 2004, 2014), but later he expanded his studies to
still lifes, oscillators, and spaceships (Beer, 2015). This expansion connects the research in autopoi-
esis to the investigations of the GoL community exploring soups and ashes.
The term symbiosis is derived from the Ancient Greek word (cid:1)(cid:3)(cid:4)(cid:5)ί !(cid:1)(cid:6)& (sumbíōsis), significado
“living together.” There are many different types of symbiosis (douglas, 2010). Martin and
Schwab (2013, pag. 32) write, “Confusion has afflicted the definition of symbiosis for over 130 years.”
They propose a set of terms based on whether the two species in a symbiotic relationship experience
a beneficial effect (+), a harmful effect (−), or a neutral effect (0). This yields six types of symbiosis:
neutralism (0/0), antagonism (−/−), amensalism (0/−), agonism (+/−), commensalism (+/0), and mutualism
(+/+) (Martín & Schwab, 2013, pag. 41). They also distinguish endosymbiosis (one organism lives within
the tissues of the other ) and ectosymbiosis (one organism lives on the body surface of the other,
including the digestive tract).
Maynard Smith and Szathmáry, studying the major transitions in evolution, found that several of
the major transitions involved symbiosis with a shift in the level of selection (Maynard Smith &
Szathmáry, 1995). The prototypical example is the transition from prokaryotes to eukaryotes
(Margulis, 1970, 1981; Maynard Smith & Szathmáry, 1995). This is an instance of endosymbiosis
Artificial Life Volume 27, Número 1
31
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
where the two organisms become so tightly bound together that they cannot live apart, so natural
selection must select them as a whole; it is no longer possible to select only one of the two parts.
Layer 4 in Model-S (fusion of seed patterns) is a model of endosymbiosis.
The current article brings together the work of the GoL community on soups and ashes
(Goucher, 2021a, 2021b; LifeWiki, 2021C, 2021d, 2021gramo); the research in theoretical biology on
autopoiesis (Beer, 2014, 2015; Varela et al., 1974); the research of the evolutionary computation
community on models of evolution (Simón, 2013; Whitley, 1989; Whitley & Kauth, 1988); y el
research in theoretical biology on symbiosis (douglas, 2010; Margulis, 1970, 1981).
3 Fondo: Model-S and Autopoiesis
Before we present the experiments (en la sección 4), we need to provide some more background in-
formation on Model-S (Sección 3.1) and autopoiesis (Sección 3.2).
3.1 Model-S
The Game of Life is a solitaire game in which the player creates an initial seed pattern in an infinite
grid of square cells (jardinero, 1970; Poundstone, 2013). The rules of GoL determine how the pat-
tern changes over time. In Model-S, we view the initial seed pattern as analogous to a genome, y el
rules determine how the phenome develops from the genome (ontogeny—the development of an or-
ganism over its lifespan).
Each cell in the GoL grid is either dead (estado 0) or alive (estado 1). The state of a cell depends on the
state of its eight nearest neighbors in the grid. The rules for GoL are concisely expressed as B3/S23:
A cell is born (it switches from state 0 to state 1) if it has exactly three live neighbors (B3). A cell
survives (it stays in state 1) if it has either two or three live neighbors (S23). De lo contrario, the cell re-
mains dead or it dies (estado 0).
Time passes in a series of discrete intervals. The states of the cells in the grid at time t = 0
constitute the initial seed pattern for the game. The states at time t uniquely determine the new states
at time t + 1. With each time step, all of the cells in the grid are updated.
The Immigration Game is a two-player game in which the two players each create an initial seed
patrón (LifeWiki, 2021mi; Wainwright, 1971). The rules for life and death are B3/S23, as in GoL,
except that there are two alive states in IG (estados 1 y 2). Conventionally, the background (estado 0)
is usually white and the living states (estados 1 y 2) are usually red and blue. There are two new rules
for determining color. (1) Live cells do not change color unless they die (switch to state 0). (2) Cuando
a new cell is born, it takes the color of the majority of its live neighbors. Since birth requires three
live neighbors, there is always a clear majority.
Model-S (Turney, 2021) uses the Golly cellular automata software (Trevorrow et al., 2021) to run
Immigration Games (LifeWiki, 2021mi; Wainwright, 1971). In Model-S, Golly is controlled by Python
scripts that implement evolution outside Golly. A seed pattern is represented in Python by a binary
matrix. The initial population consists of 200 random seed patterns. The algorithm for evolution is a
steady-state model with a constant population size (Whitley, 1989; Whitley & Kauth, 1988), en el cual
each new child replaces the least fit member of the population. Members of the population are
chosen for reproduction using tournament selection (Whitley, 1989).
In Model-S, fitness is determined by one-on-one competitions between two seed patterns in IG.
The seeds are represented as binary matrices when they are not competing. When it is time to com-
pete, one seed is assigned red (estado 1 remains state 1) and the other seed is assigned blue (estado 1
switches to state 2). The two competitors are randomly situated in a toroidal grid with some space
separating them. The game runs until a time limit is reached. The winner is the seed that grows the most.
The growth of the red seed is the number of cells in state 1 at the end of the game minus the
number of cells in state 1 at the beginning of the game. Asimismo, the growth of the blue seed is
the increase in blue states. If a seed shrinks instead of growing, its score is zero. Ties are allowed.
32
Artificial Life Volume 27, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
In these competitions, the size of the toroidal grid and the time limit for the game are both
functions of the sizes of the seeds. Larger seeds are allowed larger grids and longer time limits.
The reasoning is that any fixed limit on space and time would set a bound on the possible evolution
of the seeds over many generations, which would be less interesting than unbounded growth. El
motivation for a toroidal grid is to limit the space for growth, so that the two seeds are forced to
interact. Without a limit on space, the two seeds might simply avoid each other, which would be less
interesting than interaction. The toroid limits the space for any given competition, but the toroid
increases in size from one competition to another, if the seed patterns increase in size. Asimismo, el
time limit is fixed for any given competition, but can increase from one competition to another.
(Space and time can also decrease, if the seeds happen to decrease in size.)
In IG, if states 1 y 2 were colored black, the game would appear to be exactly the same as
GoL. The purpose of the two colors is to score the two players, to convert GoL from a solitaire
game into a two-player competitive game. In IG, we can choose to be red–blue color blind, y
view the game as GoL. In the work we describe in this article, Model-S measures the fitness of seeds
by competitions in the three-state Immigration Game (two seeds at a time), but the ashes are an-
alyzed in the two-state Game of Life (one seed at a time).
The fitness of a seed in IG is the fraction of games that it wins playing against every seed in the
población. The initial population consists of 200 randomly generated seed patterns. Tournament
selection randomly chooses two seeds from the population, and the fitter of the two is selected for
reproduction. The type of reproduction depends on which layers are enabled. The new child re-
places the least fit member of the population, so the population remains at 200 for the entire
run of Model-S. Cuando 200 children have been born, we say that one generation has passed. Nosotros
run Model-S for 100 generaciones, Resultando en 20,000 births (100 generations with 200 births =
20,000 births; the generation count goes from 0 a 100, but generation 0 is randomly generated,
not born from parents).
The fitness of a seed pattern is measured by two competitions against every seed pattern in the
población (2 competitions per competitor × 200 competitors = 400 competitions, incluido 2 self-
competitions). Each competition involves two different seed patterns in the same grid, playing IG.
The two seed patterns interact with each other, which means that their phenomes are different in
each competition, due to their interaction. The fitness of a seed pattern is defined as the average
number of games the pattern wins, playing IG. This average is not a property of the phenomes,
since each phenome is different. Fitness is a property of the given genome (the seed pattern that
is currently being evaluated), which is the same in each competition. Por otro lado, the produc-
tivity of a seed pattern is measured with a single seed pattern in a grid, playing GoL. There is no need
to repeat runs, since the final ashes will always be the same. We could say that productivity and
diversity are a product of the seed pattern (genome) or a property of the ashes (phenome).
The fitness in Model-S is an internal, relative fitness. The fitness of a seed is relative to the fitness
of the other seeds in the current population. With each new birth, the fitness of every seed in the
population is updated. It follows that the average fitness (the average fraction of games won) es
always 0.5. This means that we cannot compare the fitness of two seeds that are sampled from
different populations. We argued in our prior work (Turney, 2020b) that an internal, relative fitness
measure is more likely to result in open-ended evolution (Taylor et al., 2016) than an external, ab-
solute fitness measure, although we have not yet tested this hypothesis.
In order to evaluate the results from running Model-S, we need to create an external, absolute
fitness measure that will allow us to compare seeds from different populations. This external fitness
measure has no impact on the course of evolution in Model-S; it is only applied after a run of
Model-S, in order to allow us to interpret the results of the run. The external, absolute measure
we use is the fraction of Immigration Games that a seed wins when competing against seeds of
the same size (the same number of rows and columns) and the same density (the same number
of living cells in the initial seed pattern) (Turney, 2020b). A given seedʼs competitors are generated
by copying the given seed and then randomly shuffling the cells in the copied seed. This ensures that
the winner is winning due to the structure of the seed (the specific pattern of living and dead cells),
Artificial Life Volume 27, Número 1
33
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
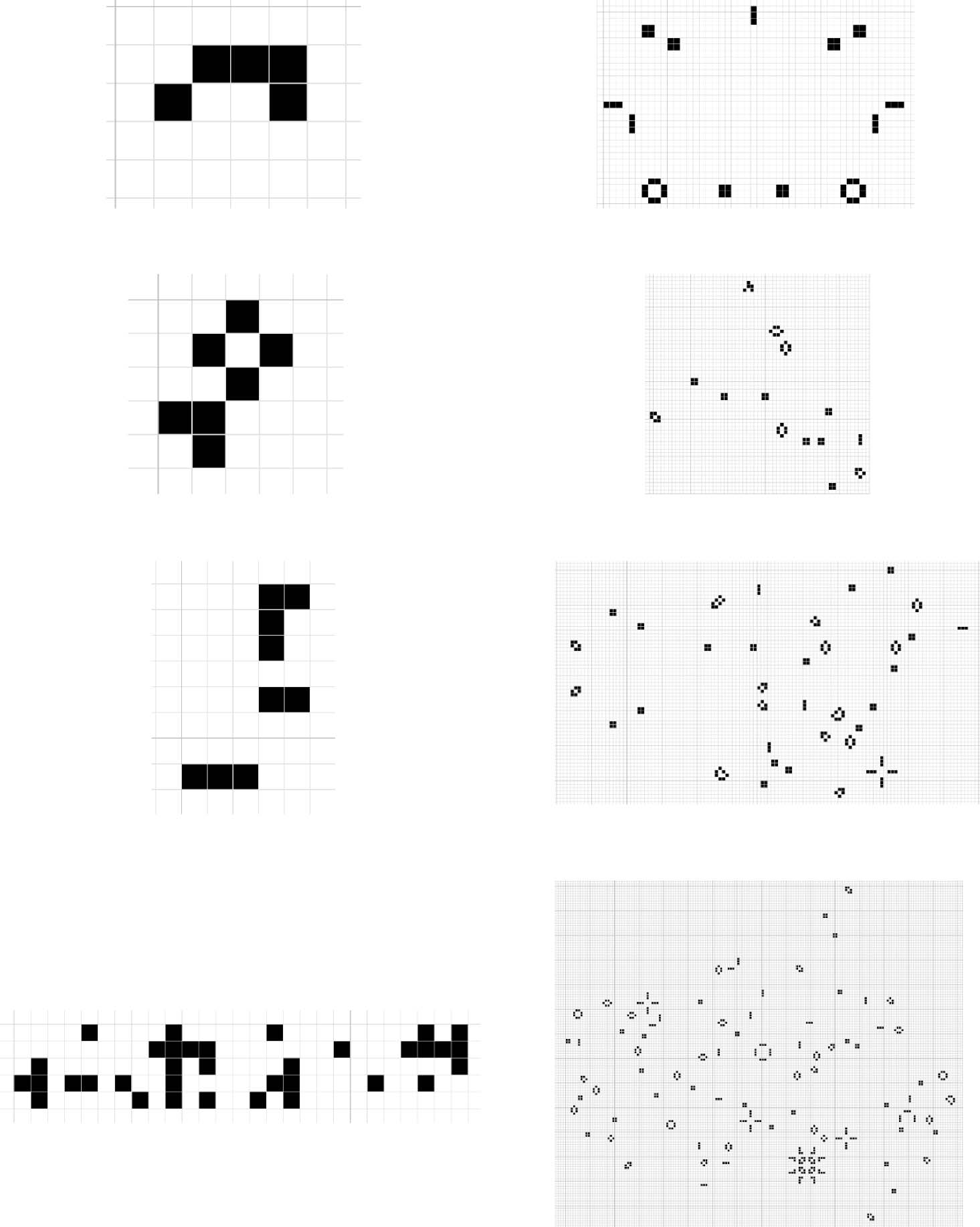
Cifra 3. Fitness and area of the four layers in the final generation. Correlation = 0.811, p value = 2.72E-12. This bar
graph presents the correlation between elite seed fitness in the final generation and elite seed area. The area of an elite
seed is measured by the product of the width and height of the seed. The correlation between fitness and area (0.811) es
high and significant. We evaluate the statistical significance of the correlation using a two-tailed Student t test for Pearson
correlations. The correlation is based on comparing two samples of 48 values each (4 layers × 12 carreras).
not the size or density of the seed. This is analogous to the sport of boxing, where fighters are
matched by weight.
Cifra 3 shows the external, absolute fitness of the elite seeds in the final generation for each of
the four layers of Model-S (blue bars). It also shows the areas (rows × columns) of the elite seeds in
the final generation for the four layers of Model-S (red bars). Each bar is the average of 12 separate
runs of Model-S. For each generation (cada 200 births), we sampled the top 50 most fit seeds (based
on their internal, relative fitness) and calculated their external, absolute fitness. We call these seeds the
elite seeds of the given generation. We saved all the elite seeds from our past work, which allows us
to use these seeds again in the current article.
We can see in Figure 3 that area is highly correlated with fitness (the correlation is 0.811 and it is
statistically significant), but increasing area does not cause increasing fitness. Increasing area is nec-
essary for increasing fitness, because increasingly complex structures require larger seeds to store
increasing amounts of information. But increasing area is not sufficient for increasing fitness, ser-
cause a specific kind of structure is required for fitness; eso es, a structure that grows quickly and
robustly during IG.
In our previous paper (Turney, 2020b), introducing Model-S, we presented three different argu-
ments in support of the claim that increasing area does not cause increasing fitness. We will only
mention one of those arguments here. Note that the external, absolute fitness is defined by com-
petitions between seeds that are matched in size and density. Por lo tanto, area cannot explain why the
average symbiotic seed in the final generation of Layer 4 has a fitness of 93.6% (ver figura 3). If area
were the deciding factor, the fitness would have to be 50%, because the two competitors have the
same area. The only difference between the symbiotic seed and its competitor is that the competitor
has been shuffled, which damages its structure but preserves its area. The structure is what deter-
mines the winner, not area.
3.2 Autopoiesis
There have been several automated censuses of ashes, counting the number of times each ash type
appears in soups (LifeWiki, 2021gramo). In the context of GoL, it seems that the term primordial soup
34
Artificial Life Volume 27, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
was first used by Robert Wainwright in 1971 (jardinero, 1971). More recently, Andrzej Okrasinski
made a census of the ashes arising from random soups from 2003 a 2008 (LifeWiki, 2021b),
Achim Flammenkamp made a census in 2004 (LifeWiki, 2021a), Nathaniel Johnston made a
census from 2009 a 2011 (LifeWiki, 2021i), and Adam Goucher has been collecting census infor-
mation since 2015, using his apgsearch software (Ash Pattern Generator Search; Goucher, 2021a;
LifeWiki, 2021C).
Goucherʼs apgsearch is currently the most popular tool for running random soups and classifying
their ashes. Apgsearch was designed for distributed computation, with many different users on the
Internet running random soups and uploading their ash censuses to a central server. Encima 100 users
have contributed to the census of ash objects (Goucher, 2021b; LifeWiki, 2021d).
Our previous work with Model-S (Turney, 2020b) demonstrated that Layer 4, the symbiotic
capa, when combined with the other three layers, resulted in a significant increase in the fitness
of the population. In that prior work, we did not analyze the specific means by which symbiosis
achieved greater fitness, except to provide evidence that the increased fitness was not merely due to
increased size in the initial seed pattern. We conjectured that autopoiesis was somehow respon-
sible (Turney, 2020b), but we did not know how to test this hypothesis at that time. The solution
became clear when we discovered Goucherʼs apgsearch software (Goucher, 2021a; LifeWiki,
2021C).
In the following experiments, we use a slightly modified version of Goucherʼs apgsearch to
identify and count the ash that is generated by evolved seed patterns from Model-S (Turney,
2021). The modification allows us to gather a census of ash for a single, specific seed pattern,
instead of accumulating counts from a large number of random soups, as the original version of
apgsearch does.
Apgsearch is currently at version 5.0, which is written in C++. We used apgsearch version 1.1,
which is written in Python and uses Golly, because Model-S is also written in Python and uses Golly.
We modified this version of apgsearch to analyze the ash from a single seed pattern, en vez de
aggregating ash statistics from very large samples of soups. The Python version of apgsearch is
slower than the C++ version, but speed was not an issue for our analysis of Model-S seeds.
Apgsearch has discovered 159,347 distinct types of ash objects so far (LifeWiki, 2021d). Beer
(2015) studied three of these objects (a block, a blinker, and a glider) in depth, to determine whether
they satisfy Maturana and Varelaʼs (1973) two conditions for autopoiesis, the closure condition and the
boundary condition, which Beer describes in this way:
The closure condition demands that the network of processes must produce the components
whose interactions generate and maintain that very same network. The boundary condition
demands that the spatial boundary that distinguishes an autopoietic system from its
background must itself be generated and maintained by the network of processes and in
turn must play a central role in enabling those same processes. (Beer, 2015, pag. 2)
Beer found that all three objects (bloques, blinkers, and gliders) satisfy the two conditions for
autopoiesis.
Blocks, blinkers, and gliders are the simplest members of their respective classes, still lifes, os-
cillators, and spaceships. Beer does not explicitly state that all still lifes, oscillators, and spaceships
satisfy the conditions for autopoiesis, but he hints that he believes this is true (Beer, 2015, pag. 15):
“This article has argued that such GoL entities as [énfasis añadido] bloques, blinkers, and gliders are
autopoietic with respect to the GoL physics because they satisfy both the closure and boundary
conditions of Maturana and Varelaʼs definition.” We believe that Beerʼs analysis generalizes to all
still lifes, oscillators, and spaceships (eso es, all ashes), but we do not have a formal proof of this.
We will proceed with the working hypothesis that Beerʼs analysis generalizes to all ashes, en el
expectation that the core ideas of our article will survive even if the working hypothesis may require
some fine-tuning.
Artificial Life Volume 27, Número 1
35
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
4 experimentos: Fitness, Productividad, Diversity, and Area
En esta sección, we present three new analyses of the data from our previous work with Model-S
(Turney, 2021). En la sección 4.1, we apply the modified apgsearch to the stored elite seeds from our
past work, in order to count the number of autopoietic structures that are produced (the quantity of
ashes). En la sección 4.2, we count the number of types of autopoietic structures the seeds produce
(the diversity of ashes). En la sección 4.3, we shuffle the seeds before we run apgsearch, to see whether the
quantity and diversity depend on the size of the seeds or the structure of the seeds (shuffling changes
structure but has no effect on size). The results show that the quantity and diversity of ash for a
given layer of Model-S is highly correlated with the fitness of that layer. Shuffling greatly reduces the
quantity and diversity of ashes, indicating that the quantity and diversity of ash is strongly affected by
the structure of the seeds. Sección 4.4 summarizes the results.
4.1 Fitness and Productivity
The productivity of a seed pattern is the quantity of ash it generates. Por ejemplo, the initial seed
pattern for Layer 4 En figura 2 yields 106 objects, as we can see in Table 1 (bottom right corner ).
For each of the four layers of Model-S, Cifra 4 shows the average quantity of ash produced by seed
patterns in the final generation. Each red bar in Figure 4 is the average productivity of 600 seed
patrones (50 elite seeds × 12 runs of Model-S).
If we compare the productivity (red bars) of the four layers in Figure 4 with the fitness (azul
barras), we see the same general pattern in productivity and fitness: The productivity and fitness of
Layer 1 (the uniform asexual layer) is low. Layer 2 (the variable asexual layer ) and Layer 3 (the sexual
capa ) have similar levels of productivity and fitness, with a slight advantage to Layer 2. Both Layers
2 y 3 are more productive and fit than Layer 1. Layer 4 (the symbiotic layer ) has the highest
productivity and fitness. Cifra 4 shows that the symbiotic layer is much more productive than
the other layers, and it is much more fit. The correlation between fitness and productivity in the
final generation is high (0.706) and statistically significant.
Cifra 4. Fitness and productivity of the four layers in the final generation. Correlation = 0.706, p value = 2.05E-08. Este
bar graph presents the correlation between elite seed fitness in the final generation and elite seed productivity in the final
generación. The productivity of an elite seed is measured by the number of ashes it creates. The correlation between
fitness and productivity (0.706) is high and significant. We evaluate the statistical significance of the correlation using a
two-tailed Student t test for Pearson correlations. The correlation is based on comparing two samples of 48 values each
(4 layers × 12 carreras).
36
Artificial Life Volume 27, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
The average seed pattern in the final generation of Layer 4 (the symbiotic layer ) generates 73.1
ashes (ver figura 4) and has an area of 94.1 (ver figura 3). Por lo tanto, the average ash generated by
the final generation of symbiotic seeds is 0.78 ashes per unit area (73.1 ashes / 94.1 área).
Catagolue reported a census on April 27, 2019, de 18,928,504,510,982 random soups, Resultando en
a total of 413,493,174,300,923 ashes (LifeWiki, 2021d). This yields an average of 21.8 ashes per
random soup. Each Catagolue soup begins with a random 16 × 16 binary matrix, which has an area
de 256 cells. Therefore the average ash generated by random soups is 0.09 ashes per unit area (21.8
ashes / 256 área).
The average final generation seed pattern of Layer 4 has an area that is 37% (94.1 área / 256 área)
of the area of Catagolueʼs random soups, and a productivity that is 335% (73.1 ashes / 21.8 ashes)
of the productivity of Catagolueʼs soups. Therefore the evolved seeds of Layer 4 son 9.1 times more
productive than random soups (335% / 37%). This shows that the evolved seeds in the final gen-
eration of Layer 4 are much more productive than random seeds. Sin embargo, Model-S was not de-
signed to produce ash; it was designed to play IG. With hindsight, it makes sense that winning IG
requires evolution to generate seeds that maximize the production of autopoietic structures.
4.2 Fitness and Diversity
The diversity of a seed pattern is the number of types of ashes it generates. Por ejemplo, the initial
seed pattern for Layer 4 En figura 2 yields 10 types of ashes, as we can see in Table 1 (bottom right
corner ). Cifra 5 shows the average diversity of ashes produced by seed patterns in the final gen-
eration of Model-S, for each of the four layers. Each red bar in Figure 5 is an average of 600 seed
patrones (50 elite seeds × 12 runs of Model-S).
If we compare the diversity (red bars) of the four layers in Figure 5 with the fitness (blue bars),
we see the same general pattern in diversity and fitness. This qualitative similarity between diversity
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 5. Fitness and diversity of the four layers in the final generation. Correlation = 0.834, p value = 1.79E-13. This bar
graph presents the correlation between elite seed fitness in the final generation and elite seed diversity in the final gen-
eration. The fitness of an elite seed is measured by competitions between the elite seed and random seeds of matched
size and density. The diversity of an elite seed is measured by the number of different types of ash that it yields. El
correlation between fitness and diversity (0.834) is high and significant. We evaluate the statistical significance of the
correlation using a two-tailed Student t test for Pearson correlations. The correlation is based on comparing two samples
de 48 values each (4 layers × 12 carreras).
Artificial Life Volume 27, Número 1
37
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
Mesa 3. This table lists the types of ash that are created in each layer of Model-S.
Rango
Layer 2
Rango
Layer 3
Rango
Layer 4
Rango
Layer 1
block
blinker
pond
beehive
glider
barco
boat
1
2
9
3
4
7
6
block
blinker
glider
beehive
pond
loaf
barco
tub
boat
pulsar
toad
1
2
4
3
9
5
7
8
6
21
11
block
blinker
pond
glider
beehive
boat
barco
loaf
long boat
tub
ship-tie
1
2
9
4
3
6
7
5
10
8
12
block
blinker
beehive
glider
boat
barco
loaf
tub
pond
toad
pulsar
loop
half-bakery
long boat
beacon
mango
ship-tie
barge
integral sign
1
2
3
4
6
7
5
8
9
11
21
49
15
10
13
16
12
14
25
19
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
total types
7
total types
11
total types
11
total types
total freq
7,799
total freq
10,490
total freq
10,187
total freq
43,865
num seeds
600
num seeds
600
num seeds
600
num seeds
600
freq / seeds
13.0
freq / seeds
17.5
freq / seeds
17.0
freq / seeds
73.1
Nota. For each layer, we collected all the ashes generated from the 600 seed patterns in generation 100 (50 elite seeds ×
12 carreras). For each type of ash in each layer of Model-S, we rank the ashes in order of decreasing frequency and we show
the rank of the type in Catagolue (LifeWiki, 2021F), where ashes are also ranked in order of decreasing frequency. En
general, the rank of each ash type in Model-S is similar to the rank in Catagolue, but we have marked some possible
outliers in italics. num = number of; freq = frequency.
38
Artificial Life Volume 27, Número 1
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
and fitness is confirmed by measuring the correlations. The correlation between diversity and fitness
es 0.834. The correlation is high and it is statistically significant.
We know from Section 4.1 that the evolved seed patterns in Layer 4 are much more productive
than random soups (there is a large quantitative difference between the number of seeds generated
in Layer 4 and the number of seeds generated in random soups—a factor of 9.1), but is there a
difference in the types of seeds in Layer 4 and the types of seeds in random soups (is there a qual-
itative difference in the distribution of seed types)?
Mesa 3 shows all the types of ash found in each layer of Model-S in the final generation. Fo-
cusing on Layer 1 as an example (the first column in Table 3), there are seven types of ash, ranked in
descending order of their frequencies in their layer in Model-S; eso es, the block is the most frequent
ash found in Layer 1, and the boat is the least frequent ash found in Layer 1. For each ash type in the
first column, there is a corresponding number in the second column that gives the rank of the ash
type in Catagolue (LifeWiki, 2021F) (from a census on July 15, 2017). Por ejemplo, the block is
ranked 1 in Catagolue, and the block is also first in Layer 1. Sin embargo, the pond is third in Layer 1,
yet it is ranked 9 in Catagolue.
For most entries in Table 3, the rank of an ash type that occurs in Model-S is approximately the
same as its rank in Catagolue. The boat is the seventh ash type in Layer 1 (see column 1), and it is the
sixth ash type in Catagolue (see column 2). The tub is the eighth ash type in Layer 4 (see column 7),
and it is the eighth type in Catagolue (see column 8). Sin embargo, although the ranks of ash types in
Model-S are mostly similar to their ranks in Catagolue, there are a few outliers. En mesa 3, tenemos
highlighted in italics one outlier in each of the four layers: the pond in Layers 1 y 3 (ranked third in
Model-S but ranked ninth in Catagolue), the pulsar in Layer 2 (ranked tenth in Model-S but ranked
twenty-first in Catagolue), and the loop in Layer 4 (ranked twelfth in Model-S but ranked forty-ninth
in Catagolue). We discuss this topic further in Section 5.
4.3 Productividad, Diversity, and Area
En la sección 3.1, we argued that, although the fitness of a seed pattern (its tendency to win Immigra-
tion Games) is correlated with the area of the seed pattern (ver figura 3), area does not cause fitness;
area merely provides room for complex structure (area allows fitness), and fitness is the result of a
certain type of complex structure (the type that wins games). In the current section, we present two
experiments that further support the claim that increasing area is not sufficient for increasing fitness.
Cifra 6 compares the productivity of the seeds before shuffling (the intact bar ) and after shuffling
(the shuffled bar). The productivity for each bar in Figure 6 is the average of 600 valores (12 runs × 50
elite seeds). The productivity of the shuffled seeds ranges from 7% a 14% of the productivity of
the intact seeds. This is evidence that the kind of structure that yields fitness is also the kind of
structure that yields productivity. By design, shuffling has no impact on area and density, so the
loss in productivity that we see here has nothing to do with area or density; it can only be caused
by the change in structure. The shuffled seeds in Layer 4 are less productive than the intact seeds in
Layer 1 (7.18 for Layer 4 shuffled versus 13.00 for Layer 1 intact—see Figure 6), although the seeds
in Layer 4 have much greater area than the seeds in Layer 1 (ver figura 3).
Cifra 7 compares the diversity of the seeds before shuffling (intact) and after shuffling (shuffled).
The diversity of the shuffled seeds ranges from 18% a 33% of the diversity of intact seeds. Shuf-
fling clearly reduces diversity, although the reduction is likely to be a consequence of the reduced
productivity of the shuffled seeds. As the number of ashes decreases, the number of types of ashes
also tends to decrease, so the reduced diversity is likely a side effect of reduced productivity. El
shuffled seeds in Layer 4 are less diverse than the intact seeds in Layer 1 (2.57 for Layer 4 shuffled
versus 3.05 for Layer 1 intact (ver figura 7), although the seeds in Layer 4 have much greater area
than the seeds in Layer 1 (ver figura 3).
The results in this section support the hypothesis that area alone is not sufficient for a seed pat-
tern to win IG. Area is required to encode complex seed patterns, but area is not sufficient for high
aptitud física. In the next section, we consider what is necessary for fitness.
Artificial Life Volume 27, Número 1
39
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
Cifra 6. The impact of shuffling on productivity. Ratio shuffled / Intact = 7% a 14%. Here we compare the productivity of
the final generations of the four layers with and without shuffling. The bar graph shows that shuffling greatly reduces pro-
ductivity. The key factor for productivity is the specific evolved structure of the seed pattern, not the area of the pattern.
4.4 Summary of Results
Mesa 4 shows the correlation between fitness and three other attributes: área, productivity, y
diversity. The table provides evidence that fitness is more correlated with diversity than it is with
area or productivity.
Cifra 7. The impact of shuffling on diversity. Ratio shuffled / Intact = 18% a 33%. Here we compare the diversity of the
final generations of the four layers with and without shuffling. The bar graph shows that shuffling greatly reduces diver-
sity. This supports the claim that, although increasing area is required for increasing diversity, the key factor for diversity
is the specific evolved structure of the seed pattern.
40
Artificial Life Volume 27, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
yo
/
_
a
_
0
0
3
3
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
9
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
Mesa 4. This table summarizes the correlations between fitness and other attributes of the four layers in Model-S.
Attributes
fitness and area
fitness and productivity
fitness and diversity
Correlación
0.811
0.706
0.834
p value
2.72E-12
2.05mi-08
1.79E-13
p value < 0.05?
Section
Yes
Yes
Yes
3.1
4.1
4.2
Figure
Figure 3
Figure 4
Figure 5
Note. Fitness was more correlated with diversity than with area or productivity. We evaluated the statistical significance
of the correlations using a two-tailed Student t test for Pearson correlations. All three correlations are statistically sig-
nificant. The correlations are based on comparing two samples of 48 values each (4 layers × 12 runs).
The results in Section 4.3 (see Figure 6 and Figure 7) tell us that, although increasing area is
necessary for increasing fitness, increasing area does not cause increasing fitness. Increasing area com-
bined with increasing diversity appears to be the mechanism by which evolution in Model-S achieves
high fitness in Layer 4. A wide range of types of ashes yields a high level of fitness, and a wide range
of types of ashes requires increasing area to accommodate new types.
In our previous work with Model-S (Turney, 2020b), we demonstrated that Layer 4 (symbiosis)
was able to evolve seed patterns with high fitness, but we did not explain what made these seed
patterns more fit than other patterns. A contribution of the current article is the insight that fitness is
the result of a diversity of autopoietic structures. This supports our view that Model-S is analogous
to biological life in eight ways (see Table 2), whereas our previous work supported only the first six
of these eight correspondences between characteristics of biological life and Model-S.
5 Future Work and Limitations
In Section 4.2, we noted that ponds, pulsars, and loops seem to be more frequent in Model-S than
we would expect from Catagolue. It seems possible that they are selected because they are more
stable than other objects, so they can survive close contact with other objects while playing IG,
whereas other objects are destroyed. We plan to test the stability of these objects relative to other
objects, and to look for other properties that might explain why ponds, pulsars, and loops seem to
be favored.
The score in IG is based on growth, which is measured by the increase in the number of living
cells of a given color (red or blue). In Section 4.1, we counted the number of ash-cells (autopoietic
structures), which is not the same as counting the number of living grid-cells (squares in the grid).
Selection in Model-S might be biased towards producing either many small ash-cells or a few large
ash-cells: For example, many small ash-cells might be less vulnerable in IG than a few large ash-cells.
We plan to look for a bias towards either smaller ash-cells or larger ash-cells. With random soups, we
see there is a bias towards smaller ash-cells; smaller ash-cells seem to occur more frequently than
larger ash-cells (LifeWiki, 2021f). To see whether IG prefers smaller ash-cells, we need to take into
account this natural bias for smaller ash-cells.
One possibility would be to alter IG so that the winner is determined by the number of ash-cells,
instead of the number of living grid-cells. If this alteration changes the distribution of ash types, then
we will learn something about how the method of scoring affects the kinds of ash we see.
Another area for future work is to explore other cellular automata rules, besides GoL, to see how
well the behaviors we have observed for GoL generalize to other rules. The rules for GoL belong to
the family of semitotalistic rules, a family with 262,144 members. Woodsʼ method for converting
GoL into a two-player game (LifeWiki, 2021e; Wainwright, 1971) generalizes to 8,192 members of
the family of semitotalistic rules. We estimate that about 4,000 of these rules will show the kind of
open-ended evolution that we have seen with Woodsʼ Immigration Game (Turney, 2020a).
Artificial Life Volume 27, Number 1
41
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
l
/
_
a
_
0
0
3
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
We mentioned in Section 3.2 that it has not yet been proven that all still lifes, oscillators, and
spaceships satisfy the conditions for autopoiesis. We expect that they all satisfy the conditions, but
the proof might be complicated. We leave this for future work.
6 Conclusion
Our previous article on Model-S (Turney, 2020b) presented a highly abstract and simplified model
of six aspects of biological life: (1) genomes, (2) phenomes, (3) natural selection, (4) competition, (5)
reproduction, and (6) mutualism. These shared abstract elements are given in the current article in
first six rows in Table 2. The previous article mentioned autopoiesis, but only as a topic for future
work. The current article presents that promised future work, adding (7) cells and (8) multicellular
organisms—the last two rows in Table 2. The key that made this extension possible was Goucherʼs
apgsearch software (Goucher, 2021a; LifeWiki, 2021c), which allows us to perform a census of ash
in experiments with Model-S. The apgsearch software permitted precise measurements of the pro-
ductivity and diversity of large quantities of ash (Section 4).
The experiments in Section 4 go beyond our previous work (Turney, 2020b) by providing some
insight into how Layer 4 is able to attain high levels of fitness. Past work demonstrated that certain
specific and relatively rare initial seed patterns were able to consistently win Immigration Games, but
we could not say exactly what property of these seed patterns enabled their success. Now we can say
that successful seed patterns are those that create a diversity of autopoietic structures (Figure 5 in
Section 4.2). It is likely that there is much more that can be said about successful seed patterns, but
we leave that as future work. Our hope is that more research will increase our understanding of the
analogies between evolving cellular automata and evolving biological life.
Acknowledgments
Thanks to the Artificial Life reviewers for their very helpful comments on the article. Thanks to
Andrew Trevorrow, Tom Rokicki, Tim Hutton, Dave Greene, Jason Summers, Maks Verver,
Robert Munafo, Brenton Bostick, and Chris Rowett, for developing Golly. Thanks to Adam Goucher
for developing apgsearch.
References
Beer, R. D. (2004). Autopoiesis and cognition in the Game of Life. Artificial Life, 10(3), 309–326. DOI: https://
doi.org/10.1162/1064546041255539, PMID: 15245630
Beer, R. D. (2014). The cognitive domain of a glider in the Game of Life. Artificial Life, 20(2), 183–206. DOI:
https://doi.org/10.1162/ARTL_a_00125, PMID: 24494612
Beer, R. D. (2015). Characterizing autopoiesis in the Game of Life. Artificial Life, 21(1), 1–19. DOI: https://doi
.org/10.1162/ARTL_a_00143, PMID: 25148547
Douglas, A. (2010). The symbiotic habit. Princeton University Press.
Gardner, M. (1970). Mathematical games: The fantastic combinations of John Conwayʼs new solitaire game
‘Life.’ Scientific American, 223(4), 120–123. DOI: https://doi.org/10.1038/scientificamerican1070-120
Gardner, M. (1971). Mathematical games: On cellular automata, self-reproduction, the Garden of Eden, and the
game ‘Life.’ Scientific American, 224(2), 112–117. DOI: https://doi.org/10.1038/scientificamerican0271-112
Goucher, A. P. (2021a, February 19). Apgsearch. Catagolue. https://catagolue.hatsya.com/apgsearch
Goucher, A. P. (2021b, February 19). Catagolue home page. https://catagolue.hatsya.com/home
LifeWiki. (2021a, February 19). Achim Flammenkampʼs census. LifeWiki. https://www.conwaylife.com/wiki
/Achim_Flammenkamp%27s_census
LifeWiki. (2021b, February 19). Andrzej Okrasinski ʼs census. LifeWiki. https://www.conwaylife.com/wiki
/Andrzej_Okrasinski%27s_census
LifeWiki. (2021c, February 19). Apgsearch. LifeWiki. https://www.conwaylife.com/wiki/Apgsearch
42
Artificial Life Volume 27, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
l
/
_
a
_
0
0
3
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Evolution of Autopoiesis and Multicellularity in the Game of Life
LifeWiki. (2021d, February 19). Catagolue. LifeWiki. https://www.conwaylife.com/wiki/Catagolue
LifeWiki. (2021e, February 19). Colourised Life: Immigration. LifeWiki. https://www.conwaylife.com/wiki
/Colourised_Life#Immigration
LifeWiki. (2021f, February 19). Most common objects on Catagolue. LifeWiki. https://www.conwaylife.com/wiki
/Most_common_objects_on_Catagolue
LifeWiki. (2021g, February 19). Soup. LifeWiki. https://www.conwaylife.com/wiki/Soup
LifeWiki. (2021h, February 19). Soup: Ash. LifeWiki. https://www.conwaylife.com/wiki/Soup#Ash
LifeWiki. (2021i, February 19). The online Life-like CA soup search. LifeWiki. https://www.conwaylife.com/wiki
/The_Online_Life-Like_CA_Soup_Search
Margulis, L. (1970). Origin of eukaryotic cells. Yale University Press.
Margulis, L. (1981). Symbiosis in cell evolution. W. H. Freeman.
Martin, B. D., & Schwab, E. (2013). Current usage of symbiosis and associated terminology. International Journal
of Biology, 5(1), 32–45. DOI: https://doi.org/10.5539/ijb.v5n1p32
Maturana, H. R., & Varela, F. J. (1973). Autopoiesis: The organization of the living. In H. R. Maturana & F. J.
Varela (Eds.), Autopoiesis and cognition (pp. 59–138). Reidel.
Maynard Smith, J., & Szathmáry, E. (1995). The major transitions in evolution. Oxford University Press.
McMullin, B. (2004). Thirty years of computational autopoiesis: A review. Artificial Life, 10(3), 277–295. DOI:
https://doi.org/10.1162/1064546041255548, PMID: 15245628
Poundstone, W. (2013). The recursive universe: Cosmic complexity and the limits of scientific knowledge. Dover
Publications.
Simon, D. (2013). Evolutionary optimization algorithms. John Wiley & Sons.
Taylor, T., Bedau, M., Channon, A., Ackley, D., Banzhaf, W., Beslon, G., Dolson, E., Froese, T.,
Hickinbotham, S., Ikegami, T., McMullin, B., Packard, N., Rasmussen, S., Virgo, N., Agmon, E., Clark, E.,
McGregor, S., Ofria, C., Ropella, G., … Wiser, M. (2016). Open-ended evolution: Perspectives from the
OEE workshop in York. Artificial Life, 22(3), 408–423. DOI: https://doi.org/10.1162/ARTL_a_00210,
PMID: 27472417.
Trevorrow, A., Rokicki, T., Rowett, C., Hutton, T., Greene, D., Summers, J., Verver, M., Munafo, R., Bostick,
B., & Lee, D. (2021, February 19). Golly. SourceForge. https://golly.sourceforge.net/
Turney, P. D. (2020a). Conditions for open-ended evolution in immigration games. arXiv, 2004(02720). https://arxiv.org
/abs/2004.02720
Turney, P. D. (2020b). Symbiosis promotes fitness improvements in the Game of Life. Artificial Life, 26(3),
338–365. DOI: https://doi.org/10.1162/artl_a_00326, PMID: 32772859
Turney, P. D. (2021, February 19). Source code for Model-S and measuring autopoiesis. GitHub. https://github.com
/pdturney/autopoiesis
Varela, F. J., Maturana, H. R., & Uribe, R. (1974). Autopoiesis: The organization of living systems, its
characterization and a model. BioSystems, 5, 187–196. DOI: https://doi.org/10.1016/0303-2647(74)90031-8
von Neumann, J. (1966). Theory of self-reproducing automata. (A. W. Burks, Ed.) University of Illinois Press.
Wainwright, R. T. (1971, June). Lifeline volume 2 [A quarterly newsletter for enthusiasts of John Conwayʼs Game
of Life]. LifeWiki. https://www.conwaylife.com/wiki/Lifeline_Volume_2
Whitley, D. (1989). The GENITOR algorithm and selective pressure: Why rank-based allocation of reproductive
trials is best. In D. Schaffer (Ed.), Third International Conference on Genetic Algorithms (pp. 116–121). Morgan
Kaufmann.
Whitley, D., & Kauth, J. (1988). GENITOR: A different genetic algorithm. In Proceedings of the Rocky Mountain
Conference on Artificial Intelligence, June 13–15, Denver, Colorado (pp. 118–130).
Artificial Life Volume 27, Number 1
43
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
7
1
2
6
2
0
2
0
4
1
5
a
r
t
l
/
_
a
_
0
0
3
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3