La simbiosis promueve el fitness
Improvements in the
Game of Life
Peter D. Turney*
Ronin Institute
peter.turney@ronininstitute.org
Palabras clave
Symbiosis, cooperation, open-ended
evolution, Game of Life, Inmigración
Juego, levels of selection
Abstract We present a computational simulation of evolving
entities that includes symbiosis with shifting levels of selection.
Evolution by natural selection shifts from the level of the original
entities to the level of the new symbiotic entity. In the simulation, el
fitness of an entity is measured by a series of one-on-one competitions
in the Immigration Game, a two-player variation of Conwayʼs Game
of Life. Mutation, reproduction, and symbiosis are implemented
as operations that are external to the Immigration Game. Porque
these operations are external to the game, we can freely manipulate
the operations and observe the effects of the manipulations. El
simulation is composed of four layers, each layer building on the
capa anterior. The first layer implements a simple form of asexual
reproduction, the second layer introduces a more sophisticated form of
asexual reproduction, the third layer adds sexual reproduction, y el
fourth layer adds symbiosis. The experiments show that a small amount
of symbiosis, added to the other layers, significantly increases the
fitness of the population. We suggest that the model may provide new
insights into symbiosis in biological and cultural evolution.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
1 Introducción
There are two main definitions of symbiosis in biology, (1) symbiosis as any association and (2) symbiosis as
persistent mutualism [7]. The first definition allows any kind of persistent contact between different
species of organisms to count as symbiosis, even if the contact is pathogenic or parasitic. El segundo
definition requires that all of the participating organisms must benefit for the persistent contact to
count as symbiosis [7]. One reason for this disagreement over definitions is that there is a large gray
zone of cases where it is difficult to know whether all of the participants benefit from the association.
Por ejemplo, in many cases the presence or absence of mutual benefit can vary over time, depending
on environmental factors. There is also a problem with deciding how long mutual benefit must persist
before we can call it persistent mutualism. The desire for simplicity explains the appeal of symbiosis as any
association. Por otro lado, the presence of mutual benefit is theoretically interesting, cual
motivates the narrower definition of symbiosis as persistent mutualism.
In this article, we introduce a model of symbiosis that focuses on symbiosis with shifting levels of selection.
In such cases the symbiotic organisms are no longer able to reproduce as separate parts; they must
* Autor correspondiente.
© 2020 Instituto de Tecnología de Massachusetts.
Publicado bajo una atribución Creative Commons
4.0 Internacional (CC POR 4.0) licencia.
Artificial Life 26: 338–365 (2020) https://doi.org/10.1162/artl_a_00326
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
reproduce as a whole. A paradigmatic example of shifting levels of selection is the merging of prokary-
otes to form eukaryotes [13-15]. Natural selection can no longer act on the component prokaryotes; él
can only act on the whole eukaryote that contains them. Selection has shifted from the parts to the
entero. We call our simulation Model-S (Model of Symbiosis). The source code for Model-S is available
for downloading [30].
Taking the perspective of the field of evolutionary optimization algorithms [21], we view symbiosis
with shifting levels of selection as the product of a genetic operator, similar to crossover, selección, o
mutation. Following lkegamiʼs [10] pioneering work, we call this genetic operator genetic fusion, o
simply fusion [29]. Fusion takes as input the genomes of two distinct entities that experience selection
separately and produces as output a genome for a merged entity that experiences natural selection as a
entero. In this article, we seek to understand what happens to the course of evolution when fusion
occurs repeatedly, in the same way that one might experiment with different forms of mutation or
crossover to understand how they affect the course of evolution.
In Model-S, we can precisely calculate the fitness of both the parts and the wholes. This allows us to
perform experiments that would be very difficult, if not impossible, in biology. When running Model-S,
we can choose between symbiosis as any association and symbiosis as persistent mutualism. Para
symbiosis as any association, the fusion operator in Model-S can arbitrarily select two organisms and force
them to work together as a whole. Selection will take place at the level of the whole, not at the level of
the parts. Even if the parts are more fit as individual parts than they are as a merged whole, evolution
with mutation and selection at the level of the whole may be able to adapt them, over many generations,
so that they work well together. For symbiosis as persistent mutualism, the fusion operator can arbitrarily
select two organisms and compare their fitness as separate parts with their fitness as a fused whole. Si
the whole is more fit than the parts, then fusion proceeds. If the whole is less fit than the parts, entonces
fusion is canceled. This ensures that fusion proceeds only when there is mutual benefit (increased
aptitud física) for both component organisms. With Model-S, we do not need to choose between the two
main definitions of symbiosis in biology. We can simulate both options.
Model-S has four layers: (1) simple asexual reproduction with genomes of constant size, (2) asexual
reproduction with genomes of variable size, (3) sexual reproduction with crossover, y (4) symbiosis
by fusion. The separation of the model into layers is not intended to reflect evolution in nature; el
motivation for the layers is to be able to measure the contributions of each layer to the evolution of a
población.
Evolution by natural selection requires variation, heredity, and differential fitness (selección) [6, 9]. En
Model-S, differential fitness is based on one-on-one competitions in the Immigration Game, cual
was invented by Don Woods and described in Lifeline in 1971 [31]. The Immigration Game is a two-player
variation of the Game of Life, invented by John Conway and presented in Scientific American in 1970 [8].
The Game of Life is played on an infinite, two-dimensional grid of square cells [19]. Each cell is either
muerto (estado 0) or alive (estado 1). The state of a cell changes with time, based on the state of its eight nearest
neighbors (the Moore neighborhood). Time passes in discrete intervals, and the states of the cells at time
t uniquely determine the states of the cells at time t + 1. The initial states at time t = 0 are chosen by
the player of the game; the initial states form a seed pattern that determines the course of the game,
analogously to the way an organismʼs genome determines its phenome. The rules for updating states
are compactly expressed as B3/S23: A cell is born (switches from state 0 to state 1) if it has exactly three
living neighbors. A cell survives (remains in state 1) if it has two or three living neighbors. Otherwise it dies.
The Immigration Game is almost the same as the Game of Life, except that there are two different
live states (estados 1 y 2) [31]. The two live states are usually represented by red and blue colors. El
rules for updating remain B3/S23, but there are new rules for determining color: (1) Live cells do not
change color unless they die. (2) When a new cell is born, it takes the color of the majority of its neigh-
bors. Since birth requires three live neighbors, there is always a clear majority. The initial states at time
t= 0 are chosen by the two players of the game; one player makes a red seed pattern and the other
player makes a blue seed pattern. The players agree on a time limit, given by a maximum value for t.
In our past work with the Immigration Game, we specified that the player with the most living
cells of their color at the end of the game is the winner. Sin embargo, this stipulation results in a bias
Artificial Life Volume 26, Número 3
339
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
towards seed patterns with many living cells. Por lo tanto, instead of counting the total number of
living cells of a given color at the end of the game, we now count the increase in the number of living
cells from the initial seed pattern to the end of the game; eso es, the final count minus the initial
count. If there is a decrease in the number of living cells, we give the player a score of zero. The player
with the largest increase in living cells of their color is the winner. Ties are allowed.
If states 1 y 2 were displayed with the same color (decir, negro), playing the Immigration Game
would appear identical to playing the Game of Life. The different colors are simply a way of keeping
puntaje, to turn the Game of Life into a competitive game.
The original rules of the Immigration Game allow the human players to intervene in the game as
it progresses [31], but we have no use for interventions in our simulations. The original rules also use
a finite toroidal grid of 25 × 25 cells instead of an infinite grid. The motivation for a finite grid is that
the limited space for growth forces the seeds to interact with each other and reduces the amount of
computation required. We use a finite toroidal grid, but we make the size of the toroid proportional
to the size of the initial seeds, so that there is no fixed limit on the size of the initial seeds.
The four layers of evolution in Model-S are external to the Immigration Game. It is possible to
build a replicator inside the Game of Life [1], but all current Game of Life replicators are much too
slow for practical experiments with simulations of evolution. An advantage of having the mecha-
nisms of evolution external to the Immigration Game is that it enables experimentation with a clean
separation of the independent variables (the external evolutionary mechanisms) and the dependent
variables (the fitness scores from the Immigration Game).
We chose the Immigration Game for our model of symbiosis because, as a two-player competitive
juego, it provides a built-in way of calculating fitness; as a Life-like game, it is well suited for modeling
biology; and it turns out that implementing fusion in the Immigration Game is simple and elegant.
Given two seeds as input to the fusion operator, we join them together side by side with a one-column
space to serve as a buffer. They are then treated the same way as any other seed; eso es, they live, die,
and reproduce as a unit. Selection takes place at the level of the whole unit (the higher level).
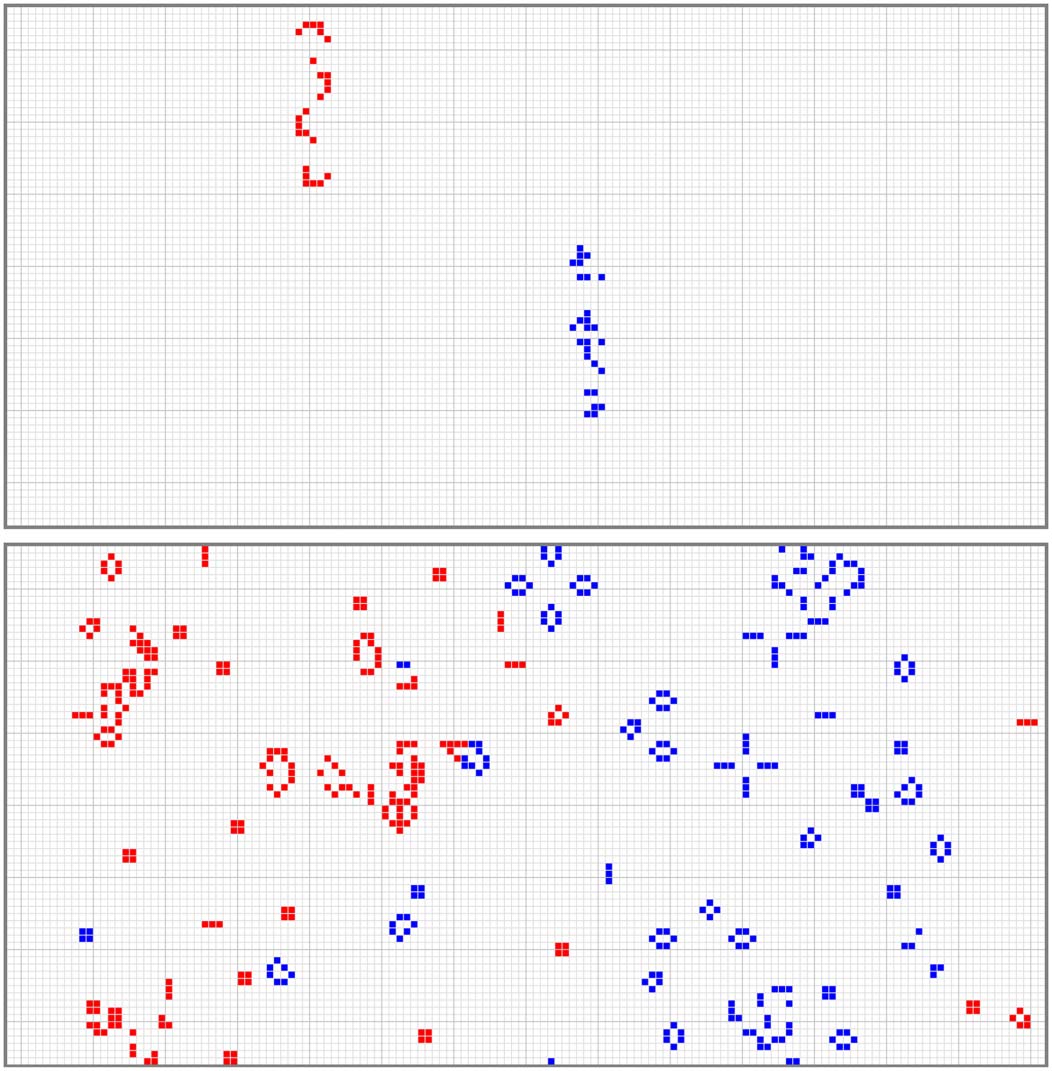
Cifra 1 shows an example of an Immigration Game. The first image shows the competing seeds
at time t = 0, and the second image shows the states of the cells when the time limit has been reached.
When they are not competing, the seeds are stored with only two states (0 y 1); they are only tem-
porarily assigned colors (estados 1 o 2) for the purpose of playing the Immigration Game to obtain a
fitness score.
As a model of biological evolution, a seed pattern in Model-S corresponds to a genome, a static
encoding of genetic information. When a seed is embedded in an Immigration Game, the dynamic
sequence of patterns created as the game runs corresponds to the dynamic development of the
phenome from the genome. When two seeds compete in an Immigration Game, this corresponds
to two living organisms (two phenomes), growing and competing for limited space. In an Immigration
Juego, the seed that grows more is the fitter of the two seeds, and the fitter seed is more likely to be
chosen by the tournament selection algorithm for reproduction, as in biological evolution, donde el
organism that grows more is likely to have more offspring.
Fusion is a genetic operator that combines two seeds to make a new genome, with a size that is
approximately the sum of its two parts. When the new fused seed is embedded in an Immigration Game,
as the game runs, the two parts interact. Model-S keeps a record of the fitness scores for the parts and
for the new fused whole, so it is possible for Model-S to distinguish between symbiosis as any asso-
ciation and symbiosis as persistent mutualism. Any fused seed, when running in an Immigration Game,
can be viewed as an instance of symbiosis as any association. At the end of an Immigration Game, nosotros
can check the final fitness score for a given fused seed, to determine whether the game counts as an
instance of symbiosis as persistent mutualism. If the score for the whole is greater than the score of
either part, then we have mutualism; eso es, both parts have benefited from their association.
Open-ended evolution is defined as evolution that is [25, pag. 409] “capable of producing a continual
stream of novel organisms rather than settling on some quasi-stable state beyond which nothing
fundamentally new occurs.” The main contributions of our article are (1) a computational simulation
of symbiosis through genetic fusion as a mechanism for evolution, (2) evidence that fusion has a
340
Artificial Life Volume 26, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Cifra 1. The first image above shows the initial state of an Immigration Game, and the second image shows the final
estado, when the game reached its time limit. The first image contains two competing seeds: a red seed (24 × 5 block, 24
live cells, density 0.200) and a blue seed (24 × 5 block, 27 live cells, density 0.225). The second image reveals that blue
won the game. Blue grew by 209 live cells (de 27 a 236), and red grew by 204 live cells (de 24 a 228). Both seeds
were the fittest seeds in the final generations of two different runs of Model-S, in which both runs used all four layers of
el modelo. The second image is a typical example of the final state of a game.
significant effect on evolution when combined with mutation and reproduction, (3) support for the
hypothesis that symbiosis by fusion can sustain open-ended evolution, y (4) source code [30] para
replicating and extending the results presented here.
En la sección 2, we discuss related work. Sección 3 outlines the principles that guided the design of
Model-S and describes each of the four layers of the model. Sección 4 presents four sets of experiments:
(1) We add the layers one by one, to see what each layer contributes to the model. (2) We test the fusion
operator to determine how much fitness is due to the increased size of fused seeds versus how much is
due to mutually beneficial interaction of fused seeds. (3) We compare human-designed Game of Life
seed patterns with evolved seed patterns from Model-S. (4) We introduce an alternative fitness measure
to evaluate whether fusion might support open-ended evolution. En la sección 5, we discuss the implica-
tions of the experimental results. Sección 6 examines limitations and possibilities for future research.
We conclude in Section 7.
2 Trabajo relacionado
The significance of hierarchical, part–whole structure in biology and culture was emphasized by Simon
[22] and Koestler [12] in the 1960s. In the 1970s, Margulis [13, 14] argued for the importance of sym-
biosis in evolution. Maynard Smith and Szathmáry [15] observed the role of symbiosis in several of the
major transitions in the evolution of life on Earth and discussed the problem of levels of selection.
Ikegami [10] introduced an influential model of symbiosis for game strategies in the Erroneous
Iterated Prisonerʼs-Dilemma game. Game strategies are represented with tree structures that choose
Artificial Life Volume 26, Número 3
341
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
to cooperate or defect, based on the past moves of an opponent. Strategy trees evolve by mutation,
selección, and symbiosis. Symbiosis involves grafting one tree onto a randomly selected leaf of another
árbol. The experiments show that there is a long-term evolutionary trend towards increasingly complex
strategy trees.
Ikegami [10] defines symbiosis as mutual cooperation (eso es, symbiosis as persistent mutualism [7]),
but his experiments do not actually test whether his genetic fusion operator results in mutual benefit.
It is not clear which of the two main definitions of symbiosis should be applied to Ikegamiʼs model.
His focus is on novelty and diversity in the population, rather than the fitness of individuals.
Watson and Pollack [32] hypothesized that symbiosis is particularly suitable for a specific type of
fitness landscape, where there is a kind of fractal structure that is evolutionarily challenging at all scales.
They demonstrated that evolution by mutation and selection without symbiosis becomes increasingly
difficult in this kind of fitness landscape, but adding symbiosis to mutation and selection allows
ongoing adaptation.
McShea and Brandon [16] assert that the increase in complexity of organisms over time is largely
due to heritable variation in part–whole hierarchies. Sin embargo, their theory of increasing complexity is
based only on the horizontal spread of the hierarchy (the number of parts at the same level), and has
nothing to do with the vertical depth of the hierarchy (the number of levels).
Banzhaf et al. [2] define a meta-model that can be used to identify levels of structure in a system.
Por ejemplo, they discuss how their meta-model could be applied to the Game of Life. A level-0 meta-
model would view the Game of Life at the level of individual cells and their states. A level-1 meta-
model would view the game at the level of common entities that appear in the game as repeating
patterns of cells and states, such as gliders, spaceships, and oscillators (these names are familiar to players
of the Game of Life [19]). A level-2 meta-model would include larger structures that are composed of
level-1 structures, etcétera. Model-S is intended to be a model, not a meta-model. In future work, el
meta-model of Banzhaf et al. [2] could be applied to analyze Model-S, but we do not pursue that here.
Moreno and Ofria [17] create a computational simulation in which cell-like organisms coordinate
their activities in ways that increase their reproduction. As the simulation runs, larger groups of or-
ganisms cooperate, sharing resources and dividing their labor. Sin embargo, their simulation is limited to
two hierarchical levels. It was not designed with a mechanism for automatically adding new levels.
The work of Beer [3–5] on modeling autopoiesis (self-production and self-maintenance) en el
Game of Life is also relevant here. Our expectation is that autopoiesis will enable a seed to maintain
itself better in the presence of disruptive competition. We conjecture that the entities that evolve in
Model-S will show increasing degrees of autopoiesis as the number of generations in the simulation
aumenta, but we have not yet tested this hypothesis.
3 Description of the Model
Model-S uses the open-source Golly software for running the Immigration Game [26]. Golly is de-
signed to support extensions using the scripting languages Lua and Python. Model-S was implemented
as an open-source Python extension of Golly [30].
Most of this section is concerned with presenting the design of the four layers of Model-S, but we
first discuss the principles behind the design. The principles should help to explain some of the design
decisiones.
3.1 Conditions for Open-Ended Evolution
Brandon [6, páginas. 5–6] states that the following three components are crucial to evolution by natural
selección:
1. Variation: There is (significativo) variation in morphological, physiological, and behavioral
traits among members of a species.
342
Artificial Life Volume 26, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
2. Heredity: Some traits are heritable, so that individuals resemble their relations more than
they resemble unrelated individuals, and offspring resemble their parents.
3. Differential fitness: Different variants (or different types of organisms) leave different numbers
of offspring in immediate or remote generations.
In the literature, differential fitness is often called selection. Godfrey-Smith [9] lists the same three compo-
nents, calling them conditions for evolution by natural selection.
Past work in artificial life has shown that, although these conditions are sufficient for evolution, ellos
are not sufficient for open-ended evolution [25]. We are particularly interested in symbiosis by fusion
because we believe that it may be one of the conditions for open-ended evolution (in addition to
variación, heredity, and differential fitness). Sin embargo, it seems likely that there may be other conditions
that are required for open-ended evolution. Since the necessary and sufficient conditions for open-
ended evolution are not yet known, we chose to use a relatively large number of additional conditions,
some of which might be unnecessary for open-ended evolution:
1. Symbiosis, fusion, and cooperation: There should be a mechanism for symbiosis with a shift in
the level of selection from the parts to the whole [15]. Hence Layer 4 adds fusion to Model-S.
2. Biotic selection: Selection can be based on an organismʼs biological environment (competitors,
predators, enfermedad, etc.) or other aspects of its environment (sunlight, agua, soil, shelter, etc.).
Our intuition is that competition with other organisms (biotic selection) is a particularly strong
likely to encourage open-ended evolution. This motivates using the
form of selection,
Immigration Game (a competitive game) for calculating differential fitness.
3. Relative fitness: The fitness of an organism is relative to the fitness of other organisms, especially
members of the same population. There is no absolute fitness. (This is related to biotic selection.)
4. Unlimited genome size: A genome with a limited size must contain a limited amount of
información, which implies a finite bound on the space of possible organisms [27, 28].
(One way around this limit is cultural evolution, where information is stored outside
the genome.) Hence Layer 2 of Model-S adds variable size for seed patterns.
5. Gene transfer: There should be some method for sharing genes (such as plasmids or sexual
reproduction) beyond replication (asexual reproduction). Hence Layer 3 introduces sexual
reproduction.
6. Genotype and phenotype: Open-ended evolution may require a distinction between genotype
and phenotype. In the Game of Life, we view the initial seed pattern as the genotype. El
growth or decline of the seed over time, as the game runs, is the development of the
phenotype from the genotype.
7. Speciation: Diversity may require reproductive boundaries (distinct species). Without sufficient
diversity, organisms may be trapped in a local optimum. Layer 3 adds reproductive boundaries
by requiring potential mates to have a certain degree of genetic similarity.
It will take much work to validate all these conditions. We leave this as future work. In this article, nosotros
have limited our scope to showing that fusion is a useful genetic operator that may contribute to achieving
open-ended evolution in a simulation. Other lists of conditions for open-ended evolution have been given
for biological evolution [23], cultural evolution [18], and natural and artificial evolutionary systems [24].
3.2 Layer 1: Uniform Asexual Layer
Model-S has several parameters for controlling its behavior. We will introduce the parameters as they
are needed in explanations. All parameter names contain an underscore symbol and use a sans serif
font. A full list of the parameters and their values is given in Table 1 en la sección 4.1.
Artificial Life Volume 26, Número 3
343
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Model-S uses a GENITOR-style algorithm [33, 34] with one-at-a-time reproduction, a constant
population size, and rank-based tournament selection. An individual in the population is represented
as an object (a data structure) containing a binary matrix that specifies a seed pattern and an array of
real values that stores a history of the results of its competitions with all other individuals in the
población. The population is an array of pop_size individuals.
Children are born one at a time. Each new child replaces the least fit member of the population,
maintaining a constant population size. When pop_size children have been born, we say that one
generation has passed. A run of Model-S begins with generation zero and lasts until generation
num_generations. A run ends when pop_size × num_generations children have been born.
In generation zero, Model-S starts with a population in which the binary matrices are randomly
initialized. The probability of ones in these matrices is given by seed_density, which we set to 0.375,
based on the advice of Johnston [11]. When all the matrices are initialized, we then initialize the
history of competition results by playing a series of Immigration Games, pairing every individual
against every other individual num_trials times. The fitness of an individual is the fraction of games
that it wins. Every win by one individual is balanced with a loss by another individual. It follows that
the average fitness of the population is always 0.5. Fitness is relative to the population, not absolute.
A new child is created by first selecting a parent, using tournament selection. We randomly select
tournament_size individuals from the population, and the fittest member of this sample is chosen as a
parent. The parent is copied to make a child. The child is then mutated by randomly flipping bits in the
binary matrix, where the probability of flipping a bit is mutation_rate. We force at least one bit to flip,
regardless of mutation_rate, so that a child is not identical to its parent, to maintain diversity in the pop-
ulación. The new child replaces the least fit member of the population, and the histories of competition
results are updated by pairing every individual against the new child in a new series of Immigration
Games. This is summarized in Figure 2.
The space and time allowed for an Immigration Game depend on the two seeds that are competing.
Open-ended evolution requires the limits on space and time to increase as the sizes of the individuals
increase: Fixed limits would set a bound on the possible variety of games. Given two seeds, let max_size
be the maximum of the number of rows and columns in the seeds; eso es, the largest width or height.
Three parameters determine the space and time allowed for the two seeds: width_factor, height_factor,
and time_factor (ver tabla 1 en la sección 4.1). The width of the Golly toroid is set to max_size times
width_factor. The height of the toroid is set to max_size times height_factor. The maximum time (el
Cifra 2. The flowchart describes the process for uniform asexual reproduction. This process is a subroutine in a loop
that produces a series of new individuals. For each individual that is added to the population, another is removed; hence
this is a steady-state model with a constant population size. Uniform asexual reproduction takes the input individual and
generates a mutated copy as the output.
344
Artificial Life Volume 26, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
number of time steps in the game) is set to the sum of the width and height of the toroid, multiplied by
time_factor.
We describe Layer 1 as the uniform asexual layer because reproduction is asexual (each child has only
one parent) and the size of the seed pattern matrix is uniform (the size is the same for every individual in
every generation). Layer 1 is intended as a minimalist baseline evolutionary system. The following layers
are expected to improve upon Layer 1.
3.3 Layer 2: Variable Asexual Layer
Layer 2 is like Layer 1, except we now have three different kinds of mutation: (1) With probability
prob_flip, the child will be mutated by flipping bits, according to mutation_rate. (2) With probability
prob_shrink, the child will be mutated by removing an outer row or column from the binary matrix.
(3) With probability prob_grow, the child will be mutated by adding an outer row or column to the
binary matrix. These three kinds of mutation are mutually exclusive; eso es, the sum of prob_flip,
prob_shrink, and prob_grow is one. This is summarized in Figure 3.
There is a minimum size for matrices (min_s_xspan columns and min_s_yspan rows), to limit how
small a matrix can become by shrinkage. If growth is selected, the newly added column or row is
initialized by randomly setting bits, where the probability of ones is given by seed_density.
We want growth in the model, so that there is no upper bound to the amount of information that can
be stored in a genome (the binary matrix of an individual). The motivation for shrinkage is to see whether
growth is a consequence of increased fitness or it is due to random drift in the space of genomes. If the
growth is due to random drift, then it should eventually flatten out as it balances with shrinkage.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 3. Layer 1 produces individuals of uniform size, whereas Layer 2 allows variable size. A random number is gen-
erated between 0 y 1. The value of the random number determines whether the individual will shrink in size, grow in
tamaño, or be passed on to Layer 1, where it will copy the size of its parent and mutate by flipping bit values.
Artificial Life Volume 26, Número 3
345
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
3.4 Layer 3: Sexual Layer
Layer 3 adds sexual reproduction to Model-S. The first parent is chosen by tournament selection,
just as in Layers 1 y 2. The second parent is chosen by looking for all individuals in the population
with a degree of similarity to the first parent that is between min_similarity and max_similarity. El
similarity of two individuals is measured by the fraction of corresponding matrix cells that have
the same binary values. The similarity of two matrices is defined as zero if the matrices have different
numbers of rows and columns. The second parent is chosen by tournament selection from this
reduced sample of potential mates. This is summarized in Figure 4.
If there are no suitable mates with the required degree of similarity, Layer 3 passes the first parent
on to Layer 2, for asexual reproduction. Many organisms in nature can reproduce either sexually or
asexually, depending on the availability of suitable mates.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 4. Layer 3 introduces sexual reproduction, where part of one individualʼs matrix is combined with part of another
individualʼs matrix. The individuals are neither male nor female; any individual can mate with any other individual, so long as
they are sufficiently similar. After mating, the child individual is passed on to Layer 2, where it grows, shrinks, or flips bits.
346
Artificial Life Volume 26, Número 3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
When two parents have been selected, they produce a child by crossover. Primero, we choose between
crossing rows and crossing columns, with equal probability. If rows are chosen, we randomly choose a
horizontal crossover point and we make a new child by combining the rows above the crossover point
from one parent and the rows below the crossover point from the other parent. Asimismo, if columns
are chosen.
There is a limit to the variety that can be produced by crossover alone, especially in the case of
small populations. Por lo tanto, after crossover takes place in Layer 3, we pass the child on to Layer 2,
where it undergoes bit flipping, shrinkage, or growth.
3.5 Layer 4: Symbiotic Layer
Layer 4 adds fusion and fission to Model-S. Primero, a seed is chosen by tournament selection, just as in
Layers 1, 2, y 3. Then there are three possibilities: (1) With probability prob_fission, the chosen seed will
be split in two. One part will enter the population and the other part will be discarded. (2) With prob-
ability prob_fusion, a second seed is chosen by tournament selection and the two seeds will be fused
together. (3) If neither fusion nor fission is chosen, then Layer 4 will pass control over to Layer 3.
We expect that prob_fusion and prob_fission will be set to values near zero, so the most likely event
is that Layer 4 will pass control on to Layer 3 for sexual reproduction, reflecting the fact that fission
and fusion are relatively rare in nature. This is summarized in Figure 5.
When fission is chosen, we look for the sparsest row or column in the binary matrix. The matrix
is then divided into two parts along the sparsest row or column. One part is discarded, including the
sparsest row or column, and the remaining part enters the population as a new individual.
When fusion is chosen, the two seeds are randomly rotated and then joined side by side with one
column of zeros between them. The column of zeros is intended to act as a buffer, to reduce the
potential for conflict or interference between the two seeds when they are joined. The column of
zeros also acts as a marker to provide a natural splitting point for possible fission events in the
future. Mutation will gradually flip some of the bits in this column of zeros, turning them into ones.
Fission and fusion in Layer 4 are somewhat analogous to shrinkage and growth in Layer 2. El
motivation of fission is to counterbalance fusion, just as shrinkage counterbalances growth. El
expectation is that, if fusion does not contribute to fitness, then any random drift towards increased
size due to fusion will eventually be limited by fission.
Layers 1, 2, y 3 are forms of reproduction, in which a child is like its parent (in the case of Layers 1 y 2)
or parents (in the case of Layer 3). Fusion in Layer 4 is analogous to sexual reproduction in Layer 3, in that
two seeds are involved in the production of a new seed, but the size of the new genome is approximately
the sum of the sizes of the two original genomes. The “child” of fusion is not similar to its “parents.” We
will see in the experiments in Section 4 that Layer 4 behaves quite differently from Layer 3.
Model-S is designed to increase the time limit for the Immigration Game when the seeds are larger, en
order to give more time for a clear winner to emerge from the game (mira la sección 3.2); de este modo, the simu-
lation slows down as the seeds become larger. Layer 4 tends to result in a rapid increase in the size of
seeds over the course of a run of Model-S. This is a positive outcome from a theoretical point of view,
since it confirms our expectations for symbiosis, but it is problematic from a practical point of view,
because the simulation runs very slowly. For this practical reason, we have designed Model-S with a
linear upper bound on the area of seeds. The area of a seed is the number of columns in the seedʼs
binary matrix multiplied by the number of rows. The upper bound is set using the parameters max_
area_first and max_area_last, where max_area_first is the maximum area of a seed in the first generation
and max_area_last is the maximum area of a seed in the last generation. For generations between the first
and last, the maximum area is determined by linear interpolation. If the fusion of two seeds would
exceed the linear upper bound on area, then Model-S prevents the fusion from happening and passes
control to Layer 3, sexual reproduction.
Layer 4 includes two binary flags for experiments with modifications to the operation of Layer 4. El
first flag, symbiosis_flag, can have the value 0 o 1. When symbiosis_flag is set to 0 (the default setting), el
fusion operator will join two seeds to make a new whole without regard to whether the component
Artificial Life Volume 26, Número 3
347
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 5. Unlike Layers 1, 2, y 3, Layer 4 is not a form of reproduction. With fusion, a new individual is created by
fusing two individuals. With fission, a new individual is created by breaking an individual into two parts. Only one of the
two parts is kept. We set the probability of fission higher than the probability of fusion in order to see whether selection
can overcome this bias towards fission. Note that fission and fusion are much less likely than sexual reproduction.
seeds benefit; eso es, Model-S will operate with symbiosis as any association. When symbiosis_flag is set to 1,
the fusion operator will temporarily join two seeds to make a new whole, to calculate the fitness of the
entero. If the fitness of the whole is greater than the fitness of either part, then the new fused seed enters
la población. De lo contrario, the new fused seed is rejected and Model-S passes control over to Layer 3,
sexual reproduction. De este modo, when symbiosis_flag is set to 1, Model-S will operate with symbiosis as persistent
mutualism [7]. Eso es, with this setting, Model-S will only allow selection to shift from the level of the
parts to the level of the whole when such a shift yields mutual benefit.
The second binary flag, fusion_test_flag, can have the value 0 o 1. When fusion_test_flag is set to 0 (el
default setting), fusion proceeds as usual. When fusion_test_flag is set to 1, one of the two seeds is randomly
shuffled before the two seeds are fused. The intention is to disrupt the structure of the seed without altering
its summary statistics, such as size, forma, and density. Shuffling takes a seed and swaps the values in the cells,
348
Artificial Life Volume 26, Número 3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
resulting in a new seed with the same size (same number of cells), the same shape (same number of rows
and columns), and the same density (same ratio of ones and zeros). The hypothesis is that size, forma, y
density are not sufficient to determine fitness: Estructura (the specific pattern of zeros and ones) is crucial to
determining fitness. Eso es, the hypothesis is that fusion with shuffling (fusion_test_flag = 1) will reduce
aptitud física, compared to fusion without shuffling (fusion_test_flag = 0), because fitness requires specific struc-
turas. Summary statistics are not sufficient to determine fitness.
4 Experiments with the Model
En esta sección, we present four sets of experiments with Model-S.
4.1 Measuring the Contributions of the Layers
In the first set of experiments, we evaluate the contributions of the four layers of Model-S. We run
Model-S twelve times with each of the four layers, yielding a total of 48 carreras. We compare the layers
with respect to the fitness of the seeds, their area, their density, and their diversity. Mesa 1 shows the
parameter settings for Model-S in these experiments.
Mesa 1. This table lists the parameters used in Model-S and their values for the experiments presented in Section 4.1.
Parameter name
experiment_type_num
pop_size
num_trials
num_generations
min_s_xspan
min_s_yspan
s_xspan
s_yspan
max_area_first
max_area_last
seed_density
width_factor
height_factor
time_factor
tournament_size
elite_size
mutation_rate
prob_flip
prob_grow
prob_shrink
min_similarity
max_similarity
prob_fission
prob_fusion
symbiosis_flag
fusion_test_flag
Parameter values
1, 2, 3, 4
200
2
100
5
5
5
5
120
170
0.375
6.0
3.0
6.0
2
50
0.01
0.6
0.2
0.2
0.8
0.99
0.01
0.005
0
0
Used in Layers
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
1, 2, 3, 4
2, 3, 4
2, 3, 4
2, 3, 4
3, 4
3, 4
4
4
4
4
Notas. One generation is defined as the birth of pop_size children; therefore the number of children born in one run is
pop_size × num_generations = 20,000. The number of runs is not a parameter in the model; each run begins by starting a new
instance of Golly.
Artificial Life Volume 26, Número 3
349
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
All the comparisons that we make here are based on samples of the populations taken during
runs of Model-S. One run of Model-S generates 20,000 niños (num_generations × pop_size). Cada
generation is defined as the birth of 200 niños (pop_size). For each generation from 0 (the initial
random population) a 100 (the final population), we store the top 50 (elite_size) fittest individuals
(where fitness is relative) in a file for later analysis.
As we discussed in Section 3, fitness in Model-S is relative to the population. The fitness of an
individual is the fraction of Immigration Games that it wins in competitions against the other indi-
viduals in the population. Por lo tanto, it does not make sense to compare the fitness value of a seed in
one population with the fitness value of a seed in another population. The fitness used in Model-S is
relative and internal. This is a consequence of conditions 2 y 3 en la sección 3.1.
To compare fitness across different layers and different populations, we need to define a fitness
measure that is absolute and external to Model-S. Given a seed from any population and any layer, nosotros
calculate its absolute fitness by competitions against randomly generated seeds with the same matrix
tamaño (the same number of rows and columns) and the same matrix density (the same fraction of ones
in the matrix). Cifra 6 gives the absolute, external fitness curves for each of the four layers.
Because absolute fitness only compares seeds of the same size and density, any statistically sig-
nificant difference in absolute fitness values for two seeds must be due to the structures of the seeds
(the pattern of zeros and ones) and to how their structures determine their development from
genome to phenome over the course of the Immigration Game. Comparing seeds that are matched
by size and density is analogous to comparing wrestlers that are matched by height and weight: Él
allows us to distinguish brute force from skill. Mesa 2 shows the statistical significance of the
differences of the fitness curves in Figure 6. All of the differences are significant, except for the
difference between Layers 2 y 3 (variable asexual reproduction and sexual reproduction).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 6. Each curve in this figure (eso es, each layer ) is the average of 12 separate runs of Model-S. The fitness of a seed
is the fraction of Immigration Game contests that it wins when competing against randomly generated seeds with the
same size (the same width and height) and the same density (the same number of live cells). This is an external measure
of fitness that does not correspond to the internal measure used in the selection process in the four layers. El interno
measure of fitness would show no progress, because it compares each individual with the population, and the population
as a whole is progressing (on average) as fast as the individuals in the population are progressing.
350
Artificial Life Volume 26, Número 3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Mesa 2. This table tests the statistical significance of the differences in the fitness curves in Figure 6.
Layers to compare
Layer 1 versus Layer 2
Layer 1 versus Layer 3
Layer 1 versus Layer 4
Layer 2 versus Layer 3
Layer 2 versus Layer 4
Layer 3 versus Layer 4
p-value
5.03E−06
4.10E−05
2.81E−10
2.79E−01
3.44E−06
2.33E−07
p-value < 0.05
Yes
Yes
Yes
No
Yes
Yes
Notes. Each layer (each curve) is summarized by the average fitness over a run, yielding a sample of
twelve values, one value for each of the twelve runs of a layer. We then compare the fitness curves
for all possible pairs of layers, using a two-tailed Welch t-test for samples with unequal variance
(heteroscedastic variance). All the pairs of curves in Figure 6 are significantly different, except for
Layers 2 and 3 (the variable asexual layer and the sexual layer ).
Figure 7 plots the growth in area for each of the four layers. Comparing Figures 6 and 7, we see
the same general trends in both cases: Layer 1 has the lowest fitness and area, Layer 4 has the highest
fitness and area, and Layers 2 and 3 are roughly like each other. Since absolute fitness is measured by
competitions between seeds that have the same number of rows and columns, increasing fitness
cannot be a direct consequence of increasing area. Increasing fitness must be an indirect conse-
quence of the greater structural complexity that is permitted by increasing area.
Figure 8 shows the density of the seeds for the four layers. Density starts off at 0.375 (as specified
by seed_density in Table 1) and then decreases to range from 0.20 to 0.25. The rate of decrease in
density over time is slower for the more fit layers (Layers 2 and 4).
Figure 9 indicates the amount of diversity in the population for the four layers. We measure the
diversity by the standard deviation of the relative fitness in the elite population sample. A low standard
Figure 7. Each curve in this figure is the average of 12 separate runs of Model-S. Comparing this figure with Figure 6
suggests that area and fitness are positively correlated, even though the fitness in Figure 6 is based on size-matched
competitions. Greater area indirectly helps fitness by allowing more information to be encoded, which permits more
complex structures and actions.
Artificial Life Volume 26, Number 3
351
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Figure 8. In all four layers, density decreases over time. Perhaps lower density allows information to travel
distances, enabling greater complexity.
longer
deviation indicates that the elite sample has little variety; all the seeds are doing approximately the same
thing. A high standard deviation indicates that the elite sample embodies a variety of different strategies.
Layer 4 appears to have a more diverse population than the other three layers.
It might be argued that the standard deviation of the relative fitness in the population does not fully
capture diversity, because two organisms might have the same relative fitness, yet they might employ
quite different strategies. We chose the standard deviation of the relative fitness as the measure of
Figure 9. The diversity of the elite seed population is measured here by the standard deviation of the internal, relative
measure of fitness. The standard deviation is then averaged over the 12 separate runs of Model-S for each layer. Layers 1,
2, and 3 appear to have approximately the same diversities, but Layer 4 appears to be more diverse.
352
Artificial Life Volume 26, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
diversity because, for evolution by natural selection to have traction, we need diversity in the relative
fitness scores. If all the organisms have the same relative fitness, the population merely undergoes random
drift. Diversity in relative fitness is exactly the kind of diversity that is required to avoid random drift.
Two organisms with the same relative fitness may employ different strategies, but it is not clear
how to compare strategies directly. On the other hand, two organisms with quite different degrees
of relative fitness are almost certainly employing different strategies. Thus, the standard deviation of
the relative fitness in the population is a reasonable surrogate for directly comparing strategies.
It seems that there are strong similarities among the graphs for fitness (Figure 6), area (Figure 7),
density (Figure 8), and diversity (Figure 9), but it is possible that these similarities are statistical artifacts.
Therefore, we look at the correlations between all pairs of these four variables and test their statistical
significance. The results are given in Table 3. All the pairs have a significant positive correlation. Greater
fitness is correlated with greater area (0.843), greater density (0.405), and greater diversity (0.566).
The high correlation between fitness and area in Table 3 (0.843) suggests that increasing area causes
increasing fitness; but that is not the case. Increasing area is a necessary condition for increasing fitness, but
not a sufficient condition. This claim is supported by a two-part argument: (1) A theoretical argument
shows that increasing area is necessary for increasing fitness. (2) Empirical evidence shows that increasing
area is not sufficient for increasing fitness.
First, increasing area is necessary for increasing fitness: Consider a 5 × 5 seed pattern with an area of
25, which can represent 225 different patterns. When evolution has explored the entire set of 225
patterns and found the fittest patterns, fitness can no longer increase. A finite seed area implies an
upper bound to fitness. If we wish to simulate open-ended evolution (unbounded fitness increase),
then we must allow seed area to increase over time, without an upper bound [27, 28]. In Figure 6, we see
that Layer 1, which has a fixed area of 25, soon reaches a generation where fitness no longer increases
(around generation 30). However, Layer 2, which has a variable area, continues to slowly improve in
fitness throughout the run.
Second, increasing area is not sufficient for increasing fitness: In biology, specific kinds of complex
structures increase fitness, by functioning in ways that enhance fitness. A structure with many
components is not necessarily complex; for example, the many components might be arranged in a
simple periodic pattern. A structure with great complexity is not necessarily fit; the complexity could be
random or irrelevant for increasing fitness. These are two ways that increasing area can fail to deliver
increasing fitness: structures that are too simple or structures that are complex in ways that are irrel-
evant for fitness.
In Figure 6, the external, absolute fitness of an evolved seed is measured by the estimated prob-
ability that the evolved seed will win competitions against random seeds. Each evolved seed is
matched against fifty random seeds with the same area (number of cells), shape (number of rows
and columns), and density (percentage of ones in the seed matrix) as the given evolved seed. The
only thing that distinguishes an evolved seed from its random opponents is the structure of their
Table 3. This table looks at the correlations between fitness, area, density, and diversity (as displayed in Figures 6 to 9).
Feature 1
Feature 2
Correlation
area
area
area
density
density
diversity
density
diversity
fitness
diversity
fitness
fitness
0.362
0.816
0.843
0.418
0.405
0.566
p-value
1.14E−02
1.64E−12
5.89E−14
3.11E−03
4.28E−03
2.72E−05
p-value < 0.05
Yes
Yes
Yes
Yes
Yes
Yes
Notes. We evaluate the statistical significance of the correlations using a two-tailed Student t-test for Pearson correlations.
All the correlations are statistically significant. Each correlation is based on comparing two samples of 48 values each
(whereas Table 2 compares two samples of 12 values each).
Artificial Life Volume 26, Number 3
353
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
matrices (the specific pattern of ones and zeros). The external fitness measure used in Figure 6 can
be viewed as a measure of the degree of nonrandom structure in a seed; that is, increasing fitness in
Figure 6 corresponds to decreasing randomness. This implies that area alone is not sufficient for in-
creasing fitness. In addition to increasing area, a specific kind of increasingly nonrandom structure is
required. Specifically, winning requires structure that is good at playing the Immigration Game.
Consider the fitness curve for Layer 4 in Figure 6. By the final generation, the average fitness of
the elite seeds in Layer 4 is 93.6%. Because the average fitness is measured by competition against
randomly generated seeds, it follows that the probability that a random seed will win against a size-
matched elite seed (the same area, shape, and density) in the final generation is 6.4% (that is, 100%
minus 93.6%). If size alone were sufficient to win a competition, then the random sample of size-
matched seeds would be equally as fit as the evolved seeds (the average fitness would be 50%). This
is clear evidence that increasing area is not sufficient for increasing fitness. Increasing area must be
combined with a specific kind of structure, which could be described as fitness-enhancing structure. In-
creasing area is a necessary condition for increasing fitness, but not a sufficient condition. Area and
fitness are highly correlated, but increasing area does not cause increasing fitness.
4.2 Size, Structure, and Symbiosis
In this subsection, we consider two questions: (1) Fusion in Layer 4 combines two evolved seeds. Would
fusion work equally well if it combined one evolved seed and one random seed? Is a second seed merely
increasing the area of the whole, or is it contributing useful structure to the whole? (2) Fusion in Layer 4
is a model of symbiosis as any association. What happens when we model symbiosis as persistent mutualism? That
is, what if we permit fusion only when the fused whole is fitter than either part? In Section 3.5, we
introduced the parameters fusion_test_flag and symbiosis_flag. We can answer the first question by setting
fusion_test_flag = 1, and we can answer the second question by setting symbiosis_flag = 1.
The advantage of the fusion operator could be due to mutually beneficial interaction between the
two entities that are fused together, or it could be due to the increased area of the fused entity,
compared with the area of other entities in the population. Here we modify the fusion operator
by randomly shuffling all the cells in one of the two selected entities before we fuse them together
(fusion_test_flag = 1). For each cell in a matrix, we randomly select another cell in the matrix and then
we swap the values in the two cells. Random shuffling changes the structure of a seed (the location
of living and dead cells in the seed matrix) but preserves the shape (the number of rows and col-
umns in the seed matrix) and the density (the number of living cells in the seed matrix divided by the
total number of cells). We call this modified fusion operator Layer 4 Shuffled.
When two seeds are fused, there are three possible results: (1) Both seeds benefit: the fitness of the
whole is greater than the fitness of the parts. (2) Only one seed benefits: one of the parts is less fit than
the whole but the other part is more fit than the whole. (3) No seeds benefit: both parts are fitter than the
whole. Symbiosis as any association includes all three of these possibilities. Symbiosis as persistent mutualism
includes only the first case, where the fused seed is fitter than either of its parts. Here we modify the
fusion operator by requiring mutual benefit (symbiosis_flag = 1). We measure the fitness of each part and
the fitness of the whole, and we only allow the new fused seed to enter the population when the fused
seed is fitter than its parts. We call this modified fusion operator Layer 4 Mutualism.
Note that the three different fusion operators (Layer 4, Layer 4 Shuffled, Layer 4 Mutualism) all
use the same method to select two seeds from the population as input to the fusion operators. The
only differences in the three operators are the ways that the two selected seeds are fused (or not
fused, as the case may be).
Figure 10 compares the fitness curves of Layer 4, Layer 4 Shuffled, and Layer 4 Mutualism. We can
see the fitness of Layer 4 Shuffled initially falls behind the fitness of Layer 4 and Layer 4 Mutualism,
but Layer 4 Shuffled eventually catches up with the other layers. There is no significant difference
between the fitness curves for Layer 4 and Layer 4 Mutualism.
The difference between Layer 4 and Layer 4 Mutualism is that Layer 4 Mutualism discards fused
seeds when the fused whole is less fit than either part, whereas Layer 4 permits fusion regardless of
354
Artificial Life Volume 26, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Figure 10. (1) Would fusion work equally well if it combined one evolved seed and one random seed? Shuffling one of the
two seeds before fusing them (Layer 4 Shuffled) reduces the fitness of the fused seeds, compared to fusing seeds without
shuffling (Layer 4 and Layer 4 Mutualism). (2) What happens when we model symbiosis as persistent mutualism? There is
no significant difference between symbiosis as any association (Layer 4) and symbiosis as persistent mutualism (Layer 4
Mutualism). This indicates that the fitness increase over the generations comes from the cases where symbiosis is
mutually beneficial.
whether the parts mutually benefit from fusion. The similarity of the two fitness curves tells us that
most of the fitness increase in Layer 4 and Layer 4 Mutualism is due to mutualism. The cases in
Layer 4 where the parts do not mutually benefit from fusion have neither a beneficial nor a detri-
mental effect on the fitness curve. Removing the cases of symbiosis that lack mutualism is neutral
with respect to its impact on fitness. These cases neither help nor harm the population.
Table 4 compares the fusion events in Layer 4, Layer 4 Shuffled, and Layer 4 Mutualism. Layer 4
Shuffled has fewer cases of mutualism than Layer 4, due to the structural damage caused by shuffling.
Layer 4 Shuffled has more fusion events than Layer 4, due to the linear bound on area, set by
max_area_first and max_area_last. The bounded area slows down the rapid growth of Layer 4 earlier
than it slows down the gradual growth of Layer 4 Shuffled. Although the three layers have similar
numbers of cases of mutualism, the cases occur in earlier generations with Layer 4 and Layer 4
Table 4. This table compares fusion events during runs of Layer 4, Layer 4 Shuffled, and Layer 4 Mutualism.
Number of fusion events
Number of mutualisms
Percent of mutualisms
Layer 4
Layer 4 Shuffled
Layer 4 Mutualism
44.3
61.5
5.4
6.6
4.9
5.4
15%
8%
100%
Notes. The numbers are averages over the 12 runs of each layer. The column “Number of fusion events” includes events with
and without mutualism. The other columns focus exclusively on fusion events with mutualism. By design, all fusion events in
Layer 4 Mutualism are cases of mutualism. The expected number of fusion events is prob_fusion × num_generations ×
pop_size = 100, but fusion is suppressed by the limits max_area_first and max_area_last in Layer 4 and Layer 4 Shuffled,
and fusion is further suppressed by the requirement of mutual benefit in Layer 4 Mutualism.
Artificial Life Volume 26, Number 3
355
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Table 5. This table compares Layer 4 with Layer 4 Shuffled in generations 30 and 100.
Generation
30
100
All
Layer 4
0.903
0.936
0.892
Layer 4 Shuffled
0.837
0.927
0.871
p-value
0.004
0.601
0.057
p-value < 0.05
Yes
No
No
Notes. The fitness of the two layers for all generations is shown in Figure 10. Layer 4 is significantly more fit than Layer 4
Shuffled in generation 30, but the difference between the two layers is no longer significant when they reach generation 100.
Averaged over all the generations, the difference is not significant. The p-values are calculated using a two-tailed Welch t-test
for samples with unequal variance (heteroscedastic variance).
Mutualism, which is why the two layers are ahead of Layer 4 Shuffled at first, but Layer 4 Shuffled
eventually catches up, due to the linear bound on area.
Table 5 shows that Layer 4 is significantly more fit than Layer 4 Shuffled in generation 30, but the
difference is no longer significant in generation 100. Averaging over all generations, the difference
between Layer 4 and Layer 4 Shuffled is not significant. Layer 4 Shuffled has a few cases where
fusion is mutually beneficial, despite the harm done by the shuffling operation. These few cases
are sufficient to allow the fitness curve for Layer 4 Shuffled to eventually catch up with Layer 4.
Table 6 tells us that, in most cases, fusion is harmful. The cases where both seeds benefit from
fusion are relatively rare. This is also true of mutation: Most mutations are harmful. We expect
harmful mutations and harmful fusions in evolution. They will eventually be eliminated by natural
selection. If there are a few cases where mutation and fusion result in increased fitness, the
population will continue to increase in fitness, despite the relative rarity of beneficial mutations
and mutualist symbiosis.
In Table 6, the cases when both parts benefit from fusion are instances of symbiosis as persistent
mutualism, whereas the other cases are instances of symbiosis as any association. Shuffling disrupts fusion
and reduces the incidence of persistent mutualism. Persistent mutualism is relatively rare (15% of the
fusions when both parts are fit; see Table 4), but shuffling makes mutualism rarer (8% of the fusions
when one part is shuffled). In Figure 10, the rise of the fitness curve for Layer 4 in generation 30,
compared to Layer 4 Shuffled in generation 30, may be explained by the greater incidence of symbiosis
as persistent mutualism. By generation 100, although Layer 4 Shuffled has fewer cases of persistent
mutualism, there are enough accumulated cases to allow Layer 4 Shuffled to catch up with Layer 4.
The results in Table 6 add further support to the argument in Section 4.1, that unbounded area
increase is not sufficient for unbounded fitness increase. Layer 4 and Layer 4 Shuffled yield the same
Table 6. In this table, we compare Layer 4 with Layer 4 shuffled.
No parts benefit
One part benefits
Both parts benefit
Layer 4
Layer 4 Shuffled
p-value of difference
p-value < 0.05
80.5%
89.3%
0.041
Yes
4.5%
2.7%
0.214
No
15.0%
8.0%
0.033
Yes
Notes. Shuffling decreases the probability that the component parts will both benefit from fusion; that is, the probability of
symbiosis as persistent mutualism. The whole is fitter than both parts 15% of the time for Layer 4, but only 8% of the time for
Layer 4 Shuffled. This shows that increased fitness does not come merely from the increase in area when fusing two seeds; a
significant part of the increased fitness comes from the structural properties of the two seeds that are fused together. Layer 4
Shuffled is less likely to have the proper structure than Layer 4. The p-values in the table are calculated using a two-tailed
Welch t-test for samples with unequal variance (heteroscedastic variance).
356
Artificial Life Volume 26, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
increase in area when fusion occurs. Shuffling is designed to have no impact on area (number of cells),
shape (number of rows and columns), and density (percentage of ones in the seed matrix). Therefore,
the only explanation for the difference between Layer 4 and Layer 4 Shuffled in Table 6 is structure.
The main lesson of this section is that mutualism is what drives ongoing fitness increase. Comparing
Layer 4 with Layer 4 Mutualism shows that the fusions that lack mutual benefit have no effect on fitness.
Layer 4 Shuffled is less likely to produce mutualism than Layer 4 and Layer 4 Mutualism, but it even-
tually produces enough mutualism to catch up with the other layers.
Layer 4 Shuffled produces more large seeds (more fusion events) than Layer 4 and Layer 4
Mutualism (Table 4), yet Layer 4 Shuffled struggles to keep up with the fitness increases of Layer 4
and Layer 4 Mutualism (Figure 10). This shows that merely producing large seeds is not enough to
promote increasing fitness. Layer 4 Mutualism produces very few large seeds (Table 4), yet surpasses
the fitness of Layer 4 Shuffled and matches the fitness of Layer 4 (Figure 10). This is further evidence
that increasing area is not sufficient for increasing fitness. Increasing area allows fitness to increase
by providing room for complex structures that enhance fitness. It is these structures that result in
ongoing fitness increase.
4.3 Comparing Evolution and Design
Table 7 summarizes the properties of the evolved seeds produced in the last generation from each of
the six layers. It might be argued that the external fitness measure in Section 4.1, based on compe-
titions against randomly generated seeds, is not sufficiently challenging. How would evolved seeds
fare against human-engineered seeds, instead of random seeds? We address that question here.
The Golly software [26] comes with a substantial collection of human-engineered Game of Life seed
patterns that can be pitted against the evolved seeds. To be fair, we focus on the human-engineered
seeds that are comparable to the evolved seeds in area, as we know from Table 3 that area and fitness are
highly correlated. Therefore, we set a limit of 10,000 on the area of human-engineered seeds. Table 8
gives the results of this contest.
The column in Table 7 that is labeled “Fitness” gives the fitness of the six different layers as mea-
sured by competition with random seeds. The bottom row in Table 8 that is labeled “Average” gives the
fitness of the six different layers as measured by competition with human-engineered seeds. Comparing
these two different external measures of fitness, we can see that the human-engineered seeds are more
challenging than the random seeds, as we might expect. The fitness scores for the competitions with
human-engineered seeds range from 47% to 73% in Table 8, whereas the fitness scores for the com-
petitions with random seeds range from 74.3% to 94.4% in Table 7. However, the two different fitness
measures give the same qualitative ranking of the six layers. For both measures, (a) Layer 1 has the
lowest fitness, (b) Layer 2 and Layer 3 are roughly similar in fitness, and they are fitter than Layer 1,
Table 7. This table summarizes the final generations of the layers (generation 100).
Layer
Layer 1
Layer 2
Layer 3
Layer 4
Layer 4 Shuffled
Layer 4 Mutualism
Average
Fitness
0.743
0.846
0.810
0.936
0.927
0.944
0.868
Area
25.0
32.8
29.5
94.1
90.5
91.3
60.5
Density
Diversity
0.217
0.229
0.212
0.245
0.245
0.233
0.230
0.0105
0.0100
0.0111
0.0139
0.0140
0.0142
0.0123
Notes. The average area of the evolved seeds (60.5 in this table) is generally smaller than the area of the human-designed seeds
(2,454 in Table 8). The evolved seeds are competing against human-designed seeds that are mostly much larger than the
evolved ones.
Artificial Life Volume 26, Number 3
357
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Table 8. This table gives the scores of each of the layers when competing against human-designed patterns of comparable
area.
Golly pattern file name
Area
Layer 1
Layer 2
Layer 3
Layer 4
Layer 4 Shuffled
Layer 4 Mutualism
Percentage of games won for each layer
100
95
100
95
100
93
100
100
100
agar-p3.rle
herringbone-agar-p14.rle
pulsars-in-tube.rle
spacefiller.rle
vacuum-cleaner.rle
acorn.lif
ark1.rle
ark2.rle
blom.rle
iwona.rle
justyna.rle
lidka-predecessor.rle
natural-LWSS.rle
rabbits-relation-17423.rle
rabbits-relation-17465.rle
rabbits.lif
temp-pulsars-big-s.rle
die658.rle
line-puffer-superstable.rle
line-puffer-unstable.rle
pi-fuse-puffer.rle
puffer-2c5.rle
puffer-train.rle
heisenblinker-30.rle
heisenburp-46-natural.rle
eaters-misc.rle
random.rle
ss-eaters.rle
stripey.rle
Average
3,456
2,304
136
1,274
8,730
21
928
2,332
60
420
374
135
40
36
24
21
64
400
4,992
1,683
1,827
8,400
90
5,032
2,346
4,851
9,604
7,298
4,290
2,454
0
0
0
58
1
0
61
11
17
32
65
66
75
69
77
97
0
2
18
1
34
17
71
100
100
100
100
47
4
0
13
57
6
2
68
27
28
26
73
72
80
71
83
95
0
3
36
5
43
27
77
98
8
0
13
59
8
2
63
28
32
38
69
73
76
71
79
95
0
2
42
8
46
31
77
99
96
81
3
75
81
42
21
82
67
87
58
79
86
81
85
84
98
3
12
90
63
75
80
90
96
100
100
100
51
100
99
100
52
100
97
100
73
97
82
3
79
72
41
26
81
69
86
56
78
85
85
82
82
98
2
19
89
57
70
75
92
96
100
97
100
72
97
73
2
56
79
41
22
84
64
85
56
80
84
86
85
83
98
3
10
83
44
68
72
88
98
100
98
100
70
Notes. We test the layers against all human-designed patterns that have an area of 10,000 or less, a total of 29 patterns. Each
human-designed Golly pattern competes 20 times against the fittest seed in the final generation of each run of the given layer.
For example, there are 12 runs for Layer 1; thus 240 (12 × 20) Immigration Games are played with each human-designed Golly
pattern. In the table, we report the percentage of competitions in which the evolved seed was the winner.
and (c) Layer 4, Layer 4 Shuffled, and Layer 4 Mutualism are roughly similar in fitness and fitter than
Layer 2 and Layer 3.
The human-engineered seeds are at a disadvantage in this contest because they were not designed to
play the Immigration Game. An exception is the class of human-engineered seeds called breeders [19].
These are seed patterns that have been engineered to fill space as quickly and densely as possible. There
is only one breeder in Table 8 (spacefiller.rle), and we can see that it won against all six evolved layers.
Golly has other breeders, but they all have areas greater than 10,000. If we raise the area limit from
10,000 to 50,000, there are five breeders below the area limit. These five breeders win in competitions
with the evolved seeds. Human engineering triumphs over Model-S evolution, but this may change if
Model-S is given the computational resources to achieve higher seed areas.
358
Artificial Life Volume 26, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
4.4 An Unbounded External Fitness Measure
The external, absolute fitness measure introduced in Section 4.1 (see Figure 6) is based on evolved
seeds competing against randomly generated seeds with the same matrix size (the same number of
rows and columns) and the same matrix density (the same fraction of ones in the matrix). Section 4.3
supports this fitness measure by showing that it agrees with the ranking produced from competitions
against human-designed seeds (see Table 8). The fitness measure of Section 4.1 works well for the
experiments presented in the preceding sections, but it has limitations.
One requirement we might impose on an external, absolute fitness measure it that it should produce
a curve that rises when the fitness of the population is improving, stays flat when the population is
neither improving nor worsening, and falls when the population is worsening. Let us call this require-
ment directional consistency. Fitness as measured by competition against random seeds (as in Figures 6 and
10) satisfies this requirement.
Another requirement we might impose on an external, absolute fitness measure is that the pace of
fitness change should correspond to the slope of the curve. Let us call this requirement slope consistency.
The absolute fitness measure in Section 4.1 (evolved seeds competing against randomly generated seeds
with the same matrix size and density) ranges between zero and one, which prevents it from satisfying
slope consistency. The upper and lower bounds on fitness do not allow the slope to remain constant for
long. As the curve gets closer to one, the slope must decrease, even if the pace of fitness change is
constant.
In this section, we present a fitness measure that satisfies both requirements, directional consistency
and slope consistency. The new fitness measure is unbounded; it ranges between negative infinity and
positive infinity. We then compare the new measure with the fitness measure in Section 4.1. The results
show that the two measures are highly correlated.
Let sn be the seed in generation n with the largest internal, relative fitness. Let pin be the probability
that sn is more fit than si, the fittest seed in generation i, where i < n and pin 2 [0, 1]. We estimate pin by
having si and sn compete against each other in g Immigration Games. If sn wins w games against si, then
we estimate the probability pin by w/g. If si and sn are equally fit, we expect pin = 0.5. To satisfy the first
requirement for an external, absolute fitness measure (directional consistency), the curve should rise
when pin > 0.5, fall when pin < 0.5, and stay flat when pin = 0.5. To achieve this behavior, we use the
formula 2pin − 1, which ranges from −1 to +1 as pin ranges from 0 to 1. The external, absolute fitness fn
of sn is then defined as follows:
fn ¼
Xi¼n−1
i¼0
ð
2pin − 1
Þ
The function fn ranges from –n to +n. The function has directional consistency: If pin reaches a gen-
eration n where the probability of winning is random ( pin = 0.5), then the curve for fn will start to flatten
out. If the probability is worse than random ( pin < 0.5), the curve will head downwards, perhaps even-
tually going below zero. If the probability is better than random ( pin > 0.5), the curve will head upwards.
The function also has slope consistency: The slope of the curve corresponds to the pace of fitness
cambiar. De este modo, this function satisfies the two requirements for an external, absolute fitness measure.
We reduce noise in our estimate of the probability pin by taking the top ten fittest seeds in generation i
and the top ten fittest seeds in generation n and making each pair of seeds compete twice, de modo que
the estimate for pin is based on the average outcome of 200 competitions (10 × 10 × 2 = 200). El
noise is further reduced by averaging over twelve separate runs of the model.
Cifra 11 shows the fitness of the six layers, as given by the new fitness measure. The new fitness
measure makes the steady fitness increase of Layer 4 more readily visible than the old fitness mea-
sure (compare Figure 11 with Figures 6 y 10).
Mesa 9 shows that the two external fitness measures, comparison with random seeds (Figures 6
y 10) and comparison with past winners (Cifra 11), are highly correlated (0.767) when we con-
sider the fitness score fn averaged over all generations. The correlation is statistically significant.
Artificial Life Volume 26, Número 3
359
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
PAG. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Cifra 11. This figure compares the six different configurations of Model-S using an unbounded external fitness measure.
Whereas the fitness measure in Figures 6 y 10 (comparing evolved seeds with random seeds of the same size and
density) is limited to ranging from 0 a 1, the fitness measure here ranges from negative infinity to positive infinity.
Comparing Figures 6 y 11, we see the same ranking of the different layers (from top to bottom: Layer 4, Layer 2, Layer 3,
Layer 1). Cifra 11 is more suitable than Figure 6 for showing the steady increase in fitness of Layer 4. En figura 11, as in
Cifra 10, Layer 4 Shuffled falls behind Layer 4 and Layer 4 Mutualism, but it eventually catches up.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
yo
/
_
a
_
0
0
3
2
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Mesa 9. This table compares the two forms of external fitness, elite seed fitness compared with random seeds of the
same size and density (see Figures 6 y 10 in Sections 4.1 y 4.2) versus current winner compared with past winners
(ver figura 11).
Fitness scores averaged over all generations
External fitness based on
comparison with past winners
External fitness based on
comparison with random seeds
Layer 1
Layer 2
Layer 3
Layer 4
Layer 4 Shuffled
Layer 4 Mutualism
Correlación
p-value of correlation
p-value < 0.05
2.3
3.2
2.4
11.7
10.8
12.0
0.739
0.814
0.799
0.892
0.871
0.897
0.767
4.15E−15
Yes
Notes. The table shows the correlation between the two measures of external fitness. The correlation is calculated from 72
random seed fitness scores and 72 past winner fitness scores (6 layers with 12 fitness values for each layer yields 72 fitness
scores). Each fitness score in this table is an average over all the generations for a given layer (an average of 12 fitness values, one
for each run, each of which is an average over the 100 generations). We evaluate the statistical significance of the correlations
using a two-tailed Student t-test for Pearson correlations. The correlation is high (0.767), and it is statistically significant.
360
Artificial Life Volume 26, Number 3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Table 10. Like Table 9, this table compares the two forms of external fitness: elite seed fitness compared with random
seeds of the same size and density, versus current winner compared with past winners.
Fitness scores in generation 100
External fitness based on
comparison with past winners
External fitness based on
comparison with random seeds
Layer 1
Layer 2
Layer 3
Layer 4
Layer 4 Shuffled
Layer 4 Mutualism
Correlation
p-value of correlation
p-value < 0.05
2.7
7.7
4.4
21.6
24.5
23.6
0.743
0.846
0.810
0.936
0.927
0.944
0.765
5.37E−15
Yes
Notes. Each fitness score in this table is an average of the final generation for a given layer (an average of 12 fitness values). The
difference between the two tables is that here we focus on the final generation, whereas the preceding table examined
the average fitness over all the generations. The similar correlation values in Table 9 (0.767) and Table 10 (0.765) indicate that
the correlations between the two forms of external fitness (comparison with past winners and comparison with random seeds)
are robust.
Table 10 shows that the two external fitness measures are also highly correlated (0.765) when we focus
on the final generation (n = 100), and the correlation is again statistically significant. All three fitness
measures (in Sections 4.1, 4.3, and 4.4) show the same general rankings of the six different configurations
of Model-S: (a) Layer 1 has the lowest fitness, (b) Layers 2 and 3 are similar, with a slight advantage to
Layer 2, and (c) Layer 4, Layer 4 Shuffled, and Layer 4 Mutualism have the highest fitness and are similar.
Open-ended evolution is defined as evolution that is [25, p. 409] “capable of producing a continual
stream of novel organisms rather than settling on some quasi-stable state beyond which nothing
fundamentally new occurs.” In Figure 11, it appears that evolution is open-ended over the course of
100 generations for Layer 4, Layer 4 Shuffled, and Layer 4 Mutualism. We hypothesize that the trends in
Figure 11 will continue indefinitely.
5 Discussion of Results
Section 4.1 shows that Layer 4, symbiosis by genetic fusion, significantly increases fitness when com-
bined with the other layers. The increase in fitness due to fusion occurs despite several obstacles: (1)
Fusion is rare, taking place in only 1 out of 200 births (prob_fusion = 0.005; see Table 1). (2) Fission is
twice as likely as fusion (prob_fission = 0.01), and fission is pushing the population towards decreasing
size, in opposition to fusion. (3) Layer 4 passes control to Layer 3 most of the time (98.5% of the time;
see Figure 5), but Layer 3 is less fit than Layer 2 (see Figure 6). Layer 4 could perform better if it passed
control directly to Layer 2 instead of Layer 3. (4) The parameters max_area_first and max_area_last impose
strong constraints on fusion (see Section 3.5). When the population reaches the limits set by these
parameters, fusion is no longer permitted (because the simulation becomes exceedingly slow).
Section 4.2 considers whether the fitness increase from fusion is due to increase in the size of the
fused seeds or due to beneficial interaction of the fused seeds. Shuffling is introduced as a way to disrupt
the structure of a seed without affecting its size. The shape, area, and density of a seed are not affected by
shuffling. The results indicate that shuffling reduces the fitness of the fused seed (see Table 6), which
slows the increase in fitness in the earlier generations, but the population recovers in the later generations
(see Figure 10 and Table 5), due to the limits on size (max_area_first and max_area_last in Section 3.5).
Artificial Life Volume 26, Number 3
361
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
Section 4.2 also examines what happens when fusion requires mutual benefit, with Layer 4 Mutualism.
The results show that the few cases of mutual benefit account for all of the increasing fitness. Elim-
inating all of the fusion events that lack mutual benefit has no impact on fitness: There is no significant
difference between the fitness curves for Layer 4 (symbiosis as any association) and Layer 4 Mutualism
(symbiosis as persistent mutualism) in Figures 10 and 11.
Section 4.3 shows that comparison of evolved seeds with human-engineered seeds (Table 8)
yields approximately the same ranking of the layers of Model-S as comparison with random seeds
of the same size and density (Table 7). This agreement between fitness measured by comparison
with random seeds (Section 4.1) and fitness measured by comparison with human-engineered seeds
(Section 4.3) suggests that both fitness measures are performing as intended.
Section 4.4 introduces a third absolute, external measure of fitness, in addition to the measures
in Sections 4.1 and 4.3. This fitness measure is designed to have both directional consistency (it rises when
the fitness of the population is improving, stays flat when the population is neither improving nor
worsening, and falls when the population is worsening) and slope consistency (the pace of fitness change
corresponds to the slope of the curve). The measure is based on comparing the fittest seed in each
generation with the fittest seed in all previous generations (where the fittest seed is determined by
the internal, relative fitness of the seeds in the given population). Tables 9 and 10 show that this
new measure of fitness is highly correlated with external fitness measured by comparison with random
seeds. Furthermore, the steady increase in the fitness of Layer 4, Layer 4 Shuffled, and Layer 4 Mutualism
that we see with this third fitness measure (Figure 11) lends support to the hypothesis that symbiosis
supports open-ended evolution.
Three measures of external, absolute fitness all produce the same general ranking of the six layers:
(a) comparison of fit seeds with random seeds of the same size and density in Figures 6 and 10,
Sections 4.1 and 4.2, (b) comparison of fit seeds with human-designed seeds in Table 8, Section 4.3,
and (c) comparison of current fit seeds with past fit seeds in Figure 11, Section 4.4. The agreement
among these three different measures shows the results are not dependent on a specific approach to
measuring external, absolute fitness.
Three different arguments support the claim that increasing area is necessary but not sufficient for
increasing fitness, and thus increasing area does not cause increasing fitness: (1) The argument at the end of
Section 4.1, comparing evolved seeds with area-matched random seeds, shows that Layer 4 generates
seeds with highly nonrandom structures, and the difference in fitness between the evolved seeds and the
random seeds cannot be due to a difference in area, because the competing seeds have the same area. (2)
The argument in Section 4.2, comparing Layer 4 and Layer 4 Mutualism, shows there is no significant
difference in their fitness curves (Figure 10), although Layer 4 produces many more large seeds than
Layer 4 Mutualism (Table 4). The relative abundance of large seeds does not boost the fitness of Layer 4,
because most of the large seeds lack mutualism. (3) The argument in Section 4.2, comparing Layer 4
and Layer 4 Shuffled, shows that shuffling reduces fitness (Tables 5 and 6), although it does not change
area. The cause of the reduction in fitness must be structural, since the only difference in the seeds is
structural.
The necessary and sufficient condition for increasing fitness is the evolution of structures that are
increasingly better at playing the Immigration Game. For unbounded fitness increase, increasing area
is necessary to accommodate increasingly complex structures, but increasing area is not sufficient.
6 Future Work and Limitations
In biology, there are many examples of symbiosis without mutualism, such as parasitism. Model-S
does not currently deal with parasitism, although it could be expanded to do so. We should not infer
from the experiments described here that evolution favors mutualism over parasitism. The genetic
fusion operator in Model-S encourages the two fused genomes to get along with each other, because
selection occurs at the level of the whole. If a parasite kills its host, selection at the level of the
whole would penalize the parasite. Genetic fusion is not the appropriate operator for modeling
362
Artificial Life Volume 26, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
the parasite-host relationship. A parasite and its host have a close relationship, but their genes are
selected separately. Fusion is essentially a mechanism for forcing the parts to either cooperate or die.
We leave parasitism for future work.
In Section 4.2, we examined the interaction between the two parts of a fused seed. Fusion is a
joining of two seeds (two genomes), which are inert, so there is no interaction at the moment of
fusion. However, there is vigorous interaction when the Immigration Game runs (the growth of the
two phenomes). Symbiosis occurs during the game, either symbiosis as any association or symbiosis
as persistent mutualism, depending on how well the seeds work together. We can easily detect the
interaction by comparing the fitness of the separate parts with the fitness of the fused whole. Table 6
tells us that the interaction is mostly negative; only 15% of the time do both parts of the fused whole
benefit (mutualism). Mostly we see association without benefit. This is expected for the fusion op-
erator, just as it is for the mutation operator: Most mutations are harmful or neutral, and only a few
are beneficial, but it is those few beneficial cases that yield greater fitness and adaptation in the long-
term evolution of the population.
We can infer the interaction between the two parts of a fused seed from its effect on fitness, but
it would be interesting to observe the interaction directly. In future work, we could visually display
the interaction between the two fused seeds. For example, suppose that a red seed that has under-
gone fusion is competing with a blue seed that has undergone fusion. We could color the red seed
in two different shades of red, corresponding to its two fused parts, and we could color the blue seed
in two different shades of blue, corresponding to its two fused parts. We would then be able to
visualize how the parts interact over the course of a game. (We need to decide how to handle
the case where an empty cell has three live neighbors with three different colors. There are many
ways to deal with this.)
A limitation of Model-S is the amount of time required to run the Immigration Game as the seeds
evolve to become larger with fusion. Addressing this problem may be a straightforward task of tuning
the parameters, width_factor, height_factor, and time_factor (see Section 3.2), but we believe a more sophis-
ticated method is required for determining the best toroid size and the best time limit for a given pair of
competing seeds. One way to set the time limit would be to use a test for quiescence to determine the end
of a game. The idea is to end the game when the score appears to be nearly stable.
A puzzle from the results presented in Section 4 is the relatively poor performance of sexual
reproduction in Layer 3. Simon [21] lists eleven different kinds of genetic crossover. It may be that
one of the other forms of crossover will perform better than the simple single-point crossover used
in Model-S. It might be helpful to introduce a form of two-dimensional crossover that exchanges
subsquares between the parent seed matrices.
We use a form of restricted mating in Model-S, controlled by the parameters min_similarity and
max_similarity (see Section 3.4). We tried to tune these parameters to improve sexual reproduction,
without success. Sexual reproduction has long been a topic for debate among evolutionary biolo-
gists, with many different theories about its role in evolution. Ridley [20] argues that a major reason
for sexual reproduction is to provide resistance against parasites. One option would be to add sim-
ulated parasites to Model-S.
Table 1 shows that the parameter space for Model-S is large. Experiments with Model-S are slow,
which makes it difficult to explore the parameter space thoroughly. Although we have run many
experiments, we have only explored a tiny fraction of the parameter space. Much exploration remains
to be done.
For those who are interested in Lamarckian evolution, Model-S could be a suitable platform.
Lamarckian evolution is based on the inheritance of acquired characteristics. We can simulate a kind
of Lamarckian evolution as follows: (1) Put a seed into the Game of Life. This seed is the genotype. (2)
Let the game run for N steps. The resulting pattern is the phenotype. (3) Take the resulting pattern out
of the game and use it as a new seed. This new seed has acquired characteristics from its time in the
Game of Life, and these characteristics are heritable.
For those who are interested in tracking the heritage of individuals, it would be easy to modify
Model-S by storing a family tree in each seed object. The nodes in the tree could be pointers into a
Artificial Life Volume 26, Number 3
363
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
database of stored seeds. This would be useful for testing hypotheses about the properties of inheritance
in Model-S.
The discussion of related work (Section 2) mentioned the meta-model of Banzhaf et al. [2], which
can be applied to the Game of Life. An interesting project for future work would be to apply the
meta-model to the Immigration Game and Model-S.
7 Conclusion
Our model of symbiosis has four layers of genetic operators. The first three layers include asexual
and sexual reproduction, with standard genetic operators such as mutation and crossover. The
fourth layer introduces two new genetic operators, fusion and fission.
In the model, the fitness of an organism is determined by competition in the Immigration Game,
a variation on the Game of Life. A key insight is that the fusion operator is easy to implement in the
Game of Life: Organisms are fused by simply joining them side by side and treating them as a new
whole. In other types of artificial life simulations, fusion may not be as straightforward to implement.
Our main result is that symbiosis by fusion is a powerful genetic operator, when combined with
the standard genetic operators (mutation and crossover ). A small amount of fusion (one birth in
200) can have a substantial impact on the course of evolution (Section 4.1). The results with the new
unbounded external fitness measure (Section 4.4, Figure 11) suggest that fusion can sustain open-
ended evolution [25].
We hope that the release of Model-S as open-source software [30] will encourage other researchers
to explore the many open questions raised in this paper. It seems likely that there are many other genetic
operators, beyond mutation, selection, crossover, fission, and fusion, awaiting artificial life models.
Acknowledgments
Thanks to Tim Taylor and Martin Brooks for helpful discussion and advice. Thanks to Andrew Trevorrow,
Tom Rokicki, Tim Hutton, Dave Greene, Jason Summers, Maks Verver, Robert Munafo, Brenton
Bostick, and Chris Rowett for developing Golly, which made this research more productive and enjoyable.
Thanks to the Artificial Life reviewers for their very thoughtful and helpful comments.
References
1. Aron, J. (2010). First replicating creature spawned in life simulator. New Scientist, Issue 2765, June. https://
www.newscientist.com/article/mg20627653-800-first-replicating-creature-spawned-in-life-simulator/.
2. Banzhaf, W., Baumgaertner, B., Beslon, G., Doursat, R., et al. (2016). Defining and simulating open-ended
novelty: Requirements, guidelines, and challenges. Theory in Biosciences, 135, 131–161.
3. Beer, R. D. (2004). Autopoiesis and cognition in the Game of Life. Artificial Life, 10(3), 309–326.
4. Beer, R. D. (2014). The cognitive domain of a glider in the Game of Life. Artificial Life, 20(2), 183–206.
5. Beer, R. D. (2015). Characterizing autopoiesis in the Game of Life. Artificial Life, 21(1), 1–19.
6. Brandon, R. N. (1996). Concepts and methods in evolutionary biology. Cambridge, UK: Cambridge University Press.
7. Douglas, A. (2010). The symbiotic habit. Princeton, NJ: Princeton University Press.
8. Gardner, M. (1970). Mathematical games—the fantastic combinations of John Conwayʼs new solitaire game
“life.” Scientific American, 223(4), 120–123.
9. Godfrey-Smith, P. (2007). Conditions for evolution by natural selection. The Journal of Philosophy, 104,
489–516.
10. Ikegami, T. (1994). From genetic evolution to emergence of game strategies. Physica D: Nonlinear Phenomena,
75(1–3), 310–327.
11. Johnston, N. (2009). Longest-lived soup density in Conwayʼs game of life. http://www.njohnston.ca/2009/06/
longest-lived-soup-density-in-conways-game-of-life/.
364
Artificial Life Volume 26, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
P. D. Turney
Symbiosis Promotes Fitness Improvements in the Game of Life
12. Koestler, A. (1967). The ghost in the machine. London, UK: Hutchinson.
13. Margulis, L. (1970). Origin of eukaryotic cells. New Haven, CT: Yale University Press.
14. Margulis, L. (1981). Symbiosis in cell evolution. San Francisco: W. H. Freeman.
15. Maynard Smith, J., & Szathmáry, E. (1995). The major transitions in evolution. Oxford, UK: Oxford University
Press.
16. McShea, D. W., & Brandon, R. N. (2011). Biologyʼs first law: The tendency for diversity and complexity to increase in
evolutionary systems. Chicago: University of Chicago Press.
17. Moreno, M. A., & Ofria, C. (2019). Toward open-ended fraternal transitions in individuality. Artificial Life,
25, 117–133.
18. Nolan, P., & Lenski, G. E. (2010). Human societies: An introduction to macrosociology (11th ed.). Boulder, CO:
Paradigm.
19. Poundstone, W. (2013). The recursive universe: Cosmic complexity and the limits of scientific knowledge. Mineola, NY:
Dover.
20. Ridley, M. (1993). The Red Queen: Sex and the evolution of human nature. New York: Harper Perennial.
21. Simon, D. (2013). Evolutionary optimization algorithms. Hoboken, NJ: Wiley.
22. Simon, H. A. (1962). The architecture of complexity. Proceedings of the American Philosophical Society, 106,
467–482.
23. Sterelny, K. (2011). Evolvability reconsidered. In B. Calcott & K. Sterelny (Eds.), The major transitions in
evolution revisited (pp. 83–100). Cambridge, MA: MIT Press.
24. Taylor, T. (2015). Requirements for open-ended evolution in natural and artificial systems. In EvoEvo
the 13th European Conference on Artificial Life (ECAL 2015), University of York, UK.
Workshop at
http://arxiv.org/abs/1507.07403.
25. Taylor, T., Bedau, M., Channon, A., Ackley, D., et al. (2016). Open-ended evolution: Perspectives from the
OEE workshop in York. Artificial Life, 22, 408–423.
26. Trevorrow, A., Rokicki, T., Hutton, T., Greene, D., Summers, J., Verver, M., Munafo, R., Bostick, B., &
Rowett, C. (2019). Golly 3.2 for Windows. http://golly.sourceforge.net/.
27. Turney, P. D. (1999). Increasing evolvability considered as a large-scale trend in evolution. In W. Banzhaf,
J. Daida, A. E. Eiben, M. H. Garzon, V. Honavar, M. Jakiela, & R. E. Smith (Eds.), Proceedings of the 1999
Genetic and Evolutionary Computation Conference (GECCO-99) Workshop Program (pp. 43–46). San Francisco,
CA: Morgan Kaufman.
28. Turney, P. D. (2000). A simple model of unbounded evolutionary versatility as a largest-scale trend in
organismal evolution. Artificial Life, 6, 109–128.
29. Turney, P. D. (2018). Conditions for major transitions in biological and cultural evolution. In Proceedings of
the Third Workshop on Open-Ended Evolution (OEE3) at the 2018 Conference on Artificial Life. http://www.alife.
org/workshops/oee3/papers/turney-oee3-final.pdf.
30. Turney, P. D. (2020). Model-S source code. https://github.com/pdturney/modeling-symbiosis-revised.
31. Wainwright, R. T. (1971). Lifeline Volume 2. http://www.conwaylife.com/wiki/Lifeline_Volume_2.
32. Watson, R. A., & Pollack, J. B. (2003). A computational model of symbiotic composition in evolutionary
transitions. Biosystems, 69, 187–209.
33. Whitley, D., & Kauth, J. (1988). GENITOR: A different genetic algorithm. In J. H. Alexander (Ed.),
Proceedings of the Rocky Mountain Conference on Artificial Intelligence. Colorado Institute for Artificial Intelligence.
34. Whitley, D. (1989). The GENITOR algorithm and selective pressure: Why rank-based allocation of
reproductive trials is best. In D. Schaffer (Ed.), Third International Conference on Genetic Algorithms (pp. 116–121).
San Francisco: Morgan Kaufmann.
Artificial Life Volume 26, Number 3
365
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
6
3
3
3
8
1
8
9
6
1
0
4
a
r
t
l
/
_
a
_
0
0
3
2
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3