Martin M. Hanczyc**
University of Southern Denmark
Takashi Ikegami*,†
University of Tokyo
Palabras clave
Chemotaxis, cognition, homeostasis,
interfacial tension, oil droplet
Chemical Basis for
Minimal Cognition
Abstract We have developed a simple chemical system capable
of self-movement in order to study the physicochemical origins of
movimiento. We propose how this system may be useful in the study
of minimal perception and cognition. The system consists simply
of an oil droplet in an aqueous environment. A chemical reaction
within the oil droplet induces an instability, the symmetry of the oil
droplet breaks, and the droplet begins to move through the aqueous
phase. The complement of physical phenomena that is then generated
indicates the presence of feedback cycles that, as will be argued,
form the basis for self-regulation, homeostasis, and perhaps an
extended form of autopoiesis. We discuss the result that simple
chemical systems are capable of sensory-motor coupling and possess
a homeodynamic state from which cognitive processes may emerge.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
1 Introducción
Percepción, intelligence, and higher-order cognitive processes as currently understood are rooted in
sensory-motor coupling in organisms [1, 2, 7, 15, 31, 45]. En efecto, this coupling is being designed into
robotic systems to create artificial “living” machines (EcoBot [27] and SlugBot [22]). Sin embargo, el
fundamental basis for cognition may already be present in simple nonliving physical systems that pos-
sess a limited suite of properties also found in living systems. Intelligence could then be traced down
to physicochemical phenomena, such as thermodynamic fluctuations in open nonequilibrium systems
of rather simple composition that form by self-assembly, as we will see in our chemical systems. So the
question is, how can we derive sophisticated intelligence from a merely thermodynamic system? En
otras palabras, when is it necessary to use the intentional stance [12] to describe a systemʼs behavior—
Por ejemplo, by using sensing, or cognition, instead of reaction, or hysteresis?
Over the past few years we have been developing self-assembling chemical systems that are capable of
motility [17, 35, 36]. The chemical systems consist of an oil droplet in an aqueous water phase. The aqueous
phase contains a surfactant that forms the interface between the water and the oil and modulates the inter-
facial tension between the droplet of oil and its environment. We embed a chemical precursor (oleic an-
hydride) in the oil phase (nitrobenzene) that hydrolyzes into more surfactant when it comes in contact with
the water phase at the oil-water interface. This reaction not only powers the droplet to move in the aqueous
phase, but also allows for sustained movement as long as enough precursor oil remains in the droplet.
By embedding a catabolic chemical reaction in a self-assembled oil droplet body, we have determined
some of the conditions necessary to establish an interactive loop that involves the global movement of the
* Contact author.
** University of Southern Denmark, Institute of Physics and Chemistry, Campusvej 55, Odense M 5230 Dinamarca. Correo electrónico: martin@ifk.sdu.dk
† University of Tokyo, Department of General Systems Studies, Komaba, Tokio 153-8902, Japón. Correo electrónico: ikeg@sacral.c.u-tokyo.ac.jp
© 2010 Instituto de Tecnología de Massachusetts
Artificial Life 16: 233–243 (2010)
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
sistema (and thus can be interpreted as sensory-motor coupling) in a wet chemistry model defined by only
five chemical components, including water. In this article we describe the phenomena involved, en par-
ticular the mechanism of motility, and we speculate on how such a bottom-up approach can define the
naturally robust architecture necessary to evolve minimal cognition on top of an elementary type of
sensory-motor coupling. By way of studying this system, we reorganize the concept of autopoiesis, el
self-regulatory mechanism of a cell, around the terms of motility and homeodynamics. These are the vo-
cabularies proposed for considering a system as a cognitive system rather than solely a chemical system.
2 Description of the System
We study the movement of an oil droplet in an aqueous phase optically. The diameter of the droplet
can range from a few hundred microns to a few centimeters. An oil phase introduced into an aqueous
phase self-assembles into an oil droplet. Without any chemical or physical perturbation, the oil drop-
let will not move. It is only by creating an instability in the system that the oil droplet becomes dy-
namic. The instability in our system arises from the hydrolysis of the oleic anhydride in the oil phase,
and within seconds after the introduction of the oil to the water phase, the droplet starts moving (see the
Appendix on materials and methods).
The movement of the droplet is accompanied by convective flow inside the droplet. The convective
flow is established in the following steps. Primero, a chemical reaction occurs on the surface of a droplet. Como
more products are accumulated, the local pH decreases. The reaction does not occur evenly along the
surface, so that the local pH varies. We have measured the interfacial tension as a function of the surfactant
to show that the surface tension increases when the pH decreases (ver figura 2 en la sección 3.1). Por lo tanto
the local interfacial tension also varies along the surface. The subsequent interfacial tension gradient defines
the Marangoni force, whose direction is tangential to the surface. The Marangoni force drives the local flow
along the surface (called the Marangoni flow). A Marangoni instability describes the flow of surfactant
(liquid) along an interface to equilibrate an imbalance in interfacial tension and is the underlying physical
principle behind the “tears of wine” effect [43]. Once the initial symmetry breaks by fluctuation, a pair of
convective flows is organized. With numerical simulation [25], we demonstrated that the initial symmetry
breakdown is followed by quadratic-vortex formation, and pair convective flow follows.
The convective flow structure in the droplet brings fresh precursor to one pole of the droplet while
controlling the release of products on the opposite pole. Experimentally, the convective flow is observed
inside the moving oil droplet, with the centerline of flow along the direction of droplet movement (ver
Cifra 1). We believe that convective flow serves to create key feedback cycles in our system, and due to
convection, the reaction and movement will be sustained.
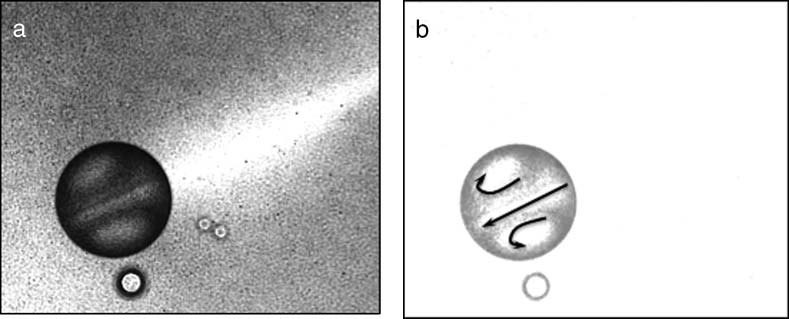
Cifra 1. DIC micrograph of a self-moving oil droplet with internal convection. (a) The droplet is moving toward the
bottom left while creating a bright trail. The characteristic flow pattern associated with convection is clearly seen within
the oil droplet. (b) Overlay of flow patterns seen in the oil droplet. The diameter of the droplet is nearly 0.1 mm.
234
Artificial Life Volume 16, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
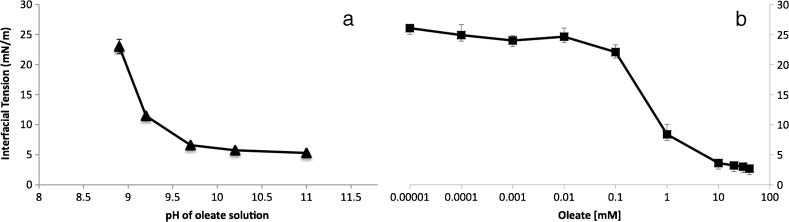
Cifra 2. Change in interfacial tension of a nitrobenzene oil droplet measured by pendant drop tensiometry and profile
análisis. (a) The interfacial tension of a pure nitrobenzene droplet in a 10 mM oleate solution with varying pH. (b) El
interfacial tension of a pure nitrobenzene droplet in solution at pH 11 with varying oleate concentration. Each measure-
ment was taken in triplicate. The interfacial tension of a drop of nitrobenzene at pH 11 with no oleate present is 27 mN/m.
The mechanism of self-motion is complex even in this simple system, with the law of reciprocal
action likely being dominant in the early stages and hydrodynamic pressure due to the fluid dynamics
in the latter stages of movement. Both the convective flow and the direction of movement are gov-
erned by unequal interfacial tension at the oil droplet boundary. The sustained imbalance in interfacial
tension as the droplet moves is likely caused by an observed local pH gradient that surrounds the
droplet. This chemical gradient is created by the droplet itself as a product of the chemical reaction.
The self-generated gradient can be overridden by an externally imposed pH gradient, and therefore the
direction of droplet motion may also be controlled. We observe that the droplet “senses” the gradient
in the environment (either self-generated or externally imposed) because the internal flow patterns and
direction of droplet movement change accordingly. When the droplet moves predictably within a pH
gradient, it exhibits chemotaxis in its most basic sense: directional movement governed by an external
chemical gradient. (Ver [17] for a complete chemical description of the system.)
3 Observations and Discussions
3.1 Sensory-Motor Coupling
Here we explain in more detail how a sensory-motor coupling arises in a simple chemical system. All the
complexity of this self-moving motion ultimately depends on the oil-water interface and the integration
of the sensor and actuator into the same dynamic structure. The interface is not a hard-shell container,
but a soft and flexible boundary under tension that can interact physically and chemically with the local
ambiente. It is the interface that is observationally responsive when exogenous chemical gradients
are added to the system. Por lo tanto, we consider the interface to be the sensor for the system.
The motor for the system arises from the flow structures observed within the oil droplet. Cuando el
interface senses a chemical gradient, such as in pH, an imbalance in the tension surrounding the droplet
results in flow structures due to a Marangoni instability. The observed flow structures with a droplet,
such as convection (ver figura 1), can act as the motor in the system.
We argue that both sensor and motor are present once the oil self-assembles into a droplet, and if so,
we can then begin to test the system for sensory-motor coupling. As the droplet ages, the anhydride
precursor fuel is hydrolyzed at the oil-water interface to produce more oleate and protons. Both prod-
ucts can affect the tension at the interface, as shown in Figure 2, and these factors modulate the re-
sponsiveness of the sensor. The oil droplets are usually added to an aqueous phase containing 10 mM
oleate at pH 11. The production of more oleate surfactant has little effect on the tension surrounding
the droplet, as the tension is already quite low (Figure 2b). Sin embargo, a local decrease in pH can have
a large effect, quickly reaching a tension maximum at pH 9 (Figure 2a). It is noted in experiments with
pH-sensitive dyes that the pH locally can decrease by several units, as low as pH 7 [17]. This change in
pH therefore can have a large effect on the tension surrounding the droplet and can cause a Marangoni
instability and induce flow. The sensing of local changes in pH by the interface induces flow, y el
Artificial Life Volume 16, Número 3
235
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
droplet starts to move. This form of pH-directed chemotaxis, as seen in our system, would be an ex-
ample of a type of sensory-motor coupling. We note that the droplets move chemotactically in pH
gradients even when the precursor fuel is not added to the system, but the movement is not sustained
and will stop once it successfully equilibrates any imbalance in tension (typically in a few seconds). Este
illustrates that instabilities in self-assembled systems induced externally can be resolved quickly. En cambio,
what we show here is that instabilities generated from within the system itself may allow for sustained
movimiento. The sustainability associated with this system is its novel feature. The system becomes sus-
tainable by circulating the reactants and products effectively as organized by the convective flow.
There are several mechanisms for chemotactic behavior in unicellular living systems. The mecha-
nisms for the motor include the rotation of the flagella in prokaryotes (p.ej., mi. coli [23]), actin-myosin
dynamics of the cytoskeleton (p.ej., Dictyostelium [6, 21, 37, 47]), movement of the cilia (in eukaryotes
such as Tetrahymena [10]), and gliding in cyanobacteria [18]. Each of these motility mechanisms is linked
to specific transmembrane receptors on the cell, acting as sensors. The sensors relay information about
the extracellular environment to the mechanical motors through intracellular signaling pathways in-
volving numerous proteins and small molecules [30, 39]. Although varying in sophistication [16, 46],
these sensory-motor couplings allow the organism to perform chemotaxis to find nutrients and avoid
waste and other poisons. The effects of sensing can be complex, with factors such as sustainability and
largo- and short-term memory coming into play. There have been some simulation studies that qualita-
tively reproduce the motion of the amoeba as well as chemotaxis [4, 29], where different diffusion rates
and production rates assigned for actin and myosin are responsible for the amoebaʼs motility. El
mechanism of movement and sensory-motor coupling in oil droplets is much simpler and does not
break down into modular units. En un sentido, the sensor and the motor are integrated into one, y
one does not make sense without the other. This kind of simple chemotaxis mechanism, as found in
our oil droplets, is not known in extant living systems.
Studying sensory-motor coupling with a simple chemical system has advantages. We do not need to
design and manufacture special devices or organs for sensor and actuator functions. En cambio, the sys-
tem self-organizes, and as a result many such oil droplets can be made simply and economically. Este
makes our chemical system attractive for those interested in studying self-organized systems that pos-
sess sensory-motor coupling, lifelike behaviors (p.ej., chemotaxis), and possibly the basic elements of
computing. The oil droplet system, because of its simplicity in composition, dynamic behavior in multi-
dimensional spaces, and possible emergent behaviors, could be used as an artificial life model system in
a chemistry laboratory, just as the game of Life is used in the virtual laboratory.
3.2 Shape Matters
By responding to a pH gradient with concomitant convective flow and movement, the droplet behaves
as if it can “perceive” the environment. (When and in what sense we can use this term “perception” will
be discussed in the following subsections.) We believe that the geometry of the interface shape can
control sensitivity to the environment. También, geometry-induced fluctuations can be the source of fluc-
tuation in motion [24, 28, 34, 38]. A coupling between fluctuations in interface geometry and fluctua-
tions in motion may be linked with the idea of biological autonomy. Por ejemplo, it has been found that
by mechanically pushing the cytoplasm of a cell (p.ej., Dictyostelium) one can elicit directional locomotion
[11]. The asymmetrical change of a boundary shape causes a polarization in actin and myosin protein
filaments, causing directional motion [44]. This internal polarization of biological chemicals may be re-
lated to our observations. In our typical experiments with droplets about 100 Am or less in diameter,
we do not observe any fluctuations in droplet shape as the droplet moves. Sin embargo, in larger droplets
hasta 0.5 cm in diameter, fluctuations in shape become readily apparent, as shown in the examples in
Cifra 3 (see also [32]). In such examples, both the distortion in shape and the fluctuation in motion
(velocity, direction) vary on the time scale of seconds (Cifra 4). The larger droplets are more easily
deformable, especially in the presence of surfactants, where the forces of gravity and perhaps flow struc-
tures counteract the Laplace pressure, which maintains the spherical shape [14]. Además, increasing
the droplet size enhances the instability of the internal flow, as the Reynolds number is proportional to
236
Artificial Life Volume 16, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
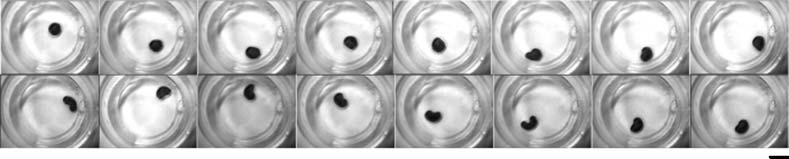
Cifra 3. Shape change in the boundary structure in moving droplets of different size. Droplets of size 1, 5, 10, y 30 AL were
added to the same aqueous phase and analyzed for fluctuations in geometry and movement (panels A, B, C, D, respectivamente).
Size bar, 1 cm.
the size. Cifra 3 shows the transition from the spherical shape to a horseshoe shape. We note that
straight directional motion is most supported by the horseshoe shape. This distorted boundary shape
may not support the convection flow seen in the smaller droplets. Por lo tanto, the mechanism of self-
movement shifts from convection-driven to shape-driven above around a few hundred microns in
droplet size. The detailed analysis of this phenomenon will be reported elsewhere [19].
3.3 Autopoiesis and Autonomy
Behavior can be treated as an extension of a basic idea of biological autonomy called autopoiesis [40].
Autopoiesis is a self-regulating mechanism of an internal metabolic network that maintains the bound-
ary of the cell. The autopoietic cell can be explicitly modeled by a simple stochastic automaton on a
Cifra 4. Fluctuations in droplet shape and movement over time. One droplet of 30-AL volume was added to the aqueous
phase, and each frame was taken at 8-s intervals. Size bar, 1 cm.
Artificial Life Volume 16, Número 3
237
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
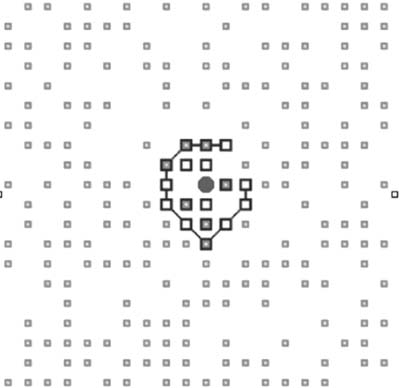
two-dimensional grid with abstract chemistry [42]. In this model, abstract chemistry is used to exemplify
el concepto. Substrates of the environment are transcribed into membrane materials by a catalytic par-
ticle. Those membrane particles will enclose the catalytic particle, so that when a membrane is broken,
it is immediately repaired by the catalytic particle; ver figura 5. This self-consistent relationship realizes
the concept of autopoiesis.
Varela and Maturana [26, 40–42] argued for organization of a minimal living system in terms of
autopoiesis. They discussed that a transition from a physicochemical system to a minimal living system
happens when a system performs an action on “what is missing on the part of the system,” in Varelaʼs
terminology. Por ejemplo, a system repairs its physicochemical boundary to maintain its “identity” (as in
our chemical system and Varelaʼs original experiment and model). This compensating process allows for
the emergence of a cognitive unit on top of the physicochemical layer. También, Varela argued that the self-
regularity, as well as the environmental regularities, is the result of coevolution between living systems
and their environment. Namely, the environment is also a part of living organisms. This is exactly what
we observed in our chemical experiments. We tend to think that the droplet becomes autonomous by its
own mechanism, but this is not completely accurate. Its environmental factors (pH and oleate concen-
tration, both of which are influenced by the action of the droplet; ver [17]) and also temporal treatment
(the freshness of the neat oil phase, pipetting actions) make autonomous movement possible. Incluso
after the autonomous droplet emerges, it is still controlled by the environment and its own temporal
cambios. This is what we consider to be the congruent regularity of the droplet motion, which is the
product of both droplet and environment.
Here we explore how autopoiesis is further developed through self-motility. The original concept of
autopoiesis does not explicitly assume the phenomenon of self-motion, nor does it argue what kinds of
self-motion are effective; but our chemical experiments and some simulated models (p.ej., [33, 34]) ex-
emplify the case where an artificial organism maintains the autopoietic system by the emergence of the
motile state. We consider the coexistence of both spontaneous and reflective (chemotactic) behavior an
extension of autopoiesis. Eso es, depending on the internal state and the environmental condition, a
droplet moves around in a certain way, at the same time changing the environmental condition (es decir.,
the modified environment can function as an external memory reservoir ). Speculatively, a droplet
may be able to “select” the action between autonomous and reflective to compensate for changes in
the velocity of the reaction. Además, there are some other types of action selection where a droplet
Cifra 5. The original simulation model of an autopoietic cell [26]. The many small squares are substrate (S) particles.
The larger squares, sometimes connected with other squares, are the link (l) particles. The single circular pattern in the
middle is the catalytic (C) particle.
238
Artificial Life Volume 16, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
spontaneously changes its direction of motion. Por ejemplo, by changing the strengths of outgoing flow
of product from the interface (through the law of reciprocal action), the droplet can navigate. El
potential reorganization of the droplet internal state as it switches between action modes is currently
under investigation.
The above dynamic component extends the notion of autopoiesis in that we can interpret the drop-
letʼs motion as action selection. Based on observation of the present droplet experiment, together with the
previous studies on autopoietic cells (p.ej., [33, 34]), we have a revised notion of autopoiesis as follows.
1. In order to have an autopoietic system, a boundary is a necessary ingredient that separates
the inside from the outside. The self-consistent relationship between the internal chemical
network and the organization of the boundary is what we call the autopoietic property of
the unit. (This is the original definition of autopoiesis.)
2. What is missing in the original picture of autopoiesis is temporal organization. The above
formal definition of autopoiesis does not tell whether autopoiesis is indeed realizable
in living systems. We need to know the dynamic stability and durability of autopoietic
properties in the real world. In the current oil droplet experiment, the droplet moves,
sometimes demonstrating chaotic itinerancy, and by doing so acquires autopoiesis.
En otras palabras, self-motility prevents the system from relaxing the nonequilibrium state,
so that it can sustain autopoietic properties over time.
3. Dynamic autopoiesis adds some new properties to a system. Primero, shape fluctuation of
the boundary assumes new degrees of freedom for the autopoietic cell. Whether the
shape is relatively round or rugged changes the compensating (therefore “intentional”)
action pattern of the cell. Segundo, the dynamic component lengthens the life span of the
autopoietic cell (in the previous models, the life span, defined by the time during which
a droplet moves, of the autopoietic cell was not controlled [34]). En otras palabras, el
dynamic autopoiesis changes the time structure of the system.
By considering the dynamic nature of autopoiesis, we noticed that a static boundary could be re-
placed by a convective flow structure. Now we have to answer the rather provocative challenge by one
of the reviewers: Why do we need to distinguish between sensing and reacting? This question naturally
leads to the question of when and where we switch the vocabulary from reaction to sensing and/or
percepción, and also, why we use the word sensory-motor coupling instead of (Por ejemplo) climbing
down the free-energy landscape. One answer is proposed by taking into account Varelaʼs work, el
emergence of compensating action to maintain the individuality of a droplet. Let us argue this in terms
of the game of Life, where we note several similarities to the chemical model described here.
In the game of Life, recurring patterns are termed gliders, and we use that term to describe the entire
dynamics instead of using the underlying bit patterns. Using that vocabulary can help to compress the
game of Life phenomena. Our first motivation was to find a gliderlike object in the chemical system, y
we have identified the droplet as such an object. Once the moving droplets appear, it is more efficient
to describe the phenomena in terms of droplets, because the reaction system and convective flow pat-
terns are hidden behind the outward phenomena. This is also true for describing robotsʼ behavior.
Their movements are nothing more than a pattern of the electric circuit and a sensory pattern hidden
behind it.
When an object emerges, the underlying constituents become unseen, and we can switch our way
of description. Por supuesto, in this droplet experiment, we see a clear connection between the underlying
chemical layer and the droplet layer, so that switching vocabulary is not strictly necessary. Sin embargo,
when we have successive hierarchical layers (p.ej., functional differentiation of the droplets) with out-
ward function overshadowing the lower-layer dynamics, we have more reason to use psychological
words in describing our system and developing our understanding of it. This droplet experiment is
one such step toward the complete vocabulary switch. Más, the droplet holds its own individuality,
a saber, a droplet becomes robust against perturbation, which is different from a glider. Por lo tanto, como
Artificial Life Volume 16, Número 3
239
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
a physicochemical entity, the droplet becomes an individual unit. In the next subsection, we consider
the underlying mechanism that holds individuality, a saber, homeodynamics.
3.4 From Homeostasis to Homeodynamics
Any perturbations in the interfacial tension of the boundary structure tend to be balanced through the
flow of surfactant. Once the tension forces around the droplet are balanced, the droplet stops moving.
Sin embargo, the emergent convective flow structure brings fresh precursor to the surface, where it be-
comes metabolized, and the system therefore maintains an imbalance in tension allowing for sustained
movimiento. This is an example of the homeodynamic state. Originally, homeostasis was described as a
property of a self-regulating system that sustains critical variables in a certain range [5, 9]. En el caso de
homeodynamics, a system changes the parameters or boundary condition to adapt to the environment.
Al hacerlo, the system dynamically organizes its own parameters. This leads to a peculiar state that we
call the homeodynamic state.
This idea of varying the systemʼs parameters is based on Ashbyʼs ultrastability [1]. In contrast with the
simple feedback loop that tunes the sensory-motor system, a second feedback loop exists to tune the
critical system parameters. The difference between variables and parameters is made explicit when writ-
ing down equations. Variables evolve temporally, and parameter values are fixed in time. But in real sys-
tems, the difference is likely not so simplistic. In case of the oil droplet, variables could be the center of
mass of the droplet, the velocity of the center mass, the amounts of chemicals, etcétera. The parameters
could be droplet size, pH, viscosity, reaction rate, etcétera. Pero, as we have seen so far, those variables
and parameters are mutually dependent. The critical parameters are changed to break up the homeostatic
state to make a system more robust against the environment, yielding a homeodynamic state.
Ashby proposed the concept of ultrastability for designing a brain system. We propose that a self-
moving oil droplet is a physical realization of homeodynamics and thus present it as a minimal
cognitive model. Self-movement regulates the chemical reaction. También, when a droplet is perturbed
externamente, it responds by resetting the flow pattern or changing the boundary shape. Shapes and flow
structures are the essential parameters of the droplet. The viable constraints on the droplet are de-
termined by those essential variables, so that positive feedback from the convection flow to the
chemical reaction is the second feedback in terms of Ashbyʼs ultrastability.
In recent studies of autonomous robotics, cognitive behaviors are characterized by a sensory-motor
coupling that can also be termed embodiment. The advantage of embodiment has been repeatedly stressed
in the field of robotics for the last two decades [3, 8, 31]. A missing notion in the robotics field is the
self-organization of self-movement and homeodynamics (a notable exception is Di Paoloʼs study of
homeoadaptation with an autonomous robot [13]). By pushing a step forward with the homeodynamic
aspect underlying any cognitive behavior, we say that the droplet can be a critical example for studying
minimal cognition. En efecto, the transition from homeostatic self (self-maintained statically) to homeo-
dynamic self (self-sustained dynamically) [20] emphasizes the potential for homeostasis as a source for
purposeful behavior even in simple systems.
4 Conclusión
Even simple chemical systems may tell us something about complex emergent phenomena such as
cognition. Using a bottom-up approach, we produced a simple oil droplet capable of sensing and mod-
ifying its environment, which results in autonomous self-movement of the droplet through an aqueous
phase. The boundary at the liquid-liquid interface serves as a highly sensitive and dynamic structure that
can perceive the environment. Once a pH gradient in the environment surrounding the oil droplet is
perceived, the droplet responds with movement within the gradient. The embedded chemistry of the
system fuels and reinforces sustained movement of the droplet. In this way the droplet maintains itself
through homeodynamic processes. We then begin to see an extended view of the self and autopoiesis
as a structure that maintains itself and its boundary through physically dynamic processes such as move-
mento. Such systems have a more active communion with their environments through perception,
240
Artificial Life Volume 16, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
Toma de decisiones, and even cognition. We hope to understand the fundamental aspects of cognition
through the intersection of simple physicochemical systems and cognitive science. Different from
the mere physicochemical process, a living system preserves its own identity and consistency with
respect to the environment by constantly readjusting the homeostatic state. Sensory-motor coupling,
rooted in homeodynamics, is the key to understanding minimal cognition and physical intelligence
conscripted and exploited by living systems [20].
When do we need to take the “intentional stance” [12] and resort to a new vocabulary? A critical
point is when a system sustains its structure by homeodynamics, eso es, a cyclical relationship be-
tween sustaining its internal process and self-motility, and thus behaviors emerge that can be named
and interpreted. Once self-motility becomes inevitable, we begin to see an extended view of the self;
autopoiesis is no longer just a stationary state that maintains itself. By realizing the homeodynamic
dimension of autopoiesis, we speculate that there would be no life without self-movement.
Expresiones de gratitud
We would like to thank the two reviewers for insightful comments and suggestions. We would also like
to thank Inman Harvey and Ezequiel Di Paolo for their discussions on Ashbyʼs ultrastability. Nosotros también
thank Tadashi Sugawara, Taro Toyota, and Naoto Horibe for their collaboration on the droplet experi-
mentos. The experimental work was supported by ProtoLife Srl, Italia, and the Center for Fundamental
Living Technology (FLinT), Dinamarca. The simulation is partially supported by the MEX project “De-
veloping Shape Language and Special Purpose Computing Systems for Simulating Abstract Chemical
Systems” (19300104) and “Emergence of Adaptive Motor Function through Interaction between Body,
Brain and Environment” (20033006).
Referencias
1. Ashby, W.. R. (1960). Design for a brain: The origin of adaptive behaviour (2y ed.). Londres: Chapman and Hall.
2. Beer, R. D. (2003). The dynamics of active categorical perception in an evolved model agent. Adaptado
Comportamiento, 11(4), 209–243.
3. Braitenberg, V. (1984). Vehicles: Experiments in synthetic psychology. Cambridge, MAMÁ: CON prensa.
4. Bentley, K., & Clack, C. (2004). The artificial cytoskeleton for lifetime adaptation of morphology. En
j. Pollak, METRO. Bedau, PAG. Husbands, t. Ikegami, & R. A. watson (Editores.), Workshop Proceedings of the 9th
International Conference on the Simulation and Synthesis of Living Systems (páginas. 13-dieciséis).
5. Bernard, C. (1957). An introduction to the study of experimental medicine. Nueva York: Dover.
6. Bottino, D., Mogilner, A., Roberts, T., Stewart, METRO., & Oster, GRAMO. (2002). How nematode sperm crawl.
Journal of Cell Science, 115, 367–384.
7. Arroyos, R. A. (1991). New approaches to robotics. Ciencia, 253(5025), 1227–1232.
8. Arroyos, R. A. (1999). Cambrian intelligence: The early history of the new AI. Cambridge, MAMÁ: CON prensa,
Bradford Books.
9. Cannon, W.. B. (1939). The wisdom of the body. Nueva York: norton.
10. Csaba, GRAMO. (1985). The unicellular Tetrahymena as a model cell for receptor research. Revisión internacional
of Cytology, 95, 327–377.
11. Dalous, J., Burghardt, MI., Muller-Taubenberger, A., Bruckert, F., Gerisch, GRAMO., & Bretschneider, t.
(2008). Reversal of cell polarity and actin-myosin cytoskeleton reorganization under mechanical
and chemical stimulation. Revista biofísica, 94, 1063–1074.
12. Dennett, D. C. (1989). The intentional stance. Cambridge, MAMÁ: CON prensa.
13. Di Paolo, mi. A. (2000). Homeostatic adaptation to inversion of the visual field and other sensorimotor
disruptions. In J.-A. Meyer, A. Berthoz, D. Floreano, h. Roitblat, & S. wilson (Editores.), From Animals to
Animats 6: Proceedings of the Sixth International Conference on the Simulation of Adaptive Behavior. Cambridge,
MAMÁ: CON prensa.
14. Fernandez, PAG., Andŕ́e, v., Riegera, J., & Kuhnlea, A. (2004). Nano-emulsion formation by emulsion
phase inversion. Colloids and Surfaces A: Physicochemical and Engineering Aspects, 251, 53–58.
Artificial Life Volume 16, Número 3
241
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
15. Fuster, j. (2003). Cortex and mind: Unifying cognition. Oxford, Reino Unido: prensa de la Universidad de Oxford.
16. Grebe, t. w., & Existencias, j. (1998). Bacterial chemotaxis: The five sensors of a bacterium. Biología actual,
8(5), R154–R157.
17. Hanczyc, METRO. METRO., Toyota, T., Ikegami, T., Packard, NORTE., & Sugawara, t. (2007). Chemistry at the oil-water
interface: Self-propelled oil droplets. Journal of the American Chemical Society, 129(30), 9386–9391.
18. Hoiczyk, MI., & Baumeister, W.. (1998). The junctional pore complex, a prokaryotic secretion organelle,
is the molecular motor underlying gliding motility in cyanobacteria. Biología actual, 8(21), 1161–1168.
19. Horibe, NORTE., Hanczyc, METRO. METRO., & Ikegami, t. (2009). Shape and motion dynamics in self-moving oil
droplets. In Proceedings of the 3rd International Conference on Mobiligence. Awaji, Japón.
20. Ikegami, T., & suzuki, k. (2008). From homeostatic to homeodynamic self. BioSystems, 91, 388–400.
21. Karakozova, METRO., Kozak, METRO., Wong, C. C., Bailey, A. o., Yates, j. r., Mogilner, A., Zebroski, h., &
Kashina, A. (2006). Arginylation of beta-actin regulates actin cytoskeleton and cell motility. Ciencia,
313, 192–196.
22. Kelly, I., & Melhuish, C. (2001). SlugBot: A robot predator. In ECAL 2001, LNAI 2159 (páginas. 519–528).
23. Larsen, S. h., Reader, R. w., Kort, mi. NORTE., Tso, W.-W., & Adler, j. (1974). Change in direction of flagellar
rotation is the basis of the chemotactic response in Escherichia coli. Naturaleza, 249, 74–77.
24. Mangome, NORTE., & Yoshikawa, k. (1996). Nonlinear oscillation and ameba-like motion in an oil/water
sistema. Journal of Physical Chemistry, 100, 19102–19105.
25. Matsuno, h., Hanczyc, METRO. METRO., & Ikegami, t. (2007). Self-maintained movements of droplets with
convection flow. In Lecture Notes in Computer Science (Including Subseries Lecture Notes in Artificial Intelligence
and Lecture Notes in Bioinformatics), 4828 LNAI (páginas. 179–188).
26. Maturana, h. r., & Varela, F. j. (1972). De máquinas y seres vivos, Santiago: Editorial Universitaria. Inglés
versión (1980). Autopoiesis: The organization of the living. In Autopoiesis and cognition: The realization of the
living. Dordrecht: Reidel.
27. Melhuish, C., Ieropoulos, I., Greenman, J., & Horsfield, I. (2006). Energetically autonomous robots:
Food for thought. Autonomous Robots, 21, 187–198.
28. Nagai, K., Sumino, y., Kitahata, h., & Yoshikawa, k. (2005). Mode selection in the spontaneous motion
of an alcohol droplet. Physical Review E, 71, 065301.
29. Nishimura, S. I., & Sasai, METRO. (2004). Inertia of chemotactic motion as an emergent property in a model of
an eukaryotic cell. In J. Pollack, METRO. Bedau, PAG. Husbands, t. Ikegami, & R. A. watson (Editores.), Artificial Life IX:
Proceeding of the 9th International Conference on the Simulation and Synthesis of Living Systems (páginas. 410–414).
Cambridge, MAMÁ: CON prensa.
30. Padre, C. A., & Devreotes, PAG. norte. (1999). A cellʼs sense of direction. Ciencia, 284, 765–770.
31. Pfeifer, r., & Bongard, j. C. (2006). How the body shapes the way we think: A new view of intelligence.
Cambridge, MAMÁ: CON prensa.
32. Sumino, y., Kitahata, h., Yoshikawa, K., Nagayama, METRO., nomura, S.-i. METRO., Magome, NORTE., & Mori, Y.
(2005). Chemosensitive running droplet. Physical Review. mi, 72, 041603.
33. suzuki, K., & Ikegami, t. (2004). Self-repairing and mobility of a simple cell. In J. Pollack (Ed.),
Artificial Life IX (páginas. 421–426). Cambridge, MAMÁ: CON prensa.
34. suzuki, K., & Ikegami, t. (2009). Shapes and self-movement in protocell systems. Artificial Life, 15(1),
59–70.
35. Toyota, T., Maru, NORTE., Hanczyc, METRO. METRO., Ikegami, T., & Sugawara, t. (2009). Self-propelled oil droplets
consuming “fuel” surfactant. Journal of the American Chemical Society, 131(14), 5012–5013.
36. Toyota, T., Tsuha, h., Yamada, K., Takakura, K., Ikegami, T., & Sugawara, t. (2006). Listeria-like motion
of oil droplets. Chemistry Letters, 35, 708–709.
37. Uchida, k. S., Kitanishi-Yumura, T., & Yumura, S. (2003). Myosin II contributes to the posterior
contraction and the anterior extension during the retraction phase in migrating Dictyostelium cells.
Journal of Cell Science, 116, 51–60.
242
Artificial Life Volume 16, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
METRO. METRO. Hanczyc and T. Ikegami
Chemical Basis for Minimal Cognition
38. Upadhyaya, A., & van Oudenaarden, A. (2003). Biomimetic systems for studying actin-based motility.
Biología actual, 13, R734–R744.
39. Van Haastert, PAG. J., & Devreotes, PAG. norte. (2004). Chemotaxis: Signalling the way forward. Reseñas de naturaleza
Biología celular molecular, 5, 626–634.
40. Varela, F. GRAMO. (1979). Principles of biological autonomy. Nueva York: Elsevier.
41. Varela, F. GRAMO. (1992). Autopoiesis and a biology of intentionality. In B. McMullin & norte. Murphy (Editores.),
Autopoiesis and perception: A workshop with ESPRIT BRA 3352 (páginas. 4–14). Dublín.
42. Varela, F. GRAMO., Maturana, h. r., & Uribe, R. (1974). Autopoiesis: The organization of living systems, es
characterization and a model. BioSystems, 5, 187–196.
43. Velarde M. GRAMO., wilson, S. K., & Helliwell, j. R. (1998). Marangoni and interfacial phenomena in materials
Procesando. Philosophical Transactions of the Royal Society A, 356, 829–844.
44. Verkhovsky, A. B., Svitkina, t. METRO., & Borisky, GRAMO. GRAMO. (1999). Self-polarization and directional motility
of cytoplasm. Biología actual, 9, 11–20.
45. von Uexküll, j. (1926). Theoretical biology. Londres: Kegan, Pablo, Trench, Tubner.
46. Yumura, S., & Fukui, Y. (1998). Spatiotemporal dynamics of actin concentration during cytokinesis
and locomotion in Dictyostelium. Journal of Cell Science, 111, 2097–2108.
47. Yumura, S., Mori, h., & Fukui, Y. (1984). Localization of actin and myosin for the study of ameboid
movement in Dictyostelium using improved immunofluorescence. Journal of Cell Biology, 99, 894–899.
Apéndice: Materials and Methods
A.1 Oleate Surfactant
Neat oleic acid oil (NuChek Prep, Elysian, Minnesota) was added to alkaline water to make varying con-
centrations of oleate micelles at pH 11. NaOH was adjusted to make oleate solutions at varying pH
for tensiometry.
A.2 Oil phase
Neat oil of oleate anhydride (Fluka) was added to nitrobenzene (Fluka) en 0.5 METRO. Pure anhydride
was used in neat oil form. Anhydride oil stocks were stored away from direct light and under argon.
A.3 Glass Slide Experiment with DIC Microscopy
Surfactant (100 mL) was added to glass slides with a concave depression (VWR, Milan, Italia) of 1.5-cm
diameter. An oil droplet was then added, and movement was monitored using microscopy. Running
droplets were analyzed in real time using an inverted Olympus differential interference contrast (DIC)
microscope, BX51, equipped with a Toshiba CCD camera connected to a video-recording system
(Sony, WV-DR9).
A.4 Glass Dish Experiment
Aqueous phase (800 mL) was added to a glass dish of 35-mm diameter with a 27-mm-diameter quartz
base. An oil droplet was then added, and movement was recorded using an iSight digital video camera
and iMovie software.
A.5 Tensiometry
Interfacial tension of the oil-water interface was determined using a PAT1D tensiometer (Sinterface) por
the pendent drop method using the Sinterface software. All samples were prepared with nitrobenzene
as the internal phase and an aqueous phase containing varying concentrations of oleate or 10 mM oleate
with varying pH. All values were taken after the tension reached a steady state, and each condition was
tested in triplicate.
Artificial Life Volume 16, Número 3
243
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
a
r
t
yo
/
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
1
6
3
2
3
3
1
6
6
2
6
7
3
a
r
t
yo
/
_
a
_
0
0
0
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3