Biología Computacional
and the Limits of Shared
Vision
Annamaria Carusi
Oxford e-Research Centre,
Universidad de Oxford
Several studies have focused on the social sharing of visual practices as consti-
tutive of evidence within a domain, while there has been relatively less atten-
tion paid to points where the social sharing of practices breaks down, or is re-
sisted. This article argues that a study of both types of cases is necessary in
order to gain a better perspective on social sharing of practices, and on what
other factors this sharing is dependent upon. The article presents the case of
currently emerging inter-disciplinary visual practices in the domain of com-
putational biology, where the sharing of visual practices would be beneªcial
to the collaborations necessary for the research. Computational biology in-
cludes sub-domains where visual practices are coming to be shared across dis-
ciplines, and those where this is not occurring, and where the practices of oth-
ers are resisted. A signiªcant point of difference between these sub-domains is
between visualizations that render the output of simulations and those which
are images taken during observations using the techniques of microscopy. A
crossing over, compromise or sharing of practices relating to these different
sub-domains is difªcult and often resisted. This resistance needs to be con-
textualised in a far richer account of the relations between the visual arti-
hechos, the scientists who use them within disciplinary domains, the theoretical
and instrumentational outlook of the disciplines in question, and that to-
wards which the science is directed, its domain of study. Social practices alone
are not sufªcient to account for the shaping of evidence. The philosophy of
Merleau-Ponty is introduced as providing an alternative framework for
thinking of the complex inter-relations between all of these factors. Este
I would like to thank all the biologists, of all the different sub-disciplines, who partici-
pated in this study, and especially Blanca Rodriguez and James Wakeªeld. The paper also
beneªted from the generous input of two anonymous reviewers for Perspectives on Science, a
whom I very grateful. Responsibility for any shortcomings is of course mine.
Perspectives on Science 2011, volumen. 19, No. 3
©2011 by The Massachusetts Institute of Technology
300
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
301
philosophy enables us to think of the inter-constitutive relations between these
different factors, which ultimately deªne an epistemological and ontological
space in which the object of study itself has an active constitutive role, and in
which the scientist as person and perceiving body within a knowledge domain
is also constituted.
Since the 1980s, several studies of visual perception have persuasively ar-
gued that important aspects of human vision are best accounted for not by
recourse to inner mental representations but rather through socially ob-
servable actions and behaviors (p.ej. Linchar 1985, Latour 1986, Linchar
1990, Goodwin 1994, Goodwin 1997, Sharrock & Coulter 1998). Mientras
there are clearly physiological mechanisms required for vision, psychologi-
cal accounts of perception in terms of inner mental representations have
been dislodged from their position as the basic term in the interface be-
tween human beings and their environment and replaced with terms such
as “social practice,” and “vernacular intelligibility.” The focus for these
theorists in accounting for vision shifts to intersubjective rather than sub-
jective accounts of vision, with demonstrations regarding the ways in
which what is seen within an environment by any one person depending
on the sharing of that environment with others and on the practices for
sharing of the group. Eso es, members of groups and communities tend
to pick out and perceive the same kinds of entities and processes, y el
same kinds of properties and features. In these accounts, the inner world of
invisible and mystical mental representations is replaced by the outer
world of visible and observable social doings. Often the later philosophy of
Wittgenstein is claimed as a philosophical framework that informs this al-
ternative view, with the notion of language game being interpreted socio-
logically as showing that the meanings of terms, utterances, and “moves”
are determined by social agreement alone. Phenomenology is generally
claimed as another important philosophical framework, in particular the
social phenomenology of Alfred Schutz which focuses on the ways in
which a subjectively shared lifeworld conditions subjective experiences
(Schutz 1973).
The work of Maurice Merleau-Ponty on the phenomenology of percep-
tion is less often explicitly drawn upon in this tradition of social account-
ing for perception but shares many of the same concerns. Merleau-Ponty’s
best known work is The Phenomenology of Perception (1962) and this work
shares the anti-representationalist and anti-subjectivist approach to vision
sketched out in the previous paragraph. Over the course of the develop-
ment of his thought, Merleau-Ponty was increasingly concerned to dis-
solve the dichotomies between subjective and objective, and between in-
ner and outer, that tend to be a feature of representationalist accounts of
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
302
Computational Biology and the Limits of Shared Vision
perception by focusing instead on the perceiving body. This shift in focus
allows for an understanding of perception in terms of the actions and in-
teractions of subjects within an environment. The extent to which the
boundaries between the two are porous, and the fact that these boundaries
are by the very same token connectors, is especially clear in the notion of
the body schema, which Merleau-Ponty had initially borrowed from neu-
rology and then psychology. The body schema is that through which
perceivers place and position themselves relative to other perceivers and to
their environments as a whole; it is a type of scaffolding or architecture
which is both “of the body” and “of the world” having been traced out
through sedimented and ongoing transactions between the two. In works
such as Visible and Invisible (1968), Merleau-Ponty goes on to further erode
the subject-object dichotomy, although in order to do so his philosophy
takes an ontological turn. The perceiving body is both subject and object,
y, more important, reversibly so: we perceive because we interchange-
ably play both these roles. Sin embargo, there is a perceptual blind-spot in
that we cannot experience ourselves as both at the same time—as my left
hand touches my right, I can experience my left hand as touching and thus
as subject of the experience; and I can experience my right hand as
touched and thus as object, but there is a lacuna, a gap in the experience of
both at the same time.Due to the way in which attention operates in expe-
rience, we cannot but swivel from the one to the other without reaching
the point of stasis where both are present to awareness. This is the limit
beyond which it is impossible to go and still remain within traditional
phenomenology. Moving beyond, Merleau-Ponty begins to unfold an on-
tology around the structures of being that make it possible for the revers-
ibility and intertwinement of subjects and objects of perception, and in
Visible and Invisible he does this through the notion of ºesh. By the term
“ºesh” Merleau-Ponty means “sensibility in general.” “Flesh” expresses
the connection that there exists between subjects and objects of perception
in the very fact of the capacity to perceive of subjects and the capacity to
be perceived of objects. It is described as the element which deªnes them
in relation to each other (Merleau-Ponty 1968, páginas. 193–194), and also as
the principle which allows the body and its environment to open onto one
another—la déhiscence—, and for their crossing over or reversal (their criss-
crossing)—le chiasme. Like the principle which allows two mirrors placed
in front of one another to reºect one another without the reºections being
“located” in either one of the mirrors, and which can be expressed as
reºectibility in general, “ºesh” is a feature of neither the subject nor of the
object, but denotes the relation between them of sensibility in general.
“Flesh” as such is not experienced; rather it frames the ways in which the
environment “answers” to shared acts of perception. Rather than shared
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
303
practicas, Merleau-Ponty often speaks of shared modes or “styles” of per-
ception which are produced through people interacting with each other
and with their environments (Merleau-Ponty 1968).1
Besides the unªnished manuscript of Visible and Invisible (1968), cuando
he died Merleau-Ponty also left a large body of notes on the life sciences,
where he continued to elaborate the detail of this ontology of ºesh. El
work that has been published in English under the title Nature: Course
Notes from the Collège de France (English trans. 2003, French original 1995)
as a collection of notes by Merleau-Ponty and of his students, giving a tan-
talising glimmer of the account which Merleau-Ponty himself might ulti-
mately have put forward. It is particularly interesting for ongoing devel-
opments in the different strands of biological research that Merleau-Ponty
turned, esta vez, to biology, just as previously he had turned to neurol-
ogia, psicología, and psychoanalysis. For Merleau-Ponty in this late work,
the ontology of vision and the ontology of nature are indissoluble. It is im-
possible within the scope of a single paper to show the full signiªcance of
Merleau-Ponty’s sketched-out thoughts and ideas for vision in general and
for vision in the domain of biological seeing in particular. en este documento, I
undertake something on a much smaller scale, and that is to show how fol-
lowing Merleau-Ponty down this avenue of thought might be helpful as a
way of deepening accounts of perception which focus on the social sharing
of practices as constitutive of important aspects of vision, opening these
accounts onto the ways in which vision is formed at the interstices be-
tween seeing/seen beings and things. It becomes clear that there is a need
to push beyond the privileging of shared social practices in the under-
standing of vision where there is a failure to share visual practices, dónde
sharing is resisted or collapses. At these junctures, it is possible to claim
that this is because different groups and cultures have different practices
which are not necessarily shared. Sin embargo, it is also possible to take a dif-
ferent, ontological, route, which is the one I explore in this paper.
This paper focuses ªrst, on accounts of vision in specialised environ-
mentos, where expertise or specialised professional skills are in use, and sec-
ond, on issues that emerge when we try to account for the failure to share
visual practices that occurs at particular junctures. Computational biology
is a domain where there are a variety of visual practices, which sometimes
converge and overlap and sometimes compete. An ontological turn is use-
ful as a way of making sense of the resistance to sharing visual practices
1. An even earlier philosophical account is that of Immanuel Kant who hinted at the
central role of shared modes of seeing by invoking the “sensus communis” or common sense
in judgement. Although the argument about the sensus communis is to be found in the Third
Critique, Kant sees it as an argument relevant to knowledge in general, including the per-
ceptual knowledge which he deals with in the First Critique. See Carusi 2008.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
304
Computational Biology and the Limits of Shared Vision
that can occur in the computational biology arena. The paper explores this
claim and shows one way—a Merleau-Pontian way—of “doing” the onto-
logical turn, and what it could offer to philosophical and sociological ac-
counts of vision in science.
The paper begins with a consideration of a paradigmatic paper in the
approach to vision in science as a matter of socially shared practices,
Charles Goodwin’s “Professional Vision” (1994), and raises questions re-
garding the extent to which an emphasis on socially sharing visual prac-
tices obscures the role of objects of vision in specifying the visual situa-
ción. Since instances of contested vision present themselves as interesting
cases for examining the limits of shared vision, the case study of computa-
tional biology is introduced in the second section. Section 2a introduces
computational biology as a sprawling area of interdisciplinary research in
biology, comprising both early stage collaborations which are often still
fraught with difªculties of sharing practices, and mature collaborations
where there is a rapprochement between disciplines. The visualizations
used are often indicative of the different stages of collaboration, with visu-
alizations in mature collaborations serving signiªcant epistemic functions
for researchers across the collaborations, whereas visualizations of early
stage collaborations are still embedded ªrmly in one or other of the differ-
ent disciplines. Section 2b describes central features of visualizations in es-
tablished computational biology, while section 2c delves deeper into a
consideration of visualizations in early stage computational biology, cual
in section 2d are related to the context of observational experimental biol-
ogia. Section 2e considers the types of exchanges across the contexts of the
different visual practices in computational biology. The discussion in sec-
ción 3 once again poses the question regarding why some exchanges seem
more available than others, and brings to the fore the very different
ontologies of that which is observed in a computational visualization as
opposed to that which is observed through a session of microscopial obser-
vación, and the way in which this ontology speciªes differently not only
the ontological nature of the object seen but the act of seeing and the
viewer. Merleau-Ponty’s notion of the circuit is drawn upon as a way of re-
thinking the relation between viewers and that which is seen, which al-
lows for an internal relation of co-constitution between them.
Sección 1: Shared visions
Social accounts of vision focus on shared practices, modes or styles of vi-
sion that enable human perceivers not merely to agree on what they per-
ceive, but to perceive the array of objects, events, and other features that
are characteristic of human existence. Several writers have discussed the
ways in which scientiªc seeing is socially mediated, constructed, or consti-
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
305
tuted through processes which essentially involve shared practices, styles,
or modes of perception. The central message of these accounts is that
scientiªc knowledge, evidencia, observación, and perception are contingent
on social or communal aspects of science.
Histórico, sociological, and philosophical accounts of vision and visual
practices in professional and scientiªc domains have yielded fascinating
results detailing the variety of ways in which convergence on ways of see-
ing is obtained through techniques, activities, practicas, routines, and cul-
tures.2 More rare are accounts which deal with a breakdown in sharing, a
lack of agreement or downright disagreement, or an inability to come to
see as others do, even when the potential for this would appear to be quite
obvious.3 Goodwin’s “Professional Vision” (1994) is an interesting exam-
por ejemplo. en este documento, Goodwin contrasts two contexts in which visual prac-
tices play a central role in enabling others to come to see important fea-
tures of visual artifacts or ªelds. It is not simply coming to see that is at
issue, but rather coming to see as someone else sees, in one case, the “someone
else” being a professor, and in the other, a provider of court testimony. El
ªrst case is that of archaeology professors training students to “see” in an
archaeological dig; the second is the case where lawyers defending the po-
lice ofªcers on trial for the beating of Rodney King use video evidence to
convince the jury that the ofªcers’ actions did not constitute an abuse.
Goodwin sees close analogies between the two cases. He writes: “It would
however be quite wrong to treat the selective vision that is so salient in
the King trial as a special, deviant case, merely a set of lawyers’ tricks de-
signed to distort what would otherwise be a clear, neutral vision of objec-
tive events unambiguously visible on the tape. All vision is perspectival
and lodged within endogenous communities of practice” (1994, pag. 606).
The juxtaposition of the King trial with the archaeology professor induct-
ing a student into archaeological ªeld excavation is meant to show the
similarities across the two situations, that is that both involve discursive
practices which “shape events” within a ªeld (pag. 606). De este modo, coming to see
as someone else sees is not only seeing in the same way (the same mode or
2. The literature on this point is too large to cite. Representative are Lynch & Woolgar
1990, Baigrie 1996, jones & Galison 1998.
3. Peter Galison’s (1997) account of the image and logic traditions of experimental and
theoretical physicists is a notable exception, showing how these traditions struggled—and
sometimes still struggle—to come to mutual comprehension. His notion of the trading
zone as a way for scientists to come to sufªcient understanding so that they can at least col-
laborate in a mutually beneªcial way is useful; sin embargo, he sees scientists as interacting
primarily through language, or gives a linguistic metaphor of their interactions, claiming
that scientists in interdisciplinary domains develop pidgins and creoles which allow them
to operate in the trading zone. On this theory, ways of seeing are contingent upon trading
zone languages, an alternative which cannot be pursued in this paper.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
306
Computational Biology and the Limits of Shared Vision
style of seeing), but seeing what others in that same endogenous commu-
nity see. Goodwin stresses this, writing of the use of video evidence in the
King trials that “[oh]pposing sides in the case used the murky pixels of the
same television image to display to the jury incommensurate events: a
brutal, savage beating of a man lying helpless on the ground versus careful
police response to a dangerous ‘PCP-crazed giant’ who was argued to be in
control of the situation,” and that “[a]n archaeologist and a farmer see
quite different phenomena in the same patch of dirt.” Goodwin writes
that the process of socially shaping vision “creates the objects of knowl-
edge that become the insignia of a profession’s craft: the theories, artifacts
and bodies of expertise that are its special and distinctive domain of com-
petence” (1994, pag. 606). He provides a fascinating account of the details of
the way in which this is achieved. Sin embargo, it is an account which ulti-
mately leaves one perplexed as to the nature of these events and objects so
powerfully formed by socially shared visual practices. From where does the
disagreement over what is seen in the video evidence—which gave rise to
two different verdicts at the two trials held—emerge? Is it simply that in
one case the jury shared the vision of one set of lawyers, and in the other,
they shared that of another set of lawyers? What makes the difference
between one or the other (besides the force of rhetoric alone, which is cer-
tainly a possibility)? Or did they never come to share the practices of
either?
If we consider the kinds of differences between what is seen (a brutal
beating or responsible police action; soil for crops or soil hiding archaeo-
logical treasures), it is clear that what is seen is not a matter of vision as an
isolated act of perception alone, but vision embedded in contexts of ac-
ciones, interactions, purposes, and motivations. Seeing is not only a matter
of shared visual practices, but a range of other complexly related practices,
comportamiento, and behaviors. Eso es, what is seen is always a matter of the
whole sphere of human actions. Sin embargo, implicit in this socially con-
textualized understanding of vision is another important role-player, eso
es, the things with which humans interact, the objects whereby they seek
to fulªll their goals, the physical and natural environments in which they
interact as social beings. To use a grammatical metaphor: Las acciones son
something like transitive verbs, in that they need objects, direct or indi-
recto. To overly stress the role of actions is like considering the meaning of
a sentence as a matter of the verb alone. This is a bad account of sentence
meaning and a truncated account of vision. A shift of focus is necessary for
the objects of vision to come back into view and to take their place in the
whole visual situation, comprising viewers as well as what they see.
Por supuesto, sociological theories of science have not been slow to per-
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
307
ceive this challenge and to try to meet it. There has been a clear shift from
the “docile objects” which were the only form in which objects were ad-
mitted in accounts of scientiªc vision, Por ejemplo, in an early paper of
Michael Lynch (1985, pag. 43). To name but two interesting developments
that give more nuanced accounts of the ontological space in which com-
plex social practices, such as the conduct of science, occur: (1) Actor Net-
work Theory developed by Bruno Latour, John Law, and Michel Callon try
to discard the assumption that in any ªeld, some things are subjects and
some things are objects, and attempt to adopt a position of neutrality. El
extent to which this can be achieved is contentious, but this is a challeng-
ing move. Y (2) Karen Barad adopts a position of agential realism for
human and non-human bodies, with phenomena produced by ontologi-
cally inseparable agential intra-actions (Barad 2003).
Merleau-Ponty did not see himself contributing speciªcally to the un-
derstanding of vision in science or other specialised domains. Además,
he did not pause long on the role of technologies in making visible objects
of science. Yet his theory is an unplumbed resource for a theory of vision in
scientiªc contexts which pushes against an ontology skewed towards so-
cial actors.
Sección 2: Failures of vision
In the previous section the question was asked, What might be the sources
of resistance to socially sharing visual practices? To pursue this line of in-
quiry it is necessary to focus on cases where practices are not shared and
where there is real resistance to their being shared, as much as where they
are shared. Emerging shifts in disciplinary boundaries in scientiªc re-
search are generally good places to look for such examples, since they pres-
ent examples where visual practices are transferred between domains, o
where instead they are not transferred but resisted. This is what is occur-
ring in the shifting boundaries around computational biology, with some
sub-domains within computational biology showing a high degree of dis-
ciplinary fusion, others showing a lesser degree of fusion but some co-
operación, and others instead resisting entering the domain altogether. Él
is a good ªeld of inquiry for this exploration, since it offers examples of
shared visions and contested visions within the same broad ªeld of study.
At stake in the different visual practices that converge or diverge in and
around computational biology is the reconstitution of the ªeld of biology.
The study of the visual practices of and around computational biology,
against their broader epistemic backgrounds, allows us to test the limits of
sharing visual practices. Accounting for these limits entails drawing on
the ontology of vision in these sub-domains.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
308
Computational Biology and the Limits of Shared Vision
Section 2a: The research world of Computational Biology
The case described in this paper centres on Computational Biologists
and their collaborators. Research in computational biology is progress-
ing across many sub-domains within biology, for example biochemistry
and physiology, though at various paces. Computational Biology can be
deªned as the application of computational methods to the study of bio-
logical processes, speciªcally the methods and techniques of modelling,
simulation, and visualization. Computational Biology is closely related to
Systems Biology, Mathematical Biology, and increasingly, Synthetic Biol-
ogia. It is a multi-disciplinary area of study, drawing on the expertise and
knowledge of a wide array of people from various disciplines: computer
científicos, numerical analysts, engineers, mathematicians, biophysicists
and a broad range of the life sciences, physiologists, developmental biolo-
gists and others. Computational biology crucially depends upon cross-
disciplinary collaborations in order to be successful. This means that it de-
pends on getting on board researchers who have fundamentally different
visual and epistemic practices. Visual artifacts play a pivotal role in com-
municating the epistemic goals and outputs of computational biology
across this range of disciplinary diversity. One of the marks of success in
establishing collaborations is the use of the same visual artifacts across the
different disciplinary groupings. Visualizations of computational simula-
tions are common currency among the members of successful collab-
orations being used for analyses, in the context of workshops, and in pub-
lications. Where instead there is no collaboration, or a greater difªculty of
establishing one, there is not a circulation of visual currency. Hay
several reasons for this of course, including social and institutional factors.
Sin embargo, the nature of visual artifacts used across disciplines, the style of
observation with which they are closely connected, and the ontological na-
ture of that which is observed also differ substantially.
Collaborations between experimental biologists and computational bi-
ologists, eso es, broadly speaking, mathematicians, computer scientists,
physicists, and engineers, are what keep the machinery of computational
research going. Some collaborations, such as those found in cardiac model-
ling, are longstanding and have a long history. Others have a far shorter
history and for various reasons—including greater biological and mathe-
matical complexity—are more difªcult. en este documento, a variety of domains
are considered: cardiac modelling, modelling of multicellular systems
such as the modelling of tumours, and domains which can be described as
qualitative observational biology, such as embryology and cell biology.
The case study emerges out of ongoing participative research with mem-
bers of both groups and includes observations, interviews, participation at
workshops and seminars, and my own teaching of interdisciplinary re-
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
309
search skills in the context of a doctoral course for computational biology
estudiantes. A Forum on Scientiªc Method in Biology was held as part of this
estudiar: this was an event to which a small group of biologists “of different
persuasions” were invited to discuss with each other what biological
method is, with the aim of understanding which methods are considered
to be likely candidates for arriving at speciªcally biological questions and
answers (Carusi et al. 2009).4
En primer lugar, something of the general domain of computational biology
needs to be understood. Computational biology attempts to model biolog-
ical processes, usually in the form of ordinary or partial differential equa-
ciones. The parameterisation of the models (through which they are said to
“gain reality”) is accomplished by using experimental data. This data can
come from published literature; sin embargo, ideally it will come from collab-
orators in experimental biology. Such collaborators are very valuable to
computational biologists because they run experiments which supply the
kinds of data needed for the construction of the models, and ultimately for
their validation. Models are a quantitatively expressed hypothesis regard-
ing the phenomena observed in the experiment and are tested at two
puntos. En primer lugar, through the simulation, which allows a testing of different
solutions of the equations and a reªnement of the initial hypothesis con-
cerning the underlying mechanism. En segundo lugar, in the comparison between
the output of the simulation and experimental data. Precisely how this
comparison is carried out is a crucial point in the validation of the models.
While there is not yet general consensus among the disciplines in-
volved concerning the epistemic status of mathematical models, one view
is that mathematical modelling is a hypothesis generating (and therefore
predictive) device; a mathematical model, sin embargo, is not computational
en si mismo, and is in fact generally worked out using pen and paper.5 The
mathematical model is in the form of differential equations; for it to be
solved, numerical analysis and computational algorithms are used to es-
tablish the possible numerical values of the derivatives. This is carried out
computationally in a simulation. The output of the simulation is visual-
ized, and the observation and analysis of the visualization is a central part
of the epistemic practice of the computational biologists.
Sin embargo, collaborations are not always successful, since computational
4. The Forum was convened by Annamaria Carusi (philosopher/STS scholar), Blanca
Rodriguez (computational biologist) and James Wakeªeld (cell biologist); other partici-
pants were: Brian Goodwin, Denis Noble, Evelyn Fox Keller, Kevin Burrage, Philip
Maini, Lynn Margulis, Derek Terrar, Eric Werner, Tim Horder.
5. And in fact, is more likely to be worked out in this very low tech fashion as was dis-
covered in our investigation into the working practices of the Integrative Biology team
miembros (Mascord, Jirotka & Carusi 2007).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
310
Computational Biology and the Limits of Shared Vision
biology is not universally accepted as a positive approach and methodol-
ogy in biological research.6 The difference in the acceptance and ease of
collaboration can result from differences in the “stages of development” of
the research program in the various pockets of biological research, Alabama-
though this assumes that all biological sciences will eventually be “ma-
ture” enough to develop into the use of full-ºedged computational meth-
probabilidades. A veces, sin embargo, non-acceptance can come from deeply held
allegiances to particular observational practices. This is what will be dis-
cussed in the next few sections.
A brief and necessarily superªcial history of heart and cancer modelling
goes some way to explaining why collaborations in the former are rela-
tively easier than collaborations in the latter.
Heart modelling has a longer history than cancer modelling, going
back to the 1953 Hodgkin and Huxley model of electrical stimulation in
excitable cells such as neurons and cardiac myocytes. Denis Noble
modiªed the model so that it could also be used “to describe the long-
lasting action and pace-maker potentials of the Purkinje ªbres of the
heart” (Noble 1962, pag. 318). Since then there has been an extensive exper-
imental program generating data to parameterise the models, and to vali-
date them. For a variety of reasons, primarily having to do with (1) differ-
ences between whole-organ and cellular processes, (2) the tractability of
these processes tractability to understanding through mathematical ap-
se acerca, y (3) the only relatively recent development of mathematical
and computational techniques capable of dealing with the levels of com-
plexity of multicellular biological processes, cancer modelling is at an ear-
lier stage of development.
In the heart modelling community, there is a long-established collabo-
ration between experimental physiologists, mathematicians, and com-
puter scientists. Heart modelling and in silico experiments started off with
individuals who embodied inter-disciplinarity in that they were experi-
mental physiologists, and therefore already geared towards the quantita-
tive analysis of experimental data. There has been a long line of experi-
mental physiologists who knew enough mathematics to develop their own
modelos. As they conducted the experiments themselves, they were also
able to test the models themselves: there was no gap between the develop-
ment and testing of a model. As the models became more complex, profes-
sional mathematicians began to work in the area, and professional com-
puter scientists also began to collaborate when there arose a need to run far
more complex algorithms at a greater speed. Sin embargo, among mathemati-
6. See for example Horder 2008, and Carusi et al. 2009.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
311
cians, computer scientists, and physiologists there was at least a shared
understanding of the need for the collaboration, and a shared commitment
to the use of quantitative methods, at least alongside observational meth-
probabilidades. This creates a natural collaboration between experimental physiolo-
gists and mathematicians/computer scientists. Because of the increasing
complexity of the problems and the need to speed up the algorithms, truly
interdisciplinary individuals become scarcer, and there is a greater need for
distribution of expertise. Cooperation and collaboration are pre-requisites
for progress (Welsh et al. 2006), and have been in place in some instances
at least since the 1980s.
Whereas computational and mathematical approaches to whole organ
procesos (such as electrical currents in the heart) require collaboration
with physiologists, in the domain of tumour growth and other multicel-
lular processes, collaboration with cell biologists is required. Sin embargo,
cancer modelling and cellular biology generally have a much shorter his-
tory of using quantitative techniques, with fewer people able to span the
divide between biology and mathematics, in view of the greater complex-
ity of the biological processes and thus also of the mathematics. De este modo, es
more difªcult to be a fully interdisciplinary practitioner. The corollary of
the absence of an overlapping space has been the entrenchment of very
different epistemic cultures, one of which is predominantly qualitative,
whereas the other is predominantly quantitative. This makes co-operation
and collaboration far more difªcult to achieve, and the computational bi-
ology program in these domains becomes more difªcult to establish.
A simpliªed computational biology research ºow is illustrated in the
diagram in ªgure 1. From this, we see that there is a ºow between wetlab
and drylab, or between laboratory experiments and computationally simu-
lated experiments. Wetlab experiments, carried out by experimental biol-
ogists and physiologists, supply data in the form of parameter values.
Mathematical models are produced by mathematical biologists in the
form of ordinary or partial differential equations and parameterised (allá
are various approaches to this: bottom up, De arriba hacia abajo, and middle out; ver
Brenner et al. 2001; Krohs & Callebaut 2007). The differential equations
of the models are solved in the simulations, in a process requiring the in-
put of numerical analysts and computer scientists. The output of the sim-
ulation is visualized, and this output is then compared against experimen-
tal results.
The research strand devoted to visualizations is a central one in the
overall computational biology program, with doctoral dissertations and
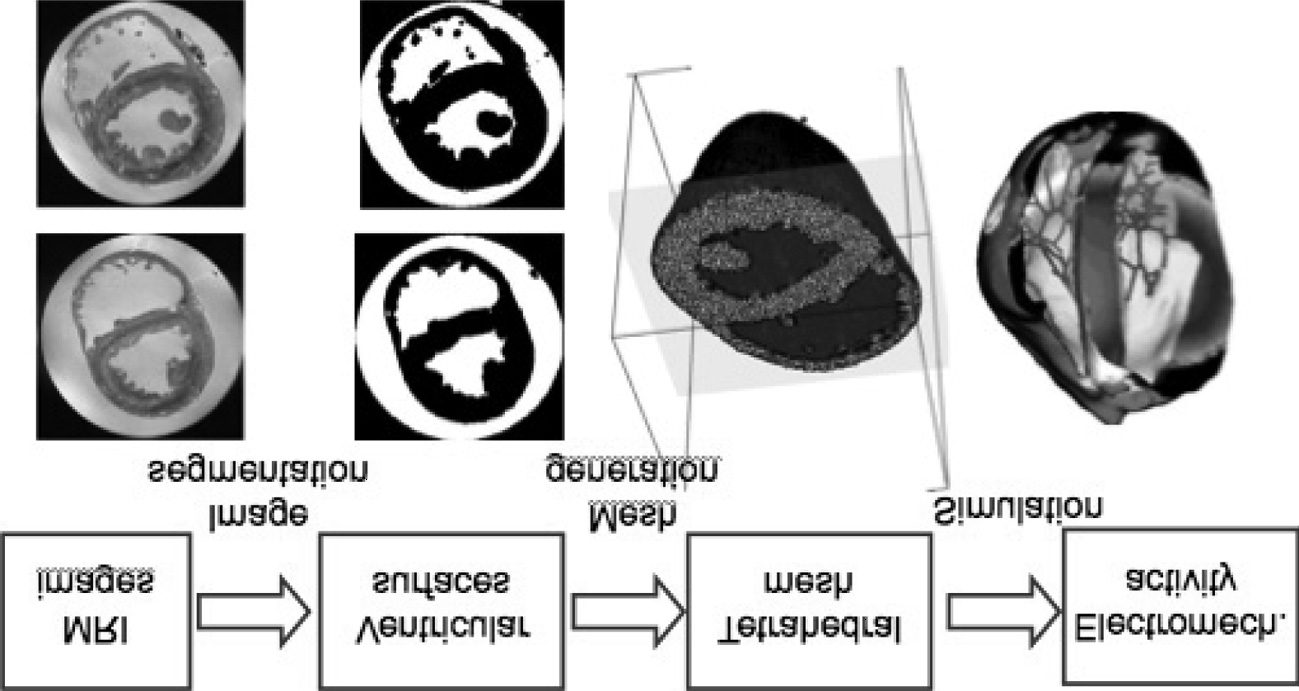
other research being devoted to it. The process of getting from MRI im-
siglos, or from histology to whole organ visualizations, is a research chal-
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
312
Computational Biology and the Limits of Shared Vision
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
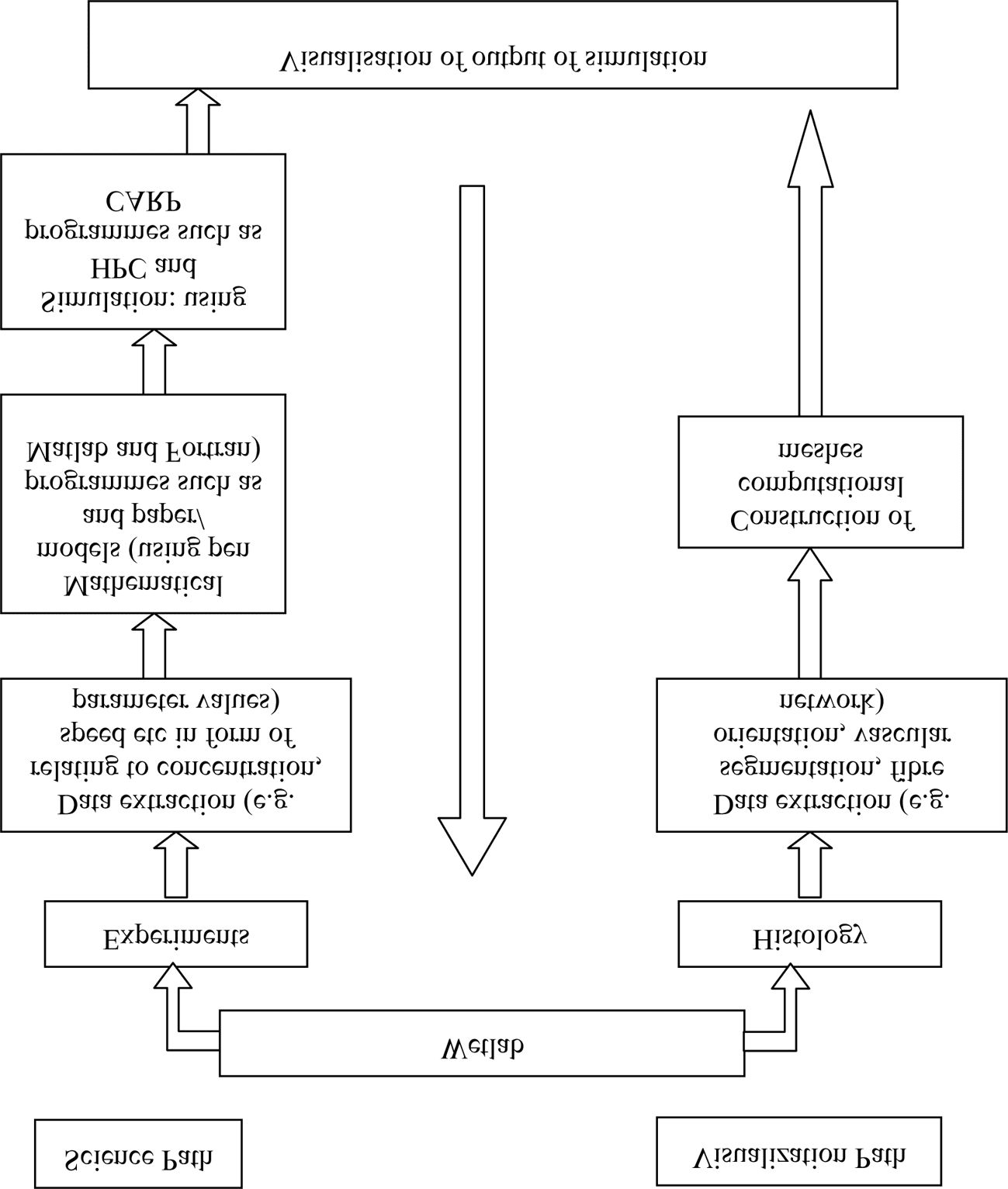
Cifra 1: Core stages of a simpliªed ‘bottom up’ workºows from wetlab to visu-
alisations and back to wetlab.
lenge in its own right (Burton et al. 2006; Plotkowiak et al. 2008), and is
demonstrated in Figure 2.
Some of the problems that emerge occur precisely around the issue of
parameter values. This is the output of the wetlab experimental process
and a crucial component of the computational biology enterprise. Para el
mathematicians and computer scientists in the project, it is essential that
Perspectives on Science
313
Cifra 2: From images to visualisations via Tetrahedral Meshes
the models they develop be parameterised with data derived from wetlab
experimentos. Data provided by experiments for the modelling process are
labelled with the term “parameter values”; sin embargo, the term can mean
different things for a mathematical computational biologist than it does
for a biologist or physiologist. A parameter value is a quantitative entity—
it can relate to temporal or spatial properties, or concentrations of mole-
cules, speeds, etcétera. Data in the form of parameter values are the only
way for models to “gain reality,” by which is meant for them to engage
with actual physical processes, and to have a chance of being tested and
possibly validated.
This is not so for other areas of biology of interest to mathematical biol-
ogia, such as cancer modelling. Several mathematicians in the study ex-
pressed frustration with biologists for not being willing to collaborate,
and commented on the difªculty of extracting parameter values from biol-
ogists. The difªculties relate to agreeing on which parameters may be of
interés, as well as the meaning of the values: Por ejemplo, whether they
are relative or absolute values. For a biologist, it may be sufªcient to know
that there is a relation between two (or more) propiedades, and they are in-
terested in the quantity of these only relative to each other. Matemático
biologists, en cambio, must know “absolute” quantities, concentrations, etc..
of these properties: eso es, what they are independently of each other.
Only with this knowledge, can they begin to pinpoint what they see as the
causal mechanisms of the processes they are attempting to understand.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
314
Computational Biology and the Limits of Shared Vision
As a computational mathematician put it:
[B]iologists often just want to know that something happens, two
proteins interact which regulate the expression of some other pro-
tein [. . .] now, that’s it, they just want to know that that’s the
mechanism. We want to know exactly how much of that protein
has to be present in order to cause exactly what degree of regulation
of this other protein . . . so we can build a model which explains
the concentration of the thing that caused the regulation of that
other thing [. . .] [por] a series of graphs [y un] model to reproduce
those graphs. Biologists don’t and can’t typically do that. They just
know that these proteins are present in the system. They just tell
you they’re there. No concentrations, rate constants, absolute con-
centrations in any particular situation. Biologists either can’t do
that or they won’t because it’s not what they’re interested in.
A very important aspect of this relationship is the fact that the research
questions of computational biologists are often geared towards interven-
tional strategies—for example, medical or pharmaceutical interventions.
Computational biology for the understanding of cardiac disease is closely
associated with pharmaceutical research. Por ejemplo, the mathematician
quoted above went on to say:
I want to be able to quantitatively explain what’s going on in terms
of molecules, forces, exactly what’s going on. And the reason I want
to explain that is I want to design new treatments and drugs and
only way to do that safely is to be able to say exactly what’s going
en.
But this involves testing for parameters that may be of little interest to
biologists—who are not medical, clinical, or pharmaceutical researchers.
This is also found in contexts where collaborations with biologists are
sought for engineering purposes, generating many questions about molec-
ular kinematics for the engineers in the collaboration, but for which “it re-
mains far from obvious in what sense they are biological questions” (Keller
2002, pag. 232). This is a crucial point of difference in the epistemic goals of
these disciplines. As a deeper exploration of the role of observation and vi-
sualization shows, it is also the point around which there are profoundly
different ontological commitments.
The broad domain of computational biology can be divided into two
categories: the ªrst category is where there are successful and entrenched
collaborations with buy-in from the various disciplines involved; the sec-
ond is where there are tentative and early stage collaborations, with ongo-
ing attempts to get collaborators on board. Within this second category
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
315
there are potential candidates for collaboration among biologists who have
not bought into the computational biology program, and who may be
sceptical and likely to resist it. For the sake of clarity, I shall label the ªrst
category “established CB,” the second category “beginning CB,” and the
sub-category “CB sceptics.”
The computational biology program of research is still relatively
young, and is still a program in need of validation and acceptance in main-
stream science. While it has been successful in garnering funding, con
several large funding programs internationally devoted to it,7 it has not
met with universal approval, and sometimes struggles to ªnd collabora-
tors within speciªc sub-domains. Sin embargo, because interdisciplinary col-
laboration is crucial for all aspects of the computational biology program,
a great deal of energy is invested into it. De este modo, the computational and
mathematical researchers, from the doctoral to professorial level, involved
in the cancer modelling programs (an example of beginning CB category)
are well aware of the need to cultivate potential partners in biology, y
spend a great deal of time in PR-type exercises. A number of strategies are
usado, including interpersonal meetings with potential partners, trabajar-
shops, and demonstrations geared towards biologists. Biologists are the
implicit audience of several journal articles by mathematical modellers de-
scribing what modelling can contribute to cancer research, and using sev-
eral persuasive means to get biologists on board. Por ejemplo, Byrne et al.
(2006) put forward an argument for the use of mathematical models in
studying tumour growth which appeals to the shared value of ªnding a
cure for cancer: “In this paper, we review a number of mathematical mod-
els that have been developed to describe some of the above aspects of tu-
mour growth. In so doing, we aim to show how mathematical modelling,
computation and analysis can generate useful insight into the mechanisms
that underpin this devastating disease” (pag. 1564).8
While important in terms of the overall strategy of computational biol-
ogia, these discursive strategies remain relatively external to the character-
istic practices of the computational and mathematical modellers. Visual
persuasive appeals are also used and to some extent the use of visualiza-
tions can be seen in a similar vein, as essentially rhetorical communica-
7. With projects such as the European funded Network of Excellence Virtual Physio-
logical Human. http://www.vph-noe.eu/ (accedido 30/05/2011).
8. Other articles along the same lines for mathematical modeling in cancer are Koma-
rova 2005, Van Leeuwen et al. 2007; others with greater emphasis on experimental physi-
ology are Kohl et al. 2000, Cazador & Borg 2003; for the most recent development of the
computational biology program towards simulating all organs of the human body: Fenner,
Brook et al. 2008.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
316
Computational Biology and the Limits of Shared Vision
ciones, trying to engage with the biologists’ highly visual culture, and try-
ing to share a common visual ground with the biologists.
In this section we have seen some of the context in which research in
computational biology is carried out, the need for interdisciplinary collab-
oration, and some of the obstacles to it stemming from the different histo-
ries of the different domains, and their different purposes and goals. En el
next section, we turn to visualizations as used within the modelling and
simulation enterprise.
Section 2b: Visualizations in the context of established CB
Computational biology in the domain of whole organ modelling is a good
example of the established CB category, with convergence sufªcient for a
collaborative interdisciplinary program of research. The collaboration be-
tween experimental physiologists and computational biologists in this
category is greatly facilitated by the fact that they share a common vocab-
ulary of parameter values: eso es, not only what they mean, but what val-
ues it is useful and desirable to test in an experiment. This convergence
around parameter values is made possible by the fact that they hold re-
search questions in common, and thus their research is driven by the same
interests and purposes—including, Por ejemplo, seeking collaborations
with industry partners such as pharmaceutical companies. The collabora-
tion is kept going through common research projects, dedicated doctoral
programas, co-supervision of doctoral students, and the increasing numbers
of interdisciplinary researchers. The willingness of experimentalists to
supply data for the simulations is a sign of the usefulness of the results of
the simulations for their own research questions; en efecto, both groups con-
verge on collaborations with external industry partners, in particular phar-
maceutical companies which are increasingly interested in using computa-
tional biology techniques for drug discovery and testing. There is
coordination, but not complete overlap between the different disciplines,
Por ejemplo, since there are still distinct roles for the different disciplines.
There is also much disagreement regarding, Por ejemplo, what a model is,
or even what computational biology as a ªeld is, and what exactly it is try-
ing to do. While there is some discussion, there is not disagreement to the
extent that it prevents the collaboration from occurring at all.
The key role of visualizations in the computational biology program of
research is evident in several ways: the visualizations are prominent in
teaching, in workshops, and in publications. The visualizations are also
cultural identiªers for computational biology groups, being used as logos,
ofªce artworks and the like. Visualizations are described—for example, por
their developers—as a powerful communicative tool in conveying com-
plex mathematical ideas in a visually compelling way. Sin embargo, the visu-
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
317
alizations are couched in a dual rhetoric: one which emphasizes their use
to communicate with experimentalists and get them on board; the other
which emphasizes their epistemic role. They are used for data analysis, como
tools of discovery and exploration of the processes under study; ellos son
used for evidence and justiªcation of claims. Visualizations are not an op-
tional add-on to modelling and simulation and they are not an illustration
of concepts and statements that are also couched in verbal terms. Bastante,
they are an integral part of the simulation process. As Winsberg puts it:
Visualization is by far the most effective means of identifying char-
acteristic features out of complex dynamical data sets, and so it is
the most, if not the only, effective means of judging the degree of
calibration a simulation enjoys with other data sets and with ana-
lytic results. De este modo, visualization plays a crucial role in sanctioning
as well as in analyzing simulation results. Not only does the episte-
mology of simulation call upon resources that are empirical, y
that come from outside of the theory, it also calls upon the faculties
of the observer. (1999, pag. 290)
This is a crucial point. The visualizations are qualitative renderings of a series
of quantitative processes. Their epistemic role, that is their use to validate the
whole process of modelling and simulation, is largely based on observation
of qualitative features. Not exclusively so, since evidently this is a mode of
qualitative observation impregnated by quantitative analysis.9 However, él
is an intriguing fact that a process that is so deeply quantitative ulti-
mately yields qualitative results. The question is what that visual render-
ing is ultimately of: eso es, a question regarding what is seen to be repre-
enviado, in terms of “content” and ontology.
Let us look at this in greater detail: Physiologists make use of various
imaging techniques, including microscopial images, MRIs, and fMRIs,
and make extensive use of images in their research. An image, sin embargo,
is a very different representation from a visualization, since it has an en-
tirely different causal history. Images are not without their algorithmic
and mathematical aspect, but they are deªned by the role in producing
them of light, sound, or other signals emitted from the object imaged.
What they are taken to represent is thus relative to the object with which
they stand in the appropriate causal relationship. Although visualizations
do start off from images, there are a number of intervening steps leading
to the construction of a computational mesh and ªnite element mesh
which then serves as the basic geometry for the visualized simulation, en
9. The fact that qualitative comparison with experimental results is used for validation
is also contested and held to be not optimal by some. See for example Parker 2008.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
318
Computational Biology and the Limits of Shared Vision
the form of an animated movie, which is at a very distant remove from any
images. The movie which is the visualization of the simulation, is itself a
rendering of the model in its predictive capacity of processes over time. En
the ªrst place, these visualizations play a role in bringing about a shared
mode of perception and set of visual practices with epistemic import—as
described in section 1, and argued for in the context of computational bi-
ology in a previous paper (Carusi 2008). Given that the mathematics is in
the ªrst place inaccessible and difªcult to comprehend by non-mathemati-
cal researchers, the visualizations may be the only direct access the
experimentalists have to the ideas and hypotheses being proposed for later
pruebas. In order to facilitate communication with experimentalists, el
style of the illustrations was initially derived from textbook illustrations
with which physiologists would have been familiar. They play a crucial
role in bridging the gap between those who speak the mathematical lan-
guage, and those who do not—or at least not as proªciently as the dedi-
cated mathematicians. For experimentalists, the context of the visualiza-
tions are their experiments, including instruments, laboratory techniques,
procesos, etcétera.
The computational biologists (mathematicians, engineers, physicists)
in the collaboration do, in principle, have an alternative point of access to
the simulation (the mathematical model itself). Sin embargo, as pointed out
arriba, even they rely on the visualization for grasping the outcomes of the
simulation, which is an initial testing of the model. Eso es, the simula-
tion has more in it than the mathematical model, and not everything can
be read off the model itself—else it would not need the simulation. Para
the computational biologists, the context of the visualizations is the
model and modelling process, parameterisation, the computation required
for the running of the simulation, etcétera. Set against these very differ-
ent contexts, the visualizations have different ranges of meaning for mod-
ellers and experimentalists. Despite this, the same visualizations are used
by both groups in the collaboration, building up a core of shared mean-
ings over the course of ongoing joint research projects. This is shown, para
ejemplo, in the numerous occasions when the visualizations are used
across disciplines. A sample of the resulting visualizations are shown in
Cifra 3.10
Sin embargo, even though this paradigmatic example of established CB ap-
10. The limitations of black and white publication do not allow a full appreciation of
these stills in their context, where for example they could be seen in a context of compari-
son with other visualizations and with the results of wetlab experiments. See for example
the online version of Rodriguez et al, 2005 en (cid:2)http://circres.ahajournals.org/cgi/content/
full/97/2/168(cid:3) [accedido 16/07/2010] for an excellent example of experiment/simulation
comparisons.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
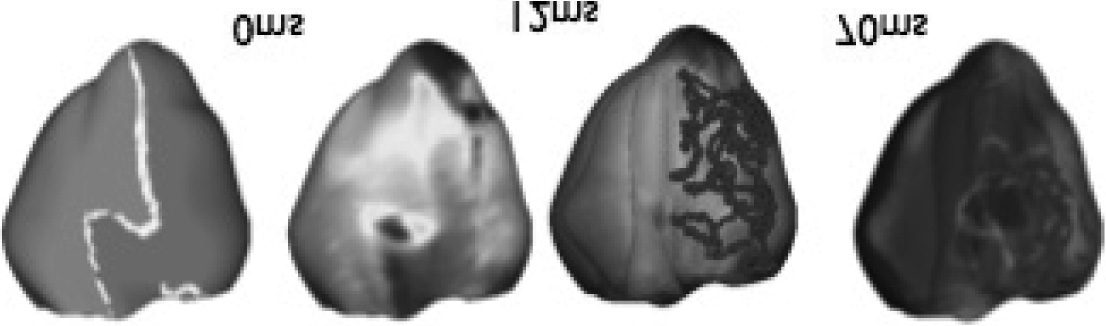
319
Cifra 3: A set of stills from a simulation of electrical activity in the heart.
pears to be one of the sharing of practices around the visualizations, es
not at all clear that they do so because the practices are socially shared, o
because of some other feature of the visualizations: such as what they are of
or about, their ontology, on which the social sharing of practices is predi-
cated. Let us look at the next category of computational biologists to see
whether any clarity can be gained by a consideration of cases where there
is a failure of shared vision.
Section 2c: Visualizations in the context of beginning CB
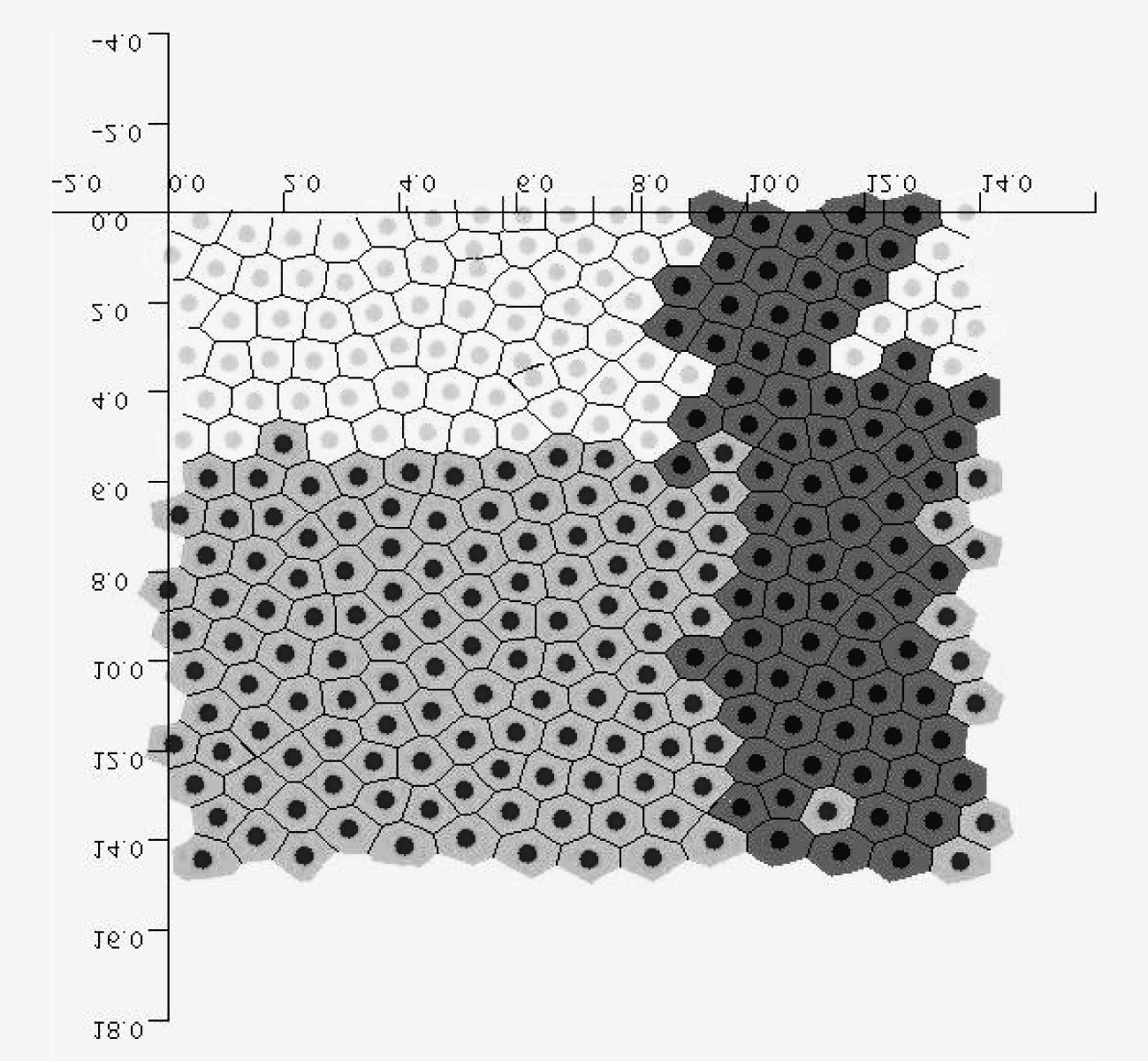
Images for the simulation of multicellular processes, such as tumour
growth, are far more problematic, for a variety of reasons, including the
fact that in these cases there is no certainty that the geometry of cells and
other entities involved in the processes is signiªcant (whereas it is clearly
signiªcant in the case of electrical currents across the heart). Sin embargo, en
beginning CB, there have also been attempts to engage with the observa-
tional preferences of biologists through visualizations, con resultados mixtos.
Visualizations such as that in Figure 4 are an example.11
Sin embargo, unlike the visualizations used for the simulations of heart
modelling, these visualizations were still relatively isolated from the rest
of the mathematical modelling process for the projects studied. Hay
a number of reasons for this: the science is at a less developed stage than,
Por ejemplo, cardiac modelling, and this results in visualizations which are
less complex, less intrinsic to the process of exploring evidence. Esto es
clear from the fact that in when it comes to cancer modelling, it is still
unclear whether the geometry of cells is a relevant factor, and this results
in the fact that the spatial properties of the visualization (borrowing from
the polygonal shape of epithelial cells) are not necessarily an essential as-
pect of the visualization for the purposes of rendering the processes at
work in the growth of cancer cells. Even from the point of view of scien-
tiªc workºows, the emphasis in these beginning CB projects is still on the
11. http://web.comlab.ox.ac.uk/chaste/cancer_index.html (accedido 30/05/2011).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
320
Computational Biology and the Limits of Shared Vision
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 4: A still from a visualization of the growth of cancer cells ‘Snapshot of
an intestinal crypt simulation. Here the crypt is modelled on a cylindrical geome-
intentar, by performing the simulation on a plane and enforcing periodicity on the left
and right edges. Transit cells are shown in yellow, differentiated cells in red, y
the blue cells are the progeny of a single cell that was dyed at the beginning of the
simulation.’ [Notice the adoption of terminology borrowed from microscopy: el
use of dyes for tracking cells].
matemáticas, cual, as an epistemic process, is not computational—in-
deed, only pen and paper may be necessary for working out the equations
of the model. Sin embargo, with the perceived need to communicate on the
biologists’ qualitative and observational terms in mind, the style for ªgure
4 was taken from the diagrams used in cell biology text books. Sin embargo,
in these textbooks, most often such diagrams are seen side by side with mic-
roscopial images,12 and those images, Sucesivamente, seen in the context of micro-
12. S. F. Gilbert’s Developmental Biology (2010) being an excellent example.
Perspectives on Science
321
scopial practice. The communicative and rhetorical roles of these visual-
izations are emphasized over their epistemic role. Thus the visualizations
are felt to be “for the biologists,” since they help to communicate the pro-
gram of research to the biologists who would otherwise not engage with
the mathematics. Sin embargo, visualizations of this sort are unlikely to ªnd
many appreciators in the biological community, and indeed this is clear by
the lack of take-up this type of visualization had received in other contexts
such as publications or presentations as compared with the cardiac visual-
izations. They failed to establish an “endogenous community” with so-
cially shared visual practices.
In the previous two sections we have seen that visualizations play a key
role in communicating the output of computational simulations. Ellos son
sometimes used in a way that cements the collaboration that makes the
whole modelling and simulation enterprise possible. Sin embargo, they some-
times fail to engage biologists and may well be resisted as representations
of biological processes. The next section establishes why this is the case by
consideration of the role of observation in cell biology.
Section 2d: Observations and images in the context of microscopy
and cell biology
The practices around any particular artifact need to be seen in the context
of other practices, instruments, and the broader conceptual context of the
discipline. There are several important differences between the epistemic
practices of biologists and mathematical biologists which have been ex-
tensively described by Evelyn Fox Keller in the book Making Sense of Life.
These differences involve having very different orientations to theory—
particularly theory couched in mathematical terms—, than to observa-
ción, and as has already been noted, to the goals of biology (Keller 2002).
The attitude expressed in the biologist Lynn Margulis’s ironic reference to
mathematical models as “numerology” (Carusi et al. 2009, pag. 32) is shared
by many who would not express it in quite this way.
From the point of view of biologists, computational biologists often
underestimate the complexity of biological processes—and indeed, de
what would be required to conduct the experiments required to build a
plausible model. Por ejemplo, one biologist interviewed commented on a
failed collaboration with a computational biologist:
[The computational biologist said ] we need to know the dissocia-
tion constant of this protein, reaction diffusion equations, concen-
trations of proteins in the cell. That sort of stuff I don’t have a clue,
it would take years to get. More generally, about systems biology as
a whole—how do you model networks, pathways, particular pro-
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
322
Computational Biology and the Limits of Shared Vision
cesses. Seems to me that you need to have these parameters. El
concentration of your protein, how fast does it move in the cell, es
it in the right location to reach with something downstream of it.
And there are really only a very few biological pathways for which
data like that are available.
One of the reasons why this type of data is unavailable is the difªculty of
abstraction in biological processes. Mathematicians sometimes refer to the
background complexity making abstraction difªcult as “noise,” whereas
for some biologists, this is precisely what makes them biological. Este
brings into play the question of modularity and which categories or “mod-
ules” are still distinctively biological in character. Por ejemplo, the self-
styled historical biologist, Lynn Margulis, warns against the “Whitehead-
ian fallacy of misplaced concreteness” (Carusi et al. 2009, pag. 27) and Brian
Goodwin, an acclaimed mathematical biologist, in his more recent work
turned towards a more holistic approach to biology, questioning the ab-
straction that necessarily goes hand in hand with mathematical models as
a general approach for understanding biological processes. In his view,
other non-mathematical terms play a signiªcant role in describing biolog-
ical phenomena:
brian: James [. . .] [y]ou used the terms “beauty,” “harmony,"
“grace.” Why not add “wholeness” and “health” and “coherence”?
Because you can see when microtubule organisation is coherent and
when it’s disorganised, and when it’s disorganised it’s a clue to
algo; some of the components are “funny.” Now, that can be
quite subtle and I would say that human intuition is bloody good
at this kind of detection. (pag. 18)
These disagreements are also expressed with respect to the visualizations.
Just because qualitative observation is such a key aspect of cell biologists’
práctica, it is speciªcally around attempts to engage them through obser-
vation that resistance is likely to be met. The failure to establish shared
practices around these artifacts stems from that to which visualizations as
opposed to microscopy are supposed to give visual access.
Microscopy is the key epistemic instrument for cell biologists’ explora-
tion and discovery of their speciªc research ªelds, and their observational
practices are formed in relation to the microscope to a large extent. El
microscope puts the biologist in what is experienced as direct contact with
tokens of that ªeld in the form of the samples being explored, even though
the experience is in fact mediated by the microscope and the theories con-
cerning the laws of physics which are built into it. This difference between
what is known about how a microscope works and using a microscope
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
323
came out clearly in several interviews, as well as in the Forum (Carusi et al.,
2009). If there is any instrument that biologists identify with, it is the
microscope—in the sense that they could not be biologists without a micro-
alcance. Por ejemplo, the quotation below is from a biologist who addressed
the question of scientiªc method in biology ªrst and foremost by describ-
ing the ritual of setting up a session with the microscope:
The afªnity of a cell biologist to his microscope shouldn’t be under-
estimated. A pesar de, I understand the physics of how a microscope
obras, it’s not the physics that interests me. It’s using the micro-
scope as an extension of my eyes in order to perceive the innermost
workings of a cell. In order to do that, I guess it becomes quite rit-
ualistic. So, I take samples, cells, tissues, and I prepare them in a
certain way. And then I’ll go to my little microscope room, little
dark room, pull the curtain, sit down on my seat, and then I’ll go
through a series of well-trod steps; I’ll take the cover off the micro-
alcance, I’ll put this little light on and that little light on, the x, y
stage. And then when I’m comfortable, when I’m feeling “in the
zone,” I’ll put the sample on there and I’ll take a very cursory
glance around what’s on that microscope slide. And instinctively I
know whether it’s a good sample or a bad sample, whether the ªxa-
tions worked properly, whether the stains worked properly, si
there are any differences between control and treated sample. (pag. 9)
A closer examination reveals that it is not simply the instrument as such;
bastante, it is the observational situation comprising person-microscope-
sample and the entire set of interactions around it that is so strongly
emblematic of what it is to be a biologist. And the reason for its being em-
blematic is the direct access to biological processes that the situation is ex-
perienced as affording. Por supuesto, this experience is not as direct, transpar-
ent, or unmediated as it appears. Vision is very closely connected with
possibilities of action: this is especially strongly brought out in Merleau-
Ponty’s philosophy of perception. Microscopy is no exception to this.
Throughout its history, the microscope has caught the imagination of nat-
ural history enthusiasts, promising a means for “exploring the inner laby-
rinths of nature” (Warner 1982) but it took some time for it to become an
instrument for serious scientiªc pursuit. The turning point occurred when
it became possible not only to look through a microscope passively, but to
interact with it and with the sample. As Evelyn Fox Keller shows, the pro-
fessional biological gaze through the microscope came into being when the
object could be no longer merely be looked at but also manipulated. Este
is especially important in establishing that what is seen through the mi-
croscope is real and not an artifact of the microscope. Keller writes:
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
324
Computational Biology and the Limits of Shared Vision
once the microscope was joined with the manual manipulations of
an experimental biology—marking, cutting and dissecting under
the scope—and the interdependency of hand and eye previously re-
served for the naked eye was extended into the microscopic realm,
the microscope became a reliable tool for veridical knowledge.
(1996, pag. 112)
What is considered to be real in both everyday and scientiªc contexts is
very closely connected to perceptions of causality. The interactions that are
possible with observed samples—isolating one part from others, marking
with dye, separating, extrayendo, injecting, etc.—and in all instances
watching and following with the eye what happens following each of these
interventions—creates a context in which causality can be experienced, en
the ªrst instance, in the very interactions themselves, including the mis-
takes. Ian Hacking’s description of learning to observe by active doing, es
vivid:
The conviction that a particular part of a cell is there as imaged is,
to say the least, reinforced when, using straightforward physical
medio, you microinject a ºuid into just that part of the cell. We see
the tiny glass needle—a tool that we have ourselves hand crafted
under the microscope—jerk through the cell wall. We see the lipid
oozing out of the end of the needle as we gently turn the microme-
ter screw on a large, thoroughly macroscopic plunger. Blast! Inept
as I am, I have just burst the cell wall, and must try again on an-
other specimen. (1983, páginas. 189–190)
As Keller points out, this “making real by touch” is the entry way to the
scientiªc realm of causal efªcacy, or “making things real by using them to
effect change in other things we know are real” (1996, pag. 114). This was
clear in interviews conducted with biologists who use microscopy as their
main instrument, as they stressed the relationship between what is seen
and the techniques used as relevant to a particular experimental hypothe-
ses. Por ejemplo, the use of fate-tracking (the use of dyes for particular
cells in order to track them through developmental processes) in embryol-
ogia, or the use of micro-scalpels and other instruments to transfer tissue,
and other similar techniques are all involved in the highly skilled hand-
eye coordination required for this form of microscopy.
What counts as real in these observations is a matter of the close inter-
relationship between observing through one kind of technology and ma-
nipulating what is observed, often by means of techniques using other
tools and technologies (micropipettes, microneedles, dyes,
labelling,
tracking, etc.). The process of vision is not static and inert, and that which
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
325
is observed is seen because it is manipulated, becomes visible through the
interactions with it.
Compare it for a moment to the visualization that renders the outcome
of a simulation. This too is a highly constructed viewing episode made
possible by a particular conªguration of viewers, instruments, and theo-
ries. The computational biologist is as interactive as the biologist with
their visualizations: Por ejemplo, they halt them, adjust parameters and
see what happens, alternate views, etc.. The visualizations used for compu-
tational biology also include social interactions, which is crucial for this
highly cooperative and collaborative research area. The visualizations as
material artifacts are fully incorporated (in the fullest sense of the word)
into research events such as seminars and workshops, where they are dis-
played, animated by gestures, and used as media for social interaction.
Physical interactions can also occur through the computational instru-
mentation that render the visualizations, or that analyse and process data:
the software as well as (or through) the screen, the mouse, and keyboard.
And of course, computational biologists experience their visualizations as
compelling too.
In this section we have seen that the mode of observations of both those
biologists who use microscopy as their main instrument, and those who
use visualizations of simulations are highly interactive, involving physical
interactions with that which is seen. In the next section we consider what
kind of exchanges there are between these two modes of seeing.
Section 2e: Exchanging and sharing modes of seeing
Observation is a key epistemic practice across the groups considered, be it
of visualizations of simulations, observations down a microscope, or obser-
vation of images: eso es, either “stills” from a simulation, or microscopial
images. What kinds of exchanges are most common between these? Nosotros
have seen that in the practice of computational biologists, comparisons are
carried out between the visualizations of simulations and experimental
resultados. Sin embargo, in the case of whole organ modelling they are carried
out using a range of different visual artifacts, ranging from 2D graphs to
the optical imaging of organs.13 The computational biologists on whom
this research was based admitted to not being able to see anything much
down a microscope, and seem to experience it as a passive mode of obser-
vación. Por ejemplo, interviewees claimed not to be able “to do anything
with it,” such as stop it so that you can look again, or change anything
while you’re looking. Este, por supuesto, does not resonate with the microsco-
pist’s experience at all, since their experience is of a highly interactive pro-
13. See for example Rodriguez et al. 2005.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
326
Computational Biology and the Limits of Shared Vision
impuesto, one which seems to involve their whole being and identiªcation as a
biologist. For a biologist committed to microscopy as a mode of observa-
ción, seeing a visualization of a simulation lacks the allure of observed pro-
cesses unfolding in the process of observation (the double-facedness of the
process of observed/observation is essential, and will be returned to in the
next section when we consider the ontology of what is seen). Thus when
mathematical biologists share with biologists visualizations such as the
one in ªgure 3 depicting the growth of cancer cells as mathematically
modelled, in a bid to share with them the qualitative practice with which
they are familiar, they are unlikely to ªnd a responsive audience. The visu-
alization simply looks uninteresting in the eyes of biologists who are used
to seeing these diagrams alongside microscopial images or in contexts
which relate to such images. Fundamentalmente, the micoscopial images in their
doblar, are only fully signiªcant when seen relative to the complex and
wholly embodied observations of processes often by means of powerful mi-
croscopes, and even more importantly, by means of a whole panoply of
techniques and tools which allow them to interact with the sample (Keller
1996).
Sin embargo, mathematicians too can exhibit a great deal of scepticism re-
garding the “visualizations” of biologists. Por ejemplo, biologists some-
times produce animations of biological processes, such as those produced
by Drew Berry,14 or by those to be found on the website of Hans Clevers’
Group at the Hubrecht Institute.15 Typically, these visualizations are pro-
duced by means of a combination of observation, microscopy images, mano
dibujo, and in depth research of current knowledge of the processes de-
picted. These movies are used mostly for communication and education,
and not for actual research. En tono rimbombante, their audience can include math-
ematical biologists (either in training or experienced researchers) en orden
to give them an idea of what the biological processes they are interested in
may “look like.” One view expressed was that it is very useful to be able to
see the movies since mathematicians working in biology do not have easy
access to this kind of process; the movies can help them to understand
their models better by putting them in context. Other views (sometimes
by the same people) were that the movies were also potentially mislead-
En g, in that they make it appear that all the problems have been solved,
that there is a complete understanding of the process. Sometimes these an-
imations are labelled as “just cartoons,” although even this view often goes
14. Walter and Eliza Hall Institute of Medical Research TV: http://www.wehi.edu.au/
education/wehi-tv/ (accedido 30/05/2011).
15. Hans Clevers Group, Hubrecht Institute, Movie Animation: http://www.hubrecht
.eu/research/clevers/research.html (accedido 30/05/2011).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
327
hand in hand with a distrust of what is compelling for the eye. A view
that is often expressed is that the movies may be misleading in that they
are seductive, and may persuade you to “buy into” a particular way of
seeing—and therefore understanding—the phenomena. One interviewee
likened this to seeing the movie adaptation of a book: the picture you had
in your mind may not be the same as in the movie and that may be jar-
ring; but equally, it may override the picture you have in your mind be-
cause it is so compelling. Es, in this regard, to be mistrusted.
The question arises as to how biologists themselves view these movies
and animations of biological processes. As has been stated, this type of an-
imation is not generally used for exploratory research; sin embargo, these ani-
mations can allow biologists to recognise, something which in principle,
they may be able to observe. Watching the movies can resonate with the
experience of observing the processes in question, and in this way, rein-
force the attachment that biologists can experience to their observational
práctica. This relationship might draw upon the historical relationship be-
tween microscopial observation and hand-drawn illustrations.
In any particular area of computational biology and the disciplines
afªliated to it either through active or potential collaboration, or simply
sharing the broad biological domain with it, there will be visual artifacts
around which exchange occurs and those instead around which exchange
does not occur. No single case study can grasp all of the variations possi-
ble. Sin embargo, in this article I have wanted to show one particular area
around which exchange seems to be strongly resisted, and that is in the
observational process of microscopy and its related visual artifacts on one
mano, and the visualizations of simulations on the other. In this area, allá
are socially shared visual practices in the groupings around established
CB, but this is very patchy and superªcial in beginning CB, and does not
occur among the CB sceptics, who resist seeing as computational biolo-
gists do. To extend the analogy from Goodwin’s paper on professional vi-
sión, we might say that the established CB group display a similar sharing
of practices evident in the case of archaeological instruction (though they
are not quite the same since the different disciplines do keep their own
practices as well); beginning CB might be likened to the groups consti-
tuted by lawyers and members of the jury who were convinced by their
demonstrations of visual practices around video evidence (if there were
cualquier); and the CB sceptics can be likened to those members of the jury who
remained unmoved by these demonstrations. The question is what makes
the difference between these groupings? It is not a question that can be
answered purely in the terms of socially shared practices, since the ques-
tion is why do some visual practices come to be shared whereas others do
no?
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
328
Computational Biology and the Limits of Shared Vision
In the observational situations of biology and computational biology
there are several structural features in common: a high degree of what we
can call “fusion” between observers and instruments directed at processes
to be made sense of. The instruments combine those which render a visual
artifact to the gaze, and those which enable interaction and manipulation.
None of these elements can be isolated from the others in the observation
of the process in question: there is nothing to be seen without the inter-
vention of viewers and instruments. The englobing context is that of the
broader epistemic and institutional context in which the entire conªg-
uration of viewers, instruments, and processes occurs, and from which it
gains meaning. There is no innocent eye, there is no neutral instrument,
there is no simply given object. There is, sin embargo, a fundamental differ-
ence in the ontology of what is seen. This is explored in the last section of
this paper.
Sección 3 Discusión
Gaps, ªssures, and breakdowns in sharing visual practices show the limits
of what can be accomplished by the social sharing of these practices alone,
and show that sharing, where it does occur, is predicated upon a much
wider set of factors. Faltering or even failing inter-disciplinary collabora-
tions where there are competing views on the same ªeld are good places to
examine these limits. The question “when can practices be shared?” is
inter-related with other questions such as: who does the observing, con
what activities, técnicas, and instruments, in what contexts, and for
what purposes? All of these converge on the question: “what is seen or ob-
served?” The observations of dry lab experiment and wet lab experiment
are undoubtedly both constructions, but this still leaves plenty of scope
for real differences regarding the epistemology and ontology of the broad
ªeld of biology. Whether progress is made in the more intractable sub-
domains of computational biology depends on whether the translation can
be made between models and simulations on one hand and the type of ex-
perimental data obtained through the speciªc techniques characteristic
of that sub-domain on the other. Visual practices—including practices of
observation—are a central aspect of these techniques, since they are often
one of the primary means through which evidence is constituted within
the sub-domain. Evidence interweaves interactions between people, en-
struments, and the entities observed, be they simulated or “real,” creating
a web that is at the same time social, instrumentational, and ontological.
When broadened out in this way, it begins to become more apparent why
visual practices cannot be shared across some domains without a concomi-
tant change or even sacriªce of the very thing that is felt to deªne that do-
main of inquiry: the thing that is studied, about which researchers want to
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
329
know and understand more, towards which their inquiry is directed.
There is a personal and subjective level in that what is studied and how it
is studied deªnes not only the domain of inquiry, but also oneself as scien-
tist in relation to it. It is this, tal vez, that partly accounts for the resis-
tance to sharing visual practices: not simply because they are different
ways of doing, but because they change the domain of inquiry, and in so
doing, also potentially change the researcher, or make a demand for
change at a very deep level of the way that researchers experience their
own identity in a domain. Sin embargo, researchers do change disciplines, entonces
this must remain a partial answer.
Between the biological sciences considered in this paper, a central dif-
ference in the nature of the visualizations, images, and microscopial
observations is the ontology of that towards which the scientists’ visual
practices are directed. No doubt in both cases, vision is not of a “pure”
object—whatever that might be—but involves constructs of practices, en-
struments, and theories. De hecho, the visual objects—that which is seen in
and through the instrumentation of visualization and of microscopy—are
both models, since the samples observed through microscopy are generally
of animal models, such as drosophila, sea urchins, or worms. But the way
in which they are constructed, the history that gave rise to them, y, im-
portantly, the role of the biological object in them, is substantially differ-
ent. In the process of modelling and simulation, the biological object is
externo, at least to the observation of the visualization that renders the
proceso. What is observed in the visualization is the simulated model; él
funciones, like a concretisation of an otherwise abstract entity. It is not
dissimilar to Kant’s proposal that metaphors and other images “body
forth” or make concrete an abstract idea, y son, for that very reason, un
object of aesthetic pleasure—and hence, to continue in an un-Kantian
way—of attachment and identiªcation.16 The biological object or process
which is modelled acts as an external constraint; it is an external validator
or data provider, not that which is seen or embodied in the visualization.
What is compelling in the visualizations is the perceptual form of a math-
ematical idea.
What is observed through the microscope is instead a biological ob-
ject—not an unadulterated pure biological object, but biological nonethe-
menos, in the form of cells and components of cells, tissues, and organisms.
The biological is not external to the process of observation. Bastante, el
16. In the production of aesthetic ideas, genius “attempts with the aid of an imagina-
tion which emulates the display of reason in its attainment of a maximum, to body forth to
sense [rational ideas] with a completeness of which nature affords no parallel” (Kant 1952,
pag. 176–7). Aesthetic ideas serve “as a substitute for logical presentation” for rational ideas
(1952:177).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
330
Computational Biology and the Limits of Shared Vision
unfolding of the processes observed are closely related to the process of ob-
servicio, through the interactions between observer, observado, and in-
strument that occurs throughout the manipulations through which the
observed becomes visible. The blurring of the boundaries between viewer
and viewed is captured very well by the exchange between Barbara
McClintock and Marcus Rhoade reported in Keller’s biography: To his re-
mark that he marvels at what she is able to see when she looks at a cell
down a microscope, she responded: “Well you know, when I look at a cell I
get down in that cell and look around” (2003, pag. 69).
In both cases, what is seen, what the visual experience is of, is that
which emerges through an ongoing series of interactions in which the ob-
server and the observed are both participants, in the sense that the shape of
the interaction comes from both. So does the ontology of the interaction
come from both: what it is of or about, biological process or mathematical
idea, emanates from the whole interaction and not from one side consti-
tuting or constructing the other.
In the Phenomenology of Perception (1962) in a section dealing with “The
Thing and the Natural World,” Merleau-Ponty put forward a framework
for thinking of the relation between perceivers and “the thing in the natu-
ral world” as one of inter-constitution. Merleau-Ponty wrote of the relation
between perceiver and perceived as one of communication and dialogue,
where each completes the other. In this relationship, the objects of science
are far from being “docile objects” (Linchar 1985, pag. 43). Natural objects
become objects for us through a certain organisation and arrangement of
their sensible aspects (ellos tienen, as Merleau-Ponty put it in Visible and In-
visible (1968) an opening onto subjects, as subjects have to objects, in a re-
versible relation of intertwinement), but at the same time we cannot ever
know them entirely—they can always surprise us. Even more, Podríamos
say that scientiªc vision offers an experience like Klee’s, of objects that
“look back” and with which the eye and hand are interwoven: “The eye
[. . .] is that which has been moved by some impact of the world, which it
then restores to the visible through the traces of a hand” (1993, pag. 127). En
this relationship with natural objects, the moves we make in the dialogue
are completed by the object and vice versa.
[mi]very perception is a communication or communion, the taking
up or completion by us of some extraneous intention or, sobre el
other hand, the complete expression outside ourselves of our per-
ceptual powers and a coition, por así decirlo, of body with things.
(Merleau-Ponty1962, pag. 320)
Perceiving things is in a sense to live them, to have them lodged within
oneself in one’s orientation towards the world:
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
331
In order to perceive things, we need to live them . . . To “live” a
thing is not to coincide with it . . . Our problem, por lo tanto, ser-
comes clear. The perceiving subject must, without relinquishing his
place and his point of view, and in the opacity of sensation, reach
out towards things to which he has, in advance, no key, and for
which he nevertheless carries within himself the project, and open
himself to an absolute Other which he is making ready in the
depths of his being. (pag. 325–6)17
For Merleau-Ponty, the freedom of the subject (or even of socially grouped
subjects) to make their world is not inªnite. Herein lies his disagreement
with Sartre for whom freedom is absolute. For Merleau-Ponty this
amounts to a blindness or to a seeing only of the self, which is the same
thing. He uses the metaphor of the circuit in his further eroding of the
subject-object distinction in the Notes on Nature. The two hands, tocar-
ing and touched, are in a circuit: if one were to trace that circuit, uno
would not be able to tell where one leaves behind the touching and enters
the touched. The relation between them is furthermore one of immanence:
the touching and the touched are touching and touched in relation to one
otro. Similarly with vision: seers and seen are in a circuit. One would
want to say, in science, but also many other domains, that it is a circuit en-
crusted with technologies (a point that also needs much elaboration). El
point to emphasize though, is that these circuits of observer and observed
demarcate profoundly different ontologies. Visualized mathematical-com-
putational ideas are an entirely different ontological species from observed
biological entities in virtue of the scope of observers by whose interactions
(comportamiento, proactions, reactions) they have been coaxed into visibility, y
by the constraints and possibilities of visibility and interaction for which
they allow. There can be no meaningful sharing of visual practices across
fundamentally different ontologies of vision. Bastante, practices are shared
when ontologies of vision themselves become inter-related: when experi-
17. The details of the implications of this account—intended for basic experience and
not for science as such—cannot be broached here (See Rouse 2005). Of course it makes a
difference that for Merleau-Ponty, science is a “second-order expression” relative to a “basic
experience of the world,” although “second-order” here does not mean that it is any less in-
tensely lived. On the contrary, if anything, we might suppose that scientists live the rela-
tion to their objects and artifacts more intensely than basic experience, and that the experi-
ence of being in a dialogue with these, of interrogating them with ones eyes and other
senses, and through one’s instruments more is keenly felt. At any rate, there is no reason to
think that the ontology of these domains is a diminished version of the ontology of basic
experiencia. There is also a particular signiªcance to the biological sciences, as is evident in
Merleau-Ponty’s elaboration of his ontology of the ºesh in his thoughts on the life sciences,
in Nature (1995). See also Hansen 2005.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
332
Computational Biology and the Limits of Shared Vision
mental practices yielding observational and image data and computational
visualizations of simulations are oriented one towards the other. In these
casos, the visibility of the experimental is interdependent with the compu-
tational, since experimental visibility answers to the computational mode
of probing, and the computational visualization models itself upon the ex-
perimental mode of seeing. They come to be in a circuit of visibility them-
selves. But the forging of these circuits does not come without ontological
costo, including a cost to observers in giving up a certain relationship to an
object of vision, to a certain way of being for themselves as much as for
that upon which they gaze. This must not be underestimated. It is not
just that a particular perspective on an object is lost or gained, it is that
the object—and with it, the entire mesh of inter-relationships through
which it is manifested, including the being or identiªcation of the ob-
server—is lost or gained.
Conclusión
Within the bounds of this paper, it is not possible to delve further into the
ontology of computational simulations and visualizations on one hand and
of microscopial observations on the other. A fuller account is necessary. Mi
aim has been to show that the sociality of visual practices—the fact of
their being shared by communities—is not sufªcient to account for what
is seen through those practices, and that this is particularly evident when
there is a resistance to come to see as others do. Merleau-Ponty offers, para
visión, and potentially for the whole range of sensory openings onto our
entornos, an ontology of inter-constitution of parts, where each par-
ticipates in the being of the other, or in the becoming phenomenon of the
otro. Drawing upon this theory, an alternative framework has been of-
fered according to which scientiªc viewers and scientiªc objects inter-
constitute each other and in this inter-constitution, deªne a mode of vi-
sion with a speciªc ontology, and a speciªc becoming of seer and seen. El
ontology identiªes at one stroke the scientist qua scientist and that which
they observe (what it is to be a biologist of a certain kind is to observe in a
certain way things of a certain kind). It is only through a framework, como
Merleau-Ponty’s, which allows for this more complex comprehension of
the ontologies of vision that we can begin to grasp the complexities of cur-
rent scientiªc visual practices. It is also a framework which places con-
straints on how much it is possible to tell about the so-called constitution
of objects from manifested social practices among scientists or other com-
munities. What is needed are new methodologies for probing the circuits
in which ontologies become manifest.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
333
Referencias
Baigrie, Brian S. 1996. Picturing Knowledge. Historical and Philosophical
Problems Concerning the Use of Art in Science. toronto: University of To-
ronto Press.
Barad, karen 2003. “Posthumanist Performativity: Toward an Under-
standing of How Matter Comes to Matter.” Signs: Journal of Women in
Culture and Society 28: 801–31.
Brenner S, D. Noble, t. Sejnowski, R. D. Campos, S. Laughlin, METRO.
Berridge, and K. Prank, l. Segel, R. mi. Dolmetsch. 2001. “Under-
standing Complex Systems: Top-Down, Bottom-up or Middle-Out?"
Pp. 150–59 in Novartis Foundation Symposium: Complexity in Biological
Information Processing, volumen. 239. Edited by G. R. Bock and J. A. Goode.
Chichester: John Wiley.
Burton, R. A., GRAMO. Plank, j. mi. Schneider, V. Grau, h. Ahammer, S. l.
Keeling, j. Sotavento, norte. PAG. Herrero, D. Gavaghan, norte. Trayanova, PAG. Kohl.
2006. “Three-Dimensional Models of Individual Cardiac Histoanat-
omy: Tools and Challenges.” Annals of the New York Academy of Sciences
1080: 301–19.
Byrne, h. METRO., t. Alarcon, METRO. R. Owen, S. D. Webb, y P. k. Maini.
2006. “Modeling Aspects of Cancer Dynamics: A Review.” Philosophical
Transactions of the Royal Society 364: 1563–78.
Carusi, Annamaria 2008. “Scientiªc Visualisations and Aesthetic Grounds
for Trust.” Ethics and Information Technology 10: 243–54.
Carusi, Annamaria, Blanca Rodriguez, James Wakeªeld, Kevin Burrage,
Evelyn Fox Keller, Brian Goodwin, Philip Maini, Lynn Margulis,
Denis Noble, Derek Terrar, Anne Trefethen, Eric Werner. 2009. Fo-
rum for Scientiªc Method in Biology: Transcript. Universidad de Oxford
e-Research Centre. http://ora.ouls.ox.ac.uk/objects/uuid%3A938f44c7-
e0c4–4b8f-a4c3-d2fac38f9044
Fenner, j. w., B. Brook, GRAMO. Clapworthyet al. 2008. “The Europhysiome,
Step and a Roadmap for the Virtual Physiological Human.” Philosophi-
cal Transactions of the Royal Society 366: 2979–99.
Galison, Peter 1997. Image and Logic: A Material Culture of Microphysics.
chicago & Londres: University of Chicago Press.
Gavaghan, D., A. Garny., PAG. Maini, y P. Kohl. 2006. “Mathematical
Models in Physiology.” Philosophical Transactions of the Royal Society 364:
1088–106.
Gilbert, Scott F. 2010. Developmental Biology, Ninth Edition. Sunderland:
Sinauer Associates Inc.
Goodwin, Charles. 1994. “Professional Vision.” American Anthropologist
96:606–33.
Goodwin, Charles. 1997. “The Blackness of Black: Color Categories and
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
334
Computational Biology and the Limits of Shared Vision
Situated Practice.” Pp. 111–142 in Discourse, Tools and Reasoning: Essays
on Situated Cognition Edited by L. B. Resnick, R. Saljö, C. Pontecorvo,
y B. Burge. Berlina, Heidelberg & Nueva York: Saltador.
Hacking, Ian. 1983. Representing and Intervening. Cambridge: Cambridge
Prensa universitaria.
Hansen, Mark B. norte. 2005. “The Embryology of the (en)Visible.” Pp. 231–
65 in Cambridge Companion to Merleau-Ponty. Edited by T. Carmen
Hansen, M.B.N. Cambridge: Prensa de la Universidad de Cambridge.
Horder, Tim. 2008. “A History of Evo-Devo in Britain: Theoretical Ideals
Confront Biological Complexity.” Annals of the History and Philosophy of
Ciencia 13: 101–74.
Cazador, Peter J. and Thomas K. Borg. 2003. “Integration from Proteins
to Organs: The Physiome Project.” Nature Reviews, 4 Marzo 2003:
237–423.
Husserl, Edmund. 1970. The Crisis of the European Sciences and Transcenden-
tal Phenomenology. Including the Origin of Geometry, translated by David
Carr. Evanston: Northwestern University Press.
jones, Caroline. A. and Peter Galison. 1998. Picturing Science, Producing
Arte. Nueva York & Londres: Routdlege.
Kant, I. 1952. Critique of Judgement, translated by J. C. Meredith. Oxford:
Letras gruesas a la media.
Keller, Evelyn Fox. 1996. “The Biological Gaze.” Pp. 107–21 in Future
Natura: Naturaleza, Ciencia, Cultura. Edited by G. Robertson, METRO. Mash, l.
Tickner, j. Bird, B. Curtis, and T. Putnam. Londres: Routledge.
———. 2002. Making Sense of Life: Explaining Biological Development with
Modelos, Metaphors and Machines. Cambridge, Mass. & Londres, Inglaterra:
Prensa de la Universidad de Harvard.
———. 2003. A Feeling for the Organism. Nueva York: Henry Holt and Co.
Kohl, Peter, Dennis Noble, Raimond L. Winslow, Peter J. Cazador. 2000.
“Computational Modeling of Biological Systems: Tools and Visions.”
Philosophical Transactions of the Royal Society 358: 579–610.
Komarova, Natalia L. 2005. “Mathematical Modelling of Tumorigenesis:
Mission Possible.” Current Opinion in Oncology, 17: 39–43.
Krohs, Ulrich. and Werner Callebaut. 2007. “Data without Models
Merging with Models without Data.” Pp. 181–214 in Philosophi-
cal Foundations of Systems Biology. Edited by F. Boogerd, F. j. Brugge-
hombre, J-H S. Hofmeyr, and H. V. Westerhoff. Ámsterdam & Oxford:
Elsevier.
Latour, Bruno. 1986. “Visualization and Cognition: Thinking with Eyes
and Hands.” Knowledge and Society 6: 1–40.
Linchar, Miguel. 1985. “Discipline and the Material Form of Images: Un
Analysis of Scientiªc Visibility.” Social Studies of Science 15: 37–66.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Perspectives on Science
335
———. 1990. “The Externalized Retina: Selection and Mathematization
in the Visual Documentation of Objects in the Life Sciences.” Pp. 153–
86 in Representation in Scientiªc Practice. Edited by M. Woolgar and
METRO. Linchar. Cambridge MA; Londres: CON prensa
Mascord, Matthew, Marina Jirotka, and Annamaria Carusi . 2007. Integra-
tive Biology Vre. Digital Pen Evaluation Report.
Merleau-Ponty, Maurice. 1968. Visible and Invisible,
A. Lingis. Evanston: Northwestern University Press.
translated by
———. 1993. “Eye and Mind.” Pp. 121–49 in The Merleau-Ponty Aesthet-
ics Reader: Philosophy and Painting. Edited by G. A. Johnson. Evanston
Northwestern University Press
———. 1962. Phenomenology of Perception, translated by C. Herrero. Londres:
Routledge & Kegan Paul.
Merleau-Ponty, Maurice and Dominique Séglard. 1995. Naturaleza. Course
Notes from the College De France, translated by Robert Vallier. Evanston:
Northwestern University Press.
Noble, Denis. 1962. “A Modiªcation of the Hodgkin-Huxley Equations
Applicable to Purkinje Fibre Action and Pacemaker Potentials.” Jour-
nal Physiology 160: 317–52.
parker, Wendy S. 2008. “Computer Simulation through an Error-
Statistical Lens.” Synthese 163: 371–84.
Plotkowiak, METRO., B. Rodriguez, GRAMO. Plank, j. Schneider, D. Gavaghan,
PAG. Kohl, and V. Grau. 2008. “High Performance Computer Simulations
of Cardiac Electrical Function Based on High Resolution Mri Data-
sets.” Computational Science—ICCS 2008 5101: 571–80.
Rodríguez, B., l. li, j. mi. Eason, I. R. Eªmov, y N. Trayanova. 2005.
“Differences between Left and Right Chambers Geometry Affect: Car-
diac Vulnerability to Electric Shock.” Circulation Research 97: 168–75.
Rouse, Joseph. 2005. “Merleau-Ponty’s Existentialist Conception of Sci-
ence.” Pp. 265–91 in Cambridge Companion to Merleau-Ponty. Edited by
t. Carmen Hansen, M.B.N Cambridge: Prensa de la Universidad de Cambridge.
Schutz, Alfred and Thomas Luckmann. 1973. The Structures of the Life-
Mundo, translated by R. METRO. Zaner and H. t. Engelhardt. Londres:
Heinemann.
Sharrock, Wes and Jeff Coulter. 1998. “On What We Can See.” Theory and
Psicología 8: 147–64.
Van Leeuwen, Ingeborg M. METRO., Carina M. Edwards, Mohammad Ilyas,
Helen M. Byrne. 2007. “Towards a Multiscale Model of Colorectal
Cancer.” World Journal of Gastroenterology 13: 1399–407.
Warner, John Harley. 1982. “Exploring the Inner Labyrinths of Creation:
Popular Microscopy in Nineteenth-Century America.” Journal of the
History of Medicine and Allied Sciences 37: 7–33.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
336
Computational Biology and the Limits of Shared Vision
galés, Elaine, Marina Jirotka, and David Gavaghan. 2006. “Post-
Genomic Science: Cross-Disciplinary and Large-Scale Collaborative Re-
search and Its Organizational and Technological Challenges for the
Scientiªc Research Process.” Philosophical Transactions of the Royal Society
364: 1533–49.
Winsberg, eric. 1999. “Sanctioning Models: The Epistemology of Simula-
tion.” Science in Context 12: 275–92.
Wittgenstein, Ludwig. 1953. Philosophical Investigations, translated by
GRAMO. mi. METRO. Anscombe. Oxford: Blackwell.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
pag
oh
s
C
/
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
1
9
3
3
0
0
1
7
8
9
7
0
8
pag
oh
s
C
_
a
_
0
0
0
4
4
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3