ARTÍCULO DE INVESTIGACIÓN
Brain Areas Critical for Picture Naming: A
Systematic Review and Meta-Analysis of
Lesion-Symptom Mapping Studies
un acceso abierto
diario
Vitória Piai1,2
and Dilys Eikelboom3
1Radboud University, Donders Centre for Cognition, Nimega, Países Bajos
2Radboudumc, Donders Centre for Medical Neuroscience, Department of Medical Psychology, Nimega, Países Bajos
3Max Planck Institute for Psycholinguistics, Nimega, Países Bajos
Palabras clave: confrontation naming, lexical semantics, object naming, oral naming, word finding
ABSTRACTO
Lesion-symptom mapping (LSM) studies have revealed brain areas critical for naming,
typically finding significant associations between damage to left temporal, inferior parietal,
and inferior fontal regions and impoverished naming performance. Sin embargo, specific
subregions found in the available literature vary. Por eso, the aim of this study was to perform a
systematic review and meta-analysis of published lesion-based findings, obtained from studies
with unique cohorts investigating brain areas critical for accuracy in naming in stroke patients
al menos 1 month post-onset. An anatomic likelihood estimation (PERO) meta-analysis of these
LSM studies was performed. Ten papers entered the ALE meta-analysis, with similar lesion
coverage over left temporal and left inferior frontal areas. This small number is a major
limitation of the present study. Clusters were found in left anterior temporal lobe, posterior
temporal lobe extending into inferior parietal areas, in line with the arcuate fasciculus, and in
pre- and postcentral gyri and middle frontal gyrus. No clusters were found in left inferior
frontal gyrus. These results were further substantiated by examining five naming studies that
investigated performance beyond global accuracy, corroborating the ALE meta-analysis
resultados. The present review and meta-analysis highlight the involvement of left temporal and
inferior parietal cortices in naming, and of mid to posterior portions of the temporal lobe in
particular in conceptual-lexical retrieval for speaking.
INTRODUCCIÓN
According to psycholinguistic models of language production, a speaker starts with a concept
they want to express and goes through several stages until their intention can be articulated.
Generally speaking, these stages can be seen as conceptual preparation, lexical selection (es decir.,
an operation at the level of “lemmas,” a semantic-syntactic representation), phonological
retrieval and encoding (es decir., the retrieval and ordering of the speech sounds associated with
that lemma), phonetic encoding (es decir., the computation of the gestural score), and articulation
(p.ej., Dell, 1986; Dell & O’Seaghdha, 1992; Levelt et al., 1999).
Producing language involves an extensive network of brain areas. Neurolinguistic models
of language production have attempted to link the proposed cognitive stages to different brain
areas (Hickok & Poeppel, 2007; Indefrey & Levelt, 2004; Roelofs, 2014). Various methods
have been used to uncover these neural substrates, Por ejemplo, by combining word
Citación: Piai, v., & Eikelboom, D.
(2023). Brain areas critical for picture
naming: A systematic review and meta-
analysis of lesion-symptom mapping
estudios. Neurobiology of Language,
4(2), 280–296. https://doi.org/10.1162
/nol_a_00097
DOI:
https://doi.org/10.1162/nol_a_00097
Supporting Information:
https://doi.org/10.1162/nol_a_00097
Recibió: 13 Puede 2022
Aceptado: 16 December 2022
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Vitória Piai
vitoria.piai@donders.ru.nl
Editor de manejo:
Stephen M. wilson
Derechos de autor: © 2023
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
production tasks with functional neuroimaging (p.ej., Price et al., 1996), electrophysiology
(p.ej., Liljeström et al., 2009), or neurostimulation techniques (p.ej., Hernandez-Pavon et al.,
2014). En general, these methods have highlighted the correlation between activity in a broad
fronto-temporo-parietal network and word production.
A different, noncorrelational approach to studying language production consists of examining
the consequences of tissue damage on performance, as done with lesion-symptom mapping
(LSM) técnicas. Although differing in the particular methodology, these techniques aim to
map the relationship between lesions and behavior, generating statistical maps. Rather than
requiring a cut-off score or binary data (p.ej., behavioral performance is either deficient or
no), a continuous behavioral score can be used, thus enabling a more sensitive approach. Fur-
thermore, these techniques can serve as a whole-brain analysis, rather than studying particular
regions-of-interest (ROI), although they are inherently limited by lesions coverage. In one par-
ticular and much used approach, voxel-based lesion-symptom mapping (VLSM), a statistical test
is run at every voxel, comparing a behavioral score between patients with and without a lesion
in that voxel, thus identifying voxels critical for the measured behavioral performance (Bates
et al., 2003). Besides VLSM, similar (and more recent) lesion-symptom mapping approaches
existir, such as voxel-based morphometry (Ashburner & Friston, 2000), voxel-based correlational
methodology (VBCM; Tyler et al., 2005), and multivariate methods (reviewed in Ivanova et al.,
2021), such as support vector regression multivariate LSM (Zhang et al., 2014). We note that a
discussion of how these methods work is beyond the scope of the present study (see for expla-
nation and comparisons between these methods, p.ej., Geva et al., 2012; Ivanova et al., 2021).
Baldo et al. (2013) applied VLSM on naming accuracy in chronic stroke patients, mientras
controlling for overall fluency in speech production and visual recognition of the items in
the naming test. Significant brain regions were predominantly found in the left mid and pos-
terior portions of the middle temporal gyrus (MTG), suggesting that naming critically depends
on this area and the adjacent white matter. A different study by Thye and Mirman (2018) también
found that damage to the left MTG was associated with deficits in naming in stroke patients, en
addition to areas in the left inferior frontal gyrus (IFG), supramarginal gyrus (SMG), and angular
gyrus (AG). Similar LSM studies confirmed the involvement of these areas but also found other
or additional areas associated with naming performance, such as the left postcentral gyrus,
inferior temporal gyrus (ITG), inferior longitudinal fasciculus, and temporal pole (p.ej., Alyahya
et al., 2018b; Faroqi-Shah et al., 2014; Piras & Marangolo, 2007).
Present Study
En suma, damage to left temporal, inferior parietal (AG and SMG) and inferior frontal areas is
typically associated with deficits in naming, though specific subregions found in the literature
vary. An informal attempt to summarize available LSM evidence may be complicated by the fact
that comparability is limited when different studies implement different experimental designs.
Studies may for example vary in employed LSM approach, task demand, and covariates used.
Además, small sample sizes may be investigated, resulting in lower reliability. Sin embargo, el
use of a formal meta-analytic approach allows for a quantitative review of a large body of LSM
datos, enabling the identification of locations in the brain that show consistent relationships to
behavior across studies (Eickhoff et al., 2012). Por eso, the aim of the present study was to per-
form a systematic review and an anatomic likelihood estimation (PERO) meta-analysis of studies
using LSM methods in combination with a naming task, to identify a pattern of consistent asso-
ciations between brain lesions and word production. Además, a more in-depth analysis was
performed attempting to align the processing stages most likely tapped into by a meta-analyzed
Neurobiology of Language
281
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
study to stages proposed by psycholinguistic models of word production (p.ej., Dell, 1986; Dell
& O’Seaghdha, 1992; Indefrey & Levelt, 2004; Levelt et al., 1999).
In our systematic review, papers using any form of LSM were considered. Papers had to imple-
ment oral naming as a behavioral task in some form. To limit heterogeneity across the researched
participant group, we only included studies in individuals beyond the acute stages of stroke (aquí
defined as at least one month post-stroke). Papers in which the dependent variable was global
accuracy in naming performance were considered for global accuracy analysis. A partir de estos, solo
papers that provided coordinates qualified for ALE meta-analysis (ver figura 1). Papers with a
dependent variable more elaborate than global accuracy, such as error type in naming or com-
bining naming with another language task, were not considered for the ALE meta-analysis since
there was not enough consistency across them, which would introduce large heterogeneity in
the dependent variable tested. En cambio, these papers were considered for a beyond-accuracy
analysis in narrative form. Dependent variables from these papers were linked to the stages of
word production described above in an attempt to elucidate the ALE meta-analysis results.
MATERIALES Y MÉTODOS
Literature Search and Selection
A systematic search was performed, using the Web of Science Core Collection (Clarivate,
2023) and APA PsycINFO (American Psychological Association, 2023) databases using the
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
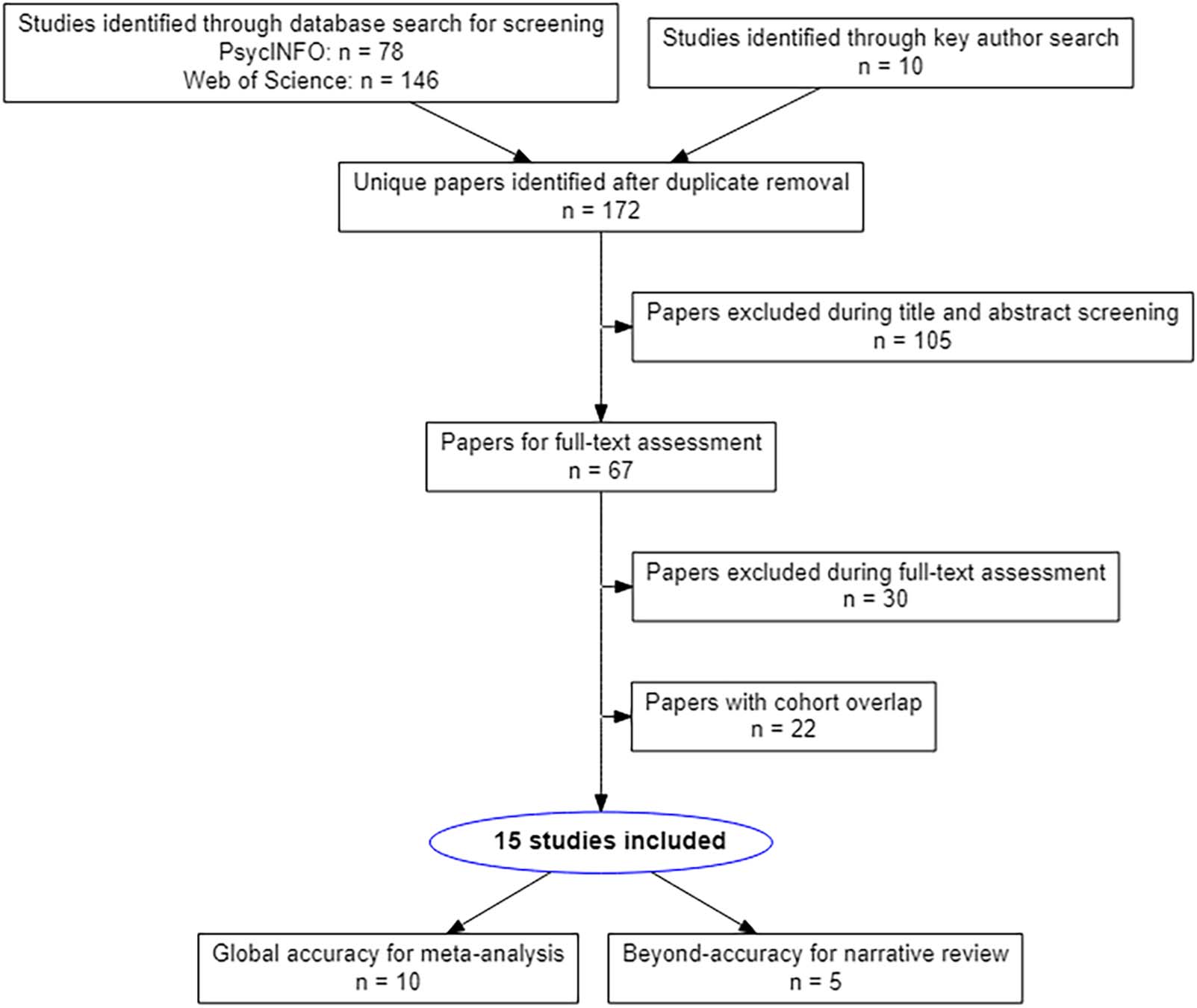
Cifra 1. PRISMA flow diagram of the literature selection process.
Neurobiology of Language
282
Meta-analysis of naming LSM studies
following keywords: [(«naming» O «producción») Y («stroke» O «poststroke» O «infarct»
O «CVA» O «cerebral vascular accident*» O «cerebrovascular accident*» O «post stroke»)
Y («lesion behavio*r mapping» O «voxel wise» O «voxel based» O «symptom mapping»
O «lesion mapping»)]. This resulted in a list of 162 unique papers (last updated on September
18, 2021). To identify other papers not picked up in the automatic search, publications from
the following authors (who have published seminal studies with LSM methodology) eran
screened: Baldo, J.; Binder, j. r.; Crinion, j. T.; Dragoy, o.; Fridriksson, J.; Hope, t. METRO. h.;
Lambon Ralph, METRO. A.; Mirman, D.; Precio, C. J.; Schwartz, METRO. MI.; and Wilson, S. METRO. Más-
más, the reference list of a review paper by Mirman and Thye (2018) was searched for rel-
evant papers. This yielded 10 additional papers. A PRISMA flowchart showing the selection of
these papers can be found in Figure 1.
Papers were independently screened by two authors (title and abstract in phase one, full text
in phase two) on whether they satisfied the following selection criteria: papers had to (1) ser
original empirical work; (2) include stroke patients only; (3) state clearly that all patients were
at least one month post onset of stroke; (4) involve LSM of (5) oral picture naming task perfor-
mance; (6) not reporting single cases; (7) not based on functional imaging or using synthetic
datos; (8) not studying predefined ROIs. During full-text screening, documentos (with corresponding
total number) were excluded according to the following criteria: Not single-word oral picture
naming (norte = 13); no LSM methodology (norte = 6); not all patients >1 month post-stroke (norte = 4);
no (solo) stroke cohort (norte = 2); no coordinates provided (norte = 3); no original or real data (norte = 2).
Paper Categorization
Papers were categorized according to the dependent variable used for LSM, resulting in a
global accuracy analysis data set (es decir., accuracy in naming performance, not further specified).
An exception was made for papers studying a compound naming score from the Western
Aphasia Battery (Kertesz, 1982). This score consists of four naming subtests, one of which
is oral naming, which we considered to suffice for our analyses, thus allowing us to include
more papers in the ALE meta-analysis. Papers in the global accuracy analysis data set had to
provide coordinates to be included for ALE meta-analysis. Authors were contacted for addi-
tional information on foci coordinates. Papers analyzing a more specific score, Por ejemplo,
the number of semantic or phonological errors or after a dimensionality reduction step (p.ej.,
using principal component analysis [PCA]), were included for a beyond-accuracy analysis.
Próximo, for each type of analysis (es decir., global accuracy or beyond-accuracy), relevant papers
were screened for potential overlap in the participant sample by checking overlap in authors
and noting where participants were recruited from. For unclear cases, authors were contacted
to gain information regarding the participants tested, but no response was received. To reduce
the risk of duplicated data, which inflates the effect size, from every subgroup of potentially
overlapping papers, the paper that suited our research purpose best was selected in the fol-
lowing way. If dependent variables used in the overlapping papers were equal, the paper with
the largest cohort was selected. Priority was given to papers providing coordinates over papers
without them. Selection priority was also given to the dependent variable reflecting the most
clear-cut measure of naming (as opposed to the naming score being used with a technique for
dimension reduction such as PCA, which is then related to lesion information), noun naming
specifically rather than verb naming (since verb naming was much less common across the
estudios). If multiple LSM analyses were performed within one paper, selection priority was
given to results from analyses of which noun naming was the largest part, as this is the most
commonly reported measure of naming across papers. También, univariate VLSM results were
Neurobiology of Language
283
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
prioritized over any other form of LSM, and so were manually rather than automatically traced
lesion maps, as these two types tend to be the most common approaches. Además, priority
was given to results controlling for all available covariates. The choice for the most commonly
used approach was meant to increase the comparability across studies and, de este modo, decrease
heterogeneity.
For the ALE meta-analysis, clusters of (potentially) overlapping papers were found. Out of
the first cluster (Alyahya et al., 2018a, 2018b; Butler et al., 2014), the paper by Alyahya et al.
(2018b) was selected for including noun naming (cf. Alyahya et al., 2018a) and a more suit-
able dependent variable to our study (cf. Butler et al., 2014). The second cluster consisted of
two papers (Piras & Marangolo, 2007, 2010), for which Piras and Marangolo (2010) provided
coordinates. For the third cluster (Pustina et al., 2016; Thye & Mirman, 2018; Zhang et al.,
2014), the paper by Thye and Mirman (2018) was selected given the availability of foci coor-
dinates. For the two papers by Lukic et al. (2017, 2021), using verb naming, el que tiene el
larger sample size was chosen (Lukic et al., 2021).
For beyond-accuracy analysis, clusters of (potentially) overlapping papers were obtained.
The first cluster contained three papers (Fridriksson et al., 2016, 2018; Stark et al., 2019), afuera
of which Fridriksson et al. (2018) was selected for studying different error types within naming
(cf. Fridriksson et al., 2016) and the larger sample size (cf. Stark et al., 2019). The second clus-
ter consisted of nine papers (Alyahya et al., 2018a, 2020a, 2020b; Butler et al., 2014; Halai
et al., 2017, 2018a, 2018b; Tochadse et al., 2018; Zhao et al., 2020), out of which Tochadse
et al. (2018) was selected. en este documento, semantic and phonological errors were studied, producir-
ing s and p parameters, respectivamente, according to the computational model of Dell (Dell,
1986; Dell et al., 2013), whereas all other papers in this cluster used PCA on a neuropsycho-
logical test battery, except for Halai et al. (2018b). This latter paper studied different error types
within naming but was excluded as Tochadse et al. (2018) provided, in our opinion, a theo-
retically better motivated distinction between error types than Halai et al. (2018b). The final
cluster consisted of nine papers (Chen et al., 2019; Dell et al., 2013; Mirman, Chen, et al.,
2015; Mirman & Graziano, 2013; Mirman, zhang, et al., 2015; Schwartz et al., 2009, 2011,
2012; Walker et al., 2011), of which Dell et al. (2013) was chosen for conducting VLSM on
parameters derived from computational modeling (as in Tochadse et al., 2018).
The selection yielded 15 original research papers that used LSM techniques in combination
con un (picture) naming task in stroke individuals >1 month post-onset with non-overlapping
cohorts. Ten papers qualified for ALE meta-analysis of global accuracy and five for beyond-
accuracy analysis. The selection and categorization procedure of the included papers can be
found in a PRISMA flowchart, como se muestra en la figura 1.
Quality Assessment
To try to chart the heterogeneity across studies included in the meta-analysis, we performed a
quality assessment of the evidence for the purpose of our systematic review by checking var-
ious parameters. We note that this does not speak to the quality of the papers themselves, pero
rather to the quality of the evidence as it impacts our synthesis and findings. Papers could
receive a maximum of 6 points on the (clarity of the description of the) studied population,
2 points for the clarity of the description of the task and how performance was scored, 4 puntos
para el (description of the) statistical analysis, and finally 1 point for the clarity of the outcome
measure, with a maximum of 13 points in total. Details regarding the parameters and weighted
distributions of points can be found in Table S1 in the Supporting Information, available at
https://doi.org/10.1162/nol_a_00097, and scoring per paper can be found in Table S2.
Neurobiology of Language
284
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
ALE Meta-analysis
ALE meta-analysis was performed using the revised version implemented in BrainMap Ginger-
PERO 3.0 software following the MNI152 template (Eickhoff et al., 2009, 2012; Turkeltaub et al.,
2012), in conjunction with anatomical data (rather than using functional neuroimaging data
for activation likelihood estimation meta-analyses, see for a similar approach Na et al., 2022;
Urgesi et al., 2014). These data concerned peak coordinates of significant clusters associated
with naming task performance as a result of LSM. All extracted coordinates were reported in
Instituto Neurológico de Montreal (MNI) espacio.
In ALE analysis, foci from a given study or experiment are modeled as Gaussian probability
density distributions with a full-width half-maximum (FWHM) calculated from the experi-
ment’s sample size and merged together to form a map. This map therefore represents a sum-
mary of the results of that study, taking into account between-subject and between-template
variabilidad (p.ej., caused by data smoothing and standardization into anatomical space), por
modeling foci as probability distributions rather than singular points. These probability distri-
butions are then taken together by calculating the voxel-wise union of the maps from different
estudios, to assign to every voxel an ALE value equal to the probability that at least one of the
foci in the data set actually lies within this voxel (Turkeltaub et al., 2002, 2012). Por último, estafa-
vergence of foci across experiments is tested by comparing the calculated ALE values against
ALE values obtained under an empirically defined null distribution reflecting random spatial
association. A whole-brain map can then be produced, showing the differential likelihood of
asociaciones (in our case, between lesion and naming score) at all brain locations afforded by
the lesion coverage. Significance was assessed using a cluster-level familywise error correction
set at p < 0.05, with a cluster forming threshold set at p < 0.01 and 1000 permutations. Ana-
tomical labels were obtained from BrainMap GingerALE 3.0, based on the Talairach Daemon
(1988 Talairach atlas). We note that the use of a Gaussian probability density distribution with
FWHM, which is commonly used for functional magnetic resonance imaging (fMRI) studies,
may not be the best option for an ALE meta-analysis of LSM studies. However, fMRI is a hae-
modynamic measure shaped by properties of the vascular system and strokes are vascular in
nature, motivating the use of this distribution. This issue remains nevertheless a limitation of
our approach, as no empirical studies exist validating the use of this probability distribution in
ALE meta-analyses for LSM data.
RESULTS
ALE Meta-analysis of Global Accuracy
Descriptions of the 10 papers used for ALE meta-analysis studying accuracy in naming are
shown in Table 1. These papers in total regarded 69 foci, acquired from 534 subjects. As
far as we could establish, within each paper coverage over left temporal and left inferior frontal
areas was similar; as such, there was in general no particular bias to frontal cortex relative to
temporal cortex.
Quality of the evidence was assessed and total score per paper can be found in Table 1.
Papers scored 9 or higher. Detailed scoring per quality parameter can be found in Table S2.
Results of the ALE analysis can be found in Figure 2 and Table 2. Here, we follow the sub-
division of the temporal lobe into anterior, mid, and posterior portions by Indefrey and Levelt
(2004), with corresponding boundary y coordinates in Talairach space at −7 and −38. Four
significant clusters were identified. Cluster 1, with 7 peaks contributed by seven studies,
had the maximal ALE value (ALE = 0.028) in left anterior temporal cortex (MNI −42, −2,
Neurobiology of Language
285
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
n
o
_
a
_
0
0
0
9
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
2
8
6
Table 1.
List of papers included for anatomic likelihood analysis meta-analysis, studying global accuracy in naming.
Study
Akinina et al.,
2019
N (#f )c
40 (21)
Age
(range or
M ± SD)
33–78
Post-stroke
time
(months)
Language
Dependent
variable
Lesion
delineation
Modality
Analysis
Lesion
volume as
covariate
>3
Russian
Verb
Manual
resonancia magnética
VLSM
Sí
mín.. subjects
with lesioned
voxeld
10% (norte = 4)
No. of foci
(and cluster
contribución)mi
1 (1)
Quality
(/13)
12
naming
exactitud
(picture)
Alyahya et al.,
48 (14)
44–87
>12
Inglés
Noun
Automated
resonancia magnética
VBCM
Sí
n.m.
8 (1, 2)
12
2018b
Baldo et al.,
2013
Faroqi-Shah
et al.,
2014
Geva et al.,
2012a
Griffis et al.,
2017
Lukic et al.,
2021
Piras &
Marangolo,
2010
naming
exactitud
(picture)
96 (21)
31–84
>3
Inglés
Noun
Manual
resonancia magnética, CT
VLSM
n.m.
5% (norte = 5)
1 (2)
12
naming
exactitud
(picture)
31 (10)
42–72
>10
Inglés
Noun
Manual
resonancia magnética
VLSM
n.m.
13% (norte = 4)
5 (2, 3)
11
naming
exactitud
(picture)
21 (7)
21–81
>6
Inglés
Noun
Manual
resonancia magnética
VLSM
Sí
5% (norte = 1)
12* (2, 3, 4)
12
naming
exactitud
(CAT)
43 (18)
23–90
>12
Inglés
Noun
Automated
resonancia magnética
SVR-LSM
Sí
23% (norte = 10)
13 (1, 2, 3)
12
naming
exactitud
(picture)
76 (26)
22–81
>8
Inglés
Verb
Semiautomated MRI
VLSM
Sí
10% (norte = 7)
2 (1, 3)
13
naming
exactitud
(NNB)
20 (7)
38–78
>6
italiano
Noun
Manual
resonancia magnética
VLSM
Sí
25% (norte = 5)
5 (1, 3)
13
naming
exactitud
(picture)
METRO
mi
t
a
–
a
norte
a
yo
y
s
i
s
oh
F
norte
a
metro
i
norte
gramo
l
S
METRO
s
t
tu
d
i
mi
s
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sul et al.,
2019
31 (15)
55.5 ±
11.5
>12
Korean
Naming
puntaje
(K-WAB)
Manual
resonancia magnética
VLSM
n.m.
10% (norte = 3)
7 (1, 2)
9
Thye &
Mirman,
2018b
128 (57)
26–79
>1
Inglés
Noun
Manual
resonancia magnética, CT
VLSM
Sí
10% (norte = 13)
15* (1, 3, 4)
10
naming
exactitud
(PNT)
Nota. Post-stroke time was the minimum time between stroke onset and scanning or testing, whichever was performed earlier. Quality assessment was performed by scoring different
parameters out of a maximum of 13 puntos (details and weighted distributions of points can be found in Table S1, scoring per paper can be found in Table S2). CAT = Comprehensive
Aphasia Test; NNB = Northwestern Naming Battery; K-WAB = Korean version of the Western Aphasia Battery; PNT = Philadelphia Naming Test; MRI = magnetic resonance imaging; CT =
computed tomography; VLSM = voxel-based lesion-symptom mapping; VBCM = voxel-based correlational methodology; SVR-LSM = support vector regression multivariate lesion-symptom
mapping; n.m. = not mentioned in paper or supplementary material.
a For Geva et al. (2012), the statistical map was cluster thresholded at z > 4.61 by the first author of that study.
b For Thye and Mirman (2018), from the statistical map made available, center coordinates of clusters with >10 voxels were selected by the authors, in a procedure blinded for cluster/voxel
ubicación.
c Number of subjects with the amount of females stated in brackets.
d Minimum number of subjects with lesion in a specific voxel before this voxel is included in statistical analysis, presented as percentage out of the full cohort.
e Number of significant foci obtained.
norte
mi
tu
r
oh
b
oh
oh
gramo
y
yo
i
oh
F
l
a
norte
gramo
tu
a
gramo
mi
2
8
7
METRO
mi
t
a
–
a
norte
a
yo
y
s
i
s
oh
F
norte
a
metro
i
norte
gramo
l
S
METRO
s
t
tu
d
i
mi
s
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
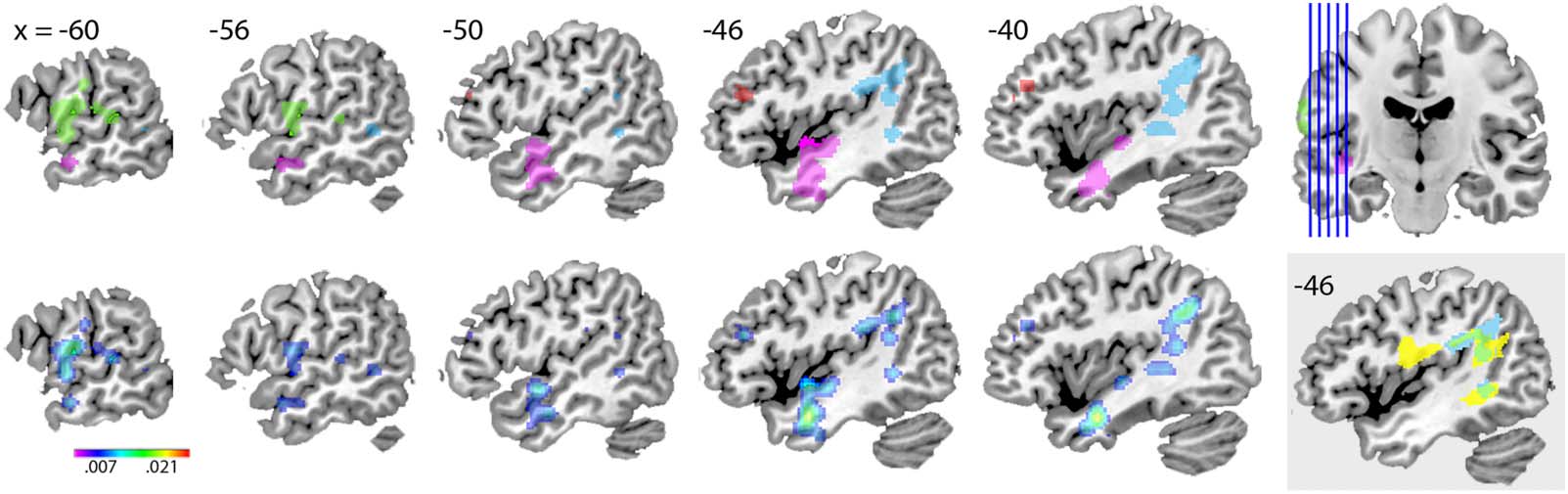
Cifra 2. Results of the anatomic likelihood estimate (PERO) análisis. (Top) Location of the four clusters (grupo 1 in pink, grupo 2 in light
azul, grupo 3 in green, grupo 4 in red), with significant association between global accuracy in naming task performance and brain lesions.
Location of all sagittal slices is indicated in the upper right corner. (Bottom) ALE maps of the four clusters, corrected for cluster-level familywise
error at an alpha level of 0.05 (following voxel-level threshold of 0.01). The color bar indicates the ALE value range. (Gray inset) Cluster 2 (en
azul), arcuate fasciculus (in yellow), and their overlap (in green). The arcuate fasciculus mask was obtained from the Natbrain atlas (Catani &
Thiebaut de Schotten, 2008).
−24), closest to MTG. Cluster 2, con 10 peaks contributed by six studies, had the maximal
ALE value (ALE = 0.016) in left inferior parietal lobule (labeled angular gyrus in the AAL atlas;
Devenyi et al., 2017). Cluster 3, con 5 peaks contributed by six studies, had the maximal ALE
valor (ALE = 0.017) in left postcentral gyrus. Cluster 4, con 5 peaks contributed by two
estudios, had the maximal ALE value (ALE = 0.011) in left.
Beyond-Accuracy Analysis
Details of the five papers considered for beyond-accuracy analysis can be found in Table 3.
These papers measured different dependent variables. We tried to link their word production
measure to proposed stages of word production (Dell, 1986; Levelt et al., 1999; Schwartz
et al., 2004). We note that on the basis of the outcome measures reported, conceptual prep-
aration and lexical selection could not be distinguished.
Out of the papers included for beyond-accuracy analysis, five papers studied a measure
most likely associated with the conceptual-lexical selection stage, and three papers also stud-
ied a measure most likely associated with the phonological encoding stage.
Fridriksson et al. (2018) studied semantic and phonological errors, linked to the conceptual-
lexical and phonological code stages, respectivamente. Lesion-symptom mapping results for
semantic errors revealed significant areas overlapping with cluster 2, the strongest predictor
of semantic error production being lesions in the left “posterior” (authors’ own terminology)
MTG. No significant regions were found to be predictive of phonological errors made in nam-
En g (cf. Schwartz et al., 2012). Of note, our cluster 3 shows overlap with their LSM results for
articulation rate.
A different way to examine lexical-semantic versus phonological stages is through formal-
ization of a computational model. Based on Dell’s computational model of word production
(Dell, 1986; Dell et al., 2013), two relevant parameters are defined: (1) The s parameter, rep-
resenting the connection weights between conceptual and lexical units (“lemma access”), y
(2) the p parameter, the connection weights between the lexical and phonological units. De
performance in picture naming and nonword repetition tests, Dell et al. (2013) derived s and p
parámetros, which were analyzed with VLSM. The s parameter was associated with left “ante-
rior” (authors’ own terminology) STG and MTG, left temporal pole, and left middle and inferior
frontal gyri, overlapping with our four clusters. Además, the association was also present
Neurobiology of Language
288
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
Mesa 2.
Significant anatomic likelihood analysis clusters and corresponding MNI coordinates of the local maxima.
Cluster
1
Volumen (mm3)
6,736
Label
Left Superior Temporal Gyrus, BA 38
ALE score
0.028
Z score
6.3
Left Superior Temporal Gyrus, BA 22
Left Sub-lobar Insula, BA 13
Left Inferior Temporal Gyrus, BA 20
Left Inferior Temporal Gyrus, BA 20
Left Inferior Temporal Gyrus, BA 21
Left Middle Temporal Gyrus, BA 21
2
6,208
Left Superior Temporal Gyrus, BA 39
Arcuate Fasciculus*
Arcuate Fasciculus*
Left Sub-lobar Insula, BA 13
Left Caudate Tail
Left Sub-lobar Insula, BA 13
Left Sub-lobar Insula, BA 13
Left Transverse Temporal Gyrus, BA 41
Left Caudate Tail
Left Middle Temporal Gyrus, BA 22
3,816
Left Transverse Temporal Gyrus, BA 42
Left Superior Temporal Gyrus, BA 22
Left Postcentral Gyrus, BA 2
Left Superior Temporal Gyrus, BA 42
Left Postcentral Gyrus, BA 40
2,360
Left Middle Frontal Gyrus, BA 9
Left Middle Frontal Gyrus, BA 9
Left Middle Frontal Gyrus, BA 10
Left Middle Frontal Gyrus, BA 46
Left Middle Frontal Gyrus, BA 9
3
4
0.013
0.012
0.010
0.010
0.010
0.008
0.016
0.011
0.011
0.011
0.010
0.010
0.010
0.009
0.009
0.008
0.017
0.012
0.010
0.010
0.008
0.011
0.010
0.010
0.010
0.010
3.9
3.8
3.5
3.5
3.4
2.9
4.5
3.6
3.6
3.6
3.6
3.5
3.3
3.2
3.2
2.9
4.5
3.8
3.4
3.4
2.9
3.7
3.5
3.5
3.5
3.5
Coordinates
y
−2
−2
−14
−10
−8
−8
2
−48
−46
−48
−30
−40
−36
−40
−38
−32
−48
−10
−6
−16
−30
−24
40
36
44
34
36
X
−42
−48
−44
−50
−44
−60
−54
−42
−40
−46
−46
−38
−44
−38
−36
−38
−56
−62
−62
−62
−60
−62
−30
−34
−36
−46
−42
z
−24
−8
−6
−22
−34
−16
−16
32
14
0
24
2
28
26
12
2
2
14
0
26
8
14
20
26
18
20
26
Nota. BA = Brodmann Area. Anatomical labeling provided by BrainMap GingerALE 3.0, based on the Talairach Daemon 1988 atlas. MNI = Montreal Neu-
rological Institute.
* No gray matter found, anatomical label derived from the Natbrain atlas (Catani & Thiebaut de Schotten, 2008).
Neurobiology of Language
289
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
Mesa 3.
List of papers studying a beyond-accuracy measure of naming.
Estudiar
Dell et al.,
2013
norte (#F )a
103 (44)
Post-stroke
tiempo
(meses)
>1
Idioma
Inglés
Dependent variable
Semantic and phonological parameter
weights via computational modelling
Stage of word production
Conceptual/lexical selection and
phonological encoding
respectivamente
Fridriksson
et al.,
2018
harvey &
Schnur,
2015
Schnur
et al.,
2009
Tochadse
et al.,
2018
105 (n.m.)
>6
Inglés
Semantic and phonological noun
Conceptual/lexical selection and
naming errors
phonological encoding
respectivamente
15 (4)
>6
Inglés
Semantic interference in noun naming
Conceptual/lexical selection
12 (n.m.)
>10
Inglés
Growth of semantic interference in noun
Conceptual/lexical selection
naming
53 (n.m.)
>12
Inglés
Semantic and phonological parameter
weights via computational modelling
Conceptual/lexical selection and
phonological encoding
respectivamente
Nota. Post-stroke time was the minimum time between stroke onset and scanning or testing, whichever was performed earlier. n.m. = not mentioned in paper or
supporting information.
a Number of subjects with the number of females stated in brackets.
posteriorly, at the temporo-parietal and parietal-temporal-occipital junctions, including AG.
The p parameter was mainly associated with left SMG and postcentral gyrus (also including
precentral gyrus and insula, similar to our cluster 3). Tochadse et al. (2018) similarly studied
naming through s and p weights from Dell’s computational model. Based on the peak coor-
dinates obtained in their VBCM analysis and coordinates for the subdivision of the temporal
lobe (based on Indefrey & Levelt, 2004), the s parameter was associated with regions in the
mid portion of the left temporal lobe. Peak coordinates associated with the p parameter were
located either in the anterior or mid portions of the temporal lobe.
Two other papers combined LSM with semantic interference, which could be linked to the
conceptual-lexical selection stages of word production (Roelofs, 2018). Harvey and Schnur
(2015) studied the areas involved in both semantic interference and growth of interference
across cycles in naming. The largest significant cluster associated with semantic interference
was located in the left posterior MTG (according to the subdivision adopted here: MNI −52,
−40, −5, Talairach y = −40), close to our cluster 2, whilst the other cluster was located in the
left mid MTG (according to the subdivision adopted here: MNI −49, −21, −8, Talairach y =
−22). No region was significantly associated with growth in interference across naming cycles.
This latter dependent variable, eso es, the growth of interference, was specifically studied in a
study by Schnur et al. (2009). VLSM analysis revealed that growth of interference was signif-
icantly related to voxels only in the “posterior” (author’s own terminology) left IFG.
To conclude the beyond-accuracy analysis, a tendency seems to be present across studies
for deficits in the conceptual preparation and/or lexical retrieval stages to be associated with
lesions in somewhat more mid to posterior temporal regions. Regarding the phonological code
retrieval stage, since only two studies obtained statistically significant results (Dell et al., 2013;
Tochadse et al., 2018) that were not converging, the evidence remains inconclusive.
Neurobiology of Language
290
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
DISCUSIÓN
To investigate the brain areas critical for word production, the present study quantitatively
compared results from papers combining lesion-symptom mapping with global accuracy
scores in naming, by performing an ALE meta-analysis. We identified four separate clusters.
One cluster was predominantly in the anterior portion of the left temporal lobe, in STG, MTG,
and ITG. The second cluster was predominantly in the posterior portion of the left temporal
lobe including the inferior parietal lobule, mostly in white matter. An overlay of this cluster
with the outline of the arcuate fasciculus indicated a large degree of overlap. The third cluster
had a peak in postcentral gyrus and the fourth cluster in middle frontal gyrus. No peaks were
identified in the left IFG. This distribution was found despite a similar lesion coverage over left
inferior frontal as over left temporal lobe areas. En general, the quality of the evidence across
studies was good for the purpose of our review and meta-analysis. The vast majority of the
studies were conducted in English-speaking countries, with only three other languages repre-
sented in our sample.
Three papers could not be included in the global accuracy due to missing coordinates. El
three strongest predictors of correct naming obtained by Fridriksson et al. (2018) were the left
“posterior” (authors’ own terminology) STG, AG, and SMG. Anterior portions of the temporal
lobe were also found in this study, though these predictors were less strong. Pillay et al.
(2017) provided a VLSM map of picture naming in their supplementary materials, cual
revealed significant areas (through visual inspection) in posterior portions of left STG, inferior
parietal lobule, lateral frontal cortex, and insula. Finalmente, in the study by Skipper-Kallal et al.
(2017), impairment in picture naming was associated with left posterior STG (authors’ own
terminology) En particular, but also with left AG, intraparietal sulcus, and parts of the pars
triangularis in the IFG.
Since overall naming scores reflect a mixture of errors, it is difficult to relate the patterns
found to particular stages of word production. Por ejemplo, while Akinina et al. (2019) took
care to try to isolate the lexical stage, and Baldo et al. (2013) reported their findings while
covarying for visual perception and overall speech fluency deficits, the global accuracy
measure in other studies is less specific to one or a couple of stages. Por lo tanto, nosotros también
synthesized studies examining measures beyond global accuracy in an attempt to elucidate
the patterns found by relating them as much as possible to particular stages of word produc-
tion as stipulated by psycholinguistic models. We found tentative evidence that conceptual
preparation and/or lexical selection are associated with lesions in somewhat more mid to
posterior temporal lobe regions, whereas the evidence for phonological encoding was less
consistent across studies.
In the course of publishing this work, another meta-analysis of lesion-symptom mapping
studies was published focusing on various language tasks (Na et al., 2022). For naming, el
authors found a cluster in the left parahippocampal gyrus and left mid STG (MNI −59, −11, 7,
Brodman Area 22, Talairach y = −12). This cluster is in the proximity of cluster 3 we identified.
Sin embargo, unlike in our meta-analysis, the authors did not differentiate between phase of the
stroke (acute, subacute, and chronic were all included) or performance measure (global accu-
racy as well as specific error types were included) in the analysis.
While two previous (semi-)systematic reviews and meta-analyses have provided evidence
on the neural substrates of more specific stages of word production based on correlational
measures (Indefrey & Levelt, 2004; Precio, 2012), here we explicitly sought to provide causal
evidencia. The meta-analysis of Indefrey and Levelt (2004) has suggested that lexical selection
is associated with left MTG (and the mid portion in particular), whereas phonological code
Neurobiology of Language
291
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
retrieval (part of phonological encoding) is associated with left posterior MTG and STG. Syl-
labification (the ordering of phonemes into syllables, part of phonological encoding) is asso-
ciated with left posterior IFG and phonetic encoding and articulation mainly with bilateral
ventral motor and sensory regions. Our results for global naming accuracy are partly in line
with this proposal. The beyond-accuracy analysis, por el contrario, provides tentative evidence in
agreement with this proposal regarding the conceptual/lexical stages, which in our findings
were related to more mid to posterior temporal lobe regions.
Our results concern stroke populations only. Sin embargo, given that different pathologies have
their intrinsic spatial biases—in the case of stroke, due to cerebrovascular organization—trying
to bridge across pathologies would be fruitful for our understanding of the neural basis of lan-
guage production. These studies are however scarcer, so no meta-analysis is possible given the
current literature. Of note, converging evidence for the present results can be found. Para examen-
por ejemplo, in an LSM study examining presurgical brain tumor cases (Faulkner & Wilshire, 2020),
conceptual/lexical selection (operationalized as semantic errors and omissions in picture nam-
ing and category fluency performance covaried for letter fluency performance) was associated
with left posterior MTG and parts of the lateral occipital cortex and ITG. Phonological encod-
En g (operationalized as accuracy on word and nonword repetition and effect of word length in
picture naming) was associated with left posterior SMG and AG. In individuals with Alzheimer
enfermedad, correlations between hypometabolism and naming errors have also been found (Isella
et al., 2020). En particular, semantic errors were related to the mid portion of the MTG and left
ITG. Formal errors (es decir., the resulting existing word resembles the target word in terms of its
forma, “rat” instead of “mat”), tapping both into the connections between lexical and phonol-
ogical units and into phonological encoding (Dell et al., 1997), were associated with left
anterior/mid MTG. Finalmente, producing neologisms and nonwords was associated with left
SMG and the mid portion of STG. In a cohort of individuals with primary progressive aphasia
producing semispontaneous speech (Wilson et al., 2010), atrophy in the mid portion of the left
temporal lobe (mainly MTG) was associated with the production of nouns of increasing lexical
frequency, a measure tapping into both lexical and phonological stages (Kittredge et al., 2008).
Sucesivamente, producing phonological paraphasias was associated with atrophy in the mid portion
of left STG. En suma, converging with the stroke-aphasia literature reviewed above,
conceptual/lexical stages tend to be associated with the mid to posterior portions of the tem-
poral lobe, and MTG in particular, in addition to ITG. This latter region is not often represented
in stroke-aphasia cases given the difference in arterial blood supply between STG and MTG on
la una mano, and ITG on the other. Once phonological representations become implicated, a
tendency is seen for associations with (mid) STG and SMG, which is also suggested by some of
the stroke studies we reviewed.
En general, the literature strongly suggests an important role for the left temporal, en vez de
frontal, lobe in naming, contrary to a perhaps more popular view of the left temporal lobe as
the site for comprehension and the frontal lobe for production. Part of this misconception may
have its roots in the fact that producing language is a motor function, which nevertheless
requires the retrieval of conceptual, lexical, and phonological information, processes that
we would argue are not particularly linked to the frontal cortex.
Admittedly, the vast majority of the studies reviewed here employed noun rather than verb
naming (with the exception of Akinina et al., 2019; Lukic et al., 2021), a deliberate choice to
increase the comparability across papers. A review of a large body of literature comparing
nouns and verbs has concluded that these two grammatical classes are processed by a largely
overlapping set of areas in production and comprehension (Vigliocco et al., 2011). Neural dif-
ferences between the two emerge, sin embargo, as a function of the task (among a few other
Neurobiology of Language
292
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
variables, see Vigliocco et al., 2011): Por ejemplo, when there is an emphasis in morphosyntax
as in verb naming, in which case the involvement of left IFG becomes more prominent. In both
Akinina et al. (2019) and Lukic et al. (2021), the accuracy score was based on any correct
morphological form of the verb, thus emphasizing more morphosyntax than noun/object nam-
ing does. De este modo, one could argue that our inclusion of more studies examining noun naming
rather than verb naming may have overemphasized the temporal rather than the frontal cortex.
Sin embargo, the frontal cortex contribution in this case would arguably not be due to conceptual-
lexical (and phonological) information being retrieved. De este modo, our conclusion remains that the
left temporal, rather than the frontal cortex, is the most critical for conceptually driven naming.
The terminology adopted by authors for subdividing the temporal lobe is not always
consistent or transparent. Aquí, we opted for adopting a coordinate-based standard used by
Indefrey and Levelt (2004), separating the temporal lobe into an anterior, a mid, and a poste-
rior portion. This system is useful for being more objective (when coordinates are available).
Sin embargo, the boundaries between these portions are not necessarily a subdivision reflecting
some form of organization of the temporal lobe (a “natural kind,” cf. cytoarchitectonics, tran-
scriptomics, etc.), but rather a convention adopted to help researchers structure results. Future
research could attempt to link findings of particular locations in the temporal lobe to physio-
logically and biologically based subdivisions, which could prove useful for integrating findings
across studies using different methods and elucidating temporal lobe functions.
A limitation of our study is the small number of comparable papers in the analyses per-
formed. We used strict criteria whereby papers were excluded if there was cohort overlap
or if we could not ascertain that there was no overlap. Por eso, more empirical studies are
required to increase robustness of the ALE meta-analysis and to strengthen our claims on
the psychological nature of the foci identified in the present study. These limitations relate
to two recommendations we can make to improve the field of language production. En primer lugar,
authors would ideally disclose cohort overlap with previous studies (this was done in some,
but not all, of the studies we reviewed). En segundo lugar, following open science practices, autores
would ideally make postprocessed data available (p.ej., Thye & Mirman, 2022). An alternative
to this solution would be to report a set of coordinates for as much as the method affords. A
second limitation of our study is the use of the ALE method without a validation for the prob-
ability distribution we employed in combination with LSM data.
En conclusión, the ALE meta-analysis of 10 lesion-symptom mapping studies of naming per-
formance yielded distinct clusters, predominantly in anterior and posterior portions of the left
temporal lobe, for which the posterior distribution seems to follow the arcuate fasciculus. Two
additional clusters were found in postcentral and middle frontal gyrus. No peaks were iden-
tified in the left IFG. Regions consistent with these foci were also revealed by examining
papers studying more detailed measures of naming or other populations than stroke, dónde
we found a tendency for lesions in mid to posterior parts of the temporal lobe to be more
consistently associated with conceptual-lexical deficits. A major limitation of the present study
remains the small number of papers included in the meta-analysis.
EXPRESIONES DE GRATITUD
The authors are indebted to Peter Indefrey for critical discussions, Sharon Geva, Daniel
Mirman, Melissa Thye, Dorian Pustina, Sladjana Lukic, and Laura Skipper-Kallal for providing
additional information for the meta-analysis, and Daniel Sharoh for a blinded procedure of
cluster threshold of one data set. The authors are also thankful to the critique provided by three
anonymous reviewers, which substantially improved the quality of the work presented here.
Neurobiology of Language
293
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
INFORMACIÓN DE FINANCIACIÓN
Vitória Piai, Dutch Research Council, Award ID: NOW, 451-17-003. Vitória Piai, Language in
Interaction Consortium, Dutch Research Council, Award ID: NOW, 024.001.006.
CONTRIBUCIONES DE AUTOR
Vitória Piai: Conceptualización; Análisis formal; Administración de proyecto; Visualización; Writing –
original draft; Escritura – revisión & edición. Dilys Eikelboom: Análisis formal; Project adminis-
tration; Visualización; Escritura – borrador original; Escritura – revisión & edición.
DATA AVAILABILITY STATEMENT
All data associated with these analyses are available via https://osf.io/8xtp9/.
REFERENCIAS
Akinina, y., Dragoy, o., Ivanova, METRO. v., Iskra, mi. v., Soloukhina,
oh. A., Petryshevsky, A. GRAMO., Fedina, oh. NORTE., Turken, A. Ud.,
Shklovsky, V. METRO., & Dronkers, norte. F. (2019). Grey and white mat-
ter substrates of action naming. Neuropsicología, 131, 249–
265. https://doi.org/10.1016/j.neuropsychologia.2019.05.015,
PubMed: 31129278
Alyahya, R. S. w., Halai, A. D., Conroy, PAG., & Lambon Ralph, METRO. A.
(2018a). The behavioural patterns and neural correlates of con-
crete and abstract verb processing in aphasia: A novel verb
semantic battery. NeuroImagen: Clinical, 17, 811–825. https://
doi.org/10.1016/j.nicl.2017.12.009, PubMed: 29619318
Alyahya, R. S. w., Halai, A. D., Conroy, PAG., & Lambon Ralph, METRO. A.
(2018b). Noun and verb processing in aphasia: Behavioural pro-
files and neural correlates. NeuroImagen: Clinical, 18, 215–230.
https://doi.org/10.1016/j.nicl.2018.01.023, PubMed: 29868446
Alyahya, R. S. w., Halai, A. D., Conroy, PAG., & Lambon Ralph, METRO. A.
(2020a). Mapping psycholinguistic features to the neuropsycho-
logical and lesion profiles in aphasia. Corteza, 124, 260–273.
https://doi.org/10.1016/j.cortex.2019.12.002, PubMed:
31958653
Alyahya, R. S. w., Halai, A. D., Conroy, PAG., & Lambon Ralph, METRO. A.
(2020b). A unified model of post-stroke language deficits
including discourse production and their neural correlates. Cerebro,
143(5), 1541–1554. https://doi.org/10.1093/ brain/awaa074,
PubMed: 32330940
American Psychological Association. (2023). APA PsycINFO (datos-
base). https://www.apa.org/pubs/databases/psycinfo/
Ashburner, J., & Friston, k. j. (2000). Voxel-based morphometry—
The methods. NeuroImagen, 11(6), 805–821. https://doi.org/10
.1006/nimg.2000.0582, PubMed: 10860804
Baldo, j. v., Arévalo, A., Patterson, j. PAG., & Dronkers, norte. F. (2013).
Grey and white matter correlates of picture naming: Evidencia
from a voxel-based lesion analysis of the Boston Naming Test.
Corteza, 49(3), 658–667. https://doi.org/10.1016/j.cortex.2012
.03.001, PubMed: 22482693
Bates, MI., wilson, S. METRO., Saygin, A. PAG., Dick, F., Sereno, METRO. I.,
Caballero, R. T., & Dronkers, norte. F. (2003). Voxel-based lesion–
symptom mapping. Neurociencia de la naturaleza, 6(5), 448–450.
https://doi.org/10.1038/nn1050, PubMed: 12704393
mayordomo, R. A., Lambon Ralph, METRO. A., & Woollams, A. METRO. (2014). Cap-
turing multidimensionality in stroke aphasia: Mapping principal
behavioural components to neural structures. Cerebro, 137(12),
3248–3266. https://doi.org/10.1093/ brain/awu286, PubMed:
25348632
Catani, METRO., & Thiebaut de Schotten, METRO. (2008). A diffusion tensor
imaging tractography atlas for virtual in vivo dissections. Corteza,
44(8), 1105–1132. https://doi.org/10.1016/j.cortex.2008.05.004,
PubMed: 18619589
Chen, P., Middleton, MI., & Mirman, D. (2019). Words fail:
Lesion-symptom mapping of errors of omission in post-stroke
aphasia. Journal of Neuropsychology, 13(2), 183–197. https://
doi.org/10.1111/jnp.12148, PubMed: 29411521
Clarivate. (2023). Web of Science Core Collection (database).
Scientific and academic research solutions – accelerate research to advance the knowledge frontier
/research-discovery-and-workflow-solutions/web-of-science/web
-of-science-core-collection/
Dell, GRAMO. S. (1986). A spreading-activation theory of retrieval in sen-
tence production. Revisión psicológica, 93(3), 283–321. https://
doi.org/10.1037/0033-295X.93.3.283, PubMed: 3749399
Dell, GRAMO. S., & O’Seaghdha, PAG. GRAMO. (1992). Stages of lexical access in
language production. Cognición, 42(1–3), 287–314. https://doi
.org/10.1016/0010-0277(92)90046-k, PubMed: 1582160
Dell, GRAMO. S., Schwartz, METRO. F., Martín, NORTE., Saffran, mi. METRO., & Gagnon,
D. A. (1997). Lexical access in aphasic and nonaphasic speakers.
Revisión psicológica, 104(4), 801–838. https://doi.org/10.1037
/0033-295X.104.4.801, PubMed: 9337631
Dell, GRAMO. S., Schwartz, METRO. F., Nozari, NORTE., Faseyitan, o., & Branch
Coslett, h. (2013). Voxel-based lesion-parameter mapping: Iden-
tifying the neural correlates of a computational model of word
producción. Cognición, 128(3), 380–396. https://doi.org/10
.1016/j.cognition.2013.05.007, PubMed: 23765000
Devenyi, GRAMO. A., Pipitone, J., & Raihaan, PAG. (2017). AAL atlas (Soft-
mercancía). https://github.com/CobraLab/documentation/wiki/AAL
-Atlas
Eickhoff, S. B., Bzdok, D., Laird, A. r., Kurth, F., & Fox, PAG. t. (2012).
Activation likelihood estimation meta-analysis revisited. Neuro-
Image, 59(3), 2349–2361. https://doi.org/10.1016/j.neuroimage
.2011.09.017, PubMed: 21963913
Eickhoff, S. B., Laird, A. r., Grefkes, C., Wang, l. MI., Zilles, K., &
Fox, PAG. t. (2009). Coordinate-based activation likelihood estima-
tion meta-analysis of neuroimaging data: A random-effects
approach based on empirical estimates of spatial uncertainty.
Mapeo del cerebro humano, 30(9), 2907–2926. https://doi.org/10
.1002/hbm.20718, PubMed: 19172646
Faroqi-Shah, y., Kling, T., Solomon, J., Liu, S., Parque, GRAMO., & Braun, A.
(2014). Lesion analysis of language production deficits in apha-
sia. Aphasiology, 28(3), 258–277. https://doi.org/10.1080
/02687038.2013.853023
Neurobiology of Language
294
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
Faulkner, j. w., & Wilshire, C. mi. (2020). Mapping eloquent cortex:
A voxel-based lesion-symptom mapping study of core speech
production capacities in brain tumour patients. Brain and Lan-
guage, 200, Article 104710. https://doi.org/10.1016/j.bandl
.2019.104710, PubMed: 31739187
Fridriksson, J., den Ouden, D.-B., Hillis, A. MI., Hickok, GRAMO., Rorden,
C., Basilakos, A., Yourganov, GRAMO., & Bonilha, l. (2018). Anatomy
of aphasia revisited. Cerebro, 141(3), 848–862. https://doi.org/10
.1093/brain/awx363, PubMed: 29360947
Fridriksson, J., Yourganov, GRAMO., Bonilha, l., Basilakos, A., den
Ouden, D.-B., & Rorden, C. (2016). Revealing the dual streams
of speech processing. Actas de la Academia Nacional de
Ciencias, 113(52), 15108–15113. https://doi.org/10.1073/pnas
.1614038114, PubMed: 27956600
Geva, S., Baron, J.-C., jones, PAG. S., Precio, C. J., & Warburton, mi. A.
(2012). A comparison of VLSM and VBM in a cohort of patients
with post-stroke aphasia. NeuroImagen: Clinical, 1(1), 37–47.
https://doi.org/10.1016/j.nicl.2012.08.003, PubMed: 24179735
Griffis, j. C., Nenert, r., Allendorfer, j. B., & Szaflarski, j. PAG. (2017).
Damage to white matter bottlenecks contributes to language
impairments after left hemispheric stroke. NeuroImagen: Clinical,
14, 552–565. https://doi.org/10.1016/j.nicl.2017.02.019,
PubMed: 28337410
Halai, A. D., Woollams, A. METRO., & Lambon Ralph, METRO. A. (2017).
Using principal component analysis to capture individual differ-
ences within a unified neuropsychological model of chronic
post-stroke aphasia: Revealing the unique neural correlates of
speech fluency, phonology and semantics. Corteza, 86, 275–
289. https://doi.org/10.1016/j.cortex.2016.04.016, PubMed:
27216359
Halai, A. D., Woollams, A. METRO., & Lambon Ralph, METRO. A. (2018a).
Predicting the pattern and severity of chronic post-stroke lan-
guage deficits from functionally-partitioned structural lesions.
NeuroImagen: Clinical, 19, 1–13. https://doi.org/10.1016/j.nicl
.2018.03.011, PubMed: 30038893
Halai, A. D., Woollams, A. METRO., & Lambon Ralph, METRO. A. (2018b).
Triangulation of language-cognitive impairments, naming errors
and their neural bases post-stroke. NeuroImagen: Clinical, 17,
465–473. https://doi.org/10.1016/j.nicl.2017.10.037, PubMed:
29159059
harvey, D. y., & Schnur, t. t. (2015). Distinct loci of lexical and
semantic access deficits in aphasia: Evidence from voxel-based
lesion-symptom mapping and diffusion tensor imaging. Corteza,
67, 37–58. https://doi.org/10.1016/j.cortex.2015.03.004,
PubMed: 25880795
Hernandez-Pavon, j. C., Makela, NORTE., Lehtinen, h., Lioumis, PAG., &
Makela, j. PAG. (2014). Effects of navigated TMS on object and
action naming. Frontiers in Human Neuroscience, 8, Article 660.
https://doi.org/10.3389/fnhum.2014.00660, PubMed: 25228868
Hickok, GRAMO., & Poeppel, D. (2007). The cortical organization of
speech processing. Naturaleza Reseñas Neurociencia, 8(5), 393–402.
https://doi.org/10.1038/nrn2113, PubMed: 17431404
Indefrey, PAG., & Levelt, W.. j. METRO. (2004). The spatial and temporal sig-
natures of word production components. Cognición, 92(1–2),
101–144. https://doi.org/10.1016/j.cognition.2002.06.001,
PubMed: 15037128
Isella, v., Rosazza, C., Gazzotti, METRO., Sala, J., Morzenti, S., Crivellaro,
C., Appollonio, I. METRO., Ferrarese, C., & Luzzatti, C. (2020). A
metabolic imaging study of lexical and phonological naming
errors in Alzheimer disease. American Journal of Alzheimer’s
Disease & Other Dementias, 35, Article 1533317520922390.
https://doi.org/10.1177/1533317520922390, PubMed:
32356456
Ivanova, METRO. v., Herron, t. J., Dronkers, norte. F., & Baldo, j. V. (2021).
An empirical comparison of univariate versus multivariate
methods for the analysis of brain–behavior mapping. Humano
Mapeo cerebral, 42(4), 1070–1101. https://doi.org/10.1002/hbm
.25278, PubMed: 33216425
Kertesz, A. (1982). The Western aphasia battery. Grune & Stratton.
Kittredge, A. K., Dell, GRAMO. S., Verkuilen, J., & Schwartz, METRO. F. (2008).
Where is the effect of frequency in word production? Insights
from aphasic picture-naming errors. Cognitive Neuropsychology,
25(4), 463–492. https://doi.org/10.1080/02643290701674851,
PubMed: 18704797
Levelt, W.. j. METRO., Roelofs, A., & Meyer, A. S. (1999). A theory of lex-
ical access in speech production. Ciencias del comportamiento y del cerebro,
22(1), 1–38. https://doi.org/10.1017/S0140525X99001776,
PubMed: 11301520
Liljeström, METRO., Hultén, A., Parkkonen, l., & Salmelin, R. (2009).
Comparing MEG and fMRI views to naming actions and objects.
Mapeo del cerebro humano, 30(6), 1845–1856. https://doi.org/10
.1002/hbm.20785, PubMed: 19378277
Lukic, S., Barbieri, MI., Wang, X., Caplan, D., Kiran, S., Rapp, B.,
Parrish, t. B., & Thompson, C. k. (2017). Right hemisphere grey
matter volume and language functions in stroke aphasia. Neural
Plasticity, 2017, Article 5601509. https://doi.org/10.1155/2017
/5601509, PubMed: 28573050
Lukic, S., Thompson, C. K., Barbieri, MI., Chiappetta, B., Bonakdarpour,
B., Kiran, S., Rapp, B., Parrish, t. B., & Caplan, D. (2021). Common
and distinct neural substrates of sentence production and com-
prehension. NeuroImagen, 224, Article 117374. https://doi.org/10
.1016/j.neuroimage.2020.117374, PubMed: 32949711
Mirman, D., Chen, P., zhang, y., Wang, Z., Faseyitan, oh. K.,
Coslett, h. B., & Schwartz, METRO. F. (2015). Neural organization
of spoken language revealed by lesion-symptom mapping.
Comunicaciones de la naturaleza, 6, Article 6762. https://doi.org/10
.1038/ncomms7762, PubMed: 25879574
Mirman, D., & Graziano, k. METRO. (2013). The neural basis of inhibi-
tory effects of semantic and phonological neighbors in spoken
word production. Revista de neurociencia cognitiva, 25(9),
1504–1516. https://doi.org/10.1162/jocn_a_00408, PubMed:
23647518
Mirman, D., & Thye, METRO. (2018). Uncovering the neuroanatomy of
core language systems using lesion-symptom mapping. Actual
Directions in Psychological Science, 27(6), 455–461. https://doi
.org/10.1177/0963721418787486
Mirman, D., zhang, y., Wang, Z., Coslett, h. B., & Schwartz, METRO. F.
(2015). The ins and outs of meaning: Behavioral and neuroana-
tomical dissociation of semantically-driven word retrieval and
multimodal semantic recognition in aphasia. Neuropsicología,
76, 208–219. https://doi.org/10.1016/j.neuropsychologia.2015
.02.014, PubMed: 25681739
Na, y., Jung, J., Tench, C. r., Auer, D. PAG., & Pyun, S.-B. (2022). Lan-
guage systems from lesion-symptom mapping in aphasia: A
meta-analysis of voxel-based lesion mapping studies. Neuro-
Image: Clinical, 35, Article 103038. https://doi.org/10.1016/j
.nicl.2022.103038, PubMed: 35569227
Pillay, S. B., Binder, j. r., Humphries, C., Bruto, W.. l., & Book,
D. S. (2017). Lesion localization of speech comprehension
deficits in chronic aphasia. Neurología, 88(10), 970–975.
https://doi.org/10.1212/ WNL.0000000000003683, PubMed:
28179469
Piras, F., & Marangolo, PAG. (2007). Noun–verb naming in aphasia: A
voxel-based lesion-symptom mapping study. NeuroReport, 18(14),
1455–1458. https://doi.org/10.1097/ WNR.0b013e3282ef6fc9,
PubMed: 17712273
Neurobiology of Language
295
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Meta-analysis of naming LSM studies
Piras, F., & Marangolo, PAG. (2010). When “Crack walnuts” lies in
different brain regions: Evidence from a voxel-based lesion-
symptom mapping study. Journal of the International Neuropsy-
chological Society, 16(3), 433–442. https://doi.org/10.1017
/S1355617710000068, PubMed: 20178682
Precio, C. j. (2012). A review and synthesis of the first 20 years of PET
and fMRI studies of heard speech, spoken language and reading.
NeuroImagen, 62(2), 816–847. https://doi.org/10.1016/j
.neuroimage.2012.04.062, PubMed: 22584224
Precio, C. J., moore, C. J., Humphreys, GRAMO. w., Frackowiak, R. S. J., &
Friston, k. j. (1996). The neural regions sustaining object recog-
nition and naming. Proceedings of the Royal Society B: Biológico
Ciencias, 263(1376), 1501–1507. https://doi.org/10.1098/rspb
.1996.0219, PubMed: 8952093
Pustina, D., Coslett, h. B., Turkeltaub, PAG. MI., Tustison, NORTE., Schwartz,
METRO. F., & Avants, B. (2016). Automated segmentation of chronic
stroke lesions using LINDA: Lesion identification with neighbor-
hood data analysis. Mapeo del cerebro humano, 37(4), 1405–1421.
https://doi.org/10.1002/hbm.23110, PubMed: 26756101
Roelofs, A. (2014). A dorsal-pathway account of aphasic language
producción: The WEAVER++/ARC model. Corteza, 59, 33–48.
https://doi.org/10.1016/j.cortex.2014.07.001, PubMed: 25128898
Roelofs, A. (2018). A unified computational account of cumulative
semantic, semantic blocking, and semantic distractor effects in
picture naming. Cognición, 172, 59–72. https://doi.org/10.1016
/j.cognition.2017.12.007, PubMed: 29232595
Schnur, t. T., Schwartz, METRO. F., Kimberg, D. y., Hirshorn, MI., Coslett,
h. B., & Thompson-Schill, S. l. (2009). Localizing interference
during naming: Convergent neuroimaging and neuropsycholog-
ical evidence for the function of Broca’s area. Actas de la
Academia Nacional de Ciencias, 106(1), 322–327. https://doi.org
/10.1073/pnas.0805874106, PubMed: 19118194
Schwartz, METRO. F., Faseyitan, o., kim, J., & Coslett, h. B. (2012). El
dorsal stream contribution to phonological retrieval in object
naming. Cerebro, 135(12), 3799–3814. https://doi.org/10.1093
/brain/aws300, PubMed: 23171662
Schwartz, METRO. F., Kimberg, D. y., Caminante, GRAMO. METRO., Brecher, A.,
Faseyitan, oh. K., Dell, GRAMO. S., Mirman, D., & Coslett, h. B.
(2011). Neuroanatomical dissociation for taxonomic and the-
matic knowledge in the human brain. Actas de la
Academia Nacional de Ciencias, 108(20), 8520–8524. https://
doi.org/10.1073/pnas.1014935108, PubMed: 21540329
Schwartz, METRO. F., Kimberg, D. y., Caminante, GRAMO. METRO., Faseyitan, o.,
Brecher, A., Dell, GRAMO. S., & Coslett, h. B. (2009). Anterior tempo-
ral involvement in semantic word retrieval: Voxel-based
lesion-symptom mapping evidence from aphasia. Cerebro,
132(12), 3411–3427. https://doi.org/10.1093/ brain/awp284,
PubMed: 19942676
Schwartz, METRO. F., Wilshire, C. MI., Gagnon, D. A., & Polansky, METRO.
(2004). Origins of nonword phonological errors in aphasic pic-
ture naming. Cognitive Neuropsychology, 21(2–4), 159–186.
https://doi.org/10.1080/02643290342000519, PubMed:
21038198
Skipper-Kallal, l. METRO., Lacey, mi. h., Xing, S., & Turkeltaub, PAG. mi.
(2017). Functional activation independently contributes to nam-
ing ability and relates to lesion site in post-stroke aphasia.
Mapeo del cerebro humano, 38(4), 2051–2066. https://doi.org/10
.1002/hbm.23504, PubMed: 28083891
Rígido, B. C., Basilakos, A., Hickok, GRAMO., Rorden, C., Bonilha, l., &
Fridriksson, j. (2019). Neural organization of speech production:
A lesion-based study of error patterns in connected speech. Cor-
tex, 117, 228–246. https://doi.org/10.1016/j.cortex.2019.02.029,
PubMed: 31005024
Sul, B., Sotavento, k. B., hong, B. y., kim, j. S., kim, J., Hwang, W.. S., &
Lim, S. h. (2019). Association of lesion location with long-term
recovery in post-stroke aphasia and language deficits. Fronteras en
Neurología, 10, Article 776. https://doi.org/10.3389/fneur.2019
.00776, PubMed: 31396146
Thye, METRO., & Mirman, D. (2018). Relative contributions of lesion
location and lesion size to predictions of varied language deficits
in post-stroke aphasia. NeuroImagen: Clinical, 20, 1129–1138.
https://doi.org/10.1016/j.nicl.2018.10.017, PubMed: 30380520
Thye, METRO., & Mirman, D. (2022). Relative contributions of lesion
location and lesion size to predictions of varied language deficits
in post-stroke aphasia [Proyecto]. https://osf.io/w4u2q/
Tochadse, METRO., Halai, A. D., Lambon Ralph, METRO. A., & Abel, S.
(2018). Unification of behavioural, computational and neural
accounts of word production errors in post-stroke aphasia. nuevo-
roImage: Clinical, 18, 952–962. https://doi.org/10.1016/j.nicl
.2018.03.031, PubMed: 29876280
Turkeltaub, PAG. MI., Eden, GRAMO. F., jones, k. METRO., & Zeffiro, t. A. (2002).
Meta-analysis of the functional neuroanatomy of single-word
lectura: Method and validation. NeuroImagen, 16(3), 765–780.
https://doi.org/10.1006/nimg.2002.1131, PubMed: 12169260
Turkeltaub, PAG. MI., Eickhoff, S. B., Laird, A. r., Fox, METRO., Wiener, METRO., &
Fox, PAG. (2012). Minimizing within-experiment and within-group
effects in activation likelihood estimation meta-analyses. Humano
Mapeo cerebral, 33(1), 1–13. https://doi.org/10.1002/hbm.21186,
PubMed: 21305667
tyler, l. K., Marslen-Wilson, w., & Stamatakis, mi. A. (2005). Disso-
ciating neuro-cognitive component processes: Voxel-based cor-
relational methodology. Neuropsicología, 43(5), 771–778.
https://doi.org/10.1016/j.neuropsychologia.2004.07.020,
PubMed: 15721189
Urgesi, C., Candidi, METRO., & Avenanti, A. (2014). Neuroanatomical
substrates of action perception and understanding: An anatomic
likelihood estimation meta-analysis of lesion-symptom mapping
studies in brain injured patients. Frontiers in Human Neurosci-
ence, 8, Article 344. https://doi.org/10.3389/fnhum.2014
.00344, PubMed: 24910603
Vigliocco, GRAMO., Vinson, D. PAG., Druks, J., Barber, h., & Cappa, S. F.
(2011). Nouns and verbs in the brain: A review of behavioural,
electrophysiological, neuropsychological and imaging studies.
Neurociencia & Revisiones de biocomportamiento, 35(3), 407–426.
https://doi.org/10.1016/j.neubiorev.2010.04.007, PubMed:
20451552
Caminante, GRAMO. METRO., Schwartz, METRO. F., Kimberg, D. y., Faseyitan, o.,
Brecher, A., Dell, GRAMO. S., & Coslett, h. B. (2011). Support for
anterior temporal involvement in semantic error production
in aphasia: New evidence from VLSM. Brain and Language,
117(3), 110–122. https://doi.org/10.1016/j.bandl.2010.09
.008, PubMed: 20961612
wilson, S. METRO., Henry, METRO. l., Besbris, METRO., Ogar, j. METRO., Dronkers,
norte. F., Jarrold, w., Molinero, B. l., & Gorno-Tempini, METRO. l. (2010).
Connected speech production in three variants of primary pro-
gressive aphasia. Cerebro, 133(7), 2069–2088. https://doi.org/10
.1093/brain/awq129, PubMed: 20542982
zhang, y., Kimberg, D. y., Coslett, h. B., Schwartz, METRO. F., & Wang,
z. (2014). Multivariate lesion-symptom mapping using support
vector regression. Mapeo del cerebro humano, 35(12), 5861–5876.
https://doi.org/10.1002/hbm.22590, PubMed: 25044213
zhao, y., Halai, A. D., & Lambon Ralph, METRO. A. (2020). Evaluating
the granularity and statistical structure of lesions and behaviour
in post-stroke aphasia. Brain Communications, 2(2), Article
fcaa062. https://doi.org/10.1093/ braincomms/fcaa062,
PubMed: 32954319
Neurobiology of Language
296
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
4
2
2
8
0
2
0
7
9
0
1
4
norte
oh
_
a
_
0
0
0
9
7
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3