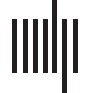ARTÍCULO DE INVESTIGACIÓN
Left Frontal White Matter Links to Rhythm
Processing Relevant to Speech Production in
Apraxia of Speech
, Manon Grube6,7
Rose Bruffaerts1,2,3,4
Silvy Gabel1, An-Sofie De Weer2, Eva Dries2, Karen Van Bouwel2, Timothy D. Griffiths6
, Jolien Schaeverbeke1
, Ahmed Radwan5
Stefan Sunaert5,8
, and Rik Vandenberghe1,2
1Laboratory for Cognitive Neurology, Department of Neurosciences & Leuven Brain Institute,
Katholieke Universiteit Leuven, Lovaina, Bélgica
2Neurology Department, University Hospitals Leuven, Lovaina, Bélgica
3Computational Neurology, Experimental Neurobiology Unit (ENU), Department of Biomedical Sciences,
University of Antwerp, Antwerp, Bélgica
4Biomedical Research Institute, Hasselt University, Diepenbeek, Bélgica
5Translational MRI, Department of Imaging and Pathology & Leuven Brain Institute,
Katholieke Universiteit Leuven, Lovaina, Bélgica
6Biosciences Institute, Medical School, Newcastle University, Newcastle-upon-Tyne, Reino Unido
7BIFOLD, Technische Universität Berlin, Alemania; Department of Psychology, Ashoka University, India
8Radiology Department, University Hospitals Leuven, Lovaina, Bélgica
Palabras clave: speech production, ritmo, apraxia of speech, structural MRI, psychoacoustics
,
,
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
ABSTRACTO
Recent mechanistic models argue for a key role of rhythm processing in both speech
production and speech perception. Patients with the non-fluent variant (NFV) of primary
progressive aphasia (PPA) with apraxia of speech (AOS) represent a specific study population
in which this link can be examined. Previously, we observed impaired rhythm processing in
NFV with AOS. We hypothesized that a shared neurocomputational mechanism structures
auditory input (sound and speech) and output (speech production) in time, a “temporal
scaffolding” mechanism. Since considerable white matter damage is observed in NFV, we test
here whether white matter changes are related to impaired rhythm processing. Forty-seven
participants performed a psychoacoustic test battery: 12 patients with NFV and AOS, 11
patients with the semantic variant of PPA, y 24 cognitively intact age- and education-
matched controls. Deformation-based morphometry was used to test whether white matter
volume correlated to rhythmic abilities. En 34 Participantes, we also obtained tract-based
metrics of the left Aslant tract, which is typically damaged in patients with NFV. Nine out
de 12 patients with NFV displayed impaired rhythmic processing. Left frontal white matter
atrophy adjacent to the supplementary motor area (SMA) correlated with poorer rhythmic
abilities. The structural integrity of the left Aslant tract also correlated with rhythmic abilities. A
colocalized and perhaps shared white matter substrate adjacent to the SMA is associated with
impaired rhythmic processing and motor speech impairment. Our results support the existence
of a temporal scaffolding mechanism structuring perceptual input and speech output.
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
un acceso abierto
diario
Citación: Bruffaerts, r., Schaeverbeke,
J., Radwan, A., Grube, METRO., Gabel, S., De
Weer, A.-S., Dries, MI., Van Bouwel, K.,
Griffiths, t. D., Sunaert, S., &
Vandenberghe, R. (2022). Left frontal
white matter links to rhythm
processing relevant to speech
production in apraxia of speech.
Neurobiology of Language, 3(4),
515–537. https://doi.org/10.1162/nol_a
_00075
DOI:
https://doi.org/10.1162/nol_a_00075
Supporting Information:
https://doi.org/10.1162/nol_a_00075
Recibió: 12 Puede 2021
Aceptado: 3 Junio 2022
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Rose Bruffaerts
rose.bruffaerts@uantwerpen.be
Editor de manejo:
Stefano Cappa
Derechos de autor: © 2022
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
Left frontal white matter & rhythm in PPA
Primary progressive aphasia (PPA):
A heterogeneous group of
neurodegenerative diseases
presenting with prominent
language deficits.
INTRODUCCIÓN
Recent neurophysiological evidence proposes a central role for rhythm processing in speech
production and speech perception (Poeppel & Assaneo, 2020). Aquí, we examine this link in
individuals with the non-fluent variant (NFV) of primary progressive aphasia (PPA) with apraxia
of speech (AOS). The rhythmicity of speech originates at the suprasegmental level as a result of
the metrical frame of words containing multiple segments or syllables (Aichert et al., 2016).
Speech rhythm also enhances perception: Speech perception is optimal between 2 y 8 Hz
(Ghitza & Greenberg, 2009), and the auditory cortex is tuned to these frequencies (Boemio
et al., 2005). The relationship between the neural processes supporting speech rhythm and
speech perception is under debate. Grube et al. (2012) have proposed a “temporal scaffolding
mechanism,” a common neurocomputational mechanism which structures both perceptual
input and speech production, in time. Específicamente, they observed that abilities to detect timing
differences in meaningless auditory stimuli correlated with phonological abilities (lectura, rep-
etition). This correlation between rhythm and phonological processing may reflect their rele-
vance in parsing the speech stream and extracting suprasegmental attributes. Patients with NFV
and AOS represent a unique opportunity to test the temporal scaffolding hypothesis because
impaired speech production co-occurs with a perceptual deficit in this population (Goll
et al., 2010; Grube et al., 2016). Clinically, NFV patients present with effortful, non-fluent
speech with simplified structures of their utterances (Grossman, 2018). A substantial number
of NFV cases also demonstrate AOS (Duffy et al., 2014), a motor speech disorder that leads
to phonetic errors, trial-and-error articulation (Ogar et al., 2007), slow speech rates, syllable
segmentation, and lengthened intersegment durations (Josephs et al., 2012). Vowel lengthening
is often observed in AOS (Ballard et al., 2014; Josephs et al., 2012). Typical AOS-related
changes in speech timing consist of relative lengthening of the first unstressed vowel compared
to the second stressed vowel (Duffy et al., 2017), impacting the suprasegmental level of speech
while often still respecting the representation of stress. Etiologically, AOS is caused by impaired
motor planning of movements for speech (Duffy et al., 2014; Grossman, 2018; Ogar et al.,
2007). Another motor speech impairment, dysarthria, is also observed in NFV with AOS in
an estimated 20–50% of patients (Duffy et al., 2014). The dysarthria co-occurring with AOS
is mainly of the spastic or hypokinetic type (Duffy et al., 2014). It is clear that the co-occurrence
of these two motor speech disorders (AOS and dysarthria) complicates the assessment of the
phenotype, as dysarthria in itself may impact neurolinguistic test scores and, especially in the
case of ataxic dysarthria, also the speech rhythm.
Co-occurring auditory perceptual abnormalities have been documented before in NFV with
AOS. Compared to controls and patients with the semantic variant (SV) of PPA, patients with
NFV were less able to discriminate between sounds with different spectral shapes (Goll et al.,
2010). Compared to controls and SV, NFV were also impaired at detecting changes in a rhyth-
mic pattern, which relies on the capacity to extract suprasegmental timing (Grube et al., 2016).
In a PPA cohort including 12 patients with NFV and AOS (Mesa 1) y 11 patients with SV
(Mesa 2), we here study the neuroanatomical correlate of impaired auditory rhythmic process-
ing in a search for supporting evidence for a temporal scaffolding mechanism. Grey matter
atrophy is consistently found in NFV in the left opercular part (BA44) of the inferior frontal
gyrus (IFG), insula, premotor and the supplementary motor areas (SMA) (Gorno-Tempini
et al., 2011; Rogalski et al., 2011). The degree of atrophy in these regions correlates with
markers of speech timing in patients with AOS (Ballard et al., 2014). SMA has been identified
as a gray matter correlate of auditory rhythmic processing in healthy volunteers (Grahn & Brett,
2007; Grahn & Schuit, 2012) and NFV (Hardy, Agustus, marshall, clark, Russell, Vínculo, et al.,
2017; Hardy, Agustus, marshall, clark, Russell, Brotherhood, et al., 2017). The correlational
Neurobiology of Language
516
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
Mesa 1.
Characteristics of NFV patients.
Case
Age
Sex
Educación
CPM (/36)
Dis. Dur.
BNT (/60)
AAT rep 1 (/30)
AAT rep 2 (/30)
AAT rep 3 (/30)
AAT rep 4 (/30)
AAT rep 5 (/30)
DS
DIAS cons (/15)
DIAS vow (/15)
DIAS dia
WEZT (/40)
Dysarthria
Extrapyr
CIT Spect
6
52
F
17
31
2
58
28
26
28
26
26
6
/
/
/
/
−
−
/
13
79
F
8
24
5
48
29
24
22
23
27
3
/
/
/
/
−
−
/
15
71
METRO
15
24
20*
78
METRO
17
30
21*
72
F
12
12
1.5
2.5
2.5
55
30
21
29
30
30
4
/
/
/
/
S
+
/
48
29
30
28
29
30
6
13
15
103
36
−
−
/
30
26
28
24
24
15
2
/
/
/
/
−
−
/
FDG-PET
SMA
L IFG L Frontal
L IFG
Nl
F
15
32
5
41
30
30
30
30
30
4
15
15
77
35
−
+
/
/
22*
63
23*
62
25
57
F
10
/
31*
69
METRO
18
29
2.5
3.5
1.5
F
12
31
46
28
30
30
29
28
5
10
14
50
36
S
+
/
Nl
/
/
/
29
20
30
29
29
28
7
13
14
115
31
h
+
/
7
27
29
28
15
17
3
14
14
24
29
−
+
+
/
36
58
F
12
21
4
52
28
28
28
28
23
5
3
15
48
20
−
−
/
37
80
F
10
4
4
27
14
25
27
14
10
3
0
6
6
11
SH
−
/
38
65
METRO
16
35
3
57
30
30
29
30
30
4
15
15
47
38
h
−
/
Cut-off
/
/
/
<28 >367
1320
1264
1139
<500 >367
<500 > 0.136.
Behavioral Testing
Confrontation naming was tested using the Boston Naming test with Dutch norms (Mariën
et al., 1998). Non-verbal executive functioning was evaluated using Raven’s Coloured Pro-
gressive Matrices. Speech repetition was assessed using the Akense Afasie test (Graets et al.,
1992). Note that AOS, agrammatism, and dysarthria were diagnosed clinically prior to enroll-
mento. To quantify the degree of AOS, the Dutch Diagnostic Instrument for AOS (Diagnostisch
Instrument voor Apraxie van de Spraak; DIAS; Feiken & Jonkers, 2012) was added when it
became available (for this reason it was not performed in 4 del 12 NFV cases). The DIAS
consists of vowel and consonant repetition (15 trials each) and diadochokinesis testing. During
the latter task, the examiner first reads three successive alternating syllables aloud, p.ej., “pa ta
ka” and asks the patient to repeat these. If successful, the patient is asked to repeat it as many
times as possible during a period of 8 s. The diadochokinesis severity score is the sum of correctly
repeated syllables across trials. To quantify grammaticality, we used a sentence comprehension
Cifra 1. Pure-tone audiograms of all participants. (A) Mean composite ear and frequency score (250–4000 Hz) data for each participant
grupo. (B) Mean thresholds (and standard error of the mean) for detection of tones at frequencies of 250, 500, 1000, 2000, y 4000 Hz para
each participant group. HC: healthy controls. NFV: participants with non-fluent variant of primary progressive aphasia. SV: participants with
semantic variant of primary progressive aphasia.
Neurobiology of Language
520
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
prueba, the Dutch verbs and sentences test ( Werkwoorden en Zinnen Test; WEZT; Bastiaanse &
Maas, 2000), consisting of 40 sentence-picture matching trials with active or passive sen-
tences containing possible role reversal (p.ej., “the horse was kicked by the cow”).
Psychoacoustic Tasks
Testing consisted of four pre-existing tasks (r1–r4; Grube et al., 2016) : the single time-interval
duration discrimination task (r1), the Isochrony deviation detection task (r2), and two metrical
pattern discrimination tasks (r3, r4). See Figure 2A and the additional audio files 1–4 in the
supplementary information. (Supporting Information can be found at https://doi.org/10.1162
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
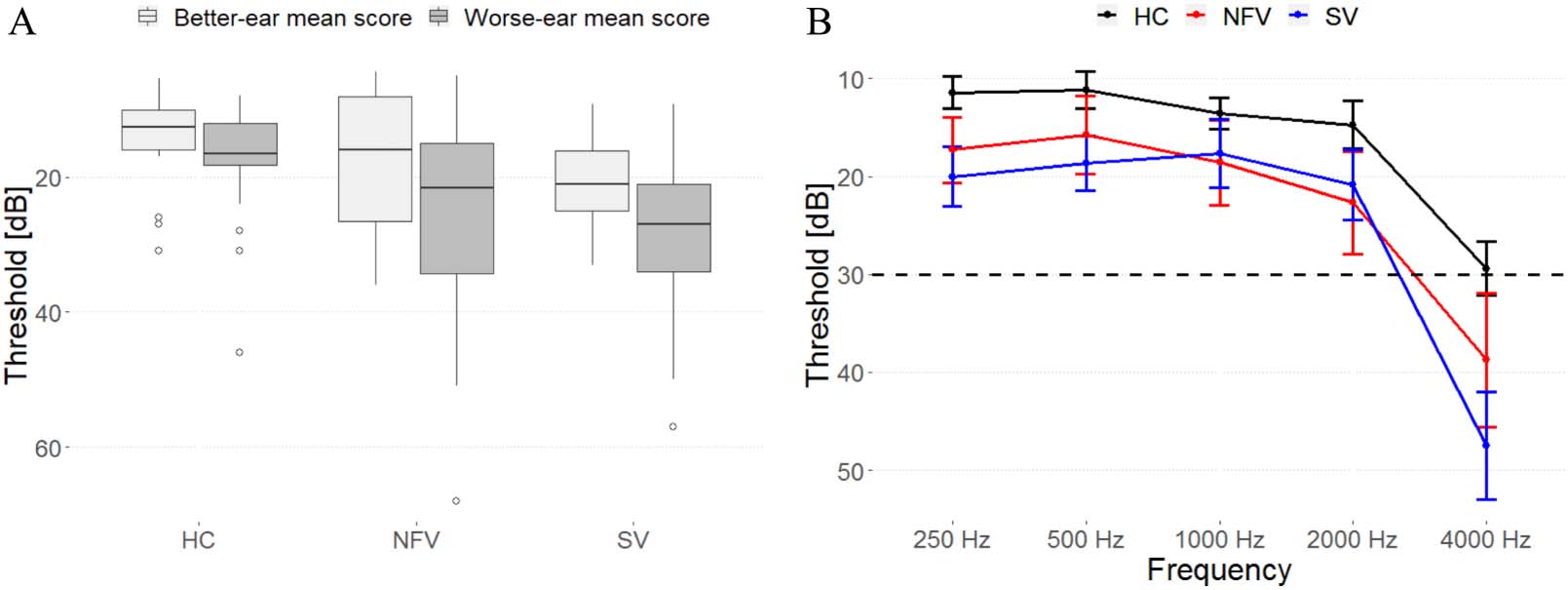
Cifra 2. Psychoacoustic tasks. (A) Diseño experimental: audio samples in supplementary information. (B) Mean raw thresholds (and standard
error of the mean) across controls, NFV, and SV patients. (C) Mean z scores (and standard error of the mean) across primary progressive aphasia
(PPA) subtypes. Dotted line represents the z cut-off for Bonferroni-corrected p < 0.05. (D) z scores for psychoacoustic tests in NFV (case num-
bers refer to Table 1). The size and color of dots reflect z score values; nonsignificant values are gray. Missing data indicates that the patient was
unable to perform the task. HC: healthy controls. NFV: participants with non-fluent variant of primary progressive aphasia. SV: participants with
semantic variant of PPA.
Neurobiology of Language
521
Left frontal white matter & rhythm in PPA
/nol_a_00075). The tasks followed a two-alternative forced-choice algorithm. Participants
responded verbally or by pointing to a graphical scheme. Instructions, verbal and graphic,
were repeated until the participant understood the task. Five practice trials were repeated until
five consecutive correct responses were recorded, and if needed, instructions were repeated
and the nature of the errors was explained. If the participant indicated during the test phase
that they had forgotten the instructions, then they were repeated, the practice trials run again,
and the test phase then restarted.
All tasks used 500 Hz 100 ms pure tones and consisted of 50 trials. The tasks were based on a
two-alternative forced-choice adaptive paradigm following a 2-down, 1-up algorithm (i.e., dif-
ficulty increasing after 2 consecutive correct responses and decreasing after every incorrect one).
A larger step size was used up to the fourth reversal and after that a smaller one. The outcome
measure was the threshold, calculated as the mean over the last six reversals measured with the
small step size, estimating the 70.9%-correct point of the psychometric function (Levitt, 1971).
The difference between the target and distractor was varied as a relative proportion of the dura-
tion or tempo of the reference. The Single time-interval duration discrimination task (r1) required
participants to indicate which of the two-tone pairs contained the “longer gap.” Initially, the
target was longer by 90% of the reference inter-onset-interval (depending on the trial, between
300 and 600 ms), and adaptively adjusted in steps of 12% and 6%. In the Isochrony deviation
detection task (r2), participants were instructed to indicate which of two otherwise isochronous
five-tone sequences contained a lengthening or “extra gap.” The reference sequence had an
isochronous inter-onset-interval ranging from 300 to 600 ms. The target had one lengthened
inter-onset-interval between the third and fourth tone. The initial default value of the lengthening
was 60% of the inter-onset-interval, adaptively adjusted in steps of 6% and 2%. In both tasks (r1,
r2), a local deviation is introduced to generate the target. As such, these tasks test the detection of
lower-order differences in timing between consecutive tones.
In the metrical pattern discrimination tasks (r3, r4), participants had to decide which of three
rhythmic sequences (the second or the third) of seven tones sounded “different,” based on a
distortion within the rhythm. The reference sequence had a strongly (r3) or a weakly (r4) met-
rical beat of four evoked by the temporal spacing of the tones over 16 time units. In the strongly
metrical sequence, accented tones occurred every four units, in the weakly metrical sequence,
two of those were silent (Grube & Griffiths, 2009). The default initial distortion in pattern (a
change in the long compared to the short intervals) was 65%, adaptively adjusted in steps of
12% and 6%. Metrical pattern discrimination (r3, r4) requires processing of the higher-order
temporal structure of the stimuli, since global deviations distributed across the sequence need
to be detected. Typical syllable rates in Dutch (the native language of the participants) are 4–5
syllables per second (period 200–250 ms), which is close to the tempi used in our tasks.
Statistical Analysis of Psychoacoustic Tasks
Depending on the distribution, outcome measures were log-transformed to allow for paramet-
ric analysis at the group level. At the individual level, each patient’s performance on the psy-
choacoustic tasks was analyzed in comparison to the group by using a modified Crawford
t test (Crawford & Garthwaite, 2007). For the comparison between each patient and the
controls, to facilitate comparison between tasks and to enable Bonferroni correction, the exact
p values (estimated percentiles) calculated according to Crawford and Garthwaite were trans-
formed into normalized z scores using the standard normal cumulative distribution function.
The significance threshold was set to a one-tailed significance level of p < 0.05, Bonferroni-
corrected for the number of tests (n = 4 for the psychoacoustic tasks, one-tailed since the a priori
Neurobiology of Language
522
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
n
o
_
a
_
0
0
0
7
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Left frontal white matter & rhythm in PPA
hypothesis is that NFV would perform worse). We compared the psychoacoustic thresholds
between NFV and SV using a Student’s t test (one-tailed significance level of p < 0.05, effect
size: Cohen’s d with Hedges correction for small samples).
Connected Speech Analysis
To obtain a tentative measure of the suprasegmental timing of speech, we determined the
normalized PVI for vowel nucleus duration in connected speech samples using Praat
6.1.02 (https://www.fon.hum.uva.nl/praat/). The samples consisted of a 2-minute “Cookie
Theft Scene” description (20 controls, 11 NFV, 9 SV, samples in Dutch). For every partici-
pant, the median PVI (Ballard et al., 2014) was determined for polysyllabic words with a
strong-weak stress pattern (e.g., COO-kie) and for words with a weak-strong stress pattern
(e.g., out-DA-ted). PVI was calculated following the procedure outlined in Ballard et al.
(2014) and Duffy et al. (2017), equaling 100 × (d1 − d2) / [(d1 + d2) / 2], where d1 and
d2 are the durations of the first and second vowel. False starts, which may occur in the
speech of NFV, were ignored for the purpose of the calculations. Normalization corrects for a
difference in speech rates. PVI values closer to zero are consistent with relatively equal stress
between the first two vowels of a word (Ballard et al., 2014). We also report absolute vowel
durations for each subgroup to provide more insight into the PVI values, as well as the results
per patient group, since PVI changes may be language-specific (Ballard et al., 2014). For NFV,
SV, and controls, we correlated PVI to the psychoacoustic tasks to test the link between auditory
rhythmic abilities and speech production (significance level of p < 0.05). In an exploratory
analysis, we also correlated the PVI metrics demonstrating a correlation to the psychoacous-
tics tasks to the white matter metrics.
Acquisition of MRI Data
Twenty-three patients (12 NFV, 11 SV) and 24 controls received a high resolution T1-weighted
structural MRI. All controls and 13 patients were scanned on a 3T Philips Intera system
equipped with an 8-channel receive-only head coil (SENSitivity Encoding head coil). Ten
patients were scanned on a 3T Philips Achieva dstream scanner equipped with a 32-channel
head volume coil. An identical 3D turbo field echo sequence was used on both systems (cor-
onal inversion recovery prepared 3D gradient-echo images, inversion time (TI) 900 ms, shot
interval = 3,000 ms, echo time (TE) = 4.6 ms, flip angle 8°, 182 slices, voxel size 0.98 ×
0.98 × 1.2 mm3). The diffusion weighted images consisted of 45 directions of diffusion weight-
ing with b = 800 as well as 1 non-diffusion weighted image (B0), acquired in the axial plane,
with isotropic voxel size of 2.2 mm, TR 9,900 ms, TE 90 ms, flip angle 90°, fold over direction
AP, fat shift direction A (anterior), in-plane parallel image acceleration (SENSE) factor 2.5.
Deformation-Based Morphometry
DBM was performed using the CAT12 toolbox (https://www.neuro.uni-jena.de/cat), an exten-
sion of SPM12 (https://www.fil.ion.ucl.ac.uk/spm). Segmentation was performed in CAT12
using a default tissue probability map. Local adaptive segmentation was used at default
strength (medium) and diffeomorphic anatomical registration through exponentiated lie alge-
bra (DARTEL) was used for registration to the default template (IXI555_MNI152). Voxel size for
normalized images was set at 1.5 mm (isotropic) after internal resampling at 1 mm. Local
deformations were estimated using the Jacobian determinant, while ignoring the affine part
of the deformation field. Thus, additional correction for total intracranial volume is not required
(Gaser & Kurth, 2019). Images were smoothed using a 8 × 8 × 8 mm3 Gaussian kernel. Jacobian
Neurobiology of Language
523
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
n
o
_
a
_
0
0
0
7
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Left frontal white matter & rhythm in PPA
maps of controls and both PPA groups were compared using a one-way between-subject
ANOVA. Multiple linear regression was used to correlate the psychoacoustic tasks (r1–r4) at
the individual level to the Jacobian maps within each PPA subtype. Scanner type and age were
introduced as nuisance variables in all analyses. Threshold of significance was set at voxel-
level uncorrected p < 0.001 and cluster-level family-wise error (FWE) corrected p < 0.05
(Grube et al., 2016; Poline et al., 1997). The resulting volume of interest was mirrored to
explore contralateral effects. To determine whether the observed DBM result was related to grey
or white matter involvement, a supplementary whole-brain VBM analysis using respectively
segmented grey and white matter maps was performed in which total intracranial volume
(TIV) was also added as a nuisance variable.
Diffusion Tensor Imaging
Diffusion images were preprocessed and analyzed using the KU Leuven neuroimaging suite
(KUL_NIS; Radwan & Sunaert, 2022), which relies on MRTrix3 (Tournier et al., 2012, 2019);
FSL (Jenkinson et al., 2012; Smith et al., 2004); and ANTs (Tustison et al., 2021). The prepro-
cessing pipeline included the following steps: First, the data were converted to the brain imag-
ing data structure convention (BIDS), then to MIF using mrconvert. Diffusion preprocessing
included denoising using dwidenoise (Veraart et al., 2016); echo planar imaging (EPI) distor-
tion correction was done using Synb0-DisCo (Schilling et al., 2019) and FSL topup (Andersson
et al., 2003). Correction for subject motion, and eddy current artefacts was performed using
MRTrix3 dwifslpreproc, which relies on FSL eddy (Andersson & Sotiropoulos, 2016).
This step was followed by imaging bias correction with dwibiascorrect. DTI (Basser et al.,
1994a, b) and CSD (Tournier et al., 2007) estimation was done using MRTrix3 dwi2tensor and
dwi2response. White matter response functions were averaged over the whole group and the
average response functions were used to calculate fiber orientation distribution (FOD) maps
for every subject using dwi2fod. FreeSurfer’s recon-all (Fischl, 2012), and the multi-scale brain
parcellator (Tourbier et al., 2020) were used for parcellating the 3D T1-weighted images,
based on the Desikan-Killiany atlas (Desikan et al., 2006) and the Lausanne parcellation
(Cammoun et al., 2012).
Next, we used the KU Leuven Fun-With-Tracts package (KUL_FWT; Radwan et al., 2022)
for calculating DTI scalar maps, and parcellation-based probabilistic CSD tractography using
second-order integration over orientation distributions (iFOD2; Tournier et al., 2010). The
bundle-specific approach of KUL_FWT was used for frontal Aslant tract tractography:
Tractograms were initially generated with 8,000 streamlines. We used the IFG pars trian-
gularis and pars opercularis labels from aparc+aseg maps generated by FreeSurfer, and the
superior frontal gyrus 5 & 6 labels from the scale-3 maps generated by the multi-scale
brain parcellator as the first and second inclusion regions, respectively. The whole corpus
callosum, brainstem, and orbitofrontal cortex labels from the aparc+aseg were used as
exclusion regions.
Smoothed fractional anisotropy (FA) and mean diffusivity (MD) maps were compared
between controls, NFV, and SV using a between-subject ANOVA (threshold same as
DBM/ VBM). Scanner type, TIV, and age were introduced as nuisance variables. A template
for the left Aslant tract was generated for healthy controls using the 75% overlap threshold
(Catani et al., 2013). FA and MD of the left Aslant tract were extracted for each patient by
averaging values from all voxels included in this template (Catani et al., 2013). We com-
pared the FA and MD between NFV and SV by means of a Student’s t test (one-tailed p <
Neurobiology of Language
524
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
n
o
_
a
_
0
0
0
7
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Left frontal white matter & rhythm in PPA
0.05). FA and MD were correlated to rhythmic abilities to confirm the DBM findings (one-
tailed p < 0.05).
RESULTS
Psychoacoustic Tasks
Two tasks (single time-interval duration discrimination (r1) and isochrony deviation detection
(r2); see Figure 2A and additional audio files 1–2) probed the detection of local deviations of
temporal structure. Two tasks (metrical pattern discrimination tasks using a strongly (r3) or
weakly (r4) metrical reference; see Figure 2A and additional audio files 3–4) probed the detec-
tion of higher-order deviations of temporal structure. The latter tasks (r3, r4) measure the
“rhythmic abilities” of each participant. Based on the suprasegmental abnormalities found
in the patients’ speech, we postulate that the tasks indexing suprasegmental timing (r3, r4) will
be most impaired in NFV with AOS.
In a group-level analysis, performance on the psychoacoustic tasks was poorer in NFV
compared to controls: Mean z scores were significantly increased (higher individual psycho-
acoustic thresholds equaling poorer abilities to detect deviations) in NFV for discrimination of
strongly metrical sequences (r3, mean z: 2.94), discrimination of weakly metrical sequences
(r4, mean: 2.93), and isochrony deviation detection (r2, mean: 2.46) (Figure 2B–C). We com-
pared the psychoacoustic test scores between the NFV and SV. This resulted in significantly
poorer scores in NFV for the discrimination of weakly metrical sequences (r4, p = 0.001,
Hedges’ g: 1.48) (Figure 2B–C). At the individual level, deficits were observed mainly in
NFV patients (z > 2.24) (Figura 2D). The weakly metrical pattern discrimination task (r4) dem-
onstrated a significant impairment in 7 NFV (Figura 2D) y 2 SV patients. Similarmente, strongly
metrical pattern discrimination (r3) was impaired in 6 NFV (Figura 2D) y 4 SV, así como
isochrony deviation detection (r2) en 6 NFV and 2 SV patients. Single time-interval discrimi-
nación (r1) was impaired in just 4 NFV and 1 SV patients. En resumen, 9/12 NFV cases and
4/11 SV cases were impaired in one or more of the tasks. No correlation was found between
better-ear hearing levels and psychoacoustic scores in patients (all ps > 0.220).
Correlation of Psychoacoustic Tasks With Speech Timing
To determine PVI as a proxy for suprasegmental timing of speech, vowel duration was first cal-
culated for all polysyllabic words with a strong-weak pattern (stress on the first syllable) y un
weak-strong pattern (stress on the second syllable). As expected in Dutch, participants gener-
ated more words with a strong-weak pattern compared to a weak-strong pattern (Figura 3A).
Lengthening of the first and second vowel (absolute duration) was observed for words with a
strong-weak pattern in NFV compared to controls (1st vowel: one-way ANOVA F(2, 37) = 4.17,
pag = 0.015; 2nd vowel: ANOVA F(2, 37) = 5.45, pag = 0.008; in both cases post hoc testing dem-
onstrated differences between the NFV with AOS and the control group; Figura 3B). Lengthen-
ing of the first vowel was observed for words with a weak-strong pattern in NFV compared to
controls (one-way ANOVA F(2, 35) = 5.11, pag = 0.011; Figura 3C). A trend was observed for
between-group differences of the duration of the second vowel in these words (one-way
ANOVA F(2, 35) = 2.9, pag = 0.068; Figura 3C).
PVI reflects the relative duration of the stressed versus unstressed syllable. PVI values were
closer to zero (relatively equal stress) for words with a weak-strong stress pattern in NFV com-
pared to controls and SV (one-way ANOVA F(2, 35) = 5.37, pag = 0.009; Figura 3D). No significant
between-group differences were found for strong-weak words. Across all participants, PVI for
Neurobiology of Language
525
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
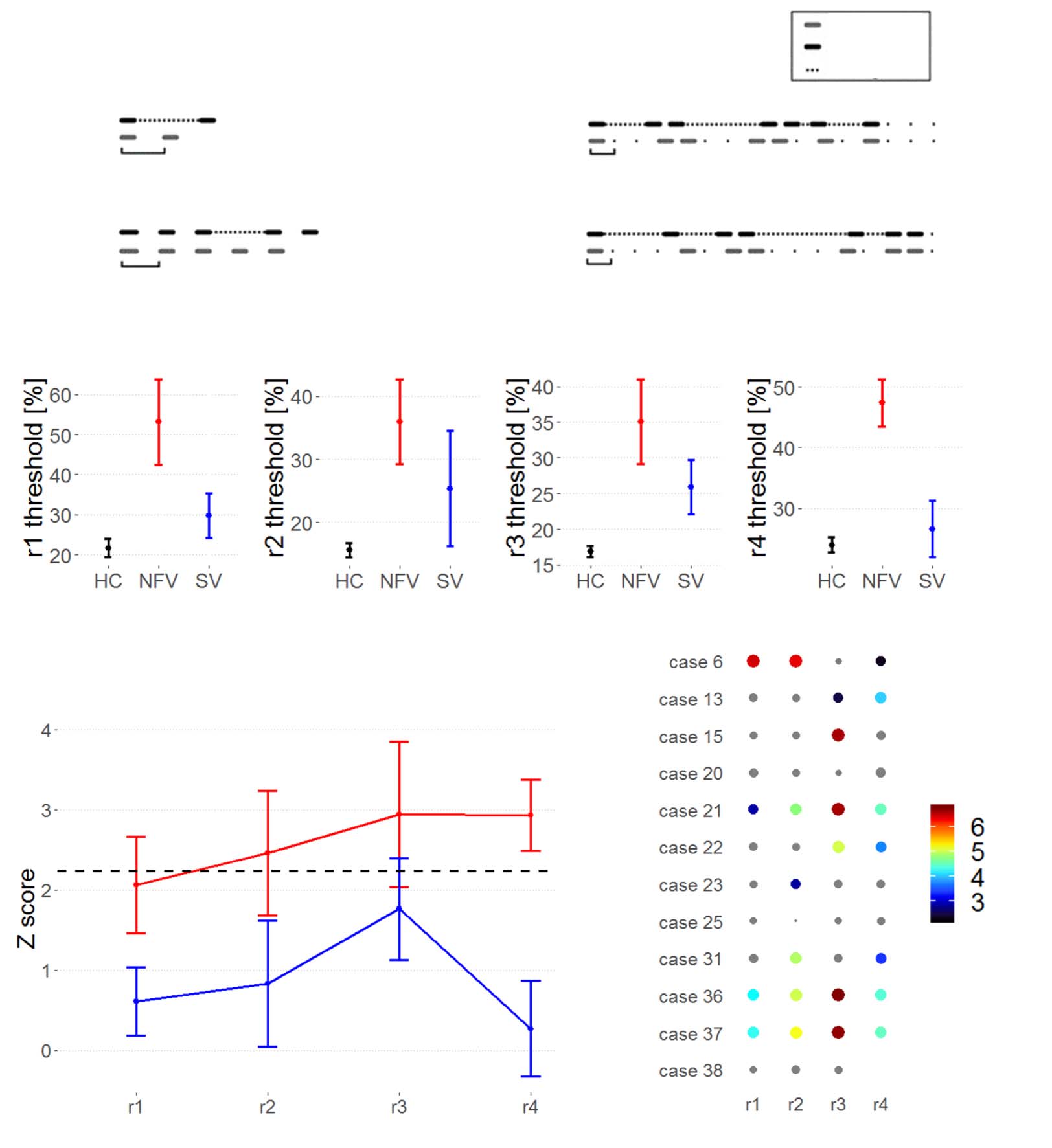
Cifra 3. Suprasegmental timing of speech. (A) Frequency of words with a strong-weak pattern and a weak-strong pattern in the three
participant groups. (B–C) Vowel durations for words with (B) a strong-weak pattern and (C) a weak-strong pattern. (D) PVI of words with a
strong-weak and a weak-strong pattern. (E–F) Correlation of PVI for (mi) strong-weak and (F) weak-strong words and strongly metrical
sequence thresholds (r3, log-transformed, %); regression line indicates a significant correlation at the subgroup level. PVI: pairwise variability
índice. HC = healthy controls. NFV: participants with non-fluent variant of primary progressive aphasia. SV: participants with semantic variant of
primary progressive aphasia.
strong-weak words correlated with the psychoacoustic threshold for strongly metrical pattern
discriminación (r3) (r= 0.444, pag = 0.004; Figure 6E). In an analysis per subgroup, this correlation
was found for NFV (r= 0.634, pag = 0.036), SV (r= 0.761, pag = 0.017), and controls (r= 0.531, pag =
0.016). This means that participants with poorer perceptual rhythmic abilities displayed greater
duration differences between the first and second vowels of words with a strong-weak stress
patrón. No correlation was found with any of the other psychoacoustic tasks (r1, r2, r4), y
no correlation was found with the PVI for weak-strong words (all p > 0.1, Figura 3F).
White Matter Changes: Deformation-Based Morphometry
In the SV group, atrophy was localized to the anterior temporal
A whole-brain DBM analysis was conducted to characterize the atrophy patterns for each PPA
group and to test whether rhythmic processing correlated with atrophy. The expected atrophy
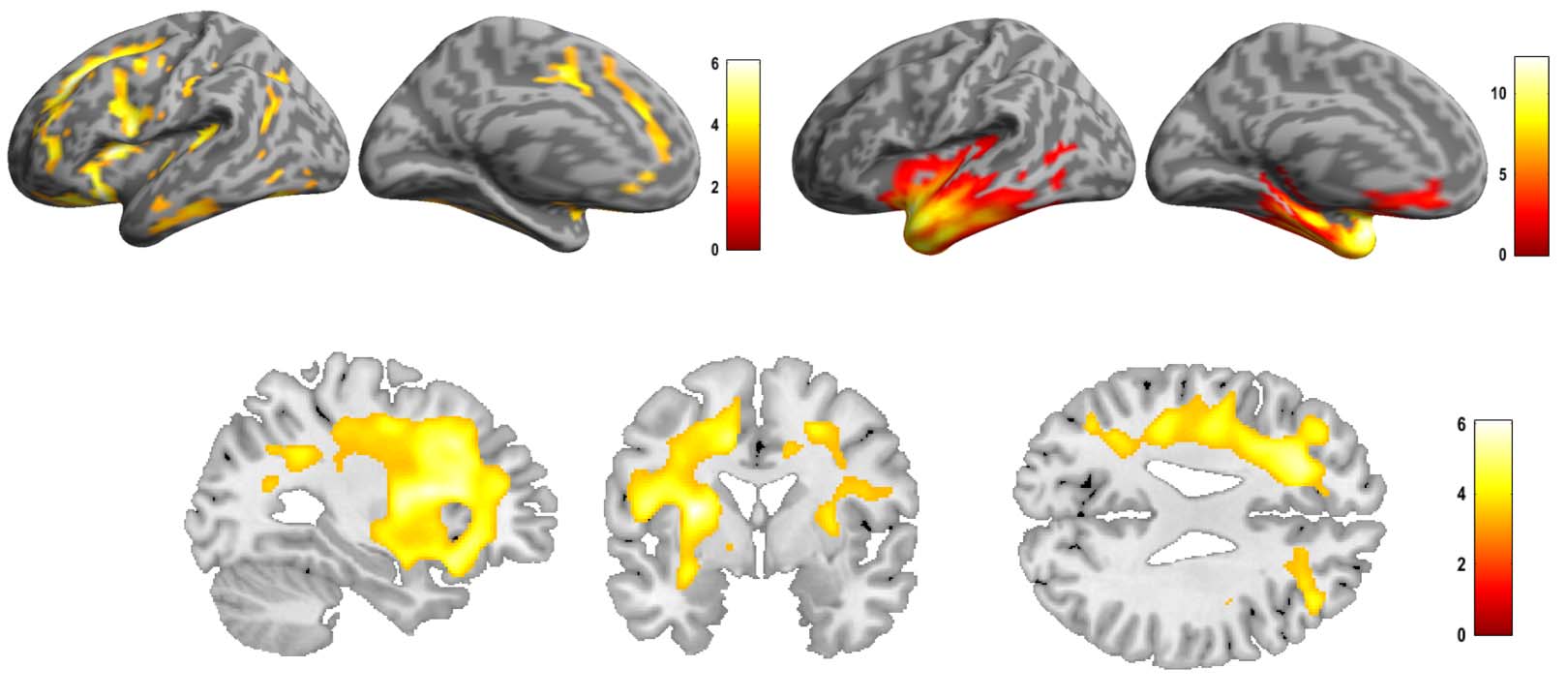
patterns per PPA subgroup were observed using DBM (Cifra 4). In the NFV group, atrophy
was observed mainly in the frontal lobes compared to controls, with a left-sided predominance
(Figure 4A–B).
lobes
(Figura 4A). In the NFV group, voxel-wise multiple linear regression showed that the strongly
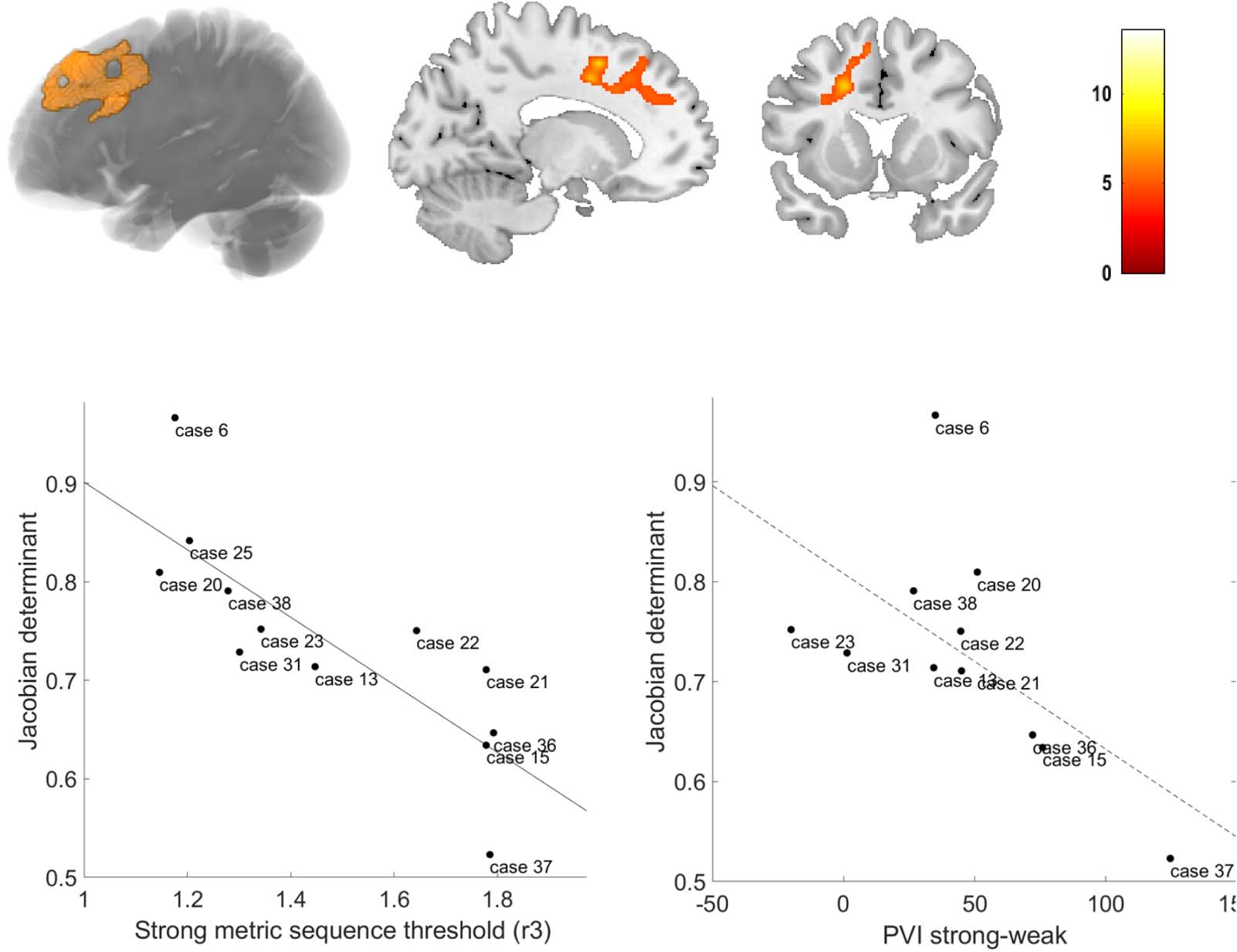
metrical rhythm discrimination task (r3) negatively correlated with volume changes in the left
frontal white matter (Instituto Neurológico de Montreal (MNI) = −20, 20, −36; −17, 8, 48; −9, 39,
50; kE 2,426 vóxeles, z score: 4.92) (Figure 5A–B). This negative correlation indicates that
poorer rhythmic abilities (es decir., larger psychoacoustic thresholds) were linked to more atrophy.
For illustrative purposes, we plotted the individual NFV thresholds for the strongly metrical
Neurobiology of Language
526
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
Cifra 4. Deformation-based morphometry (DBM) análisis: comparison of controls, the NFV, and SV patients. (A) Renderings shows atrophy
de 12 NFV and 11 SV compared to 24 controls (cluster-level FWE-corrected p < 0.05). (B) Slices in NFV. NFV: participants with non-fluent
variant of primary progressive aphasia. SV: participants with semantic variant of primary progressive aphasia.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
n
o
_
a
_
0
0
0
7
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Deformation-based morphometry (DBM) of rhythmic abilities in NFV cases. (A) Rendering of the DBM result (orange) projected
onto structural template. (B) Slices. (C) Correlation between volume loss and strongly metrical sequence thresholds (r3, log-transformed, %) in
NFV patients in the region of interest (A–B), exploratory plot for illustrative purposes. (D) Correlation between volume loss and PVI for words
with a strong-weak pattern in NFV patients in the region of interest (A–B) (dotted regression line since there was a trend for significance). Case
numbers refer to Table 1. PVI: pairwise variability index.
Neurobiology of Language
527
Left frontal white matter & rhythm in PPA
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
n
o
_
a
_
0
0
0
7
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
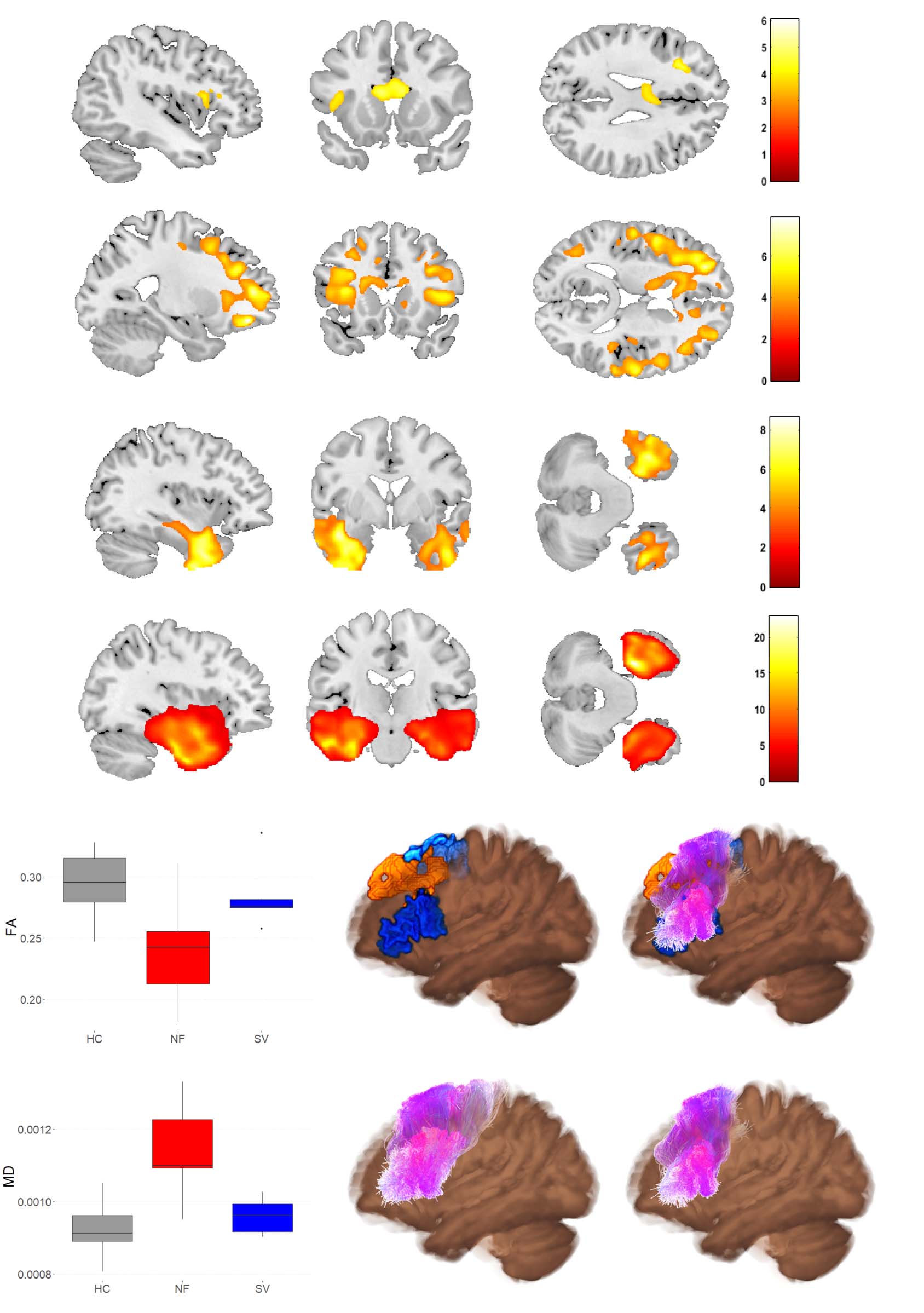
Figure 6. Diffusion tensor imaging (DTI) metrics. (A) Fractional anisotropy (FA) and (B) Mean diffusivity (MD) in NFV patients versus controls
(cluster-level FWE-corrected p < 0.05). (C) FA and (D) MD in SV patients versus controls (cluster-level FWE-corrected p < 0.05). Comparison of
(E) FA and (F) MD in the left frontal Aslant tract between the NFV and SV subtypes and healthy controls (HC). (G) 3D visualization of the left
Aslant tract projected on the average FA map—Top left: seed regions (blue) and result of deformation-based morphometry (DBM) (orange); Top
right: seed regions (blue) and result of DBM (orange) combined with Aslant tract streamlines of NFV patients (purple); Bottom left: streamlines
of controls (purple); Bottom right: streamlines of NFV patients (purple). ROI: region of interest. HC: healthy controls. NFV: participants with
non-fluent variant of primary progressive aphasia. SV: participants with semantic variant of primary progressive aphasia.
Neurobiology of Language
528
Left frontal white matter & rhythm in PPA
discrimination task (r3) versus volume loss in this region (r = −0.316, p < 0.001) (Figure 5C).
The correlation between the PVI for words with a strong-weak pattern and volume loss in this
region showed a trend for significance (r = −0.600, p = 0.051) (Figure 5D). No correlation was
observed with the strongly metrical discrimination task (r3) in the right-sided homologue vol-
ume of interest ( VOI) (r = −0.347, p = 0.267). Finally, DBM analysis yielded no other signif-
icant correlations with the psychoacoustic tasks in the NFV or SV subgroups. The additional
VBM analysis using segmented white matter maps of the NFV patients confirmed that the loss
of volume was related to white matter changes: When correlating the strongly metrical dis-
crimination task (r3) with white matter intensity, 2 clusters survived the preset significance
threshold and the largest cluster (peak coordinates MNI = −26, 18, 30; kE 232 voxels, z =
3.74) overlapped with the result of the DBM analysis (Figure 5A–B). In the VBM analysis using
grey matter maps, no clusters survived the preset threshold.
White Matter Changes: Diffusion Tensor Imaging
A whole-brain comparison between NFV, SV, and controls showed reduced FA in NFV in the
left inferior frontal region, the corpus callosum, and the anterior cingulate (Figure 6A). MD was
widely increased in NFV, with a predominance in both frontal lobes (Figure 6B). In SV, FA was
reduced and MD was increased in both anterior temporal lobes (Figure 6C–D). We then com-
pared FA and MD between NFV and SV specifically within the template of the left Aslant tract
derived from the controls. FA was lower in NFV compared to SV ( p = 0.038, Hedges’ g: −1.17;
Figure 6E), and MD was higher in NFV compared to SV ( p = 0.003, Hedges’ g: 1.85; Figure 6F).
Across all participants, FA and MD in the left Aslant tract correlated to the performance on the
strongly metrical rhythm discrimination task (r3) (FA: r = −0.535, p = 0.001; MD: r = 0.652, p <
0.001). This means that FA was lower and MD was higher when performance on the metrical
rhythm discrimination task (r3) was weaker. Within the relatively small NFV group, a trend in
the expected direction was observed when correlating FA and MD in the left Aslant tract with
the performance on the strongly metrical rhythm discrimination task (r3) (FA: r = −0.670, p =
0.099; MD: r = 0.745, p = 0.053). In the contralateral homologue VOI, no correlation was
observed (FA: r = −0.517, p = 0.235; MD: r = 0.245, p = 0.597). Neither FA nor MD in the
left Aslant tract correlated with performance on any other psychoacoustic task in NFV (r1, r2,
r4, all ps > 0.271), but there was a correlation between the DTI metrics and PVI for words with a
strong-weak pattern (FA: r = −0.791, pag = 0.034; Maryland: r= 0.905, pag = 0.005). Visual inspection of
the left Aslant tract in NFV showed that this tract overlapped with the region where there were
white matter volume changes identified by DBM (Figure 6G).
DISCUSIÓN
In patients with NFV and AOS, we investigated the neural correlate of impaired auditory rhyth-
mic processing. In a group of 12 consecutive cases with NFV, we confirmed that rhythmic
abilities are overall poorer compared to controls and patients with SV (Grube et al., 2016).
Behaviorally, we observed a correlation between rhythmic abilities and a marker for the supra-
segmental timing of speech for NFV as well as for SV and healthy controls, in agreement with a
coupling between auditory perception and speech production. DBM demonstrated that atro-
phy in the left frontal lobe correlated with the individual patients’ rhythmic abilities. We com-
plemented DBM with DTI to provide an independent measure of white matter changes. DTI
confirmed a correlation between the left Aslant tract MD and FA and rhythmic abilities. Given
the prior work implicating the left Aslant tract in non-fluent speech and AOS (Canu et al.,
2019; Catani et al., 2013; Mandelli et al., 2014), our results indicate it may be a part of
common anatomical substrate for rhythmic abilities and speech production. Whilst our
Neurobiology of Language
529
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
findings are correlational, the results for speech timing complement the two independent
white matter metrics. Our findings strengthen the evidence base for a well-defined neurocom-
putational mechanism of temporal scaffolding linking perception and speech production. En
NFV, the concept of impaired temporal scaffolding ties both core auditory deficits and AOS to
left frontal atrophy.
Although four psychoacoustic tasks were performed, both the marker for the suprasegmental
timing of speech and left frontal lobe atrophy in NFV were linked to impaired performance on
the strongly metrical rhythm discrimination task (r3). This task is conceptually different from the
single time-interval duration discrimination task (r1) and the isochrony deviation detection task
(r2): Determining the metricality of a tone sequence (r3) requires processing of the higher-order
temporal structure determined by the grouping of salvos of notes that induce the sense of a
regularly occurring metrical “beat” (Grube & Griffiths, 2009). Metricality-based rhythm discrim-
ination (r3) necessitates detecting global deviations distributed across the entire sequence. Nuestro
results are in agreement with prior work: The detection of temporal changes between syllables
was more impaired in PPA compared to controls when stimuli contained a higher number of
syllables (Rohrer et al., 2012). A diferencia de, the single time-interval duration discrimination task
(r1) and the isochrony deviation detection task (r2) test lower-order differences in timing in a
simple isochronous sequence based on a local deviation. The weakly metrical rhythm discrim-
ination task (r4) is more challenging as it does not rely on a clear metrical beat (Grube & Griffiths,
2009) (higher thresholds for r4 versus r3 in controls, pag < 0.001). Perhaps domain-general pro-
cesses play a larger role in this task compared to the strongly metrical rhythm discrimination task
(r3), but additional manipulations are required to support this hypothesis. Specifically, the rela-
tion between rhythm processing and working memory deficits in NFV should be investigated
more thoroughly using tasks indexing auditory working memory, phonological working mem-
ory, and working memory tasks not grounded in the auditory modality. Decreased auditory
working memory (measured by digit span) is usually observed in NFV and has been linked to
agrammatism (Rohrer et al., 2010). In our NFV cases, we did not find a strong correlation
between metrical pattern discrimination (r3, r4) and digit span (both p > 0.093). The prior musi-
cal experience of the individual participants may also have influenced their performance on the
psychoacoustic tasks. Keeping in mind the labor-intensive administration of our tasks, nosotros
reached a considerable albeit modest sample size. We acknowledge that further validation of
our findings requires a larger multicenter sample given the relative rarity of PPA.
Using two independent white matter metrics, we identified an overlapping white matter sub-
strate in the left frontal lobe that may play a role in both rhythmic abilities and speech produc-
tion and is thus a candidate component of the temporal scaffolding mechanism. We observed
white matter degeneration close to the SMA. The Aslant tract connects the superior frontal
gyrus/SMA to the IFG, cortical regions that have previously been implicated in rhythm process-
En g (Poeppel & Assaneo, 2020). SMA was linked to auditory rhythmic processing (Hardy, Agustus,
marshall, clark, Russell, Vínculo, et al., 2017) and AOS (Tetzloff et al., 2018; Whitwell et al.,
2013) to NFV. IFG plays a role in speech rhythm (Long et al., 2016) and synchronizing speech
to external auditory stimuli (Assaneo et al., 2019). The correlation between rhythmic abilities
and white matter integrity in the left frontal lobe aligns with the contemporary view that speech
rhythm production and perception are sustained by a left hemispheric network rather than a
single cortical region (Mandelli et al., 2016; Poeppel & Assaneo, 2020). Besides the Aslant tract,
other white matter tracts, p.ej., tracts connecting to the auditory cortices (Assaneo et al., 2019),
may also play an important role in this network. Respectivamente, the DBM analysis also demon-
strated that the region in which atrophy correlated to rhythmic abilities extends beyond the
boundaries of the left Aslant tract. While we focus here on white matter damage, the existing
Neurobiology of Language
530
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
literature suggests that cortical damage also contributes to abnormal speech rhythm production
and perception in NFV (Hardy, Agustus, marshall, clark, Russell, Vínculo, et al., 2017). Mientras
VBM did not detect a correlation between cortical atrophy and rhythmic abilities in our NFV
grupo, this does not preclude functional abnormalities at the cortical level. The unique contri-
butions of grey and white matter to rhythmic processing require further study in a larger cohort.
An important consideration is whether the white matter changes reflect tau pathology,
which is found in up to 88% of NFV patients (Spinelli et al., 2017). En particular, DTI metrics
have been put forward as a marker of tauopathy and other proteinopathies (Downey et al.,
2015; Mahoney et al., 2013). DTI imaging is sensitive to changes caused by tau pathology
at the single-subject level (Sajjadi et al., 2013), presumably because of underlying glial pathol-
ogia (Forman et al., 2002), p.ej., by myelin injury or changes in other structures that affect water
difusión (Galantucci et al., 2011). Aquí, we performed DTI imaging in a small subset of the
study population (7 NFV, 7 SV, y 20 controls) and because of the small sample size, nosotros
interpret these preliminary findings with appropriate caution. Diffusion MRI and fiber tracto-
graphy are known to have a number of methodological drawbacks: The ones most relevant to
our work are volume averaging and lack of accuracy in voxels with complex fiber architecture
such as the capsules (internal, externo. and extreme), as well as the centrum semiovale. Nosotros
attempted to maximize the accuracy of voxel-wise comparisons by relying on CSD iFOD2
tractography, and by using streamline filtering to minimize false positive streamlines. Our pre-
liminary DTI results are in alignment with prior work in PPA (Powers et al., 2013): MD changes
were more pronounced than FA changes in NFV patients (Whitwell et al., 2010). In our study,
neuropathological data is lacking for most patients, thus prohibiting us from making strong
claims in relation to pathology. We would not advocate linking tauopathy to a simple DTI
parameter. Bastante, our findings advance the broader characterization of the possible
disease-specific involvement of white matter tracts. Our results align with the “molecular
nexopathy” paradigm (Warren et al., 2013): The left frontal network containing IFG and
SMA as nodes demonstrates a selective vulnerability to tau protein, which could spread locally
through the left Aslant tract in a prion-like fashion. Even if certain proteinopathies are strongly
linked to predictable phenotypes of network disruption, the molecular nexopathy paradigm
does not propose complete specificity. Respectivamente, we also observed impaired perceptual
rhythmic abilities in some SV patients. As the disease progresses, PPA subtypes exhibit con-
vergence of their atrophy patterns (Bruffaerts et al., 2020; Leyton et al., 2019), even though the
underlying neuropathology is different. This convergence of atrophy patterns may explain that
some SV demonstrated impaired rhythmic abilities.
In an attempt to understand how impaired auditory rhythmic processing relates to AOS, nosotros
correlated each participant’s rhythmic abilities to a suprasegmental marker of speech timing,
the PVI. Curiosamente, we observed a correlation between speech timing for strong-weak words
and metrical pattern discrimination ability in all three participant groups. Such a coupling
aligns with the temporal scaffolding hypothesis because it supports the idea that both speech
production and perception tap into a common neural mechanism sustaining rhythm process-
ing at the input and output level. The fact that the coupling even holds in NFV patients further
supports this notion. At the segmental level, we observed that the first and second vowel dura-
tion are both longer in words with a strong-weak pattern in NFV compared to controls. Cómo-
alguna vez, this did not lead to significant changes to the speech rhythm properties in the relative
sense at the suprasegmental level, which replicates the findings of Vergis et al. (2014). Como
the dominant rhythmic structure, the production of words with a strong-weak pattern appears
to be relatively spared in AOS in German (Aichert et al., 2016) e inglés (Bailey et al.,
2019). We showed here that this is also the case in Dutch. We moreover observed a positive
Neurobiology of Language
531
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
correlation between the suprasegmental timing of speech (reflecting the contrast between the
length of the first stressed versus second unstressed vowel) and rhythmic abilities in all groups
for strong-weak words. Hipotéticamente, increased contrast in these spoken words co-occurring
with poorer rhythmic abilities may be interpreted as a “coarser” or more un-even resolution of
the temporal scaffolding system and may give rise to an overall slower, and less fluent produc-
tion of speech. En otras palabras, the more accurate the system, the more even the syllables are
being produced in words with strong-weak pattern, while pathology results in abnormal
lengthening of the first, strong syllable. For words with a weak-strong pattern, we replicated
the lower PVI values typically observed in AOS (Ballard et al., 2014; Duffy et al., 2017) y
also observed abnormal lengthening of the first vowel in these words. Our behavioral findings
cannot explain the full extent of speech abnormalities in AOS and evidence the need for auto-
acoplado, broadly validated speech-based markers in PPA.
We opted here to study the PVI for duration because of its intuitive link to suprasegmental
timing, but it is clearly an imperfect marker of AOS and speech rhythm. Alternative markers (p.ej.,
PVI for intensity/fundamental frequency, measures of silence duration or variability) may provide
additional information. We acknowledge that we could not in our analysis take into account the
position of the word in the sentence or other word characteristics such as word length, cual
may also influence the timing of speech (Aichert et al., 2016; Bailey et al., 2019; Vergis et al.,
2014). Additional variability was possibly introduced by using connected speech samples
resulting in different outputs across participants, while other studies used tasks that elicited
an identical response from all participants. The presence of moderate dysarthria (with impact
on intelligibility) en 3 out of 12 NFV cases may have also affected our marker of speech timing
(Duffy et al., 2017). While PVI is a sensitive measure to identify AOS in NFV (Ballard et al.,
2016), PVI should be considered an aspecific marker of motor speech impairment. Por eso,
when the PVI is abnormal in a patient with both dysarthria and AOS, it is not straightforward
to pinpoint which motor speech abnormality drives the changes in the PVI (Duffy et al., 2017).
Idealmente, AOS should be studied using the PVI in patients without co-occurring dysarthria.
Whilst our results can thus not be conclusive, they build on and expand findings from the
existing literature on AOS.
Finalmente, we acknowledge that even though all our NFV cases exhibited clear AOS, the clinical
picture included other symptoms often observed in NFV such as agrammatism, dysarthria, o
extrapyramidal signs. We cannot quantify how these findings relate to the observed white matter
cambios. Idealmente, our results should be replicated in patients with isolated AOS, por ejemplo, en
patients with primary progressive AOS (ppAOS) (Josephs et al., 2012), but this phenotype is even
more rare than NFV (Duffy et al., 2021). Neuroimaging analyses of patients with ppAOS are in
agreement with our results: Reduced FA was observed in the left SMA in ppAOS (Utianski et al.,
2018), and functional connectivity analysis demonstrated that SMA is disconnected from the
speech and language network in ppAOS (Botha et al., 2018). The atrophy pattern germane to
ppAOS differs from the atrophy pattern reported in NFV without AOS, with bilateral volume loss
more anteriorly in the frontal lobes compared to ppAOS and also in the left temporal lobe
(Tetzloff et al., 2019). Por último, an important non-neurodegenerative cause of AOS is stroke. Lesion
mapping in stroke patients with AOS demonstrated maximal overlap in the left (pre)motor cortices
and adjacent white matter (Graff-Radford et al., 2014), which coincides with our neuroimaging
findings in white matter. Además, using resting-state fMRI abnormal functional connectivity
of the left premotor cortex was observed in stroke patients with AOS compared to those without
AOS (New et al., 2015), indicative of white matter damage in the left frontal lobe in the patients
with AOS. En resumen, the regions identified as neural correlates of rhythmic processing in our
study are consistent with prior neuroimaging studies on the neural correlates of AOS.
Neurobiology of Language
532
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
Conclusions
Co-occurring impaired rhythmic abilities and AOS in NFV prompted our search for the neu-
roanatomical correlate of rhythmic processing in NFV. Our DBM and DTI findings in NFV
showed concordant evidence that rhythmic abilities correlate with left frontal white matter
atrophy, overlapping with the substrate for AOS. En general, we provided initial evidence for a
common neurocomputational mechanism for speech production and perception with its neu-
ral basis located in the left frontal lobe. Future studies should target larger multicentric patient
cohorts with pure AOS to characterize this mechanism in greater detail. A better understand-
ing of the neurobiological link between speech production and perception may contribute to
the development of tailored rehabilitation strategies (Ballard et al., 2015).
EXPRESIONES DE GRATITUD
The authors thank Bruno Bergmans, Maryland, PhD, Charlotte Swinnen, Maryland, Anne Sieben, Maryland,
PhD, and Yolande A. Pijnenburg, Maryland, PhD, for the referral of patients. We thank Emma
Luckett, MSc, for copyediting. RB is a senior and JS is a junior postdoctoral fellow of the
Research Foundation Flanders (FWO).
INFORMACIÓN DE FINANCIACIÓN
Rose Bruffaerts, Fonds Wetenschappelijk Onderzoek (https://dx.doi.org/10.13039
/501100003130), Award ID: 12I2121N. Rik Vandenberghe, Fonds Wetenschappelijk
Onderzoek (https://dx.doi.org/10.13039/501100003130), Award ID: G0925.15. Rik Vandenberghe,
Belgian Federal Science Policy Office (https://dx.doi.org/10.13039/501100002749), Award
ID: 7/11. Rik Vandenberghe, Onderzoeksraad, KU Lovaina (https://dx.doi.org/10.13039
/501100004497), Award ID: OT/12/097, C14/17/108.
CONTRIBUCIONES DE AUTOR
Rose Bruffaerts: Conceptualización: Equal; Curación de datos: Equal; Análisis formal: Lead;
Adquisición de financiación: Equal; Investigación: Equal; Metodología: Equal; Administración de proyecto:
Equal; Supervisión: Equal; Validación: Equal; Visualización: Lead; Escritura – borrador original:
Lead. Jolien Schaeverbeke: Curación de datos: Equal; Análisis formal: Secundario; Investigación:
Equal; Administración de proyecto: Equal; Validación: Equal; Escritura – revisión & edición: Equal.
Ahmed Radwan: Análisis formal: Equal; Metodología: Equal; Software: Equal; Visualización:
Secundario; Escritura – revisión & edición: Equal. Manon Grube: Conceptualización: Equal; Para-
mal analysis: Secundario; Metodología: Equal; Escritura – revisión & edición: Equal. Silvy Gabel:
Curación de datos: Secundario; Análisis formal: Secundario; Investigación: Equal; Project admin-
istración: Secundario; Escritura – revisión & edición: Secundario. An-Sofie De Weer: Data cura-
ción: Secundario; Análisis formal: Secundario; Investigación: Equal; Administración de proyecto:
Secundario; Escritura – revisión & edición: Secundario. Eva Dries: Curación de datos: Secundario;
Análisis formal: Secundario; Investigación: Equal; Escritura – revisión & edición: Secundario.
Karen Van Bouwel: Curación de datos: Secundario; Análisis formal: Secundario; Investigación:
Equal; Escritura – revisión & edición: Secundario. Timothy D. Griffiths: Conceptualización:
Equal; Metodología: Equal; Escritura – revisión & edición: Equal. Stefan Sunaert: Formal anal-
ysis: Equal; Metodología: Equal; Recursos: Equal; Software: Equal; Escritura – revisión & edit-
En g: Equal. Rik Vandenberghe: Conceptualización: Equal; Curación de datos: Secundario; Fondos
adquisición: Equal; Administración de proyecto: Equal; Recursos: Equal; Supervisión: Equal; Vali-
dación: Equal; Escritura – revisión & edición: Equal.
Neurobiology of Language
533
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
REFERENCIAS
Aichert, I., Späth, METRO., & Ziegler, W.. (2016). The role of metrical
information in apraxia of speech. Perceptual and acoustic analy-
ses of word stress. Neuropsicología, 82, 171–178. https://doi.org
/10.1016/j.neuropsychologia.2016.01.009, PubMed: 26792367
andersson, j. l. r., Skare, S., & Ashburner, j. (2003). How to correct
susceptibility distortions in spin-echo echo-planar images: Application
to diffusion tensor imaging. NeuroImagen, 20(2), 870–888. https://
doi.org/10.1016/S1053-8119(03)00336-7, PubMed: 14568458
andersson, j. l. r., & Sotiropoulos, S. norte. (2016). Un integrado
approach to correction for off-resonance effects and subject move-
ment in diffusion MR imaging. NeuroImagen, 125, 1063–1078.
https://doi.org/10.1016/j.neuroimage.2015.10.019, PubMed:
26481672
Assaneo, METRO. F., & Poeppel, D. (2018). The coupling between audi-
tory and motor cortices is rate-restricted: Evidence for an intrinsic
speech-motor rhythm. Science Advances, 4(2), Article eaao3842.
https://doi.org/10.1126/sciadv.aao3842, PubMed: 29441362
Assaneo, METRO. F., Ripollés, PAG., Orpella, J., lin, W.. METRO., de Diego-Balaguer,
r., & Poeppel, D. (2019). Spontaneous synchronization to speech
reveals neural mechanisms facilitating language learning. Naturaleza
Neurociencia, 22(4), 627–632. https://doi.org/10.1038/s41593
-019-0353-z, PubMed: 30833700
Bailey, D. J., Bunker, l., Mauszycki, S., & Wambaugh, j. l. (2019).
Reliability and stability of the metrical stress effect on segmental
production accuracy in persons with apraxia of speech. interna-
tional Journal of Language & Communication Disorders, 54(6),
902–913. https://doi.org/10.1111/1460-6984.12493, PubMed:
31338954
Ballard, k. J., Azizi, l., Duffy, j. r., McNeil, METRO. r., Halaki, METRO.,
O’Dwyer, NORTE., Layfield, C., Scholl, D. I., Vogel, A. PAG., & Robin,
D. A. (2016). A predictive model for diagnosing stroke-related
apraxia of speech. Neuropsicología, 81, 129–139. https://doi.org
/10.1016/j.neuropsychologia.2015.12.010, PubMed: 26707715
Ballard, k. J., Savage, S., Leyton, C. MI., Vogel, A. PAG., Hornberger, METRO.,
& Hodges, j. R. (2014). Logopenic and nonfluent variants of pri-
mary progressive aphasia are differentiated by acoustic measures
of speech production. MÁS UNO, 9(2), Article e89864. https://
doi.org/10.1371/journal.pone.0089864, PubMed: 24587083
Ballard, k. J., Wambaugh, j. l., Duffy, j. r., Layfield, C., Maas, MI.,
Mauszycki, S., & McNeil, METRO. R. (2015). Treatment for acquired
apraxia of speech: A systematic review of intervention research
entre 2004 y 2012. American Journal of Speech-Language
Pathology, 24(2), 316–337. https://doi.org/10.1044/2015_AJSLP
-14-0118, PubMed: 25815778
Bajos, PAG. J., mattiello, J., & LeBihan, D. (1994a). Estimation of the
effective self-diffusion Tensor from the NMR spin echo. Diario de
Resonancia magnetica, Serie B, 103(3), 247–254. https://doi.org
/10.1006/jmrb.1994.1037, PubMed: 8019776
Bajos, PAG. J., mattiello, J., & LeBihan, D. (1994b). MR diffusion
tensor spectroscopy and imaging. Revista biofísica, 66(1),
259–267. https://doi.org/10.1016/S0006-3495(94)80775-1,
PubMed: 8130344
Bastiaanse, r., & Maas, mi. (2000). Werkwoorden en zinnentest
[Verb and sentence test] WEZT. Swets & Zeitlinger.
Boemio, A., Fromm, S., Braun, A., & Poeppel, D. (2005). Hierarchi-
cal and asymmetric temporal sensitivity in human auditory cor-
tices. Neurociencia de la naturaleza, 8(3), 389–395. https://doi.org/10
.1038/nn1409, PubMed: 15723061
Botha, h., Utianski, R. l., Whitwell, j. l., Duffy, j. r., clark, h. METRO.,
Strand, mi. A., Machulda, METRO. METRO., Tosakulwong, NORTE., Knopman,
D. S., Petersen, R. C., Jacobo, C. r., Josephs, k. A., & jones, D. t.
(2018). Disrupted functional connectivity in primary progressive
apraxia of speech. NeuroImagen: Clinical, 18, 617–629. https://
doi.org/10.1016/j.nicl.2018.02.036, PubMed: 29845010
Bruffaerts, r., Schaeverbeke, J., De Weer, A.-S., Nelissen, NORTE., Dries,
MI., Van Bouwel, K., Sieben, A., Bergmans, B., Swinnen, C.,
Pijnenburg, y., Sunaert, S., Vandenbulcke, METRO., & Vandenberghe,
R. (2020). Multivariate analysis reveals anatomical correlates of
naming errors in primary progressive aphasia. Neurobiology of
Aging, 88, 71–82. https://doi.org/10.1016/j.neurobiolaging
.2019.12.016, PubMed: 31955981
Calamuneri, A., Arrigo, A., Mormina, MI., Milardi, D., Cacciola, A.,
Chillemi, GRAMO., Marino, S., Gaeta, METRO., & Quartarone, A. (2018).
White matter tissue quantification at low b-values within con-
strained spherical deconvolution framework. Frontiers in Neurol-
ogia, 9, Article 716. https://doi.org/10.3389/fneur.2018.00716,
PubMed: 30210438
Cammoun, l., Gigandet, X., Meskaldji, D., Thiran, j. PAG., despreciar, o.,
Do, k. P., Maeder, PAG., Meuli, r., & Hagmann, PAG. (2012). Cartografía
the human connectome at multiple scales with diffusion spec-
trum MRI. Journal of Neuroscience Methods, 203(2), 386–397.
https://doi.org/10.1016/j.jneumeth.2011.09.031, PubMed:
22001222
Canu, MI., Agosta, F., Imperiale, F., Fontana, A., Caso, F., Spinelli,
mi. GRAMO., Magnani, GRAMO., Falini, A., Comi, GRAMO., & Filippi, METRO. (2019).
Added value of multimodal MRI to the clinical diagnosis of pri-
mary progressive aphasia variants. Corteza, 113, 58–66. https://doi
.org/10.1016/j.cortex.2018.11.025, PubMed: 30605869
Cardenas, V. A., Boxer, A. l., chao, l. l., Gorno-Tempini, METRO. l., Molinero,
B. l., Weiner, METRO. w., & Studholme, C. (2007). Deformation-based
morphometry reveals brain atrophy in frontotemporal dementia.
Archives of Neurology, 64(6), 873–877. https://doi.org/10.1001
/archneur.64.6.873, PubMed: 17562936
Catani, METRO., Mesulam, METRO. METRO., Jakobsen, MI., Malik, F., Martersteck,
A., Wieneke, C., Thompson, C. K., Thiebaut de Schotten, METRO.,
Dell’Acqua, F., Weintraub, S., & Rogalski, mi. (2013). A novel
frontal pathway underlies verbal fluency in primary progressive
aphasia. Cerebro, 136(8), 2619–2628. https://doi.org/10.1093/brain
/awt163, PubMed: 23820597
Crawford, j. r., & Garthwaite, PAG. h. (2007). Comparison of a single
case to a control or normative sample in neuropsychology:
Development of a Bayesian approach. Cognitive Neuropsychology,
24(4), 343–372. https://doi.org/10.1080/02643290701290146,
PubMed: 18416496
Desikan, R. S., Ségonne, F., pescado, B., quinn, B. T., Dickerson,
B. C., Blacker, D., Buckner, R. l., Valle, A. METRO., Maguire, R. PAG.,
Hyman, B. T., Alberto, METRO. S., & Killiany, R. j. (2006). An auto-
mated labeling system for subdividing the human cerebral cortex
on MRI scans into gyral based regions of interest. NeuroImagen,
31(3), 968–980. https://doi.org/10.1016/j.neuroimage.2006.01
.021, PubMed: 16530430
Downey, l. MI., Mahoney, C. J., Buckley, A. h., Golden, h. l., Henley,
S. METRO., Schmitz, NORTE., Schott, j. METRO., Simpson, I. J., Ourselin, S., Fox,
norte. C., Crutch, S. J., & Warren, j. D. (2015). White matter tract sig-
natures of impaired social cognition in frontotemporal lobar
degeneration. NeuroImagen: Clinical, 8, 640–651. https://doi.org
/10.1016/j.nicl.2015.06.005, PubMed: 26236629
Duffy, j. r., Hanley, h., Utianski, r., clark, h., Strand, MI., Josephs,
k. A., & Whitwell, j. l. (2017). Temporal acoustic measures dis-
tinguish primary progressive apraxia of speech from primary pro-
gressive aphasia. Brain and Language, 168, 84–94. https://doi.org
/10.1016/j.bandl.2017.01.012, PubMed: 28187331
Neurobiology of Language
534
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
Duffy, j. r., Strand, mi. A., & Josephs, k. A. (2014). Motor speech
disorders associated with primary progressive aphasia. Aphasiol-
ogia, 28(8–9), 1004–1017. https://doi.org/10.1080/02687038
.2013.869307, PubMed: 25309017
Duffy, j. r., Utianski, R. l., & Josephs, k. A. (2021). Primary pro-
gressive apraxia of speech: From recognition to diagnosis and
care. Aphasiology, 35(4), 1–32. https://doi.org/10.1080
/02687038.2020.1787732, PubMed: 34219857
Feiken, J., & Jonkers, R. (2012). DIAS: Diagnostisch instrument voor
apraxie van de spraak [DIAS: Diagnostic tool for speech apraxia].
Bohn Stafleu van Loghum. https://doi.org/10.1007/978-90-313
-9060-1
pescado, B. (2012). FreeSurfer. NeuroImagen, 62(2), 774–781. https://
doi.org/10.1016/j.neuroimage.2012.01.021, PubMed: 22248573
Forman, METRO. S., Zhukareva, v., Bergeron, C., Chin, S. S.-M.,
Grossman, METRO., clark, C., Sotavento, V. M.-Y., & Trojanowski, j. q.
(2002). Signature tau neuropathology in gray and white matter
of corticobasal degeneration. American Journal of Pathology,
160(6), 2045–2053. https://doi.org/10.1016/S0002-9440(10)
61154-6, PubMed: 12057909
Galantucci, S., Tartaglia, METRO. C., wilson, S. METRO., Henry, METRO. l., Filippi,
METRO., Agosta, F., Dronkers, norte. F., Henry, R. GRAMO., Ogar, j. METRO., Molinero,
B. l., & Gorno-Tempini, METRO. l. (2011). White matter damage in
primary progressive aphasias: A diffusion tensor tractography
estudiar. Cerebro: A Journal of Neurology, 134(10), 3011–3029.
https://doi.org/10.1093/brain/awr099, PubMed: 21666264
Gaser, C., & Kurth, F. (2019). Manual computational anatomy
toolbox—CAT12. https://www.neuro.uni-jena.de/cat12/CAT12
-Manual.pdf
Ghitza, o., & Greenberg, S. (2009). On the possible role of brain
rhythms in speech perception: Intelligibility of time-compressed
speech with periodic and aperiodic insertions of silence. Phone-
tica, 66(1–2), 113–126. https://doi.org/10.1159/000208934,
PubMed: 19390234
Goll, j. C., Crutch, S. J., Loo, j. h. y., Rohrer, j. D., Frost, C.,
Bamiou, D.-E., & Warren, j. D. (2010). Non-verbal sound process-
ing in the primary progressive aphasias. Cerebro, 133(1), 272–285.
https://doi.org/10.1093/brain/awp235, PubMed: 19797352
Gorno-Tempini, METRO. l., Hillis, A. MI., Weintraub, S., Kertesz, A., Mendez,
METRO., Cappa, S. F., Ogar, j. METRO., Rohrer, j. D., Negro, S., Boeve, B. F.,
Manes, F., Dronkers, norte. F., Vandenberghe, r., Rascovsky, K., Patterson,
K., Molinero, B. l., Knopman, D. S., Hodges, j. r., Mesulam, METRO. METRO., &
Grossman, METRO. (2011). Classification of primary progressive apha-
sia and its variants. Neurología, 76(11), 1006–1014. https://doi.org
/10.1212/ WNL.0b013e31821103e6, PubMed: 21325651
Graets, PAG., De Bleser, r., & Willmes, k. (1992). Akense Afasie test.
Swets and Zeitlinger.
Graff-Radford, J., jones, D. T., Strand, mi. A., Rabinstein, A. A., Duffy,
j. r., & Josephs, k. A. (2014). The neuroanatomy of pure apraxia
of speech in stroke. Brain and Language, 129, 43–46. https://doi
.org/10.1016/j.bandl.2014.01.004, PubMed: 24556336
Grahn, j. A., & Brett, METRO. (2007). Rhythm and beat perception in
motor areas of the brain. Revista de neurociencia cognitiva,
19(5), 893–906. https://doi.org/10.1162/jocn.2007.19.5.893,
PubMed: 17488212
Grahn, j. A., & Schuit, D. (2012). Individual differences in rhythmic
capacidad: Behavioral and neuroimaging investigations. Psychomu-
sicology: Music, Mente, and Brain, 22(2), 105–121. https://doi.org
/10.1037/a0031188
Grossman, METRO. (2018). Linguistic aspects of primary progressive
aphasia. Annual Review of Linguistics, 4, 377–403. https://doi
.org/10.1146/annurev-linguistics-011516-034253, PubMed:
30112427
Grube, METRO., Bruffaerts, r., Schaeverbeke, J., Neyens, v., De Weer,
A.-S., Seghers, A., Bergmans, B., Dries, MI., Griffiths, t. D., &
Vandenberghe, R. (2016). Core auditory processing deficits in
primary progressive aphasia. Cerebro, 139(6), 1817–1829.
https://doi.org/10.1093/brain/aww067, PubMed: 27060523
Grube, METRO., & Griffiths, t. D. (2009). Metricality-enhanced temporal
encoding and the subjective perception of rhythmic sequences.
Corteza, 45(1), 72–79. https://doi.org/10.1016/j.cortex.2008.01
.006, PubMed: 19058797
Grube, METRO., Kumar, S., Cooper, F. MI., Turton, S., & Griffiths, t. D.
(2012). Auditory sequence analysis and phonological skill. Pro-
ceedings of the Royal Society B, Ciencias Biologicas, 279(1746),
4496–4504. https://doi.org/10.1098/rspb.2012.1817, PubMed:
22951739
Hardy, C. j. D., Agustus, j. l., marshall, C. r., clark, C. NORTE., Russell,
l. l., Vínculo, R. l., Brotherhood, mi. v., tomás, D. l., Crutch, S. J.,
Rohrer, j. D., & Warren, j. D. (2017). Behavioural and neuro-
anatomical correlates of auditory speech analysis in primary
progressive aphasias. Alzheimer’s Research & Therapy, 9(1),
Article 53. https://doi.org/10.1186/s13195-017-0278-2,
PubMed: 28750682
Hardy, C. j. D., Agustus, j. l., marshall, C. r., clark, C. NORTE., Russell,
l. l., Brotherhood, mi. v., Vínculo, R. l., Fiford, C. METRO., Ondobaka, S.,
tomás, D. l., Crutch, S. J., Rohrer, j. D., & Warren, j. D. (2017).
Functional neuroanatomy of speech signal decoding in primary
progressive aphasias. Neurobiology of Aging, 56, 190–201.
https://doi.org/10.1016/j.neurobiolaging.2017.04.026, PubMed:
28571652
Hardy, C. j. D., Frost, C., Sivasathiaseelan, h., Johnson, j. C. S.,
Agustus, j. l., Vínculo, R. l., Benhamou, MI., Russell, l. l., marshall,
C. r., Rohrer, j. D., Bamiou, D.-E., & Warren, j. D. (2019). Find-
ings of impaired hearing in patients with nonfluent/agrammatic
variant primary progressive aphasia. JAMA Neurology, 76(5),
607–611. https://doi.org/10.1001/jamaneurol.2018.4799,
PubMed: 30742208
Jenkinson, METRO., beckman, C. F., Behrens, t. mi. J., lana rica, METRO. w., &
Herrero, S. METRO. (2012). FSL. NeuroImagen, 62(2), 782–790. https://doi
.org/10.1016/j.neuroimage.2011.09.015, PubMed: 21979382
Josephs, k. A., Duffy, j. r., Strand, mi. A., Machulda, METRO. METRO., Senjem,
METRO. l., Master, A. v., Lowe, V. J., Jacobo, C. r., & Whitwell, j. l.
(2012). Characterizing a neurodegenerative syndrome: Primary
progressive apraxia of speech. Cerebro, 135(5), 1522–1536.
https://doi.org/10.1093/brain/aws032, PubMed: 22382356
Keitel, A., Bruto, J., & Kayser, C. (2018). Perceptually relevant
speech tracking in auditory and motor cortex reflects distinct lin-
guistic features. PLOS Biology, 16(3), Article e2004473. https://
doi.org/10.1371/journal.pbio.2004473, PubMed: 29529019
Levitt, h. (1971). Transformed up-down methods in psychoacous-
tics. Journal of the Acoustical Society of America, 49(2, punto. 2),
467–477. https://doi.org/10.1121/1.1912375
Leyton, C. MI., Landin-Romero, r., Liang, C. T., Burrell, j. r., Kumfor,
F., Hodges, j. r., & Piguet, oh. (2019). Correlates of anomia in
non-semantic variants of primary progressive aphasia converge
con el tiempo. Corteza, 120, 201–211. https://doi.org/10.1016/j
.cortex.2019.06.008, PubMed: 31325799
Largo, METRO. A., Katlowitz, k. A., Svirsky, METRO. A., Clary, R. C., Byun,
t. METRO., Majaj, NORTE., Oya, h., Howard, METRO. A., & verdelee,
j. D. W.. (2016). Functional segregation of cortical regions under-
lying speech timing and articulation. Neurona, 89(6), 1187–1193.
https://doi.org/10.1016/j.neuron.2016.01.032, PubMed:
26924439
Magrassi, l., Aromataris, GRAMO., Cabrini, A., Annovazzi-Lodi, v., &
Moro, A. (2015). Sound representation in higher language areas
Neurobiology of Language
535
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
/
.
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
during language generation. Proceedings of the National Acad-
emy of Sciences, 112(6), 1868–1873. https://doi.org/10.1073
/pnas.1418162112, PubMed: 25624479
Mahoney, C. J., Malone, I. B., Ridgway, GRAMO. r., Buckley, A. h.,
Downey, l. MI., Golden, h. l., ryan, norte. S., Ourselin, S., Schott,
j. METRO., Rossor, METRO. NORTE., Fox, norte. C., & Warren, j. D. (2013). Blanco
matter tract signatures of the progressive aphasias. Neurobiología
of Aging, 34(6), 1687–1699. https://doi.org/10.1016/j
.neurobiolaging.2012.12.002, PubMed: 23312804
Mandelli, METRO. l., Caverzasi, MI., Binney, R. J., Henry, METRO. l., Lobach,
I., Block, NORTE., Amirbekian, B., Dronkers, NORTE., Molinero, B. l., Henry,
R. GRAMO., & Gorno-Tempini, METRO. l. (2014). Frontal white matter tracts
sustaining speech production in primary progressive aphasia.
Revista de neurociencia, 34(29), 9754–9767. https://doi.org/10
.1523/JNEUROSCI.3464-13.2014, PubMed: 25031413
Mandelli, METRO. l., Vilaplana, MI., Marrón, j. A., Hubbard, h. I., Binney,
R. J., Attygalle, S., Santos-Santos, METRO. A., Molinero, z. A., Pakvasa, METRO.,
Henry, METRO. l., rosa, h. J., Henry, R. GRAMO., Rabinovici, GRAMO. D., Molinero,
B. l., Seeley, W.. w., & Gorno-Tempini, METRO. l.
(2016).
Healthy brain connectivity predicts atrophy progression in
non-fluent variant of primary progressive aphasia. Cerebro, 139(10),
2778–2791. https://doi.org/10.1093/ brain/aww195, PubMed:
27497488
Mariën, PAG., Mampaey, MI., Vervaet, A., Saerens, J., & De Deyn, PAG. PAG.
(1998). Normative data for the Boston naming test in native
Dutch-speaking Belgian elderly. Brain and Language, 65(3),
447–467. https://doi.org/10.1006/ brln.1998.2000, PubMed:
9843614
Mesulam, M.-M., Rogalski, mi. J., Wieneke, C., Hurley, R. S., Geula,
C., Bigio, mi. h., Thompson, C. K., & Weintraub, S. (2014).
Primary progressive aphasia and the evolving neurology of
the language network. Nature Reviews Neurology, 10(10),
554–569. https://doi.org/10.1038/nrneurol.2014.159, PubMed:
25179257
Nuevo, A. B., Robin, D. A., Parkinson, A. l., Duffy, j. r., McNeil,
METRO. r., Piguet, o., Hornberger, METRO., Precio, C. J., Eickhoff, S. B.,
& Ballard, k. j. (2015). Altered resting-state network connectivity
in stroke patients with and without apraxia of speech. Neuro-
Image: Clinical, 8, 429–439. https://doi.org/10.1016/j.nicl.2015
.03.013, PubMed: 26106568
Ogar, j. METRO., Dronkers, norte. F., Brambati, S. METRO., Molinero, B. l., & Gorno-
Tempini, METRO. l. (2007). Progressive nonfluent aphasia and its
characteristic motor speech deficits. Alzheimer Disease & también-
ciated Disorders, 21(4), S23–S30. https://doi.org/10.1097/ WAD
.0b013e31815d19fe, PubMed: 18090419
Poeppel, D., & Assaneo, METRO. F. (2020). Speech rhythms and their
neural foundations. Naturaleza Reseñas Neurociencia, 21(6),
322–334. https://doi.org/10.1038/s41583-020-0304-4, PubMed:
32376899
Poline, j. B., Worsley, k. J., evans, A. C., & Friston, k. j. (1997).
Combining spatial extent and peak intensity to test for activations
in functional imaging. NeuroImagen, 5(2), 83–96. https://doi.org
/10.1006/nimg.1996.0248, PubMed: 9345540
Powers, j. PAG., McMillan, C. T., Brun, C. C., Yushkevich, PAG. A., zhang,
h., Gee, j. C., & Grossman, METRO. (2013). White matter disease cor-
relates with lexical retrieval deficits in primary progressive apha-
sia. Frontiers in Neurology, 4, Article 212. https://doi.org/10.3389
/fneur.2013.00212, PubMed: 24409166
Radwan, A. [METRO.], & Sunaert, S. (2022, Febrero). KU Lovaina
NeuroImaging suite: A set of tools for brain and spine MRI anal-
ysis [Computer software]. https://github.com/treanus/KUL_NIS
Radwan, A. METRO., Sunaert, S., Schilling, K., Descoteaux, METRO., Landman,
B. A., Vandenbulcke, METRO., Theys, T., Dupont, PAG., & Emsell, l.
(2022). An atlas of white matter anatomy, its variability, y
reproducibility based on constrained spherical deconvolution
of diffusion MRI. NeuroImagen, 254, Article 119029. https://doi
.org/10.1016/j.neuroimage.2022.119029, PubMed: 35231632
Rogalski, MI., Cobia, D., harrison, t. METRO., Wieneke, C., Thompson,
C. K., Weintraub, S., & Mesulam, M.-M. (2011). Anatomy of lan-
guage impairments in primary progressive aphasia. Diario de
Neurociencia, 31(9), 3344–3350. https://doi.org/10.1523
/JNEUROSCI.5544-10.2011, PubMed: 21368046
Rohrer, j. D., Rossor, METRO. NORTE., & Warren, j. D. (2010). Syndromes of
nonfluent primary progressive aphasia: A clinical and neurolin-
guistic analysis. Neurología, 75(7), 603–610. https://doi.org/10
.1212/ WNL.0b013e3181ed9c6b, PubMed: 20713949
Rohrer, j. D., Sauter, D., Scott, S., Rossor, METRO. NORTE., & Warren, j. D.
(2012). Receptive prosody in nonfluent primary progressive
aphasias. Corteza, 48(3), 308–316. https://doi.org/10.1016/j
.cortex.2010.09.004, PubMed: 21047627
Sajjadi, S. A., Acosta-Cabronero, J., Patterson, K., Diaz-de-Grenu,
l. Z., williams, GRAMO. B., & Nestor, PAG. j. (2013). Diffusion tensor
magnetic resonance imaging for single subject diagnosis in neu-
rodegenerative diseases. Cerebro, 136(7), 2253–2261. https://doi
.org/10.1093/brain/awt118, PubMed: 23729473
Schaeverbeke, J., Gabel, S., Meersmans, K., Bruffaerts, r., Liuzzi,
A. GRAMO., Evenepoel, C., Dries, MI., Van Bouwel, K., Sieben, A.,
Pijnenburg, y., Peeters, r., Bormans, GRAMO., Van Laere, K., Koole,
METRO., Dupont, PAG., & Vandenberghe, R. (2018). Single-word com-
prehension deficits in the nonfluent variant of primary progres-
sive aphasia. Alzheimer’s Research & Therapy, 10(1), Article 68.
https://doi.org/10.1186/s13195-018-0393-8 , PubMed:
30021613
Schilling, k. GRAMO., Blaber, J., Huo, y., Newton, A., Hansen, C., Nath,
v., Shafer, A. T., williams, o., Resnick, S. METRO., Rogers, B., anderson,
A. w., & Landman, B. A. (2019). Synthesized b0 for diffusion dis-
tortion correction (Synb0-DisCo). Imagen de resonancia magnética,
64, 62–70. https://doi.org/10.1016/j.mri.2019.05.008, PubMed:
31075422
Herrero, S. METRO., Jenkinson, METRO., lana rica, METRO. w., beckman, C. F.,
Behrens, t. mi. J., Johansen-Berg, h., Bannister, PAG. r., De Luca,
METRO., Drobnjak, I., Flitney, D. MI., Niazy, R. K., Saunders, J., Vickers,
J., zhang, y., De Stefano, NORTE., Brady, j. METRO., & Matthews, PAG. METRO.
(2004). Advances in functional and structural MR image analysis
and implementation as FSL. NeuroImagen, 23(S1), S208–S219.
https://doi.org/10.1016/j.neuroimage.2004.07.051, PubMed:
15501092
Spinelli, mi. GRAMO., Mandelli, METRO. l., Molinero, z. A., Santos-Santos, METRO. A.,
wilson, S. METRO., Agosta, F., Grinberg, l. T., Huang, mi. J., Trojanowski,
j. P., Meyer, METRO., Henry, METRO. l., Comi, GRAMO., Rabinovici, GRAMO., rosa,
h. J., Filippi, METRO., Molinero, B. l., Seeley, W.. w., & Gorno-Tempini,
METRO. l. (2017). Typical and atypical pathology in primary progres-
sive aphasia variants: Pathology in PPA Variants. Annals of Neu-
rology, 81(3), 430–443. https://doi.org/10.1002/ana.24885,
PubMed: 28133816
Tetzloff, k. A., Duffy, j. r., clark, h. METRO., Strand, mi. A., Machulda,
METRO. METRO., Schwarz, C. GRAMO., Senjem, METRO. l., Reid, R. I., Spychalla, A. J.,
Tosakulwong, NORTE., Lowe, V. J., Jacobo, Jr., C. r., Josephs, k. A., &
Whitwell, j. l. (2018). Longitudinal structural and molecular neu-
roimaging in agrammatic primary progressive aphasia. Cerebro,
141(1), 302–317. https://doi.org/10.1093/ brain/awx293,
PubMed: 29228180
Tetzloff, k. A., Duffy, j. r., clark, h. METRO., Utianski, R. l., Strand,
mi. A., Machulda, METRO. METRO., Botha, h., Martín, PAG. r., Schwarz,
C. GRAMO., Senjem, METRO. l., Reid, R. I., Gunter, j. l., Spychalla, A. J.,
Knopman, D. S., Petersen, R. C., Jacobo, C. r., Lowe, V. J., Josephs,
Neurobiology of Language
536
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Left frontal white matter & rhythm in PPA
k. A., & Whitwell, j. l. (2019). Progressive agrammatic aphasia
without apraxia of speech as a distinct syndrome. Cerebro, 142(8),
2466–2482. https://doi.org/10.1093/ brain/awz157, PubMed:
31199471
Tourbier, S., Alemán-Gómez, y., Griffa, A., Bach Cuadra, METRO., &
Hagmann, PAG. (2020). Multi-scale brain parcellator: A BIDS App
for the Lausanne connectome parcellation [Computer software].
https://doi.org/10.7490/F1000RESEARCH.1117844.1
Tournier, J.-D., Calamante, F., & connelly, A. (2007). Robust deter-
mination of the fibre orientation distribution in diffusion MRI:
Non-negativity constrained super-resolved spherical deconvolu-
ción. NeuroImagen, 35(4), 1459–1472. https://doi.org/10.1016/j
.neuroimage.2007.02.016, PubMed: 17379540
Tournier, J.-D., Calamante, F., & connelly, A. (2010). Improved
probabilistic streamlines tractography by 2nd order integration
over fibre orientation distributions. ISMRM: Actas de la
International Society for Magnetic Resonance in Medicine 18,
1670.
Tournier, J.-D., Calamante, F., & connelly, A. (2012). MRtrix: Diffu-
sion tractography in crossing fiber regions. Imaging Systems and
Tecnología, 22(1), 53–66. https://doi.org/10.1002/ima.22005
Tournier, J.-D., Herrero, r., Raffelt, D., Tabbara, r., Dhollander, T.,
Pietsch, METRO., Christiaens, D., Jeurissen, B., Yeh, C.-H., & connelly,
A. (2019). MRtrix3: A fast, flexible and open software framework
for medical image processing and visualisation. NeuroImagen,
202, Article 116137. https://doi.org/10.1016/j.neuroimage.2019
.116137, PubMed: 31473352
Tustison, norte. J., Cocinar, PAG. A., Holbrook, A. J., Johnson, h. J.,
Muschelli, J., Devenyi, GRAMO. A., Duda, j. T., El, S. r., Cullen,
norte. C., Gillen, D. l., Yassa, METRO. A., Piedra, j. r., Gee, j. C., &
Avants, B. B. (2021). The ANTsX ecosystem for quantitative biolog-
ical and medical imaging. Informes Científicos, 11(1), Article 9068.
https://doi.org/10.1038/s41598-021-87564-6, PubMed:
33907199
Utianski, R. l., Duffy, j. r., clark, h. METRO., Strand, mi. A., Botha, h.,
Schwarz, C. GRAMO., Machulda, METRO. METRO., Senjem, METRO. l., Spychalla,
A. J., Jacobo, C. r., Petersen, R. C., Lowe, V. J., Whitwell, j. l., &
Josephs, k. A. (2018). Prosodic and phonetic subtypes of primary
progressive apraxia of speech. Brain and Language, 184, 54–65.
https://doi.org/10.1016/j.bandl.2018.06.004, PubMed:
29980072
Veraart, J., Nóvikov, D. S., Christiaens, D., Ades-aron, B., Sijbers, J.,
& Fieremans, mi. (2016). Denoising of diffusion MRI using random
matrix theory. NeuroImagen, 142, 394–406. https://doi.org/10
.1016/j.neuroimage.2016.08.016, PubMed: 27523449
Vergis, METRO. K., Ballard, k. J., Duffy, j. r., McNeil, METRO. r., Scholl, D., &
Layfield, C. (2014). An acoustic measure of lexical stress differ-
entiates aphasia and aphasia plus apraxia of speech after stroke.
Aphasiology, 28(5), 554–575. https://doi.org/10.1080/02687038
.2014.889275
Warren, j. D., Rohrer, j. D., Schott, j. METRO., Fox, norte. C., Hardy, J., &
Rossor, METRO. norte. (2013). Molecular nexopathies: A new paradigm of
neurodegenerative disease. Trends in Neurosciences, 36(10),
561–569. https://doi.org/10.1016/j.tins.2013.06.007, PubMed:
23876425
Whitwell, j. l., Avula, r., Senjem, METRO. l., Kantarci, K., Weigand,
S. D., Samikoglu, A., Edmonson, h. A., Vemuri, PAG., Knopman,
D. S., Boeve, B. F., Petersen, R. C., Josephs, k. A., & Jacobo,
C. R. (2010). Gray and white matter water diffusion in the syn-
dromic variants of frontotemporal dementia. Neurología, 74(16),
1279–1287. https://doi.org/10.1212/ WNL.0b013e3181d9edde,
PubMed: 20404309
Whitwell, j. l., Duffy, j. r., Strand, mi. A., Xia, r., Mandrekar, J.,
Machulda, METRO. METRO., Senjem, METRO. l., Lowe, V. J., Jacobo, C. r., &
Josephs, k. A. (2013). Distinct regional anatomic and functional
correlates of neurodegenerative apraxia of speech and aphasia: Un
MRI and FDG-PET study. Brain and Language, 125(3), 245–252.
https://doi.org/10.1016/j.bandl.2013.02.005, PubMed: 23542727
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
norte
oh
/
yo
/
yo
a
r
t
i
C
mi
–
pag
d
F
/
/
/
/
3
4
5
1
5
2
0
4
4
1
1
4
norte
oh
_
a
_
0
0
0
7
5
pag
d
.
/
yo
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurobiology of Language
537