Taran, NORTE., Farah, r., Gashri, C., Gitman, MI., Rosch, K., Schlaggar, B.L. & Horowitz-Kraus, t. (2023). Executive functions-
based reading training engages the cingulo-opercular and dorsal attention networks. Neurociencia en red, Advance
publicación. https://doi.org/10.1162/netn_a_00335.
Executive functions-based reading training engages the cingulo-
opercular and dorsal attention networks
Título corto: Executive functions-based reading training: neurobiological effect
Nikolay Taran1, Rola Farah1, Carmel Gashri1, Ester Gitman1, Keri Rosch2,3,
Bradley L. Schlaggar2,4,5, and Tzipi Horowitz-Kraus1,2,3
1Educational Neuroimaging Group, Faculty of Education in Science and
Tecnología, Faculty of Biomedical Engineering, Technion Israel Institute of
Tecnología, Haifa, Israel
2Kennedy Krieger Institute, baltimore, Maryland, EE.UU
3Department of Psychiatry and Behavioral Science, Universidad Johns Hopkins
School of Medicine, baltimore, Maryland, EE.UU
4Department of Pediatrics, Johns Hopkins University School of Medicine,
baltimore, Maryland, EE.UU
5Department of Neurology, Johns Hopkins University School of Medicine,
baltimore, Maryland, EE.UU
Expresiones de gratitud: This study was supported by the National Institute of
Child Health and Human Development (R01 HD086011; PI: t. H.-K.).
Conflict of interest: The authors declare no potential conflict of interest
1
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
t
/
.
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
t
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Abstracto
The aim of this study was to determine the effect of a computerized executive
funciones (EFs)-based reading intervention on neural circuits supporting EFs
and visual attention. Seed-to-voxel functional connectivity analysis was
conducted focusing on large-scale attention system brain networks, during an
fMRI reading fluency task. Los participantes fueron 8-12 year-old English-speaking
children with dyslexia (n=43) and typical readers (n=36) trained on an EFs-
based reading training (n=40) vs math training (n=39). Training duration was 8
semanas.
1
2
3
4
5
6
7
8
9
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
/
t
.
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
.
t
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
After the EFs-based reading intervention, children with dyslexia improved their
10
scores in reading rate and visual attention (compared to math intervention).
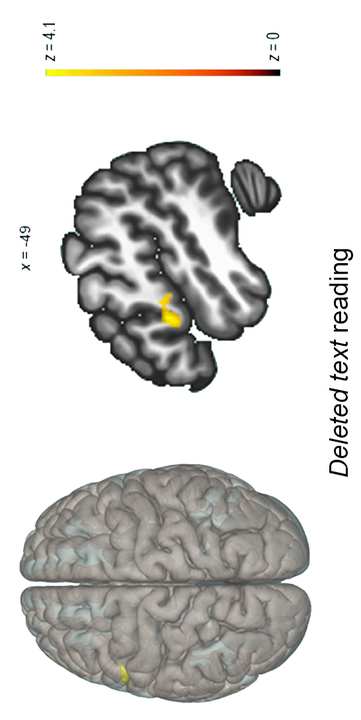
Neurobiologically, children with dyslexia displayed an increase in functional
connectivity strength after the intervention between the cingulo-opercular
network and occipital and precentral regions. Noteworthy, the functional
connectivity indices between these brain regions showed a positive
correlation with speed of processing and visual attention scores in both pre-
test and post-test. The results suggest that reading improvement following an
EFs-based reading intervention involves neuroplastic connectivity changes in
brain areas related to EFs and primary visual processing in children with
dyslexia. Our results highlight the need for training underlying cognitive
abilities supporting reading, such as EFs and visual attention, in order to
enhance reading abilities in dyslexia.
11
12
13
14
15
16
17
18
19
20
21
22
Palabras clave: Dyslexia, Executive functions, Visual attention, Intervention, resonancia magnética funcional,
23
Functional Connectivity.
24
2
Introducción
Dyslexia: definition and explanatory theories
Developmental dyslexia (henceforth, dyslexia) is classified as one type of
specific learning disorder, with different studies reporting a prevalence
entre 5 y 20% (Schulte-Körne 2010, norton, Black et al. 2014, Wagner,
Zirps et al. 2020). This disability is a heritable, life-long condition with early
onset (Snowling, Hulme et al. 2020). Dyslexia is described as a difficulty in
accurate and fluent word recognition and spelling (Peterson and Pennington
2012) that cannot be explained by sensorial deficits, insufficient literature
exposure, delayed development of cognitive abilities or low intelligence
(Schulte-Körne 2010).
For the last several decades, the scientific consensus regards dyslexia as a
language disorder in which, for alphabetic-based written language, el
proximate cause is a phonological processing deficit (Peterson and
Pennington, 2012, pag. 3). A close relationship exists between children’s
phonological skills (es decir. phonological awareness) and the mastering of word
lectura (Melby-Lervag, Lyster et al. 2012). According to the mentioned
theory, children with reading difficulties manifest a neural processing deficit in
the representation of the sounds in language. More recently, the role of
Executive Functions (EFs) in dyslexia has been highlighted (Horowitz-Kraus
2012, Varvara, Varuzza et al. 2014). EFs are a set of higher-order cognitive
abilities (inhibition, switching, updating, ver (Miyake 2000)) that allow
individuals to adapt and overcome different challenging environmental
condiciones (galés, Pennington et al. 1991, Diamante 2020). Recent studies
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
.
t
/
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
t
/
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
have highlighted the critical role EFs have in intact and impaired reading as
supporting all three components of the Simple View of Reading model (RVS):
word decoding (Kieffer and Christodoulou 2020, Nguyen, Del Tufo et al.
2020), comprensión (Washburn 2022) and critically, reading fluency –
defined as fast and accurate reading (Silverman, Speece et al. 2013). El
three main EFs (inhibition, switching, updating (Miyake 2012)) seem to have a
direct effect on reading fluency (Kieffer and Christodoulou 2020, Nguyen, Del
Tufo et al. 2020).
Children with dyslexia show dysfunctions in both verbal and visuo-spatial
1
2
3
4
5
6
7
8
9
working memory, switching, and in the inhibition of irrelevant information
10
(Booth, Boyle et al. 2010, Horowitz-Kraus 2012, Varvara, Varuzza et al. 2014,
11
Barbosa, Rodrigues et al. 2019). Además, children with dyslexia display
12
below-average performance in speed of processing (Booth, Boyle et al. 2010),
13
which raised a theory regarding a slow speed of processing and a lack of
synchronization between visual and auditory sensory modalities in these
readers (Breznitz 2003, Breznitz 2006). Por último, visuo-spatial attention
difficulties were also reported (Franceschini, Gori et al. 2012, Varvara,
Varuzza et al. 2014). These different theories aiming to explain the source of
reading difficulties in dyslexia emphasize the complexity of the reading
proceso, as outlined in the SVR (Hoover and Gough 1990) and its refined
extensions (Catts 2018, Spencer, Richmond et al. 2020).
Dyslexia presents thus, a multifaceted nature with tight ontogenetic relation
between the underlying neural systems (Dehaene 2009), and variabilility
related to the different types of orthographies (Siok, Jia et al. 2020).
14
15
16
17
18
19
20
21
22
23
24
4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
.
/
t
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
t
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurobiological correlates of dyslexia
The neural circuits associated with word decoding (the factor underlying the
reading comprehension deficit in dyslexia, in terms of the SVR) encompass
mostly left hemisphere areas, including the left inferior frontal gyrus or left IFG
(norton, Beach et al. 2015), inferior occipito-temporal regions (p.ej., the Visual
Word Form Area or VWFA), and regions around the tempo-parietal junction,
including the angular (AG) and supramarginal gyri (SMG). Studies in adults
report an engagement of the VWFA not only in decoding tasks (Dehaene and
cohen 2011, Cutting, Clements-Stephens et al. 2013), but also in
phonological tasks (Yarkoni, Speer et al. 2008, Conant, Liebenthal et al.
2020), providing support for a role of this region in linking phonology and
orthography. The SMG was found to be activated during auditory processing
of syllabic sequences (Deschamps and Tremblay 2014) as well as word
lectura (Weiss, Katzir et al. 2016). The processing features of the mentioned
areas seem to be especially relevant for the decoding (es decir., visual-to-
phonological translation) of graphemes and words (Vogel, Petersen et al.
2014). Sin embargo, the specific mechanistic contributions of each of these
cortical regions to the process of reading are not yet fully understood.
It has been suggested that the functional connectivity of four different
cognitive networks comprising the attention system are affected in dyslexia:
the cingulo-opercular (CO), fronto-parietal (FP), ventral and dorsal attention
redes (VAN, and DAN respectively) (Corbetta and Shulman 2002,
Freedman, Zivan et al. 2020, Farah, Ionta et al. 2021, Taran, Farah et al.
2021). The FP network seems to predispose to switching and goal–directed
comportamiento, while the CO is related to error monitoring and feedback control
5
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
/
.
t
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
.
t
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(Dösenbach, Fair et al. 2007, graton, Sun et al. 2018). The VAN and DAN
are mainly involved in visuo-spatial attention, in both bottom-up and top-down
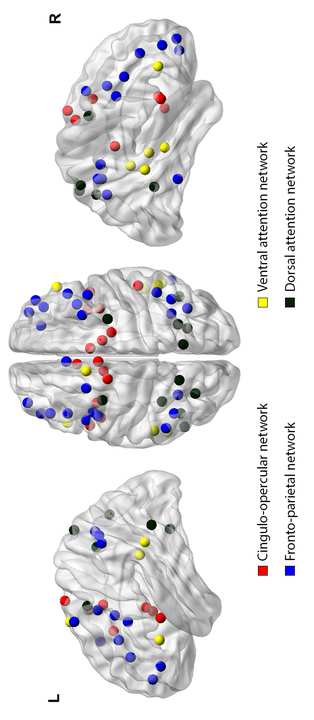
procesos (ver figura 1) (Corbetta and Shulman 2002, Szczepanski and
Kastner 2013, Liu, Bengson et al. 2016). We have previously suggested that
artificially accelerated reading vs reading at a natural reading speed in
English-speaking 8-12 years old children with dyslexia is associated with
increased synchronization between the CO-FP networks and visual-auditory
redes (Horowitz-Kraus 2022). These results echoed findings from Hebrew-
speaking adults showing that during natural reading speed, there is a reduced
synchronization between ERPs associated with visual and auditory
Procesando, which was also associated with reduced speed of processing
during word reading, supporting the asynchronization theory (Breznitz 2003,
1
2
3
4
5
6
7
8
9
10
11
12
Breznitz 2006). These findings support the neural noise hypothesis in children
13
with dyslexia as an explanation for their slow reading speed (Hancock 2017),
but also open up possibilities for reading improvements, especially in the
fluency domain, to reduce this “noise”.
14
15
16
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
/
t
.
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
.
t
/
Fundamentalmente, the VWFA displays specific patterns of connectivity with the angular
17
gyrus and the IFG, making it a candidate to be one of the links between
linguistic (temporal language network) and attentional circuitry (FP) (stevens,
Kravitz et al. 2017, Chen, Wassermann et al. 2019, López-Barroso, Thiebaut
18
19
20
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
de Schotten et al. 2020). This region is functionally connected to the DAN and
21
is highly involved in general visual processing (Vogel, Miezin et al. 2011).
Además, the angular gyrus and the IFG seem to play a role in language
processing and reading as integrative, multimodal hubs, es decir., recruiting and
synchronizing large-scale whole-brain networks (Rosselli, Ardila et al. 2015,
22
23
24
25
6
Xu, Lin et al. 2016, Taran, Farah et al. 2021). En resumen, convergent
evidence points towards the mentioned brain networks and regions as the
primary neurobiological correlates of reading.
Cifra 1. Graphical representation of the CO, the FP, the VAN, y el
DAN
[Insert Figure 1 aquí]
Cifra 1. Graphical representation of the CO, the FP, the VAN, and the DAN,
as described based on anatomical and functional meta-analyses (Fuerza,
Cohen et al. 2011). From left to right: lateral view of the left hemisphere,
superior view of the brain, lateral view of the right hemisphere.
Reading interventions for dyslexia
1
2
3
4
5
6
7
8
9
10
11
In line with the traditional, phonological understanding of dyslexia, most of the
12
current interventions are focused on the explicit training of phonological
awareness and grapheme-phoneme correspondence skills or spelling ability
13
14
(Galuschka, Görgen et al. 2020). Although these interventions generally show
15
a positive effect on reading efficiency in children (morris, Lovett et al. 2012,
16
Savage, Georgiou et al. 2018), investigating the effectiveness of phonological
17
based-interventions suggests that additional sub-components of reading can
be trained in order to improve reading abilities (Strong, Torgerson et al. 2011,
Snowling 2013, williams, Walker y otros. 2017, Snowling, Hulme et al. 2020).
Multiple empirical as well as review studies, meta-analyses, and theoretical
works suggest that future treatments of dyslexia ought to be multi-sensorial
(Snowling and Hulme 2012), and focused not only on explicit phonological
18
19
20
21
22
23
skills but also underlying cognitive abilities, such as EFs, speed of processing
24
and visuo-spatial attention skills (Horowitz-Kraus, Vannest et al. 2014, Peters,
25
7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
t
/
/
.
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
.
t
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
De Losa et al. 2019, piedra 2019, Vidyasagar 2019). Además, novedoso
perspectives on reading instruction and remediation highlight the importance
of improving sublexical skills and general aspects of language and knowledge
in combination with phonological training (Fletcher, Savage et al. 2021).
In the past years, several studies have demonstrated the effect of an EFs-
based reading intervention targeting working memory, inhibition, visuo-spatial
atención, and speed of processing while exposed to written materials (es decir.
oraciones) (Breznitz, Shaul et al. 2013, Horowitz-Kraus 2013, Horowitz-Kraus
2014, Horowitz-Kraus 2014, Horowitz-Kraus and Breznitz 2014, Horowitz-
1
2
3
4
5
6
7
8
9
Kraus 2015, Horowitz-Kraus, DiFrancesco et al. 2015, Horowitz-Kraus 2015b,
10
Horowitz-Kraus 2015c, Cecil, Brunst et al. 2021) on reading, EFs, and brain
structure and function. This training program forces the reader to visually
follow the letters (engaging visual attention) as they are erased from the
11
12
13
pantalla (reliance on working memory) and replaced by asterisks at a gradually
14
increasing speed (speed of processing) without the ability to regress to the
beginning of the sentence (inhibition) (Breznitz 1992, Cecil, Brunst et al.
2021). This training was found to improve reading rate, accuracy and reading
comprensión (Horowitz-Kraus, Vannest et al. 2014, Peters, De Losa et al.
2019), as well as working memory, switching, shifting, visual attention, y
speed of processing (Horowitz-Kraus 2014, Horowitz-Kraus, Cicchino et al.
2014, Horowitz-Kraus, Hershey et al. 2019, Peters, De Losa et al. 2019). On
the neurobiological level, this training was found to increase the connectivity
15
16
17
18
19
20
21
22
strength between EFs, attention and sensory networks (visual processing and
23
auditory networks) during word reading tasks (Horowitz-Kraus, DiFrancesco
24
et al. 2015, Horowitz-Kraus and Holland 2015, Horowitz-Kraus, Hershey et al.
25
8
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
t
/
.
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
.
/
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
2019), increase the within-network connectivity of the CO network during rest
(Horowitz-Kraus, Toro-Serey et al. 2015), increase the magnitude of error-
detection ERPs during word reading errors (Horowitz-Kraus 2016), y era
associated with lower GLX/Glu concentration in the anterior cingulate cortex
(Cecil, Brunst et al. 2021). It was suggested that the speeded deletion of
letters from the screen (artificially inducing fluent reading), engages EFs and
attention neural-circuits and hence reduces the asynchrony/neural noise in
the visual-auditory circuits (Cecil, Brunst et al. 2021). Sin embargo, the effect of
this EFs-based reading intervention both behaviorally and neurobiologically
compared to active control training, is yet to be resolved. More specifically,
the effect of this EFs-based reading training on brain network connectivity
during a reading fluency task against an active control group remains
unknown.
Aims and hypotheses
1
2
3
4
5
6
7
8
9
10
11
12
13
14
The goal of the present study was to determine the neurobiological systems
15
underlying the reading improvement following EFs-based reading intervention
16
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
t
/
/
.
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
/
t
.
while performing a contextual reading fluency task. En la misma vena, nosotros
aimed to measure the effects of the hypothesized neuroplastic changes on
specific cognitive domains. Para tal fin, functional MRI was acquired while
children with dyslexia and typically reading children performed a contextual
reading fluency task before and after the intervention. The experimental
design included an active control group that trained using the math program.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
17
18
19
20
21
22
Children with and without dyslexia were included in the experiment, given that
23
the goal of the study was to explore the behavioral and neurobiological effect
24
9
of the EFs-based reading training on both TR and children with dyslexia.
Específicamente, it was hypothesized that children with dyslexia would benefit
more from the intervention both in terms of behavioral measures (visual
atención, EFs, lectura) and neurobiological changes, ultimately leading to a
reduction in cognititve and functional connectivity disparities between the two
grupos.
We expected both TRs and children with dyslexia who underwent the EFs-
based reading intervention to improve their contextual reading fluency, EFs,
and attention abilities compared to those undergoing the control intervention.
Neurobiologically, we hypothesized that higher functional connectivity in both
typical readers (TR) and children with dyslexia would be found between EFs
redes (FP, CO), attention networks (VAN, DAN), and sensory networks
(visual, auditory) after the EFs-based reading intervention, relative to the
improvement with math training. Además, children with dyslexia when
1
2
3
4
5
6
7
8
9
10
11
12
13
14
compared with typical readers were expected to display greater changes both
15
in functional connectivity (stronger connections) y desempeño conductual
(higher scores).
Métodos
Study Procedure
16
17
18
19
The studies were conducted at Cincinnati Children’s Hospital Medical Center,
20
Ohio, EE.UU. The experimental procedure was designed in accordance with the
21
Declaration of Helsinki, it was reviewed and approved by the institutional
review board. Participants’ parents signed informed consent before enrolling
to the study. Children were compensated for their participation in the study
22
23
24
10
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
.
t
/
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
/
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(en general $150). Exclusion criteria included: comorbidity with attention
difficulties, intellectual disability or any other neurodevelopmental disorders,
neurological or psychiatric conditions.
Primero, all participants underwent a battery of cognitive tests. fMRI data were
acquired while performing the contextual reading fluency task (ver figura 2
for an overview of the experimental design). Después, all children were
randomly divided into two intervention groups. One group underwent an EFs-
based reading training, another group underwent a control (matemáticas) training.
Both interventions had a duration of 8 semanas (3 sessions per week, 20-25
minutes per session). Finalmente, a post-test cognitive test battery was
administered and a second fMRI session was conducted.
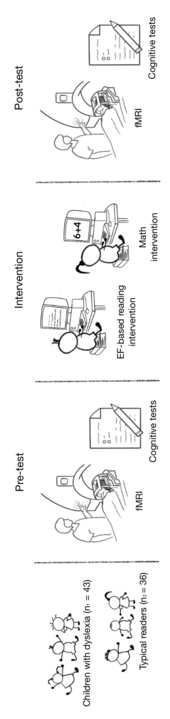
Cifra 2. Overview of the experimental design.
[Insert Figure 2 aquí]
Cifra 2. Seventy-nine children participated in the research project. Todo
participants underwent behavioral assessment and resting-state fMRI
scanning. Después, they were randomly divided into two intervention
grupos: the Executive Functions (EFs)-based reading training (experimental
grupo) and math training (active control group). After the intervention
sessions, they underwent the same behavioral tests performed in the pre-test
and a second resting-state fMRI session.
Participantes
Un total de 79 English-speaking children participated in the experiment: 43
typical readers (TR, edad media = 10.04 ± 1.45, 18 hembras), y 36 niños
with dyslexia (children with dyslexia, edad media = 9.3 ± 1.36, 22 hembras).
11
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
t
/
.
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
t
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Participants from both groups were randomly divided into two intervention
grupos: one group underwent the EFs-based reading training (21 TR, 19
children with dyslexia) and the second group performed computerized math
training (22 TR, 17 children with dyslexia), which served as an active control
measure. There were no significant differences in nonverbal reasoning
abilities between the reading groups (TR mean percentile = 67.27 ± 19.33,
mean percentile = 56.83 ± 22.82, t(77)=2.20, p>.05) nor the intervention
grupos (EFs-based reading training group mean percentile = 59.9 ± 21.9,
math training group mean = 62.67 ± 19.9, t(77)=.581, p>.05). Similarmente, allá
were no significant differences between the groups in age or sex (niños
with dyslexia mean age = 9.65 ± 1.42, TR mean age = 9.96 ± 1.34,
t(77)=1.23, p>.05; EFs-based reading training group mean age = 9.8 ± 1.4,
math training group mean age = 9.6 ± 1.4, t(77)=.329, df=77, p>.05). El
present study experienced a sample attrition rate of 16.8%: 16 out of 95
participants did not complete the training (6 dropped out from the EFs-based
reading intervention and 10 from the math intervention) and were
subsequently excluded from further analysis. The final number of participants
(79) provided a statistical power above 95% for both independent samples t-
tests and 2×2 repeated measures ANOVA (Rosner 2011).
Behavioral measures
General cognitive abilities
Non-verbal intelligence was evaluated using the Test of Nonverbal
Inteligencia (TONI) (Marrón, Sherbenou et al. 2010). General verbal abilities
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
12
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
.
/
t
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
t
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(receptive vocabulary) were evaluated using the Peabody Picture Vocabulary
Test – 4th Edition (PPVT-4) (Dunn and Dunn 2007).
Reading abilities
Reading abilities in both TR and children with dyslexia were evaluated using a
battery of normative English tests: orthographic processing: Test of Word
Reading Efficiency (TOWRE-Sight Word Efficiency) (Torgesen, Wagner et al.
1999), phonological processing: Comprehensive Test of Phonological
Procesando (CTOPP) (Wagner, Torgesen et al. 2013), reading accuracy
(number of correctly read words), reading rate (reading speed) y
comprehension -Gray Oral Reading Test (GORT) for reading rate, exactitud,
and comprehension (Wiederholt and Bryant 2012) and Test of Silent Reading
Efficiency and Comprehension (TOSREC) for reading comprehension
(Schrank, Mather et al. 2014). Orthographic and phonological processing
were measured using isolated word/non-word reading. Reading accuracy,
rate were measured in contextual reading.
Executive functions and attention abilities
Executive functions were assessed using several tasks designed to address
the three main EFs: 1) Working memory was assessed using both forward
and backward digit recall as implemented in the Digits span subtest of the
Wechsler Intelligence Scale for Children (WISC-IV) (Wechsler 2011); 2)
switching: using the letter-number sequencing subtest from the Trail Making
Test from the Delis – Kaplan Executive Function System (DKEFS) (Delis
2011); y 3) inhibition: using the Color-word subtest (condición 3) del
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
13
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
t
/
.
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
.
t
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
DKEFS. Speed of processing was tested using the Coding and Symbol
search subtests of the WISC.
Selective visual attention was assessed using the Sky-search subtest of the
Test of Everyday Attention for Children (TEA-Ch) (Manly, Anderson et al.
2001).
Neuroimaging data
Neuroimaging data acquisition
The fMRI images were acquired using a Philips Ingenia 3 Tesla MRI scanner
(Philips Healthcare, Best, Países Bajos). Each fMRI session (pre-test and post-
1
2
3
4
5
6
7
8
9
prueba) era 13 minutes long and the repetition time (TR) era 1 segundo: un todo-
10
brain T2* functional volume was acquired every 1 second for a total of 780
volumes per session. The echo time (EL) era 30 EM. A field of view (FOV) de
20 X 20 X 14.4 cm, matrix of 80 X 80, and slice thickness of 3 mm were
11
12
13
utilized. Además, for each participant, whole-brain T1 images were acquired
14
in order to co-register the functional images to a high-resolution anatomical
imagen. The TR for the T1 scan was 8.1 EM, with a TE of 3.7 EM, inversion
time of 940 ms and a flip angle of 8 º. The FOV was 22.4 X 25.6 X 16 cm,
matrix of 224 X 256, and slice thickness of 1 mm.
Before the first fMRI session, all children were invited to explore the MRI
scanner environment and to practice laying down on the scanner bed “as still
15
16
17
18
19
20
as a statue” (Vannest, Rajagopal et al. 2014, Taran, Farah et al. 2021). Foam
21
pads were placed on either side of the head-coil apparatus in order to
minimize motion. Presentation of the stimuli was possible using an MRI-
compatible audiovisual system (Avotec, SS3150/SS7100).
22
23
24
14
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
t
/
/
.
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
t
/
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neuroimaging task
Participants performed the fluency task inside the scanner before and after
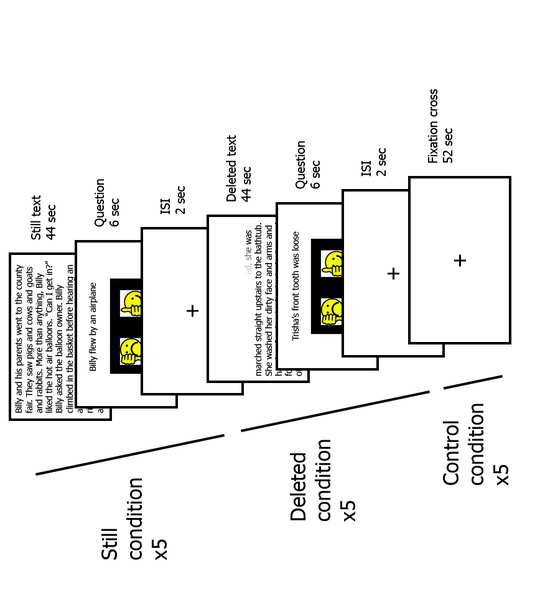
la intervención (ver figura 3). The fluency task is a reading task including
two different reading conditions: “Still condition”, in which a written story
appears on the screen for 44 artículos de segunda clase, and “Deleted condition”, in which the
presented text is deleted letter by letter starting from the first letter and is
completely deleted after 44 artículos de segunda clase. After the story, participants are
presented with a yes/no question based on the text. Response times and
accuracy are recorded. The Deleted condition was developed based on
accelerated reading in resemblance to the EFs-based reading intervention. A
constant deletion rate of 119 ms per letter was utilized in the Deleted
condición, which was previously reported to be the average reading rate of
children with and without dyslexia between the ages of 8 y 12 (Horowitz-
Kraus 2014, Taran, Farah et al. 2022). This reading rate was selected to
ensure that all participants could read the text passages without encountering
significant difficulties.
The written stories were between 200 y 250 characters in length. The fMRI
experiment was divided into 15 bloques (5 Still stories, 5 Deleted stories and 5
control blocks). The length of each block was 52 artículos de segunda clase: 44 seconds for the
story, 6 seconds for reading and answering the question and a 2 second-long
Inter Stimulus Interval (ISI) during which a fixation cross was presented in the
center of the screen. The control condition consisted in a fixation cross
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
presented in the center of the screen for 52 artículos de segunda clase. There were two different
23
sets of stories, one for the pre-intervention fMRI session and another one for
the post-intervention fMRI session. The text passages presented in the pre-
24
25
15
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
t
.
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
t
/
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
intervention and post-intervention did not differ in difficulty: text difficulty was
calculated based on sentence length, repetition of words, word length and
frequency (mean difficulty pre-test ± SD = 602 ± 90, post-test = 654 ± 102;
t(9)=1.32, p=.22).
1
2
3
4
Cifra 3. Graphical representation of the experimental fluency fMRI task
5
[Insert Figure 3 aquí]
Cifra 3. Graphical representation of the experimental fluency fMRI task
(Taran, Farah et al. 2021). Three different conditions were presented in an
interleaved fashion: Still, in which the participants were asked to read a still
texto, Deleted, which consisted in accelerated reading, and a Control condition,
consisting in a fixation cross presented in the middle of the screen. Cada
condition was presented 5 times in each session.
EFs-based reading training
The EFs-based reading training was developed based on the finding that
reading a text that is being deleted from the screen improves reading fluency
6
7
8
9
10
11
12
13
14
15
16
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
/
.
t
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
t
.
/
in both TR and individuals with dyslexia –fluency defined as fast and accurate
17
lectura (Breznitz, Shaul et al. 2013). Previous research suggests that this
kind of training improves reading fluency by boosting different cognitive
functions within the EFs domain such as working memory and inhibition, pero
also speed of processing and visual attention (Breznitz, Shaul et al. 2013).
Aquí, children trained three times per week (for eight weeks) for a total of 24
sessions: each session was 20 a 25 minutes long. In an initial session, el
reading speed of each participant was calculated. This was done by
presenting sentences on the screen (without deleting them) and once the
18
19
20
21
22
23
24
25
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
16
participant finished reading the sentences, they were instructed to push the
space bar which prompted a multiple-choice comprehension question.
Automated calculation of reading speed was performed by dividing the
reading time of each text passage by the number of characters in that specific
fragment of text. It is important to note that only the passages for which
participants answered the reading comprehension questions correctly were
included in this calculation. Once the reading pace was established, el
training phase started with a series of text passages that were being erased
from the screen at that individual’s reading speed. The training phase
consisted in reading short sentences (~100 characters) and answering a
multiple-choice reading comprehension question presented immediately after
each sentence. The characters within the sentence were being deleted from
1
2
3
4
5
6
7
8
9
10
11
12
the screen and replaced by asterisks starting from the first letter at a gradually
13
increasing pace; the erasure rate would increase only when the participant’s
14
answer to the reading comprehension question was correct on 10 consecutive
15
ensayos (Cecil, Brunst et al. 2021).
Math training (control group)
16
17
METRO
ath training was administered using an online math
tool
C
ustomized for each
18
participant’s grade level. The training encompassed various topics for third to
sexto
–
grade participants, including number properties, operaciones, y más.
Tercero
–
grade tra
ining covered number sense, addition, subtraction,
multiplicación, división, propiedades, mixed operations, fractions, geometría,
19
20
21
22
datos, and measurement. Cuatro
–
grade training incorporated all of the previous
23
temas, along with decimals and operations on frac
ciones. Quinto
–
grade training
included all previous topics, as well as exponents, powers of 10, datos,
24
25
17
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
.
/
/
t
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
/
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
medición, and operations on decimals. Sixth
–
grade training covered all
previous topics and more: number theory, estimations, ratios and rates,
ecuaciones,
percents, dinero, tiempo, graphs, and statistics. The training platform
was accessed through the website https://www.ixl.com/math.
The intervention
duration was similar to that of the
-sEF
based
reading training
:
three sessions
per week for eight weeks (totaling 24 sessions), with each session lasting
aproximadamente 20 a 25 minutos. This training did not have a speed
component in it
.
Data analyses
Behavioral data
In order to find baseline performance differences between TR and children
with dyslexia, independent two-sided Student’s t-tests were utilized. A
determine intervention-related differences in the behavioral tests (outlined in
the behavioral measures session), 2 Capacitación (Reading, Matemáticas) X 2 Time (Pre-
prueba, Post-test) repeated-measures ANOVA were conducted (Toffalini, Giofrè
et al. 2021). These analyses were conducted for the whole group as well as
for children with dyslexia and TR separately. All statistical analyses included
edad, sexo, and socioeconomic status as covariates.
Raw p-values were adjusted using Bonferroni’s procedure for multiple
comparisons. Significance value (alfa) was set to 0.05. The statistical
analysis was conducted using IBM SPSS statistics version 28 (IBM 2021).
Neuroimaging data
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
Image pre-processing was performed using Statistical Parametric Mapping 12
23
(SPM) (Friston, Penny et al. 2006) and CONN (version 20b) (Whitfield-
24
18
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
.
t
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
.
/
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Gabrieli and Nieto-Castanon 2012). The pre-processing of the fMRI data
included five different steps performed in order to adjust the fMRI volumes
and to increase the Signal-to-Noise Ratio (SNR). These steps included: 1)
realignment and motion correction, 2) outlier identification, 3) segmentation, 4)
normalization and co-registration and 5) smoothing. Motion is an especially
relevant noise source in young children (Power et al., 2012). De este modo, adicional
motion correction tools were utilized: aComprCor method for nuisance
regression was combined with scrubbing of consecutive functional volumes
with global signal changes of intensity above z=3 and/or framewise
1
2
3
4
5
6
7
8
9
displacement above 0.5 mm, in line with recent benchmarking
10
recomendaciones (círico, Wolf et al. 2017). It was confirmed that there were no
11
significant differences between the framewise displacements of the two
grupos. High pass and low pass filters were utilized in order to keep the fMRI
12
13
signal of interest for the fluency task: high pass filter 0.0096 Hz, low pass filter
14
0.165 Hz (Bijsterbosch, Smith et al. 2017). Only the Still and Deleted reading
conditions were included in the analysis (específicamente, only the 44 artículos de segunda clase
15
16
during which text was being read, leaving out the time of question reading and
17
respuesta). The rest condition was not included in the analysis.
Seed-to-voxel analyses were conducted on the fMRI data. Boxcar
hemodynamic response functions (HRFs) were generated to align with the
distinct blocks comprising the experimental design. These functions were
18
19
20
21
incorporated in the denoising and first-level analysis stages as implemented in
22
CONN version 20b (Whitfield-Gabrieli and Nieto-Castanon 2012) para el
calculation of task-residual functional connectivity between the chosen seed
regions and the rest of voxels in the brain (Tran, McGregor et al. 2018). A
23
24
25
19
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
t
/
.
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
.
t
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
previously described brain atlas developed based on anatomical and
functional meta-analyses was utilized for the definition of regions and
networks of interest (Fuerza, Cohen et al. 2011). The Power264 atlas includes
264 regions grouped in 14 redes. Four cognitive brain networks involved in
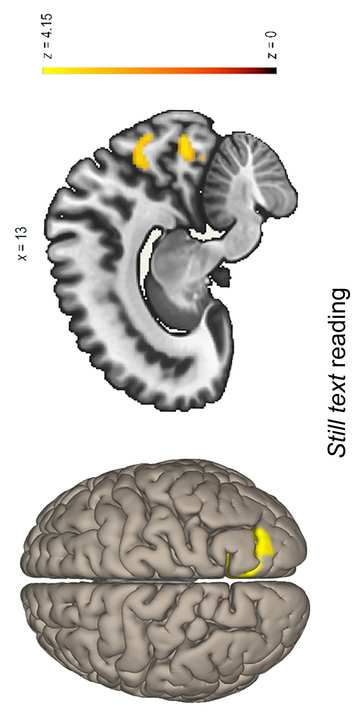
visual attention processing and EFs were chosen as the seeds (es decir., the DAN,
VAN, FP, and CO). One network was taken at a time and all of its nodes were
defined as one seed. Después, the correlation between the averaged
timeseries of all voxels within the seed and every other voxel in the brain was
computed, before exploring whether any brain region showed significant
correlations with the seed (for single level models) or whether any significant
change in connectivity existed between one condition and another (para
multilevel models). The voxels were reconstructed in 2 X 2 X 2 mm for the
análisis. Voxel-wise statistics throughout the whole brain were performed at
an FDR-corrected cluster level p<0.05. This procedure yielded multiple statistical parametrical functional connectivity maps for each seed. All neuroimaging results were corrected for multiple comparisons. The validity of the relations between neurobiological variables and behavioral traits drawn by brain-wide association studies (BWAS) has been questioned in a recent benchmarking paper (Marek, Tervo-Clemmens et al. 2022). Even though the present paper cannot be considered a BWAS, it does report of univariate brain-behavior associations, the reliability of which seems to be 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 lower than was initially thought (Marek, Tervo-Clemmens et al. 2022). Here, a 22 control permutation analysis was performed to increase the validity of the reported results: the sample (79 pre-post fMRI datasets) was randomly divided into four sub-groups (resembling the four experimental groups in the 23 24 25 20 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / / t e d u n e n a r t i c e - p d l f / d o i / / / . t 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d t . / f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 original 2 x 2 –group x condition– design) before seed-to-voxel analysis using the DAN as a seed. The 79 participants were divided into four groups using the rand MATLAB command. Marek and colleagues (2022), conclude that a random division of the subjects into different subgroups could induce spurious results that can contradict the actual obtained results. Here, null results were expected when comparing the randomized groups. Correlation between neuroimaging and behavioral data In order to evaluate the strength of the linear relationship between cognitive test results and functional connectivity values, correlative analyses were conducted. Functional connectivity values between different brain regions in which increases in connectivity strength were dected were tested as 1 2 3 4 5 6 7 8 9 10 11 predictors of cognitive performance. This was calculated: first, for the group in 12 which the increase was detected and second, for the whole sample. Linear correlation was examined using Pearson’s r correlation coefficient. Spurious significance resulting from multiple comparisons was controlled using Bonferroni’s correction. Structural equation modelling-based moderation analyses were conducted as well. Specifically, the areas showing enhanced synchronization after the reading training in the neuroimaging analysis were tested as moderators of the improvement in reading performance. Results Baseline behavioral measures (pre-test) Reading and verbal abilities 21 13 14 15 16 17 18 19 20 21 22 23 24 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . t / / e d u n e n a r t i c e - p d l f / d o i / / . t / 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d t / . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 The TR group consistently scored higher across reading measures, including phonological awareness, word reading, pseudoword reading, and reading comprehension (see Table 1). No significant differences were found between groups in receptive vocabulary or non-verbal intelligence (PPVT-4 number of errors, t=.905, p=.36; TONI percentile, t=1.8, p=.1). Executive function and attention abilities TR outperformed children with dyslexia in several EFs, including working memory, switching, and inhibition (see Table 1). Furthermore, there were significant differences between the groups in speed of processing and visual attention, with children with dyslexia displaying poorer performance. Table 1. Cognitive abilities before intervention. SD – standard deviation, TR – typical readers. Significant p-values are reported following APA guidelines: * p<0.05, ** p<0.01, *** p<0.001. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / t / e d u n e n a r t i c e - p d l f / d o i / . / t / 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d / t . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 22 The effect of the intervention on behavioral measures (pre-test vs post-test) 1 2 Ability Test TR group (n=43) Dyslexia group (n=36) Student’s t (p-value) Degrees of freedom Cohen’s d Mean SD Mean SD 11.35 1.94 8.00 2.77 6.11*** 61.07 1.4 8.83 2.241 6.78 2.11 4.15*** 104.37 11.14 81.94 12.52 8.42*** 105.33 9.91 84.75 10.88 8.79*** TOSREC (Index) 104.07 17.46 85.64 14.49 5.04*** GORT Comprehension (Percentile) 65.68 26.89 27.27 22.27 7.10*** TONI (Percentile) 66 19 55 21 1.8 Updating/Working Memory WISC Digit Span (Std. score) 11.4 2.72 9.72 2.17 2.98** 76 77 77 77 77 77 77 0.94 1.9 1.9 1.1 2 0.2 0.6 CTOPP Elision (Std. score) CTOPP Letter naming (Scaled score) TOWRE - SWE (Scaled score) TOWRE - PDE (Scaled score) Phonological Awareness Phonological Awareness Word reading Pseudoword reading Reading comprehension Reading comprehension Non-verbal intelligence Switching Inhibition Visual Attention Speed of processing DKEFS Trail Making Test Condition 4 (Std. Score) DKEFS Color Word Condition 3 (Std. Score) TEA-Ch Sky Search Attention (Percentile) WISC Symbol Search (Std. score) Reading abilities 10.72 2.75 7.39 4.27 3.89*** 51.56 0.9 10.65 2.76 8.18 3.42 3.53*** 77 35.16 25.9 23.79 19.72 2.13* 73 0.8 0.5 11.16 3.02 9.5 2.18 2.83** 75.48 0.6 Whole group analysis: ANOVA analysis of the pre-test and post-test scores showed a significant main effect of Time for contextual reading rate (F(1,78)=5.64, p<.05, ηp 2=.07, see Table 2), suggesting overall improvement in reading rate between the pre-test and post-test for all participants. No significant main effect of training was observed. Furthermore, the analysis revealed a significant Time × Training interaction for all participants (F(1,78)=4.88, p<.05, ηp 2=.06). This result suggests that children undergoing 10 23 3 4 5 6 7 8 9 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . t / / e d u n e n a r t i c e - p d l f / d o i / / / t . 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d t / . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 the EFs-based intervention improved their contextual reading rate more than those undergoing the math intervention, regardless of the presence of dyslexia. A partial Eta Squared of .06 and a Cohen’s f equal to .38 indicated a moderate-to-strong effect size. However, there was no significant Time, Training, nor Time × Training interaction effects on reading accuracy nor comprehension (when considering both children with dyslexia and TR together). Children with dyslexia: The results of the statistical analysis revealed significant effects of Time on word reading and reading rate for children with dyslexia, indicating that all participants with dyslexia improved their single word reading abilities and contextual reading speed, regardless of the administered intervention (F(1,34)=4.23, p<.05, ηp 2=.11, F(1,33)=5.9,p<.05, ηp 2=.15 for word reading and reading rate, respectively). Partial Eta Squared indices between .11 and .15 and Cohen’s f between .4 and .6 indicated a moderate-to-strong effect size. Furthermore, an analysis of variance 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 conducted on reading accuracy in children with dyslexia revealed a significant 16 interaction effect (Time × Training), indicating that participants with dyslexia who underwent EFs-based reading training improved their contextual reading accuracy scores more than those undergoing math training (F(1,33)=6.5, p<.05, ηp 2=.16). A partial Eta Squared of .16 and a Cohen’s f equal to .69 indicated a strong effect size. Typical readers: Children without dyslexia who underwent the EFs-based reading training improved their performance in reading rate more than those undergoing math training, as revealed by a significant ANOVA interaction 17 18 19 20 21 22 23 24 24 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . t / / e d u n e n a r t i c e - p d l f / d o i / / . t / 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d / t . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 effect (Time × Training F(1,41)=5.78, p<.05, ηp 2=.12). A partial Eta Squared equal to .12 and a Cohen’s f equal to .38 indicated a moderate effect size. EFs and attention abilities Whole group analysis : Significant main effects of Time were revealed for inhibition, switching, processing speed, and visual attention, indicating that all participants improved their performance, regardless of the training condition (F(1,75)=18.83, p<.001, 2 pη =.2; F(1,76)=11.87, p<.001, 2 pη =.14; and F(1,76)=11.87, p<.001, 2 pη =.14; and F(1,72)=29.4, p<.001, 2 pη =.29, respe ctively). However, there were no significant main effects of training nor was there evidence of differential . effects of training condition for the RD and TR groups (see Table 2) Statistical analyses of the pre-post scores on EFs tests revealed no significant differential effects of the training for the EFs-based reading training vs Math training on any EFs: working memory, switching, and . inhibition were tested (see Table 2) In order to test the effect of training on speed of processing, a Time x Training repeated measures ANOVA was conducted on WISC Symbol . Search scores The effect of the EFs-based reading intervention on visual attention scores was not significant as tested by 2x2 repeated measures ANOVA: Sky search Time × Training interaction F(1,72)=3.01, p=.09). Children with dyslexia: The subgroup of children with dyslexia displayed 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 significant improvements in inhibion and switching over time regardless of the 24 25 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . t / / e d u n e n a r t i c e - p d l f / d o i / / . t / 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d / t . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 intervention group that the belonged to (as revelaed by a significant ANOVA main effect of Time, see Table 2). ANOVA analysis of visual attention scores revealed that the interaction term (Time × Training) was significant for children with dyslexia (F(1,34)=5.127, p<.05, ηp 2=.14), suggesting larger improvements in visual attention scores after the EFs-based reading intervention when compared to the active control math intervention. The observed effect size for children with dyslexia was large (partial eta squared equaled .14, Cohen’s d was equal to .46). Typical readers: The subgroup of children without a diagnosis of dyslexia displayed significant increases in working memory performance, as revealed by a significant ANOVA main effect of Time (F(1,40)=5.72, p<.05, ηp 2=.13). Noteworthy, all participants displayed this improvement, regardless of the received treatment. Improvements in inhibition and speed of processing were 1 2 3 4 5 6 7 8 9 10 11 12 13 apparent as well, as revealed by significant main effects of Time (F(1,41)=9.3, 14 15 16 17 p<.01, ηp 2=.18 and F(1,41)=10.64, p<.01, ηp 2=.21, respectively). Table 2. The effect of the intervention on reading and cognitive abilities. Cohen’s f measure of effect size is reported for the significant Time x Training interactions 26 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . t / / e d u n e n a r t i c e - p d l f / d o i / . t / / 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d . / t f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 Ability Test Word reading TOWRE SWE Group Whole sample (79) TR (43) Math int. pre-test (mean, SD) 93.9(15.9) Math int. post-test (mean, SD) 93(20) EFs-based int. pre-test (mean, SD) 94.4(16.8) EFs-based int. post-test (mean, SD) 95(16.2) Main effect of Time Main effect of Training Time*Training Interaction F(1,77)=.19, p=.66, ηp 2=.002 F(1, 77)=.01, p=.9, ηp 2=.001 F(1,77)=.01, p=.91, ηp 2=.001 104.8(9.2) 100.8(22.8) 103.9(13.1) 103.3(14) DD (36) 79.9(10.8) 73(8.7) 83.7(13.8) 85.7(13.5) Pseudoword reading TOWRE PDE Whole sample TR 96.6(14.6) 94.9(16.1) 95.3(14.7) 94.3(22.5) 105.5(8.9) 105.7(10.3) 105.1(11.1) 105.7(11) DD 85(12.1) 81(9.7) 84.5(10) 84.5(15.5) Contextual reading rate GORT Reading Rate Contextual reading accuracy GORT Reading Accuracy Reading comprehension GORT reading comprehension Whole sample TR DD Whole sample TR 41.9(28.9) 41.7(27.6) 41.1(31.8) 49(28.6) 57.3(27.6) 61.6(27.6) 58.8(25.7) 65.8(19) 21.3(20.3) 24.5(18.2) 20.1(14.8) 27.2(20) 50.9(31.7) 50.1(32.5) 48.8(32.9) 49.4(28.7) 70.2(23.8) 70.9(24) 73.3(24.5) 65.6(26.2) DD 26(22) 23.7(20.4) 25.1(18.9) 32.3(20.4) Whole sample TR 50.3(31.8) 47.5(29.1) 46.5(31.7) 45.2(29.2) 65.8(28.6) 61.8(24.2) 68.2(25.2) 62(22.1) DD 30.1(23.5) 29.1(24.4) 27.4(22.8) 27.4(25.1) Working memory WISC Digit Span Whole sample 11.1(2.8) 11.3(2.9) 10.2(2.3) 10.5(3.3) 27 2=.11 2=.01 2=.02 F(1,41)=.66, p=.42, ηp F(1,34)=4.23, p=.04*, ηp F(1,77)=.76, p=.38, ηp F(1,41)=.13, p=.73, ηp F(1,34)=1.87, p=.18, ηp F(1,77)=5.64, p=.04*, ηp 2=.003 2=.07 2=.05 2=.02 2=.001 2=.001 F(1,41)=.05, p=.82, ηp F(1,34)=.73, p=.4, ηp F(1,77)=.04, p=.83, ηp F(1,41)=.15, p=.69, ηp F(1,34)=1.97, p=.17, ηp F(1,77)=.395, p=.53, ηp 2=.005 2=.004 2=.004 2=.005 2=.001 2=.009 F(1,41)=.36, p=.55, ηp F(1,34)=.18, p=.67, ηp F(1,77)=.05, p=.83, ηp F(1,41)=.18, p=.67, ηp F(1,34)=.17, p=.68, ηp F(1,77)=4.88, p=.02*, ηp 2=.005 2=.004 2=.06 2=.008 F(1,41)=.33, p=.57, ηp F(1,34)=5.9, p=.02*, ηp 2=.15 2=.001 F(1,41)=.02, p=.93, ηp F(1,34)=.02, p=.93, ηp 2=.001 2=.05 2=.04 2=.001 F(1,77)=.08, p=.78, ηp F(1,41)=1.7, p=.19, ηp F(1,34)=1.8, p=.18, ηp F(1,77)=2.2, p=.14, ηp F(1,41)=2.5, p=.12, ηp F(1,34)=.06, p=.8, ηp F(1,76)=1.1, p=.29, ηp 2=.002 2=.03 2=.06 2=.01 2=.01 2=.001 2=.001 F(1,77)=.02, p=.88, ηp F(1,41)=.03, p=.88, ηp F(1,34)=.34, p=.56, ηp F(1,77)=.1, p=.75, ηp F(1,41)=.03, p=.86, ηp F(1,34)=.07, p=.79, ηp F(1,76)=1.99, 2=.03 p=.16, ηp 2=.001 2=.001 2=.002 2=.12 F(1,41)=5.78, p=.02*, ηp F(1,34)=.83, p=.37, ηp 2=.03 2=.16 2=.06 2=.001 F(1,77)=.001, p=.97, ηp F(1,41)=2.5, p=.17, ηp F(1,34)=6.5, p=.01*, ηp F(1,77)=.01, p=.91, ηp F(1,41)=.1, p=.75, ηp F(1,34)=.06, p=.8, ηp F(1,76)=.01, p=.94, ηp 2=.003 2=.001 2=.002 2=.001 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . t / / e d u n e n a r t i c e - p d l f / d o i / / . t / 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d t . / f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 Inhibition DKEFS Color Word TR DD Whole sample TR 11.9(3) 12.3(3) 10.8(2.3) 12.1(3.2) 9.5(2.2) 8.7(2.2) 9.9(2.5) 10.1(2.5) 9.7(2.6) 10.5(2.5) 9.5(3.8) 10.5(3.6) 10.9(2) 11.6(1.9) 10.4(3.4) 11.4(2.9) DD 7.9(2.4) 9.2(2.6) 8.5(4.1) 9.5(3.9) Switching Trail Making Test Whole sample 9.5(3.8) 10.8(3.8) 9.1(3.9) 10.2(3.8) Speed of processing WISC Symbol Search Visual attention TEA-Ch Sky search TR DD Whole sample TR DD Whole sample TR 10.6(3.2) 12(2.8) 10.8(2.2) 11.14(3.3) 7.8(4.2) 10.2(3.5) 7.3(4.4) 9(3.9) 10.3(2.5) 11.5(2.7) 10.5(3.1) 11.4(3.4) 10.9(2.6) 12.6(2.8) 11.4(3.5) 12.4(3.7) 9.6(2) 10.1(2) 9.5(2.4) 10.3(2.4) 7.6(3.1) 10(6.4) 7.4(2.7) 10(3) 8.2(3.1) 11.3(4.1) 8(2.5) 10.6(2.7) DD 6.9(3) 8.4(3) 6.7(2.7) 9.5(3.4) Notes: F=ANOVA F term. *p<.05, **p<.01, ***p<.001 28 2=.13 2=.03 F(1,40)=5.72, p=.02*, ηp F(1,34)=.99, p=.33, ηp F(1,75)=18.83, p<.001***, ηp F(1,41)=9.3, p=.004**, ηp F(1,32)=9.54, p=.004**, ηp F(1,73)=12.6, p<.001***, ηp 2=.15 2=.18 2=.23 2=.2 2=.02 2=.001 F(1,40)=.66, p=.19, ηp F(1,34)=1.77, 2=.05 p=.19, ηp F(1,75)=.05, p=.82, ηp F(1,41)=.21, p=.65, ηp F(1,32)=.15, p=.69, ηp F(1,73)=.95, p=.33, ηp 2=.005 2=.005 2=.01 2=.04 2=.05 2=.001 F(1,40)=1.77, p=.19, ηp F(1,34)=1.75, p=.19, ηp F(1,75)=.08, p=.78, ηp F(1,41)=.42, p=.52, ηp F(1,32)=.083, p=.77, ηp F(1,73)=1.16, p=.28, ηp 2=.003 2=.02 2=.01 2=.08 F(1,41)=3.8, p=.06, ηp F(1,30)=8.89, p=.006**, ηp F(1,76)=11.87, p<.001***, ηp 2=.23 2=.14 2=.005 F(1,41)=.19, p=.66, ηp F(1,30)=.43, p=.52, ηp F(1,76)=.007, p=.93, ηp 2=.001 2=.01 2=.03 F(1,41)=1.4, p=.24, ηp F(1,30)=.31, p=.58, ηp F(1,76)=.12, p=.73, ηp 2=.002 2=.01 F(1,41)=10.64, p=.002**, ηp 2=.21 F(1,41)=.03, p=.86, ηp 2=.001 F(1,41)=.45, p=.51, ηp 2=.01 F(1,33)=2.19, p=.15, ηp 2=.06 F(1,33)=.003, p=.96, ηp 2=.001 F(1,33)=.08, p=.78, ηp 2=.002 2=.29 F(1,72)=29.4, p<.001***, ηp F(1,36)=14.68, p<.001***, ηp F(1,34)=17.53, p<.001***, ηp 2=.29 2=.34 2=.002 F(1,72)=.12, p=.73, ηp F(1,36)=.57, p=.46, ηp F(1,34)=.217, p=.64, ηp 2=.006 2=.02 2=.001 F(1,72)=.08, p=.78, ηp F(1,36)=.77, p=.39, ηp F(1,34)=5.13, p=.02*, ηp 2=.14 2=.02 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / / t e d u n e n a r t i c e - p d l f / d o i / t / / . 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d t / . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 1 2 3 Fluency fMRI task Baseline (pre-test) Accuracy: TR outperformed children with dyslexia in both Still and Deleted conditions (see Table 3). The number of correct responses was statistically lower in children with dyslexia when compared to TR in both conditions. There were no significant differences between the children who later underwent EFs-based reading training and those in the math group (p>.05).
Reaction time: TR showed shorter response times when compared to children
with dyslexia in both conditions (ver tabla 3). No significant differences were
found between the intervention groups (EFs-based reading intervention vs
math intervention) in the baseline reaction time (p>.05).
Mesa 3. Baseline performance in the fluency task; significant differences between
children with dyslexia and TR.
Children with
dyslexia
Typical
Readers
Student’s t (p-value)
Cohen’s d
Still
Correct
3.710 ± 1.29
4.27 ± 1.11
Response
Time
4136 ± 1020
3580 ± 961
Deleted
Correct
4.05 ± .97
4.79 ± .67
Response
Time
4107 ± 856
3422 ± 859
2.06, .02*
Cohen’s d=.47
2.47, .008*
Cohen’s d=.56
3.93, <.001*** Cohen’s d=.89 3.5, <.001*** Cohen’s d=.79 Independent samples t-tests indicate significant differences between TR and children with dyslexia in the task. *p<.05, **p<.01, ***p<.001. Cohen’s d measure of effect size is reported. Pre-intervention vs post-intervention Accuracy: No significant differences in accuracy were found in any of the four subgroups. Two by two (2 x 2) repeated measures ANOVA –Time (pre- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 29 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / t / e d u n e n a r t i c e - p d l f / d o i / . / t / 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d . t / f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 test/post-test) x Training (EFs-based reading int./Math int.) – did not show any significant main effects nor significant interaction effects (see Table 4). Reaction time: Overall, all participants undergoing EFs-based reading intervention showed significantly greater response times to the Still condition reading comprehension questions after the intervention (see Table 4). Furthermore, TR (but not children with dyslexia) undergoing Math intervention showed increased response times to the Still condition comprehension questions in the post-test when compared to the pre-test. Longer response times in the post-test were associated with increases in accuracy, measured as the number of correct responses (Pearson’s r=-.43, r2=.18, p<.001). In other terms, children demonstrating increases in response time to the still condition (between the pre-test and the post-test) displayed higher accuracies in the same condition (in the post-test). This association suggested a possible relation between the change in response times and the intervention-related change in EFs. In line with the a posteriori formulated hypothesis, a significant correlation was found between pre-post change in 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 response time during the Still condition and pre-post change in inhibition in all 17 participants (Pearson’s r=-.31, r2=.09, p=.01). Specifically, the participants showing larger improvements in inhibition after the intervention displayed longer response times after the intervention. Table 4. Pre-post differences in Fluency task performance. 18 19 20 21 22 23 Pre-test Still Response time (ms) Correct answers Deleted Response time (ms) Correct answers Post-test Still Response time (ms) Correct answers Deleted Response time (ms) Correct answers Contrast t-test (p-value), Cohen’s d 30 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / t / e d u n e n a r t i c e - p d l f / d o i / / t . / 1 0 1 1 6 2 n e n _ a _ 0 0 3 3 5 2 1 5 6 8 1 1 n e n _ a _ 0 0 3 3 5 p d t . / f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 Typical readers EFs-based reading intervention 3553 ± 1097 (A) 4.09 ± 1.37 3330 ± 1014 4.7 ± .9 4147 ± 1012 (B) 3.95 ± 1.09 3236 ± 845 4.7 ± .47 Math intervention 3606 ± 836 (C) Children with dyslexia EFs-based reading intervention 4267 ± 1019 (E) Math intervention 3980 ± 1032 4.45 ± .8 3.47 ± 1.42 4 ± 1.1 3510 ± 693 4204 ± 721 3703 ± 971 4.86 ± .35 4.1 ± .9 4055 ± 693 (D) 4833 ± 965 (F) 3.95 ± 1.14 3 ± 1 3296 ± 981 3936 ± 953 4.6 ± .94 4.6 ± .91 4 ± .96 4423 ± 1239 3.33 ± 1.15 3703 ± 971 4.58 ± .9 B>A
D>C
F>E
3.15**
Cohen’s d=.71
2.5*
Cohen’s d=.55
2.47*
Cohen’s d=.63
Paired t-tests indicate higher response times in the fluency task after the EFs-based
reading intervention. *pag<.05, **p<.01, ***p<.001. Cohen’s d measure of effect size is
reported.
Neuroimaging results
Baseline connectivity analysis (pre-test)
Baseline differences found between children with dyslexia and TR in the
connectivity patterns of attention-related brain networks were reported
previously (Taran et al., 2021), and are outside the scope of the present
paper, which focuses on intervention-related brain connectivity changes. For
this reason, only pre-post results are presented here.
The effect of intervention on functional connectivity (pre-test vs post-test)
Visual Attention networks
Dorsal attention network (DAN)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
Still condition: No differences in brain connectivity while reading Still text were
17
found in the DAN when comparing baseline and post-intervention seed-to-
voxel statistical parametric maps.
Deleted condition: Following intervention, TR who underwent the EFs-based
reading training showed an increase in functional connectivity strength
between the DAN and the left IFG while reading Deleted text, in comparison
18
19
20
21
22
31
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
to TR who underwent the math intervention, as revealed using 2x2 (Time x
Training) repeated measures ANOVA (T(41)=3.54, cluster p-FDR<.05, Table
5, Figure 4). No differences were found between the connectivity patterns of
the DAN in children with dyslexia when comparing the pre-intervention and
the post-intervention.
A complete graphical representation of the seed-to-voxel results in the DAN
can be found in Supplementary Figure 1.
Ventral attention network (VAN)
No results were found in the VAN when comparing the connectivity pattern of
this network with the rest of the brain before and after the trainings neither in
Still nor Deleted conditions.
EFs networks
Cingulo-opercular network (CO)
Still: Children with dyslexia who underwent the EFs-based reading
1
2
3
4
5
6
7
8
9
10
11
12
13
14
intervention showed a higher functional connectivity between the CO, the right
15
cuneus, and the right lingual gyrus in the Still condition, in comparison to
those undergoing math intervention (T(34)=3.6, cluster p-FDR corrected<.05,
see Table 5, Figure 5). In addition, in children with dyslexia who underwent
the EFs-based reading intervention there was a higher functional connectivity
between the CO and the right and left precentral gyrus after the intervention
(Supplementary Figure 1).
Deleted: No significant results were found in the seed-to-voxel analysis of the
Deleted condition.
Fronto-parietal network (FP)
32
16
17
18
19
20
21
22
23
24
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
.
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
No significant results were found when analyzing the FP network, neither in
the Still nor the Deleted conditions.
Table 5. Seed-to-voxel results.
1
2
3
4
Seed
Group
Contrast
Functional connectivity
MNI Coordinates
x
y
z
Cluster size
(voxels)
DAN
TR
EFs-based int. > Matemáticas
int.,
Deleted T2 > Deleted T1
Left inferior frontal gyrus
-48
10
8
313
VAN
Children
con
dyslexia
TR
Children
con
dyslexia
FP
TR
Children
con
dyslexia
CO
TR
Children
con
dyslexia
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
EFs-based int.
Still T2 > Still T1
EFs-based int. > Math
int.
Still T2 > Still T1
Right/Left precentral gyrus
-12
-24
72
178
Right lingual gyrus
20
-76
-4
Right cuneus
10
-82
28
299
245
Statistical threshold; cluster-level p-FDR corrected <.05, voxel-level uncorrected p<.001. Significant increases in functional connectivity were found in the CO and the DAN after the EFs-based reading intervention, but not the math intervention (active control group). [Insert Figure 4 here] Figure 4. Seed to voxel analysis (DAN)– Typical Readers. Seed to voxel analysis result. Cluster-level p-FDR corrected<.05. The seed is the DAN, contrast is EFs-based reading intervention>Math intervention, Deleted
Post>Deleted Pre. Left: 3D brain render with a 50% transparencia, superior
vista. Right: sagittal slice.
[Insert Figure 5 aquí]
33
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
.
/
/
t
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
norte
mi
norte
_
a
_
0
0
3
3
5
pag
d
t
/
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 5. Seed to voxel analysis (CO) – Children with dyslexia. Seed to
voxel analysis result. Cluster-level p-FDR corrected<.05. The seed is the CO network, contrast is EFs-based reading intervention> Math intervention, Still
Post>Still Pre. Left: 3D brain render, superior view. Right: sagittal slice.
Random classification
After randomly dividing the participants into four groups, the connectivity
between the DAN and the rest of the brain was calculated. There was an
increase in functional connectivity strength between the DAN and an occipital
cluster comprising the left lingual gyrus in one of the random groups when
comparing the pre-test and post-test Still text reading conditions (ver
Supplementary figure 1).
When comparing the pre-test and post-test data of the random groups, No
l
PAG<0.001
differences between random groups 1 and 2 were found (all groups included
both TR and children with dyslexia undergoing both interventions, see
Supplementary figure 1). In the same vein, no significant differences were
found between groups 3 and 4. Crucially, no differences between the
randomly created groups were observed.
Correlation between behavioral and neuroimaging data
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
A positive correlation between the DAN and the left IFG during Deleted text
20
reading and two cognitive measures was found in the group of TR –in the two
21
training groups and the two timepoints: working memory (r=.27, R2=.03,
p<.05), and speed of processing (r = .24, R2= .06, p<.05) (see Figure 6). This
functional connectivity index was not correlated with working memory nor
speed of processing in the group of children with dyslexia (working memory r
22
23
24
25
34
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
.
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
= .15, R2= .02, p=.24, speed of processing r = .04, R2= .001, p=.71). In the
same vein, the increase in connectivity strength DAN-left IFG after the
intervention was found to be a significant moderator of the change in reading
fluency for all TR (beta = 2.02, se=.94, t(3)=2.14, p<.05, see Figure 7). In this
moderation model, the main effect of both independent variables on the
outcome variable were significant (GORT fluency before intervention t=9.55,
p<.001; delta DAN – left IFG t = 2.51, p<.05). This model was not significant in
the group of participants with dyslexia.
When exploring the behavioral correlates of the increased functional
connectivity between the CO, the right cuneus, and the right lingual gyrus
during the Deleted condition in children with dyslexia, a positive correlation
was found with two cognitive variables: speed of processing (r=.27, R2=.073,
p<0.05), and visual attention (r=.31, R2=.095, p<0.05, Figure 8). The increase
1
2
3
4
5
6
7
8
9
10
11
12
13
14
in functional connectivity between the CO and the mentioned occipital regions
15
was not a statistical predictor of the change in reading fluency in children with
dyslexia (beta = .156, se=.13, t(3)=1.3, p=.21). No significant correlations
were found between the CO-right occipital cortex functional connectivity and
behavioral measures in the TR group.
[Insert Figure 6 here]
Figure 6. Correlation between the neuroimaging and behavioral results
in Typical Readers. Higher synchronization between the DAN and the left
IFG was related to increased performance in working memory and speed of
processing.
[Insert Figure 7 here]
35
16
17
18
19
20
21
22
23
24
25
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Simple moderation model. The change in connectivity between
the DAN and the left IFG between the pre-test and the post-test was a
significant moderator of the reading improvement in Typical Readers,
regardless of the intervention they received. Beta expressed as
unstandardized regression coefficients. *p<.05, ***p <.001.
[Insert Figure 8 here]
Figure 8. Correlation between the neuroimaging and behavioral results
in children with dyslexia. Higher functional connectivity between the CO, the
right lingual gyrus, and the right cuneus was related to increased performance
in visual attention and processing speed tasks.
Discussion
Here, the specific effect of an EFs-based reading intervention vs a control
1
2
3
4
5
6
7
8
9
10
11
12
math training were examined on the behavioral level and the neural correlates
13
associated with EFs, visual attention, and reading in 8-12 years old children
with dyslexia and TR. In line with our hypotheses, we found significant
improvements after the EFs-based reading intervention in reading rate and
14
15
16
visual attention. However, we could not find a specific effect of the EFs-based
17
reading intervention on relevant EFs (working memory, switching, inhibition),
when comparing it to math training. On the neurobiological level, we found
two different mechanisms related to the EFs-based reading intervention; in
TR, higher connectivity was observed between the DAN and the left IFG.
Children with dyslexia displayed an increase in integration between a
cognitive control network (CO) and visual processing related-areas. In both
18
19
20
21
22
23
36
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
cases, the increase in connectivity positively correlated with performance in
different cognitive tasks.
Improvements in reading and visual attention after the EFs-based reading
intervention
At the baseline, children with dyslexia displayed deficits in visual attention,
executive functions, phonological processing, word reading, and contextual
reading when compared to TR. These impairments were not explained by
reduced scores in standardized intelligence tests.
After the EFs-based reading intervention (compared to math intervention),
improvements in children with dyslexia were apparent in both contextual
reading rate and visual attention. Higher scores were observed as well in the
TR group after the EFs-based reading intervention, but only in contextual
reading rate (when compared to those undergoing math training). These
results suggest that the EFs-based reading intervention effectively improves
the reading skills and visual attention abilities of children with dyslexia.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
However, it was not possible to find a specific effect of the EFs-based reading
17
intervention on EFs when comparing it to math intervention. Mathematical
18
training, and arguably any kind of complex cognitive training, may involve one
19
or more EFs given that these skills are the very basic psychological abilities
underlying complex behavioral tasks –those that result from the combination
of more basic mental processes (American Psychiatric Association 2007).
Mathematical reasoning, long-term planning, writing, or reading are some
examples of these complex behavioral tasks that rely on the harmonic
20
21
22
23
24
37
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
functioning of fundamental cognitive processes, many of which fall within the
umbrella term Executive Functions.
The improvement in visual attention abilities after the EFs-based reading
intervention in children with dyslexia confirms two of our assumptions. First,
the EFs-based reading intervention triggers not only EFs but also basic
features of visual processing (Horowitz-Kraus, Vannest et al. 2014). Second,
the training of visual attention abilities may be of high relevance in children
with dyslexia and could lead to improvements in reading (Vidyasagar 2019).
This manuscript presents evidence of improvements in visual attention and
reading rate following an EFs-based reading intervention. Visual attention
and EFs were found to be correlated with the intervention-related change in
functional connectivity (in children with dyslexia). However, no causal
relations were found between improvements in visual attention or EFs and
improvements in reading ability. As aforementioned, the math intervention
may have also targeted basic cognitive abilities within the EFs and visual
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
attention domains as well as the EFs-based reading intervention, which could
16
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
/
t
.
be the reason that prevented determining more specific effects of the EFs-
based intervention. Another limitation was the use of a single visual attention
measure (visual search or selective visual attention). The utilization of
17
18
19
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
different visual attention tasks with a closer relation to decoding may result in
20
the determination of better associations between improvement in visual
attention and reading performance. Future intervention studies, including a
waiting list (passive) control group might be able to draw robust links
between the training of EFs and visual attention and improvements in
21
22
23
24
38
reading. Alternatively, the investigation of non-linguistic cognitive training
aimed at improving EFs and/or visual attention can shed light on this issue.
Participants with dyslexia showed a significant improvement in reading
accuracy after the EFs-based reading intervention (compared to the control
training), but not in reading fluency or reading comprehension. This might be
due to the control training utilized here (math) and the well-established
importance of EFs in math performance (Bull and Lee 2014, Cragg and
Gilmore 2014). It is possible that Math training improved EFs, which, per the
current study’s hypothesis, had an effect on reading abilities. Future studies,
including active and passive control groups may be able to test the effect of
EFs-based reading training and visual training for dyslexia. Experimental
studies investigating interventions for dyslexia with variable loads of EFs and
visual attention (perhaps adopting a parametric approach with different
subjects receiving different weights for each component) might enhance our
understanding of the importance of each one of these factors in the treatment
of dyslexia.
Longer response times in the Fluency task after the intervention
Significant increases in response time were observed in the Still condition in
all participants who underwent EFs-based reading intervention. Furthermore,
only TR (not children with dyslexia) displayed longer response times in the
post-test compared to the pre-test after the math intervention. It has been
previously suggested that in tasks involving EFs, longer response times are
typically associated with superior performance (Partchev, De Boeck et al.
2013, Goldhammer, Naumann et al. 2014, Su and Davison 2019). On the
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
39
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
contrary, longer response times in tasks that involve lower-level processing
indicate lower ability (Partchev, De Boeck et al. 2013). These studies imply
that increased response times during reading tasks can reflect a deliberate
effort by participants to perform adequately (greater involvement of EFs). The
present results confirmed this hypothesis, given the significant association
between longer response times (when comparing the pre-test and the post-
test) and enhanced performance in the post-test. Here, a significant
correlation was found between pre-post changes in inhibition and pre-post
changes in response time to the still condition in all participants. That is, the
1
2
3
4
5
6
7
8
9
participants showing larger score improvements in inhibition after the
10
intervention displayed longer response times after the intervention, which is in
11
line with the presented explanation. In this vein, longer response times after
the intervention may be related to increases in cognitive inhibition, which is
involved in response monitoring (Kilian, Bröckel et al. 2020). In the present
study, all groups who were expected to demonstrate reading improvement in
response to the EFs-based reading intervention, including both typically
developing readers and those with dyslexia, exhibited longer response times.
In contrast, the group of children with dyslexia who received the Math
intervention did not improve, which is consistent with the presented post-hoc
explanation.
Notably, this effect was not observed in the Deleted condition, where no
significant differences in response time nor accuracy were found for any
12
13
14
15
16
17
18
19
20
21
22
group. It is plausible that participants dedicated more time to ensure accuracy
23
in the Still condition due to a reduced sense of urgency to respond rapidly,
24
40
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
while the accelerated presentation pace of the Deleted condition induced an
equally fast response.
Greater synchronization between EFs networks and visual processing regions
after the EFs-based reading intervention
Stronger functional connectivity was found in children with dyslexia between
the CO, the right lingual gyrus, and the right cuneus after the EFs-based
reading training. The cuneus and lingual gyrus comprise the medial occipital
lobe and have a role in basic and higher-level visual processing (Allison,
Begleiter et al. 1993, Mai and Paxinos 2012, Palejwala, Dadario et al. 2021).
More specifically, the cuneus and lingual gyrus seem to have special
relevance in visual memory, linguistic processing (written words), direction
1
2
3
4
5
6
7
8
9
10
11
discrimination, and motion perception (Palejwala, Dadario et al. 2021). Taking
12
into account the role of these areas in word decoding and movement-related
features of visual processing, it seems reasonable to assume that the fast-
paced deleted letters characterizing the EFs-based reading intervention
triggered a higher synchronization between the mentioned visual processing
13
14
15
16
regions and a higher-order cognitive control network in order to achieve better
17
performance at the task. This increase in functional connectivity is interpreted
as an indicator of higher integration of the basic (and not-so-basic) cognitive
processes underlying fluent reading, i.e., visual attention, working memory,
speed of processing, and more (Dehaene 2009, Sporns 2012). Here, a
positive correlation was found between these regions’ connectivity and the
scores in visual attention and speed of processing.
18
19
20
21
22
23
41
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Despite the major role of the left fusiform gyrus (or VWFA) in word decoding,
we did not find an association between this region and the EFs-based
intervention-related cognitive gain. Here, children with dyslexia displayed an
increase in connectivity strength between the CO and occipital areas involved
in visual processing, but not the the VWFA. The lingual gyrus and the right
cuneus possess certain processing features that allow them to engage in the
primary processing of visual information, further feeding into the
(hierarchichally superior) occipito-temporal VWFA. The present results
suggest that the decoding deficit in dyslexia may arise in early stages of visual
processing, which is not in synchrony with higher-order cognitive control
networks. This benefitial pattern of connectivity was reinforced after the EFs-
based reading training. How are the interactions between the VWFA and the
rest of the brain affected in dyslexia and how can a treatment address this
connections is a matter of further research.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
Recent studies suggest that one of the mechanisms of brain maturation along
15
development is a higher synchronization between whole-brain networks and
versatile neural hubs such as the IFG (Wierenga, van den Heuvel et al. 2016,
Hermosillo 2022). The medial occipital lobe, and specifically the bilateral
lingual gyri, seem to adopt a major role as an association hub along
development (Chen, Liu et al. 2013, Oldham and Fornito 2019). By boosting
the synchrony between the CO and the medial occipital lobe, the EFs-based
reading intervention might be enforcing a pattern of brain maturation in
16
17
18
19
20
21
22
children with dyslexia (Hermosillo 2022), which might help them overcome the
23
asynchronization reported earlier (Breznitz 2003) and reduce neural noise in
their sensory systems (Hancock 2017).
42
24
25
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
.
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Greater engagement of attention and linguistic/multimodal regions in TR after
the EFs-based reading intervention
A different neuroplasticity mechanism was found in TR children who
underwent the EFs-based reading intervention. In this subgroup, higher
functional connectivity was detected between the DAN and the left IFG while
reading deleted text. The left IFG is associated with phonological and
semantic functions (Klaus and Hartwigsen 2019). The role of this region in
word comprehension and production is undisputed (Costafreda, Fu et al.
2006, Klaus and Hartwigsen 2019). Furthermore, it is now well established
that the left IFG is a neuronal hub playing a role in multiple interrelated lower-
and higher-order cognitive operations such as multisensory integration
(Pulvermüller 2013, Li, Seger et al. 2020), verbal working memory
(Costafreda, Fu et al. 2006, Emch, von Bastian et al. 2019), creativity (Khalil,
Karim et al. 2020), and inhibitory control (Swick, Ashley et al. 2008,
Tomiyama, Murayama et al. 2022). In the same vein, the bilateral inferior
frontal gyri participate in distinct large-scale networks such as the language
network (Pulvermüller 2013, Tomasello, Garagnani et al. 2017, Gao, Guo et
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
al. 2020), the VAN (Corbetta and Shulman 2002, Bernard, Lemee et al. 2020),
18
and the limbic network (Cha, DeDora et al. 2016, Rolls, Cheng et al. 2020),
with broad connections to the temporal and parietal cortices (Nakae,
Matsumoto et al. 2020). The increase in strength of functional connectivity
between the DAN and the left IFG found after the EFs-based reading
intervention suggests that our training paradigm enforced in TR a maturation
19
20
21
22
23
in brain network interactions typical of the late childhood period (Sporns 2011,
24
Oldham and Fornito 2019).
25
43
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Higher synchrony between the DAN and the left IFG was related to higher
scores in verbal working memory, which is a cognitive ability that heavily
relies on the frontal lobe and the language network, including the left IFG
(Dehaene 2009, Emch, von Bastian et al. 2019). The involvement of the DAN
in visuo-attentional processing (Szczepanski and Kastner 2013, Zhao, Wang
et al. 2022) makes it a major candidate to contribute to the reading process,
and several studies have indeed highlighted the role of (especially posterior)
DAN areas in reading (Cohen, Dehaene et al. 2008, Qian, Bi et al. 2016).
However, the specific role of the different areas comprising the dorsal
1
2
3
4
5
6
7
8
9
attention system in the processing of printed words is not yet completely clear.
10
Our results suggest that higher synchrony between the DAN and the left IFG
is related to higher cognitive performance in speed of processing, working
memory and reading in TR, and that the EFs-based reading intervention is
11
12
13
capable of inducing increases in functional connectivity between these areas.
14
No significant differences when randomly dividing the sample
Here, we included an additional analysis in which random groups were
created and the seed-to-voxel analysis was repeated in order to check for the
validity of our results. One pre-post test contrast reached significance in one
of the subgroups. An increase in connectivity after the training in both TR and
15
16
17
18
19
participants with dyslexia undergoing either intervention could be representing
20
a mechanism common to both interventions. Alternatively, it could be a
neurobiological change related to normative development. In any case, it
remains unanswered why this change in connectivity is visible in one of the
subgroups only and this result is arguably an indicator of the spurious effects
that can be found performing fMRI analyses when running undirected
21
22
23
24
25
44
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
contrasts. Crucially, no differences were found between any of the random
groups, arguing in favor of the empirical strength of the results presented in
the present paper.
Limitations
Recent benchmarking papers have criticized the poor reliability of BWAS
(Baykara, Könen et al. 2021, Marek, Tervo-Clemmens et al. 2022). Here, we
did not explicitly conduct a BWAS but we did include univariate prediction
models for brain-behavioral phenotypes. Even though we tried addressing this
limitation by performing an extra control analysis (group permutation), the
need for larger sample sizes in order to increase the reliability and
generalizability of the associations in the field of cognitive neuroscience is an
1
2
3
4
5
6
7
8
9
10
11
unavoidable reality. Furthermore, current recommendations highlight the need
12
to move towards individualized treatment approaches when addressing
cognitive traits in clinical populations (Baykara, Könen et al. 2021). The
utilization of different active as well as passive (waiting list) control groups
might produce better outcomes when studying the neuropsychological effect
of an intervention program; we were not able here to isolate the effect of the
EFs-reading intervention on executive functions themselves because of the
recruitment of EFs in mathematical thinking. In summary, individualized
programs combining features of the EFs-based reading intervention (and
other phonological and visual trainings), might be the most fruitful research
direction for the development of treatment plans for dyslexia in upcoming
years.
Conclusion
45
13
14
15
16
17
18
19
20
21
22
23
24
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The EFs-based reading intervention improved the reading skills of children
with dyslexia and TR, and two different neuroplasticity mechanisms were
observed. In TR, the development of more mature brain connectivity patterns
between a large-scale network (i.e., DAN) and a multimodal hub (i.e., left IFG)
seemed to be mediating the reading fluency improvement. In children with
dyslexia, the synchronized activity of visual processing areas (which have
also been suggested to act as integration hubs) (Chen, Liu et al. 2013), and
performance monitoring cognitive control systems was related to higher
performance in low-level (i.e., visual attention) and higher-order cognitive
tasks (i.e., working memory) that are foundational for reading. These findings
point at the importance of addressing EFs and visual attention in the
1
2
3
4
5
6
7
8
9
10
11
development of interventions aimed at the improvement of reading in dyslexia.
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
46
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Supplementary figure 1. Seed to voxel analysis results. Significant results were
found for the DAN and the CO network. A control randomized analysis was carried
out on the DAN. See next page.
[Insert Supplementary Figure 1 here]
References
Allison, T., A. Begleiter, G. McCarthy, E. Roessler, A. C. Nobre and D. D.
Spencer (1993). "Electrophysiological studies of color processing in human
visual cortex." Electroencephalography and Clinical Neurophysiology/Evoked
Potentials Section 88(5): 343-355.
American Psychiatric Association (2007). APA Dictionary of Psychology.
Washington, DC, US, American Psychological Association.
Barbosa, T., C. C. Rodrigues, C. B. Mello, M. Silva and O. F. A. Bueno
(2019). "Executive functions in children with dyslexia." Arq Neuropsiquiatr
77(4): 254-259.
Baykara, E., T. Könen, K. Unger and J. Karbach (2021). "MRI Predictors of
Cognitive Training Outcomes." Journal of Cognitive Enhancement 5(2): 245-
258.
Bernard, F., J.-M. Lemee, E. Mazerand, L.-M. Leiber, P. Menei and A. Ter
Minassian (2020). "The ventral attention network: the mirror of the language
network in the right brain hemisphere." Journal of Anatomy 237(4): 632-642.
Bijsterbosch, J., S. M. Smith and C. Beckmann (2017). An Introduction to
Resting State FMRI Functional Connectivity. Oxford, UK, Oxford University
Press.
Booth, J., J. Boyle and K. Steve (2010). "Do tasks make a difference?
Accounting for heterogeneity of performance of children with reading
difficulties on tasks of executive function: Findings from a meta‐analysis -
Booth - 2010 - British Journal of Developmental Psychology - Wiley Online
Library." British Journal of Developmental Psychology 28(1): 133-176.
Breznitz, Z. (2006). Fluency in Reading: Synchronization of Processes.
Mahwah, New Jersey, Lawrence Erlbaum Associates.
Breznitz, Z., & Misra, M. (2003). "Speed of processing of the visual-
orthographic and auditory-phonological systems in adult dyslexics: the
contribution of "asynchrony" to word recognition deficits." Brain Lang 85(3):
486-502.
Breznitz, Z., & Share, D. L. (1992). "Effects of accelerated reading rate on
memory for text." Journal of Educational Psychology 84(2): 193-199.
Breznitz, Z., S. Shaul, T. Horowitz-Kraus, I. Sela, M. Nevat and A. Karni
(2013). "Enhanced reading by training with imposed time constraint in typical
and dyslexic adults." Nature Communications 4(1): 1-6.
Breznitz, Z., S. Shaul, T. Horowitz-Kraus, I. Sela, M. Nevat and A. Karni
(2013). "Enhanced reading by training with imposed time constraint in typical
and dyslexic adults." Nat Commun 4: 1486.
Brown, L., R. Sherbenou and S. Johnsen (2010). Test of Nonverbal
Intelligence. Fourth Edition. Austin, TX, PRO-ED.
47
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bull, R. and K. Lee (2014). "Executive Functioning and Mathematics
Achievement." Child Development Perspectives 8(1): 36-41.
Catts, H. W. (2018). "The Simple View of Reading: Advancements and False
Impressions." Remedial and Special Education 39(5): 317-323.
Cecil, K. M., K. J. Brunst and T. Horowitz-Kraus (2021). "Greater reading gain
following intervention is associated with low magnetic resonance
spectroscopy derived concentrations in the anterior cingulate cortex in
children with dyslexia." Brain Res 1759: 147386.
Cecil, K. M., K. J. Brunst and T. Horowitz-Kraus (2021). "Greater reading gain
following intervention is associated with low magnetic resonance
spectroscopy derived concentrations in the anterior cingulate cortex in
children with dyslexia." Brain Research 1759: 147386.
Cha, J., D. DeDora, S. Nedic, J. Ide, T. Greenberg, G. Hajcak and L. R.
Mujica-Parodi (2016). "Clinically Anxious Individuals Show Disrupted
Feedback between Inferior Frontal Gyrus and Prefrontal-Limbic Control
Circuit." J Neurosci 36(17): 4708-4718.
Chen, L., D. Wassermann, D. A. Abrams, J. Kochalka, G. Gallardo-Diez and
V. Menon (2019). "The visual word form area (VWFA) is part of both language
and attention circuitry." Nat Commun 10(1): 5601.
Chen, Z., M. Liu, D. W. Gross and C. Beaulieu (2013). "Graph theoretical
analysis of developmental patterns of the white matter network." Front Hum
Neurosci 7: 716.
Ciric, R., D. H. Wolf, J. D. Power, D. R. Roalf, G. L. Baum, K. Ruparel, R. T.
Shinohara, M. A. Elliott, S. B. Eickhoff, C. Davatzikos, R. C. Gur, R. E. Gur, D.
S. Bassett and T. D. Satterthwaite (2017). "Benchmarking of participant-level
confound regression strategies for the control of motion artifact in studies of
functional connectivity." Neuroimage 154: 174-187.
Cohen, L., S. Dehaene, F. Vinckier, A. Jobert and A. Montavont (2008).
"Reading normal and degraded words: Contribution of the dorsal and ventral
visual pathways." NeuroImage 40(1): 353-366.
Conant, L. L., E. Liebenthal, A. Desai, M. S. Seidenberg and J. R. Binder
(2020). "Differential activation of the visual word form area during auditory
phoneme perception in youth with dyslexia." Neuropsychologia 146: 107543.
Corbetta, M. and G. L. Shulman (2002). "Control of goal-directed and
stimulus-driven attention in the brain." Nat Rev Neurosci 3(3): 201-215.
Costafreda, S. G., C. H. Fu, L. Lee, B. Everitt, M. J. Brammer and A. S. David
(2006). "A systematic review and quantitative appraisal of fMRI studies of
verbal fluency: role of the left inferior frontal gyrus." Hum Brain Mapp 27(10):
799-810.
Cragg, L. and C. Gilmore (2014). "Skills underlying mathematics: The role of
executive function in the development of mathematics proficiency." Trends in
Neuroscience and Education 3(2): 63-68.
Cutting, L. E., A. Clements-Stephens, K. R. Pugh, S. Burns, A. Cao, J. J.
Pekar, N. Davis and S. L. Rimrodt (2013). "Not all reading disabilities are
dyslexia: distinct neurobiology of specific comprehension deficits." Brain
Connect 3(2): 199-211.
Dehaene, S. (2009). Reading in the brain: The new science of how we read.
New York, NY, Penguin.
Dehaene, S. (2009). Reading in the Brain: The science and evolution of a
human invention. USA, Penguin Group.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
48
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Dehaene, S. and L. Cohen (2011). "The unique role of the visual word form
area in reading." Trends Cogn Sci 15(6): 254-262.
Delis, D. C., Kaplan, E., & Kramer, J. H. (2011). Delis-Kaplan Executive
Function System (D–KEFS).
Deschamps, I. and P. Tremblay (2014). "Sequencing at the syllabic and
supra-syllabic levels during speech perception: an fMRI study." Front Hum
Neurosci 8: 492.
Diamond, A. (2020). "Executive functions." Handb Clin Neurol 173: 225-240.
Dosenbach, N. U., D. A. Fair, F. M. Miezin, A. L. Cohen, K. K. Wenger, R. A.
Dosenbach, M. D. Fox, A. Z. Snyder, J. L. Vincent, M. E. Raichle, B. L.
Schlaggar and S. E. Petersen (2007). "Distinct brain networks for adaptive
and stable task control in humans." Proc Natl Acad Sci U S A 104(26): 11073-
11078.
Dunn, L. M. and D. M. Dunn (2007). Peabody Picture Vocabulary Test, Fourth
Edition (PPVT-4), Pearson.
Emch, M., C. C. von Bastian and K. Koch (2019). "Neural Correlates of Verbal
Working Memory: An fMRI Meta-Analysis." Frontiers in Human Neuroscience
13.
Farah, R., S. Ionta and T. Horowitz-Kraus (2021). "Neuro-Behavioral
Correlates of Executive Dysfunctions in Dyslexia Over Development From
Childhood to Adulthood." Front Psychol 12: 708863.
Fletcher, J. M., R. Savage and S. Vaughn (2021). "A Commentary on Bowers
(2020) and the Role of Phonics Instruction in Reading." Educ Psychol Rev
33(3): 1249-1274.
Franceschini, S., S. Gori, M. Ruffino, K. Pedrolli and A. Facoetti (2012). "A
causal link between visual spatial attention and reading acquisition." Curr Biol
22(9): 814-819.
Freedman, L., M. Zivan, R. Farah and T. Horowitz-Kraus (2020). "Greater
functional connectivity within the cingulo-opercular and ventral attention
networks is related to better fluent reading: A resting-state functional
connectivity study." Neuroimage Clin 26: 102214.
Friston, K., W. Penny, J. Ashburner, S. Kiebel and T. Nichols (2006).
Statistical Parametric Mapping: The Analysis of Functional Brain Images,
Academic Press.
Galuschka, K., R. Görgen, J. Kalmar, S. Haberstroh, X. Schmalz and G.
Schulte-Körne (2020). "Effectiveness of spelling interventions for learners with
dyslexia: A meta-analysis and systematic review." Educational Psychologist
55(1): 1-20.
Gao, Z., X. Guo, C. Liu, Y. Mo and J. Wang (2020). "Right inferior frontal
gyrus: An integrative hub in tonal bilinguals." Human Brain Mapping 41(8):
2152-2159.
Goldhammer, F., J. Naumann, A. Stelter, K. Tóth, H. Rölke and E. Klieme
(2014). "The time on task effect in reading and problem solving is moderated
by task difficulty and skill: Insights from a computer-based large-scale
assessment." Journal of Educational Psychology 106: 608-626.
Gratton, C., H. Sun and S. E. Petersen (2018). "Control networks and hubs."
Psychophysiology 55(3).
Hancock, R., Pugh, K. R., Hoeft, F. (2017). "Neural Noise Hypothesis of
Developmental Dyslexia." Trends in Cognitive Neuroscience Volume 21,
Issue 6, June 2017, Pages(6): 434-448.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
49
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hermosillo, R. (2022). Applying Precision Functional Mapping Techniques to
Quantify Age-Related Changes in Network Topography. Flux Congress, Paris,
France.
Hoover, W. A. and P. B. Gough (1990). "The simple view of reading." Reading
and Writing 2(2): 127-160.
Horowitz-Kraus, T. (2012). "Pinpointing the Deficit in Executive Functions in
Adolescents With Dyslexia Performing the Wisconsin Card Sorting Test: An
ERP Study." Journal of Learning DIsabilities.
Horowitz-Kraus, T. (2012). "Pinpointing the deficit in executive functions in
teenage dyslexic readers: an ERP study using the Wisconsin Card Sorting
Test." Journal of Learning Disabilities. Aug 18.
Horowitz-Kraus, T. (2013). Can we train the dyslexic readers to read like a
typical readers? EEG and fMRI study using the Reading Acceleration
Program. Brain, Mind and Fluency Conference. Haifa, Israel, Edmond J. Safra
Brain Research Center for the Study of Learning Disabilities.
Horowitz-Kraus, T. (2016). "Improvement of the Error-detection Mechanism in
Adults with Dyslexia Following Reading Acceleration Training." Dyslexia
22(2): 173-189.
Horowitz-Kraus, T., & Breznitz, Z. (2014). "Can reading rate acceleration
improve error monitoring and cognitive abilities underlying reading in
adolescents with reading difficulties and in typical readers?" Brain Res 1544:
1-14.
Horowitz-Kraus, T., & Holland, S.K. (2015). "Greater Functional Connectivity
between Reading and Error-Detection Regions Following Training with the
Reading Acceleration Program in Children with Reading Difficulties." Annals
of Dyslexia 10.1007/s11881-015-0096-9
Horowitz-Kraus, T. and Z. Breznitz (2014). "Can reading rate acceleration
improve error monitoring and cognitive abilities underlying reading in
adolescents with reading difficulties and in typical readers?" Brain research
1544: 1-14.
Horowitz-Kraus, T., N. Cicchino, M. Amiel, S. K. Holland and Z. Breznitz
(2014). "Reading improvement in English- and Hebrew-speaking children with
reading difficulties after reading acceleration training." Annals of Dyslexia
64(3): 183-201.
Horowitz-Kraus, T., M. DiFrancesco, B. Kay, Y. Wang and S. K. Holland
(2015). "Increased resting-state functional connectivity of visual- and
cognitive-control brain networks after training in children with reading
difficulties." Neuroimage Clin 8: 619-630.
Horowitz-Kraus, T., DiFrancesco, M., Kay, B., Wang, Y., & Holland, S.K.
(2015c). "Increased functional connectivity of specific brain networks after
reading training in dyslexic children." Clinical Neuroimage 8 619-630.
Horowitz-Kraus, T., A. Hershey, B. Kay and M. DiFrancesco (2019).
"Differential effect of reading training on functional connectivity in children with
reading difficulties with and without ADHD comorbidity." J Neurolinguistics 49:
93-108.
Horowitz-Kraus, T. and S. K. Holland (2015). "Greater functional connectivity
between reading and error-detection regions following training with the
reading acceleration program in children with reading difficulties." Annals of
dyslexia 65(1): 1-23.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Horowitz-Kraus, T., Rosch, K., Fotang, J., Mostofsky, S., Schlaggar, B. L.,
Pekar, J., DiFrancesco, M., Altaye, M., Farah, R. (2022). Accelerated, but not
still, text is associated with stronger functional connectivity of visual and
auditory networks with cognitive control networks in both dyslexic and typical
readers. Israeli Society for Neuroscience. Eilat, Israel, FENS.
Horowitz-Kraus, T., C. Toro-Serey and M. DiFrancesco (2015). "Increased
Resting-State Functional Connectivity in the Cingulo-Opercular Cognitive-
Control Network after Intervention in Children with Reading Difficulties." PLoS
One 10(7): e0133762.
Horowitz-Kraus, T., Toro-Serey, C., & DiFrancesco, M. (2015b). "Increased
Resting-state Functional Connectivity in the Cingulo-opercular Cognitive-
control Network after Intervention in Children with Reading Difficulties."
PLOSONE 10(7 ): e0133762.
Horowitz-Kraus, T., J. J. Vannest, D. S. Kadis, N. D. Cicchino, Y. Y. Wang
and S. K. Holland (2014). "Reading acceleration training changes brain
circuitry in children with reading difficulties." Brain and Behavior 4(6): 886-902.
Horowitz-Kraus, T., Vannest, J. J., Kadis, D., Cicchino, N., Wang, Y. Y.,
Holland, S. K. (2014). "Reading acceleration training changes brain circuitry in
children with reading difficulties." Brain Behav.
IBM, C. (2021). IBM SPSS Statistics for Windows, Version 28.0. Armonk, NY,
IBM Corp.
Khalil, R., A. A. Karim, A. Kondinska and B. Godde (2020). "Effects of
transcranial direct current stimulation of left and right inferior frontal gyrus on
creative divergent thinking are moderated by changes in inhibition control."
Brain Structure and Function 225(6): 1691-1704.
Kieffer, M. J. and J. A. Christodoulou (2020). "Automaticity and control: how
do executive functions and reding fluency interact in predicting reading
comprehension?" Reading Research Quarterly 55(1): 147-166.
Kilian, C., K. L. Bröckel, R. Overmeyer, R. Dieterich and T. Endrass (2020).
"Neural correlates of response inhibition and performance monitoring in binge
watching." International Journal of Psychophysiology 158: 1-8.
Klaus, J. and G. Hartwigsen (2019). "Dissociating semantic and phonological
contributions of the left inferior frontal gyrus to language production." Human
Brain Mapping 40(11): 3279-3287.
Li, Y., C. Seger, Q. Chen and L. Mo (2020). "Left Inferior Frontal Gyrus
Integrates Multisensory Information in Category Learning." Cereb Cortex
30(8): 4410-4423.
Liu, Y., J. Bengson, H. Huang, G. R. Mangun and M. Ding (2016). "Top-down
Modulation of Neural Activity in Anticipatory Visual Attention: Control
Mechanisms Revealed by Simultaneous EEG-fMRI." Cereb Cortex 26(2):
517-529.
López-Barroso, D., M. Thiebaut de Schotten, J. Morais, R. Kolinsky, L. W.
Braga, A. Guerreiro-Tauil, S. Dehaene and L. Cohen (2020). "Impact of
literacy on the functional connectivity of vision and language related
networks." NeuroImage 213: 116722.
Mai, J. and G. Paxinos (2012). The Human Nervous System. Amsterdam,
Netherlands, Elsevier Academic Press.
Manly, T., V. Anderson, I. Nimmo-Smith, A. Turner, P. Watson and I. H.
Robertson (2001). "The differential assessment of children's attention: the
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
51
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Test of Everyday Attention for Children (TEA-Ch), normative sample and
ADHD performance." J Child Psychol Psychiatry 42(8): 1065-1081.
Marek, S., B. Tervo-Clemmens, F. J. Calabro, D. F. Montez, B. P. Kay, A. S.
Hatoum, M. R. Donohue, W. Foran, R. L. Miller, T. J. Hendrickson, S. M.
Malone, S. Kandala, E. Feczko, O. Miranda-Dominguez, A. M. Graham, E. A.
Earl, A. J. Perrone, M. Cordova, O. Doyle, L. A. Moore, G. M. Conan, J.
Uriarte, K. Snider, B. J. Lynch, J. C. Wilgenbusch, T. Pengo, A. Tam, J. Chen,
D. J. Newbold, A. Zheng, N. A. Seider, A. N. Van, A. Metoki, R. J. Chauvin, T.
O. Laumann, D. J. Greene, S. E. Petersen, H. Garavan, W. K. Thompson, T.
E. Nichols, B. T. T. Yeo, D. M. Barch, B. Luna, D. A. Fair and N. U. F.
Dosenbach (2022). "Reproducible brain-wide association studies require
thousands of individuals." Nature 603(7902): 654-660.
Melby-Lervag, M., S. A. Lyster and C. Hulme (2012). "Phonological skills and
their role in learning to read: a meta-analytic review." Psychol Bull 138(2):
322-352.
Miyake, A., Friedman , N.P., Emerson. M.J., Witzki . A.H.,Howerter , A. &
Wager, T.D. (2000). "The Unity and Diversity of Executive Functions and
Their Contributions to Complex “Frontal Lobe” Tasks: A Latent Variable
Analysis. ." Cognitive Psychology 41: 49-100.
Miyake, A., Friedman, N. P. (2012). "The Nature and Organization of
Individual Differences in Executive Functions: Four General Conclusions."
Current Directions in Psychological Science 21(1): 8-14.
Morris, R. D., M. W. Lovett, M. Wolf, R. A. Sevcik, K. A. Steinbach, J. C.
Frijters and M. B. Shapiro (2012). "Multiple-component remediation for
developmental reading disabilities: IQ, socioeconomic status, and race as
factors in remedial outcome." J Learn Disabil 45(2): 99-127.
Nakae, T., R. Matsumoto, T. Kunieda, Y. Arakawa, K. Kobayashi, A.
Shimotake, Y. Yamao, T. Kikuchi, T. Aso, M. Matsuhashi, K. Yoshida, A.
Ikeda, R. Takahashi, M. A. Lambon Ralph and S. Miyamoto (2020).
"Connectivity Gradient in the Human Left Inferior Frontal Gyrus: Intraoperative
Cortico-Cortical Evoked Potential Study." Cerebral Cortex 30(8): 4633-4650.
Nguyen, T. Q., S. N. Del Tufo and L. E. Cutting (2020). "Readers Recruit
Executive Functions to Self-Correct Miscues during Oral Reading Fluency."
Scientific Studies of Reading 24(6): 462-483.
Norton, E. S., S. D. Beach and J. D. Gabrieli (2015). "Neurobiology of
dyslexia." Curr Opin Neurobiol 30: 73-78.
Norton, E. S., J. M. Black, L. M. Stanley, H. Tanaka, J. D. Gabrieli, C. Sawyer
and F. Hoeft (2014). "Functional neuroanatomical evidence for the double-
deficit hypothesis of developmental dyslexia." Neuropsychologia 61: 235-246.
Oldham, S. and A. Fornito (2019). "The development of brain network hubs."
Dev Cogn Neurosci 36: 100607.
Palejwala, A. H., N. B. Dadario, I. M. Young, K. O’Connor, R. G. Briggs, A. K.
Conner, D. L. O’Donoghue and M. E. Sughrue (2021). "Anatomy and White
Matter Connections of the Lingual Gyrus and Cuneus." World Neurosurgery
151: e426-e437.
Partchev, I., P. De Boeck and R. Steyer (2013). "How much power and speed
is measured in this test?" Assessment 20: 242-252.
Peters, J. L., L. De Losa, E. L. Bavin and S. G. Crewther (2019). "Efficacy of
dynamic visuo-attentional interventions for reading in dyslexic and
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
52
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
neurotypical children: A systematic review." Neurosci Biobehav Rev 100: 58-
76.
Peterson, R. L. and B. F. Pennington (2012). "Developmental dyslexia."
Lancet 379(9830): 1997-2007.
Power, J. D., A. L. Cohen, S. M. Nelson, G. S. Wig, K. A. Barnes, J. A.
Church, A. C. Vogel, T. O. Laumann, F. M. Miezin, B. L. Schlaggar and S. E.
Petersen (2011). "Functional network organization of the human brain."
Neuron 72(4): 665-678.
Pulvermüller, F. (2013). "How neurons make meaning: brain mechanisms for
embodied and abstract-symbolic semantics." Trends Cogn Sci 17(9): 458-
470.
Qian, Y., Y. Bi, X. Wang, Y.-W. Zhang and H.-Y. Bi (2016). "Visual dorsal
stream is associated with Chinese reading skills: A resting-state fMRI study."
Brain and Language 160: 42-49.
Rolls, E. T., W. Cheng, J. Du, D. Wei, J. Qiu, D. Dai, Q. Zhou, P. Xie and J.
Feng (2020). "Functional connectivity of the right inferior frontal gyrus and
orbitofrontal cortex in depression." Soc Cogn Affect Neurosci 15(1): 75-86.
Rosner, B. (2011). Fundamentals of Biostatistics. Boston, MA, Brooks/Cole.
Rosselli, M., A. Ardila and B. Bernal (2015). "[Angular gyrus connectivity
model for language: a functional neuroimaging meta-analysis]." Rev Neurol
60(11): 495-503.
Savage, R., G. Georgiou, R. Parrila and K. Maiorino (2018). "Preventative
Reading Interventions Teaching Direct Mapping of Graphemes in Texts and
Set-for-Variability Aid At-Risk Learners." Scientific Studies of Reading 22(3):
225-247.
Schrank, F., N. Mather and K. McGrew (2014). Woodcock-Johnson IV Tests
of Achievement. Rolling Meadows, IL, USA, Riverside.
Schulte-Körne, G. (2010). "The prevention, diagnosis, and treatment of
dyslexia." Dtsch Arztebl Int 107(41): 718-726; quiz 727.
Silverman, R. D., D. L. Speece, J. R. Harring and K. D. Ritchey (2013).
"Fluency Has a Role in the Simple View of Reading." Scientific Studies of
Reading 17(2): 108-133.
Siok, W. T., F. Jia, C. Y. Liu, C. A. Perfetti and L. H. Tan (2020). "A Lifespan
fMRI Study of Neurodevelopment Associated with Reading Chinese." Cereb
Cortex 30(7): 4140-4157.
Snowling, M. J. (2013). "Early identification and interventions for dyslexia: a
contemporary view." Journal of Research in Special Educational Needs 13(1):
7-14.
Snowling, M. J. and C. Hulme (2012). "Annual research review: the nature
and classification of reading disorders--a commentary on proposals for DSM-
5." J Child Psychol Psychiatry 53(5): 593-607.
Snowling, M. J., C. Hulme and K. Nation (2020). "Defining and understanding
dyslexia: past, present and future." Oxf Rev Educ 46(4): 501-513.
Spencer, M., M. C. Richmond and L. E. Cutting (2020). "Considering the Role
of Executive Function in Reading Comprehension: A Structural Equation
Modeling Approach." Sci Stud Read 24(3): 179-199.
Sporns, O. (2011). Networks of the brain. Cambridge, MA, US, MIT Press.
Sporns, O. (2012). Discovering the Human Connectome, The MIT Press.
Stein, J. (2019). "The current status of the magnocellular theory of
developmental dyslexia." Neuropsychologia 130: 66-77.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
53
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Stevens, W. D., D. J. Kravitz, C. S. Peng, M. H. Tessler and A. Martin (2017).
"Privileged Functional Connectivity between the Visual Word Form Area and
the Language System." J Neurosci 37(21): 5288-5297.
Strong, G. K., C. J. Torgerson, D. Torgerson and C. Hulme (2011). "A
systematic meta-analytic review of evidence for the effectiveness of the 'Fast
ForWord' language intervention program." J Child Psychol Psychiatry 52(3):
224-235.
Su, S. and M. Davison (2019). "Improving the Predictive Validity of Reading
Comprehension Using Response Times of Correct Item Responses." Applied
Measurement in Education 32: 166-182.
Swick, D., V. Ashley, Turken and U (2008). "Left inferior frontal gyrus is critical
for response inhibition." BMC Neuroscience 9(1): 102.
Szczepanski, S. and S. Kastner (2013). "Shifting Attentional Priorities: Control
of Spatial Attention through Hemispheric Competition." The Journal of
Neuroscience 33: 5411 - 5421.
Taran, N., R. Farah, M. DiFrancesco, M. Altaye, J. Vannest, S. Holland, K.
Rosch, B. L. Schlaggar and T. Horowitz-Kraus (2021). "The role of visual
attention in dyslexia: Behavioral and neurobiological evidence." Hum Brain
Mapp.
Taran, N., R. Farah, M. DiFrancesco, M. Altaye, J. Vannest, S. Holland, K.
Rosch, B. L. Schlaggar and T. Horowitz-Kraus (2022). "The role of visual
attention in dyslexia: Behavioral and neurobiological evidence." Human brain
mapping 43(5): 1720-1737.
Toffalini, E., D. Giofrè, M. Pastore, B. Carretti, F. Fraccadori and D. Szűcs
(2021). "Dyslexia treatment studies: A systematic review and suggestions on
testing treatment efficacy with small effects and small samples." Behav Res
Methods 53(5): 1954-1972.
Tomasello, R., M. Garagnani, T. Wennekers and F. Pulvermüller (2017).
"Brain connections of words, perceptions and actions: A neurobiological
model of spatio-temporal semantic activation in the human cortex."
Neuropsychologia 98: 111-129.
Tomiyama, H., K. Murayama, K. Nemoto, M. Tomita, S. Hasuzawa, T.
Mizobe, K. Kato, A. Ohno, S. Tsuruta, O. Togao, A. Hiwatashi and T. Nakao
(2022). "Increased functional connectivity between presupplementary motor
area and inferior frontal gyrus associated with the ability of motor response
inhibition in obsessive–compulsive disorder." Human Brain Mapping 43(3):
974-984.
Torgesen, J., R. K. Wagner and C. Rashotte (1999). "Test of word reading
efficiency (TOWRE) Pro-Ed." Austin, Tx.
Tran, S. M., K. M. McGregor, G. A. James, K. Gopinath, V. Krishnamurthy, L.
C. Krishnamurthy and B. Crosson (2018). "Task-residual functional
connectivity of language and attention networks." Brain Cogn 122: 52-58.
Vannest, J., A. Rajagopal, N. D. Cicchino, J. Franks-Henry, S. M. Simpson, G.
Lee, M. Altaye, C. Sroka and S. K. Holland (2014). "Factors determining
success of awake and asleep magnetic resonance imaging scans in
nonsedated children." Neuropediatrics 45(6): 370-377.
Varvara, P., C. Varuzza, A. C. Sorrentino, S. Vicari and D. Menghini (2014).
"Executive functions in developmental dyslexia." Front Hum Neurosci 8: 120.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
54
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Vidyasagar, T. R. (2019). "Visual attention and neural oscillations in reading
and dyslexia: Are they possible targets for remediation?" Neuropsychologia
130: 59-65.
Vogel, A. C., F. M. Miezin, S. E. Petersen and B. L. Schlaggar (2011). "The
Putative Visual Word Form Area Is Functionally Connected to the Dorsal
Attention Network." Cerebral Cortex 22(3): 537-549.
Vogel, A. C., S. E. Petersen and B. L. Schlaggar (2014). "The VWFA: it's not
just for words anymore." Front Hum Neurosci 8: 88.
Wagner, R. K., J. Torgesen, C. Rashotte and N. Pearson (2013). "The
Comprehensive Test of Phonological Processing, (CTOPP-2) Austin." Texas:
Pro-Ed.
Wagner, R. K., F. A. Zirps, A. A. Edwards, S. G. Wood, R. E. Joyner, B. J.
Becker, G. Liu and B. Beal (2020). "The Prevalence of Dyslexia: A New
Approach to Its Estimation." J Learn Disabil 53(5): 354-365.
Washburn, J. (2022). "Reviewing Evidence on the Relations Between Oral
Reading Fluency and Reading Comprehension for Adolescents." Journal of
Learning Disabilities 55(1): 22-42.
Wechsler, D. (2011). Wechsler Intelligence Scale for Children, Fourth Edition.
Encyclopedia of Child Behavior and Development. S. Goldstein and J. A.
Naglieri. Boston, MA, Springer US: 1553-1555.
Weiss, Y., T. Katzir and T. Bitan (2016). "When transparency is opaque:
Effects of diacritic marks and vowel letters on dyslexic Hebrew readers."
Cortex 83: 145-159.
Welsh, M. C., B. F. Pennington and D. B. Groisser (1991). "A normative-
developmental study of executive function: A window on prefrontal function in
children." Developmental Neuropsychology 7: 131-149.
Whitfield-Gabrieli, S. and A. Nieto-Castanon (2012). "Conn: a functional
connectivity toolbox for correlated and anticorrelated brain networks." Brain
Connect 2(3): 125-141.
Wiederholt, J. L. and B. R. Bryant (2012). Gray Oral Reading Tests. Fifth
Edition. (GORT-5). Austin, TX, Pro-Ed.
Wierenga, L. M., M. P. van den Heuvel, S. van Dijk, Y. Rijks, M. A. de Reus
and S. Durston (2016). "The development of brain network architecture." Hum
Brain Mapp 37(2): 717-729.
Williams, K. J., M. A. Walker, S. Vaughn and J. Wanzek (2017). "A Synthesis
of Reading and Spelling Interventions and Their Effects on Spelling Outcomes
for Students With Learning Disabilities." J Learn Disabil 50(3): 286-297.
Xu, Y., Q. Lin, Z. Han, Y. He and Y. Bi (2016). "Intrinsic functional network
architecture of human semantic processing: Modules and hubs." Neuroimage
132: 542-555.
Yarkoni, T., N. K. Speer, D. A. Balota, M. P. McAvoy and J. M. Zacks (2008).
"Pictures of a thousand words: investigating the neural mechanisms of
reading with extremely rapid event-related fMRI." Neuroimage 42(2): 973-987.
Zhao, J., J. Wang, C. Huang and P. Liang (2022). "Involvement of the dorsal
and ventral attention networks in visual attention span." Hum Brain Mapp
43(6): 1941-1954.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
55
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure
Click here to access/download;Figure;FIG1.tif
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure
Click here to access/download;Figure;FIG2.tiff
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure
Click here to access/download;Figure;FIG3.tif
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure
Click here to access/download;Figure;FIG4.tif
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure
Click here to access/download;Figure;FIG5.tiff
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure
Click here to access/download;Figure;FIG6.tiff
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure
Click here to access/download;Figure;FIG7.tif
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure
Click here to access/download;Figure;FIG8.tif
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Author Summary
EF-based reading training engages the cingulo-opercular and
dorsal attention networks
Nikolay Taran, Rola Farah, Carmel Gashri, Ester Gitman, Keri Rosch, Bradley L.
Schlaggar, and Tzipi Horowitz-Kraus
Author summary
The aim of the study was to determine the behavioral and neural effects of a
computerized executive functions (EFs)-based reading intervention. A total of 79
participants (8-12-year-olds, English-speaking) with and without dyslexia trained either
on an EFs-based reading training or a math training.
After the EFs-based reading intervention, children with dyslexia improved their scores in
reading rate and visual attention. Intervention-related increases in fMRI functional
connectivity were observed between the cingulo-opercular network and occipital
regions. Higher indices of connectivity were related to better speed of processing and
visual attention. The reading improvement involved neuroplastic connectivity changes in
brain areas related to EFs and primary visual processing. The importance of training the
cognitive abilities supporting reading in dyslexia (e.g., EFs and visual attention) is
highlighted.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
.
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
5
2
1
5
6
8
1
1
n
e
n
_
a
_
0
0
3
3
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3