INVESTIGACIÓN
Coactivation pattern analysis reveals altered
salience network dynamics in children with
autism spectrum disorder
Emily Marshall
1∗
, Jason S. Nomi
1∗
, Bryce Dirks1, Celia Romero1,
Lauren Kupis1, Catie Chang2,3,4, and Lucina Q. Uddin1,5
1Department of Psychology, University of Miami, Coral Gables, Florida, EE.UU
2Department of Electrical Engineering and Computer Science, Vanderbilt University, Nashville, TN, EE.UU
3Department of Biomedical Engineering, Vanderbilt University, Nashville, TN, EE.UU
4Vanderbilt University Institute of Imaging Science, Vanderbilt University, Nashville, TN, EE.UU
5Neuroscience Program, University of Miami Miller School of Medicine, Miami, Florida, EE.UU
These authors contributed equally.
∗
Palabras clave: Anterior insula, Dynamic coactivation patterns, Conectividad funcional, Lateral
fronto-parietal network, Medial fronto-parietal network, Midcingulo-insular network
ABSTRACTO
Brain connectivity studies of autism spectrum disorder (ASD) have historically relied on static
measures of functional connectivity. Recent work has focused on identifying transient
configurations of brain activity, yet several open questions remain regarding the nature of
specific brain network dynamics in ASD. We used a dynamic coactivation pattern (CAP)
approach to investigate the salience/midcingulo-insular (M-CIN) network, a locus of
dysfunction in ASD, in a large multisite resting-state fMRI dataset collected from 172 niños
(ages 6–13 years; norte = 75 ASD; norte = 138 masculino). Following brain parcellation by using
independent component analysis, dynamic CAP analyses were conducted and k-means
clustering was used to determine transient activation patterns of the M-CIN. The frequency of
occurrence of different dynamic CAP brain states was then compared between children with
ASD and typically developing (TD) niños. Dynamic brain configurations characterized by
coactivation of the M-CIN with central executive/lateral fronto-parietal and default
mode/medial fronto-parietal networks appeared less frequently in children with ASD
compared with TD children. This study highlights the utility of time-varying approaches for
studying altered M-CIN function in prevalent neurodevelopmental disorders. We speculate
that altered M-CIN dynamics in ASD may underlie the inflexible behaviors commonly
observed in children with the disorder.
RESUMEN DEL AUTOR
Autism spectrum disorder (ASD) is a neurodevelopmental disorder associated with
altered patterns of functional brain connectivity. Little is currently known about how
moment-to-moment brain dynamics differ in children with ASD and typically developing
(TD) niños. Altered functional integrity of the midcingulo-insular network (M-CIN) tiene
been implicated in the neurobiology of ASD. Here we use a novel coactivation analysis
approach applied to a large sample of resting-state fMRI data collected from children with
ASD and TD children to demonstrate altered patterns of M-CIN dynamics in children with
the disorder. We speculate that these atypical patterns of brain dynamics may underlie
behavioral inflexibility in ASD.
un acceso abierto
diario
Citación: marshall, MI., Nomi, j. S.,
Dirks, B., Romero, C., Kupis, l., Chang,
C., & Uddin, l. q. (2020). Coactivation
pattern analysis reveals altered
salience network dynamics in children
with autism spectrum disorder.
Neurociencia en red, 4(4), 1219–1234.
https://doi.org/10.1162/netn_a_00163
DOI:
https://doi.org/10.1162/netn_a_00163
Supporting Information:
https://doi.org/10.1162/netn_a_00163
Recibió: 27 Abril 2020
Aceptado: 29 Julio 2020
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Lucina Q. Uddin
l.uddin@miami.edu
Editor de manejo:
Caterina Gratton
Derechos de autor: © 2020
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
norte
mi
norte
_
a
_
0
0
1
6
3
pag
d
.
t
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Salience network dynamics in autism
INTRODUCCIÓN
Autism spectrum disorders (ASDs) are neurodevelopmental disorders with central features of
atypical social communication and restricted and repetitive behavioral patterns (Americano
Psychiatric Association, 2013). These disorders have a high prevalence among school-aged
children and adolescents in the United States, and have recently been estimated to affect
alrededor 1 en 40 individuals between the ages of 3 y 17 (Kogan et al., 2018). Because of
the broad range and individual variability of the associated symptoms and etiologies of these
disorders, they are difficult to understand and to treat. Sin embargo, neuroimaging research in
the past decade has identified brain network functional connectivity (FC) as a metric with
which to potentially develop diagnostic indicators for ASD (Uddin et al., 2017). These stud-
ies have leveraged advances in machine learning to provide initial evidence that FC of the
salience/midcingulo-insular (M-CIN) network (Uddin, yo, & Spreng, 2019), comprising the
bilateral anterior insula and anterior cingulate cortices (Uddin, 2015), can discriminate ASD
from typical development (Anderson et al., 2011; Uddin et al., 2013).
Previous FC and activation studies of ASD have indicated that atypical functioning of the
M-CIN, default mode/medial fronto-parietal (M-FPN), and central executive/lateral fronto-
parietal (L-FPN) is associated with the disorder (Abbott et al., 2016; Padmanabhan et al., 2017;
Di Martino et al., 2009; Green et al., 2016). Because these brain networks support social
and emotional behavior and executive functions, it is possible that altered relationships be-
tween them may underlie the social communication deficits and inflexible behaviors associ-
ated with ASD. Within the broader M-CIN, the anterior insula is a key node that orchestrates
switching between the M-FPN and L-FPN (menón & Uddin, 2010) and is posited to be a spe-
cific locus of dysfunction in ASD (Uddin & menón, 2009; Nomi, Molnar-Szakacs, & Uddin,
2019). In studies of neurotypical individuals, the anterior insula has been shown to exhibit
patterns of dynamic FC that link it transiently with the M-FPN (Nomi et al., 2016; Chang &
guantero, 2010). Connectivity studies focusing on the anterior insula and the broader M-CIN in
ASD have produced mixed findings, with some indicating that this network may be hypercon-
nected in children with the disorder (Uddin et al., 2013) while others report hypoconnectivity
of this network (Abbott et al., 2016; Ebisch et al., 2011).
Traditional FC approaches operate under the assumption that the brain’s functional archi-
tecture remains relatively static throughout an entire fMRI scan. Although these static FC meth-
ods have provided informative estimates of the functional architecture of the brain, a growing
body of evidence suggests that time-varying analysis of functional networks may reveal addi-
tional, dynamic aspects of brain function that have been previously overlooked (Calhoun et al.,
2014; Hutchison et al., 2013). Because of this, dynamic or time-varying approaches have re-
cently become an area of interest for characterizing brain function and dysfunction (Uddin &
Karlsgodt, 2018; Lurie et al., 2020). In contrast to the single, average FC estimate produced by
static approaches, dynamic analyses can identify multiple transient brain “states” or coactiva-
tion patterns (CAPs) that recur throughout an fMRI scan (Liu et al., 2018; Chen et al., 2015).
The current study uses a dynamic CAP approach based on previous research demonstrat-
ing that whole-brain patterns, captured at the peaks of a brain region’s signal, can resolve a
traditional region of interest (ROI)–based FC map into multiple transient patterns occurring at
different points in time. Liu and Duyn first showed that applying k-means clustering to spatial
patterns derived from averaging across all brain voxels related to the highest ∼15% of activa-
tion time frames from the BOLD signal of a posterior cingulate cortex (PCC) ROI permits the
identification of multiple different coactivation patterns, breaking down traditional static FC
findings into multiple contributing patterns (Liu & duyn, 2013). Chen and colleagues showed
Midcingulo-insular network:
Core cortical brain regions within the
midcingulo-insular network (M-CIN)
include bilateral anterior insula and
anterior midcingulate cortex. Este
network encapsulates the previously
characterized “salience,” “ventral
atención,” and “cingulo- opercular”
redes.
Medial fronto-parietal network:
Core cortical brain regions within
the medial fronto-parietal network
(M-FPN) include medial prefrontal
and posterior cingulate cortex. Este
network is also referred to as the
“default mode” network.
Lateral fronto-parietal network:
Core cortical brain regions within
the lateral fronto-parietal network
(L-FPN) include lateral prefrontal and
anterior inferior parietal cortex. Este
network is also referred to as the
“control” or “central executive”
network.
Coactivation pattern analysis:
Coactivation pattern (CAP) análisis
tracks brain state alterations at the
individual time frame level, and is
one approach for quantifying brain
dinámica.
Neurociencia en red
1220
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
norte
mi
norte
_
a
_
0
0
1
6
3
pag
d
t
/
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Salience network dynamics in autism
that a dynamic CAP analysis of a PCC node that employs k-means clustering across the highest
activation time frames (instead of averaging across such time frames) produces dynamic CAPs
that are less variable (es decir., more stable) than dynamic brain states identified using a sliding win-
dow correlation analysis (Chen et al., 2015). Además, unlike sliding window correlations,
dynamic CAP analyses do not require the arbitrary selection of a window size over which to
average FC measures (Allen et al., 2014). De este modo, dynamic CAP analyses offer some advantages
over sliding window dynamic FC approaches.
Compared with static FC methods, dynamic time-varying approaches may exhibit superior
accuracy in characterizing the behavior of typically developing (TD) individuals, así como
in discriminating clinical from nonclinical populations including autism, schizophrenia, de-
presion, and bipolar disorder (Chen et al., 2017; Rashid et al., 2018; Damaraju et al., 2014;
Demirtas et al., 2016; Kaiser et al., 2019). Recent studies have used dynamic FC approaches to
investigate ASD, revealing abnormal patterns of whole-brain network configurations (Uddin,
2020b). De Lacy et al. (2017) and Watanabe and Rees (2017) both reported a reduction in the
number of transitions between brain state configurations in ASD, suggesting decreased func-
tional flexibility or overly stable dynamic properties of the autistic brain. In both global and
local brain networks, ASD has also been associated with reductions in other measures of dy-
namism, including functional fluidity and dynamic range (Fu et al., 2019). In a large sample of
niños, Rashid et al. (2018) found that those with autistic traits spent more time in a “globally
disconnected” state. Similarmente, Chen et al. (2017) found that individuals with ASD had gener-
ally weaker dynamic functional connections within whole-brain states. Individuals with ASD
have also been shown to have longer mean dwell times in these whole-brain configurations
with weaker connectivity (Yao et al., 2016). These studies provide evidence for consistent ab-
normalities in dynamic brain configurations in ASD, but have focused mostly on whole-brain
network organization. Además, no previous study has used a CAP analysis to probe brain
dynamics in ASD. Further investigation is needed to determine whether the findings reported
to date are related to specific brain networks implicated in ASD pathology such as the M-CIN
(Uddin & menón, 2009).
En el presente estudio, we applied independent component analysis (ICA) to resting-state fMRI
data collected from three cohorts of age- and IQ-matched children with ASD and TD children
and conducted a dynamic CAP analysis focusing on the M-CIN. The goals of the study were
a (a) identify patterns of dynamic CAPs related to the M-CIN, y (b) compare these dynamic
M-CIN coactivation patterns between TD and ASD groups. We hypothesized that children
with ASD would show altered M-CIN dynamics compared with TD children.
METHODS AND MATERIALS
Participantes
Participant data were obtained from the Autism Brain Imaging Data Exchange (ABIDE I and II)
public databases (Di Martino et al., 2014, 2017), combined with data collected by the Brain
Connectivity and Cognition Laboratory at the University of Miami (UM). Institutional review
board approval was provided by UM and each data contributor in the ABIDE databases. El
sample selected from the databases met the following inclusion criteria: (a) subjects were be-
tween ages 6 y 13; (b) subjects had more than 160 volumes (5 minutes and 20 artículos de segunda clase)
of resting-state fMRI data acquisition; (C) subjects had fMRI data collected using a repetition
tiempo (TR) = 2 artículos de segunda clase; y (d) subjects had their eyes closed during the resting-state scan
(Nair et al., 2018). Un total de 70 children scanned at UM met these criteria. Data for an addi-
tional 136 subjects were downloaded from the ABIDE databases. We selected the following
Neurociencia en red
1221
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
norte
mi
norte
_
a
_
0
0
1
6
3
pag
d
t
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Salience network dynamics in autism
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
norte
mi
norte
_
a
_
0
0
1
6
3
pag
d
.
t
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. Participant selection process. Neuroimaging and phenotypic data from three sites
(University of Miami, Erasmus University Medical Center Rotterdam, and Stanford University) eran
incluido.
sites that met these criteria: Universidad Stanford (ABIDE I and II) and Erasmus University Medi-
cal Center Rotterdam (ABIDE II). After quality control and removal of subjects with high levels
of head motion (mean framewise displacement (FD) > 0.5 mm and/or more than 35 marcos
with FD > 0.5 mm), the final sample consisted of 172 subjects (138 machos) (Cifra 1). El
ASD (norte = 75) and TD (norte = 97) groups did not differ in terms of age (pag = 0.285), full-scale
IQ (pag = 0.127), or mean head motion (pag = 0.917). For participants from UM and Stanford
Universidad, ASD diagnosis was confirmed by a licensed psychologist using the Autism Diag-
nostic Observation Schedule (ADOS-2) (Lord et al., 2012). For the Erasmus Medical Center
Participantes, an ASD diagnosis was confirmed from central medical records of the children’s
general practitioners in the Netherlands. Additional demographic information for participants
can be found in Table 1.
MRI Data Acquisition
At all three data collection sites, participants underwent a mock scanning session to familiarize
them with the MRI session procedure and offer them the opportunity to opt out before under-
going the MRI scan. For the resting-state fMRI scan, participants were instructed to keep their
Neurociencia en red
1222
Salience network dynamics in autism
Number of participants
Age (años)
Sex
Handedness
Full IQ
Verbal IQ
Performance IQ
Mesa 1. Participant demographics
TD
97
ASD
75
9.79 ± 1.74 (6.3−13.2)
74 machos, 23 hembras
10.07 ± 1.70 (6.5−13.0)
64 machos, 11 hembras
88 bien, 4 izquierda, 5 ambi
112.9 ± 13.9 (79−151)
113.1 ± 15.7 (67−153)
107.5 ± 16.0 (71−152)
0.157 ± 0.088 (0.046−0.453)
66 bien, 6 izquierda, 3 ambi
108.8 ± 17.1 (67−141)
108.3 ± 18.4 (72−149)
106.3 ± 15.5 (66−133)
0.155 ± 0.071 (0.055−0.430)
P value
−
0.285
−
−
0.127
0.100
0.632
Mean head motion (mm)
Nota. Mean ± standard deviation (minimum-maximum). IQ was measured based on Wechsler abbrevi-
ated scale of intelligence (WASI, WASI-II) or Snijders–Oomen Nonverbal Intelligence Test (SON-R). TD,
typically developing; ASD, autism spectrum disorder; IQ, intelligence quotient.
0.917
eyes closed while remaining awake. The scanner information and data acquisition parameters
for each site are detailed in Supporting Information Table S1.
Preprocessing of Resting-State fMRI Data
As the three sites utilized different scan lengths, each scan was trimmed to 155 timepoints,
which was the length of the data collected at the Erasmus site. We removed the first five
volumes from the beginning of the scans to remove any scanner initiation artifacts, and re-
moved additional volumes from the end of the scans, as needed, to make each scan 5 minutos
y 10 seconds in duration. The scans were then subjected to quality control procedures as
follows. All neuroimaging data underwent a visual quality control procedure conducted by
trained research assistants to identify scanner- or motion-induced artifacts. Artifacts included,
but were not limited to, signal loss, head coverage, motion slicing, ringing, blurring, ghosting,
and wrapping. Data were rated as either pass, qualified pass, or fail. All images included
in the analysis received either a pass or a qualified pass rating. The resting-state scans were
then preprocessed using DPABI version 3.1 (http://rfmri.org/dpabi) (Yan et al., 2016) and SPM
versión 12 (https://www.fil.ion.ucl.ac.uk/spm/software/spm12/). Preprocessing steps included
realignment, normalization to the 3-mm MNI template, y alisado (6 mm at full width at
half maximum).
We did not apply global signal regression (GSR) in the current analysis following previous
CAP studies (Chen et al., 2015; Kaiser et al., 2019). It is worth noting that in CAP analysis,
functional networks are defined on the bases of instantaneous regional synchrony at each
volume of data, and network definition does not rely on FC and thus may be less sensi-
tive to the motion concerns inherent to FC analysis (Liu et al., 2018). The global signal has
been demonstrated to contain both nuisance signals and neural signals (Murphy & Fox, 2017;
Liu, Nalci, & Falahpour, 2017). Although removal of the global signal as a preprocessing step
significantly mitigates artifacts from a variety of sources (Power et al., 2017; Ciric et al., 2018),
it also results in removal of neuronal signal to a degree that is unknown in any given dataset
(Uddin, 2017). Evidence from electrophysiological recordings in macaques (Scholvinck et al.,
2010) and magnetic resonance spectroscopy studies in rodents (Hyder & Rothman, 2010)
clearly demonstrates that the global signal also includes neural signals. The global signal
has been further shown to have a direct neuronal source in studies of macaques (Turchi et al.,
En el estudio actual, we were concerned that GSR might
2018) and rats (Ma et al., 2020).
Global signal regression:
Global signal regression (GSR) refers
to the removal of the average value of
all whole-brain or gray matter signals
via regression as a resting state fMRI
data denoising strategy.
Neurociencia en red
1223
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
norte
mi
norte
_
a
_
0
0
1
6
3
pag
d
/
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Salience network dynamics in autism
differentially affect the ASD and TD groups under investigation. As previously noted in sim-
ulation studies (Saad et al., 2012) and empirical studies of ASD, GSR can lead to a reversal
in the direction of group correlation differences relative to other preprocessing approaches
(Gotts et al., 2013). It is for these multiple reasons that we did not apply GSR in the current
análisis, though we acknowledge that the costs and benefits associated with doing so depend
on the research question and thus may differ from study to study (Uddin, 2020a).
Group Independent Component Analysis
The resting-state fMRI data from all 172 participants were subjected to a high model order
ICA by using the Group ICA of fMRI Toolbox (GIFT) (https://trendscenter.org/software/gift/).
We used a model order of 100 independent components (ICs) as in previous similar work
(Allen et al., 2014; Nomi et al., 2016), as individual brain networks are not always effectively
separated when using lower model orders such as 25 o 50 (Ray et al., 2013), pero puede ser
too finely parcellated at higher model orders above 100 (Kiviniemi et al., 2009). The info-
max algorithm was utilized for the ICA (http://mialab.mrn.org/software/gift) (Calhoun et al.,
2001). To ensure the stability of this estimation, the ICA algorithm was repeated 20 times us-
ing ICASSO (http://www.cis.hut.fi/projects/ica/icasso). Todo 100 ICs were visually inspected and
classified as noise or non-noise by two of the authors (JN & EM). The ICA components related
to movement, white matter, or cerebrospinal fluid were removed from further analysis. El
remaining 69 components were grouped into 12 functional domains to facilitate CAP interpre-
tation: salience/M-CIN, default mode/M-FPN, central executive/L-FPN, sensorimotor, frontal,
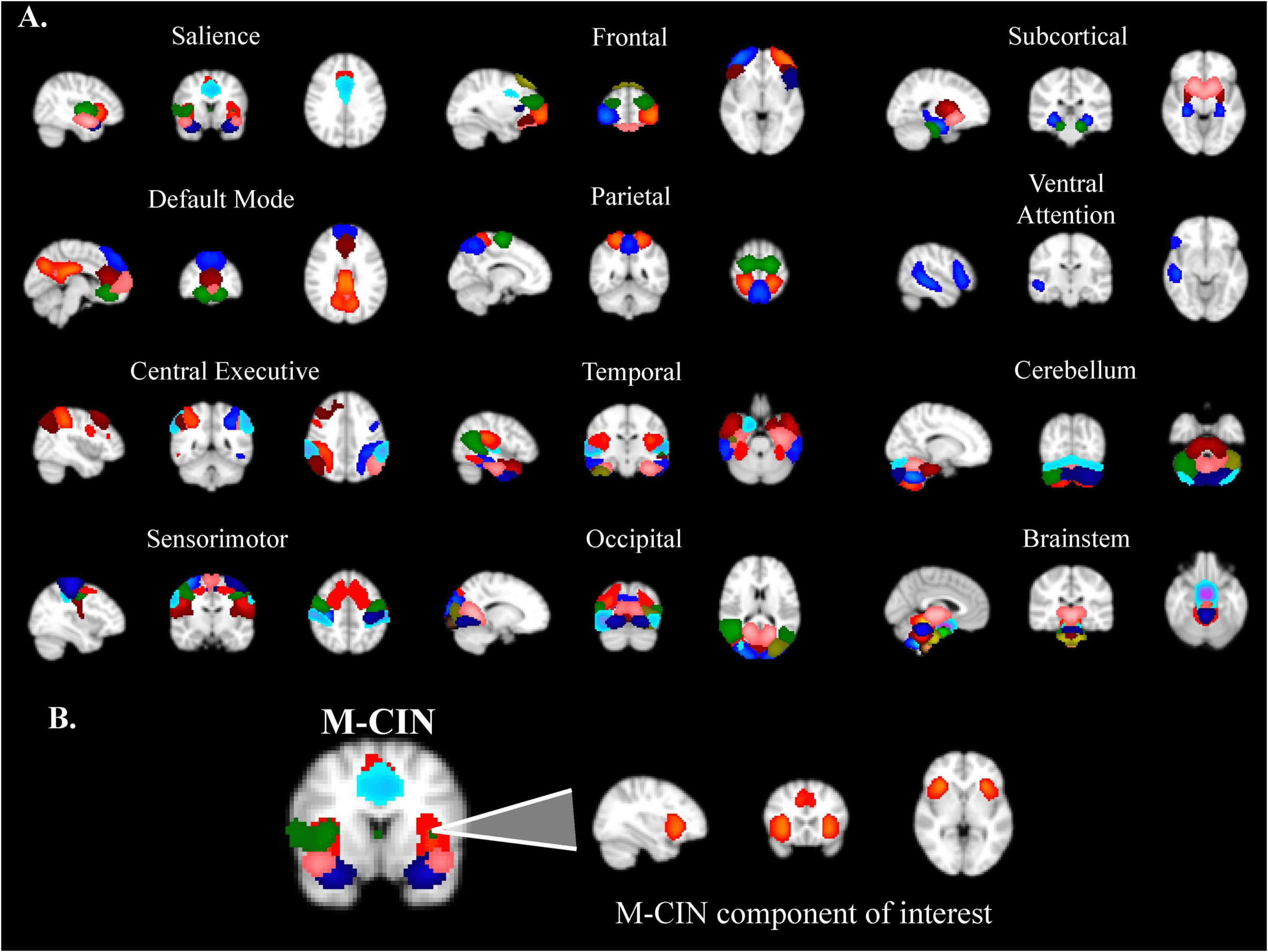
parietal, temporal, occipital, subcortical, atención, cerebellum, and brainstem (Cifra 2).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
norte
mi
norte
_
a
_
0
0
1
6
3
pag
d
.
/
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 2.
(A) Organization of ICA components into 12 functional networks. (B) Sagittal, coronal,
and axial views of the salience/midcingulo-insular (M-CIN) component of interest in the analyses.
Neurociencia en red
1224
Salience network dynamics in autism
The network of interest in this study was the salience/M-CIN, with key nodes in the bi-
lateral anterior insula and anterior cingulate cortices (Uddin, 2015), chosen because of its
previous designation as a major locus of dysfunction in ASD (Uddin & menón, 2009; Nomi,
Molnar-Szakacs, & Uddin, 2019).
Post-ICA Processing of fMRI Data
The time series were analyzed as a 155 (volumes) × 100 (ICs) matrix for each of the 172
Participantes, and were postprocessed using Matlab code from the GIFT toolbox. Postprocessing
included linear detrending, despiking using the AFNI 3D despike command, and bandpass
filtering (0.01–0.1 Hz) Allen et al. (2014).
Statistical Analyses
A k-means clustering algorithm was applied to a concatenated matrix of all subjects’ time-
points of non-noise components to extract dynamic CAPs corresponding to different resting-
redes estatales (Liu et al., 2018). For the anterior insula/anterior cingulate component we
identified as the salience/M-CIN for each participant, timepoints with signal intensity among
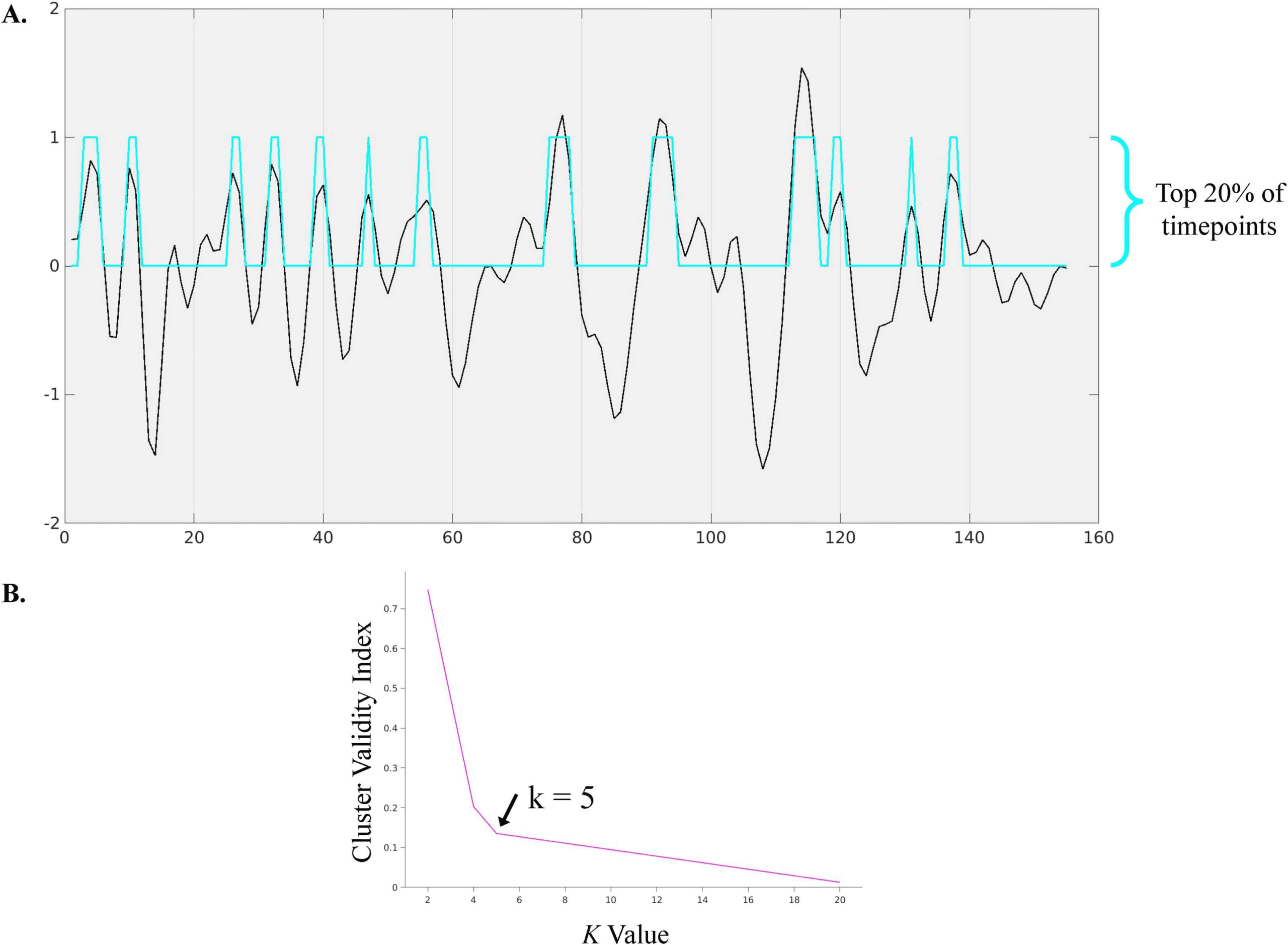
the top 20% (31 timepoints) y 30% (47 timepoints) of activation strength were extracted for
análisis (Chen et al., 2015). This resulted in thresholding at the subject level. The optimal
number of clusters was determined to be k = 5 by running k-means clustering with k-values 2
a través de 20 on a concatenated data matrix of all non-noise components for the selected time-
points across all participants. This optimal k was identified using the cluster validity index,
defined as the elbow point (Cifra 3) of a least-squares fit line plotted across the cluster validity
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
norte
mi
norte
_
a
_
0
0
1
6
3
pag
d
t
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 3.
(A) Sample time course depicting extraction of top 20% of timepoints. Blue represents
arriba 20% activation timepoints for salience/M-CIN component. (B) Elbow criteria graph to identify
optimal k as 5.
Neurociencia en red
1225
Salience network dynamics in autism
índice (the ratio between the within-cluster distance and the between-cluster distance) across
all values of k (Allen et al., 2014). Following k-means clustering, the frequency of occur-
rence of each brain configuration was calculated for each participant and compared between
grupos.
Participants were divided into TD and ASD groups. For the top 20% and top 30% analiza,
the mean frequencies of occurrence of CAPs were determined for the two groups. Frecuencia
was computed as the proportion of occurrence for a specific brain state out of all possible
brain states. Multiple linear regression models were used to compare the frequency of dynamic
CAP occurrence between groups while controlling for data collection site (UM, Erasmus, o
stanford), mean head motion, edad, and handedness.
Additional follow-up analyses were run to further explore patterns of activity when each
group was clustered separately, to examine the relationship between head motion and CAPs,
and to compare the current results against surrogate phase-randomized data. To determine
if each group could be characterized by distinct CAPs rather than differences in frequency of
the same CAPs, the ASD and TD groups were clustered separately using the optimal value
of k as determined for all data. To determine if there was a relationship between specific
CAPs, head motion and frequency of occurrence, average FD was computed for all TRs as-
signed to each CAP. Finalmente, to ensure that the identified CAPs were not driven by random
differences in the data rather than brain activity, k-means clustering was conducted on phase-
randomized surrogate data according to the procedure in (Lancaster et al., 2018). Los datos
were phase randomized producing surrogate data with the same mean, variance, and autocor-
relation of the original time series. K-means clustering was then conducted using the optimal
k value previously identified for the top 20% and top 30% of time points as in the previous
analiza.
RESULTADOS
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
norte
mi
norte
_
a
_
0
0
1
6
3
pag
d
/
.
t
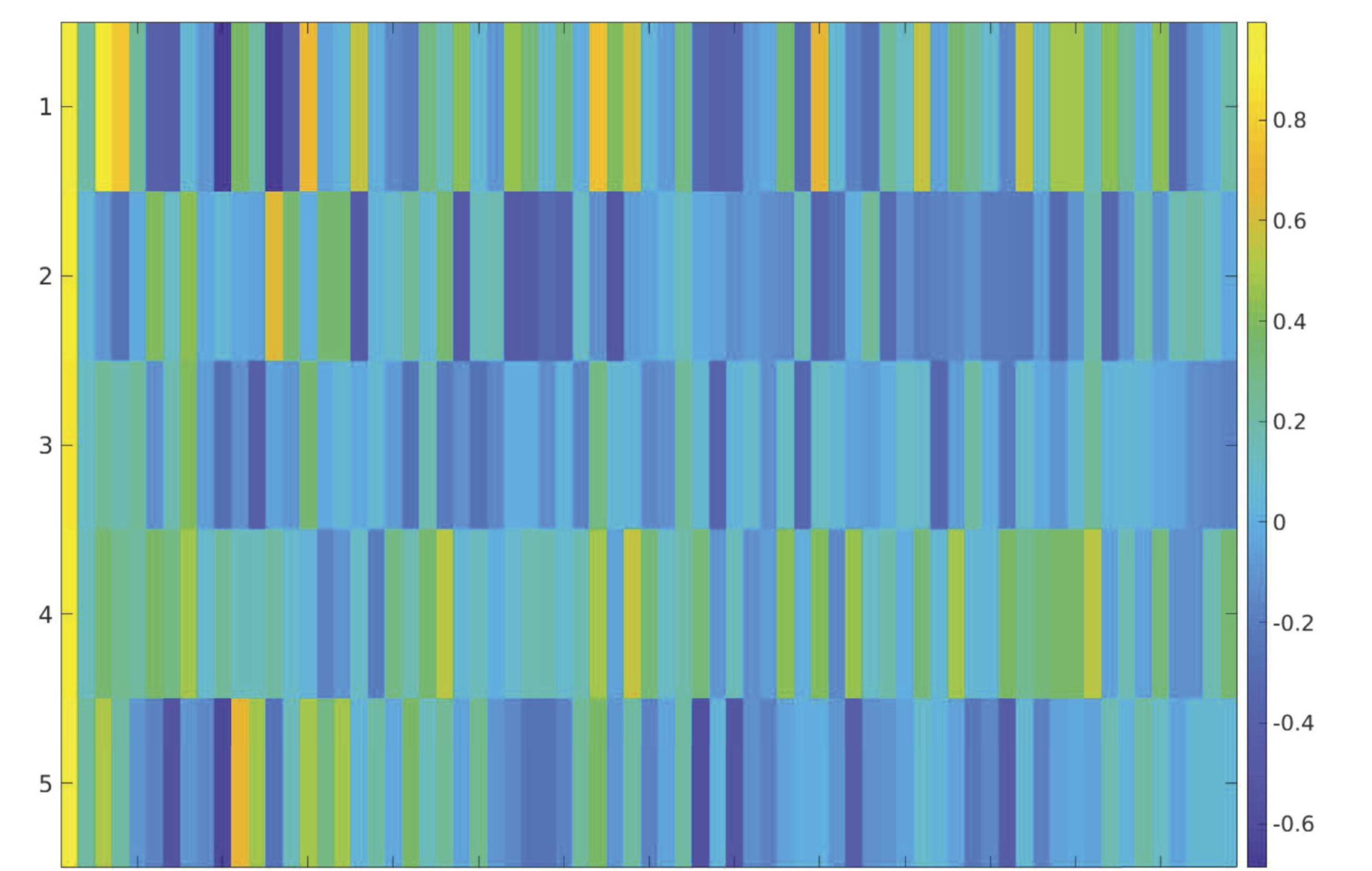
For the optimal k of five clusters, the activation patterns for the top 20% of time points are
como se muestra en la figura 4. For each of the five clusters, el 10 components with the highest coacti-
vation with the M-CIN component of interest were identified and are listed according to their
brain network (Cifra 5). The most notable brain configuration patterns were States 1 y 2.
Estado 1 was characterized by strong coactivation with other components of the M-CIN. En
both the top 20% and top 30% analiza, children with ASD entered this brain state more fre-
quently than their TD counterparts, although this difference was not statistically significant (pag =
0.1698, pag = 0.1856) (Cifra 6). Estado 2 was characterized by coactivation with the posterior
cingulate cortex (PCC) and medial prefrontal cortex (mPFC) of the M-FPN, as well as bilateral
regions of the L-FPN. This pattern of coactivation was significantly more frequently observed in
the TD children compared with ASD in both analyses (pag = 0.0198, k = 0.0232) (Cifra 6). Sig-
nificant group differences were not observed for frequency of occurrence of states 3–5. Resultados
for the top 30% of time points are presented in Supporting Information Figures S1 and S2.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
To ensure that our k-means clustering algorithm produced similar clusters in the ASD and TD
Participantes, the above steps were run on the two groups, separately. There was no noticeable
difference between the CAPs when the groups were clustered separately. The results of these
analyses are detailed in Supporting Information Figures S3 and S4, and Supporting Information
Table S2.
Neurociencia en red
1226
Salience network dynamics in autism
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
norte
mi
norte
_
a
_
0
0
1
6
3
pag
d
.
t
/
Cifra 4. Activation of components within each brain network for Top 20% análisis. COI, component of interest; SN, salience network;
DMN, default mode network; cen, central executive network; Front., frontal; Par., parietal; SM, sensorimotor; Sub., subcortical; Temp.,
temporal; Occ., occipital; Att., atención; CB, cerebellum.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
There was no identifiable systematic relationship between head motion, the frequency of
occurrence, or amount of TRs in each CAP (Supporting Information Figure S5). The most
frequently occurring CAP (p.ej., the most TRs) did not have the highest average motion (CAP 4).
The CAP with the lowest frequency of occurrence (CAP 1) did not have the average lowest
movimiento (CAP 2). The distribution of TR FD for each CAP also did not appear to drive the
clustering results (p.ej., some CAPs with all TRs being high motion while other TRs were all
low motion) while the average FD for each CAP was low (average FD < 0.2 mm).
Finally, phase-randomized surrogate data produced no noticeable CAPs of brain activity
(Supporting Information Figure S6). The CAPs from the primary analysis show strong patterns
of activation related to distinct brain networks. The CAPs from the phase-randomized surrogate
data show weak activation across network nodes that does not appear to be related to distinct
brain networks.
Network Neuroscience
1227
Salience network dynamics in autism
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
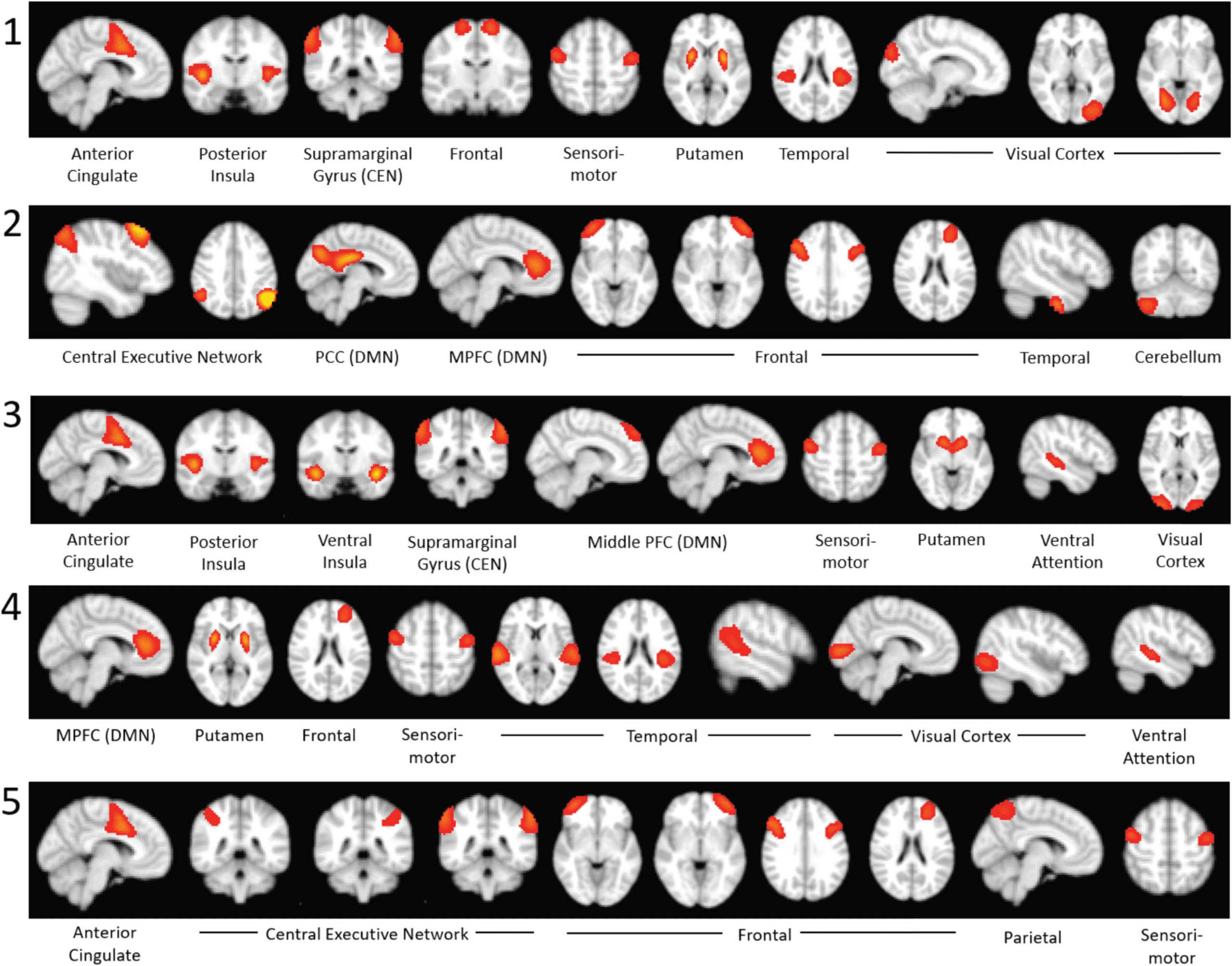
Figure 5. Top 10 most strongly activated components of each brain state in the top 20% analysis.
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
n
e
n
_
a
_
0
0
1
6
3
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Observed frequency of occurrence of coactivation patterns 1–5 for typically developing
(TD) and autism spectrum disorder (ASD) groups in the top 20% and top 30% analyses; *p < .05
in t test; **p < .05 in t test and multiple regression analyses.
Network Neuroscience
1228
Salience network dynamics in autism
DISCUSSION
ASDs are widely thought to be associated with atypical patterns of functional brain connectivity
(Uddin, Supekar, & Menon, 2013; Vissers, Cohen, & Geurts, 2012; Müller et al., 2011) within
and between large-scale brain networks important for high-level cognitive and emotional
processes (Padmanabhan et al., 2017; Nomi, Molnar-Szakacs, & Uddin, 2019). Analysis of
brain dynamics is beginning to reveal insights into neurodevelopmental disorders affecting
brain connectivity (Uddin & Karlsgodt, 2018; White & Calhoun, 2019). This emerging lit-
erature provides initial evidence for alterations in ASD related to the number of transitions
between brain states (Uddin, 2020a). Most of these studies have focused on characterizing
whole-brain dynamic patterns and utilized sliding window dynamic FC approaches that have
known potential pitfalls such as arbitrary window sizes (Preti, Bolton, & Van De Ville, 2017;
Lurie et al., 2020). Here we focus specifically on dynamics of a brain network known as the
salience network, or M-CIN, with key nodes in the bilateral anterior insula and anterior cin-
gulate cortices (Uddin, 2015), thought to be a locus of dysfunction in ASD (Uddin & Menon,
2009; Nomi, Molnar-Szakacs, & Uddin, 2019). We use dynamic coactivation pattern (CAP)
analysis, which relies on fewer assumptions than the sliding window approach, and permits
the examination of brain state alterations closer to the temporal resolution of individual time
frames (Chen et al., 2015). Using a combined ICA and CAP analytic approach, we investi-
gated the dynamic nature of M-CIN organization in children with ASD. By utilizing only time
points with the highest overall activation in the M-CIN in our analyses, we were able to focus
on this key network and note differences in the frequency of its coactivation with other major
large-scale networks of the brain.
In three of the five dynamic states or CAPs identified across TD and ASD participants, the
component of interest was consistently coactivated with other components of the M-CIN, in-
cluding the anterior cingulate and the posterior and ventral regions of the insular cortex. This
coactivation was particularly strong in State 1 and was also observed to some extent in States 3
and 4, which showed similar coactivation of the M-CIN components. Previous static FC work
has demonstrated altered M-CIN network properties in children and adolescents with ASD
(Ebisch et al., 2011; Abbott et al., 2016), and FC of the M-CIN can be used to discriminate
ASD from typical development (Uddin et al., 2013; Anderson et al., 2011).
The relative frequency of occurrence of State 2 differed significantly between the TD and
ASD groups.
In this state, the component of interest was not strongly coactivated with any
other components of the M-CIN, and its coactivation with components of other networks
was relatively weak, except a single component in the central executive/lateral fronto-parietal
network (L-FPN) and two components in the default mode/medial fronto-parietal network
(M-FPN). Children with ASD exhibited this CAP significantly less frequently than TD children.
The lower frequency of occurrence of this state for children with ASD indicates less coactiva-
tion of the M-CIN with components of the M-FPN and L-FPN, especially the medial prefrontal
cortex (mPFC) and PCC. Previous work has demonstrated that effective connectivity among
nodes of these three networks can be used to discriminate task-evoked and resting states in TD
children to a greater extent than in children with ASD. The same study found that this brain
state discriminability was related to symptom severity in the domain of restricted and repeti-
tive behaviors in children with ASD (Uddin et al., 2015). The reduced communication among
these three networks observed in our ASD samples could be related to symptoms of cognitive
and behavioral inflexibility commonly observed in children with the disorder.
With regard to the L-FPN and M-FPN, two other major networks that have been impli-
cated in autism and other neurodevelopmental disorders, components of these networks were
Network Neuroscience
1229
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
n
e
n
_
a
_
0
0
1
6
3
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Salience network dynamics in autism
consistently among the 10 with strongest coactivation with the M-CIN in all 5 observed CAPs.
Within the M-FPN, the components with high activation included the mPFC in States 2, 3,
and 4 as well as the PCC in State 2. For the L-FPN, the supramarginal gyrus (SMG) was a top
10 component in States 1 and 3, and bilateral regions of the L-FPN were strongly activated
in States 2 and 5. As activation of the L-FPN and M-FPN has been shown to be influenced
and controlled by the M-CIN (Uddin, 2015), this commonly observed CAP is consistent with
previous research examining interactions among these three networks (Goulden et al., 2014).
Only one previous study has specifically examined anterior insula dynamics in ASD. Us-
ing a large ABIDE sample, Guo and colleagues identified an anterior insula ROI using Neu-
rosynth (Yarkoni et al., 2011); they then used a sliding window analytic approach to show that
this ROI exhibited decreased FC with the mPFC and PCC in certain brain states compared
with TD individuals (Guo et al., 2019). Our current findings of reduced coactivation between
M-CIN and M-FPN in ASD is in line with this observation, and employs a method that over-
comes some of the limitations of sliding window correlation analyses (Lurie et al., 2020;
Preti, Bolton, & Van De Ville, 2017).
Limitations
A few limitations of the current work should be noted. First, our sample includes data collected
at three different sites. While this increases the sample size and statistical power, the use of
data across multiple sites presents its own limitations in that intersite variability may affect
the analyses. Though we selected data from a limited age range by using similar acquisition
parameters and participant eye status (eyes closed), and attempted to control for acquisition
site across all analyses, we cannot be sure that inherent between-site effects are completely
accounted for. In addition, we cannot ascertain whether diagnostic procedures and scanner
sessions were conducted in the same way for all participants at the three sites. There were also
some discrepancies in the information that was reported to the ABIDE database for the different
sites, eliminating the possibility of using behavioral measures like the Repetitive Behavior Scale
or the Social Communication Questionnaire to explore relationships between dynamic FC
parameters and symptom severity in the ASD group. Furthermore, similarly to most studies of
ASD, our sample consisted mostly of males, with only 34 female participants included in the
analyses. Although sex was included as a variable in our statistical analyses and had minimal
effect on the significant results reported, this imbalance of males to females may fail to account
for differences in the brain activity of the two sexes (Lai et al., 2017).
Despite these limitations, this study identifies the M-CIN, including the bilateral anterior
insula and anterior cingulate, as a network of particular interest for further investigation with
dynamic FC approaches in children with ASD. Further research could use tasks known to
elicit activation in this network to investigate the differential activation in children and ado-
lescents with ASD (Odriozola et al., 2016), as well as adults and older individuals, as patterns
of dynamic FC may change significantly with age (Hutchison & Morton, 2016, 2015). The
relationship between the M-CIN, M-FPN, and L-FPN in ASD also merits further investigation,
as the identification of connections or dynamic coactivations (or lack thereof) between these
networks may be utilized in the future as potential diagnostic identifiers of ASD and other
neurodevelopmental disorders (Uddin et al., 2017).
Conclusion
The findings of this study build on the growing body of literature on the use of dynamic connec-
tivity approaches in the investigation of neurodevelopmental disorders, and ASD in particular.
Network Neuroscience
1230
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
n
e
n
_
a
_
0
0
1
6
3
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Salience network dynamics in autism
Our combination of ICA and CAP allowed for the identification of highly specific networks
of interest, particularly those centered on the bilateral insular and anterior cingulate cortices.
The results of this study provide further insight into the dynamic FC abnormalities that may
underlie the clinical presentation of ASD, but future studies are needed to identify the neural
mechanisms by which these brain abnormalities are related to ASD symptoms, particularly
those related to inflexible behaviors.
ACKNOWLEDGMENTS
The authors gratefully acknowledge Willa Voorhies, Paola Odriozola, Kristafor Farrant, Dina
Dajani, Casey Burrows, Taylor Bolt and Shruti Vij for assistance with MRI data collection,
as well as Amy Beaumont, Sandra Cardona, Meaghan Parlade, and Michael Alessandri for
assistance with clinical assessments.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00163.
AUTHOR CONTRIBUTIONS
Emily Marshall: Data curation; Formal analysis; Visualization; Writing - Original Draft. Jason
Nomi: Conceptualization; Formal analysis; Funding acquisition; Investigation; Supervision;
Visualization; Writing - Original Draft; Writing - Review & Editing. Bryce Dirks: Data curation;
Supervision; Writing - Review & Editing. Celia Romero: Conceptualization; Data curation;
Supervision; Writing - Review & Editing. Lauren Kupis: Formal analysis; Supervision; Writing -
Review & Editing. Catie Chang: Formal analysis; Methodology; Resources; Writing - Review
& Editing. Lucina Q. Uddin: Conceptualization; Funding acquisition; Project administration;
Resources; Supervision; Writing - Original Draft; Writing - Review & Editing.
FUNDING INFORMATION
Lucina Q. Uddin, National Institute of Mental Health (http://dx.doi.org/10.13039/100000025),
Award ID: R01MH107549. Lucina Q. Uddin, Canadian Institute for Advanced Research.
Lucina Q. Uddin, University of Miami Gabelli Senior Scholar Award. Jason Nomi, National In-
stitute of Mental Health (http://dx.doi.org/10.13039/100000025), Award ID: R03MH121668.
Jason Nomi, Brain & Behavior Research Foundation.
REFERENCES
Abbott, A. E., Nair, A., Keown, C. L., Datko, M.,
Jahedi, A.,
Fishman, I., & Muller, R. A. (2016). Patterns of atypical functional
connectivity and behavioral links in autism differ between de-
fault, salience, and executive networks. Cerebral Cortex, 26(10),
4034–4045. DOI: https://doi.org/10.1093/cercor/bhv191, PMID:
26351318, PMCID: PMC5027998
Allen, E. A., Damaraju, E., Plis, S. M., Erhardt, E. B., Eichele, T.,
& Calhoun, V. D.
(2014). Tracking whole-brain connectivity
dynamics in the resting state. Cerebral Cortex, 24(3), 663–676.
DOI: https://doi.org/10.1093/cercor/bhs352, PMID: 23146964,
PMCID: PMC3920766
American Psychiatric Association. (2013). Diagnostic and Statistical
Manual of Mental Disorders (DSM-5). Washington, D.C: American
Psychiatric Publishing. DOI: https://doi.org/10.1176/appi.books
.9780890425596
Anderson, J. S., Nielsen, J. A., Froehlich, A. L., DuBray, M. B.,
Druzgal, T. J., Cariello, A. N., . . . Lainhart, J. E.
(2011). Func-
tional connectivity magnetic resonance imaging classification of
autism. Brain: A Journal of Neurology, 134(Pt 12), 3742–3754.
DOI: https://doi.org/10.1093/brain/awr263, PMID: 22006979,
PMCID: PMC3235557
Calhoun, V. D., Adali, T., Pearlson, G. D., & Pekar, J. J. (2001). A
method for making group inferences from functional MRI data
using independent component analysis. Human Brain Mapping,
14(3), 140–151. DOI: https://doi.org/10.1002/hbm.1048, PMID:
11559959, PMCID: PMC6871952
Calhoun, V. D., Miller, R., Pearlson, G., & Adali, T. (2014). The
chronnectome: Time-varying connectivity networks as the next
frontier in fMRI data discovery. Neuron, 84(2), 262–274. DOI:
Network Neuroscience
1231
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
n
e
n
_
a
_
0
0
1
6
3
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Salience network dynamics in autism
https://doi.org/10.1016/j.neuron.2014.10.015, PMID: 25374354,
PMCID: PMC4372723
Chang, C., & Glover, G. H.
(2010). Time–frequency dynamics of
resting-state brain connectivity measured with fMRI. Neuro-
Image, 50(1), 81–98. DOI: https://doi.org/10.1016/j.neuroimage
.2009.12.011, PMID: 20006716, PMCID: PMC2827259
Chen, H., Nomi, J. S., Uddin, L. Q., Duan, X., & Chen, H. (2017).
Intrinsic functional connectivity variance and state-specific
under-connectivity in autism. Human Brain Mapping, 38(11),
5740–5755. DOI: https://doi.org/10.1002/hbm.23764, PMID:
28792117, PMCID: PMC5783325
Chen, J. E., Chang, C., Greicius, M. D., & Glover, G. H. (2015).
Introducing co-activation pattern metrics to quantify sponta-
neous brain network dynamics. NeuroImage, 111, 476–488.
DOI: https://doi.org/10.1016/j.neuroimage.2015.01.057, PMID:
25662866, PMCID: PMC4386757
Ciric, R., Rosen, A. F. G., Erus, G., Cieslak, M., Adebimpe,
A., Cook, P. A., Bassett, D. S., Davatzikos, C., Wolf, D. H.,
& Satterthwaite, T. D.
(2018). Mitigating head motion arti-
fact in functional connectivity MRI. Nature Protocols, 13(12),
2801–2826. DOI: https://doi.org/10.1038/s41596-018-0065-y,
PMID: 30446748
Damaraju, E., Allen, E. A., Belger, A., Ford, J. M., McEwen, S.,
Mathalon, D. H., . . . Calhoun, V. D. (2014). Dynamic func-
tional connectivity analysis reveals transient states of dysconnec-
tivity in schizophrenia. NeuroImage Clinical, 5, 298–308. DOI:
https://doi.org/10.1016/j.nicl.2014.07.003, PMID: 25161896,
PMCID: PMC4141977
de Lacy, N., Doherty, D., King, B. H., Rachakonda, S., & Calhoun,
V. D. (2017). Disruption to control network function correlates
with altered dynamic connectivity in the wider autism spectrum.
NeuroImage Clinical, 15, 513–524. DOI: https://doi.org/10.1016
/j.nicl.2017.05.024, PMID: 28652966, PMCID: PMC5473646
Demirtas, M., Tornador, C., Falcón, C., López-Solà, M., Hernández-
Ribas, R., Pujol, J., . . . Deco, G. (2016). Dynamic functional con-
nectivity reveals altered variability in functional connectivity
among patients with major depressive disorder. Human Brain
Mapping, 37(8), 2918–2930. DOI: https://doi.org/10.1002/hbm
.23215, PMID: 27120982, PMCID: PMC5074271
Di Martino, A., O’Connor, D., Chen, B., Alaerts, K., Anderson, J. S.,
Assaf, M., . . . Milham, M. P. (2017). Enhancing studies of the
connectome in autism using the autism brain imaging data
exchange II. Science Data, 4, 170010. DOI: https://doi.org/10.1038
/sdata.2017.10, PMID: 28291247, PMCID: PMC5349246
Di Martino, A., Ross, K., Uddin, L. Q., Sklar, A. B., Castellanos, F. X.,
& Milham, M. P. (2009). Functional brain correlates of social and
nonsocial processes in autism spectrum disorders: An activation
likelihood estimation meta-analysis. Biological Psychiatry, 65(1),
63–74. DOI: https://doi.org/10.1016/j.biopsych.2008.09.022,
PMID: 18996505, PMCID: PMC2993772
Di Martino, A., Yan, C.-G., Li, Q., Denio, E., Castellanos, F. X.,
Alaerts, K., . . . Milham, M. P. (2014). The autism brain imag-
ing data exchange: Towards a large-scale evaluation of the intrin-
sic brain architecture in autism. Molecular Psychiatry, 19(6),
659–667. DOI: https://doi.org/10.1038/mp.2013.78, PMID:
23774715, PMCID: PMC4162310
Ebisch, S. J., Gallese, V., Willems, R. M., Mantini, D., Groen, W. B.,
Romani, G. L., Buitelaar, J. K., & Bekkering, H. (2011). Altered
intrinsic functional connectivity of anterior and posterior insula
regions in high-functioning participants with autism spectrum
disorder. Human Brain Mapping, 32(7), 1013–1028. DOI:
https://doi.org/10.1002/hbm.21085, PMID: 20645311, PMCID:
PMC6870194
Fu, Z., Tu, Y., Di, X., Du, Y., Sui, J., Biswal, B. B., Zhang, Z.,
de Lacy, N., & Calhoun, V. D.
increased
thalamic-sensory connectivity and decreased whole-brain dyna-
mism in autism. NeuroImage, 190, 191–204. DOI: https://doi
.org/10.1016/j.neuroimage.2018.06.003, PMID: 29883735,
PMCID: PMC6281849
(2019). Transient
Gotts, S. J., Saad, Z. S., Jo, H. J., Wallace, G. L., Cox, R. W., &
Martin, A. (2013). The perils of global signal regression for group
comparisons: A case study of autism spectrum disorders. Fron-
tiers in Human Neuroscience, 7, 356. DOI: https://doi.org/10.3389
/fnhum.2013.00356, PMID: 23874279, PMCID: PMC3709423
Goulden, N., Khusnulina, A., Davis, N. J., Bracewell, R. M., Bokde,
A. L., McNulty, J. P., & Mullins, P. G. (2014). The salience net-
work is responsible for switching between the default mode net-
work and the central executive network: Replication from DCM.
NeuroImage. DOI: https://doi.org/10.1016/j.neuroimage.2014.05
.052, PMID: 24862074
Green, S. A., Hernandez, L., Bookheimer, S. Y., & Dapretto, M.
(2016). Salience network connectivity in autism is related to
brain and behavioral markers of sensory overresponsivity. Journal
of the American Academy of Child and Adolescent Psychiatry,
55(7), 618–626. DOI: https://doi.org/10.1016/j.jaac.2016.04.013,
PMID: 27343889, PMCID: PMC4924541
Guo, X., Duan, X., Suckling, J., Chen, H., Liao, W., Cui, Q., &
Chen, H. (2019). Partially impaired functional connectivity states
between right anterior insula and default mode network in autism
spectrum disorder. Human Brain Mapping, 40(4), 1264–1275.
DOI: https://doi.org/10.1002/hbm.24447, PMID: 30367744,
PMCID: PMC3951650
Hutchison, R. M., & Morton, J. B.
(2015). Tracking the brain’s
Journal of
functional coupling dynamics over development.
Neuroscience, 35(17), 6849–6859. DOI: https://doi.org/10.1523
/JNEUROSCI.4638-14.2015,
PMCID:
PMC6605187
25926460,
PMID:
Hutchison, R. M., & Morton, J. B. (2016). It’s a matter of time: Re-
framing the development of cognitive control as a modification
of the brain’s temporal dynamics. Developmental Cognitive Neu-
roscience, 18, 70–77. DOI: https://doi.org/10.1016/j.dcn.2015
.08.006, PMID: 26375924, PMCID: PMC6990064
Hutchison, R. M., Womelsdorf, T., Allen, E. A., Bandettini, P. A.,
Calhoun, V. D., Corbetta, M., . . . Chang, C. (2013). Dynamic
functional connectivity: Promise, issues, and interpretations. Neuro-
Image, 80, 360–378. DOI: https://doi.org/10.1016/j.neuroimage
.2013.05.079, PMID: 23707587, PMCID: PMC3807588
Hyder, F., & Rothman, D. L. (2010). Neuronal correlate of BOLD
signal fluctuations at rest: ERR on the side of the baseline [Review
of Neuronal correlate of BOLD signal fluctuations at rest: ERR on
Network Neuroscience
1232
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
n
e
n
_
a
_
0
0
1
6
3
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Salience network dynamics in autism
the side of the baseline]. Proceedings of the National Academy of
Sciences of the United States of America, 107(24), 10773–10774.
DOI: https://doi.org/10.1073/pnas.1005135107, PMID:
20534504, PMCID: PMC2890714
Kaiser, R. H., Kang, M. S., Lew, Y., Van Der Feen, J., Aguirre, B.,
Clegg, R., . . . Pizzagalli, D. A. (2019). Abnormal frontoinsular-
default network dynamics in adolescent depression and rumina-
tion: A preliminary resting-state co-activation pattern analysis.
Neuropsychopharmacology, 44(9), 1604–1612. DOI: https://doi
.org/10.1038/s41386-019-0399-3, PMID: 31035283, PMCID:
PMC6784913
Kiviniemi, V., Starck, T., Remes,
J.,
Haapea, M., . . . Tervonen, O. (2009). Functional segmentation
of the brain cortex using high model order group PICA. Human
Brain Mapping, 30(12), 3865–3886. DOI: https://doi.org/10.1002
/hbm.20813, PMID: 19507160, PMCID: PMC6870574
J., Long, X., Nikkinen,
Kogan, M. D., Vladutiu, C. J., Schieve, L. A., Ghandour, R. M.,
Blumberg, S. J., Zablotsky, B., . . . Lu, M. C. (2018). The preva-
lence of parent-reported autism spectrum disorder among US
children. Pediatrics, 142, e20174161. DOI: https://doi.org/10
.1542/peds.2017-4161, PMID: 30478241, PMCID: PMC6317762
Lai, M.-C., Lerch, J. P., Floris, D. L., Ruigrok, A. N. V., Pohl, A.,
Lombardo, M. V., & Baron-Cohen, S. (2017). Imaging sex/gender
and autism in the brain: Etiological implications. Journal of Neuro-
science Research, 95, 380–397. DOI: https://doi.org/10.1002/jnr
.23948, PMID: 27870420
Lancaster, G., Iatsenko, D., Pidde, A., Ticcinelli, V., & Stefanovska,
A. (2018). Surrogate data for hypothesis testing of physical sys-
tems. Physics Reports, 748(18), 1–60. DOI: https://doi.org/10
.1016/j.physrep.2018.06.001
Liu, T. T., Nalci, A., & Falahpour, M.
(2017). The global signal in
fMRI: Nuisance or information? NeuroImage, 150, 213–229.
DOI: https://doi.org/10.1016/j.neuroimage.2017.02.036, PMID:
28213118, PMCID: PMC5406229
Liu, X., & Duyn, J. H.
(2013). Time-varying functional network
information extracted from brief instances of spontaneous brain
activity. Proceedings of the National Academy of Sciences of the
United States of America, 110(11), 4392–4397. DOI: https://doi
.org/10.1073/pnas.1216856110, PMID: 23440216, PMCID:
PMC3600481
Liu, X., Zhang, N., Chang, C., & Duyn, J. H. (2018). Co-activation
patterns in resting-state fMRI signals. NeuroImage, 180(Pt B),
485–494. DOI: https://doi.org/10.1016/j.neuroimage.2018.01.041,
PMID: 29355767, PMCID: PMC6082734
Lord, C., Rutter, M., DiLavore, P., Risi, S., Gotham, K., & Bishop,
(2012). Autism diagnostic observation schedule, second
S. L.
edition. Torrance, CA: Western Psychological Services.
Lurie, D. J., Kessler, D., Bassett, D. S., Betzel, R. F., Breakspear,
M., Kheilholz, S., . . . Calhoun, V. D.
(2020). Questions and
controversies in the study of time-varying functional connectiv-
ity in resting fMRI. Network Neuroscience, 4(1), 30–69. DOI:
https://doi.org/10.1162/netn_a_00116, PMID:
32043043,
PMCID: PMC7006871
Ma, Y., Ma, Z., Liang, Z., Neuberger, T., & Zhang, N. (2020). Global
brain signal in awake rats. Brain Structure & Function, 225(1),
227–240. DOI: https://doi.org/10.1007/s00429-019-01996-5,
PMID: 31802256, PMCID: PMC7391791
Menon, V., & Uddin, L. Q.
(2010). Saliency, switching, attention
and control: A network model of insula function. Brain Structure
& Function, 214(5–6), 655–667. DOI: https://doi.org/10.1007
/s00429-010-0262-0, PMID: 20512370, PMCID: PMC2899886
Müller, R.-A., Shih, P., Keehn, B., Deyoe, J. R., Leyden, K. M., &
(2011). Underconnected, but how? A survey of
Shukla, D. K.
functional connectivity MRI studies in autism spectrum disorders.
Cerebral Cortex, 21, 2233–2243. DOI: https://doi.org/10.1093
/cercor/bhq296, PMID: 21378114, PMCID: PMC3169656
Nair, S.,
Murphy, K., & Fox, M. D. (2017). Towards a consensus regarding
global signal regression for resting state functional connectivity
MRI. NeuroImage, 154, 169–173. DOI: https://doi.org/10.1016/j
.neuroimage.2016.11.052, PMID: 27888059, PMCID: PMC5489207
J. O.,
Jao Keehn, R. J., Berkebile, M. M., Maximo,
Witkowska, N., & Müller, R.-A. (2018). Local resting state func-
tional connectivity in autism: Site and cohort variability and the
effect of eye status. Brain Imaging and Behavior, 12(1), 168–179.
DOI: https://doi.org/10.1007/s11682-017-9678-y, PMID:
28197860, PMCID: PMC5628079
Nomi, J. S., Farrant, K., Damaraju, E., Rachakonda, S., Calhoun,
V. D., & Uddin, L. Q. (2016). Dynamic functional network
connectivity reveals unique and overlapping profiles of insula
subdivisions. Human Brain Mapping, 37(5), 1770–1787. DOI:
https://doi.org/10.1002/hbm.23135, PMID: 26880689, PMCID:
PMC4837017
Nomi, J. S., Molnar-Szakacs, I., & Uddin, L. Q. (2019). Insular func-
tion in autism: Update and future directions in neuroimaging
and interventions. Progress in Neuro-Psychopharmacology &
Biological Psychiatry, 89, 412–426. DOI: https://doi.org/10.1016
/j.pnpbp.2018.10.015, PMID: 30381235
Odriozola, P., Uddin, L. Q., Lynch, C. J., Kochalka, J., Chen, T., &
Menon, V. (2016). Insula response and connectivity during social
and non-social attention in children with autism. Social Cognitive
and Affective Neuroscience, 11(3), 433–444. DOI: https://doi
.org/10.1093/scan/nsv126, PMID: 26454817, PMCID:
PMC4769628
Padmanabhan, A., Lynch, C. J., Schaer, M., & Menon, V.
(2017).
The default mode network in autism. Biological Psychiatry. Cog-
nitive Neuroscience and Neuroimaging, 2(6), 476–486. DOI:
https://doi.org/10.1016/j.bpsc.2017.04.004, PMID: 29034353,
PMCID: PMC5635856
Power,
J. D., Plitt, M., Laumann, T. O., & Martin, A. (2017).
Sources and implications of whole-brain fMRI signals in humans.
NeuroImage, 146, 609–625. DOI: https://doi.org/10.1016/j
.neuroimage.2016.09.038, PMID: 27751941, PMCID:
PMC5321814
Preti, M. G., Bolton, T. A. W., & Van De Ville, D.
(2017). The
dynamic functional connectome: State-of-the-art and perspec-
tives. In NeuroImage, 160, 41–54. DOI: https://doi.org/10.1016
/j.neuroimage.2016.12.061, PMID: 28034766
Rashid, B., Blanken, L. M. E., Muetzel, R. L., Miller, R., Damaraju,
E., Arbabshirani, M. R., . . . Calhoun, V. (2018). Connectivity dy-
namics in typical development and its relationship to autistic
traits and autism spectrum disorder. Human Brain Mapping,
Network Neuroscience
1233
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
n
e
n
_
a
_
0
0
1
6
3
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Salience network dynamics in autism
39(8), 3127–3142. DOI: https://doi.org/10.1002/hbm.24064,
PMID: 29602272, PMCID: PMC6045960
Ray, K. L., McKay, D. R., Fox, P. M., Riedel, M. C., Uecker, A. M.,
Beckmann, C. F., . . . Laird, A. R. (2013). ICA model order selec-
tion of task co-activation networks. Frontiers in Neuroscience,
7, 237. DOI: https://doi.org/10.3389/fnins.2013.00237, PMID:
24339802, PMCID: PMC3857551
Saad, Z. S., Gotts, S. J., Murphy, K., Chen, G., Jo, H. J., Martin, A.,
& Cox, R. W. (2012). Trouble at rest: How correlation patterns
and group differences become distorted after global signal regres-
sion. Brain Connectivity, 2(1), 25–32. DOI: https://doi.org/10
.1089/brain.2012.0080,
PMCID:
PMC3484684
22432927,
PMID:
Scholvinck, M. L., Maier, A., Ye, F. Q., Duyn, J. H., & Leopold,
D. A. (2010). Neural basis of global resting-state fMRI activity.
Proceedings of the National Academy of Sciences of the United
States of America, 107(22), 10238–10243. DOI: https://doi.org
/10.1073/pnas.0913110107, PMID: 20439733, PMCID:
PMC2890438
Turchi, J., Chang, C., Ye, F. Q., Russ, B. E., Yu, D. K., Cortes, C. R.,
Monosov, I. E., Duyn, J. H., & Leopold, D. A. (2018). The basal
forebrain regulates global resting-state fMRI fluctuations. Neuron,
97(4), 940–952. DOI: https://doi.org/10.1016/j.neuron.2018.01
.032, PMID: 29398365, PMCID: PMC5823771
Uddin, L. Q. (2015). Salience processing and insular cortical func-
tion and dysfunction. Nature Reviews Neuroscience, 16(1), 55–61.
DOI: https://doi.org/10.1038/nrn3857, PMID: 25406711
Uddin, L. Q. (2017). Mixed signals: On separating brain signal from
noise. Trends in Cognitive Sciences, 21(6), 405–406. DOI:
https://doi.org/10.1016/j.tics.2017.04.002, PMID: 28461113,
PMCID: PMC6033047
Uddin, L. Q. (2020a). Bring the noise: Reconceptualizing spon-
taneous neural activity. Trends in Cognitive Sciences. DOI:
https://doi.org/10.1016/j.tics.2020.06.003, PMID: 32600967
Uddin, L. Q. (2020b). Brain mechanisms supporting flexible cogni-
tion and behavior in adolescents with autism spectrum disorder.
Biological Psychiatry. 2020 May 20:S0006-3223(20)31597-3.
DOI: https://doi.org/10.1016/j.biopsych.2020.05.010, PMID:
32709415, PMCID: PMC7677208
Uddin, L. Q., Dajani, D. R., Voorhies, W., Bednarz, H., & Kana,
R. K. (2017). Progress and roadblocks in the search for brain-
based biomarkers of autism and attention-deficit/hyperactivity
disorder. Translational Psychiatry, 7(8), e1218. DOI: https://doi
.org/10.1038/tp.2017.164, PMID:
28892073, PMCID:
PMC5611731
Uddin, L. Q., & Karlsgodt, K. H. (2018). Future directions for exam-
ination of brain networks in neurodevelopmental disorders. Jour-
nal of Clinical Child and Adolescent Psychology, 53, 47(3), 483–497.
DOI: https://doi.org/10.1080/15374416.2018.1443461, PMID:
29634380, PMCID: PMC6842321
Uddin, L. Q., & Menon, V. (2009). The anterior insula in autism:
Under-connected and under-examined. Neuroscience and Bio-
behavioral Reviews, 33(8), 1198–1203. DOI: https://doi.org/10
.1016/j.neubiorev.2009.06.002, PMID: 19538989, PMCID:
PMC2743776
Uddin, L. Q., Supekar, K., Lynch, C. J., Cheng, K. M., Odriozola,
P., Barth, M. E., Phillips, J., Feinstein, C., Abrams, D. A., &
Menon, V. (2015). Brain state differentiation and Behavioral In-
flexibility in Autism. Cerebral Cortex, 25(12), 4740–4747. DOI:
https://doi.org/10.1093/cercor/bhu161, PMID: 25073720, PM-
CID: PMC4635916
Uddin, L. Q., Supekar, K., Lynch, C. J., Khouzam, A., Phillips,
J., Feinstein, C., Ryali, S., & Menon, V.
Salience
network–based classification and prediction of symptom sever-
JAMA Psychiatry, 70(8), 869–879.
ity in children with autism.
DOI: https://doi.org/10.1001/jamapsychiatry.2013.104, PMID:
23803651, PMCID: PMC3951904
(2013).
Uddin, L. Q., Supekar, K., & Menon, V.
(2013). Reconceptualiz-
ing functional brain connectivity in autism from a developmental
perspective. Frontiers in Human Neuroscience, 7, 458. DOI:
https://doi.org/10.3389/fnhum.2013.00458, PMID: 23966925,
PMCID: PMC3735986
Uddin, L. Q., Thomas Yeo, B. T., & Nathan Spreng, R. (2019). To-
wards a universal taxonomy of macro-scale functional human
brain networks. Brain Topography, 926–942. DOI: https://doi.org
/10.1007/s10548-019-00744-6, PMID: 31707621
Vissers, M. E., Cohen, M. X., & Geurts, H. M. (2012). Brain connec-
tivity and high functioning autism: A promising path of research
that needs refined models, methodological convergence, and
stronger behavioral links. Neuroscience & Biobehavioral Re-
views, 36, 604–625. DOI: https://doi.org/10.1016/j.neubiorev
.2011.09.003, PMID: 21963441
Watanabe, T., & Rees, G. (2017). Brain network dynamics in high-
functioning individuals with autism. Nature Communications,
8, 16048. DOI: https://doi.org/10.1038/ncomms16048, PMID:
28677689, PMCID: PMC5504272
White, T., & Calhoun, V. D.
(2019). Dissecting static and dyna-
mic functional connectivity: Example from the autism spectrum.
Journal of Experimental Neuroscience. DOI: https://doi.org/10.1177
/1179069519851809, PMID: 31210734, PMCID: PMC6545633
Yan, C. G., Wang, X. D., Zuo, X. N., & Zang, Y. F. (2016). DPABI:
Data processing & Analysis for (Resting-State) Brain Imaging.
Neuroinformatics, 14(3), 339–351. DOI: https://doi.org/10.1007
/s12021-016-9299-4, PMID: 27075850
Yao, Z., Hu, B., Xie, Y., Zheng, F., Liu, G., Chen, X., & Zheng, W.
(2016). Resting-state time-varying analysis reveals aberrant varia-
tions of functional connectivity in autism. Frontiers in Human
Neuroscience, 10, 463. DOI: https://doi.org/10.3389/fnhum.2016
.00463, PMID: 27695408, PMCID: PMC5025431
Yarkoni, T., Poldrack, R. A., Nichols, T. E., Van Essen, D. C., &
Wager, T. D. (2011). Large-scale automated synthesis of human
functional neuroimaging data. Nature Methods, 8(8), 665–670.
DOI: https://doi.org/10.1038/nmeth.1635, PMID: 21706013,
PMCID: PMC3146590
Network Neuroscience
1234
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
4
4
1
2
1
9
1
8
6
7
0
8
2
n
e
n
_
a
_
0
0
1
6
3
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3