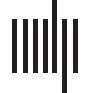INVESTIGACIÓN
Sleep spindles in primates: Modeling the
effects of distinct laminar thalamocortical
connectivity in core, matrix, y
reticular thalamic circuits
Arash Yazdanbakhsh1,2,3
, Helen Barbas2,3,4,5, and Basilis Zikopoulos2,3,5,6
1Computational Neuroscience and Vision Lab, Department of Psychological and Brain Sciences,
Boston University, Bostón, MAMÁ, EE.UU
2Graduate Program for Neuroscience, Boston University, Bostón, MAMÁ, EE.UU
3Center for Systems Neuroscience, Bostón, MAMÁ, EE.UU
4Neural Systems Laboratory, Program in Human Physiology, Department of Health Sciences,
College of Health and Rehabilitation Sciences (Sargent College), Boston University, Bostón, MAMÁ, EE.UU
5Department of Anatomy and Neurobiology, Boston University School of Medicine, Boston University, Bostón, MAMÁ, EE.UU
6Human Systems Neuroscience Laboratory, Program in Human Physiology, Department of Health Sciences,
College of Health and Rehabilitation Sciences (Sargent College), Boston University, Bostón, MAMÁ, EE.UU
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Palabras clave: First-order thalamic nuclei, High-order thalamic nuclei, Schizophrenia, Laminar
conectividad, Spindle dynamics, Spindle propagation
ABSTRACTO
Sleep spindles are associated with the beginning of deep sleep and memory consolidation
and are disrupted in schizophrenia and autism. In primates, distinct core and matrix
thalamocortical (TC) circuits regulate sleep spindle activity through communications that are
filtered by the inhibitory thalamic reticular nucleus (TRN); sin embargo, little is known about
typical TC network interactions and the mechanisms that are disrupted in brain disorders.
We developed a primate-specific, circuit-based TC computational model with distinct core
and matrix loops that can simulate sleep spindles. We implemented novel multilevel cortical
and thalamic mixing, and included local thalamic inhibitory interneurons, and direct
capa 5 projections of variable density to TRN and thalamus to investigate the functional
consequences of different ratios of core and matrix node connectivity contribution to spindle
dinámica. Our simulations showed that spindle power in primates can be modulated based
on the level of cortical feedback, thalamic inhibition, and engagement of model core versus
matrix, with the latter having a greater role in spindle dynamics. The study of the distinct
spatial and temporal dynamics of core-, matrix-, and mix-generated sleep spindles establishes
a framework to study disruption of TC circuit balance underlying deficits in sleep and
attentional gating seen in autism and schizophrenia.
RESUMEN DEL AUTOR
Sleep spindles are slow brain oscillations that are associated with the beginning of deep sleep,
aprendiendo, and memory storage and are disrupted in brain disorders. We developed a
computational model that can simulate sleep spindles in humans, using novel data on the
organization and connectivity of circuits that link the thalamus and cortex, and generate
spindles. Our model sheds light on the role of excitatory and inhibitory neurons in the network
dynamics and the functional consequences of differential engagement and connectivity of
un acceso abierto
diario
Citación: Yazdanbakhsh, A., Barbas, h.,
& Zikopoulos, B. (2023). Sleep spindles
in primates: Modeling the effects of
distinct laminar thalamocortical
connectivity in core, matrix, y
reticular thalamic circuits. Red
Neurociencia, 7(2), 743–768. https://doi
.org/10.1162/netn_a_00311
DOI:
https://doi.org/10.1162/netn_a_00311
Supporting Information:
https://doi.org/10.1162/netn_a_00311
Recibió: 19 Julio 2022
Aceptado: 1 Marzo 2023
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autores correspondientes:
Arash Yazdanbakhsh
yazdan@bu.edu
Basilis Zikopoulos
zikopoul@bu.edu
Editor de manejo:
Olaf Sporns
Derechos de autor: © 2023
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
Sleep spindles in core/matrix thalamocortical circuits
thalamic and cortical areas involved in typical brain function. Our work also establishes a
framework for the future study of the dynamics of variable types of sleep spindles and their
disruption that can lead to deficits in sleep, memory, and attention.
INTRODUCCIÓN
Sleep spindles:
Widespread brain oscillations with a
frequency of 7–15 Hz and duration
of 0.5–3 sec, associated with the
beginning of deep NREM sleep,
aprendiendo, and memory consolidation.
Sleep spindles are widespread oscillations associated with the beginning of deep NREM sleep,
aprendiendo, and memory consolidation (Lüthi, 2014; Steriade, 2005; Steriade, Domich, Oakson,
& Deschenes, 1987). Because sleep spindles are disrupted in a variety of neurological and
psychiatric conditions, they are an important clinical marker of atypical brain function in sev-
eral disorders, including schizophrenia and autism (Farmer et al., 2018; Ferrarelli et al., 2007;
Manoach et al., 2010; Mylonas et al., 2022).
Thalamic reticular nucleus (TRN):
Inhibitory nucleus that surrounds
the entire thalamus filtering
thalamocortical communications
through selective inhibition of
thalamic neurons.
Core thalamocortical (TC) circuitos:
Circuits between parvalbumin-
expressing thalamic projection
neuronas (PV+) that innervate focally
the middle cortical layers and receive
input from cortical pyramidal
neurons in layer 6.
Matrix thalamocortical (TC) circuitos:
Circuits between calbindin-
expressing thalamic projection
neuronas (CB+) that send widespread
innervation predominantly to the
upper cortical layers and receive
input from cortical pyramidal
neurons in layer 5.
Local thalamic inhibition:
Inhibition of thalamic projection
neurons by local thalamic
interneurons that are present in the
primate thalamus.
Neurociencia en red
Reciprocal connections between thalamic nuclei and cortical areas that are gated by the
inhibitory thalamic reticular nucleus (TRN) form thalamocortical (TC) circuits that regulate
sleep spindle activity (jones, 2007). Sin embargo, in mammals two anatomically and functionally
distinct TC circuits can be identified, the ‘core’ and the ‘matrix’ loops (jones, 1998; Müller
et al., 2020; Rovo, Ulbert, & Acsady, 2012; Zikopoulos & Barbas, 2007b), and it is not clear
how spindle activity and associated functions are regulated by each circuit through synaptic,
cellular, or regional interactions at the level of the cortex or thalamus. The core TC circuits,
prevalent in sensory thalamus, drive activity focally in the middle cortical layers. Sucesivamente, estos
core thalamic neurons are innervated by small ‘modulatory’ cortical axon terminals from pyra-
midal neurons in layer 6 (L6). The matrix TC circuits, prevalent in high-order thalamus, have a
complementary organization: a mix of large and small axon terminals from cortical layer 5 (L5)
pyramidal neurons drive activity of matrix thalamic neurons that, Sucesivamente, innervate broadly
and modulate the superficial cortical layers. The relative prevalence of the two TC loops dif-
fers, with core circuits prevailing in primary and unimodal association cortical areas and first-
order relay nuclei, whereas matrix circuits predominate in association and limbic cortical
areas and high-order thalamic nuclei (harris & Shepherd, 2015; jones, 1998; Rouiller &
Welker, 2000; Rovo et al., 2012; sherman & Guillery, 1996, 2006). Sin embargo, there is consid-
erable overlap of these parallel loops across TC networks, increasing the complexity of the
sistema (jones, 1998; Müller et al., 2020; Rovo et al., 2012; Zikopoulos & Barbas, 2007b).
En tono rimbombante, in addition to the excitatory connections in the thalamus, both TC circuits
likely engage an extensive network of local thalamic inhibitory interneurons that have
expanded significantly in evolution and constitute a hallmark of the primate thalamus (Arcelli,
Frassoni, Regondi, De Biasi, & Spreafico, 1997), increasing the complexity of potential inter-
actions in primates. Además, the connectivity of the inhibitory TRN with core and matrix TC
loops is not well studied. The TRN is a key generator of spindle oscillations that intercepts all
TC communications, but sends inhibitory projections only back to the thalamus (Lüthi, 2014;
Steriade, 2005; Steriade et al., 1987). The TRN gets most of its cortical input from L6 pyramidal
neurons that participate in core TC loops (Bourassa & Deschenes, 1995; Bourassa, Pinault, &
Deschenes, 1995; Kakei, Na, & Shinoda, 2001), but is predicted to get substantial input from
L5 pyramidal neurons of matrix TC loops in primates (Zikopoulos & Barbas, 2006, 2007a), como
was recently shown to some extent in mice (Hádinger et al., 2022; Hádinger et al., 2019;
Prasad, Carroll, & sherman, 2020).
Based on this evidence, we set out to test the impact of local thalamic inhibition, variable
levels of cortical L6 and L5 pyramidal neuron terminations in TRN, and the effects of these
744
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
connections in core and matrix TC circuits and spindle activity through the construction of a
rate-based computational TC model. Our model successfully simulated relay and filtered sig-
nals to sustain and propagate spindle oscillations with different powers, depending on the
level of cortical feedback, thalamic inhibition, and involvement of model core versus matrix
circuitos. Our simulations additionally revealed differences in the spatiotemporal dynamics of
core-generated, matrix-generated, or mixed types of spindles, highlighting novel circuit mech-
anisms involved in typical TC functions, like the sleep–wake cycle, sensory processing, atten-
tional gating, and memory consolidation. En tono rimbombante, characterization of novel interactions
between key nodes of TC circuits points to potential disruptions of mechanisms that may
underlie atypical spindle dynamics in disorders, including autism, and schizophrenia.
MÉTODOS, MODELS, AND THEORY
Model Design: Basic Connectivity Frame
The model is based on key molecular, anatómico, and connectivity features of distinct TC
circuits and their specialized interactions with the inhibitory TRN in primates that frame a
basic TC circuit, as elaborated below, and summarized in Figure 1.
There are two parallel functionally, structurally, and neurochemically distinct TC loops: el
core and matrix (jones, 2007). Thalamic neurons in core and matrix TC circuits of primates can
be distinguished neurochemically (Clasca, Rubio-Garrido, & Jabaudon, 2012; Münkle,
Waldvogel, & Faull, 2000). In core loops, excitatory TC projection neurons express the
calcium-binding protein parvalbumin (PV). Core PV+ excitatory thalamic neurons project
focally and drive activity of the middle cortical layers (principalmente 4, but also 3b, and 5a) y
get feedback from cortical layer 6. In the parallel matrix loops, excitatory TC projection
neurons express the calcium-binding protein calbindin (CB). Matrix CB+ thalamic neurons
innervate broadly and modulate primarily the superficial cortical layers (1–3a), recruiting a
broad horizontal spread that targets apical dendrites of pyramidal cortical neurons from layers 2,
3, y 5, which extend to the upper cortical layers. Cortical layer 5 neurons target and drive
activity of matrix CB+ thalamic neurons.
The entirely inhibitory TRN in primates covers almost the entire thalamus and gates all
reciprocal connections between the cortex and the thalamus (Zikopoulos & Barbas, 2007a).
The TRN receives input from excitatory thalamic projection neurons and pyramidal neurons in
the cortex that project to the thalamus. Anatomical and physiological data suggest that cortical
input drives TRN activity (Liu & jones, 1999; zhang & jones, 2004). Sucesivamente, the TRN sends
inhibitory projections only back to the thalamus. Until recently, it was thought that layer 6, pero
not layer 5, cortical pyramidal neurons project to TRN (Guillery, 1995, 2005; Guillery, Feig, &
Lozsádi, 1998; Guillery & Harting, 2003; Guillery & sherman, 2002; Kakei et al., 2001;
Rouiller & Durif, 2004; Rouiller & Welker, 1991, 2000; sherman & Guillery, 2002). We first
provided strong indirect evidence for a projection from layer 5 to TRN when we studied
prefrontal corticothalamic projections and compared them with projections from sensory asso-
ciation cortices and other corticosubcortical projections in primates (Zikopoulos & Barbas,
2006, 2007a). Like other cortices, prefrontal areas project to the thalamus mainly from layer
6, but also issue significant projections from layer 5 (xiao, Zikopoulos, & Barbas, 2009). Nosotros
found that prefrontal layer 5 axon terminations in the thalamus constituted a mix of large and
small boutons (thin and thick connections in Figure 1), and overall they were larger than
terminals from sensory association areas known to originate from layer 6. Recent studies in
mice confirmed that there are direct projections of variable density from L5 pyramidal neurons
to TRN from some cortical areas, especially association areas that likely participate in matrix
Neurociencia en red
745
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
Cifra 1. The proposed basic thalamocortical (TC) core, matrix, and mix circuit. The model circuit was based on two parallel thalamocortical
loops. Core: PV+ excitatory thalamic neurons project focally to the middle cortical layers and get feedback from cortical layer 6. Matrix: CB+
thalamic neurons innervate broadly the superficial cortical layers and receive projections from cortical layer 5. Layer 5 terminals in the thalamus
are larger (thick arrowheads) than terminals from layer 6. Thalamic and cortical neurons project to TRN (TRNC and TRNM). Mix: The two
parallel loops cross-connect at the level of the thalamus and/or cortex. The mix dynamics were implemented in three different ways, indicated
by dashed lines cross-linking core and matrix loops in panels A–C: (1) corticoreticular, where L6 and L5 concurrently excited TRNM and TRNC;
(2) thalamoreticular, where PV+ and CB+ concurrently excited TRNM and TRNC; y (3) cortical, where mixing of core and matrix networks
occurred at the level of the cortex, through columnar laminar interactions, simplified as a cross-connection between L5 and L6 (dashed line);
this way, we could modify core (L6 and PV+) versus matrix (L5 and CB+) contribution in the mix. (A) In closed loop connections, TRN neurons
directly inhibit the thalamic neurons that excite them. (B) The anatomical circuits underlying open loop connections are shown, in which TRN
neurons do not directly inhibit the thalamic neurons that excited them. (C) In the model we simulated the open loop architecture through
implementation of the closed loop connections in which TRN neurons directly inhibit the thalamic neurons that excited them, following a
symmetric Gaussian spread with the peak at the reciprocal thalamic neuron. Respectivamente, the bilateral and symmetric Gaussian connectivity
strength from TRN to thalamus resembles bilateral and symmetric open loops with opposite open directions (balanced). We called this the
Hybrid Loop design. Inhibitory model neurons are shown in red, including TRN (TRNC and TRNM for core and matrix), PV−, and CB− local
thalamic inhibitory interneurons. The local inhibitory thalamic neurons PV− and CB− send GABAergic projections to thalamic projection neu-
ron (PV+ and CB+; shown in panels B and C) and can receive excitatory input from the periphery, subcortical nuclei, including TRN, y
cortical areas. For simplification of the schematic diagram, the local inhibitory thalamic neurons PV− and CB− in B are shown to receive inhib-
itory input only from TRN to highlight their potential role in open loop TRN-TC architecture, whereas in C they are shown to receive input from
TRN or from cortex and they send inhibitory projections to TC neurons. Blue units are core excitatory elements, eso es, neurons in cortical layers
4 y 6 (L3b–5a and L6), as well as thalamic PV+ neurons. Green units are matrix excitatory units, eso es, neurons in cortical layer 5 (L5), cuyo
apical dendrites reach upper layers (L1–3a) and receive widespread input from excitatory thalamic CB+ neurons. Note that in the primate
thalamus, PV and CB stain cell bodies and dendrites of core and matrix TC projection neurons, respectivamente. There is some neuropil staining,
especially PV axons and terminals, that come mainly from TRN and basal ganglia; sin embargo, the cellular label is predominant and clearly
distinguishes the two systems. In several nuclei, Por ejemplo, the ventral anterior, PV, and CB neurons are intermingled and at equal numbers,
so it is not possible to define pure core or matrix regions (Zikopoulos & Barbas, 2007b). (D) When the Gaussian peak of connectivity strength is
to the thalamic neuron that directly excites the TRN neuron, due to spatially symmetric bilateral Gaussian spread of connectivity strength, el
circuit is composed of two open loops in opposite directions (balanced), then our architecture is functionally similar to a closed loop.
Mixing of core and matrix networks:
The two parallel core and matrix
loops can be isolated or cross-
connect at multiple nodes either
within the thalamus (thalamoreticular
and corticoreticular cross-
connections) or the cortex
(corticocortical cross-connections).
circuitos (Hádinger et al., 2019, 2022; Prasad et al., 2020). Since the presence of L5 projections
to TRN is not ubiquitous, we represent it with a dotted line from L5 to TRN in the simplified
diagram in Figure 1, which shows the main nodes of core and matrix circuits.
There is evidence that TRN neurons are paired with thalamic neurons, forming reciprocal,
closed loop circuits, in which TRN neurons send GABAergic input to the thalamic neurons
that innervate them directly (Figura 1A) (Marrón, Taheri, Kenyon, Berger-Wolf, & Llano,
2020; FitzGibbon, Solomon, & Goodchild, 2000; Gentet & Ulrich, 2003; Hale, Sefton, Baur,
& Cottee, 1982; Lo & sherman, 1994; McAlonan, Cavanaugh, & Wurtz, 2006; Pinault, 2004;
Pinault & Deschenes, 1998; sherman & Guillery, 1996; Shosaku, 1986; Steriade, McCormick,
& Sejnowski, 1993; Warren, Agmon, & jones, 1994; willis, Slater, Gribkova, & Llano, 2015),
Neurociencia en red
746
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
Open loop circuits:
In open loop connections, TRN
neurons do not directly inhibit the
thalamic neurons that excited them.
Gaussian connectivity strength
from TRN to thalamus resembles
bilateral but asymmetric open loops
with opposite open directions
(not balanced). Due to spatially
asymmetric bilateral Gaussian spread
of connectivity strength, the circuit is
composed of two unbalanced open
loops in opposite directions therefore
our architecture is functionally
similar to an open loop.
as reviewed in Jones (2007). A diferencia de, in open loop circuits, TRN neurons innervate other
thalamic neurons that do not directly innervate them (Figura 1B) (Crabtree, 1998; Crabtree,
Collingridge, & Isaac, 1998; Crabtree & Isaac, 2002; Kimura, 2014; Justicia & sherman, 2005;
Sotavento, Cruikshank, & Connors, 2010; McAlonan et al., 2006; Pinault & Deschenes, 1998).
In our model circuit we accounted for the TRN-thalamic loop architecture complexity by
representing innervation strength with Gaussian curves that have variable spread. We called
this architecture Hybrid Loop Design (Figura 1C). When the Gaussian peak of connectivity
strength is to the thalamic neuron, which directly excites the TRN neuron due to spatially sym-
metric bilateral Gaussian spread of connectivity strength, the loop is composed of two open
loops in opposite directions (balanced; Figure 1D), then our architecture is functionally similar
to a closed loop. En cambio, when the connectivity spread is either spatially asymmetrical or
follows a shifted Gaussian, then our circuit architecture resembles an open loop. Además,
in primates, there is extensive presence of local inhibitory neurons in the thalamus, shown as
PV− and CB− in Figure 1 (jones, 2007), which may participate in open loop circuits when
innervated by TRN and, Sucesivamente, innervate TC neurons. Little is known about the connectivity
of local inhibitory thalamic neurons, but studies have shown that they receive input from the
sensory periphery, the cortex, and the amygdala and form synaptic triads in the thalamus
(Kultas-Ilinsky, Hacer, & Ilinsky, 1995; Tai, Hacer, Ilinsky, & Kultas-Ilinsky, 1995; Timbie, Garcia-
Cabezas, Zikopoulos, & Barbas, 2020), as reviewed in Jones (2007).
The two parallel core and matrix circuits are not necessarily isolated, but can overlap across
TC networks. This overlap can be extensive in high-order TC networks, like the ones linking
the mediodorsal nucleus (Maryland) or the ventral anterior nucleus with the prefrontal cortex (jones,
1998; Müller et al., 2020; Rovo et al., 2012; Zikopoulos & Barbas, 2007b). Sin embargo, it is not
known whether this overlap is limited at the level of the thalamus, or if it is also present at the
level of the cortex and TRN. To account for the potential overlap of core and matrix circuits at
each TC node, we included cross-connections (dashed lines in Figure 1) that can facilitate
mixing of the two parallel loops at all levels. For cortical core contribution in the mix, L6
directly projects to TRNM, and for cortical matrix contribution in the mixing, L5 directly pro-
jects to TRNC. For thalamic core contribution in the mix, PV+ directly projects to TRNM, y
for thalamic matrix contribution in the mix, CB+ directly projects to TRNC. We also include
mixing of core and matrix networks at the level of the cortex through several laminar interac-
ciones, simplified in Figure 1 as a cross-connection between L5 and L6 (dashed line). The source
codes for the neural model are available in the Supporting Information.
Mathematical Specification of the Rate Model: Dynamics of Model Neurons
We constructed a computational model of core and matrix TC circuits based on the connec-
tivity above. We expressed each simulated neuron’s activity, based on inhibitory and excit-
atory inputs, con el tiempo, as a first-order differential equation. Model neurons were represented
by a single-compartment voltage (v) con el tiempo (Layton, Mingolla, & Yazdanbakhsh, 2012,
2014; Layton & Yazdanbakhsh, 2015) that obeys the following shunting equation:
dv
dt
¼ −Av − v − Eext
d
d
ÞIext − v − Einh
ÞIinh
(1)
In this equation, A denotes the constant decay (leakage) tasa, which brings v back to zero
when there is no excitatory or inhibitory input to the neuron to fulfill the physiological con-
straint that a neuron without input finally goes back to its resting potential. Iext and Iinh specify
the summed excitatory and inhibitory inputs to the neuron at each time. The terms Eext and Einh
refer to excitatory and inhibitory reversal potentials, respectivamente, which keep v within Eext and
Neurociencia en red
747
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
Einh range to fulfill the physiological constraint of the limited dynamical range of each neuron
actividad. We set Eext and Einh equal to 1 and −1; por lo tanto, our model neurons activities fell
dentro [−1, 1] range. Mesa 1 shows Iext and Iinh for each model neuron type as illustrated in
Cifra 1.
To facilitate description of excitatory/inhibitory interactions (Iext and Iinh), we constructed
thalamic, reticular, and cortical networks as a distance-dependent on-center- off-surround
shunting network, as we have described in the past (Layton et al., 2012, 2014; Layton &
Yazdanbakhsh, 2015; Sherbakov & Yazdanbakhsh, 2013; Wurbs, Mingolla, & Yazdanbakhsh,
2013) and recently in computational models of cortico-thalamic-amygdalar circuits (John,
Bullock, Zikopoulos, & Barbas, 2013; John, Zikopoulos, Bullock, & Barbas, 2016). Semejante
distant-dependent on-center interareal interactions are modeled by bell-shaped (Gaussian)
profiles reflected in Iext column of Table 1 por (X Ò Y ) symbols, in which X and Y indicate
the presynaptic and postsynaptic regions, Por ejemplo, in row 1 de mesa 1, L5 and CB+, respetar-
activamente. Similarmente, the off-surround distant dependent reflected in the Iinh column. Networks of
this type offer a simple, biologically plausible means to implement contrast enhancement and
a host of other processes by varying the strength of the off-surround inhibition to modulate the
tuning curve of each cell (Cao, Mingolla, & Yazdanbakhsh, 2015; Qian & Yazdanbakhsh,
2015). Strong inhibition yields sharp contrast, fine tuning, and high attention. Weak inhibition
leads to spreading activity, lower contrast, and inattentiveness. These processes may be altered
in autism and schizophrenia, leading to deficits in attentional gating (Medalla & Barbas, 2009).
De este modo, modulating the balance between excitation and inhibition in various nodes of the circuit
can provide a way to investigate the effects of various network elements on signal processing
and attention.
In the model, we took into account both anatomical (conectividad) and physiological dif-
ferences in L5 and L6 neurons and their terminals. In our simulations, these differences were
reflected by the parameterization of the model, so that the L5 Ò CB connection was set to be
20 veces (∼1 order of magnitude) stronger than L6 Ò PV. This is in line with physiology data
suggesting more burst firing and larger excitatory postsynaptic potentials with more paired
pulse depression for L5 neurons and terminals (Agmon & Connors, 1992; Chagnac-Amitai,
Luhmann, & Príncipe, 1990; de Kock et al., 2021; Greenhill, Ranson, & Fox, 2015; Larkum,
Zhu, & Sakmann, 1999; Livingstone, Hombre libre, & Hubel, 1996; Llano & sherman, 2009; Shai,
Anastassiou, Larkum, & Koch, 2015), but also with the anatomy, which has shown that L5
terminals in the thalamus are much larger than L6 terminals and can drive CB neuron activity
(Bourassa et al., 1995; Hoogland, Wouterlood, Welker, & Van der Loos, 1991; Llano &
sherman, 2009; Zikopoulos & Barbas, 2007b). Varying the relative strength of L5 Ò CB con-
nection changed the oscillatory tendency of the matrix loop (shown in the Results).
Equations for Model Neurons
Ecuación 1 is the general equation for all neurons of the model circuit of Figure 1. The key
distinction between model neurons is based on their excitatory (Iext) and inhibitory (Iinh) inputs
stemming from the neuroanatomy of the connectivity; por lo tanto, the current neural model
reflects the impact of neuroanatomy in generating distinct spatiotemporal dynamics of core
and matrix TC loop spindles (Piantoni, Halgren, & Cash, 2016), based on the position and
distribution of neurons rather than differences of reticular, thalamic, and cortical neuron types.
Each row of Table 1 represents the presynaptic total excitatory and inhibitory input to each
model neuron (es decir., CB+, TRNM, etc.). Indices i and j indicate the spatial position of model
neurons i and j on a one-dimensional array. Pre- to postsynaptic connections are characterized
Neurociencia en red
748
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
S
yo
mi
mi
pag
s
pag
i
norte
d
yo
mi
s
i
norte
C
oh
r
mi
/
metro
a
t
r
i
X
t
h
a
yo
a
metro
oh
C
oh
r
t
i
C
a
yo
C
i
r
C
tu
i
t
s
Þji TRNj
METRO
ÞjiTRNj
C
t
norte
mi
w
oh
r
k
norte
mi
tu
r
oh
s
C
mi
norte
C
mi
i
Model neuron
Symbol
Excitatory input (Iext)
Inhibitory input (Iinh)
Mesa 1. Model neurons and their excitatory and inhibitory inputs (ver figura 1)
Matrix excitatory thalamic neurons
Core excitatory thalamic neurons
Matrix inhibitory thalamic neurons
Matrix inhibitory thalamic neurons
TRN matrix
TRN core
Superficial layer neurons (SLN)
Middle layer neurons (MLN)
Cortical layer 5 neuronas
Cortical layer 6 neuronas
CBþ
i
PV þ
i
CB−
i
PV −
i
TRNi
METRO
TRNi
C
li
1−3a
li
3b−4
li
5
li
6
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
L5 → CBþ
d
ji Lj
Þ
5
L6 → PV þ
d
Þ
jiLj
6
CBþ → CB−
d
Þ
jiCBþ
j
PV þ → PV −
d
Þ
jiPV þ
j
CBþ → TRNM
d
PV þ → TRNC
d
ji CBþ
Þ
jþη þ L5 → TRNM
d
Þ
jiLj
5
jiPV þ
Þ
jþη þ L6 → TRNC
d
Þ
jiLj
6
CBþ → L1−3a
d
ji CBþ
Þ
jþη
PV þ → L3b−4
d
jiPV þ
Þ
jþη
d
L1−3a → L5
Þ
jiLj
1−3a
d
L3b−4 → L6
Þ
jiLj
3b−4
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
PN
j¼1
CB− → CBþ
d
Þ
jiCB−
j−η þ TRNM → CBþ
d
PV − → PV þ
d
ji PV −
Þ
j−η þ TRNC → PV þ
d
TRNM → CB−
d
jiTRNj
Þ
METRO
TRNC → PV −
d
Þ
jiTRNj
C
d
TRNM → TRNM
Þ
jiTRNj
METRO
d
TRNC → TRNC
jiTRNj
Þ
C
Nota. η = offset to simulate open loop (ver figura 1); → = intercortical, thalamocortical, thalamoreticular, or corticoreticular connection.
7
4
9
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
por (Pre Ò Post) symbols subindexed by ji indicating the jth presynaptic to ith postsynaptic
neuron strength. The relative strength of jth presynaptic to ith postsynaptic neuron follows
a normal distribution centered at i (es decir., the ith pre- to ith postsynaptic connection is the
strongest ).
d
pre → post
Þ
¼
ji
pag
1
p e−1
ffiffiffiffiffiffi
2Pi
2
j−i
σð Þ2
(2)
Spindles in the Model TRN
Neurons in the TRN send inhibitory GABAergic projections to thalamus (Cifra 1). TRN
neurons can fire in bursts, generating inhibitory postsynaptic potentials (IPSPs) in excitatory
thalamic neurons (PV+ and CB+), which in turn exhibit rebound bursts, activating the TRN
neuronas. By simulating neurons using the rate-based approach detailed above, we first intro-
duced burst-like activity input in our model TRN neurons (Figura 2A), which mimics the tem-
poral dynamic of constituent bursts in a spindle. The y-axis in Figure 2A shows normalized
values with the lower and upper limits of −1 and +1, similar to the lower and upper bounds
of all of the model neurons’ activities. The interburst interval was set to 100 ms to replicate a
10 Hz burst rate in 0.5 segundo. Such a voltage induction in model neurons is within the physio-
logical range of sleep spindles with a frequency of 7–15 Hz and duration of 0.5–3 sec (Lüthi,
2014; Steriade, 2005; Steriade et al., 1987). In order to have the input temporal dynamics of
Figure 2A as close as possible to the burst sequence of spindles, we approximated physiolog-
ical voltage patterns shown in previous work (McCormick & Bal, 1997) by a sixth-degree poly-
nomial to offer a slow initial increase and then a sharp raise mimicking T-current before the
burst spikes. After this stage, the burst is approximated by a densely packed spike-like pattern
entre 0 y 1. For visibility, the burst spike peaks are slightly decreasing sequentially with
no effect on the model activity dynamics. We therefore approximated the activity of TRN neu-
rons recorded intracellularly, in vivo, which communicate with each other through GABAA
receptor-mediated synapses, inducing chloride ion channel-mediated IPSPs. The chloride
reversal potential is approximately −71 mV, which is relatively depolarized compared to the
−78 mV resting membrane potential of TRN neurons resulting in IPSP-triggered low-threshold
spikes (LTSs) (Bazhenov, Timofeev, Steriade, & Sejnowski, 1999). We approximated the −78 to
−71 mV dynamic leading to LTS by a sixth-degree polynomial (slow to fast upward curving up
before burst in Figure 2A).
Through injection of spindle-pattern input in TRN, we bypassed sleep spindle generation in
the TRN and focused on the examination of the dynamics of core and matrix loops, in terms of
oscillatory activity and spindle tendency in the cortex and thalamus. Sin embargo, since the
modelo, based on its architecture, could oscillate independently, we additionally provided
the model response to tonic depolarizing, hyperpolarizing, or 0.5-sec square inputs to show
that the model also generates spindle-like oscillation in response to tonic inputs (Figure 2C–E).
We used this approach, to additionally test oscillatory activity and spindle tendency in our
simulations.
TRN Induced IPSPs in TC Neurons and Rebound
In the model circuit of Figure 1, TRN neurons project to the excitatory thalamic neurons in
core (PV+) and matrix (CB+) by inhibitory connections (representing GABA synapses). Cuando
model TRN neurons receive the spindle inducing input (Figura 2A), they generate IPSPs in TC
neuronas (Lüthi, 2014; McCormick & Bal, 1997). The postsynaptic potential in TC neurons
induces a hyperpolarizing H-current leading to cation low-threshold Ca2+ channel T-currents
Neurociencia en red
750
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
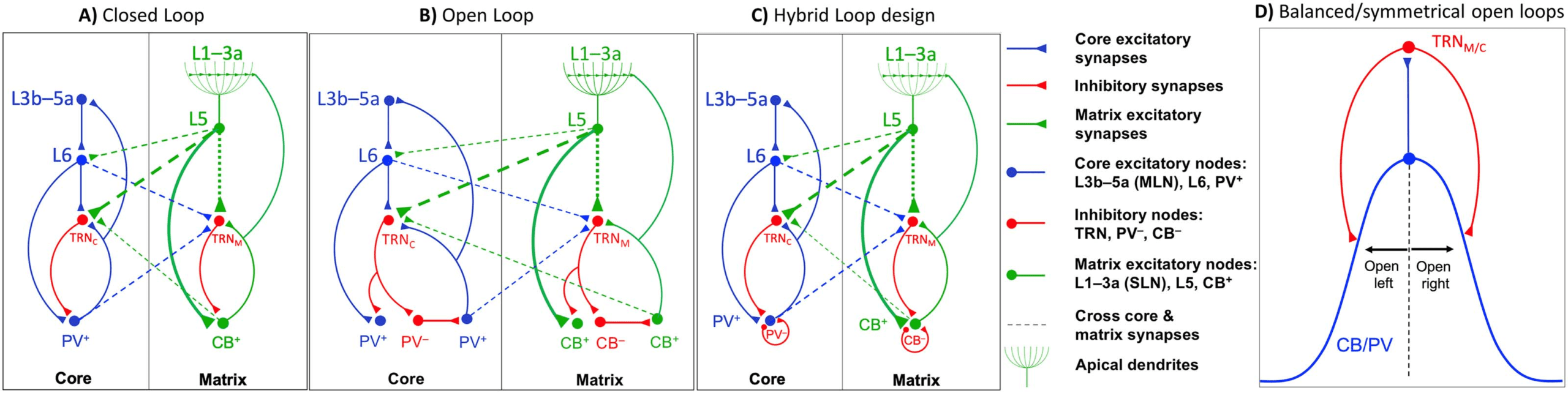
Cifra 2. Model inputs and response. (A) Model TRN input resembling spindle burst temporal pattern. Spindle generation in the model is
based on inducing TRN neurons by inputs resembling the spindle spike temporal patterns, which can entrain the model thalamocortical loop
depending on the tendency of rebound depolarization and oscillation. Throughout the reported simulations, we chose a 10 Hz (100-ms inter-
burst interval) spindle-pattern input with a 500-ms duration to induce the model TRN units. To set the TRN spindle inducing pattern as similar
to a physiological spindle as possible, we considered physiological properties of TRN neurons (reviewed in McCormick & Bal, 1997) y
approximated the initial membrane potential raise and then an accelerated upturn (T-current) before the spikes burst by a sixth-degree poly-
nomial. We also approximated the bursts by densely packed spike-like pattern between 0 y 1 to have the model neuron activities within the
normalized values throughout the simulation. (B) Model response to 0.5-sec tonic input to PV. Each layer neuron activity is amplitude scaled to
prevent overlap for clear visibility. During the input onset, each model neuron depolarizes, reaches its peak, reverses, and after a few oscil-
lations settles. After turning off the square input at 0.5 segundo, the activities drop sharply toward baseline followed by hyperpolarization, rebound
depolarization, and oscillations to finally settle on the resting state. Response to a 0.5-sec square input illustrates the dynamics of the rate-
based model and its tendency for rebound depolarization. (C) Model response to a depolarizing input pulse. After PV depolarization, hay
hyperpolarization and a rebound depolarization. (D) Model response to a hyperpolarizing input (due to a brief input pulse to TRN). PV neurons
exhibit hyperpolarization first and then they rebound depolarize. (mi) Model response to 0.5-sec depolarizing tonic input, while the network is
in high spindle tendency, due to increased levels of TC inhibition by TRN, shows oscillatory activity of higher amplitude and longer duration
compared to the response in panel B with the network in lower spindle tendency; here too, each layer neuron activity is amplitude scaled to
highlight PV oscillations. (F) Band-pass filtering typically used to filter the signal in the relevant frequency bands; in this example, 5–15 Hz
produces output similar to the original model output.
before the rebound bursts. The bursts of TC neurons activate the TRN neurons through the
excitatory synapses of the model circuit (Cifra 1; representing glutamatergic input to gluta-
mate receptors of TRN neurons). Figure 2B shows the mutual interaction of model TRN and TC
(es decir., PV+) neurons induces rebound activity of TC neurons after hyperpolarization (ver, p.ej.,
red PV+ curve right after 0.5 segundo). The rebound in the simple compartment rate-based model
stems from the decay term −Av in Equation 1 which brings the hyperpolarized and/or depo-
larized v back to zero with the rate A combined with the excitatory and inhibitory neurons’
Neurociencia en red
751
Sleep spindles in core/matrix thalamocortical circuits
interactions in the model circuit. Although the simplified mechanisms in the model are by far
less sophisticated than the actual physiology of membrane voltage modulation through the
variety of hyperpolarization activated cationic H-currents, low-threshold Ca2+ channels
T-current, neurotransmitter gated and voltage gated channels, they nevertheless, approximate
the hyperpolarization and rebound of TC neurons.
Análisis de los datos
Figure 2B illustrates oscillation tendency of the model core thalamocortical loop; after the
input is discontinued, activity drops sharply to rest, further reduces toward hyperpolarization,
after which activity rebounds toward depolarization. Such a dynamic can cycle a few more
veces, and more cycles indicate more rebound depolarizations after input shutdown, cual
are substrates for spindle sustaining. Por lo tanto, we define spindle tendency index (STI ) como
the total duration (in seconds) of the sequence of depolarization rebounds after hyperpolari-
zación. We consider 1% of maximum activity level of a model neuron as the threshold for
the peak of a depolarization to be counted as a rebound for inclusion within the
hyperpolarization-depolarization duration after input shutdown. El 1% thresholding
stemmed from the minor, subthreshold depolarization needed to activate the H-current in neu-
rons in vivo to initially and partially depolarize the membrane before the T-current taking over
and leading to bursting. Changing the threshold did not change the ordinal relations of spindle
tendencies across conditions. We considered the unit of time in seconds because average
spindle duration is ∼1 sec, and therefore the number of seconds conveys how many times
of an average spindle duration a hyperpolarization-depolarization sequence lasts as an indi-
cator of spindle tendency. Por ejemplo, in Figure 2B, the duration of the above threshold
hyperpolarization-depolarization sequence after the input shutdown is 0.37 segundo; hence,
STI = 0.37. The index enables us to compare the tendency of model core, matrix, and mix
TC circuits to sustain spindles, as reported in the Results section. We additionally checked
for oscillatory spindle tendency through filtering of signals within the relevant frequency band
being thresholded (band-pass filtering 5–15 Hz; Figura 2F).
Además, we visualized the spatiotemporal characteristics of core, matrix, and mixed TC
circuits using heat map matrices, y, to facilitate comparisons, we calculated dot products of
vectorized matrices and then normalized the dot product by the absolute value of each vector,
→ : B
→= Aj
j Bj
→
j, in which A
→
y B
eso es, A
are vectorized heat map matrices and |A| y |B| son
their size (escalar). This normalized value is between −1 and 1 (in all of our cases between 0
→
y 1); closer to 1 means more similarity (parallel) between A
and closer to 0 medio
more dissimilarity (orthogonal). In order to show the similarity and difference with a finer grain,
we calculated the ArcCos of cosine similarity, eso es, θ = ArcCos
to obtain the
angle θ between spatiotemporal patterns. θ = 0 means complete similarity (parallel), y el
more θ deviates from zero the similarity decreases.
→ = Aj
→
y B
→ : B
A
(cid:4)
j
j Bj
(cid:3)
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
RESULTADOS
Rate-Based Model Simulations: Ability to Sustain and Propagate Activity
We constructed a computational TC circuit that included core and matrix components with an
optional and variable cortical L5 to TRN projection (L5-TRN ON/OFF). Based on the features
of TC circuits, our model was able to simulate relay and filtering of signals and could propa-
gate and sustain spindle oscillations. Tal como, throughout the simulation results reported in the
following sections, the state of the network supporting the spindle was based on the presence
Neurociencia en red
752
Sleep spindles in core/matrix thalamocortical circuits
and amplitude of single vs sustained oscillations. Además, parameterization of the model,
so that the L5 Ò CB connection was set to be 20 veces (∼1 order of magnitude) stronger than
L6 Ò PV to reflect anatomical (conectividad) and physiological differences in L5 and L6 neu-
rons and their terminals, appeared to be necessary to keep the oscillatory tendency of the
matrix loop. This is due to major connectivity pattern differences of the matrix loop, cual
has spatially broader modulatory input to the upper layers of the cortex, compared to the spa-
tially narrower, feedforward driving input of the core TC in the middle layers. Two orders of
magnitude stronger L5 Ò CB connection compared to L6 Ò PV also worked, showing a wider
connectivity strength tolerance for the matrix loop. The limit in our rate-based model was 3
orders of magnitude in which matrix lost its oscillatory tendency.
To evaluate the readiness of the network to generate or maintain spindles, we gave the
model TRN neurons of the model core TRNc the spindle-like input depicted in Figure 2 para
500 ms with starting time at 0 reflected on the x-axis. As indicated in Figure 2 and elaborated
in the Data Analysis section, we shut down the spindle-like input at time 0.5 segundo. After the
spindle input induction, we left the network on its own, to see how the model TC loop could
sustain postspindle oscillations (sequential hyperpolarizations and depolarization rebounds) a
estimate the STI. STI is a relative measure of network spindle tendency that can be used to
compare the model core, matrix, and mix in supporting spindle generation and sustaining.
Using STI, we can also compare model spindle tendency with and without layer 5 corticotha-
lamic projections to TRN, local thalamic inhibition, and with mixing of parallel loops. Nosotros
additionally checked for oscillatory spindle tendency through band-pass filtering (5–15 Hz;
Figura 2F). The result of the band-pass filtering metric was consistent with the STI metric,
because the rate-based model in response to tonic or oscillatory inputs generated regular
and consistent frequency. Por lo tanto, the band-pass filtering output and the original output
were consistent and power thresholding was the same (Figura 2F).
The simulated oscillatory activity was subthreshold (Figure 2B–E) and in the case of TC
neurons resembled minor rebound depolarizations that can be considered as an
indicator/marker of a burst (McCormick & Bal, 1997; McCormick & Pape, 1990). Esto es
because due to the nature of rate-based models, a burst cannot be directly generated and visu-
alized (there is no TC sequence of H-current and T-current built into rate-based models).
En cambio, a minor subthreshold rebound depolarization is an indicator of the tendency of the
network to generate a burst and the subthreshold oscillatory activity after the initial input is an
indicator of spindle oscillations. This is supported by previous physiological work that showed
the subthreshold pacemaker potential of TC neurons, demonstrating that a TC neuron burst
starts with a minor subthreshold depolarization with H-current activation, which slightly depo-
larizes the neuron (Deschenes, Paradis, roy, & Steriade, 1984; McCormick & Bal, 1997;
McCormick & Pape, 1990; Steriade & Deschenes, 1984). After this initial phase, the T-current
then takes over and pushes the membrane voltage from its minor initial depolarization further
up to cross the threshold leading to an action potential followed by a burst.
Además, as can be seen in Figure 2, the model has an oscillatory tendency with similar
general features, regardless of the input: (a) in response to a depolarizing input pulse to TC
neuronas (Figura 2C), where after PV depolarization, there is hyperpolarization and a rebound
depolarization, o (b) in response to a hyperpolarizing input on TC neurons (due to a brief
input pulse to TRN), where after PV hyperpolarization,
there is rebound depolarization
(Figura 2D). Use of a tonic depolarizing input to TC neurons instead of the initial oscillatory
input to TRN for simulations did not change the model dynamics and findings of the study:
injecting a tonic input of longer duration (0.5 segundo), as shown in Figure 2E, led to high oscilla-
tory activity and high spindle tendency.
Neurociencia en red
753
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
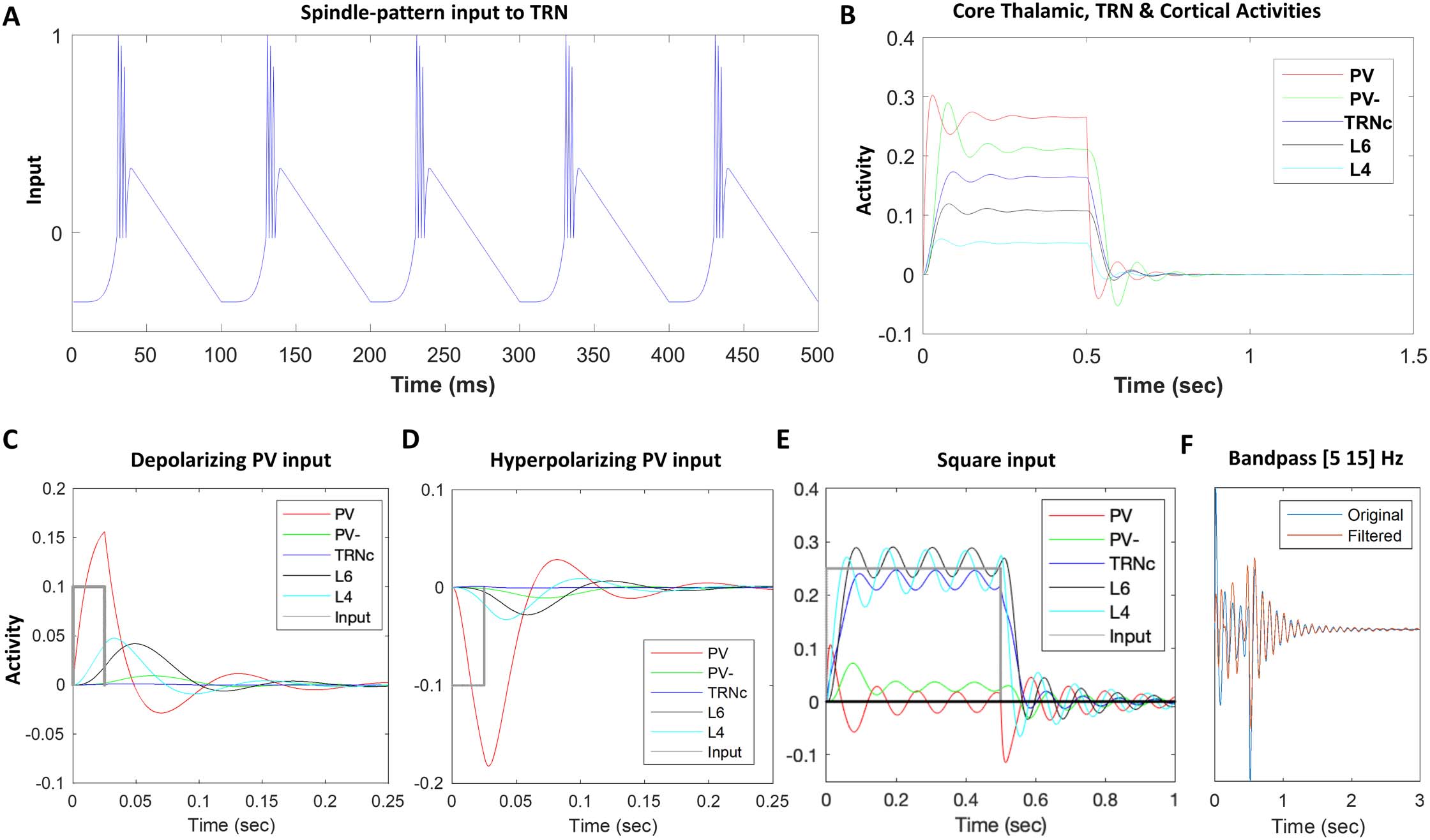
Impact of L5 to TRNM Connections
Projections from L5 to TRN have only recently been shown for some cortical areas and at
variable levels (p.ej., Prasad et al., 2020; Zikopoulos & Barbas, 2006), with L6 projections to
TRN being considered the norm, and several studies showing absence of direct L5 synaptic
terminals in TRN (p.ej., Bourassa & Deschenes, 1995). This prompted us to investigate the effects
of direct L5 to TRNM projection on spindle dynamics. Figure 3A shows the spindle dynamics in
matrix TC loop when the connection between model L5 and TRNM is absent. We considered
this condition the baseline. In Figure 3B and C, we turned on synaptic connections between L5
and TRNM at a level equivalent of 5–10% of L6-to-TRNC model connection strengths. Com-
pared to Figure 3A, the single-compartment voltage dynamics of matrix TC loop model neurons
in Figure 3B and C showed a temporal extension of hyperpolarization/depolarization beyond
0.5 segundo (when the spindle-like input to TRN was shut down) that gradually increased with
increased strength of L5 to TRNM input (from Figure 3B to 3C). This suggests that the presence
of direct input from L5 to TRN promotes temporal sustaining of spindles.
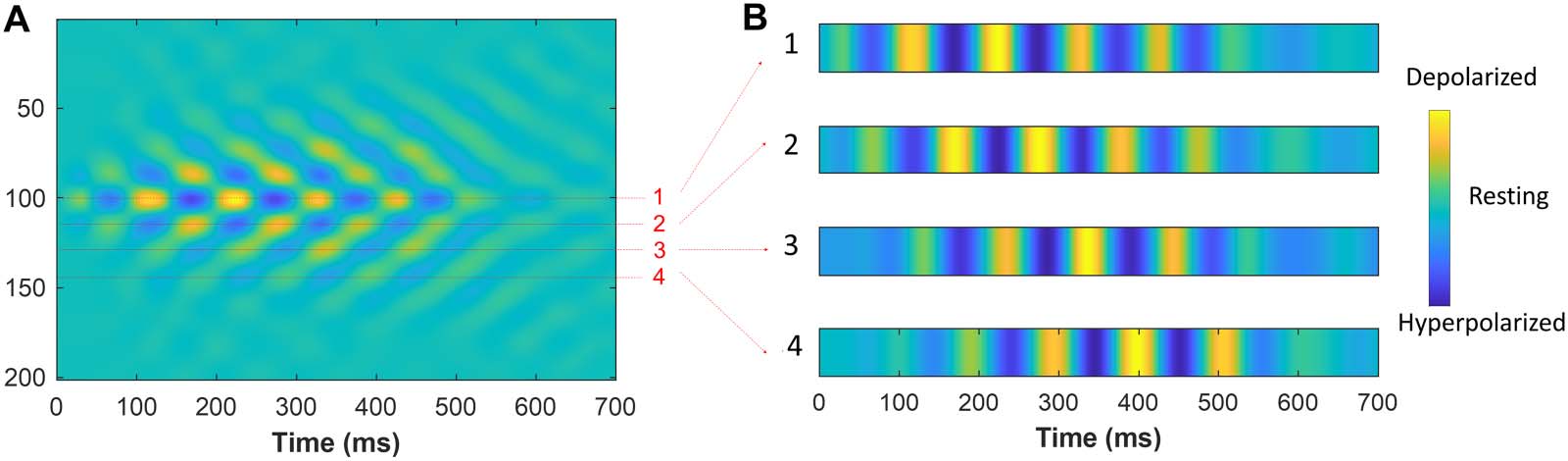
Spindle Wave Propagation
kim, Bal, and McCormick (1995) recorded simultaneously from multiple sites in the ferret dor-
solateral geniculate nucleus (LGNd) in vitro and found that spindle oscillations propagate
across the slice. The hyperpolarization and depolarization waves in Figure 4A show the same
trend; spindling tendency propagated across locations where model array neurons numbered
from 1–200 (y-axes) are distributed. Figure 4B shows the hyperpolarization-depolarization
dynamics of model TC neurons for four locations indicated in Figure 4A: first being center
(neuron no. 100), second location (neuron no. 113), third location (neuron no. 128), y
fourth location (neuron no. 147). Following first to fourth locations in order shows that the
initiation of spindle tendency propagated from the first to the fourth location. This provided
a mechanistic platform to further investigate the propagation dynamics based on the involve-
ment of different constituent parts of the TC loop. The example in Figure 4B is based on a
mixed core-matrix circuit with only core (PV+) neurons’ membrane potentials shown.
Cifra 3. The effect of model TRN input resembles spindle temporal pattern. The model is based on direct and indirect evidence for the
presence of L5 neuron projections to TRNM. (A) No direct L5 to TRNM results in STI < 0.1, which does not promote spindle tendency in the
network. (B) In comparison, implementation of partial, small L5 to TRNM connection (equivalent to 5% of L6 to TRNC model connection
strength) yields STI = 0.57, which shows a facilitating effect of direct (L5 to TRNM) connection in promoting spindle tendency of the network.
(C) the efficacy of L5-TRN connection is further increased from 5% to 10% resulting in STI > 1 that can contribute to the elongation of spindling
time and can facilitate continuous or repetitive spindles instead of isolated ones.
Neurociencia en red
754
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
Cifra 4. Spindle wave propagation across model neuron arrays. (A) The activity of 200 model TC neurons (PV+) con el tiempo. Yellow, depo-
larization; verde, resting potential; and blue, hyperpolarization. Model recording sites are indicated by 1, 2, 3, y 4 corresponding to neurons
100, 113, 128, y 147, respectivamente. (B) From top to bottom, four recording locations 1–4 from the model neurons are shown. Similar to the
Kim et al. (1995) recording simultaneously from multiple sites in the ferret LGNd, spindle oscillations propagate across locations 1–4 in the
modelo. Each subpanel of panel B is normalized within its range, and therefore the depolarization and hyperpolarization colors are identical,
merely showing whether the membrane potential is depolarized or hyperpolarized. Por lo tanto, the aim of panels 1–4 is to indicate the voltage
change direction either toward depolarization or hyperpolarization independent of the color map magnitude, which was adjusted to enhance
visibility.
Impact of Thalamic Local Inhibition on Spindle Dynamics
By tracing the behavior of the model TC neurons under different conditions, we evaluated the
tendency of the network to sustain or terminate spindles. Figure 5A shows the model core
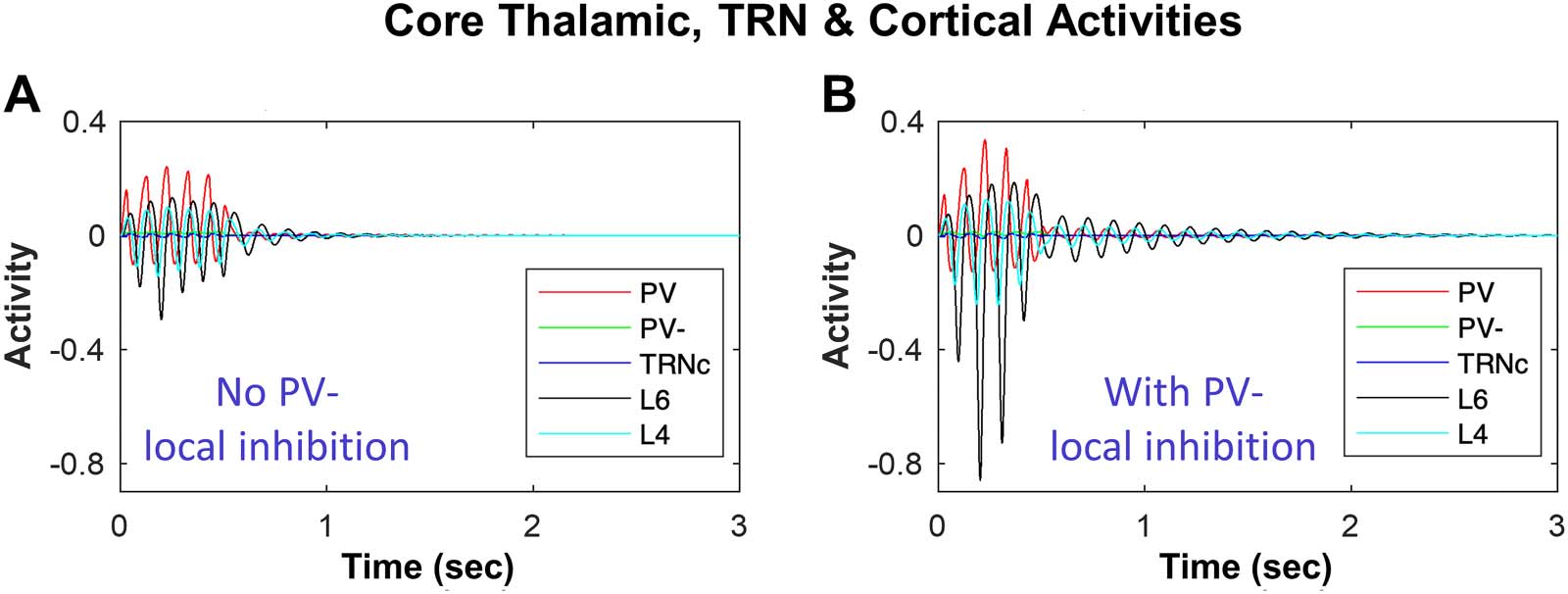
neuron activities when there is no local inhibition in the thalamus (no PV− Ò PV ).
Figure 5B shows the effects of local inhibition in the thalamus on the oscillatory TC activity.
The presence of local inhibition in thalamus indicates a higher spindle probability in the
model neurons of core TC loop, as seen in the comparison of the traces of activity of PV
and L4 neurons on and after 0.5 segundo, which lead to STI >1 in Figure 5B compared to STI =
0.55 in Figure 5A. Similarmente, local inhibition (CB−) in the matrix TC loop increased the spindle
probability when the L5-TRNM connection was present. With no L5-TRNM (STI < 0.1;
Figure 3A), presence or absence of local inhibition did not make a difference and STI remained
<0.1, indicating the necessity of L5-TRNM connection for the gating effect of local inhibition in
promoting spindling tendency.
Impact of local thalamic inhibition in biasing the network toward sustaining or terminating spindles. (A) shows the dynamics of core
Figure 5.
model neurons with the absence of local inhibition (no PV− inhibition of PV+), and (B) with the presence of local inhibition. The spindle-like
input (shown in Figure 2) is ON for 0.5 sec starting at time 0 and turned OFF at time 0.5 sec. By tracing the behavior of the model TC neurons,
we evaluate the tendency of the network to sustain or terminate spindles. Tracing the activity of PV and L4 neurons in panel A and B on and
after 0.5 sec indicates longer duration and higher amplitude of hyperpolarization and rebound polarization in panel B compared to A, indi-
cating a higher spindle probability in the model neurons of TC loop with the presence of local inhibition in thalamus.
Network Neuroscience
755
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
n
e
n
_
a
_
0
0
3
1
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
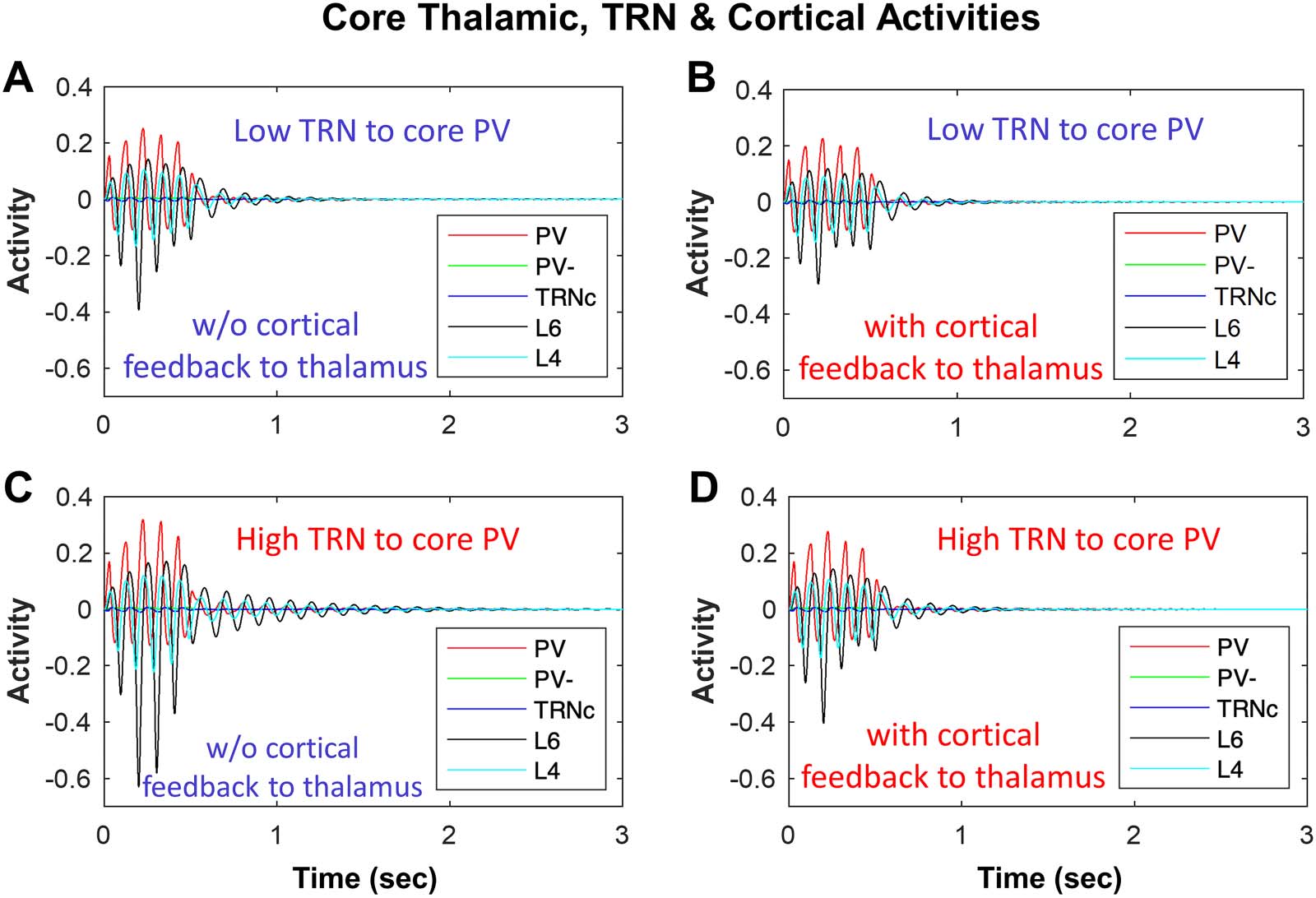
Impact of TRN Inhibition of Thalamus on Spindle Dynamics
Following the same approach, we tested the effect of different levels of TRN inhibition and cor-
tical feedback on the TC loop tendency to generate and sustain spindles (Figure 6A–D). Low
versus high levels of TRNc input to model excitatory core PV thalamic neurons (Figure 6A–C)
showed that during the initial spindle-like input to model TRNc (as shown in Figure 2) from
time 0–0.5 sec, the amplitude of hyperpolarization and rebound in core model neurons were
higher with increased TRNc inhibition of PV neurons. After the spindle-like input to model
TRNc was shut down (after 0.5 sec), the sequence of hyperpolarization and rebound events
due to model core TC loop sustained longer over time with higher TRNc inhibition of PV
neurons (Figure 6C with STI >1) compared to lower TRN inhibition (Figure 6A with STI =
0.55). A similar trend was present for matrix TC loops; higher TRNM input to excitatory
thalamic neurons (CB+) increased the STI.
Impact of Cortical Input to Thalamus on Spindle Dynamics
Corticothalamic feedback in the TC loop controls sleep spindle duration in vivo (Bonjean et al.,
2011). In line with this, cortical feedback in our model was a key player in the spindling time and
tendency to keep spindles unitary, repetitive, or sustained. In Figure 6A and C we illustrate an exam-
ple of the dependence of spindle dynamics on synaptic interactions within the thalamoreticular
loop, without any cortical feedback. Comparison of Figure 6C (STI > 1) with 6D (STI = 0.55) poder
provide an example of the impact of model cortex feedback to TC neurons. We found that the effect
of cortex was toward reducing the spindling time, and after the spindle-like input was shut down at
500 EM, there was less tendency for spindles, in the form of fewer hyperpolarization and rebound
depolarization events. Cortical feedback can be channeled in two forms: cortico-thalamic (mostrado
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Impact of TRN inhibition and cortical feedback to thalamus in biasing the network toward sustaining or terminating spindles. (A) El
Cifra 6.
dynamics of core model neurons without cortical feedback to thalamus (STI = 0.55) compared to (B) with cortical feedback to thalamus (STI <
0.55), indicating a decreased spindling tendency. (C) Similar to panel A, but with increased TRN inhibition of PV, which increased STI to >1
compared to panel A with STI = 0.55; (D) Similar to panel C, but with cortical feedback to thalamus, which reduced STI to 0.55 from >1 in C.
Neurociencia en red
756
Sleep spindles in core/matrix thalamocortical circuits
here in Figure 6, L6 Ò PV) and corticoreticular (not shown). Consistent with what Bonjean et al.
(2011) have shown (see their Figure 4), we also found that in our single-compartment voltage
rate-based model, more feedback from L6 to PV (cortico-thalamic) reduces the spindle duration
(Figure 6D; compare with 6C with no cortical feedback). Por otro lado, greater cortical feedback
to TRN in our model leads to greater input from TRN to thalamus and has the opposite effect, increas-
ing spindle tendency, in line with previous findings (Bonjean et al., 2011). The effects of the model
matrix cortical feedback to CB and TRNM were similar to the effects observed in the model core.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
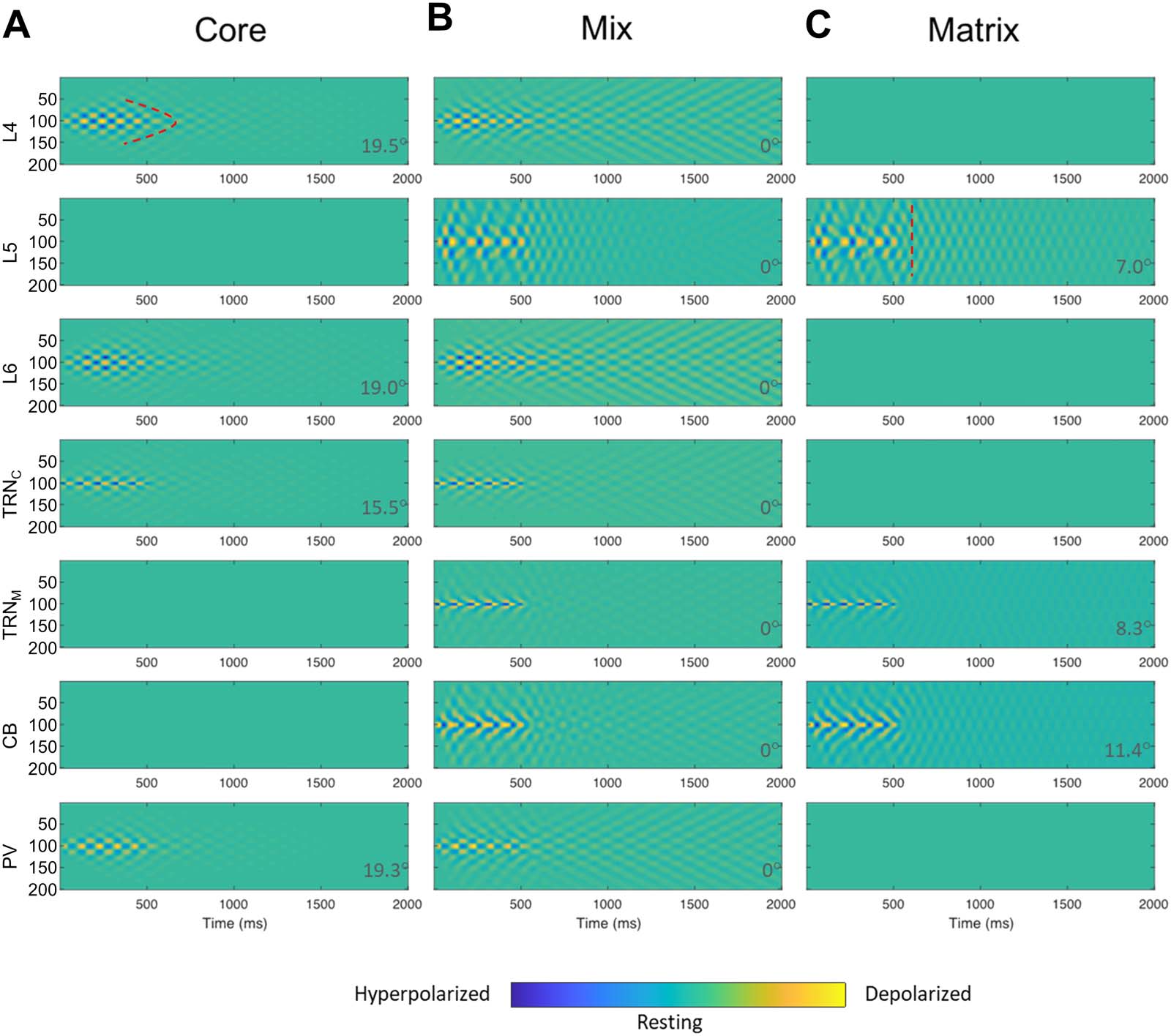
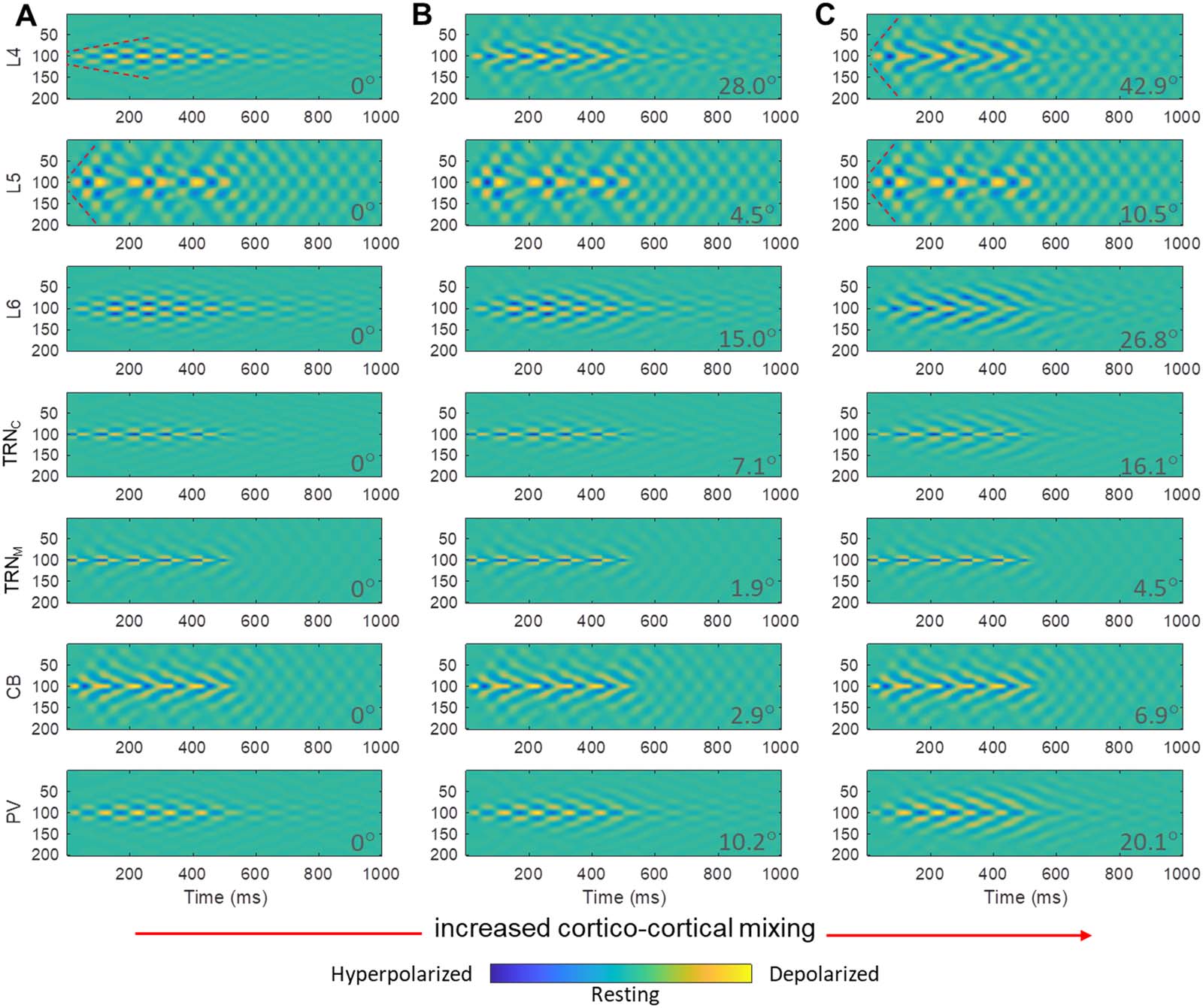
Cifra 7. Core, Matrix, and mixed TC model spatiotemporal dynamics. (A) Only model L6, TRNC, and PV (core TC loop) were involved in
spindle entrainment, induced by inputting spindle-like input (Cifra 2) to TRNC, while L5, TRNM, and CB (matrix) remained inactive. (B) El
mix (50–50% thalamoreticular mixing) of core and matrix, engaged all model areas. (C) Only model L5, TRNM, and CB (matrix TC loop) eran
involved in spindle entrainment, induced by inputting spindle-like input (Cifra 2) to TRNM, while L6, TRNC, and PV (core) remained inactive.
Spindle-like input (Cifra 2), was delivered to neuron no. 100 (y-axis) in the TRNC and TRNM array of model neurons, lasting for 500 ms and was
then shut down. The presence of color map ripples in the figure panels are equivalent to sequences of hyperpolarization and depolarization events
shown in previous figures and indicate spindle tendency of the network (más allá de 500 ms based on the TC model dynamics). Comparison of cor-
responding panels in A and C revealed a broader spatial spread and temporal extent in matrix than core. Notablemente, the mix circuit spatiotemporal
dynamics of spindles in each site was a hybrid of core and matrix spindles in terms of broadness of spatial smear and temporal extent. In panel A, el
model core cortical layer 4, L4, showed a focal tendency for spindle sustaining indicated by a curved dotted marking. Compare with panel C model
matrix cortical layer 5 (L5), in which the spindle tendency across model neurons after propagation appeared synchronized and therefore the
depolarization iso-amplitude traces aligned vertically at each time bin after spindle propagation for a few hundred milliseconds. A vertical dotted
mark as an example is shown in panel C, L5. To further compare the effects of mixing, we set 50–50% mixing level in panel B as the baseline (angle
i: ArcCos of cosine similarity = 0°). Comparison of θ values in panel B with pure core in A and pure matrix in C provides an estimate of the blending
efectos, expressed as rising deviation of the angle θ from 0°. Higher deviation of the angle θ from 0° indicated increased dissimilarity from B.
Neurociencia en red
757
Sleep spindles in core/matrix thalamocortical circuits
The Impact of Mixed Core and Matrix Circuits on Spindle Spatiotemporal Dynamics
Most TC loops in primate brains appear to include a mixture of core and matrix components
(jones, 2007; Müller et al., 2020; Piantoni et al., 2016; Zikopoulos & Barbas, 2007b). Para esto
reason, we compared the spatiotemporal dynamics of spindles in the mix circuit with those of
isolated core and matrix TC circuits (Cifra 7). There are multiple ways of mixing core and
matrix circuits, a saber, through thalamus, corteza, o ambos. In thalamic mixing (Cifra 7), nosotros
connected model PV and CB to TRNM and TRNC, respectivamente. In cortico-thalamic mixing (no
mostrado), we connected model L6 and L5 to TRNM and TRNC, respectivamente. Finalmente, nosotros también
included in the model laminar interactions occurring at the cortical level, which constitute
the most widely inferred, modeled, and studied mixing of core and matrix TC circuits. El
mixed spindle spatiotemporal dynamics turned out to be a hybrid of the core and matrix spin-
dles in all cases. These results highlight the impact of circuit connectivity on the core and
matrix spindle patterns (Piantoni et al., 2016), which blended seamlessly in mixed designs,
despite the constituent regional neural unit differences in core and matrix TC loops.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 8. Spatiotemporal dynamics of cortical mixed TC model with different ratios of involved core and matrix. (A–D) The spatiotemporal
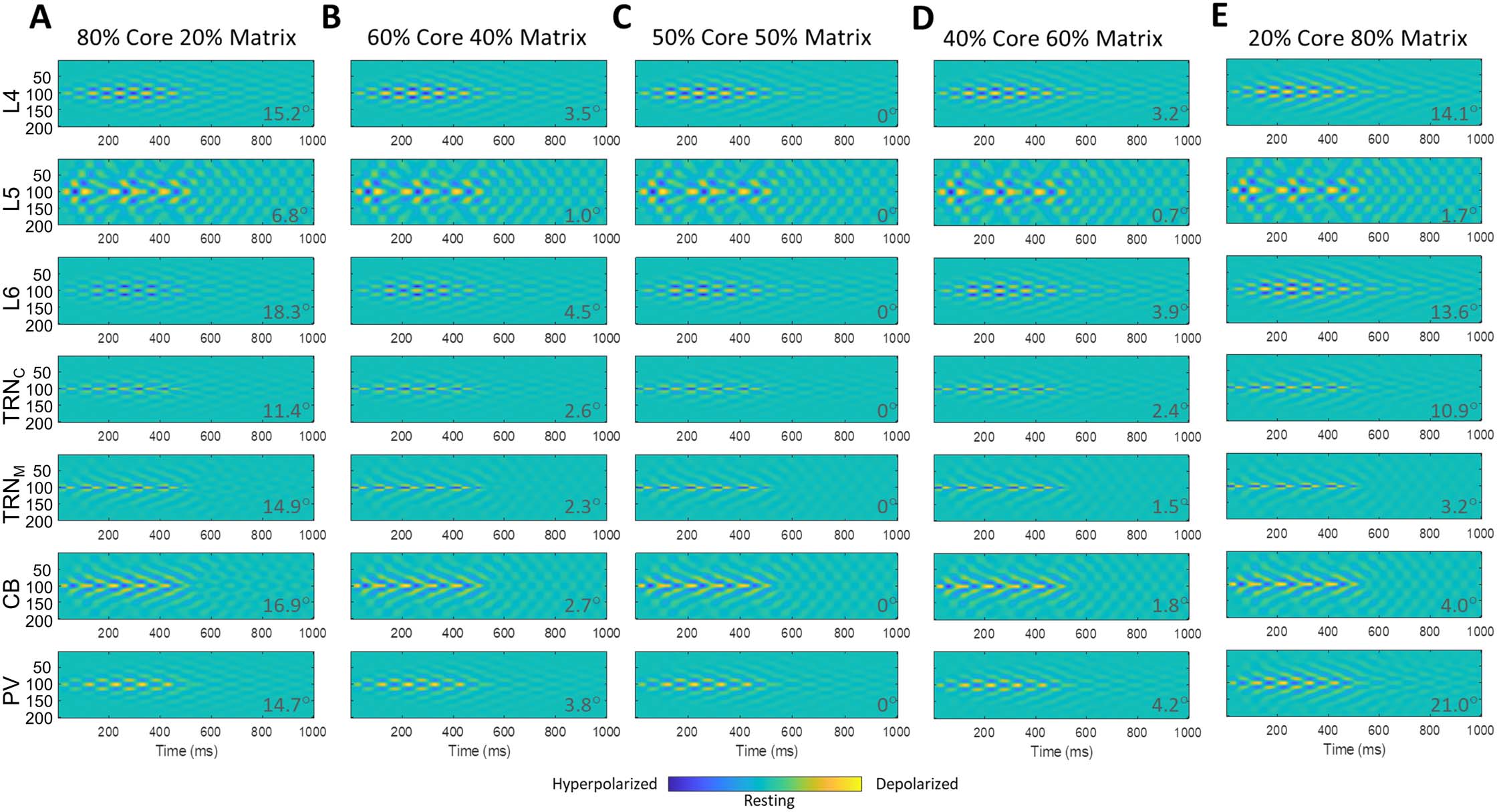
dynamics of TC neuronal activity with different ratios of mixed core/matrix: 80/20, 60/40, 40/60, y 20/80 are illustrated, with mixing occur-
ring at the level of the cortex. The higher the ratio of core/matrix, the more core-like were the spatiotemporal dynamics, eso es, focal in space
and less smeared over time. En cambio, the higher the ratio of matrix/core, the more matrix-like were the spatiotemporal dynamics, eso es,
more diffuse in space and smeared over time. This trend could be seen in all (in particular core) regiones (L4, L6, TRNC, and PV) involved in the
mix TC loop; the spatiotemporal dynamics of matrix specific regions (L5, TRNM, and CB), more or less, remained the same, eso es, diffuse and
smeared across different ratios of mixing, as if matrix kept its diffuse spatiotemporal dynamics due to its wider horizontal (within cortical
laminae) connectivity and depending on the ratio of core/matrix in the mix, the activity of core regions could be more or less matrix-like.
To further compare the effects of mixing, we set 50–50% mixing level in panel C as the baseline (angle θ: ArcCos of cosine similarity = 0°).
Comparison of θ values between panels provides an estimate of the magnitude of blending effects, expressed as rising deviation of the angle θ
from 0°. Higher deviation of the angle θ from 0° indicated increased dissimilarity from the baseline in panel C.
Neurociencia en red
758
Sleep spindles in core/matrix thalamocortical circuits
Cifra 8 shows a range of cortical core/matrix mixing ratios from 80% core/20% matrix to
20% core/80% matrix in five steps. The level of core and matrix involvement influenced the
spatiotemporal dynamics of spindling tendency in the network. Mixed TC loops were mostly
influenced by matrix toward a diffuse spatial and smeared over time spindling tendency
(hyperpolarizations and rebound depolarizations), highlighted by the increased deviation from
0° (50–50% mix) of the angle θ (ArcCos of cosine similarity), at each panel bottom right, cual
indicates increased dissimilarity. Neurons of the model matrix TC loops (p.ej., in L5 or CB)
influenced the spindling of core neurons (p.ej., in L4 or PV) in the mix, while retaining a rel-
atively unaffected diffuse and smeared in time pattern of spindling. The same relationship of
increased dissimilarity with increased deviation of the angle θ from 0° was observed in all
mixing scenarios examined (Cifra 7, Cifra 8, y figura 9).
Synaptic efficacy:
The weight or strength of a synaptic
connection quantified as a single
number, which can be adjusted to
increase or decrease.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 9. Spatiotemporal dynamics of TC model with different synaptic efficacy of cortical mixing of core and matrix. The spatiotemporal
dynamics of the TC model plateaued after reaching increased synaptic efficacy in cortical mixing. The plateau synaptic efficacy being used
era 30% (A), 60% (B), y 90% (C). Similar to Figure 8, matrix model neurons kept their spatiotemporal dynamics, and the response of core
model neurons resembled the response of matrix model neurons as the efficacy of mixing gradually increased. Model core cortical layer 4 (L4),
en 90% mixing level (C), compared to 30% mixing level (A), showed faster activity propagation across neurons indicated by higher slope (es decir.,
number of neurons per millisecond) of red dotted marks. This shows that mixing core with matrix impacts core signal propagation rate. Sobre el
other hand, matrix fast propagation rate remained relatively the same across different mixing levels of core and matrix, indicated by the same
slope of red dotted marks in panels A and C L5 panels. To further compare the effects of mixing, we set cortical mixing level in panel A as the
base (30% of the plateau of the synaptic efficacy), with an estimated angle θ (ArcCos of cosine similarity) = 0°. Comparison of θ values in
panel A with B and C provides an estimate of the cumulative blending effects of gradually increasing mixing levels in B and C, expressed as
rising deviation of the angle θ from 0°. Higher deviation of the angle θ from 0° indicated incremental increasing dissimilarity of B and C from A.
Neurociencia en red
759
Sleep spindles in core/matrix thalamocortical circuits
Thalamic or cortical mixing produced relatively similar results. The difference was the gran-
ularity of the impact of mixing. In cortical mixing, the effect was fine grained (Figures 8 y 9),
in which the graded increment of efficacy or ratio of mixing resulted in graded change in the
spatiotemporal dynamic of response. Por otro lado, in thalamic mixing, the initial incre-
ment of efficacy or ratio of mixing brought the spatiotemporal dynamic of mix to the
max/plateau level quickly, acting as a coarse tuning knob (Cifra 7).
DISCUSIÓN
We developed a circuit-based TC computational model, with distinct core and matrix loops,
based on detailed neuroanatomical organization and connectivity data of TC circuits in pri-
mates that can simulate sleep spindles. We additionally simulated mixed TC loops through
novel multilevel cortical, thalamoreticular, and corticoreticular mixing to investigate the func-
tional consequences of different ratios of core and matrix node connectivity contribution to
spindle dynamics. En tono rimbombante, our rate-based model, for the first time to our knowledge,
included and examined the role of local thalamic inhibitory interneurons, and direct layer 5
projections of variable density to TRN (L5-TRN) and thalamus. Simulations of our rate-based
model circuit showed the following: (a) increased local inhibition in the thalamus or (b)
increased TRN inhibition of core and matrix thalamic neurons could enhance spindle gener-
ation and sustain spindle activity for longer periods; (C) the nature of spindles in matrix was
more diffuse compared to core, with the mix type showing intermediate properties in agree-
ment with hypotheses that spindles can be classified in core-generated, matrix-generated or
mixed types, depending on the neuroanatomy of pathways involved in their generation
(Piantoni et al., 2016); (d) the L5-TRN projection enhanced spindle generation and propaga-
ción; (mi) spindle power could be modulated based on the level of cortical feedback and
involvement in model core versus matrix; y (F ) matrix TC spindles synchronized their spatial
propagation early on, whereas core TC spindles tended to remain spatially focal. In the mix
modelo, the activity of core neurons synchronized and inherited matrix synchronization.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Connections From Cortical L5 to TRN Are Necessary for Heterogeneity of Spindle Oscillations,
Especially for Matrix and Mix Spindle Generation
Projections from L5 to TRN have only recently been shown directly or indirectly for some cor-
tical areas and at variable levels (Hádinger et al., 2022; Prasad et al., 2020; Zikopoulos &
Barbas, 2006, 2007a). In primates, several thalamic nuclei, especially those connected with
prefrontal cortices, receive anywhere from 20 a 50% of their cortical projections from layer 5
pyramidal neurons (Xiao et al., 2009). These thalamic nuclei, which receive projections from
pyramidal neurons in cortical layers 5 y 6, participate in matrix TC loops, in addition to
typical core TC circuits (Zikopoulos & Barbas, 2007b). These robust projections from layer
5 terminate as axonal branches with a mix of small and large axon terminals that form synap-
ses with thalamic projection neurons and interact with ionotropic glutamate receptors
(Reichova & sherman, 2004; Zikopoulos & Barbas, 2007b). In primates, these types of axons
that contain large or a mix of large and small axon terminals are also found in TRN regions that
are connected with prefrontal cortices, account for about 38% of all prefrontal axons, y
most likely originate in cortical layer 5. These types of axon terminals are not seen in pathways
from sensory cortices to TRN in primates (Zikopoulos & Barbas, 2006).
Our simulations showed that the cortical projection to TRN is a critical circuit component
that determines the spatiotemporal dynamics of spindle propagation and underlies the
Neurociencia en red
760
Sleep spindles in core/matrix thalamocortical circuits
heterogeneity of spindles, observed in functional and physiological studies of the human brain
(Bonjean et al., 2011, 2012; Mak-McCully et al., 2014, 2017). The L5-TRN connection, en par-
particular, appears essential for the generation of matrix and consequently mix spindles. Con
regard to mix spindle dynamics, we examined, por primera vez, mixing of the core and matrix
connections at multiple levels. Our findings suggest that mixing, both at the thalamoreticular
nivel (CB+ and PV+ thalamic projection neurons sending their signals to TRNc and TRNm,
respectivamente), or at the corticoreticular level (L5 and L6 pyramidal projection neurons innervat-
ing TRNc and TRNm, respectivamente), can act as a coarse knob for spindle control. That is because
the spindle generator (TRN) is targeted in the mix, y, tal como, the spindle dynamics change
de modo significativo, even after low levels of mixing; hence, we have a narrow dynamical range for
mixing. En cambio, cortical mixing can act as a fine knob. This is because, mixing at a level
not directly involved in the generation of spindles (es decir., corteza), can be done with a broad
dynamical range (fine knob), because the cortex is entrained by the spindle generator level
(thalamoreticular), rather than being a generator itself. Cortical mixing is likely the prevalent
mode of mixing of TC circuits, because it relies on interlaminar connectivity that constitutes
the main component of cortical microcircuitry, eso es, projections from neurons in L3/4 to pyra-
midal neurons in L5, or core cortical nodes in L4 and L6 receiving L1-3 signals that are also
influenced by a wide kernel of matrix inputs (Krishnan et al., 2018; Thomson & lamy, 2007;
Yoshimura, Dantzker, & Callaway, 2005). To this effect, recent physiological studies have high-
lighted the laminar heterogeneity of human sleep spindles (Hagler et al., 2018; Krishnan et al.,
2018; Thomson & lamy, 2007; Yoshimura et al., 2005).
Local Thalamic Inhibition Increases Ability to Generate, Sustain, and Propagate Spindle Activity
Our simulations showed that increased local inhibition in thalamic nuclei, which can hyper-
polarize either core or matrix TC projection neurons, could enhance spindle generation and
sustain spindle activity for longer periods. En tono rimbombante, GABAergic local circuit neurons are
abundant in all the dorsal thalamic nuclei of primates, and may constitute about 40% de todo
neurons in the human thalamus (Arcelli et al., 1997; Hunt, Angustia, & jones, 1991; Herrero,
Seguela, & Padre, 1987), but are not present in all the thalamic nuclei of different mammalian
species, and are virtually absent in the thalamus of rodents, with the exception of few
first-order thalamic nuclei like the lateral geniculate nucleus (Arcelli et al., 1997; Barbaresi,
Spreafico, Frassoni, & Rustioni, 1986; Gabbott, Somogyi, Stewart, & Hamori, 1986) y el
ventral posterior nuclei (Simko & Markram, 2021). The importance of local thalamic inhibition
has not been explored in most experimental and computational studies of TC circuits or sleep
spindles, with few exceptions (Arcelli et al., 1997; Joyce et al., 2022; sherman, 2004; Timbie
et al., 2020). There is agreement that local inhibition, which is especially abundant in primates,
adds complexity to the synaptology and circuitry of the thalamus, in part by facilitating
widespread formation of triadic synaptic glomeruli in most thalamic nuclei (Arcelli et al.,
1997; Joyce et al., 2022; sherman, 2004; Timbie et al., 2020), and likely affects the functional
properties of TC circuits that can modulate attention, memory consolidation, and spindle
tendency.
En tono rimbombante, the ubiquitous presence of GABAergic local circuit neurons in the primate thala-
mus may provide alternative wiring avenues for the construction of open loop circuitry linking
TRN with the thalamus (Figura 1B; also see, p.ej., Cifra 9 of Barbas & Zikopoulos, 2007, y
Figures 5 y 9 of Zikopoulos & Barbas, 2007a). Several studies suggest that the open loop model
of TRN-thalamic interactions, in which TRN neurons are excited by a thalamic neuron and, Sucesivamente,
inhibit a different thalamic neuron or an inhibitory interneuron, can create a tunable filter that may
be modified by extra-TRN influences, like corticoreticular pathways (Brown y cols., 2020; willis
761
Triadic synaptic glomeruli:
Specialized synaptic arrangements
seen in the thalamus where an
extrathalamic excitatory terminal
innervates the dendrite of a thalamic
projection neuron and the dendrite of
a local inhibitory interneuron, cual
es, Sucesivamente, presynaptic to the same
thalamic projection neuron dendrite.
Note that dendrites of local thalamic
inhibitory interneurons contain
vesicles and can form inhibitory
synapses.
Neurociencia en red
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
Closed loop TRN-thalamic
conectividad:
In closed loop connections, TRN
neurons directly inhibit the thalamic
neurons that excite them. Gaussian
connectivity strength from TRN to
thalamus resembles bilateral and
symmetric open loops with opposite
open directions (balanced). Due to
spatially symmetric bilateral
Gaussian spread of connectivity
strength, the circuit is composed of
two open loops in opposite
directions (balanced) por lo tanto, nuestro
architecture is functionally similar to
a closed loop.
Hybrid loop TRN-thalamic
conectividad:
Bilateral and symmetric or
asymmetric Gaussian connectivity
design was used to simulate TRN-
Thalamic connectivity (closed or
open loops, respectivamente).
et al., 2015). Por otro lado, closed loop TRN-thalamic connectivity, in which TRN-thalamic
neuron pairs are reciprocally connected, allow a short window for initial excitation of a TC neuron
followed by inhibition, preventing summation and faithfully transmitting high-frequency signals to
the cortex (Steriade & Deschenes, 1984). Since both circuits are found in the primate thalamus, nosotros
modeled TRN-thalamic interactions using a novel hybrid loop design (Figura 1C) that can flexibly
simulate either open or closed loops. The hybrid loop design we implemented represents the
spread of TRN-thalamic connectivity strength using a bilateral Gaussian curve system. Our archi-
tecture resembles a closed loop functionally, when the bilateral Gaussian spread of connectivity
strength is spatially symmetric, which means that the resulting Gaussian peak of connectivity
strength is to the thalamic neuron which directly excites the TRN neuron. En otras palabras, a closed
loop in our model is composed of two open loops in opposite directions (balanced). En cambio,
when the connectivity spread is either spatially asymmetrical or follows a shifted Gaussian, entonces
our circuit architecture resembles an open loop. The hybrid loop TRN-thalamic connectivity
design used in this study can be a powerful tool to further explore open versus closed loop archi-
tecture and interactions in the thalamus and other systems in future studies.
Increased Cortical Influence on TRN Versus Thalamus Affects the Tendency to Generate Spindles in
Opposite Ways
Previous studies have shown that TRN neurons are tuned to be more responsive to cortical
than thalamic inputs, based on specialized synaptic-receptor interactions (Golshani, Liu, &
jones, 2001; Liu & jones, 1999). This suggests that changes in cortical input may affect differ-
entially TC versus TRN neurons. Our simulations show that increased ratio of core or matrix
TC neuron activation, due to increased cortical feedback to TC that is not accompanied by
equivalent increase of cortical feedback to TRN, can reduce a reverberating tendency to pro-
mote spindle duration and sustaining. A diferencia de, increased cortical feedback to TRN neurons
can reverse the balance, and increase the spindle tendency (not shown), consistent with find-
ings showing that increasing the corticoreticular conductance (es decir., gramo(PYR Ò TRN)) can result
in total spindle duration increase (Bonjean et al., 2011), and in line with the spindle generator
properties of TRN neurons. Sin embargo, recent studies have also suggested that depending on the
state of cortical and thalamic neuronal activity, high levels of cortical input to TRN may, en
some cases, disrupt thalamic spindling (Bonjean et al., 2011, 2012; Mak-McCully et al.,
2014, 2017), highlighting the complexity of the TC circuit interactions in primates.
Conclusions and Implications for Future Studies
Thalamocortical circuits are organized into diffuse matrix and spatially selective core compo-
nents that are mixed in different ratios throughout the brain (Piantoni et al., 2016). TC circuits
can indirectly propagate signals from one cortical area to another, in concert with or in addition
to direct corticocortical pathways, forming a framework for rhythmic brain activity, incluido
signal and spindle propagation across cortex (Brown y cols., 2020; en un abrazo & sherman, 2019;
jones, 2001; sherman, 2012; sherman & Guillery, 2013). Además, thalamocortical and cortical
connectivity influence sleep spindle properties (Krishnan et al., 2018). Por lo tanto, changes in
spindle dynamics can be considered a litmus test for typical thalamocortical circuit connectivity
and for disruptions underlying neuropathology, because the latter can also impact spindling ten-
dency. Por ejemplo, medicated individuals with chronic schizophrenia show deficits in spindle
density (Ferrarelli et al., 2007; Manoach et al., 2010), which parallel impaired memory consol-
idation during stage 2 sleep (Goder et al., 2015; Wamsley et al., 2012) and increased
thalamocortical functional connectivity (Baran et al., 2019; Manoach & Stickgold, 2019). Más-
encima, spindle density and duration are significantly decreased in autism spectrum disorders (ASD)
Neurociencia en red
762
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
and show negative correlation with sleep-dependent memory consolidation (Farmer et al., 2018;
Mylonas et al., 2022). Both disorders differentially affect sensory processing, atención, y
emotional processing likely involving at variable degrees first-order primary networks that participate
in core TC circuits, or high-order association networks that participate in matrix or mix TC circuits.
Por lo tanto, changes in spindle dynamics can also point to the imbalance of core and matrix involve-
ment in disorders such as schizophrenia and ASD, pinpointing specific TC loops, circuit nodes, cell
types, and underlying mechanisms that are likely disrupted. These disruptions are manifested
through distinct symptomatology, Por ejemplo, distractibility and difficulty focusing attention
is a hallmark of schizophrenia (Braff, 1993; Luck & Gold, 2008), whereas difficulty switching
attentional focus and sensory overresponsivity are often seen in ASD (allen & Courchesne,
2001; Cheung & Lau, 2020; Fan et al., 2012; Verde & Ben-Sasson, 2010; Keehn, Müller, &
Townsend, 2013; Marco, Hinkley, Colina, & Nagarajan, 2011). Future studies can use the modeling
framework we developed to further investigate how key circuit organization features in primates,
including thalamic inhibition, locally or through TRN, and extensive L5-TRN projections, poder
change spindle generation and their spatiotemporal propagation in health and in disease.
EXPRESIONES DE GRATITUD
We would like to thank Dr. Yohan John and Natalia Matuk for useful discussions and comments.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00311.
CONTRIBUCIONES DE AUTOR
Arash Yazdanbakhsh: Conceptualización; Curación de datos; Análisis formal; Adquisición de financiación;
Investigación; Metodología; Administración de proyecto; Recursos; Software; Supervisión; Valida-
ción; Visualización; Escritura – borrador original; Escritura – revisión & edición. Helen Barbas: Concep-
tualization; Investigación; Escritura – revisión & edición. Basilis Zikopoulos: Conceptualización;
Curación de datos; Análisis formal; Adquisición de financiación; Investigación; Metodología; Proyecto
administración; Recursos; Supervisión; Validación; Visualización; Escritura – borrador original;
Escritura – revisión & edición.
INFORMACIÓN DE FINANCIACIÓN
Basilis Zikopoulos, National Institute of Mental Health (https://dx.doi.org/10.13039
/100000025), Award ID: R01 MH 118500. Arash Yazdanbakhsh, National Institute of Mental
Salud (https://dx.doi.org/10.13039/100000025), Award ID: R01 MH 118500.
REFERENCIAS
Agmon, A., & Connors, B. W.. (1992). Correlation between intrinsic
firing patterns and thalamocortical synaptic responses of neurons
in mouse barrel cortex. Revista de neurociencia, 12(1), 319–329.
https://doi.org/10.1523/ JNEUROSCI.12-01-00319.1992,
PubMed: 1729440
allen, GRAMO., & Courchesne, mi. (2001). Attention function and dysfunc-
tion in autism. Frontiers in Bioscience, 6, D105–D119. https://doi
.org/10.2741/allen, PubMed: 11171544
Arcelli, PAG., Frassoni, C., Regondi, METRO. C., De Biasi, S., & Spreafico, R.
(1997). GABAergic neurons in mammalian thalamus: A marker
of thalamic complexity? Brain Research Bulletin, 42(1), 27–37.
https://doi.org/10.1016/S0361-9230(96)00107-4, PubMed:
8978932
Baran, B., Karahanoglu, F. I., Mylonas, D., Demanuele, C., Vangel,
METRO., Stickgold, r., … Manoach, D. S. (2019). Increased thalamocor-
tical connectivity in schizophrenia correlates with sleep spindle
deficits: Evidence for a common pathophysiology. Biológico
Psiquiatría: Cognitive Neuroscience and Neuroimaging, 4(8),
706–714. https://doi.org/10.1016/j.bpsc.2019.04.012, PubMed:
31262708
Neurociencia en red
763
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
Barbaresi, PAG., Spreafico, r., Frassoni, C., & Rustioni, A. (1986).
GABAergic neurons are present in the dorsal column nuclei
but not in the ventroposterior complex of rats. Brain Research,
382(2), 305–326. https://doi.org/10.1016/0006-8993(86)91340-5,
PubMed: 2428443
Barbas, h., & Zikopoulos, B. (2007). The prefrontal cortex and flex-
ible behavior. neurocientífico, 13(5), 532–545. https://doi.org/10
.1177/1073858407301369, PubMed: 17901261
Bazhenov, METRO., Timofeev, I., Steriade, METRO., & Sejnowski, t. j. (1999).
Self-sustained rhythmic activity in the thalamic reticular nucleus
mediated by depolarizing GABAA receptor potentials. Naturaleza
Neurociencia, 2(2), 168–174. https://doi.org/10.1038/5729,
PubMed: 10195202
Bonjean, METRO., Panadero, T., Bazhenov, METRO., Cash, S., Halgren, MI., &
Sejnowski, t. (2012). Interactions between core and matrix
thalamocortical projections in human sleep spindle synchroniza-
ción. Revista de neurociencia, 32(15), 5250–5263. https://doi.org
/10.1523/JNEUROSCI.6141-11.2012, PubMed: 22496571
Bonjean, METRO., Panadero, T., Lemieux, METRO., Timofeev, I., Sejnowski, T., &
Bazhenov, METRO. (2011). Corticothalamic feedback controls sleep
spindle duration in vivo. Revista de neurociencia, 31(25),
9124–9134. https://doi.org/10.1523/JNEUROSCI.0077-11.2011,
PubMed: 21697364
Bourassa, J., & Deschenes, METRO. (1995). Corticothalamic projections
from the primary visual cortex in rats: A single fiber study using
biocytin as an anterograde tracer. Neurociencia, 66(2), 253–263.
https://doi.org/10.1016/0306-4522(95)00009-8, PubMed:
7477870
Bourassa, J., Pinault, D., & Deschenes, METRO. (1995). Corticothalamic
projections from the cortical barrel field to the somatosensory
thalamus in rats: A single-fibre study using biocytin as an anter-
ograde tracer. European Journal of Neuroscience, 7(1), 19–30.
https://doi.org/10.1111/j.1460-9568.1995.tb01016.x, PubMed:
7711933
Braff, D. l. (1993). Information processing and attention dysfunc-
tions in schizophrenia. Schizophrenia Bulletin, 19(2), 233–259.
https://doi.org/10.1093/schbul/19.2.233, PubMed: 8322034
Marrón, j. w., Taheri, A., Kenyon, R. v., Berger-Wolf, t. y., & Llano,
D. A. (2020). Signal propagation via open-loop intrathalamic
architectures: A computational model. eNeuro, 7(1). https://doi
.org/10.1523/ENEURO.0441-19.2020, PubMed: 32005750
Cao, B., Mingolla, MI., & Yazdanbakhsh, A. (2015). Tuning proper-
ties of MT and MSTd and divisive interactions for eye-movement
compensación. Más uno, 10(11), e0142964. https://doi.org/10
.1371/diario.pone.0142964, PubMed: 26575648
Chagnac-Amitai, y., Luhmann, h. J., & Príncipe, D. A. (1990). Burst
generating and regular spiking layer 5 pyramidal neurons of rat
neocortex have different morphological features. Journal of Com-
parative Neurology, 296(4), 598–613. https://doi.org/10.1002
/cne.902960407, PubMed: 2358553
Cheung, PAG. PAG. PAG., & Lau, B. W.. METRO. (2020). Neurobiology of sensory
processing in autism spectrum disorder. Progress in Molecular
Biology and Translational Science, 173, 161–181. https://doi
.org/10.1016/bs.pmbts.2020.04.020, PubMed: 32711809
Clasca, F., Rubio-Garrido, PAG., & Jabaudon, D. (2012). Unveiling the
diversity of thalamocortical neuron subtypes. European Journal of
Neurociencia, 35(10), 1524–1532. https://doi.org/10.1111/j
.1460-9568.2012.08033.X, PubMed: 22606998
Crabtree, j. W.. (1998). Organization in the auditory sector of the catʼs
thalamic reticular nucleus. Journal of Comparative Neurology,
390(2), 167–182. https://doi.org/10.1002/(CIENCIA )1096-9861
(19980112)390:2<167::AID-CNE1>3.0.CO;2-#, PubMed: 9453662
Crabtree, j. w., Collingridge, GRAMO. l., & Isaac, j. t. R. (1998). A new
intrathalamic pathway linking modality-related nuclei in the dor-
sal thalamus. Neurociencia de la naturaleza, 1(5), 389–394. https://doi.org
/10.1038/1603, PubMed: 10196529
Crabtree, j. w., & Isaac, j. t. (2002). New intrathalamic pathways
allowing modality-related and cross-modality switching in the
dorsal thalamus. Revista de neurociencia, 22(19), 8754–8761.
https://doi.org/10.1523/ JNEUROSCI.22-19-08754.2002,
PubMed: 12351751
de Kock, C. PAG. J., Pie, J., Pieneman, A. w., Mease, R. A., Bast, A., Guest,
j. METRO., … Sakmann, B. (2021). High-frequency burst spiking in layer 5
thick-tufted pyramids of rat primary somatosensory cortex encodes
exploratory touch. Communications Biology, 4(1), 709. https://doi
.org/10.1038/s42003-021-02241-8, PubMed: 34112934
Deschenes, METRO., Paradis, METRO., roy, j. PAG., & Steriade, METRO. (1984). Elec-
trophysiology of neurons of lateral thalamic nuclei in cat: descansando
properties and burst discharges. Revista de neurofisiología,
51(6), 1196–1219. https://doi.org/10.1152/jn.1984.51.6.1196,
PubMed: 6737028
Admirador, J., Bernardi, S., Dam, norte. T., Anagnostou, MI., Gu, X., Martín, l.,
… Hof, PAG. R. (2012). Functional deficits of the attentional net-
works in autism. Brain and Behavior, 2(5), 647–660. https://doi
.org/10.1002/brb3.90, PubMed: 23139910
Farmer, C. A., Chilakamarri, PAG., Torre, A. MI., Swedo, S. MI., holmes,
GRAMO. l., & Buckley, A. W.. (2018). Spindle activity in young children
with autism, developmental delay, or typical development.
Neurología, 91(2), e112–e122. https://doi.org/10.1212/ WNL
.0000000000005759, PubMed: 29875224
Ferrarelli, F., Huber, r., Peterson, METRO. J., Massimini, METRO., Murphy, METRO.,
Riedner, B. A., … Tononi, GRAMO. (2007). Reduced sleep spindle
activity in schizophrenia patients. American Journal of Psychia-
intentar, 164(3), 483–492. https://doi.org/10.1176/ajp.2007.164.3
.483, PubMed: 17329474
FitzGibbon, T., Solomon, S. GRAMO., & Goodchild, A. k. (2000). Distri-
bution of calbindin, parvalbumin, and calretinin immunoreacti-
vity in the reticular thalamic nucleus of the marmoset: Evidencia
for a medial leaflet of incertal neurons. Experimental Neurology,
164(2), 371–383. https://doi.org/10.1006/exnr.2000.7436,
PubMed: 10915576
Gabbott, PAG. l., Somogyi, J., Stewart, METRO. GRAMO., & Hamori, j. (1986). A
quantitative investigation of the neuronal composition of the rat dor-
sal lateral geniculate nucleus using GABA-immunocytochemistry.
Neurociencia, 19(1), 101–111. https://doi.org/10.1016/0306
-4522(86)90008-4, PubMed: 3537838
Gentet, l. J., & Ulrich, D. (2003). Strong, reliable and precise syn-
aptic connections between thalamic relay cells and neurones of
the nucleus reticularis in juvenile rats. Journal of Physiololgy,
546(punto 3), 801–811. https://doi.org/10.1113/jphysiol.2002
.032730, PubMed: 12563005
Goder, r., Graf, A., Ballhausen, F., Weinhold, S., Baier, PAG. C.,
Junghanns, K., & Prehn-Kristensen, A. (2015). Impairment of
sleep-related memory consolidation in schizophrenia: Relevance
of sleep spindles? Sleep Medicine, 16(5), 564–569. https://doi.org
/10.1016/j.sleep.2014.12.022, PubMed: 25912599
Neurociencia en red
764
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
Golshani, PAG., Liu, X. B., & jones, mi. GRAMO. (2001). Differences in quantal
amplitude reflect GluR4- subunit number at corticothalamic syn-
apses on two populations of thalamic neurons. Actas de
the National Academy of Sciences of the United States of Amer-
ica, 98(7), 4172–4177. https://doi.org/10.1073/pnas.061013698,
PubMed: 11274440
Verde, S. A., & Ben-Sasson, A. (2010). Anxiety disorders and
sensory over-responsivity in children with autism spectrum
disorders: Is there a causal relationship? Journal of Autism and
Developmental Disorders, 40(12), 1495–1504. https://doi.org
/10.1007/s10803-010-1007-x, PubMed: 20383658
Greenhill, S. D., Ranson, A., & Fox, k. (2015). Hebbian and
homeostatic plasticity mechanisms in regular spiking and intrinsic
bursting cells of cortical layer 5. Neurona, 88(3), 539–552. https://
doi.org/10.1016/j.neuron.2015.09.025, PubMed: 26481037
Guillery, R. W.. (1995). Anatomical evidence concerning the role of
the thalamus in corticocortical communication: A brief review.
Journal of Anatomy, 187(punto 3), 583–592. PubMed: 8586557
Guillery, R. W.. (2005). Anatomical pathways that link perception
and action. Progress in Brain Research, 149, 235–256. https://
doi.org/10.1016/S0079-6123(05)49017-2, PubMed: 16226588
Guillery, R. w., Feig, S. l., & Lozsádi, D. A. (1998). Paying attention
to the thalamic reticular nucleus. Trends in Neuroscience, 21(1),
28–32. https://doi.org/10.1016/S0166-2236(97 )01157-0,
PubMed: 9464683
Guillery, R. w., & Harting, j. k. (2003). Structure and connections
of the thalamic reticular nucleus: Advancing views over half a
siglo. Journal of Comparative Neurology, 463(4), 360–371.
https://doi.org/10.1002/cne.10738, PubMed: 12836172
Guillery, R. w., & sherman, S. METRO. (2002). Thalamic relay functions
and their role in corticocortical communication: Generalizations
from the visual system. Neurona, 33(2), 163–175. https://doi.org
/10.1016/S0896-6273(01)00582-7, PubMed: 11804565
Hádinger, NORTE., Bősz, MI., Tóth, B., Vantomme, GRAMO., Lüthi, A., & Acsády, l.
(2022). Region selective cortical control of the thalamic reticular
núcleo. bioRxiv. https://doi.org/10.1101/2022.01.17.476335
Hádinger, NORTE., Bősz, MI., Vantomme, GRAMO., Tóth, B., Lüthi, A., & Acsády,
l. (2019). Specialized frontal cortical control over the anterior
thalamic reticular nucleus [Paper presentation]. The Society for
Neurociencia, chicago, IL.
Hagler, D. J., Jr., Ulbert, I., Wittner, l., Eross, l., Madsen, j. r.,
Devinsky, o., … Halgren, mi. (2018). Heterogeneous origins of
human sleep spindles in different cortical layers. Diario de
Neurociencia, 38(12), 3013–3025. https://doi.org/10.1523
/JNEUROSCI.2241-17.2018, PubMed: 29449429
en un abrazo, METRO. METRO., & sherman, S. METRO. (2019). Thalamocortical circuit
motifs: A general framework. Neurona, 103(5), 762–770. https://
doi.org/10.1016/j.neuron.2019.06.005, PubMed: 31487527
Hale, PAG. T., Sefton, A. J., Baur, l. A., & Cottee, l. j. (1982). Interre-
lations of the ratʼs thalamic reticular and dorsal lateral geniculate
nuclei. Experimental Brain Research, 45(1–2), 217–229. https://
doi.org/10.1007/BF00235781, PubMed: 6173248
harris, k. D., & Shepherd, GRAMO. METRO. (2015). The neocortical circuit:
Themes and variations. Neurociencia de la naturaleza, 18(2), 170–181.
https://doi.org/10.1038/nn.3917, PubMed: 25622573
Hoogland, PAG. v., Wouterlood, F. GRAMO., Welker, MI., & Van der Loos, h.
(1991). Ultrastructure of giant and small thalamic terminals of
cortical origin: A study of the projections from the barrel cortex
in mice using Phaseolus vulgaris leuco-agglutinin (PHA-L).
Experimental Brain Research, 87(1), 159–172. https://doi.org/10
.1007/BF00228517, PubMed: 1721878
Hunt, C. A., Angustia, D. Z., & jones, mi. GRAMO. (1991). Distribution and
density of GABA cells in intralaminar and adjacent nuclei of
monkey thalamus. Neurociencia, 43(1), 185–196. https://doi
.org/10.1016/0306-4522(91)90426-oh, PubMed: 1922765
John, Y. J., Bullock, D., Zikopoulos, B., & Barbas, h. (2013).
Anatomy and computational modeling of networks underlying
cognitive-emotional interaction. Frontiers in Human Neurosci-
ence, 7, 101. https://doi.org/10.3389/fnhum.2013.00101,
PubMed: 23565082
John, Y. J., Zikopoulos, B., Bullock, D., & Barbas, h. (2016). El
emotional gatekeeper: A computational model of attentional
selection and supression through the pathway from the amygdala
to the thalamic reticular nucleus. Biología Computacional PLoS,
12(2), e1004722. https://doi.org/10.1371/journal.pcbi.1004722,
PubMed: 26828203
jones, mi. GRAMO. (1998). Viewpoint: The core and matrix of thalamic
organización. Neurociencia, 85(2), 331–345. https://doi.org/10
.1016/S0306-4522(97)00581-2, PubMed: 9622234
jones, mi. GRAMO. (2001). The thalamic matrix and thalamocortical syn-
chrony. Trends in Neuroscience, 24(10), 595–601. https://doi.org
/10.1016/S0166-2236(00)01922-6, PubMed: 11576674
jones, mi. GRAMO. (2007). The thalamus. Nueva York, Nueva York: Cambridge Uni-
versity Press.
Joyce, METRO. k. PAG., marshall, l. GRAMO., Banik, S. l., Wang, J., xiao, D.,
Bunce, j. GRAMO., & Barbas, h. (2022). Pathways for memory, saber-
tion and emotional context: Hippocampal, subgenual area 25,
and amygdalar axons show unique interactions in the primate
thalamic reuniens nucleus. Revista de neurociencia, 42(6),
1068–1089. https://doi.org/10.1523/JNEUROSCI.1724-21.2021,
PubMed: 34903572
Kakei, S., Na, J., & Shinoda, Y. (2001). Thalamic terminal morphol-
ogy and distribution of single corticothalamic axons originating
from layers 5 y 6 of the cat motor cortex. Journal of Compar-
ative Neurology, 437(2), 170–185. https://doi.org/10.1002/cne
.1277, PubMed: 11494250
Keehn, B., Müller, R.-A., & Townsend, j. (2013). Atypical atten-
tional networks and the emergence of autism. Neurociencia
and Biobehavioral Reviews, 37(2), 164–183. https://doi.org/10
.1016/j.neubiorev.2012.11.014, PubMed: 23206665
kim, Ud., Bal, T., & McCormick, D. A. (1995). Spindle waves are
propagating synchronized oscillations in the ferret LGNd in vitro.
Revista de neurofisiología, 74(3), 1301–1323. https://doi.org/10
.1152/jn.1995.74.3.1301, PubMed: 7500152
Kimura, A. (2014). Diverse subthreshold cross-modal sensory inter-
actions in the thalamic reticular nucleus: Implications for new
pathways of cross-modal attentional gating function. European
Revista de neurociencia, 39(9), 1405–1418. https://doi.org/10
.1111/ejn.12545, PubMed: 24646412
Krishnan, GRAMO. PAG., rosa, B. P., Chen, J.-Y., Muller, l., Sejnowski,
t. J., Cash, S. S., … Bazhenov, METRO. (2018). Thalamocortical and
intracortical laminar connectivity determines sleep spindle prop-
erties. Biología Computacional PLoS, 14(6), e1006171. https://doi
.org/10.1371/journal.pcbi.1006171, PubMed: 29949575
Kultas-Ilinsky, K., Hacer, h., & Ilinsky, I. A. (1995). Nucleus reticularis
thalami input to the anterior thalamic nuclei in the monkey: A
Neurociencia en red
765
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
light and electron microscopic study. Neuroscience Letters,
186(1), 25–28. https://doi.org/10.1016/0304-3940(95)11273-Y,
PubMed: 7540266
Justicia, Y. w., & sherman, S. METRO. (2005). Mapping by laser photosti-
mulation of connections between the thalamic reticular and ven-
tral posterior lateral nuclei in the rat. Revista de neurofisiología,
94(4), 2472–2483. https://doi.org/10.1152/jn.00206.2005,
PubMed: 16160090
Larkum, METRO. MI., Zhu, j. J., & Sakmann, B. (1999). A new cellular
mechanism for coupling inputs arriving at different cortical
capas. Naturaleza, 398(6725), 338–341. https://doi.org/10.1038
/18686, PubMed: 10192334
Layton, oh. w., Mingolla, MI., & Yazdanbakhsh, A. (2012). Dynamic
coding of border-ownership in visual cortex. Revista de visión,
12(13), 8. https://doi.org/10.1167/12.13.8, PubMed: 23220579
Layton, oh. w., Mingolla, MI., & Yazdanbakhsh, A. (2014). Neural
dynamics of
feedforward and feedback processing in
figure-ground segregation. Fronteras en psicología, 5, 972.
https://doi.org/10.3389/fpsyg.2014.00972, PubMed: 25346703
Layton, oh. w., & Yazdanbakhsh, A. (2015). A neural model of
border-ownership from kinetic occlusion. Vision Research, 106,
64–80. https://doi.org/10.1016/j.visres.2014.11.002, PubMed:
25448117
Sotavento, S.-C., Cruikshank, S. J., & Connors, B. W.. (2010). Electrical and
chemical synapses between relay neurons in developing thala-
mus. Revista de fisiología, 588(punto 13), 2403–2415. https://doi
.org/10.1113/jphysiol.2010.187096, PubMed: 20457735
Liu, X. B., & jones, mi. GRAMO. (1999). Predominance of corticothalamic
synaptic inputs to thalamic reticular nucleus neurons in the rat.
Journal of Comparative Neurology, 414(1), 67–79. https://doi.org
/10.1002/(CIENCIA)1096-9861(19991108)414:1<67::AID-CNE6>3.0
.CO;2-z, PubMed: 10494079
Livingstone, METRO. S., Hombre libre, D. C., & Hubel, D. h. (1996). Visual
responses in V1 of freely viewing monkeys. Cold Spring Harbor
Symposia on Quantitative Biology, 61, 27–37. https://doi.org/10
.1101/SQB.1996.061.01.006, PubMed: 9246432
Llano, D. A., & sherman, S. METRO. (2009). Differences in intrinsic prop-
erties and local network connectivity of identified layer 5 y
capa 6 adult mouse auditory corticothalamic neurons support
a dual corticothalamic projection hypothesis. Corteza cerebral,
19(12), 2810–2826. https://doi.org/10.1093/cercor/ bhp050,
PubMed: 19351905
Lo, F. S., & sherman, S. METRO. (1994). Feedback inhibition in the catʼs
lateral geniculate nucleus. Experimental Brain Research, 100(2),
365–368. https://doi.org/10.1007/ BF00227207, PubMed:
7813674
Luck, S. J., & Gold, j. METRO. (2008). The construct of attention in
schizophrenia. Biological Psychiatry, 64(1), 34–39. https://doi
.org/10.1016/j.biopsych.2008.02.014, PubMed: 18374901
Lüthi, A. (2014). Sleep spindles: Where they come from, what they
hacer. neurocientífico, 20(3), 243–256. https://doi.org/10.1177
/1073858413500854, PubMed: 23981852
Mak-McCully, R. A., Deiss, S. r., rosa, B. P., Jung, K.-Y.,
Sejnowski, t. J., Bastuji, h., … Halgren, mi. (2014). Synchroniza-
tion of isolated downstates (K-complexes) may be caused by
cortically-induced disruption of thalamic spindling. PLoS Com-
putational Biology, 10(9), e1003855. https://doi.org/10.1371
/journal.pcbi.1003855, PubMed: 25255217
Mak-McCully, R. A., Rolland, METRO., Sargsyan, A., González, C.,
Magnin, METRO., Chauvel, PAG., … Halgren, mi. (2017). Coordination of
cortical and thalamic activity during non-REM sleep in humans.
Comunicaciones de la naturaleza, 8, 15499. https://doi.org/10.1038
/ncomms15499, PubMed: 28541306
Manoach, D. S., & Stickgold, R. (2019). Abnormal sleep spindles,
memory consolidation, and schizophrenia. Annual Review of
Clinical Psychology, 15, 451–479. https://doi.org/10.1146
/annurev-clinpsy-050718-095754, PubMed: 30786245
Manoach, D. S., Thakkar, k. NORTE., Stroynowski, MI., Ely, A., McKinley,
S. K., Wamsley, MI., … Stickgold, R. (2010). Reduced overnight
consolidation of procedural learning in chronic medicated
schizophrenia is related to specific sleep stages. Diario de
Psychiatric Research, 44(2), 112–120. https://doi.org/10.1016/j
.jpsychires.2009.06.011, PubMed: 19665729
Marco, mi. J., Hinkley, l. B. NORTE., Colina, S. S., & Nagarajan, S. S. (2011).
Sensory processing in autism: A review of neurophysiologic
findings. Pediatric Research, 69(5 punto 2), 48–54. https://doi.org
/10.1203/PDR.0b013e3182130c54, PubMed: 21289533
McAlonan, K., Cavanaugh, J., & Wurtz, R. h. (2006). Attentional
modulation of thalamic reticular neurons. Journal of Neurosci-
ence, 26(16), 4444–4450. https://doi.org/10.1523/JNEUROSCI
.5602-05.2006, PubMed: 16624964
McCormick, D. A., & Bal, t. (1997). Sleep and arousal: Thalamo-
cortical mechanisms. Revisión anual de neurociencia, 20,
185–215. https://doi.org/10.1146/annurev.neuro.20.1.185,
PubMed: 9056712
McCormick, D. A., & Pape, h. C. (1990). Properties of a
hyperpolarization-activated cation current and its role in rhyth-
mic oscillation in thalamic relay neurones. Revista de fisiología,
431, 291–318. https://doi.org/10.1113/jphysiol.1990.sp018331,
PubMed: 1712843
Medalla, METRO., & Barbas, h. (2009). Synapses with inhibitory neurons
differentiate anterior cingulate from dorsolateral prefrontal path-
ways associated with cognitive control. Neurona, 61(4), 609–620.
https://doi.org/10.1016/j.neuron.2009.01.006, PubMed: 19249280
Müller, mi. J., Munn, B., Hearne, l. J., Herrero, j. B., Fulcher, B.,
Arnatkeviciute, A., … Shine, j. METRO. (2020). Core and matrix
thalamic sub-populations relate to spatio-temporal cortical con-
nectivity gradients. NeuroImagen, 222, 117224. https://doi.org/10
.1016/j.neuroimage.2020.117224, PubMed: 32795658.
Münkle, METRO. C., Waldvogel, h. J., & Faull, R. l. (2000). The distri-
bution of calbindin, calretinin and parvalbumin immunoreacti-
vity in the human thalamus. Journal of Chemical Neuroanatomy,
19(3), 155–173. https://doi.org/10.1016/S0891-0618(00)00060
-0, PubMed: 10989260
Mylonas, D., Machado, S., larson, o., patel, r., Cox, r., Vangel,
METRO., … Manoach, D. S. (2022). Dyscoordination of non-rapid
eye movement sleep oscillations in autism spectrum disorder.
Sleep, 45(3), zsac010. https://doi.org/10.1093/sleep/zsac010,
PubMed: 35022792
Piantoni, GRAMO., Halgren, MI., & Cash, S. S. (2016). The contribution of
thalamocortical core and matrix pathways to sleep spindles.
Neural Plasticity, 2016, 3024342. https://doi.org/10.1155/2016
/3024342, PubMed: 27144033
Pinault, D. (2004). The thalamic reticular nucleus: Estructura, func-
tion and concept. Brain Research Reviews, 46(1), 1–31. https://
doi.org/10.1016/j.brainresrev.2004.04.008, PubMed: 15297152
Neurociencia en red
766
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
Pinault, D., & Deschenes, METRO. (1998). Anatomical evidence for a
mechanism of lateral inhibition in the rat thalamus. European
Revista de neurociencia, 10(11), 3462–3469. https://doi.org/10
.1046/j.1460-9568.1998.00362.x, PubMed: 9824459
Prasad, j. A., Carroll, B. J., & sherman, S. METRO. (2020). Layer 5 corti-
cofugal projections from diverse cortical areas: Variations on a
pattern of thalamic and extrathalamic targets. Journal of Neuro-
s c i en c e , 4 0 (3 0), 5 78 5 –5 7 9 6. h t t ps : / / d o i . o rg / 1 0. 1 5 2 3
/JNEUROSCI.0529-20.2020, PubMed: 32532890
Qian, J., & Yazdanbakhsh, A. (2015). A neural model of
distance-dependent percept of object size constancy. PLoS
Uno, 10(7), e0129377. https://doi.org/10.1371/journal.pone
.0129377, PubMed: 26132106
Reichova, I., & sherman, S. METRO. (2004). Somatosensory corticotha-
lamic projections: Distinguishing drivers from modulators. Jour-
nal of Neurophysiology, 92(4), 2185–2197. https://doi.org/10
.1152/jn.00322.2004, PubMed: 15140908
Rouiller, mi. METRO., & Durif, C. (2004). The dual pattern of corticotha-
lamic projection of the primary auditory cortex in macaque mon-
key. Neuroscience Letters, 358(1), 49–52. https://doi.org/10.1016
/j.neulet.2004.01.008, PubMed: 15016432
Rouiller, mi. METRO., & Welker, mi. (1991). Morphology of corticothalamic
terminals arising from the auditory cortex of the rat: A Phaseolus
vulgaris-leucoagglutinin (PHA-L) tracing study. Hearing
Investigación, 56(1–2), 179–190. https://doi.org/10.1016/0378
-5955(91)90168-9, PubMed: 1769912
Rouiller, mi. METRO., & Welker, mi. (2000). A comparative analysis of the
morphology of corticothalamic projections in mammals. Cerebro
Research Bulletin, 53(6), 727–741. https://doi.org/10.1016
/S0361-9230(00)00364-6, PubMed: 11179837
Rovo, Z., Ulbert, I., & Acsady, l. (2012). Drivers of the primate
thalamus. Revista de neurociencia, 32(49), 17894–17908.
https://doi.org/10.1523/ JNEUROSCI.2815-12.2012, PubMed:
23223308
Shai, A. S., Anastassiou, C. A., Larkum, METRO. MI., & Koch, C. (2015).
Physiology of layer 5 pyramidal neurons in mouse primary visual
corteza: Coincidence detection through bursting. PLoS Computa-
tional Biology, 11(3), e1004090. https://doi.org/10.1371/journal
.pcbi.1004090, PubMed: 25768881
Sherbakov, l., & Yazdanbakhsh, A. (2013). Multiscale sampling
model for motion integration. Revista de visión, 13(11), 18.
https://doi.org/10.1167/13.11.18, PubMed: 24080519
sherman, S. METRO. (2004). Interneurons and triadic circuitry of the thal-
amus. Trends in Neurosciences, 27(11), 670–675. https://doi.org
/10.1016/j.tins.2004.08.003, PubMed: 15474167
sherman, S. METRO. (2012). Thalamocortical interactions. Current Opin-
ion in Neurobiology, 22(4), 575–579. https://doi.org/10.1016/j
.conb.2012.03.005, PubMed: 22498715
sherman, S. METRO., & Guillery, R. W.. (1996). Functional organization
of thalamocortical relays. Revista de neurofisiología, 76(3),
1367–1395. https://doi.org/10.1152/jn.1996.76.3.1367,
PubMed: 8890259
sherman, S. METRO., & Guillery, R. W.. (2002). The role of the thalamus
in the flow of information to the cortex. Philosophical Transac-
tions of the Royal Society B, 357(1428), 1695–1708. https://doi
.org/10.1098/rstb.2002.1161, PubMed: 12626004
sherman, S. METRO., & Guillery, R. W.. (2006). Exploring the thalamus
and its role in cortical function. Cambridge, MAMÁ: CON prensa.
sherman, S. METRO., & Guillery, R. W.. (2013). Functional connections of
cortical areas: A new view from the thalamus. Cambridge, MAMÁ: CON
Prensa. https://doi.org/10.7551/mitpress/9780262019309.001.0001
Shosaku, A. (1986). Cross-correlation analysis of a recurrent inhib-
itory circuit in the rat thalamus. Revista de neurofisiología,
55(5), 1030–1043. https://doi.org/10.1152/jn.1986.55.5.1030,
PubMed: 3711965
Simko, J., & Markram, h. (2021). Morphology, physiology and syn-
aptic connectivity of local interneurons in the mouse somatosen-
sory thalamus. Revista de fisiología, 599(22), 5085–5101.
https://doi.org/10.1113/JP281711, PubMed: 34591324
Herrero, y., Seguela, PAG., & Padre, A. (1987). Distribution of
GABA-immunoreactive neurons in the thalamus of the squirrel
monkey (Saimiri sciureus). Neurociencia, 22(2), 579–591. https://
doi.org/10.1016/0306-4522(87)90355-1, PubMed: 3670598
Steriade, METRO. (2005). Sleep, epilepsy and thalamic reticular inhibi-
tory neurons. Trends in Neurosciences, 28(6), 317–324. https://
doi.org/10.1016/j.tins.2005.03.007, PubMed: 15927688
Steriade, METRO., & Deschenes, METRO. (1984). The thalamus as a neuronal
oscillator. Brain Research, 320(1), 1–63. https://doi.org/10.1016
/0165-0173(84)90017-1, PubMed: 6440659
Steriade, METRO., Domich, l., Oakson, GRAMO., & Deschenes, METRO. (1987).
The deafferented reticular thalamic nucleus generates spindle
rhythmicity. Revista de neurofisiología, 57(1), 260–273.
https://doi.org/10.1152/jn.1987.57.1.260, PubMed: 3559675
Steriade, METRO., McCormick, D. A., & Sejnowski, t. j. (1993).
Thalamocortical oscillations in the sleeping and aroused brain.
Ciencia, 262(5134), 679–685. https://doi.org/10.1126/science
.8235588, PubMed: 8235588
Tai, y., Hacer, h., Ilinsky, I. A., & Kultas-Ilinsky, k. (1995). Nucleus reti-
cularis thalami connections with the mediodorsal thalamic
núcleo: A light and electron microscopic study in the monkey.
Brain Research Bulletin, 38(5), 475–488. https://doi.org/10.1016
/0361-9230(95)02018-METRO, PubMed: 8665272
Thomson, A. METRO., & lamy, C. (2007). Functional maps of neocorti-
cal local circuitry. Frontiers in Neuroscience, 1(1), 19–42. https://
doi.org/10.3389/neuro.01.1.1.002.2007, PubMed: 18982117
Timbie, C., Garcia-Cabezas, METRO. A., Zikopoulos, B., & Barbas, h.
(2020). Organization of primate amygdalar-thalamic pathways
for emotions. Biología PLoS, 18(2), e3000639. https://doi.org/10
.1371/journal.pbio.3000639, PubMed: 32106269
Wamsley, mi. J., Tucker, METRO. A., Shinn, A. K., Ono, k. MI., McKinley,
S. K., Ely, A. v., … Manoach, D. S. (2012). Reduced sleep spin-
dles and spindle coherence in schizophrenia: Mechanisms of
impaired memory consolidation? Biological Psychiatry, 71(2),
154–161. https://doi.org/10.1016/j.biopsych.2011.08.008,
PubMed: 21967958
Warren, R. A., Agmon, A., & jones, mi. GRAMO. (1994). Oscillatory synaptic
interactions between ventroposterior and reticular neurons in mouse
thalamus in vitro. Revista de neurofisiología, 72(4), 1993–2003.
https://doi.org/10.1152/jn.1994.72.4.1993, PubMed: 7823113
willis, A. METRO., Slater, B. J., Gribkova, mi. D., & Llano, D. A. (2015).
Open-loop organization of thalamic reticular nucleus and dorsal
thalamus: A computational model. Revista de neurofisiología,
114(4), 2353–2367. https://doi.org/10.1152/jn.00926.2014,
PubMed: 26289472
Wurbs, J., Mingolla, MI., & Yazdanbakhsh, A. (2013). Modeling a
space-variant cortical representation for apparent motion. Diario
Neurociencia en red
767
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sleep spindles in core/matrix thalamocortical circuits
of Vision, 13(10), 2. https://doi.org/10.1167/13.10.2, PubMed:
23922444
xiao, D., Zikopoulos, B., & Barbas, h. (2009). Laminar and modu-
lar organization of prefrontal projections to multiple thalamic
nuclei. Neurociencia, 161(4), 1067–1081. https://doi.org/10
.1016/j.neuroscience.2009.04.034, PubMed: 19376204
Yoshimura, y., Dantzker, j. l. METRO., & Callaway, mi. METRO. (2005). Excit-
atory cortical neurons form fine-scale functional networks.
Naturaleza, 433(7028), 868–873. https://doi.org/10.1038
/nature03252, PubMed: 15729343
zhang, l., & jones, mi. GRAMO. (2004). Corticothalamic inhibition in the
thalamic reticular nucleus. Revista de neurofisiología, 91(2),
759–766. https://doi.org/10.1152/jn.00624.2003, PubMed:
14586030
Zikopoulos, B., & Barbas, h. (2006). Prefrontal projections to the
thalamic reticular nucleus form a unique circuit for attentional
mechanisms. Revista de neurociencia, 26(28), 7348–7361.
https://doi.org/10.1523/ JNEUROSCI.5511-05.2006, PubMed:
16837581
Zikopoulos, B., & Barbas, h. (2007a). Circuits for multisensory inte-
gration and attentional modulation through the prefrontal cortex
and the thalamic reticular nucleus in primates. Reviews in the
Neurosciences, 18(6), 417–438. https://doi.org/10.1515
/REVNEURO.2007.18.6.417, PubMed: 18330211
Zikopoulos, B., & Barbas, h. (2007b). Parallel driving and modula-
tory pathways link the prefrontal cortex and thalamus PLoS One,
2(9), e848. https://doi.org/10.1371/journal.pone.0000848,
PubMed: 17786219
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
7
4
3
2
1
1
8
3
6
6
norte
mi
norte
_
a
_
0
0
3
1
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurociencia en red
768