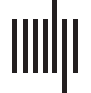INVESTIGACIÓN
Age differences in functional brain networks
associated with loneliness and empathy
Laetitia Mwilambwe-Tshilobo1, Roni Setton2, Danilo Bzdok1,3,4,5,6,
Gary R. Turner7, y r. Nathan Spreng1,4,8,9
1Instituto Neurológico de Montreal, Departamento de Neurología y Neurocirugía,
Universidad McGill, Montréal, QC, Canada
2Department of Psychology, Harvard University, Bostón, MAMÁ, EE.UU
3Department of Biomedical Engineering, Universidad McGill, Montréal, QC, Canada
4Centro de imágenes cerebrales McConnell, Universidad McGill, Montréal, QC, Canada
5School of Computer Science, Universidad McGill, Montréal, QC, Canada
6Mila–Quebec Artificial Intelligence Institute, Montréal, QC, Canada
7Department of Psychology, York University, toronto, ON, Canada
8Departments of Psychiatry and Psychology, Universidad McGill, Montréal, QC, Canada
9Douglas Mental Health University Institute, Verdun, QC, Canada
Palabras clave: Loneliness, Empathy, Social cognition, Default network, Aging, Resting-state functional
conectividad
ABSTRACTO
Loneliness is associated with differences in resting-state functional connectivity (RSFC) dentro
and between large-scale networks in early- and middle-aged adult cohorts. Sin embargo, edad-
related changes in associations between sociality and brain function into late adulthood are
not well understood. Aquí, we examined age differences in the association between two
dimensions of sociality—loneliness and empathic responding—and RSFC of the cerebral
corteza. Self-report measures of loneliness and empathy were inversely related across the
entire sample of younger (mean age = 22.6y, norte = 128) and older (mean age = 69.0y,
norte = 92) adultos. Using multivariate analyses of multi-echo fMRI RSFC, we identified distinct
functional connectivity patterns for individual and age group differences associated with
loneliness and empathic responding. Loneliness in young and empathy in both age groups
was related to greater visual network integration with association networks (p.ej., default,
fronto-parietal control). A diferencia de, loneliness was positively related to within- and between-
network integration of association networks for older adults. These results extend our previous
findings in early- and middle-aged cohorts, demonstrating that brain systems associated
with loneliness, as well as empathy, differ in older age. Más, the findings suggest that these
two aspects of social experience engage different neurocognitive processes across human
life-span development.
RESUMEN DEL AUTOR
Feelings of loneliness emerge when a person’s desire or need for an interpersonal relationship
is unmet. This state of perceived social isolation can influence social-cognitive processes
that are critical for connecting with others, such as empathy. Neuroimaging studies have
shown diverging functional connectivity patterns among functional brain networks between
lonely younger and middle-aged adults. Aquí, we take a targeted approach to directly assess
age-related differences in functional connectivity associated with loneliness and empathic
responding in younger and older adults. We find evidence that individual differences in
un acceso abierto
diario
Citación: Mwilambwe-Tshilobo, l.,
Setton, r., Bzdok, D., Tornero, GRAMO. r., &
Spreng, R. norte. (2023). Age differences in
functional brain networks associated
with loneliness and empathy. Red
Neurociencia, 7(2), 496–521. https://
doi.org/10.1162/netn_a_00293
DOI:
https://doi.org/10.1162/netn_a_00293
Supporting Information:
https://doi.org/10.1162/netn_a_00293
Recibió: 10 Febrero 2022
Aceptado: 18 Noviembre 2022
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autores correspondientes:
Laetitia Mwilambwe-Tshilobo
laetitia.mwilambwe-tshilobo
@mail.mcgill.ca
R. Nathan Spreng
nathan.spreng@gmail.com
Editor de manejo:
Olaf Sporns
Derechos de autor: © 2023
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
norte
mi
norte
_
a
_
0
0
2
9
3
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
functional connectivity related to loneliness and empathic responding differ with age. Nosotros
discuss possible mechanisms underlying these associations and their implications for brain
and social functioning across the adult life-span.
INTRODUCCIÓN
Forming and maintaining social bonds are among the most complex of human abilities. Soci-
ality and the emergence of social collaboration within species have been linked to larger brain
sizes, with humans at the peak of this evolutionary continuum (Dunbar & Shultz, 2007). Social
functioning is related to functional activation and connectivity among multiple large-scale
brain systems (Mars et al., 2012; Moran et al., 2012; Mwilambwe-Tshilobo et al., 2019; Spreng
et al., 2020). The importance of sociality as a determinant of brain health is most evident when
social needs go unmet. Perceived social isolation, or loneliness, has a significant negative
impact on mental and physical health (Cacioppo et al., 2014; Ong et al., 2016; Shankar
et al., 2013; Tilvis et al., 2011). Lonely individuals experience increased risk for cognitive
decline (Boss et al., 2015; Wilson et al., 2015), neuropathological burden (d’Oleire Uquillas
et al., 2018; Donovan et al., 2016), and Alzheimer’s disease (Wilson et al., 2007). A pesar de
loneliness is related to adverse cognitive sequelae in age-related brain disease, much of the
research investigating the impact of loneliness on brain structure and function has been con-
ducted in younger or middle-aged adults (see Lam et al., 2021, para una revisión). Loneliness poses
significant health risks and is a burden, particularly for older adults. Sin embargo, its differential
impact on brain function in early and late adulthood remains largely unexplored.
Although the experience of loneliness varies between people, it emerges because one’s
need for social connection is unfulfilled (Cacioppo & Hawkley, 2009). The felt absence of
connection has marked effects on the cognitive and affective processing of social signals
(Bangee et al., 2014; Cacioppo et al., 2015). Loneliness influences perception and attention,
resulting in negatively biased social perception and altered social functioning (Cacioppo &
Hawkley, 2009). Poor perception of social cues associated with feeling lonely may hinder
the ability to recognize and accurately interpret others’ thoughts and feelings, both core
features of empathic responding. Preliminary evidence in younger adults indicates that this
interaction may alter the impact of loneliness on the brain. Loneliness is inversely related to
white matter integrity in brain regions implicated in social-cognitive processes, with higher
empathy moderating this relationship (Nakagawa et al., 2015). This finding suggests that the
negative behavioral association between loneliness and empathy may have a direct neural
correlate, with each exerting opposing brain effects.
A growing body of neuroimaging studies now link individual differences in loneliness
(Cacioppo et al., 2009a, 2009b; Düzel et al., 2019; Kong et al., 2015; Layden et al., 2017;
Nakagawa et al., 2015; Wong et al., 2016) and empathy (Schurz et al., 2021a, 2021b; Völlm
et al., 2006) to structural and functional changes in brain regions spanning multiple neurocog-
nitive systems. Resting-state functional connectivity (RSFC) has demonstrated that interactions
among spatially distributed brain regions underlie individual differences in loneliness (feng
et al., 2019; Mwilambwe-Tshilobo et al., 2019; Spreng et al., 2020) and empathy (Christov-
Moore et al., 2020; Finn et al., 2015; Katsumi et al., 2021).
Sin embargo, many open questions remain regarding the neural associations between loneli-
ness and empathy. Primero, empathy is a multidimensional construct consisting of cognitive and
affective components, each with distinct neural patterns (Cox et al., 2012; Schurz et al.,
Loneliness:
The subjective feeling of social
isolation when one’s social needs are
unmet by the quantity or quality of
one’s social relationships.
Empathy:
The ability to infer, understand, o
simulate the feelings and thoughts of
other people.
Neurocognitive systems:
Spatially distributed large-scale brain
networks associated with cognition.
Neurociencia en red
497
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
norte
mi
norte
_
a
_
0
0
2
9
3
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
2021a). While behavioral evidence suggests that associations with loneliness vary in strength
between components (Beadle et al., 2012), this finding may not extend to all aspects of
empathic responding (es decir., empathic concern and perspective taking; Kanai et al., 2012).
Segundo, associations between empathic responding and loneliness may change as people
edad. As people age, the quality of social relationships becomes more important than quantity
(Carstensen, 1992). Older adults experience higher risks for loneliness (es decir., social isolation;
Steptoe et al., 2013; Luhmann & Hawkley, 2016). Because loneliness impacts social percep-
ción (Cacioppo et al., 2009a, 2009b), lonely older adults may have more difficulties forming
meaningful social bonds. Variability in the impact of aging on the neural correlates of cognitive
and affective components of empathy (Beadle & de la Vega, 2019) may also influence age
differences in the relationship between loneliness and empathic responding. Por lo tanto, bajo-
standing the role aging plays in both behaviors may provide more insights into their individual
and combined effects on the intrinsic functional connectivity of the brain.
Two studies investigating the relationship between RSFC and loneliness provide early evi-
dence for putative age differences in brain-loneliness associations. In a large cohort of younger
adultos, Mwilambwe-Tshilobo et al. (2019) identified that loneliness was associated with greater
RSFC between default network regions and visual and attention networks. These associations
are consistent with loneliness-related neural changes in externally directed perceptual and
attention networks (Schurz et al., 2021a) and support the altered social perception hypothesis
of loneliness and empathy (Cacioppo et al., 2015). A diferencia de, a population-based study of late
middle-aged adults revealed that loneliness was positively related to RSFC between the default
network and fronto-parietal control and limbic regions, but not the visual network (Spreng et al.,
2020). Además, while higher default network integration was negatively correlated with lone-
liness in young adults (Mwilambwe-Tshilobo et al., 2019), it was positively associated with lone-
liness in middle-aged adults. The authors provided evidence that loneliness in middle age may
precipitate more internally directed cognitive processes, mediated by the default network, como
lonely individuals mentalize about desired but unmet social interactions. Combined, estos
two studies suggest a shift in the impact of loneliness from changes occurring in brain networks
associated with externally directed cognitive processes in early adulthood to an upregulation in
brain networks associated with internally directed cognitive processes in middle adulthood.
The relationship between loneliness and age is U-shaped, with peaks at 30 y 60 years of
edad (Luhmann & Hawkley, 2016). This relationship corresponds to the average ages in the youn-
ger and middle-aged studies described above (Mwilambwe-Tshilobo et al., 2019; Spreng et al.,
2020), raising the intriguing possibility that, while the prevalence of loneliness may be similar,
the social, cognitivo, and neural sequelae may shift across the adult life-span. En el presente estudio,
we directly examine how individual and age differences in sociality—loneliness and empathic
responding—relate to the intrinsic network architecture of the brain. We focus our analysis spe-
cifically on brain regions within six networks previously implicated in loneliness in younger and
adultos mayores (Mwilambwe-Tshilobo et al., 2019; Spreng et al., 2020; Lam et al., 2021): visual,
atención dorsal, atención ventral, límbico, fronto-parietal, and default networks. We test the pre-
diction that diverging loneliness-related RSFC patterns previously identified in young and
middle-aged adults will be observed when directly comparing younger and older adults. Specif-
icamente, we hypothesize that younger adults will show a consistent pattern as Mwilambwe-
Tshilobo et al. (2019) characterized by greater functional integration of the default network with
visual and attention networks. A diferencia de, older adults will show greater functional integration of
the default network with fronto-parietal and limbic networks that more closely aligns with the
patterns observed in middle-aged adults (Spreng et al., 2020). Consistent with research indicating
that older adults prioritize close social relationships (Carstensen, 1992) and that loneliness alters
Internally directed cognitive
procesos:
Mental processes that rely upon
attentional allocation to internally
generated information (p.ej., memory,
emotions).
Externally directed cognitive
procesos:
Mental processes that rely upon
attentional allocation to
extrapersonal space and/or stimuli
(p.ej., sensory perception).
Neurociencia en red
498
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
norte
mi
norte
_
a
_
0
0
2
9
3
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
brain regions implicated in social functioning (Cacioppo et al., 2009a, 2009b; Kanai et al., 2012),
we hypothesized that the RSFC patterns positively associated with loneliness would be inversely
related to empathic responding and more robustly expressed in older adults than younger adults.
MÉTODOS
Participantes
Data from 220 participants were analyzed in the present study. Participants were part of a
larger cohort (Spreng et al., 2022), where inclusion required both loneliness and empathy
evaluaciones, and two resting-state fMRI runs of data. Participants in the final sample included
128 younger (Mage = 22.6 años, DE = 3.3; range = 18–34; 75% femenino) y 92 older (Mage =
69 años, DE = 6.6, range = 60–89; 47% femenino) healthy adults recruited in Ithaca, Nueva York,
and Toronto, Canada (Mesa 1). All participants were right-handed, que van desde 18 a 89 años
Mesa 1. Descriptive data (mean and standard deviations) and inferential statistics for behavioral measures in younger and older adults
En general
Younger adults
Older adults
Significance
Demographics
norte
Age, significar (Dakota del Sur)
Gender, norte (%)
Educación, significar (Dakota del Sur)
Social measures
220
128
92
42.0 (23.5)
22.6 (3.3)
69.0 (6.6)
F
METRO
122 (55.5)
98 (44.5)
16.1 (2.6)
75 (58.6)
53 (41.4)
47 (51.1)
45 (48.9)
15.2 (1.8)
17.5 (2.9)
< 0.001***
UCLA Loneliness Scale, mean (SD)
39.6 (9.1)
40.6 (9.4)
38.2 (8.5)
SNI Size, mean (SD)
22.5 (12.5)
23.7 (12.4)
20.9 (12.4)
Instrumental support, mean (SD)
Emotional support, mean (SD)
Friendship, mean (SD)
Empathic functioning
31.1 (7.6)
33.9 (5.4)
31.3 (6.3)
31.1 (8.7)
32.8 (5.4)
29.9 (6.6)
30.6 (6.8)
34.7 (5.4)
32.3 (5.9)
0.06
0.16
0.30
0.02*
0.01**
Reading the Mind in the Eyes (RMIE), mean (SD)
72.2 (10.0)
74.4 (9.8)
69.2 (9.7)
<0.001***
Toronto Empathy Questionnaire (TEQ), mean (SD)
39.0 (3.9)
39.1 (4.1)
38.8 (3.6)
IRI perspective taking (PT), mean (SD)
IRI empathic concern (EC), mean (SD)
2.8 (0.6)
3.0 (0.5)
2.7 (0.6)
2.9 (0.5)
2.8 (0.6)
3.1 (0.5)
0.65
0.18
0.003**
Personality
Neuroticism, mean (SD)
Cognition
2.5 (0.7)
2.7 (0.7)
2.2 (0.6)
<0.001***
NIH Cognitive Composite Score, mean (SD)
126.6 (14.4)
131.4 (14.7)
119.9 (10.9)
<0.001***
Note. IRI = Interpersonal Reactivity Index, SNI = Social Network Index, SD = standard deviation.
*p < 0.05, **p < 0.01, ***p < 0.001.
Network Neuroscience
499
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
(M = 42, SD = 23.5). All participants provided informed consent following the guidelines set
by the Institutional Review Board at Cornell University and York University.
Behavioral Measures
Loneliness measures. Loneliness was measured using the Revised UCLA Loneliness Scale
(UCLA-LS; Russell, 1996). The UCLA-LS is a 20-item questionnaire that measures subjective
feelings of loneliness and perceived social isolation (Russell, 1996). This measure is well-
established in the literature and highly reliable (Russell, 1996). One of the advantages of
the UCLA-LS questionnaire is that it assesses loneliness indirectly, which diminishes potential
response bias (Shiovitz-Ezra & Ayalon, 2012). For example, participants are asked to respond
to statements such as “How often do you feel like there is no one you can turn to?” or “How
often do you feel isolated from others.” Responses were provided on a 4-point Likert scale
ranging from 1 (Never) to 3 (Always). Negatively worded items were reverse-scored. Higher
scores reflect higher self-reported loneliness.
Aspects of empathic responding. Empathy is not a unitary concept but a multidimensional
construct that can be broken down into cognitive and affective components. The cognitive
components of empathy describe processes that underlie the ability to understand and make
inferences regarding another person’s mental state. The affective components of empathy
describe the emotional reaction toward the observed experiences of another. While both com-
ponents are conceptually distinct, recent work suggests that overlapping and unique brain
activation patterns support the ability to understand how other people think and feel (Schurz
et al., 2021a). To ensure that our assessment of empathic responding reflects cognitive and
affective neurocognitive processes, participants completed a performance-based assessment
of emotional recognition in addition to two self-report questionnaires that represent subdo-
mains of empathy along these two dimensions:
1. The Reading the Mind in the Eyes (RMIE) task was originally conceptualized as a theory
of mind questionnaire (Baron-Cohen et al., 2001). However, recent work suggests that
the RMIE measures emotional recognition, not theory of mind (Oakley et al., 2016). We
included the RMIE as a task-based measure in our analysis because emotional recogni-
tion is a critical aspect of empathic responding that is also predictive of prosociality
(Bailey et al., 2020). The RMIE task consists of 36 photos of the eye region of adults
expressing different emotional states. Participants were asked to choose one adjective
from a list of four that best expresses the internal state depicted in the photo. One point
was assigned for each correct response, 0 points for incorrect. Individual items were
summed to give a maximum of 36, with higher scores indicating better emotional
decoding.
2. The Toronto Empathy Questionnaire (TEQ) is a self-report measure that primarily
assesses emotional empathy (Spreng et al., 2009). It consists of 16 items in which par-
ticipants respond on a 5-point Likert scale ranging from 0 (Never) to 4 (Always). Neg-
atively worded items were scored in reverse. Examples of items in the TEQ include “I
can tell when others are sad even when they do not say anything” and “When I see
someone being treated unfairly, I do not feel very much pity for them.”
3. The Interpersonal Reactivity Index (Davis, 1980) is a self-report questionnaire that con-
sists of four subscales that assess different aspects of empathy: (1) Perspective taking
(PT), the ability to take another person’s psychological point of view; (2) Fantasy, the
ability to project oneself onto fictional characters; (3) Empathic Concern (EC), the ten-
dency to experience feelings of sympathy and compassion for others; and (4) Personal
Network Neuroscience
500
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
Distress, a measure of the aversive response one feels when observing the negative
experience of others. For this study, we only included measures of PT and EC because
we were specifically interested in assessing cognitive and affective aspects of empathic
responding that were other-focused (i.e., PT, EC) rather than self-focused (Fantasy,
Personal Distress). Each subscale had seven items with responses made on a 5-point
Likert scale ranging from 1 = “Does not describe me well” and 5 = “Describes me well”.
Negatively worded items were scored in reverse.
Covariates. Several demographic, social, cognitive, and personality variables associated with
loneliness and aging were included as covariates in our analyses. Demographic variables
included age, gender, and educational attainment. We also included the study site as a covar-
iate since participants were part of a multisite cohort study (Spreng et al., 2022). Since our
study focused on identifying age-related differences in the behavioral and neural associations
between loneliness and empathy, we needed to account for known age-dependent factors that
influence social and brain functioning. Age differences in loneliness are due to differences in
the distribution of risk factors. For example, older adults accumulate disproportionate risk
factors contributing to loneliness (Luhmann & Hawkley, 2016), two of which are poor cogni-
tive functioning and social isolation. Evidence suggests that loneliness may accelerate cogni-
tive decline among older adults (Shankar et al., 2013; Wong et al., 2016), and previous work
highlights the need to account for the confounding effects of objective social isolation when
examining loneliness among older adults (Steptoe et al., 2013). Age differences in loneliness
may also be due to normative age-related changes in the quantity and quality of social rela-
tionships. Aging is marked by significant transitions in the size and composition of social rela-
tionships that lead to shrinking social network size to prioritize close social ties (Carstensen,
1992). Social relationship quantity and quality are negatively correlated with loneliness.
However, having few high-quality relationships is a much stronger predictor of loneliness
(Luhmann & Hawkley, 2016). Thus, accounting for the quantity and quality of social relation-
ships may be important factors in how both age groups experience loneliness. Therefore, we
controlled for objective social isolation, relationship quality, and global cognitive function
measures to account for normative social network size and cognitive declines. We used the
Social Network Index (Cohen et al., 1997), NIH Toolbox Emotion Battery, and Cognition
Battery (https://www.nihtoolbox.org), respectively:
1. The Social Network Index is a self-report questionnaire that assesses various aspects of
social engagement with 12 different types of social relationships (e.g., spouse, children,
relative, friend, neighbor, coworker). Participants were asked to indicate the number of
people they regularly talk to or see at least once every two weeks for each relationship
type. The total number of people identified was summed to estimate social network size.
2. The NIH Toolbox Emotion Battery included three measures where participants were
asked to report on their perception of social support and friendship available to them
by others in their social networks (Salsman et al., 2013): (1) Instrumental Support: the
subjective perception that others in their social network are available to provide advice
in times of need; (2) Emotional Support: the subjective perception that people in their
social network are available to listen to one’s concerns with understanding and caring;
and (3) Friendship: the subjective perception that they have companions/friends avail-
able to them with which they can interact.
3. The NIH Toolbox Cognition Battery included a global composite score of overall cog-
nition, which was automatically computed by averaging scores across seven cognitive
function tests: the Picture Vocabulary Test and Oral Reading Recognition Test,
Network Neuroscience
501
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
Dimensional Change Card Sort Test, the Flanker Inhibitory Control and Attention Test,
the Picture Sequence Memory Test, the List Sorting Working Memory Test, and the
Pattern Comparison Processing Speed Test (Gershon et al., 2013). Higher scores repre-
sent better performance.
Beyond social isolation and cognition, certain personality traits may be risk factors for lone-
liness. Neuroticism is a personality trait that strongly positively correlates with loneliness
(Abdellaoui et al., 2019). In addition, neuroticism has been associated with cognitive decline
(D’Iorio et al., 2018). It mediates the relationship between loneliness and structural changes to
dorsolateral prefrontal cortex (Kong et al., 2015). To account for the potential contribution of
neuroticism when examining age differences in our analyses, we included neuroticism as a
covariate. Participants completed The Big Five Aspect Scale (DeYoung et al., 2007), which
is a 100-item self-report questionnaire that assesses facets of personality traits.
Behavioral Data Analysis
We first conducted an independent samples t test to compare younger and older adults on all
behavioral measures, including covariates. This allowed us to determine whether there were
any age-related differences in self-reported loneliness and empathy. We then performed
product-moment and partial correlations analyses to characterize the associations among all
behavioral measures. Next, we examined associations between loneliness and each measure
of empathic responding (RMIE, TEQ, IRI perspective taking, IRI empathic concern) in the full
sample and separately within each age group. All covariates were included in partial correla-
tion models (age, gender, site, education, neuroticism, cognitive composite score). The partial
correlation analysis excluded participants with missing data on any of the covariate measures.
In addition, given that a sizable portion of participants had missing data on the social network
size measure (young adults: n = 32; older adults: n = 20), additional partial correlation anal-
yses that included social network size as a covariate were conducted only in participants with
complete behavioral data. Subjective measures of instrumental support, emotional support,
and friendship were also included. Statistical significance was set at p < 0.05.
Neuroimaging
Imaging data were acquired on a 3T GE750 Discovery series MRI scanner with a 32-channel
head coil at the Cornell Magnetic Resonance Imaging Facility in Ithaca, NY, or on a 3T
Siemens Tim Trio MRI scanner with a 32-channel head coil at the York University Neuroim-
aging Center in Toronto, Canada. Scanning protocols were closely matched across sites.
Anatomical scans at Cornell were acquired using a T1-weighted volumetric magnetization
prepared rapid gradient echo sequence (TR = 2,530 ms; TE = 3.4 ms; 7° flip angle; 1-mm iso-
tropic voxels, 176 slices, 5 min 25 s) with 2× acceleration with sensitivity encoding. At York,
anatomical scans were acquired using a T1-weighted volumetric magnetization prepared
rapid gradient echo sequence (TR = 1,900 ms; TE = 2.52 ms; 9° flip angle; 1-mm isotropic
voxels, 192 slices, 4 min 26 s) with 2× acceleration and generalized auto calibrating partially
parallel acquisition (GRAPPA) encoding at an iPAT acceleration factor of 2. Two 10 min 06 s
resting-state runs were acquired using a multi-echo (ME) EPI sequence at Cornell University
(TR = 3,000 ms; TE1 = 13.7 ms, TE2 = 30 ms, TE3 = 47 ms; 83° flip angle; matrix size = 72 × 72;
field of view (FOV) = 210 mm; 46 axial slices; 3-mm isotropic voxels; 204 volumes, 2.5×
acceleration with sensitivity encoding) and York University (TR = 3,000 ms; TE1 = 14 ms,
TE2 = 29.96 ms, TE3 = 45.92 ms; 83° flip angle; matrix size = 64 × 64; FOV = 216 mm; 43
axial slices; 3.4 × 3.4 × 3 mm voxels; 200 volumes, 3× acceleration and GRAPPA encoding).
Network Neuroscience
502
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
Participants were instructed to stay awake and lie still with their eyes open, breathing and
blinking normally in the darkened scanner bay.
Processing
Anatomical images were skull stripped using the default parameters in FSL BET (Smith, 2002).
Brain-extracted anatomical and functional images were submitted to ME independent compo-
nent analysis (ME-ICA; version 3.2 beta; https://github.com/ME-ICA/me-ica; Kundu et al.,
2012, 2013). ME-ICA relies on the TE-dependence model of the BOLD signal to determine
T2* in every voxel and separates the BOLD signal from non-BOLD sources of noise. Before
TE-dependent denoising, time series data were minimally preprocessed: the first four volumes
were discarded, images were computed for de-obliquing, motion correction, and anatomical-
functional coregistration, and volumes were brought into spatial alignment across TEs. The T2*
maps were then used for anatomical-functional coregistration. Gray matter and cerebrospinal
fluid compartments are more precisely delineated by the T2* map than by raw EPI images
(Speck et al., 2001; Kundu et al., 2017), which is an important consideration in aging research
where enlarged ventricles and greater subarachnoid space often blur these boundaries.
Volumes were then optimally combined across TEs and denoised. The outputs of interest
included (1) spatial maps consisting of the BOLD components, (2) reconstructed time series
containing only BOLD components, and (3) the BOLD component coefficient sets.
ME-ICA effectively removes distant-dependent motion-related artifacts in the fMRI data
(Power et al., 2018). To retain all trials and maintain the same time series length across
participants, we did not implement any additional denoising steps, such as scrubbing. Instead,
we perform an image quality assessment on the denoised time series. In native space, we iden-
tified and excluded participants with unsuccessful coregistration, residual noise (framewise
displacement (FD) > .50 mm coupled with denoised time series showing DVARS > 1; Fuerza
et al., 2012), temporal signal to noise ratio < 50, or fewer than ten retained BOLD-like
components. Forty participants were excluded after the image quality assessment (younger
adults: n = 12; older adults: n = 28). Age group and site differences in residual motion for
included participants were assessed using FD calculated on the middle echo prior to pro-
cessing. Statistical results are reported in Supporting Information Table S3.
The denoised BOLD component coefficient sets in native space, optimized for multi-echo fMRI
RSFC analyses (Kundu et al., 2013), were used in subsequent steps. We refer to these as multi-echo
functional connectivity (MEFC) data. Additional measures were taken to account for variation in
the number of independent components from ME-ICA once connectivity matrices were estimated,
as detailed below. MEFC neuroimages were mapped to a common cortical surface for each
participant using FreeSurfer v6.0.1 (Fischl, 2012). To maximize alignment between intensity
gradients of structural and functional data (Greve & Fischl, 2009), MEFC data were first linearly
registered to the T1-weighted image by run. The inverse of this registration was used to project
the T1-weighted image to native space and resample the MEFC data onto a cortical surface
(fsaverage5) with trilinear volume-to-surface interpolation. This produces a cortical surface map
where each vertex, or surface point, is interpolated from the voxel data. Once on the surface, runs
were concatenated, and MEFC data at each vertex were normalized to zero mean and unit variance.
Individualized RSFC Parcellation
We generated participant-specific functional connectomes to examine individual differences in
functional brain network organization using the Group Prior Individual Parcellation (GPIP;
Chong et al., 2017). This approach enables a more accurate estimation of participant-specific
503
Multi-echo fMRI:
An fMRI acquisition approach that
collects multiple echo times (TEs)
after each excitation pulse. Based
upon the TE dependence of BOLD
signal across the brain, multi-echo
fMRI can remove signal dropout
zones observed with single-echo
fMRI, and quantitatively separate
BOLD from non-BOLD fMRI signals,
to improve image quality.
Group Prior Individual Parcellation
(GPIP):
An individualized parcellation
approach that identifies subject-
specific functional neuroanatomy,
while preserving parcel labels, across
a group of participants. From
standardized parcellation schemes,
parcel boundaries are tailored to
each person’s resting-state functional
connectivity data.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
individual functional areas (Chong et al., 2017) and is more sensitive to detecting RSFC asso-
ciations with behavior (e.g., Kong et al., 2021). The main advantage of this approach is that the
correspondence among parcel labels is preserved across participants, while the parcel bound-
aries are allowed to shift based on the individual-specific functional network organization of
each participant—thus providing a similar RSFC pattern that is shared across the population.
Starting from an initial predefined group parcellation atlas, GPIP first refines each individual’s
parcel boundaries relative to their resting-state fMRI data. Next, the concentration (inverse
covariance/partial correlation) matrices from all subjects are jointly estimated using a group
sparsity constraint. GPIP iterates between these two steps to continuously update the parcel
labels until convergence, defined as no more than one vertex changing per parcel or 40 itera-
tions. Compared to other group-based parcellation approaches, GPIP has been shown to
improve the homogeneity of the BOLD signal within parcels and the delineation between
regions of functional specialization (Chong et al., 2017).
Using this method, we used the MEFC data from each participant and parcellated the cortex
into 400 functionally defined regions. We initialized all participants to a group parcellation
atlas developed by Schaefer et al. (2018). Each parcel was matched to a corresponding net-
work in the seven network parcellation by Yeo et al. (2011). In the present report, we included
the visual, dorsal attention, ventral attention, limbic, fronto-parietal control, and default
networks given their reliable associations with loneliness across the neuroimaging literature
(Lam et al., 2021). In addition, as described in the Introduction, these networks have been
associated with loneliness in younger (Mwilambwe-Tshilobo et al., 2019) and late middle-
aged older adults (Spreng et al., 2020). Results for the full 7-network analysis, including
the somatomotor network, are reported in supplemental material (Supporting Information
Figures S1 and S2).
Partial Least Squares Analysis
PLS is a data-driven multivariate statistical technique used to decompose relationships
between two datasets (functional connections and behavioral measures) into orthogonal sets
of latent variables (LVs) that maximally covary together across participants (McIntosh &
Lobaugh, 2004). The latent variables can be interpreted as optimally paired functional networks
and behavioral phenotypes, respectively. We used PLS to identify age-related differences and
similarities in RSFC that were directly correlated to loneliness and empathy (Figure 1).
Two datasets were constructed: a Y matrix containing participants’ behavioral scores on
loneliness and empathy measures and an X matrix consisting of participants’ functional
connectomes. Each row of the Y and X matrices represents the number of participants
organized by group. The columns of matrix X correspond to the edges of the vectorized
lower triangle of the RSFC matrix. The X and Y matrices were mean-centered and normal-
0
ized. A correlation matrix (R = X
Y) was submitted to singular value decomposition (SVD)
as follows:
R ¼ X0Y ¼ USV0
SVD of the cross-correlation matrix X
consisting of three elements:
0
Y produced multiple mutually orthogonal LVs, each
1. A left singular vector (U) containing weights for each of the behavioral measures.
2. A right singular vector (V) containing weights for each of the functional connections that
best characterize the relationship between RSFC among younger and older adults.
3. A scalar singular value (S).
Partial least squares (PLS):
A multivariate decomposition
method that can be used to identify
patterns of covariance between a
set of brain variables and a set of
behaviors.
Latent variables (LVs):
In PLS, these are the linear
combinations of the input variables
that optimally explain orthogonal
patterns of covariance.
Network Neuroscience
504
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Analytic workflow of individual and age differences in functional connectivity related to loneliness and empathic responding. (1)
BOLD resting-state data were extracted from subject-specific individual parcellation in six networks of interest: visual, dorsal attention, ventral
attention, limbic, fronto-parietal control, and default networks. (2) Functional connectivity between parcels were constructed, forming a 323 ×
323 matrix. The lower triangle of each subject’s matrix was vectorized and arranged by group assignment into a larger RSFC matrix. (3) Youn-
ger and older adults’ scores on behavioral measures of loneliness and empathic responding were combined into a matrix. (4) Partial least
squares (PLS) was used to identify patterns of RSFC that maximally covary with the behavioral measures across subjects. A cross-
correlation matrix generated by multiplying the RSFC and behavioral matrix was submitted to singular value decomposition. (5) Network
contribution plots were used as a metric of the most reliable intra- and internetwork connections by summarizing the interregional connections
from the PLS matrix.
Squared singular values reflect effect sizes which are proportional to the covariance
between RSFC and behavior that is accounted for by each latent variable. The number of latent
variables is sorted in order of proportion of covariance between the RSFC and behavior
measures.
Brain scores:
A PLS output variable that quantifies
the extent to which a participant
expresses the group connectivity
pattern. The value is calculated by
taking the dot product of the PLS-
derived matrix and the individual
subject resting-state functional
connectivity matrix.
Participant-specific brain scores. For each latent variable, we derived participant-specific brain
scores that assess the extent to which each participant contributes to the group covariance
RSFC pattern. The brain scores were calculated by multiplying the original matrix of partici-
pants’ functional connectomes (X) with the PLS-derived right singular vector (V). Partial corre-
lations between the brain scores and each behavioral measure were conducted to account for
possible confounds in the brain-behavior correlation by controlling for covariates of no interest
(age, gender, site, education, neuroticism, and cognitive composite score). Covariates were
partialled out of both the brain scores and behavioral measures.
Permutation tests. The significance of each latent variable was assessed using permutation
testing. Rows of X were randomly reordered and subjected to SVD iteratively, as described
above. This was done 10,000 times, creating a distribution of singular values under the null
distribution (McIntosh & Mišić, 2013). A p value was computed for each latent variable as the
proportion of permuted singular values greater than or equal to the original singular value.
Critically, permutation tests involve the entire multivariate pattern and are performed in a
Network Neuroscience
505
Functional connectivity related to loneliness and empathy in aging
single analytic step, so correction for multiple comparisons is not required (McIntosh &
Lobaugh, 2004).
Bootstrap resampling. The reliability of the weights of individual RSFC connections and behav-
ior was assessed using bootstrap resampling (Krishnan et al., 2011; McIntosh & Mišić, 2013).
The brain-behavior correlations were calculated using 10,000 bootstrap samples. To identify
individual connections that made a statistically significant contribution to the overall RSFC
pattern, we calculated the ratio between each weight in the singular vector and its
bootstrap-estimated standard error. Bootstrap ratios are equivalent to z-scores if the bootstrap
distribution is approximately unit normal (Efron & Tibshirani, 1986). Bootstrap ratios were
therefore thresholded at values of ±1.96, corresponding to the 95% CI.
Cross-validation. To assess the reliability of our PLS analysis, we conducted a train-test valida-
tion of the PLS results using 5-fold cross-validation (Kebets et al., 2019). We assigned 80% of
the participant data in each age group to a train set and the remaining 20% to a test set. For
each fold, we used PLS to compute the RSFC (Utrain) and behavioral (Vtrain) singular vectors.
Then we projected the test data onto the singular vectors from the training data, allowing us to
estimate participant-specific brain scores and correlation for the test set (corr(Xtest Utrain, Ytest
Vtrain)). This was done over five folds, and the correlations between the test set original X
(RSFC) and Y (behavior) matrices were performed for LV1 and LV2. The significance of the
correlation was assessed using permutation tests (1,000 repetitions on the behavioral data
within each group).
Supplementary control analyses. We performed three additional analyses to account for pos-
sible confounding effects of
the quantity and quality of social relationships, age, and
motion on the primary PLS findings. First, we confirm that age group differences in the
relationship between loneliness and RSFC were not due to differences in either the quan-
tity or quality of relationships among younger and older adult participants. Two partial
correlation analyses were included using social network size (quantity) and subjective
measures of social support and friendship (quality). Brain-behavior correlations for the pri-
mary PLS results were computed and reported in Supporting Information Table S2B and
S2C, respectively.
The second control analysis was performed to confirm that participants’ age did not
influence the age differences captured in the primary PLS analysis. Age was used as a con-
tinuous variable and partialled out from the original X and Y input matrices. The two
matrices were then used to run a new PLS analysis (see Supporting Information Figures S5
and S6 and Supporting Information Results 1.3). Next, we compared the covariance of each
LV before and after partialling out age (Figure S7) to evaluate whether partialling out age
decreased the effect size, which would be indicative of the confounding influence of age
in our findings.
The last control analyses examined residual motion’s impact on RSFC in our sample. First,
two independent PLS analyses were performed: (1) examining the association between RSFC
and mean FD (preprocessing) and (2) identifying age differences in whole-brain RSFC (no
behavior). To confirm that the RSFC pattern covarying with FD was not associated with age
differences in RSFC, we correlated the brain scores derived from each PLS analysis. Relation-
ships are plotted for the entire sample and separately for younger and older adults (Supporting
Information Figure S8). Finally, to account for the effects of motion on the primary PLS analysis,
mean FD post-processing was included as an additional covariate (Table S4). Results are
reported in the Supporting Information.
Network Neuroscience
506
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
Network Contribution Analysis
In addition to assessing the contribution of interregional connections to the group differences,
we also evaluated the extent to which network-level RSFC within and between functional net-
works contributed to group differences. We summarized the network contributions using the
salience weights of the right singular vector (V). Two separate weighted adjacency matrices
were constructed from positive and negative salience weights by quantifying the network-level
contributions to the PLS-derived RSFC pattern. For both matrices, nodes represent parcels
defined by the individual parcellation, while edges correspond to the squared salience weights
of each pairwise connection. A summary of the network-level effects was estimated by assign-
ing each parcel of the Schaefer atlas according to their respective network label based on the
assignment reported by Yeo et al. (2011) and taking the average of all the squared saliences in
a given network, thereby generating a 6 × 6 matrix.
In order to statistically assess the network-level effects, we took a similar approach as
described above. However, we used the bootstrap ratios from the PLS-derived RSFC pattern.
Here, the edges of the positive and negative adjacency matrices correspond to the thresholded
bootstrap ratios of each pairwise connection of the RSFC pattern. Permutation tests were per-
formed for statistical assessment of the pairwise networks. During each permutation, network
labels for each node were randomly reordered and the mean intra- and inter- network boot-
strap ratio were recalculated. This process was repeated 1,000 times to generate an empirical
null sampling distribution that indicates no relationship between network assignment and
RSFC pattern (Mirchi et al., 2019). The mean contribution for all intra- and internetwork
network connections expressed as z-scores relative to the permuted null model are shown
in Supporting Information Figure S4A–B. The significance of the pairwise connections to the
network matrix was determined by estimating the proportion of times the value of the sampling
distribution was greater than or equal to the original value.
RESULTS
We measured self-reported loneliness and used self-report measures and task performance to
assess cognitive and emotional aspects of empathic responding (see Table 1). We hypothe-
sized that loneliness would be inversely related to empathic responding across the life-span
(Beadle et al., 2012; Nakagawa et al., 2015). Further, we predicted that these associations
might be more robust in later life as the detection of social cues declines (Moran et al.,
2012; Denburg et al., 2005) and socioemotional goals become increasingly salient
(Carstensen et al., 1999). Next, we examined age-related differences in the association
between loneliness, aspects of empathic responding, and cortical RSFC. We acquired 20
minutes of multi-echo resting-state fMRI data (Kundu et al., 2017) and applied individualized
parcellation to a subset of individuals previously examined to assess age differences in the
functional architecture of the brain (Setton et al., 2023). Multivariate PLS (McIntosh & Mišić,
2013; Spreng et al., 2020; Schurz et al., 2021b) was used to identify patterns of RSFC related to
individual differences in loneliness and empathic responding, as well as differences between
younger and older adult age groups. Based on our previous findings from independent samples
of young (Human Connectome Project; Mwilambwe-Tshilobo et al., 2019) and late middle-
aged adults (UK Biobank; Spreng et al., 2020), we predicted robust age differences in the asso-
ciation between loneliness, aspects of empathic responding, and RSFC. Specifically, we
hypothesized that age differences would arise within the default network and default network
interactions with other association networks implicated in internally directed cognitive pro-
cesses (Andrews-Hanna et al., 2014).
Network Neuroscience
507
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
Demographics and Descriptive Statistics
To examine whether the relationship between loneliness and empathic responding showed
similar associations as prior studies (Beadle et al., 2012; Nakagawa et al., 2015), we first char-
acterized individual and age-related differences in loneliness and subdomains of empathy
within our cohort. The behavioral measures included self-reported loneliness, emotional rec-
ognition accuracy on the RMIE task, self-reported empathy, perspective taking, and empathic
concern. Additionally, we controlled for nuisance or confounding variables, including scan-
ning site, gender, education, social network size, instrumental support, emotional support,
friendship, neuroticism, and global cognition (see Methods for full rationale). Table 1 summa-
rizes the means and standard deviations of loneliness and empathic responding measures,
along with all covariates included in subsequent analyses.
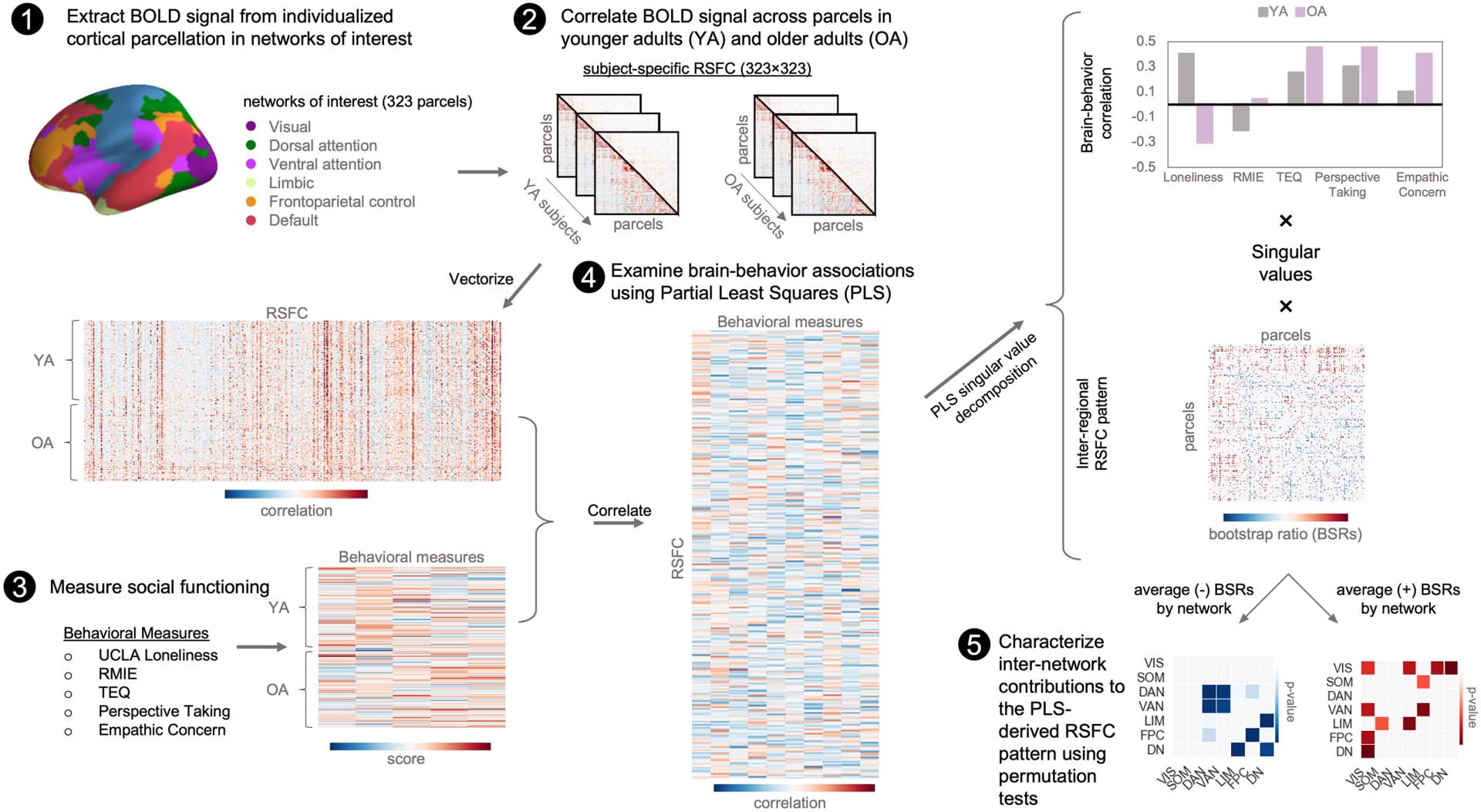
Violin plots illustrating age differences on behavioral measures in each age group are
shown in Figure 2. Scores on the UCLA loneliness scale trended lower for older, compared
to younger adults (t(218) = 1.88, p = 0.06; Cohen’s d = 0.26; Figure 2A). Younger and older
adults significantly differed on some, but not all, measures of empathy. Older adults were less
accurate at emotional recognition of others’ facial expressions based on performance on the
RMIE (t(218) = 3.94, p < 0.0001; Cohen’s d = 0.54; Figure 2F). Older adults reported greater
empathic concern than younger adults (t(218) = −3.00, p = 0.003; Cohen’s d = 0.41;
Figure 2I). No significant age differences were found on other measures of empathic function-
ing (TEQ: t(218) = 0.45, p = 0.65; Cohen’s d = 0.06; Perspective Taking: t(218) = −1.35, p =
0.18; Cohen’s d = 0.19; Figure 2G and 2H).
Neuroticism and normative cognition declines may influence the relationship between
loneliness, empathic responding, and RSFC. Therefore, we assessed age-related differences
in neuroticism and global cognition based on the NIH cognitive composite score. We also
assessed whether younger and older adults differed across covariates incorporated in subse-
quent analyses. Older adults scored lower on neuroticism (t(207.79) = 5.32, p < 0.001;
Cohen’s d = 0.72; Figure 2K) and had lower overall cognitive function (t(210) = 6.2, p <
0.001; Cohen’s d = 0.86; Figure 2J). Social networks tend to shrink with age, and evidence
from longitudinal work has found that objective social isolation may confound the effects of
loneliness among older adults (Steptoe et al., 2013). To determine whether such differences
were present within our sample, we compared the network sizes between younger and older
adults and found no age difference (t(166) = 1.42, p = 0.16; Cohen’s d = 0.22; Figure 2B). We
also included measures of perceived social support and friendship to assess the quality of
social relationships participants felt they had access to. Older adults reported greater per-
ceived emotional support (t(215) = 2.45, p = 0.02; Cohen’s d = 0.34; Figure 2D) and friend-
ship (t(215) = 2.83, p = 0.01; Cohen’s d = 0.39; Figure 2E), but no significant differences
were found for instrumental support (t(215) = −1.08, p = 0.28; Cohen’s d = 0.15; Figure 2C).
Next, we assessed the association among all behavioral variables across the full sample (see
Table 2). Scores on the UCLA loneliness scale correlated negatively with perspective taking,
empathic concern, social network size, instrumental support, emotional support, and friend-
ship. Scores on the UCLA loneliness scale were positively associated with neuroticism and
cognitive function. Accuracy on the RMIE was not significantly associated with loneliness
or other empathy subdomain measures. RMIE accuracy in this sample was positively corre-
lated with emotional support and cognitive function. The TEQ was correlated with perspective
taking and empathic concern subscales of the IRI and with participant social network size.
Empathic concern was positively correlated with perspective taking, emotional support, and
friendship, and negatively associated with neuroticism and cognitive function. Social network
Network Neuroscience
508
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
Figure 2. Group comparison on behavioral measures. Violin plots showing the distribution of behavioral scores in younger and older adults
for (A) loneliness, (B) social network size, (C–D) social support, (E) friendship, (F–I) empathic responding measures, ( J) global cognitive func-
tion, (K) and neuroticism. For comparisons on behavioral measures of interest (loneliness and empathic responding), although self-reported
loneliness was similar among age groups, significant age-related differences can be observed for task-based performance of emotional recog-
nition and in self-reported empathic concern. RMIE = Reading the Mind in the Eyes Task; TEQ = Toronto Empathy Questionnaire; **p < 0.01.
***p < 0.001.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
size was positively correlated with friendship. Emotional support was positively correlated
with friendship. Neuroticism was negatively correlated with emotional support and friendship,
and contrary to expectation, positively correlated with global cognitive function.
Behavioral Associations Between Loneliness and Empathic Responding in Younger and Older Adults
Our previous results showed significant age differences on some aspects of empathic respond-
ing. We therefore examined the association between loneliness and empathy measures in both
younger and older adults separately (controlling for gender, site, education, neuroticism, and
global cognitive function). The partial correlations between the gold standard UCLA loneliness
scale and the four measures of empathic responding are shown in Supporting Information
Table S1 for both age groups. In younger adults, loneliness was not significantly correlated
Network Neuroscience
509
t
N
e
w
o
r
k
N
e
u
r
o
s
c
e
n
c
e
i
Table 2.
Correlation of measures across all participants
1
–
0.09
−0.12
−0.17*
2
–
0.09
0.03
3
–
0.22***
4
–
5
6
7
8
9
10
11
12
−0.17*
−0.02
0.41****
0.42****
–
1. Loneliness
2. RMIE
3. TEQ
4. IRI Perspective
Taking
5. IRI Empathic
Concern
6. Social network
−0.21**
0.02
0.24**
0.1
0.17*
–
size
7. Instrumental
support
8. Emotional
support
9. Friendship
−0.34****
0
0.04
0.02
0.04
−0.03
–
−0.57****
0.15*
0.09
0.19**
0.06
0.01
0.44****
–
10. Neuroticism
0.45****
−0.68****
0.03
0.13
11. Education
−0.12
−0.05
0.08
0.04
0
12. Cognition
composite score
0.14*
0.42****
0.01
0.17*
0.06
0.27***
0.30****
0.59****
–
−0.33****
0.18**
0.03
−0.09
−0.25***
−0.23***
–
0.03
−0.02
0.13
−0.03
0.19**
−0.17*
0.07
−0.12
−0.01
−0.08
−0.11
−0.05
−0.1
–
0.27****
−0.07
–
Notes. Correlation values in boldface are statistically significant. RMIE = Reading the Mind in the Eyes Task; TEQ = Toronto Empathy Questionnaire. Social network size, neuroticism,
education, and cognition composite scores are included as covariates in analyses.
*p < 0.05, **p < 0.01, ***p < 0.001.
5
1
0
F
u
n
c
t
i
o
n
a
l
c
o
n
n
e
c
t
i
v
i
t
y
r
e
l
a
t
e
d
t
o
l
o
n
e
l
i
n
e
s
s
a
n
d
e
m
p
a
t
h
y
i
n
a
g
i
n
g
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
with any empathic responding measures. In contrast, loneliness in older adults was signifi-
cantly and negatively associated with TEQ, perspective taking and empathic concern, but
not accuracy on the RMIE task. Two additional partial correlation correlations were performed
to assess the influence of social network quantity and quality on the behavioral associations
observed in younger and older adults (Table S1B). Although not all participants in the cohort
completed the objective social network size measure, we reanalyzed the associations between
loneliness and empathic responding by including social network size as a covariate and found
that no significant associations remained (Table S1C). However, when we included instrumen-
tal support, emotional support, and friendship as proxy measures of social relationship quality,
loneliness across the full sample was significantly correlated with accuracy on the RMIE while
all remaining measures of empathic functioning were no longer significant (Table S1D).
RSFC Associations With Loneliness and Empathic Responding in Younger and Older Adults
Next, we implemented a data-driven multivariate approach to identify patterns of RSFC related
to loneliness and empathic responding in younger and older adults (Figure 1). RSFC was exam-
ined among the visual, dorsal attention, ventral attention, limbic, fronto-parietal, and default
networks. Two significant LVs capturing distinct RSFC patterns reflecting age-related differ-
ences and similarities in social behavior were observed. A scree plot showing the covariance
explained for all LVs is shown in Supporting Information Figure S3. Detailed results examining
the impact of social relationship quantity and quality on brain-behavior associations identified
by each LV are provided in Supporting Information Results 1.1.
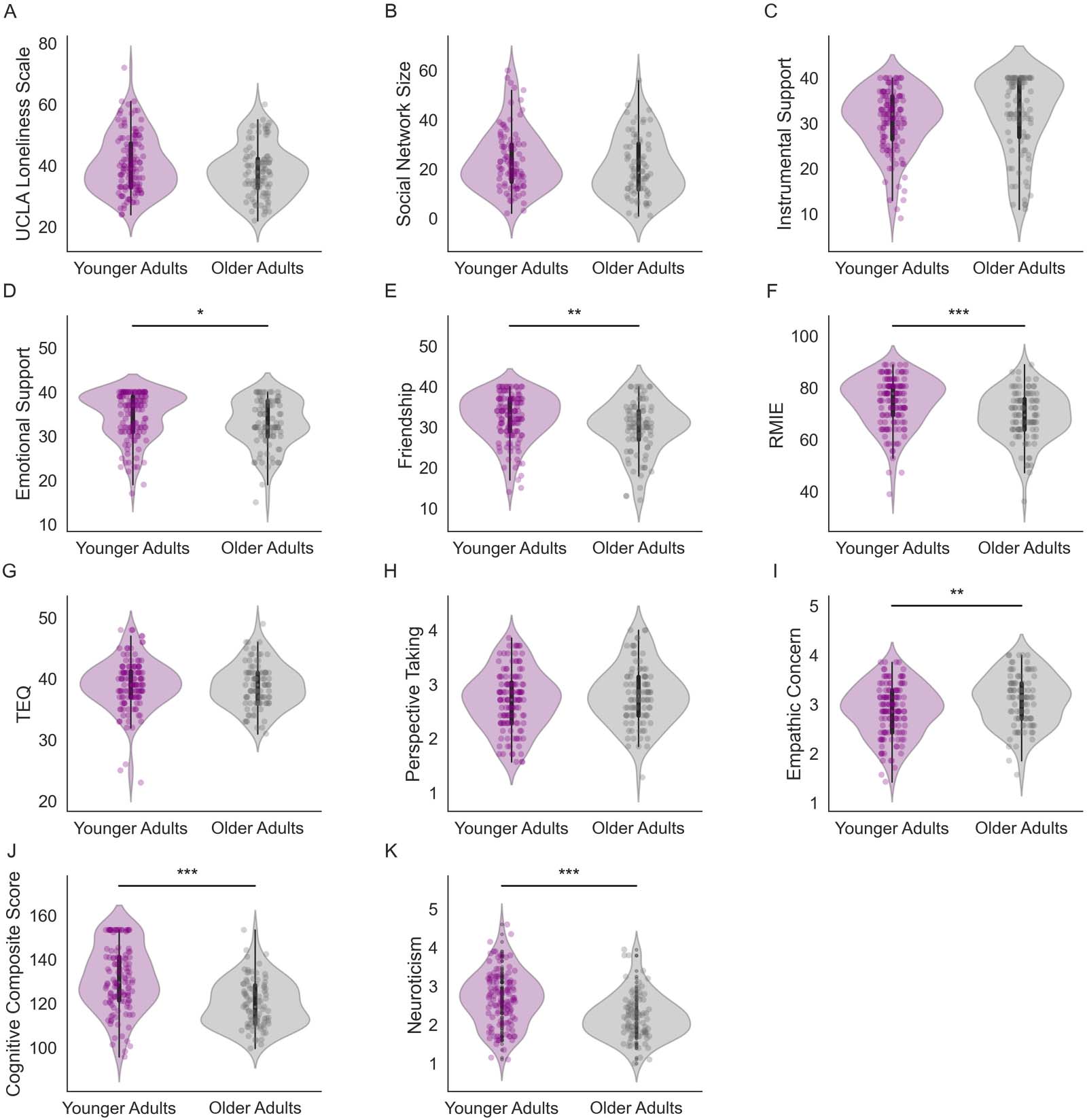
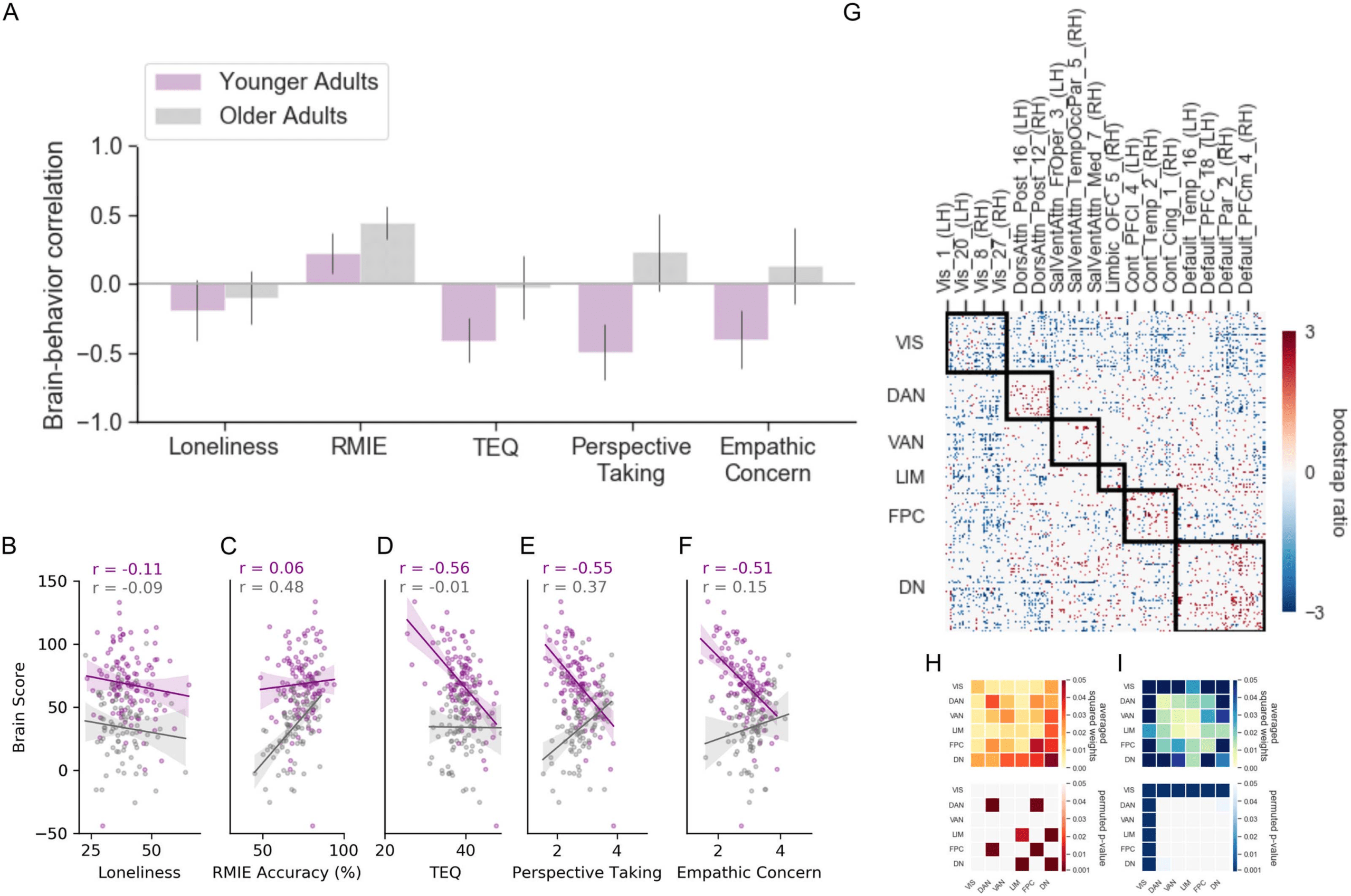
Age differences in RSFC related to loneliness. The first LV revealed a pattern of RSFC that disso-
ciated younger and older adult RSFC associated with loneliness (p = 0.04; 26.02% covariance
explained; Figure 3). Additionally, self-reported empathy covaried in both groups with a
pattern of RSFC observed for loneliness in young adults. No reliable relationship between
emotional recognition on the RMIE task and RSFC was found in either age group. To assess
the specificity of the brain-behavior correlations in each group, we performed partial correla-
tion analyses controlling for the effects of gender, site, neuroticism, and cognitive function on
participants’ brain score. Figure 3B–3F depicts scatterplots of the relationship between partic-
ipant brains scores, representing the weighted values of the RSFC pattern of the LV controlling
for covariates and all five behavioral measures. Results indicate that significant brain-behavior
correlations for LV1 were robust, as they remained significant after controlling for covariates in
both age groups (see Supporting Information Table S2A for statistical results).
Next, we summarized the average connectivity pattern within and between networks. We
examined the significance of the pairwise connections using permutation testing (Figure 3H
and 3I). The most notable feature that emerged was a dissociation between the connectivity of
the visual network and heteromodal association regions. This dissociation reflects the age
interaction in loneliness on RSFC. In younger adults, higher loneliness was associated with
greater visual network connectivity with ventral attention, fronto-parietal control, and default
networks (Figure 3H). This pattern of RSFC was also associated with self-reported empathy
(TEQ, perspective taking, and empathy) in both age groups. In contrast, higher loneliness in
older adults was associated with more intranetwork RSFC of attention, fronto-parietal, and
default networks, and greater RSFC between default and fronto-parietal, limbic, and dorsal
attention networks (Figure 3I).
RSFC related to subdomains of empathic responding. A second significant pattern revealed shared
and diverging associations related to various facets of empathic responding (p < 0.01; 16.53%
covariance explained; Figure 4). The brain-behavioral correlations for both groups are shown
Network Neuroscience
511
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. PLS analysis of brain-behavior covariance for LV1. (A) Correlation between behavioral loneliness, empathic responding measures,
and RSFC in younger and older adults. Error bars show 95% confidence intervals determined by bootstrap resampling. (B–F) Scatterplots show
correlations between participant brain scores and behavioral measures controlling for age, site, gender, education, neuroticism, and cognition
as a function of each behavioral measure. (G) Correlation matrix of the reliable pairwise functional connections associated with behavior. The
matrix bootstrap ratios are thresholded at ±2 to 3. Network-level contributions to the positive (H) and negative (I) connectivity pattern for LV1:
top matrices show the averaged squared salience weights, which reflects a summary of the connectivity pattern; bottom matrices show sig-
nificant network contribution estimated using permutation testing on the correlation matrix in panel G. Behaviors that correlate positively with
the pattern are represented in warm colors, and negative brain-behavior correlations in cool colors. VIS = Visual network; DAN = Dorsal
attention network; VAN = Ventral attention network; LIM = Limbic network; FPC = Frontoparietal control network; DN = Default network.
in Figure 4A. Across both age groups, better performance on emotional recognition on the
RMIE task correlated positively with greater intra- and internetwork RSFC among regions in
heteromodal association cortex (Figure 4G). This pattern was particularly prominent within
the dorsal attention, fronto-parietal, and default networks, and between default to other
heteromodal networks and fronto-parietal to dorsal attention networks (Figure 4H). LV2 also
captured a RSFC pattern of age group differences in the relationship between RSFC and all
empathic responding measures. Younger adults with higher scores on the TEQ, perspective
taking, and empathic concern showed strong intranetwork connectivity of the visual network
and connectivity between the visual network with the other five networks (Figure 4I). No
significant associations between RSFC and these three self-report measures of empathic
responding were found in older adults. When controlling for covariates of no interest, significant
brain-behavior correlations remained in younger adults for TEQ, perspective taking, and
empathic concern (Supporting Information Table S2A). For older adults, a significant positive
Network Neuroscience
512
Functional connectivity related to loneliness and empathy in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. PLS analysis of brain-behavior covariance for LV2. (A) Correlation between behavioral loneliness, empathic responding, and RSFC
in younger and older adults. Error bars show 95% confidence intervals determined by bootstrap through bootstrap resampling. (B–F) Scatter-
plots show correlations between participant brain scores and behavioral measures controlling for age, site, gender, education, neuroticism, and
cognition as a function of each behavioral measure. (G) Correlation matrix of the reliable pairwise functional connections associated with
behavior. The matrix bootstrap ratios are thresholded at ±2 to 3. Network-level contributions to the positive (H) and negative (I) connectivity
pattern for LV1: top matrices show the averaged squared salience weights, which reflects a summary of the connectivity pattern; bottom matri-
ces show significant network contribution estimated using permutation testing on the correlation matrix in panel G. Behaviors that correlate
positively with the pattern are represented in warm colors and negative brain-behavior correlations in cool colors.
brain-behavior correlation emerged for perspective taking. Scatterplots of the relationship
between the corrected brain scores and each behavioral measure can be found in Figure 4B–4F.
Cross-Validation of PLS Results
To account for potential overfitting from our PLS analysis, we conducted a second analysis
to assess the stability of the identified patterns (see Methods for more details). A fivefold
cross-validation was performed on the two LVs by correlating the RSFC-behavior associa-
tions of each LV in the training set and calculating the mean correlation across folds. The
mean correlation was strongly correlated (r = 0.54). RSFC-behavior correlations in the test set
representing 20% of the sample for the test set were lower but remained significantly corre-
lated across fold (r = 0.19; p = 0.003), suggesting that PLS LVs estimated from train data were
stable in the testing set.
Network Neuroscience
513
Functional connectivity related to loneliness and empathy in aging
DISCUSSION
We examined the relationship between loneliness, empathic responding, and RSFC in younger
and older adults to identify age group differences in the associations between sociality and
brain function. Older adults reported feeling less lonely and more empathic, yet scored lower
on a performance-based measure of emotional recognition (RMIE). Negative associations
between loneliness and empathy were observed across the life-span, with more robust asso-
ciations detected for older versus younger adults. Predicted age differences were observed in
the association between loneliness, empathic responding, and RSFC. Brain and behavioral
associations did not differ for loneliness and empathy in younger adults. Positive associations
were observed for both aspects of sociality and RSFC between visual regions and spatially
distributed brain systems. Older adults showed a divergence in RSFC associations between
loneliness and empathy. Higher self-reported loneliness in older adults was associated with
greater RSFC within heteromodal association networks and between attention (dorsal and ven-
tral) and the default and fronto-parietal and limbic networks. In contrast, and consistent with
younger adults, higher self-reported empathy for older adults was associated with greater
visual network connectivity to the cortex. These findings adjudicate previous reports
(Mwilambwe-Tshilobo et al., 2019; Spreng et al., 2020) and reveal that sociality and RSFC
associations differ for young and older adults. Our findings also show that age differences
are specific to loneliness and involve cortical association networks related to internally
directed cognition and socioemotional processing.
Age Differences in RSFC Related to Loneliness
We observed a difference in the relationship between RSFC and loneliness between younger
and older adults. Integration of visual and association networks was related to higher loneli-
ness in young. In contrast, higher loneliness in older adults was marked by lower RSFC of
visual regions and greater intra- and internetwork RSFC among higher order association net-
works. These findings support our hypothesis that age-related differences in the association
between loneliness and brain function reflect a shift from externally to internally oriented pro-
cessing regions, reconciling previous reports (Mwilambwe-Tshilobo et al., 2019; Spreng et al.,
2020). Importantly, no shared pattern relating loneliness to RSFC was observed between age
groups, suggesting a qualitatively different pattern in the neural basis of loneliness across the
life-span. Although we could not test this directly, we suggest that these differences reflect a
shift in the perception and experience of loneliness into older age.
In younger adults, integration of visual and association networks may reflect increased
social perception demands to monitor for threatening social cues or seek new opportunities
for social connection (Cacioppo & Hawkley, 2009). In contrast, for older adults, functional
segregation of the visual network and increased integration within and between higher order
association networks related to loneliness may reflect a shift toward more internally directed
processing (cf. Spreng et al., 2020), consistent with an age-related shift toward prioritizing
socioemotional goals. Instead of searching for new social contacts, older adults have smaller
social networks that prioritize close social connections (Carstensen, 1992). As the pursuit of
new social experiences declines with age, lonely older adults may rely more on reminiscing
about past experiences (Ross & Inagaki, 2023) or mentalizing about future social engagements
(Spreng et al., 2020). Autobiographical recollection and future thinking are robustly related to
the default network and its interactions with other association networks (Andrews-Hanna
et al., 2014; Schacter et al., 2012), which closely converges with the connectivity pattern asso-
ciated with loneliness in older but not younger adults in our study. In the context of previous
Network Neuroscience
514
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
reports (Mwilambwe-Tshilobo et al., 2019; Spreng et al., 2020), our findings suggest that
the experience of loneliness shifts over the adult life-span. However, given that our study was
cross-sectional, an alternative explanation for the age differences found could be due to older
adults experiencing loneliness more chronically than younger adults. Future studies are neces-
sary to examine the experience of loneliness and associated cognitive, social, and neural ante-
cedents and sequelae into older age (Bzdok & Dunbar, 2020, 2022; Spreng & Bzdok, 2021).
Another possible explanation for the age differences in RSFC associated with loneliness is
that healthy aging is characterized by brain network dedifferentiation (Chan et al., 2014;
Malagurski et al., 2020; Setton et al., 2023). Dedifferentiation in older adulthood may, in part,
compensate for the functional reorganization of the aging brain (Reuter-Lorenz & Cappell,
2008), although some aspects of dedifferentiation are also associated with declining brain
health, such as the accumulation of white matter hyperintensities (Kantarovich et al., 2022).
However, unlike healthy aging, our findings in lonely older adults indicate greater within-
network connectivity in higher association networks. We previously reported reduced network
modularity associated with loneliness in younger adults (Mwilambwe-Tshilobo et al., 2019).
While speculative, lonely older adults may compensate for these age and loneliness-related
functional changes by increasing within-network connectivity of higher association networks.
Future research would benefit from disentangling the combined effects of aging and loneliness
on functional brain reorganization.
Finally, our two earlier studies reported divergent associations between loneliness and brain
function for younger and middle-aged adults. Interestingly, we only partially replicated the
findings from our study involving a sizable middle-aged cohort drawn from the UK Biobank
(Spreng et al., 2020). Consistent with Spreng et al. (2020), loneliness in older adults was
related to greater RSFC within the default, fronto-parietal control and limbic networks, and
associations between the default and limbic network. Unlike Spreng et al. (2020), this pattern
extended to greater connectivity within and among ventral and dorsal attention networks in
our older adult cohort, indicating that the impact of loneliness on brain function may continue
to shift beyond midlife into older age. Our findings provide further evidence that the UK
Biobank, representing a large population-based cohort, is a developmentally unique sample
(Kiesow et al., 2021) that may not capture brain and behavioral associations observed in early
or late adulthood. Future research including an adult life-span sample is needed to fully char-
acterize differences in loneliness and brain associations across the broad continuum of adult
human development.
Shared RSFC Pattern Related to Empathy Across Age Groups
We observed age-invariant associations between subdomains of empathy and RSFC, charac-
terized by greater interactions within the visual network and connections with ventral atten-
tion, fronto-parietal control, and default networks. We did not predict this robust age-invariant
association given limited evidence relating visual network functioning to empathy (Katsumi
et al., 2021; Schurz et al., 2021a, 2021b). We speculate that the dependence of empathic
ability on the perception of social cues (Cacioppo & Hawkley, 2009) may underlie the neural
patterns observed here.
Few studies have examined the neural correlates of empathy in aging. Decreased activation
in the insula and anterior/mid-cingulate (core nodes of the ventral attention network; Chen
et al., 2014; Riva et al., 2018) have been associated with affective empathy in older adults.
However, recent work failed to find similar age differences (Ziaei et al., 2021). More robust
age-related brain differences have been observed for cognitive empathy, specifically
Network Neuroscience
515
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
implicating the dorsal medial prefrontal cortex, a key node of the default network related to
social cognition (Beadle & de la Vega, 2019; Moran et al., 2012). Our observations suggest
differing age-related trajectories in empathy and loneliness. The association between empathic
functioning and RSFC was age-invariant. In contrast, the association between loneliness
and RSFC differed with age, as we report here (Figure 3) and in other age cohorts (e.g.,
Mwilambwe-Tshilobo et al., 2019; Spreng et al., 2020).
Differences in RSFC Across Dimensions of Empathic Responding
We included performance-based emotional recognition (RMIE) and self-reported trait empathy
measures, allowing us to examine how these different expressions of empathic responding
relate to RSFC (Ziaei et al., 2021). As revealed in the second LV (Figure 4), these two aspects
of empathic responding are associated with divergent neurocognitive systems, consistent with
previous reports that empathic responding encompasses affective and cognitive processes
(Christov-Moore et al., 2020; Schurz et al., 2021a, 2021b). Specifically, we identified an
age-invariant difference in the relationship between emotion recognition ability using the
RMIE task and self-reported trait empathy measures. Younger and older adults shared a com-
mon RSFC pattern associated with better performance on the RMIE characterized by greater
intra- and internetwork connectivity of association networks. In contrast, self-reported trait
empathy measures in younger adults were associated with greater visual network connectivity
with the rest of the brain. This divergence in intrinsic network connectivity patterns may reflect
functional organizational features of brain networks that enable specialized and flexible
social-cognitive functioning. Recent work on brain network interactions related to social-
cognitive functioning by Schurz et al. (2020) proposes that differences in network segregation
and integration may account for differences in connectivity patterns across theory of mind and
empathy tasks. Specifically, interactions between the default network and attention and fronto-
parietal networks. However, further evidence is needed to disambiguate differences in net-
work interactions underlying task versus trait-based measures of empathic responding.
Behavioral Associations Between Loneliness and Empathy
Finally, while not a central aim of the current study, our behavioral findings confirmed previ-
ous reports of an inverse association between loneliness and empathy (Nakagawa et al., 2015)
observed in younger and older adults (Beadle et al., 2012). While we used a different
self-report measure to assess affective and cognitive aspects of empathy (TEQ vs. Empathy
Quotient), we also observed a negative association across the entire sample. However, when
we examined the relationship between loneliness and subdomains of empathic responding
separately in each age group, reliable associations were only observed for older adults on
self-report measures assessing the affective features of empathy. This finding may reflect shifts
in motivational goals that occur as people age. Socioemotional selectivity theory states that
socioemotional goals become salient for older adults (Carstensen, 2006). This change in goal
hierarchies in later life shifts cognitive resources toward emotional regulation to meet height-
ened socioemotional needs (Mather, 2016; Mather & Carstensen, 2005). Thus, the relationship
between loneliness and empathy may be heightened in older adulthood, reflecting the impor-
tance of maintaining adaptive socioemotional functioning in late-life development. Our results
further underscore this point by demonstrating that aspects of relationship quality are impor-
tant factors to consider when investigating age differences related to loneliness. We show that
loneliness was inversely related to the quality of social relationships, and that controlling for
social support and friendship attenuated the association between loneliness and RSFC in
younger adults, but not older adults.
Network Neuroscience
516
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
Conclusion
Loneliness is a modifiable risk factor associated with various health problems in older adult-
hood (Ong et al., 2016). Further, the experience of loneliness is negatively related to empathic
responding, which is necessary for fostering and maintaining close relationships (Beadle & de
la Vega, 2019; Morelli et al., 2017). Here we examined associations between these essential
dimensions of sociality, brain function, and differences with age. Our findings revealed that the
negative association between loneliness and empathy, observable across the life-span, was
greater in older adults. While longitudinal studies are needed to determine causal associations,
it is possible that experiencing loneliness in later life may precipitate a cascade of adverse
changes in social functioning that could exacerbate feelings of social isolation. We also
identified a pattern of age differences in brain function that is differentially related to loneliness
and empathy in older, but not younger, adults. Extending our previous work in young
(Mwilambwe-Tshilobo et al., 2019) and middle-aged (Spreng et al., 2020) cohorts, the current
results demonstrate that loneliness impacts different neurocognitive systems across the adult
life-span. Early theoretical accounts implicating loneliness in disordered social perception
(Cacioppo & Hawkley, 2009) may not fully capture the experience of loneliness in later life.
Lower motivation to form new social bonds and access to a larger store of lived social expe-
riences may shift the impact of loneliness toward more internally directed cognitive processes
and associated neural networks, as older adults mentalize and reminiscence to fulfill unmet
social desires. Whether and how such a shift may precipitate the adverse cognitive sequelae
associated with loneliness in later life is an important direction for future research (Bzdok &
Dunbar, 2020, 2022; Spreng & Bzdok, 2021).
ACKNOWLEDGMENTS
The authors thank Dr. Bratislav Misic for his insight on the PLS analysis.
SUPPORTING INFORMATION
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00293. All
data from the current report are open access and publicly available (see Spreng et al., 2022, for
data descriptor). Demographic and behavioral data are available within the Open Science
Framework project “Goal-Directed Cognition in Older and Younger Adults” (https://osf.io
/yhzxe/); neuroimaging data are available on OpenNeuro (https://openneuro.org/datasets
/ds003592).
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
AUTHOR CONTRIBUTIONS
Laetitia Mwilambwe-Tshilobo: Conceptualization; Data curation; Formal analysis; Investigation;
Methodology; Visualization; Writing – original draft. Roni Setton: Data curation; Writing – review
& editing. Danilo Bzdok: Funding acquisition; Writing – review & editing. Gary R. Turner: Funding
acquisition; Supervision; Writing – review & editing. R. Nathan Spreng: Conceptualization;
Funding acquisition; Supervision; Writing – review & editing.
FUNDING INFORMATION
R. Nathan Spreng and Danilo Bzdok, National Institutes of Health, Award ID:
R01AG068563A. R. Nathan Spreng, Canadian Institute of Health Research.
Network Neuroscience
517
Functional connectivity related to loneliness and empathy in aging
REFERENCES
Abdellaoui, A., Chen, H.-Y., Willemsen, G., Ehli, E. A., Davies,
G. E., Verweij, K. J. H., Nivard, M. G., de Geus, E. J. C., Boomsma,
D. I., & Cacioppo, J. T. (2019). Associations between loneliness
and personality are mostly driven by a genetic association with
neuroticism. Journal of Personality, 87(2), 386–397. https://doi
.org/10.1111/jopy.12397, PubMed: 29752830
Andrews-Hanna, J. R., Smallwood, J., & Spreng, R. N. (2014). The
default network and self-generated thought: Component pro-
cesses, dynamic control, and clinical relevance. Annals of the
New York Academy of Sciences, 1316(1), 29–52. https://doi.org
/10.1111/nyas.12360, PubMed: 24502540
Bailey, P. E., Brady, B., Ebner, N. C., & Ruffman, T. (2020). Effects of
age on emotion regulation, emotional empathy, and prosocial
behavior. The Journals of Gerontology: Series B, 75(4), 802–810.
https://doi.org/10.1093/geronb/gby084, PubMed: 30016531
Bangee, M., Harris, R. A., Bridges, N., Rotenberg, K. J., & Qualter, P.
(2014). Loneliness and attention to social threat in young adults:
Findings from an eye tracker study. Personality and Individual Dif-
ferences, 63, 16–23. https://doi.org/10.1016/j.paid.2014.01.039
Baron-Cohen, S., Wheelwright, S., Hill, J., Raste, Y., & Plumb, I.
(2001). The “Reading the Mind in the Eyes” test revised version:
A study with normal adults, and adults with Asperger syndrome
or high-functioning autism. The Journal of Child Psychology and
Psychiatry and Allied Disciplines, 42(2), 241–251. https://doi.org
/10.1111/1469-7610.00715, PubMed: 11280420
Beadle, J. N., & de la Vega, C. E. (2019). Impact of aging on
empathy: Review of psychological and neural mechanisms.
Frontiers in Psychiatry, 10, 331. https://doi.org/10.3389/fpsyt
.2019.00331, PubMed: 31244684
Beadle, J. N., Keady, B., Brown, V., Tranel, D., & Paradiso, S.
(2012). Trait empathy as a predictor of individual differences in
perceived loneliness. Psychological Reports, 110(1), 3–15.
https://doi.org/10.2466/07.09.20.PR0.110.1.3-15, PubMed:
22489373
Boss, L., Kang, D.-H., & Branson, S. (2015). Loneliness and cogni-
tive function in the older adult: A systematic review. International
Psychogeriatrics, 27(4), 541–553. https://doi.org/10.1017
/S1041610214002749, PubMed: 25554219
Bzdok, D., & Dunbar R. (2020). The neurobiology of social dis-
tance. Trends in Cognitive Sciences, 24(9), 717–733. https://doi
.org/10.1016/j.tics.2020.05.016, PubMed: 32561254
Bzdok, D., & Dunbar R. (2022). Social isolation and the brain in the
pandemic era. Nature Human Behaviour, 6(10), 1333–1343.
https://doi.org/10.1038/s41562-022-01453-0, PubMed: 36258130
Cacioppo, J. T., & Hawkley, L. C. (2009). Perceived social isolation
and cognition. Trends in Cognitive Sciences, 13(10), 447–454.
https://doi.org/10.1016/j.tics.2009.06.005, PubMed: 19726219
Cacioppo, J. T., Fowler, J. H., & Christakis, N. A. (2009a). Alone in
the crowd: The structure and spread of loneliness in a large social
network. Journal of Personality and Social Psychology, 97(6),
977–991. https://doi.org/10.1037/a0016076, PubMed:
19968414
Cacioppo, J. T., Norris, C. J., Decety, J., Monteleone, G., &
Nusbaum, H. (2009b). In the eye of the beholder: Individual
differences in perceived social isolation predict regional brain
activation to social stimuli. Journal of Cognitive Neuroscience,
21(1), 83–92. https://doi.org/10.1162/jocn.2009.21007,
PubMed: 18476760
Cacioppo, S., Balogh, S., & Cacioppo, J. T. (2015). Implicit attention
to negative social, in contrast to nonsocial, words in the Stroop
task differs between individuals high and low in loneliness:
Evidence from event-related brain microstates. Cortex, 70,
213–233. https://doi.org/10.1016/j.cortex.2015.05.032,
PubMed: 26195152
Cacioppo, S., Capitanio, J. P., & Cacioppo, J. T. (2014). Toward a
neurology of loneliness. Psychological Bulletin, 140(6), 1464–1504.
https://doi.org/10.1037/a0037618, PubMed: 25222636
Carstensen, L. L. (1992). Social and emotional patterns in adult-
hood: Support for socioemotional selectivity theory. Psychology
and Aging, 7(3), 331–338. https://doi.org/10.1037/0882-7974.7
.3.331, PubMed: 1388852
Carstensen, L. L., Isaacowitz, D. M., & Charles, S. T. (1999). Taking
time seriously. A theory of socioemotional selectivity. The Amer-
ican Psychologist, 54(3), 165–181. https://doi.org/10.1037/0003
-066X.54.3.165, PubMed: 10199217
Carstensen, L. L. (2006). The influence of a sense of time on human
development. Science, 312(5782), 1913–1915. https://doi.org/10
.1126/science.1127488, PubMed: 16809530
Chan, M. Y., Park, D. C., Savalia, N. K., Petersen, S. E., & Wig, G. S.
(2014). Decreased segregation of brain systems across the
healthy adult lifespan. Proceedings of the National Academy of
Sciences of the United States of America, 111(46), E4997–E5006.
https://doi.org/10.1073/pnas.1415122111, PubMed: 25368199
Chen, Y. C., Chen, C. C., Decety, J., & Cheng, Y. (2014). Aging is
associated with changes in the neural circuits underlying empa-
thy. Neurobiology of Aging, 35(4), 827–836. https://doi.org/10
.1016/j.neurobiolaging.2013.10.080, PubMed: 24211010
Chong, M., Bhushan, C., Joshi, A. A., Choi, S., Haldar, J. P.,
Shattuck, D. W., Spreng, R. N., & Leahy, R. M. (2017). Individual
parcellation of resting fMRI with a group functional connectivity
prior. NeuroImage, 156, 87–100. https://doi.org/10.1016/j
.neuroimage.2017.04.054, PubMed: 28478226
Christov-Moore, L., Reggente, N., Douglas, P. K., Feusner, J. D., &
Iacoboni, M. (2020). Predicting empathy from resting state brain
connectivity: A multivariate approach. Frontiers in Integrative
Neuroscience, 14, 3. https://doi.org/10.3389/fnint.2020.00003,
PubMed: 32116582
Cohen, S., Doyle, W. J., Skoner, D. P., Rabin, B. S., & Gwaltney,
J. M., Jr. (1997). Social ties and susceptibility to the common
cold. JAMA, 277(24), 1940–1944, https://doi.org/10.1001/jama
.1997.03540480040036, PubMed: 9200634
Cox, C. L., Uddin, L. Q., Di Martino, A., Castellanos, F. X., Milham,
M. P., & Kelly, C. (2012). The balance between feeling and
knowing: Affective and cognitive empathy are reflected in the
brain’s intrinsic functional dynamics. Social Cognitive and Affec-
tive Neuroscience, 7(6), 727–737. https://doi.org/10.1093/scan
/nsr051, PubMed: 21896497
Davis, M. H. (1980). A multidimensional approach to individual
differences in empathy. Catalog of Selected Documents in Psy-
chology, 10, 85.
Denburg, N. L., Tranel, D., & Bechara, A. (2005). The ability to decide
advantageously declines prematurely in some normal older
Network Neuroscience
518
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
persons. Neuropsychologia, 43(7), 1099–1106. https://doi.org/10
.1016/j.neuropsychologia.2004.09.012, PubMed: 15769495
DeYoung, C. G., Quilty, L. C., & Peterson, J. B. (2007). Between
facets and domains: 10 aspects of the Big Five. Journal of Person-
ality and Social Psychology, 93(5), 880–896. https://doi.org/10
.1037/0022-3514.93.5.880, PubMed: 17983306
D’Iorio, A., Garramone, F., Piscopo, F., Baiano, C., Raimo, S., &
Santangelo, G. (2018). Meta-analysis of personality traits in
Alzheimer’s disease: A comparison with healthy subjects. Journal
of Alzheimer’s Disease, 62(2), 773–787. https://doi.org/10.3233
/JAD-170901, PubMed: 29480186
d’Oleire Uquillas, F., Jacobs, H. I. L., Biddle, K. D., Properzi, M.,
Hanseeuw, B., Schultz, A. P., Rentz, D. M., Johnson, K. A.,
Sperling, R. A., & Donovan, N. J. (2018). Regional tau pathology
and loneliness in cognitively normal older adults. Translational
Psychiatry, 8(1), 282. https://doi.org/10.1038/s41398-018-0345-x,
PubMed: 30563962
Donovan, N. J., Okereke, O. I., Vannini, P., Amariglio, R. E., Rentz,
D. M., Marshall, G. A., Johnson, K. A., & Sperling, R. A. (2016).
Association of higher cortical amyloid burden with loneliness
in cognitively normal older adults. JAMA Psychiatry, 73(12),
1230–1237. https://doi.org/10.1001/jamapsychiatry.2016.2657,
PubMed: 27806159
Dunbar, R. I. M., & Shultz, S. (2007). Evolution in the social brain.
Science, 317(5843), 1344–1347. https://doi.org/10.1126/science
.1145463, PubMed: 17823343
Düzel, S., Drewelies, J., Gerstorf, D., Demuth, I., Steinhagen-
Thiessen, E., Lindenberger, U., & Kühn, S. (2019). Structural brain
correlates of loneliness among older adults. Scientific Reports, 9(1),
13569. https://doi.org/10.1038/s41598-019-49888-2, PubMed:
31537846
Efron, B., & Tibshirani, R. (1986). Bootstrap methods for standard
errors, confidence intervals, and other measures of statistical
accuracy. Statistical Science, 1(1), 54–75. https://doi.org/10
.1214/ss/1177013815
Feng, C., Wang, L., Li, T., & Xu, P. (2019). Connectome-based
individualized prediction of loneliness. Social Cognitive and
Affective Neuroscience, 14(4), 353–365. https://doi.org/10.1093
/scan/nsz020, PubMed: 30874805
Finn, E. S., Shen, X., Scheinost, D., Rosenberg, M. D., Huang, J.,
Chun, M. M., Papademetris, X., & Constable, R. T. (2015). Func-
tional connectome fingerprinting: Identifying individuals using
patterns of brain connectivity. Nature Neuroscience, 18(11),
1664–1671. https://doi.org/10.1038/nn.4135, PubMed:
26457551
Fischl, B. (2012). FreeSurfer. NeuroImage, 62(2), 774–781. https://
doi.org/10.1016/j.neuroimage.2012.01.021, PubMed: 22248573
Gershon, R. C., Wagster, M. V., Hendrie, H. C., Fox, N. A., Cook,
K. F., & Nowinski, C. J. (2013). NIH toolbox for assessment of
neurological and behavioral function. Neurology, 80(11 Supple-
ment 3), S2–S6. https://doi.org/10.1212/ WNL.0b013e3182872e5f,
PubMed: 23479538
Greve, D. N., & Fischl, B. (2009). Accurate and robust brain image
alignment using boundary-based registration. NeuroImage, 48(1),
63–72. https://doi.org/10.1016/j.neuroimage.2009.06.060,
PubMed: 19573611
Kanai, R., Bahrami, B., Duchaine, B., Janik, A., Banissy, M. J., & Rees,
G. (2012). Brain structure links loneliness to social perception.
Current Biology, 22(20), 1975–1979. https://doi.org/10.1016/j
.cub.2012.08.045, PubMed: 23041193
Kantarovich, K., Mwilambwe-Tshilobo, L., Fernández-Cabello, S.,
Setton, R., Baracchini, G., Lockrow, A. W., Spreng, R. N., &
Turner, G. R. (2022). White matter lesion load is associated with
lower within- and greater between- network connectivity across
older age. Neurobiology of Aging, 112, 170–180. https://doi.org
/10.1016/j.neurobiolaging.2022.01.005, PubMed: 35219126
Katsumi, Y., Kondo, N., Dolcos, S., Dolcos, F., & Tsukiura, T.
(2021). Intrinsic functional network contributions to the relation-
ship between trait empathy and subjective happiness. Neuro-
Image, 227, 117650. https://doi.org/10.1016/j.neuroimage.2020
.117650, PubMed: 33338612
Kebets, V., Holmes, A. J., Orban, C., Tang, S., Li, J., Sun, N., Kong, R.,
Poldrack, R. A., & Yeo, B. T. T. (2019). Somatosensory-motor dys-
connectivity spans multiple transdiagnostic dimensions of psy-
chopathology. Biological Psychiatry, 86(10), 779–791. https://
doi.org/10.1016/j.biopsych.2019.06.013, PubMed: 31515054
Kiesow, H., Uddin, L. Q., Bernhardt, B. C., Kable, J., & Bzdok, D.
(2021). Dissecting the midlife crisis: Disentangling social, per-
sonality and demographic determinants in social brain anatomy.
Communications Biology, 4(1), 728. https://doi.org/10.1038
/s42003-021-02206-x, PubMed: 34140617
Kong, X., Wei, D., Li, W., Cun, L., Xue, S., Zhang, Q., & Qiu, J.
(2015). Neuroticism and extraversion mediate the association
between loneliness and the dorsolateral prefrontal cortex. Exper-
imental Brain Research, 233(1), 157–164. https://doi.org/10.1007
/s00221-014-4097-4, PubMed: 25234401
Kong, R., Yang, Q., Gordon, E., Xue, A., Yan, X., Orban, C., Zuo,
X. N., Spreng, N., Ge, T., Holmes, A., Eickhoff, S., & Yeo, B. T. T.
(2021). Individual-specific areal-level parcellations improve
functional connectivity prediction of behavior. Cerebral Cortex,
31(10), 4477–4500. https://doi.org/10.1093/cercor/ bhab101,
PubMed: 33942058
Krishnan, A., Williams, L. J., McIntosh, A. R., & Abdi, H. (2011).
Partial least squares (PLS) methods for neuroimaging: A tutorial
and review. NeuroImage, 56(2), 455–475. https://doi.org/10
.1016/j.neuroimage.2010.07.034, PubMed: 20656037
Kundu, P., Inati, S. J., Evans, J. W., Luh, W. M., & Bandettini, P. A.
(2012). Differentiating BOLD and non-BOLD signals in fMRI time
series using multi-echo EPI. NeuroImage, 60(3), 1759–1770.
https://doi.org/10.1016/j.neuroimage.2011.12.028, PubMed:
22209809
Kundu, P., Brenowitz, N. D., Voon, V., Worbe, Y., Vértes, P. E.,
Inati, S. J., Saad, Z. S., Bandettini, P. A., & Bullmore, E. T.
(2013). Integrated strategy for improving functional connectivity
mapping using multiecho fMRI. Proceedings of the National
Academy of Sciences of the United States of America, 110(40),
16187–16192. https://doi.org/10.1073/pnas.1301725110,
PubMed: 24038744
Kundu, P., Voon, V., Balchandani, P., Lombardo, M. V., Poser, B. A.,
& Bandettini, P. A. (2017). Multi-echo fMRI: A review of applica-
tions in fMRI denoising and analysis of BOLD signals. Neuro-
Image, 154, 59–80. https://doi.org/10.1016/j.neuroimage.2017
.03.033, PubMed: 28363836
Lam, J. A., Murray, E. R., Yu, K. E., Ramsey, M., Nguyen, T. T., Mishra,
J., Martis, B., Thomas, M. L., & Lee, E. E. (2021). Neurobiology
of loneliness: A systematic review. Neuropsychopharmacology,
Network Neuroscience
519
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
46(11), 1873–1887. https://doi.org/10.1038/s41386-021-01058-7,
PubMed: 34230607
Layden, E. A., Cacioppo, J. T., Cacioppo, S., Cappa, S. F., Dodich,
A., Falini, A., & Canessa, N. (2017). Perceived social isolation is
associated with altered functional connectivity in neural net-
works associated with tonic alertness and executive control.
NeuroImage, 145(Pt A), 58–73. https://doi.org/10.1016/j
.neuroimage.2016.09.050, PubMed: 27664824
Luhmann, M., & Hawkley, L. C. (2016). Age differences in loneli-
ness from late adolescence to oldest old age. Developmental Psy-
chology, 52(6), 943–959. https://doi.org/10.1037/dev0000117,
PubMed: 27148782
Malagurski, B., Liem, F., Oschwald, J., Mérillat, S., & Jäncke, L.
(2020). Functional dedifferentiation of associative resting state
networks in older adults—A longitudinal study. NeuroImage,
214, 116680. https://doi.org/10.1016/j.neuroimage.2020
.116680, PubMed: 32105885
Mars, R. B., Neubert, F.-X., Noonan, M. P., Sallet, J., Toni, I., &
Rushworth, M. F. S. (2012). On the relationship between the
“default mode network” and the “social brain.” Frontiers in
Human Neuroscience, 6, 189. https://doi.org/10.3389/fnhum
.2012.00189, PubMed: 22737119
Mather, M. (2016). The affective neuroscience of aging. Annual
Review of Psychology, 67(1), 213–238. https://doi.org/10.1146
/annurev-psych-122414-033540, PubMed: 26436717
Mather, M., & Carstensen, L. L. (2005). Aging and motivated cog-
nition: The positivity effect in attention and memory. Trends in
Cognitive Sciences, 9(10), 496–502. https://doi.org/10.1016/j
.tics.2005.08.005, PubMed: 16154382
McIntosh, A. R., & Lobaugh, N. J. (2004). Partial least squares
analysis of neuroimaging data: Applications and advances. Neu-
roImage, 23, S250–S263. https://doi.org/10.1016/j.neuroimage
.2004.07.020, PubMed: 15501095
McIntosh, A. R., & Mišić, B. (2013). Multivariate statistical analyses
for neuroimaging data. Annual Review of Psychology, 64,
499–525. https://doi.org/10.1146/annurev-psych-113011
-143804, PubMed: 22804773
Mirchi, N., Betzel, R. F., Bernhardt, B. C., Dagher, A., & Mišic, B.
(2019). Tracking mood fluctuations with functional network
patterns. Social Cognitive and Affective Neuroscience, 14(1),
47–57. https://doi.org/10.1093/scan/nsy107, PubMed: 30481361
Moran, J. M., Jolly, E., & Mitchell, J. P. (2012). Social-cognitive
deficits in normal aging. Journal of Neuroscience, 32(16),
5553–5561. https://doi.org/10.1523/JNEUROSCI.5511-11.2012,
PubMed: 22514317
Morelli, S. A., Ong, D. C., Makati, R., Jackson, M. O., & Zaki, J.
(2017). Empathy and well-being correlate with centrality in dif-
ferent social networks. Proceedings of the National Academy of
Sciences, 114(37), 9843–9847. https://doi.org/10.1073/pnas
.1702155114, PubMed: 28851835
Mwilambwe-Tshilobo, L., Ge, T., Chong, M., Ferguson, M. A.,
Misic, B., Burrow, A. L., Leahy, R. M., & Spreng, R. N. (2019).
Loneliness and meaning in life are reflected in the intrinsic net-
work architecture of the brain. Social Cognitive and Affective
Neuroscience, 14(4), 423–433. https://doi.org/10.1093/scan
/nsz021, PubMed: 30924854
Nakagawa, S., Takeuchi, H., Taki, Y., Nouchi, R., Sekiguchi, A.,
Kotozaki, Y., Miyauchi, C. M., Iizuka, K., Yokoyama, R., Shinada,
T., Yamamoto, Y., Hanawa, S., Araki, T., Hashizume, H.,
Kunitoki, K., Sassa, Y., & Kawashima, R. (2015). White matter
structures associated with loneliness in young adults. Scientific
Reports, 5, 17001. https://doi.org/10.1038/srep17001, PubMed:
26585372
Oakley, B. F. M., Brewer, R., Bird, G., & Catmur, C. (2016). Theory
of mind is not theory of emotion: A cautionary note on the Read-
ing the Mind in the Eyes Test. Journal of Abnormal Psychology,
125(6), 818–823. https://doi.org/10.1037/abn0000182, PubMed:
27505409
Ong, A. D., Uchino, B. N., & Wethington, E. (2016). Loneliness and
health in older adults: A mini-review and synthesis. Gerontology,
62(4), 443–449. https://doi.org/10.1159/000441651, PubMed:
26539997
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., &
Petersen, S. E. (2012). Spurious but systematic correlations in
functional connectivity MRI networks arise from subject motion.
NeuroImage, 59(3), 2142–2154. https://doi.org/10.1016/j
.neuroimage.2011.10.018, PubMed: 22019881
Power, J. D., Plitt, M., Gotts, S. J., Kundu, P., Voon, V., Bandettini,
P. A., & Martin, A. (2018). Ridding fMRI data of motion-
related influences: Removal of signals with distinct spatial
and physical bases in multiecho data. Proceedings of the
National Academy of Sciences of the United States of America,
11 5 ( 9 ) , E 2 1 0 5 – E 2 11 4 . h t t p s : / / d o i . o r g / 1 0 . 1 0 7 3 / p n a s
.1720985115, PubMed: 29440410
Reuter-Lorenz, P. A., & Cappell, K. A. (2008). Neurocognitive aging
and the compensation hypothesis. Current Directions in Psycho-
logical Science, 17(3), 177–182. https://doi.org/10.1111/j.1467
-8721.2008.00570.x
Riva, F., Tschernegg, M., Chiesa, P. A., Wagner, I. C., Kronbichler,
M., Lamm, C., & Silani, G. (2018). Age-related differences in the
neural correlates of empathy for pleasant and unpleasant touch in
a female sample. Neurobiology of Aging, 65, 7–17. https://doi.org
/10.1016/j.neurobiolaging.2017.12.028, PubMed: 29407469
Ross, L. P., & Inagaki, T. K. (2023). Recalling prior experiences with
a close other can fulfill the need for social connection. Emotion,
23(2), 321–331. https://doi.org/10.1037/emo0001103, PubMed:
35446056
Russell, D. W. (1996). UCLA Loneliness Scale ( Version 3): Reliability,
validity, and factor structure. Journal of Personality Assessment,
66(1), 20–40. https://doi.org/10.1207/s15327752jpa6601_2,
PubMed: 8576833
Salsman, J. M., Butt, Z., Pilkonis, P. A., Cyranowski, J. M., Zill, N.,
Hendrie, H. C., Kupst, M. J., Kelly, M. A., Bode, R. K., Choi,
S. W., Lai, J. S., Griffith, J. W., Stoney, C. M., Brouwers, P., Knox,
S. S., & Cella, D. (2013). Emotion assessment using the NIH
Toolbox. Neurology, 80(11 Suppl 3), S76–S86. https://doi.org
/10.1212/ WNL.0b013e3182872e11, PubMed: 23479549
Schacter, D. L., Addis, D. R., Hassabis, D., Martin, V. C., Spreng,
R. N., & Szpunar, K. K. (2012). The future of memory: Remember-
ing, imagining, and the brain. Neuron, 76(4), 677–694. https://doi
.org/10.1016/j.neuron.2012.11.001, PubMed: 23177955
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X. N.,
Holmes, A. J., Eickhoff, S. B., & Yeo, B. T. T. (2018). Local-global
parcellation of the human cerebral cortex from intrinsic functional
connectivity MRI. Cerebral Cortex, 28(9), 3095–3114. https://doi
.org/10.1093/cercor/bhx179, PubMed: 28981612
Network Neuroscience
520
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity related to loneliness and empathy in aging
Schurz, M., Maliske, L., & Kanske, P. (2020). Cross-network inter-
actions in social cognition: A review of findings on task related
brain activation and connectivity. Cortex, 130, 142–157.
https://doi.org/10.1016/j.cortex.2020.05.006, PubMed:
32653744
Schurz, M., Radua, J., Tholen, M. G., Maliske, L., Margulies, D. S.,
Mars, R. B., Sallet, J., & Kanske, P. (2021a). Toward a hierarchical
model of social cognition: A neuroimaging meta-analysis and
integrative review of empathy and theory of mind. Psychological
Bulletin, 147(3), 293–327. https://doi.org/10.1037/bul0000303,
PubMed: 33151703
Schurz, M., Uddin, L.Q., Kanske, P., Lamm, C., Sallet, J., Bernhardt,
B. C., Mars, R. B., & Bzdok, D. (2021b). Variability in brain struc-
ture and function reflects lack of peer support. Cerebral Cortex,
31(10), 4612–4627. https://doi.org/10.1093/cercor/ bhab109,
PubMed: 33982758
Setton, R., Mwilambwe-Tshilobo, L., Girn, M. Lockrow, A. W.,
Baracchini, G., Hughes, C., Lowe, A. J., Cassidy, B. N., Li, J.,
Luh, W.-M., Bzdok, D., Leahy, R. M., Ge, T., Margulies, D. S.,
Misic, B., Bernhardt, B. C., Stevens, W. D., De Brigard, F.,
Kundu, P., … Spreng, R. N. (2023). Age differences in the func-
tional architecture of the human brain. Cerebral Cortex, 33(1),
114–134. https://doi.org/10.1093/cercor/ bhac056, PubMed:
35231927
Shankar, A., Hamer, M., McMunn, A., & Steptoe, A. (2013).
Social isolation and loneliness: Relationships with cognitive
function during 4 years of follow-up in the English Longitudinal
Study of Ageing. Psychosomatic Medicine, 75(2), 161–170.
https://doi.org/10.1097/ PSY.0b013e31827f09cd, PubMed:
23362501
Shiovitz-Ezra, S., & Ayalon, L. (2012). Use of direct versus indirect
approaches to measure loneliness in later life. Research on Aging,
34(5), 572–591. https://doi.org/10.1177/0164027511423258
Smith S. M. (2002). Fast robust automated brain extraction. Human
Brain Mapping, 17(3), 143–155. https://doi.org/10.1002/ hbm
.10062, PubMed: 12391568
Speck, O., Ernst, T., & Chang, L. (2001). Biexponential modeling of
multigradient-echo MRI data of the brain. Magnetic Resonance in
Medicine, 45(6), 1116–1121. https://doi.org/10.1002/mrm.1147,
PubMed: 11378891
Spreng, R. N., & Bzdok, D. (2021). Loneliness and neurocognitive
aging. Advances in Geriatric Medicine and Research, 3(2),
e210009. https://doi.org/10.20900/agmr20210009, PubMed:
33880462
Spreng, R. N., Dimas, E., Mwilambwe-Tshilobo, L., Dagher, A.,
Koellinger, P., Nave, G., Ong, A., Kernbach, J. M., Wiecki,
T. V., Ge, T., Li, Y., Holmes, A. J., Yeo, B. T. T., Turner, G. R.,
Dunbar, R. I. M., & Bzdok, D. (2020). The default network of
the human brain is associated with perceived social isolation.
Nature Communications, 11(1), 6393. https://doi.org/10.1038
/s41467-020-20039-w, PubMed: 33319780
Spreng, R. N., McKinnon, M. C., Mar, R. A., & Levine, B. (2009).
The Toronto Empathy Questionnaire: Scale development and
initial validation of a factor-analytic solution to multiple empathy
measures. Journal of Personality Assessment, 91(1), 62–71.
https://doi.org/10.1080/00223890802484381, PubMed:
19085285
Spreng, R. N., Setton, R., Alter, U., Cassidy, B. N., Darboh, B.,
DuPre, E., Kantarovich, K., Lockrow, A. W., Mwilambwe-
Tshilobo, L., Luh, W.-M., Kundu, P., & Turner, G. R. (2022).
Neurocognitive aging data release with behavioral, structural
and multi-echo functional MRI measures. Scientific Data, 9(1),
119. https://doi.org/10.1038/s41597-022-01231-7, PubMed:
35351925
Steptoe, A., Shankar, A., Demakakos, P., & Wardle, J. (2013). Social
isolation, loneliness, and all-cause mortality in older men and
women. Proceedings of the National Academy of Sciences of
the United States of America, 110(15), 5797–5801. https://doi
.org/10.1073/pnas.1219686110, PubMed: 23530191
Tilvis, R. S., Laitala, V., Routasalo, P. E., & Pitkälä, K. H. (2011). Suf-
fering from loneliness indicates significant mortality risk of older
people. Journal of Aging Research, 2011, 534781. https://doi.org
/10.4061/2011/534781, PubMed: 21423600
Völlm, B. A., Taylor, A. N. W., Richardson, P., Corcoran, R., Stirling,
J., McKie, S., Deakin, J. F. W., & Elliott, R. (2006). Neuronal cor-
relates of theory of mind and empathy: A functional magnetic
resonance imaging study in a nonverbal task. NeuroImage,
29(1), 90–98. https://doi.org/10.1016/j.neuroimage.2005.07
.022, PubMed: 16122944
Wilson, R. S., Boyle, P. A., James, B. D., Leurgans, S. E., Buchman,
A. S., & Bennett, D. A. (2015). Negative social interactions and
risk of mild cognitive impairment in old age. Neuropsychology,
29(4), 561–570. https://doi.org/10.1037/neu0000154, PubMed:
25495828
Wilson, R. S., Krueger, K. R., Arnold, S. E., Schneider, J. A., Kelly,
J. F., Barnes, L. L., Tang, Y., & Bennett, D. A. (2007). Loneliness
and risk of Alzheimer disease. Archives of General Psychiatry,
64(2), 234–240. https://doi.org/10.1001/archpsyc.64.2.234,
PubMed: 17283291
Wong, N. M. L., Liu, H.-L., Lin, C., Huang, C.-M., Wai, Y.-Y., Lee,
S.-H., & Lee, T. M. C. (2016). Loneliness in late-life depression:
Structural and functional connectivity during affective process-
ing. Psychological Medicine, 46(12), 2485–2499. https://doi.org
/10.1017/S0033291716001033, PubMed: 27328861
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari,
D., Hollinshead, M., Roffman, J. L., Smoller, J. W., Zöllei, L.,
Polimeni, J. R., Fischl, B., Liu, H., & Buckner, R. L. (2011). The
organization of the human cerebral cortex estimated by intrinsic
functional connectivity. Journal of Neurophysiology, 106(3),
1125–1165. https://doi.org/10.1152/jn.00338.2011, PubMed:
21653723
Ziaei, M., Oestreich, L., Reutens, D. C., & Ebner, N. C. (2021). Age-
related differences in negative cognitive empathy but similarities
in positive affective empathy. Brain Structure & Function, 226(6),
1823–1840. https://doi.org/10.1007/s00429-021-02291-y,
PubMed: 34037867
Network Neuroscience
521
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
9
6
2
1
1
8
4
0
9
n
e
n
_
a
_
0
0
2
9
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3