INVESTIGACIÓN
Altered large-scale brain network interactions
associated with HIV infection and error processing
Jessica S. Flannery1
, miguel c.. Riedel2
, Lauren D. Hill-Bowen3
, Ranjita Poudel3
,
Katherine L. Bottenhorn4
, Taylor Salo3
, Angela R. Laird2
,
Raul Gonzalez3, and Matthew T. Sutherland3
1Department of Psychology and Neuroscience, University of North Carolina at Chapel Hill, Chapel Hill, CAROLINA DEL NORTE, EE.UU
2Department of Physics, Florida International University, Miami, Florida, EE.UU
3Department of Psychology, Florida International University, Miami, Florida, EE.UU
4Department of Population and Public Health Sciences, University of Southern California, Los Angeles, California, EE.UU
un acceso abierto
diario
Palabras clave: HIV, Resting-state functional connectivity, Error awareness, Cannabis, Default mode
network, Central executive network, Salience network
ABSTRACTO
Altered activity within and between large-scale brain networks has been implicated across
various neuropsychiatric conditions. Sin embargo, patterns of network dysregulation associated
with human immunodeficiency virus (HIV), and further impacted by cannabis (CB) usar,
remain to be delineated. We examined the impact of HIV and CB on resting-state functional
conectividad (rsFC) between brain networks and associations with error awareness and error-
related network responsivity. Participantes (norte = 106), stratified into four groups (HIV+/CB+, HIV+/
CB−, HIV−/CB+, HIV−/CB−), underwent fMRI scanning while completing a resting-state scan
and a modified Go/NoGo paradigm assessing brain responsivity to errors and explicit error
awareness. We examined separate and interactive effects of HIV and CB on resource allocation
indexes (RAIs), a measure quantifying rsFC strength between the default mode network (DMN),
central executive network (cen), and salience network (SN). We observed reduced RAIs among
HIV+ (vs. HIV−) Participantes, which was driven by increased SN-DMN rsFC. No group
differences were detected for SN-CEN rsFC. Increased SN-DMN rsFC correlated with diminished
error awareness, but not with error-related network responsivity. These outcomes highlight
altered network interactions among participants with HIV and suggest such rsFC dysregulation
may persist during task performance, reflecting an inability to disengage irrelevant mental
operaciones, ultimately hindering error processing.
RESUMEN DEL AUTOR
One systems-level framework proposes that dysregulated brain network interactions may
underlie neurocognitive aspects of addiction, neurodegeneration, and other neuropsychiatric
condiciones. Three specific networks have been highlighted: the default mode network (DMN),
the central executive network (cen), and the salience network (SN). To examine alterations
linked with human immunodeficiency virus (HIV) and cannabis (CB) use across these three
canonical brain networks, we utilized a composite network association index, previously used
to assess drug-related effects on brain networks. This study provides a novel perspective on
dysregulated DMN and SN rsFC among participants living with HIV and delineates potential
implications for error-monitoring behavior.
Citación: Flannery, j. S., Riedel,
METRO. C., Hill-Bowen, l. D., Poudel, r.,
Bottenhorn, k. l., Salo, T., Laird, A. r.,
González, r., & Sutherland, METRO. t.
(2022). Altered large-scale brain
network interactions associated with
HIV infection and error processing.
Neurociencia en red, 6(3), 791–815.
https://doi.org/10.1162/netn_a_00241
DOI:
https://doi.org/10.1162/netn_a_00241
Supporting Information:
https://doi.org/10.1162/netn_a_00241;
https://github.com/ Flanneryg3/ HIVCB
_ProjectCode
Recibió: 25 Octubre 2021
Aceptado: 14 Febrero 2022
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Matthew T. Sutherland
masuther@fiu.edu
Editor de manejo:
Andrew Zalesky
Derechos de autor: © 2022
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
INTRODUCCIÓN
The human immunodeficiency virus (HIV) is a chronic infection that dysregulates the immune
system and enters the central nervous system (Williams et al., 2020). Neuroinflammation due
to HIV, combined with the additive and/or interactive effects of opportunistic infections, aging,
and substance use, contributes to neurocognitive impairments that persist in the antiretroviral
therapy era (Gao et al., 2020; hong & Banks, 2015; Rich et al., 2020). En particular, the impact
of cannabis (CB) use in the context of HIV infection may be especially relevant when consid-
ering neurocognitive function given the prevalence of use among people living with HIV
(PLWH; D’Souza et al., 2012; Montgomery et al., 2019; Pacek et al., 2018; Rizzo et al.,
2018) and frequent reports of CB use to relieve HIV-related symptoms (Harris et al., 2014;
Towe et al., 2020). As neurocognitive impairments among PLWH negatively impact disease
management, employment, and quality of life (Blackstone et al., 2012; Cattie et al., 2012),
there is a need to more fully characterize neurobiological contributors to such symptoms
(Robertson et al., 2020; Williams et al., 2020).
One systems-level model proposes that the neurobiological underpinnings of many neuro-
psychiatric conditions can be conceptualized in terms of dysregulated interactions between
large-scale brain networks (B. menón, 2019; V. menón, 2011). This framework has proved use-
ful for understanding altered brain function associated with addiction (Bednarski et al., 2011;
Sutherland et al., 2012; R. zhang & Volkow, 2019), Alzheimer’s disease (ANUNCIO; Damoiseaux
et al., 2012; Jones et al., 2016), attention deficit hyperactivity disorder (TDAH; Liddle et al.,
2011; Peterson et al., 2009), frontotemporal dementia, mood disorders (Bartova et al., 2015),
and schizophrenia (Zhou y cols., 2016). The three widely recognized brain networks implicated
in this model are the default mode network (DMN), the central executive network (cen), y
the salience network (SN; p.ej., V. menón, 2011; Moradi et al., 2020; Sutherland et al., 2012).
The DMN is primarily engaged during intrinsic thought processes that arise independent of
external stimuli and is thought to be involved with internal and self-referential information pro-
cesando (Buckner & DiNicola, 2019; Di & Biswal, 2014; Moradi et al., 2020), whereas the CEN
is engaged during tasks demanding attention and cognitive control and is thought to be
involved with processing external stimuli (Seeley et al., 2007; Sridharan et al., 2008). DMN
and CEN activity are generally anticorrelated, and the SN is thought to toggle neurocognitive
resources between these two networks, thereby prioritizing processing of the currently most
pertinent information (Chong et al., 2017; V. menón, 2011; menón & Uddin, 2010).
The resource allocation index (RAI) is a previously developed metric quantifying network-
level interactions based on the theorized role of the SN in toggling activity between the DMN
and CEN such that higher RAI values indicate increased synchrony between the SN and CEN
and/or decreased synchrony between the SN and DMN (Lerman et al., 2014). The RAI has
been used to evaluate dysfunction in this relative balance between SN-DMN and SN-CEN
coupling (Choi et al., 2013; Lerman et al., 2014; Reese et al., 2019; j. t. Zhang et al.,
2017). Por ejemplo, the attentional and cognitive deficits characterizing nicotine withdrawal
have been linked to reduced activation in brain regions comprising the CEN and less suppres-
sion of activity in regions comprising the DMN (Falcone et al., 2014; Hahn et al., 2007;
Lerman et al., 2014; Loughead et al., 2010; Tanabe et al., 2011). These observations support
the hypothesis that the SN may allocate attentional resources toward nicotine withdrawal–
related processes (p.ej., craving), thereby biasing activity toward the DMN and away from
the CEN (Sutherland et al., 2012; Sutherland & piedra, 2018; Sutherland et al., 2015). Como
HIV is characterized by progressive cognitive and attentional deficits (Dawes et al., 2008;
Heaton et al., 2010; Mothobi & Brew, 2012; Woods et al., 2009), and has recently been linked to
792
Default mode network (DMN):
Large-scale brain network most
active during internally focused
thought and deactivated when
focused on external tasks.
Central executive network (cen):
Large-scale brain network most
active during externally oriented,
attentionally demanding cognitive
tareas, generally anticorrelated with
DMN activity.
Salience network (SN):
Large-scale brain network thought to
dynamically prioritize internally
versus externally oriented cognition
by toggling relative activity between
the DMN and CEN, respectivamente.
Resource allocation index (RAI):
Previously developed metric
quantifying synchrony between the
DMN and SN relative to that
between the CEN and SN.
Default mode network (DMN)
suppression:
Commonly observed phenomena of
reduced DMN activity when
performing neuroimaging tasks,
where greater activity decreases
scale with increasing task difficulty.
Neurociencia en red
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
Resting-state functional
conectividad (rsFC):
Quantification of synchrony between
activity of two or more brain regions
over a period of “rest” (es decir., una tarea-
free state).
Triple network model of
psychopathy:
Framework describing multiple
neuropsychiatric conditions in terms
of dysregulated interactions between
three large-scale brain networks
(DMN, ECN, SN).
Error awareness:
Ability to explicitly recognize and
acknowledge one’s own mistakes
when performing a task.
diminished error-related suppression in some regions of the DMN (Flannery, Riedel, Salo, Poudel,
et al., 2021), we hypothesized that HIV may be similarly associated with a SN-DMN bias and that
this bias may have implications for certain cognitive functions. Más, recent work has demon-
strated interacting HIV and CB use effects on both task-based activity (Meade et al., 2019) y
resting-state functional connectivity (rsFC) of a major hub of the SN, the insula (Flannery, Riedel,
Salo, Hill-Bowen, et al., 2021), and abnormal functioning in regions comprising the CEN has
been linked to executive functioning deficits among PLWH (Castelo et al., 2006; Ernst et al.,
2003; Ipser et al., 2015). If the SN biases neurocognitive resources toward internal processes asso-
ciated with DMN function, some salient task-related events may go undetected. En efecto, evidencia
indicates that reduced RAIs correlate with diminished task performance and reduced task-
induced suppression of DMN regions suggesting that alterations in rsFC, as indexed by the
RAI, are linked with alterations in task-based performance and brain activity (Lerman et al., 2014).
In line with the triple network model (B. menón, 2019; V. menón, 2011), accumulating
neuroimaging evidence links neuropsychiatric conditions not only with altered network-level
rsFC (Alexopoulos et al., 2012; Bartova et al., 2015; Bednarski et al., 2011; Bonavita et al.,
2017; Chang et al., 2014; Gauffin et al., 2013; Høgestøl et al., 2019; Lee et al., 2017; Liddle
et al., 2011; Liu et al., 2018; Oyegbile et al., 2019; Peterson et al., 2009; Schilbach et al.,
2016; Sutherland et al., 2012; Verfaillie et al., 2018; k. Wang y cols., 2019; Y. Wang y cols., 2013;
Whitfield-Gabrieli & Vado, 2012; Wu et al., 2011; Yin et al., 2016; R. zhang & Volkow, 2019; zhou
et al., 2016), but also with altered task-based network activity (Bartova et al., 2015; Bednarski
et al., 2011; Liddle et al., 2011; Peterson et al., 2009). While small, albeit consistent, changes in
brain network interactions distinguish task from resting states, task-evoked activity is closely
related to resting-state organization (Cole y col., 2014; Smith et al., 2009), suggesting an intrinsic
architecture of functional brain organization. Tal como, examining both resting-state network
interactions and task-induced network activity may facilitate a more complete understanding
of neurobiological contributors to cognitive alterations among neuropsychiatric conditions. En
particular, lack of task-induced DMN suppression may represent a common endophenotype
across various conditions (Bartova et al., 2015; Bednarski et al., 2011; Gauffin et al., 2013;
Liddle et al., 2011; Oyegbile et al., 2019; Peterson et al., 2009; Sutherland et al., 2012;
Whitfield-Gabrieli & Vado, 2012; R. zhang & Volkow, 2019; Zhou y cols., 2016); sin embargo,
altered DMN suppression has rarely been examined among PLWH (Flannery, Riedel, Salo,
Poudel, et al., 2021). As DMN suppression may facilitate task attention (Hinds et al., 2013)
and detection of salient stimuli (singh & Fawcett, 2008), insufficient suppression likely con-
tributes to errors and/or a lack of error recognition. Proper error recognition is vital for every-
day functioning, as it facilitates behavioral adaptation to minimize future negative outcomes.
Indicative of metacognitive difficulties, PLWH often underreport their cognitive failures when
compared with objective behavioral measures (Bassel et al., 2002; Hinkin et al., 1996; Van
Gorp et al., 1991; Vance et al., 2008), suggesting compromised error recognition. Además,
CB use may exacerbate error processing deficits among PLWH, as evidence has also demon-
strated diminished error awareness among chronic CB users (Hester et al., 2009). Given that
poor error awareness may impact disease management (p.ej., taking medications) y cada-
day functioning, we sought to clarify the interrelations between network-level rsFC interac-
ciones, error awareness, and error-related brain activity among PLWH.
MÉTODOS
Summary
We first examined the independent and combined effects of HIV and CB on rsFC utilizing the
RAI as a measure of relative SN-DMN and SN-CEN interactions. Segundo, to delineate the
Neurociencia en red
793
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
behavioral implications linked with altered rsFC, we examined relations between rsFC and a
task-based behavioral measure of error awareness. Tercero, to understand the relation between
resting-state and task-based network functioning, we examined relations between rsFC and
error-related brain activity during a Go/ NoGo task variant. Regarding group effects, nosotros
expected to observe reduced RAI values among PLWH and CB users when compared with
controls indicative of altered rsFC. Regarding behavioral and brain activity implications, nosotros
expected rsFC alterations to be linked with decreased error awareness and correlate with error-
related brain activity.
Participantes
A sample of 106 participants was stratified into four groups based on HIV serostatus and CB
use history (co-occurring: HIV+/CB+, norte = 32; HIV-only: HIV+/CB−, norte = 28; CB-only: HIV−/
CB+, norte = 24; controls: HIV−/CB−, norte = 22). Demographic, descriptive, and fMRI data from
this sample are also reported elsewhere (Flannery, Riedel, Salo, Hill-Bowen, et al., 2021;
Flannery, Riedel, Salo, Poudel, et al., 2021). Participants were recruited from community-based
organizations providing health-care services throughout Miami-Dade County. Participantes
were 18–60 years old to minimize the presence of other chronic conditions (p.ej., hypertension,
diabetes), as well as the potential interactive effect of HIV and aging on neurocognition
(Morgan et al., 2012; Saloner et al., 2019; Seider et al., 2016; Valcour et al., 2011; Wendelken
& Valcour, 2012). Additional exclusionary criteria included the following: current hepatitis C
infection, English nonfluency or illiteracy, less than an eighth-grade education level, severe
learning disability, significant neurological conditions (p.ej., cerebrovascular issues, cerebro
tumor, brain lymphoma, seizures, multiple sclerosis), severe head trauma with loss of
consciousness >30 min, severe mental illness with psychotic or paranoid symptoms, or MRI
contraindications.
PLWH in this study were diagnosed with HIV 9.3 ± 8.9 (mean ± SD) years prior to assess-
ment and had no history of opportunistic infections affecting the central nervous system, y
the majority (94.3%) were taking antiretroviral medications. All CB-using participants reported
a history of regular use (operationalized as at least once per week for three straight months)
and used at least 20 times in the past year. CB-non-using participants met the following cri-
teria: no history of CB dependency, no CB use in the past month, and a negative urine THC
pantalla. Past use of and dependence on other substances, including alcohol, nicotina, cocaína,
amphetamines, benzodiazepines, or opioids was permitted across groups to provide a more
representative and generalizable sample. Sin embargo, participants were excluded if meeting cri-
teria for current substance dependence (except CB and nicotine) as assessed via the substance
use module of the Diagnostic and Statistical Manual of Mental Disorders (DSM)-5 Structured
Clinical Interview (Primero, 2014).
Procedures
Study procedures were reviewed and approved by the Institutional Review Board of Florida
International University. Following informed consent, we collected blood, behavioral, self-
informe, and MRI data across two study visits on different days. Participants were instructed
to refrain from any substance use (including CB use among CB+ participants) para 24 hr before
study visits to minimize acute pharmacological effects. Upon arrival at both visits, Participantes
completed substance use screening including urine toxicology (Drug Check Cup, NXStep) y
breathalyzer testing (AlcoMate Premium Breathalyzer). During the first visit, blood specimens
were collected, and participants completed a battery of behavioral tests and self-report ques-
tionnaires. Among PLWH, blood samples were used to quantify HIV disease severity (HIV-1
Neurociencia en red
794
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
viral load), immune function (CD4+ T-cell count, lymphocyte subset counts, total white blood
cell count). The second visit occurred less than one month after the first and participants com-
pleted a 1-hr MRI scanning session and additional self-reports. Participants were compensated
at the end of each visit.
MRI Data Acquisition
MRI data were collected on a GE Healthcare Signa MR750, 3-Tesla scanner with 32-channel
head coil. T1-weighted structural images were obtained using a magnetization-prepared rapid
gradient-echo (MPRAGE) secuencia (repetition time [TR] = 2,500 EM; tiempo de eco [EL] = 3.7 EM;
flip angle [FA] = 12°; voxel size = 1 mm3). An 8-min resting-state scan with eyes closed was
collected with 42 slices (3.4 mm thick) obtained in the axial plane using a T2*-weighted,
single-shot, gradient-echo, echo-planar imaging (EPI) sequence sensitive to blood oxygen
level–dependent (BOLD) efectos (245 volumes, TR = 2,000 EM, TE = 30 EM, FA = 75°, campo
of view = 220 × 220 mm, 64 × 64 matrix, voxel size = 3.44 × 3.44 × 3.40 mm). These same
EPI parameters were also used to collect six functional runs (169 volumes/run) while partici-
pants completed a Go/NoGo motor inhibition paradigm called the error awareness task (EAT;
Hester et al., 2005; Hester et al., 2012; Hester et al., 2009; Hester et al., 2007). In the EAT,
participants committed NoGo-errors (es decir., incorrectly pressed a button following a NoGo cue)
of which they were either aware or unaware. Participants subsequently indicated error aware-
ness by pressing an error signaling button on the trial following the error. The EAT allows for
assessment of distinct brain activity linked with cognitive failures (es decir., NoGo-errors) y
explicit error awareness. To achieve enough successful and unsuccessful NoGo trials for a
sufficiently powered study, task difficulty was individually and dynamically adapted to main-
tain participants’ average NoGo-error rate between 45% y 50%. Participants performed this
task during the four runs preceding and two runs following the resting-state scan. While EAT-
associated brain activity has been reported elsewhere (Flannery, Riedel, Salo, Poudel, et al.,
2021), here we focused on the EAT’s behavioral measure of error awareness and examined
network responsivity to task errors.
Error awareness task (EAT):
Computerized paradigm assessing
error awareness and error-related
brain responsivity in which
participants try to inhibit button-press
responses and acknowledge their
errores.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
MRI Data Processing
Resting-state functional MRI (rs-fMRI) data were first denoised using dwidenoise (MRtrix3;
Adhikari et al., 2018; Cordero-Grande et al., 2019; Tournier et al., 2019; Veraart, Fieremans,
et al., 2016; Veraart, Nóvikov, et al., 2016), which utilizes Marchenko-Pastur principal com-
ponent analysis (MPPCA) to estimate and remove Gaussian thermal noise from MRI data,
including fMRI data (Adhikari et al., 2019), based on random matrix theory. The data were
organized in BIDS format and additional preprocessing was performed with FMRIPrep
v1.5.0 (Esteban et al., 2018), a Nipype-based tool (Gorgolewski et al., 2011) often employing
Nilearn (Abraham et al., 2014). T1-weighted structural volumes were corrected for intensity
nonuniformity (N4BiasFieldCorrection v2.1.0; Tustison et al., 2010) and skull-stripped
(antsBrainExtraction.sh v2.1.0). Nonlinear registration (ANTs v2.1.0) was performed to spa-
tially normalize T1-weighted volumes to the ICBM-152 asymmetrical template v2009c (fonov
et al., 2009). Functional data were slice-time corrected to the middle of each TR using 3dTshift
(AFNI v16.2.07; Cox, 1996) and motion corrected using MCFLIRT (FSL v5.0.9; Jenkinson et al.,
2002). Boundary-based registration (bbregister, FreeSurfer v6.0.1) was used to coregister func-
tional images to corresponding T1-weighted volumes (2 × 2 × 2 mm isotropic voxels; Greve &
pescado, 2009) con 9 degrees of freedom. Lanczos interpolation (antsApplyTransforms ANTs
v2.1.0) concatenated all motion-correction transformations (functional-to-anatomical,
anatomical-to-template) and applied them in a single step. Physiological noise regressors were
Neurociencia en red
795
Large-scale brain network interactions associated with HIV infection
calculated applying aCompCor (Behzadi et al., 2007). Específicamente, cerebral spinal fluid (CSF)
and white matter ( W.M.) masks were calculated in T1w space, within a mask excluding signal
with cortical origin. Three aCompCor principal components were then calculated for both the
CSF and the WM masks. Frame-wise displacement (FD; Power et al., 2013) was also calcu-
lated for each functional run using the Nipype implementation.
3dTproject (AFNI) was used to perform simultaneous nuisance regression and bandpass
filtering. Nuisance regressors included the six aCompCor components (3 CSF, 3 W.M.;
Muschelli et al., 2014), the six motion parameters, their derivatives, and TRs acquired during
MRI stabilization (non-steady state) as determined by FMRIPrep. A 0.01 a 0.1 Hz bandpass
filter was applied and TRs with FD greater than 0.35 mm were censored along with time points
immediately preceding and following. Not all participants could be further processed because
of temporal degrees of freedom violations in the denoising procedure (caused by nuisance
regressors outnumbering observations), Resultando en 93 remaining participants (co-occurring:
HIV+/CB+, norte = 28; HIV-only: HIV+/CB−, norte = 27; CB-only: HIV−/CB+, norte = 21; controls:
HIV−/CB−, norte = 17). An average of 6.2 ± 6.0% of volumes were excluded from each partici-
pant’s resting-state scan (Table S1 in the Supporting Information). Groups did not significantly
differ in the number of censored volumes (HIV: F[1, 92] = 0.7, pag = 0.4; CB: F[1, 92] = 1.0, pag =
0.3; HIV × CB: F[1, 92] = 0.1, pag = 0.8) or in mean FD (HIV: F[1, 92] = 0.04, pag = 0.9; CB: F[1,
92] = 0.81, pag = 0.4; HIV × CB: F[1, 92] = 0.001, pag = 0.98). As motion is known to influence
functional connectivity measures (Burgess et al., 2016; Power et al., 2012), mean FD was
included as a covariate in group-level rsFC assessments.
Time series were then standardized (shifted to a zero mean and scaled to a unit variance)
and averaged across voxels within four separate network masks (Nilearn, NiftiLabelsMasker.
fit_transform). These network masks were defined with the Functional Imaging in Neuropsy-
chiatric Disorders (FIND) atlas, which includes masks for an anterior salience network (SN), a
dorsal default mode network (DMN), a left executive control network (labeled here as the
central executive network; L.CEN), and a right executive control network (R.CEN) (Altmann
et al., 2015; Shirer et al., 2012). Networks in this functional atlas were identified by applying
independent component analysis (ICA; MELODIC, FSL) to resting-state data and visually iden-
tifying 14 canonical intrinsic functional connectivity networks based on prior work out of the
30 generated; the creation of this atlas is described in detail elsewhere (Shirer et al., 2012).
While we did not have specific hypotheses regarding CEN laterality, we calculated separate
RAIs for the left and right hemisphere consistent with prior work (Lerman et al., 2014; Reese
et al., 2019; j. t. Zhang et al., 2017). We note that the RAI metric in these prior studies was
calculated using network masks derived via an ICA-based (as opposed to an atlas-based)
acercarse (Lerman et al., 2014; Moradi et al., 2020). Tal como, we also conducted ancillary
analyses using an ICA-based approach to define the network masks of interest for calculating
the RAI metrics (see Supplemental Text and Figures S1–S2 in the Supporting Information for
methodological details and results).
Correlation coefficients between the four networks’ average time series were computed
for each participant (Nilearn, ConnectivityMeasure.fit_transform) and used as a measure of
functional coupling between networks. We then computed RAI values to quantify network
interactions (Lerman et al., 2014) based on the hypothesized role of the SN toggling
resources between the CEN and DMN. Específicamente, the RAI metric integrates a positive
SN-CEN correlation and a negative SN-DMN correlation such that higher RAI values indi-
cate either positive synchronization of SN with CEN and/or negative synchronization of SN
with DMN (Lerman et al., 2014). As done previously (Lerman et al., 2014), we calculated
the RAI by first applying Fisher’s transform to correlation coefficients (CC ) between the SN
Neurociencia en red
796
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
and the R.CEN (CC SN−R. ECN), L.CEN (CC SN−L. ECN), and DMN (CC SN−DMN) using formula 1.
We then computed the RAI for the left and right CEN using formula 2 and formula 3,
respectivamente.
Formula 1.
Formula 2.
Formula 3.
(cid:1)
f CCð
Þ ¼ 0:5 (cid:2) ln
(cid:3)
1 þ CC
d
d
1 − CC
Þ
Þ
(cid:4)
l:RAI ¼ f CC SN−L:cen
(cid:5)
(cid:4)
− f CC SN−DMN
(cid:5)
(cid:4)
R:RAI ¼ f CC SN−R:cen
(cid:5)
(cid:4)
− f CC SN−DMN
(cid:5)
Network-Level rsFC: Group Effects
To assess HIV and CB main and interactive effects on RAI values, we performed HIV × CB
general linear models (GLMs) including age, sexo, mean FD, and whether the participant
was a current cigarette smoker (NIC status; defined as smoking at least eight times in the last
mes) as covariates. We then performed follow-up analyses examining whether group differ-
ences in RAI values were driven by altered SN-CEN and/or SN-DMN rsFC. Específicamente, nosotros
performed HIV × CB GLMs on participants’ standardized SN-L.CEN, SN-R.CEN, and SN-
DMN correlation coefficients (z-scores) while controlling for the above covariates.
Network-Level rsFC: Relation With Error Awareness Behavior
To link rsFC and behavior measures, we then considered relations between RAI values, SN-
CEN rsFC, SN-DMN rsFC, and a behavioral measure of explicit error awareness from the EAT.
Participants who did not meet the task performance criterion (>50% Go-errors) were excluded,
resulting in a sample of 103 participants that had viable task data and a sample of 86 partícipe-
ipants with both viable task-based and resting-state fMRI data. We first assessed group effects
on error awareness (es decir., frequency of unaware errors) by performing an HIV × CB GLM with
edad, sexo, and NIC status as covariates. One error awareness outlier was removed from all anal-
yses including this variable. As the error awareness variable was positively skewed and
included zero values, it was log10 transformed and a constant was added (log10[X + 1]) for this
análisis. We then considered relations between RAI values, network-level (SN-CEN, SN-
DMN) rsFC strength, and error awareness (norte = 92). As the frequency of unaware errors variable
also did not meet assumptions required for linear regression (Atkins & Gallop, 2007), nosotros
employed a negative binomial model (R, v.4.0.2) including age, sexo, mean FD, and NIC status
as covariates. A negative binomial model was selected over a zero-inflated or Poisson model
as the test of dispersion indicated overdispersion (pag < 0.00013) (Atkins & Gallop, 2007).
Network-Level rsFC: Relation With Error-Related Brain Activity
To link resting-state and task-related brain activity, we then considered the relations between
SN-CEN or SN-DMN rsFC and error-related activity during the EAT. First, we assessed whether
network-level responsivity to EAT errors demonstrated HIV × CB effects and/or relationships
with error awareness (paralleling the rsFC analyses above). To do so, the six EAT runs were
preprocessed with FMRIPrep (reported in more detail elsewhere; see Flannery, Riedel, Salo,
Network Neuroscience
797
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
n
e
n
_
a
_
0
0
2
4
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
Hill-Bowen, et al., 2021). Time series were scaled to the voxel-wise mean (3dcalc), thereby
allowing regression (β) coefficients to be interpreted as an approximation of percentage BOLD
signal change (% BOLD Δ; Chen et al., 2017) from the implicit baseline. Data were entered
into subject-level GLMs (3dDeconvolve, 3dREMLfit) that concatenated runs and modeled nui-
sance regressors (i.e., six motion-correction parameters and fourth-order polynomials capturing
residual head motion and baseline trends in the BOLD signal) and three task-related regressors
(NoGo-correct, NoGo-error, and Go-error) as impulse functions time-locked to stimulus onset
and convolved with a hemodynamic response (gamma) function. As such, activity associated
with task events represents activity over and above that of the ongoing Go trial period, similar to
prior work (Hester et al., 2005; Hester et al., 2012; Hester et al., 2009). Average β coefficients
associated with NoGo-error task events were extracted for each participant by averaging across
all nonzero voxels within the four network masks.
To assess group effects on the error-related responsivity, we conducted HIV × CB GLMs on
averaged NoGo-error β coefficients within each network of interest (DMN, SN, R.CEN, L.CEN),
among participants with viable task data (n = 103). Age, sex, and NIC status were included as
covariates. Second, we considered whether error-related network activity correlated with error
awareness by again employing a negative binomial model among all participants with both via-
ble task and rest data (n = 85, one outlier was removed), while controlling for the same covariates
listed above. Finally, to directly link rsFC and task-related brain activity, we conducted partial
Pearson’s correlations between rsFC values and error-related network activity controlling for
mean FD during rest.
RESULTS
Group Characteristics
Demographic and drug use characteristics of this sample have been reported elsewhere
(Flannery, Riedel, Salo, Hill-Bowen, et al., 2021), and are summarized below. Groups did
not differ in terms of age, education, race, ethnicity (Table 1; p > 0.3), or history of major
depressive episodes (Table S2 in the Supporting Information; p > 0.3). Sin embargo, the HIV+
groups included a higher percentage of self-reported males (80% masculino) than did the HIV−
grupos (55.3% masculino; pag = 0.006), consistent with national estimates regarding the sex distri-
bution (81% masculino) of new HIV diagnoses (Centers for Disease Control and Prevention,
2020). This difference was driven by the female/male composition among the CB+ groups
(HIV+/CB+ vs. HIV−/CB+: χ2[1, 92] = 6.6, pag = 0.014), but not the CB− groups (HIV+/CB−
vs. HIV−/CB−: χ2[1, 92] = 3.3, pag = 0.1). Self-reported sex was included as a covariate in all
group-level analyses. Of the PLWH in the study (norte = 54), 64.8% had an undetectable viral
load (es decir., <50 mRNA viral copies> 0.1), and groups were largely matched on other drug use characteristics includ-
ing past dependence (Tables S3–S5 in the Supporting Information); sin embargo, CB-using
groups reported more past month nicotine use (F[1, 92] = 9.7, pag = 0.002). De este modo, si
a participant was a current cigarette smoker (NIC) was included as a covariate in all group-
level analyses.
Neurociencia en red
798
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
t
norte
mi
w
oh
r
k
norte
mi
tu
r
oh
s
C
mi
norte
C
mi
i
Participant demographic, cannabis, HIV disease characteristics, and other drug use. Data are expressed as either mean (standard deviation) or frequency
Mesa 1.
across all participants or within specific groups. Drug use is the self-reported number of times using each drug in the given timeframe (es decir., past month, lifetime). HIV-1
viral load was assessed via the Abbott RealTime HIV-1 assay. Group effects were assessed via either an HIV × CB ANOVA or, for categorical variables, via chi-square
pruebas (one comparing HIV+ vs. HIV− groups and one comparing CB+ vs. CB− groups). AA: African American, C: Caucasian, A: asiático, >1: more than one race.
☨Independent sample t test between CB+ groups. δAll group effects in section assessed with independent sample t test between HIV+ groups, or for categorical variables,
a chi-square test. ξEstimator of general cognitive impairment: t score < 40 for both: total recall on the Hopkins Verbal Learning Test–Revised (HVLT-R; Benedict et al.,
1998; Brandt, 1991) and symbol search scores on the Wechsler Adult Intelligence Scale–Fourth edition ( WAIS-IV; Wechsler, 2008), or t score < 35 for either test (Carey
et al., 2004).
All participants
n = 93
HIV+/CB+
n = 28
HIV+/CB−
n = 26
HIV−/CB+
n = 22
HIV−/CB−
n = 17
Group effects (p)
HIV × CB
HIV
CB
Demographic
Age
34.8 (10.3)
Education (years)
13.8 (2.3)
Male, female
AA, C, >1
65, 28
48, 41, 4
Hispanic/Latinx
35
Cannabis use
Age regular use
19.6 (6.7)
Years regular use
14.8 (11.5)
Past month (veces)
12.8 (14.0)
33.3 (7.4)
13.8 (2.0)
26, 2
14, 14, 0
10
21.1 (6.5)
12.1 (8.9)
23.4 (9.8)
37.5 (13.2)
32.8 (10.1)
35.8 (9.3)
0.7
0.4
13.8 (2.7)
14.4 (1.2)
14, 8
7, 10
11, 10, 1
9, 7, 1
8
6
−
−
18.2 (7.1)
15.5 (12.1)
−
−
24.4 (10.1)
0
13.6 (2.8)
18, 8
14, 10, 2
9
0
Lifetime (veces)
2,141 (3,112)
3,556 (2,969)
282 (1,024)
4,182 (3,852)
7.8 (23)
0.4
HIV disease characteristics
δ
Years since HIV
diagnosis
9.3 (8.9)
7.8 (6.9)
10.8 (10.6)
n with undetectable
35
viral load
(<50 copies> 0.3). We also performed follow-up exploratory GLMs,
among HIV+ participants (norte = 54), assessing relationships between RAI values, rsFC, y
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
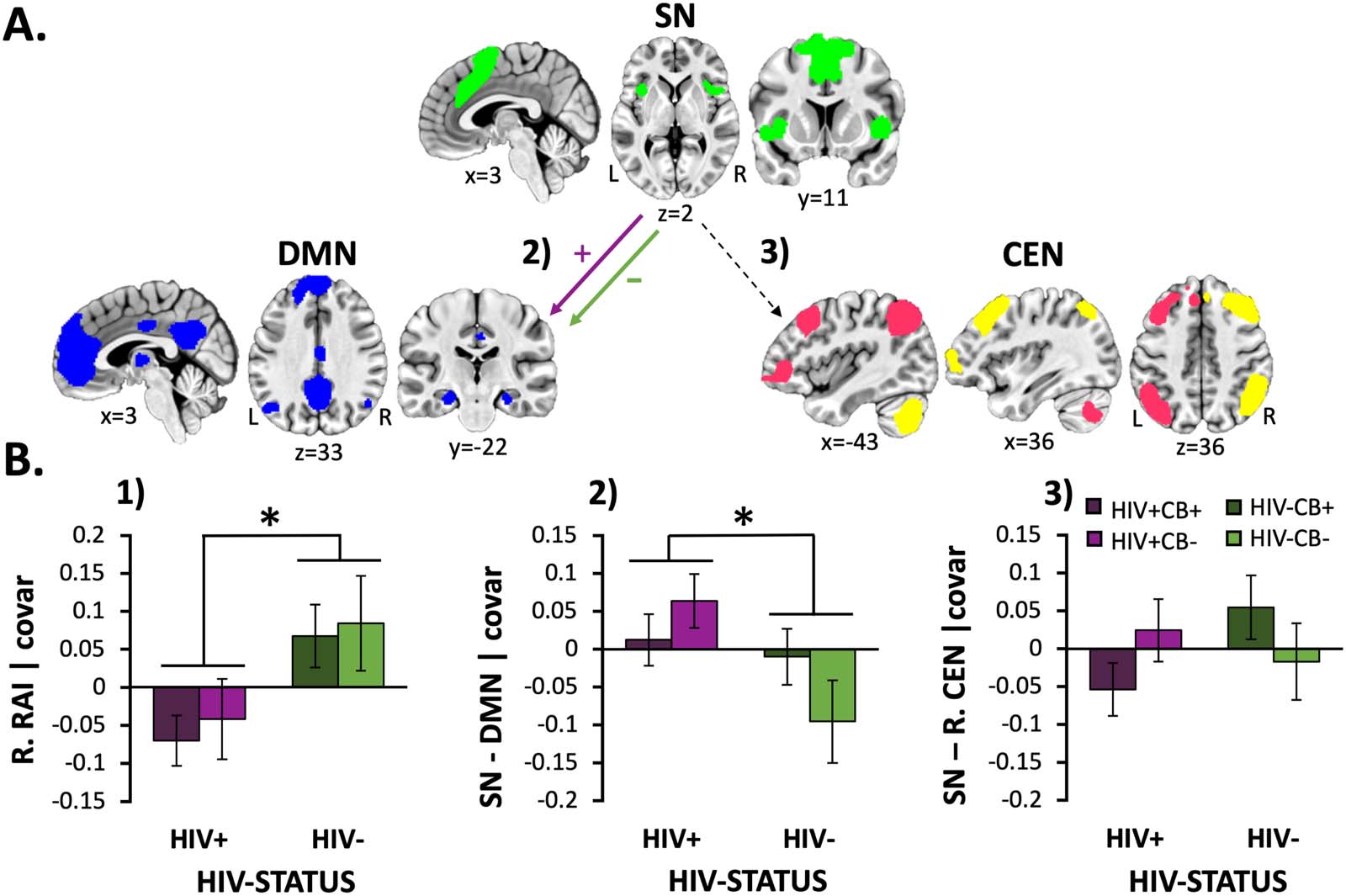
Cifra 1. HIV-associated effects on network-level resource allocation index and rsFC (SN-DMN and SN-CEN) métrica. (A) Schematic of
network-level resting-state functional connectivity (rsFC) differences between HIV+ and HIV− participants. People living with HIV (PLWH)
showed increased rsFC (purple [+] arrow) between the salience network (SN; verde) and the default mode network (DMN; azul) relative to that
from HIV− participants (verde [−] arrow). The gray dashed arrow indicates nonsignificant group differences when considering rsFC between
the SN and central executive network (cen; pink = L.CEN, yellow = R.CEN). (B.1) PLWH (vs. HIV− participants) showed reduced resource
allocation index (RAI) values when considering both the right (pag = 0.008) and left hemisphere (data not displayed, F[1, 92] = 4.3, pag = 0.042).
(B.2) HIV-associated RAI reductions were driven by increased SN-DMN rsFC among PLWH (vs. HIV− participants; pag = 0.027). (B.3) Sobre el
other hand, significant group differences were not detected when considering SN-R.CEN or SN-L.CEN (data not displayed) rsFC values (p >
0.3), which were the second aspect contributing to the composite RAI value. Unstandardized residuals are plotted after regressing effects of
edad, sexo, mean FD, and NIC status. Error bars = standard error of the mean.
Neurociencia en red
801
Large-scale brain network interactions associated with HIV infection
HIV disease characteristics (viral load, duration since diagnosis, CD4+ T-cell count, CD8+
T-cell count, total T-lymphocytes, total white blood cell count), when controlling for CB
group membership, edad, sexo, mean FD, and NIC status. We did not observe any significant
relationships between network rsFC measures and HIV disease characteristics (p > 0.07).
En general, similar outcomes and interpretations were obtained when employing an ICA-based
approach to define the four network masks of interest and reassessing rsFC metrics across
grupos; sin embargo, PLWH displayed significantly reduced rsFC between the SN and L.CEN com-
pared with HIV− participants (F[1, 92] = 4.2, pag = 0.04), while this same effect was not observed
using the atlas-based approach (Figures S1 and S2 in the Supporting Information). As this incon-
sistency may be due to the relatively more bilateral nature of the ICA-based L.CEN network
mask when compared with the atlas-based L.CEN mask, we performed exploratory follow-
up analyses assessing group differences in SN-CEN rsFC when merging both atlas-based
CEN masks to create a bilateral CEN mask. Sin embargo, we did not find any significant group
effects on SN-CEN rsFC when utilizing this bilateral CEN mask (p > 0.1).
Network-Level rsFC: Relation With Error Awareness
When considering a behavioral measure of error awareness during the modified Go/NoGo
tarea, we observed a significant HIV × CB interaction (F[1, 101] = 5.9, pag = 0.017, ηp
2 =
0.059) when controlling for covariates (Figura 2A). Específicamente, visual inspection and
follow-up t tests indicated that both the HIV-only (pag = 0.041; HIV+/CB−) and CB-only (pag =
0.020; HIV−/CB+) groups committed more unaware errors, compared with controls (HIV−/
CB−), whereas the co-occurring (HIV+/CB+) group had fewer unaware errors more similar to
that of controls (pag = 0.767). En tono rimbombante, negative binomial models examining relations
between network-level rsFC and error awareness indicated that higher SN-DMN rsFC was
linked with more unaware errors (es decir., reduced error awareness; Figura 2B; b = 1.7 [0.8], z =
2.1, pag = 0.037). Neither right RAI, left RAI (Figure S3A in the Supporting Information) nor
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
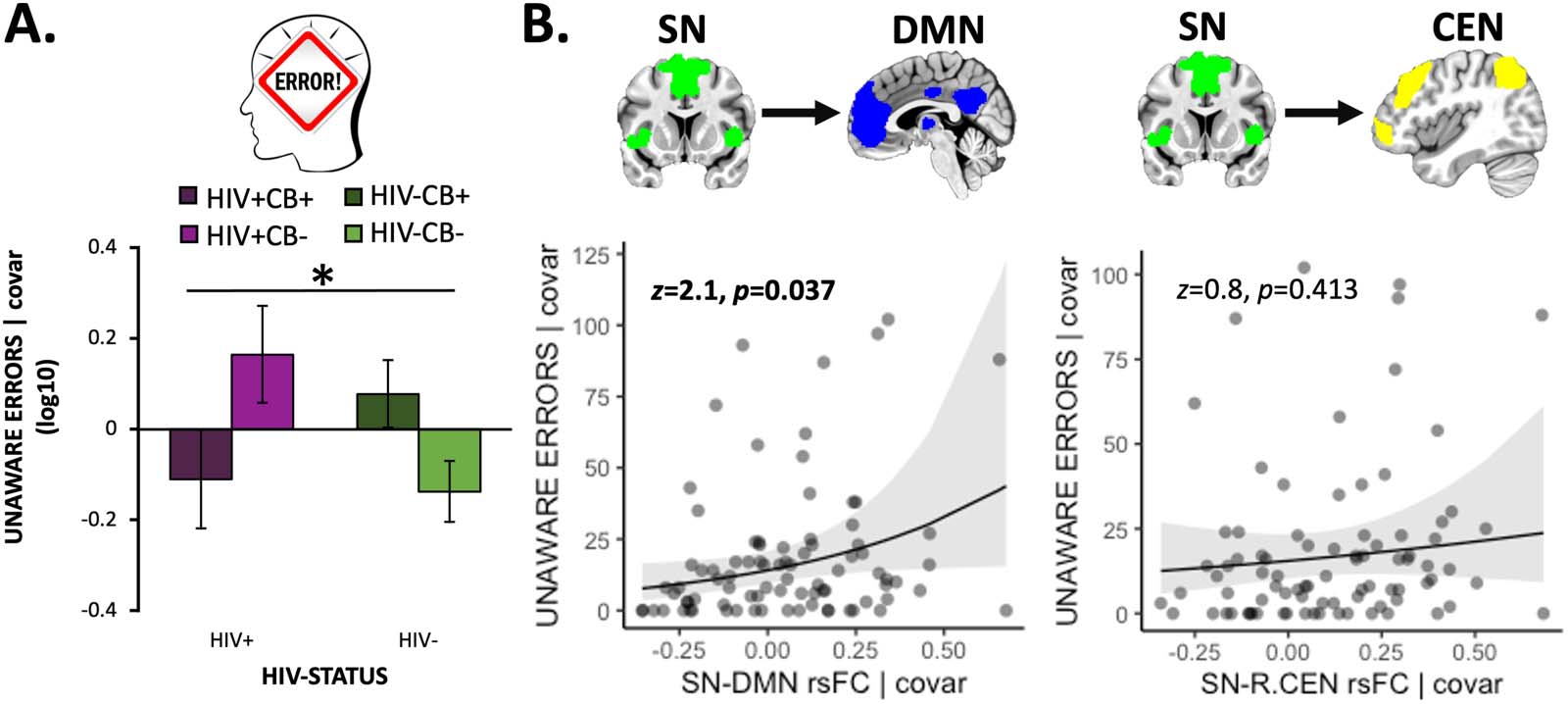
Cifra 2. Error awareness behavior was linked with SN-DMN rsFC strength. (A) We observed an HIV × CB interaction (F[1, 101] = 5.9,
pag = 0.017) when considering error awareness (operationalized as the log-transformed number of unaware errors [log10(X + 1)]) such that
both the HIV-only (HIV+/CB−) and CB-only (HIV−/CB+) grupos (relative to controls, HIV−/CB−) failed to indicate awareness of more
errores, whereas the co-occurring (HIV+/CB+) group displayed fewer unaware errors more akin to that of controls. (B) Increased salience
network–default mode network (SN-DMN) resting-state functional connectivity (rsFC) correlated with more unaware errors in the
Go/NoGo task (norte = 85, b = 1.7 [0.8], z = 2.1, pag = 0.037). A diferencia de, SN rsFC with the left central executive network (L.CEN, datos
not shown: b = 0.6 [0.7], z = 0.8, pag = 0.430) or right CEN (b = 0.6 [0.8], z = 0.8, pag = 0.413) was not linked with error awareness.
Unstandardized residuals are plotted after regressing out effects of covariates. Error bars = standard error of the mean.
Neurociencia en red
802
Large-scale brain network interactions associated with HIV infection
SN-CEN rsFC (Figura 2B) displayed a significant relationship with error awareness (p > 0.4).
These results did not significantly change when including HIV and CB group membership as
covariables. These outcomes suggest that more SN-DMN rsFC was linked with consequences
for task performance, namely reduced awareness of commission errors. Given the severe
non-normality of the unaware error variable, we believe that the negative binomial model
is the proper model for this analysis; sin embargo, to increase transparency, we also reran this anal-
ysis employing a general linear regression while controlling for the same covariates (Figure S3B
in the Supporting Information). The relationship between SN-DMN rsFC and unaware errors
no longer reached significance (pag = 0.082). We also performed follow-up exploratory analy-
ses, among HIV+ participants, assessing relationships between error awareness and HIV dis-
ease characteristics when controlling for CB group membership, edad, sexo, mean FD, and NIC
estado; sin embargo, no significant relationships were observed (p > 0.05).
Network-Level rsFC: Relation With Error-Related Brain Activity
When examining error-related β coefficients from the Go/NoGo task within each network of
interés (DMN, SN, R.CEN, L.CEN), we observed a significant main effect of HIV for the DMN,
such that PLWH (vs. HIV− participants) showed reduced DMN suppression (F[1, 102] = 5.1,
pag = 0.026; Figura 3A). This reduced error-related DMN suppression among PLWH was not
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
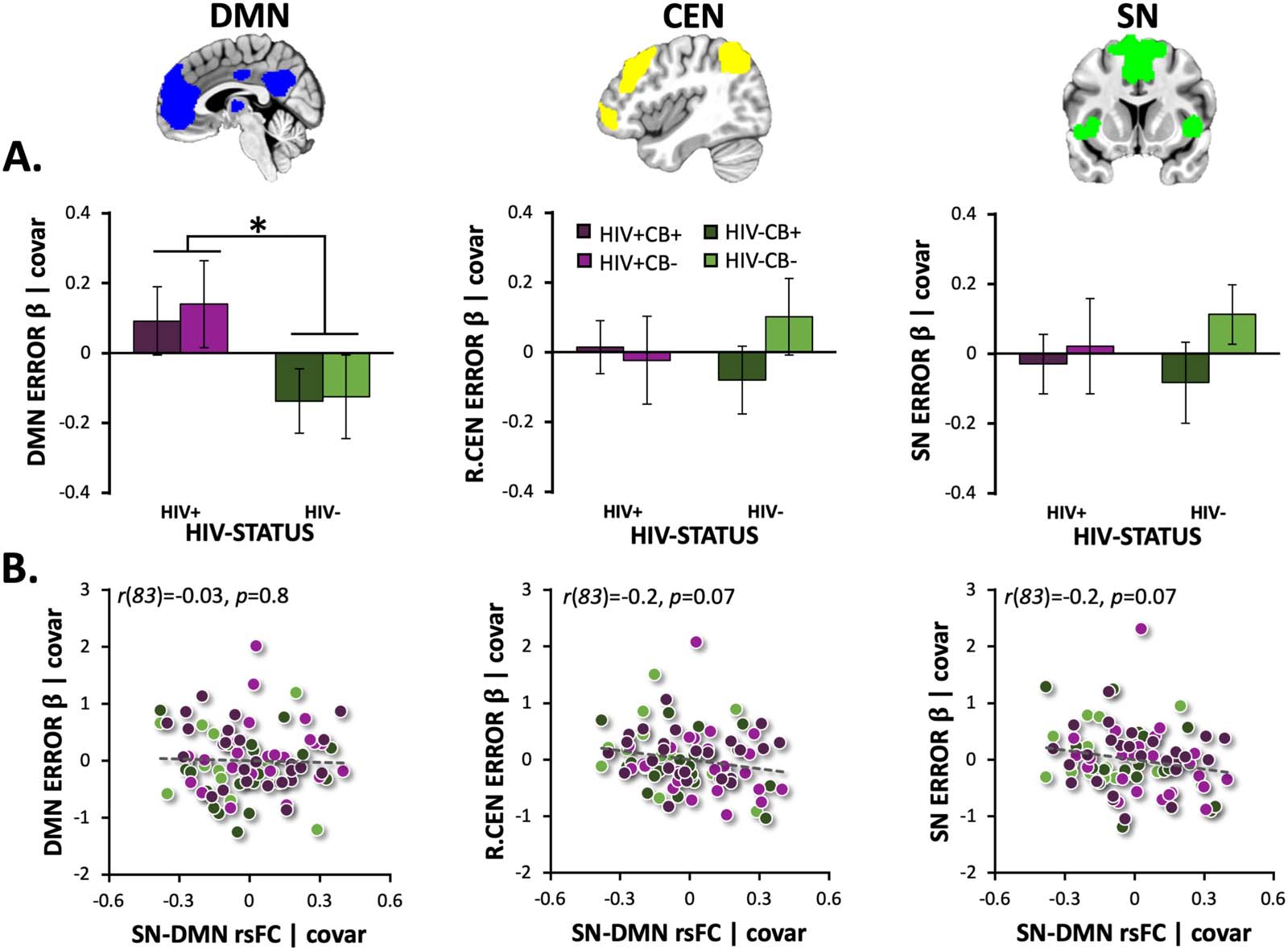
Cifra 3. HIV-associated alterations in error-related DMN suppression and relationships with SN-DMN rsFC strength. (A) People living with
HIV (PLWH) had significantly higher error-related default mode network (DMN) actividad (es decir., reduced DMN suppression; average NoGo-error
β coefficients) than did HIV− participants (F[1, 102] = 5.1, pag = 0.026; norte = 103). Error-related right central executive network (R.CEN), left CEN
(L.CEN, data not displayed), and salience network (SN) activity did not differ between HIV groups (p > 0.3). (B) Resting-state functional
conectividad (rsFC) between the SN and DMN did not correlate with error-related DMN (r(83) = −0.03, pag = 0.8), R.CEN (r(83) = −0.2, pag =
0.07), L.CEN (r(83) = −0.1, pag = 0.2; data not shown), or SN activity (r(83) = −0.2, pag = 0.07). Unstandardized residuals are plotted after
regressing out effects of covariates. Error bars = standard error of the mean.
Neurociencia en red
803
Large-scale brain network interactions associated with HIV infection
impacted by CB use (HIV × CB: F[1, 102] = 0.3, pag = 0.865; CB: F[1, 102] = 0.1, pag = 0.739).
Error-related DMN (z = −1.4, pag = 0.167), R.CEN (z = −1.6, pag = 0.112), L.CEN (data not
mostrado: z = −1.4, pag = 0.169), and SN (z = 0.2, pag = 0.813) responsivity in the error awareness
tarea (EAT) was not significantly associated with error awareness, while controlling for sex,
edad, and NIC status (Figure S4 in the Supporting Information). Finalmente, when controlling for
mean FD (during rest), we did not detect any relations between rsFC (SN-DMN, SN-CEN)
and error-related network activity (p > 0.8; Figura 3B), contrary to our hypothesis.
DISCUSIÓN
We characterized large-scale brain network interaction patterns linked with HIV and CB use
and examined implications for behavior and task-based brain function. We assessed HIV and
CB-related effects on RAIs, a measure previously used to quantify interactions across three
large-scale networks: the SN, cen, and DMN. We observed bilaterally reduced RAIs among
PLWH driven by increased SN-DMN rsFC, but not SN-CEN rsFC. No significant CB-related
effects on RAI and rsFC measures were detected. Our findings link SN-DMN rsFC strength with
an objectively measured behavioral metric; tal como, rsFC was associated with error awareness
during a Go/NoGo task variant. Específicamente, increased SN-DMN coupling correlated with
reduced error awareness (es decir., more unaware errors) across all participants. Contrary to our
hypothesis, SN-DMN rsFC did not correlate with error-related DMN suppression. Sin embargo,
we observed that PLWH displayed reduced error-related DMN suppression compared with
HIV− controls. These results demonstrate a pattern of dysregulated network function among
PLWH and highlight implications for error awareness.
Reduced RAI Among PLWH
The bilaterally reduced RAIs observed among PLWH are similar to those previously observed
across substance use and neuropsychiatric disorders (Alexopoulos et al., 2012; Bartova et al.,
2015; Bednarski et al., 2011; Bonavita et al., 2017; Chang et al., 2014; Gauffin et al., 2013;
Høgestøl et al., 2019; Lee et al., 2017; Liddle et al., 2011; Liu et al., 2018; Oyegbile et al.,
2019; Peterson et al., 2009; Schilbach et al., 2016; Sutherland et al., 2012; Verfaillie et al.,
2018; k. Wang y cols., 2019; Y. Wang y cols., 2013; Whitfield-Gabrieli & Vado, 2012; Wu
et al., 2011; Yin et al., 2016; R. zhang & Volkow, 2019; Zhou y cols., 2016). Nicotine and other
drug-dependent individuals have displayed reduced RAIs during acute withdrawal (Lerman
et al., 2014; Reese et al., 2019). Sin embargo, Moradi et al.’s (2020) recent work questioned
the RAI as a reliable biomarker for substance use disorders following null effects among stim-
ulant and/or opiate users that had been abstinent for, en algunos casos, multiple months (significado de
108 días, que van desde 4 a 365 días). Sin embargo, we note that prior examinations of RAI
changes among dependent substance users suggest that they may be linked with certain cog-
nitive symptoms of an acute withdrawal state (cognitive control among cigarette smokers
[Lerman et al., 2014], craving among individuals with internet gaming disorder [j. t. zhang
et al., 2017], and distress tolerance among cocaine users [Reese et al., 2019]), and these
symptoms were not considered in Moradi et al.’s (2020) estudiar (Lerman et al., 2014; Reese
et al., 2019; j. t. Zhang et al., 2017). Many of the cognitive deficits characterizing with-
drawal are hypothesized to stem from an inability to suppress attention toward internal craving
and aversive somatic withdrawal symptoms (Ashare et al., 2014; Lerman et al., 2014; Shoaib
& Bizarro, 2005; Sutherland et al., 2012; Wesnes et al., 2013). Our results point toward a
similar mechanism at play among PLWH that could account for certain cognitive deficits that
persist in the post–antiretroviral therapy era. Específicamente, deficits reported in learning, mem-
ory, and performance on cognitive tasks involving executive function may be manifestations of
Neurociencia en red
804
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
a more general attentional impairment characterized by an inability to suppress attention
toward physical/emotional HIV symptomology and/or other task-irrelevant, intrusive thoughts
linked with the DMN.
Increased SN-DMN rsFC Among PLWH
This interpretation is further supported by our observation that HIV-associated RAI reductions
were primarily driven by PLWH (vs. HIV− participants) presenting with higher SN-DMN syn-
chrony. Prior work demonstrating altered network organization among PLWH (Abidin et al.,
2018; Hall et al., 2021; Minosse et al., 2021) has also often highlighted alterations involving
the DMN (Thomas et al., 2015; Zhuang et al., 2017). Por ejemplo, one study found that treat-
ment naïve PLWH displayed significantly reduced rsFC within the DMN when compared with
controls (Zhuang et al., 2017), while another study utilizing graph theoretic metrics found
alterations in closeness centrality (a metric indicating connectiveness with the rest of the brain)
within the DMN and frontoparietal network among PLWH (Thomas et al., 2015). Our findings
are also generally consistent with prior work among individuals living with other conditions
leading to chronic inflammation in the central nervous system (es decir., multiple sclerosis [MS]).
Específicamente, people living with MS displayed increased rsFC between DMN regions and those
comprising the SN, and such rsFC alterations correlated with symptom severity (Bonavita
et al., 2017; Høgestøl et al., 2019). Altered DMN rsFC also has been consistently linked with
Alzheimer’s disease (ANUNCIO; Damoiseaux et al., 2012; Jones et al., 2016) and other neurodegen-
erative disorders (Alexopoulos et al., 2012; Liu et al., 2018; Yin et al., 2016). Por ejemplo,
emerging evidence suggests that mild cognitive declines that precede the onset of progressive
deterioration in AD may be associated with an initial increase in DMN rsFC followed by sub-
sequent decreases (Damoiseaux et al., 2012; Jones et al., 2016; Wu et al., 2011). Respectivamente,
changes in DMN rsFC may be predictive of future cognitive impairments before neuropsycho-
logical performance falls outside a normative range (Verfaillie et al., 2018; Y. Wang y cols.,
2013). En efecto, when considering individuals with a family history of AD, early subjective cog-
nitive decline is associated with increased rsFC between the DMN and regions of the medial
temporal lobe memory system (Verfaillie et al., 2018). A diferencia de, individuals with mild
cognitive impairments and those reporting cognitive complaints (despite normal neuropsy-
chological performance) appear to display decreased rsFC between the DMN and right
hippocampus relative to age-matched controls (Y. Wang y cols., 2013). In light of such AD
findings, we suggest that future work could consider longitudinal changes in DMN rsFC across
HIV disease phases with attention towards potential nonlinear associations with cognitive
decline (Damoiseaux et al., 2012; Jones et al., 2016; Wu et al., 2011).
Despite prior reports of reduced RAI values associated with certain symptoms of addiction
disorders, we did not observe significant CB effects on RAI values or SN-DMN rsFC. This null
resultado, corresponding with Moradi et al.’s (2020) null findings, may be due to CB users in our
sample not being in an acute withdrawal state, or interacting influences of HIV that we did not
have the power to detect. Curiosamente, while we did not detect significant HIV × CB inter-
action effects, visual inspection and exploratory follow-up t tests indicated that the CB-using
HIV+ group displayed reduced SN-DMN rsFC, more similar to that of the control group,
when compared with the non-using PLWH group. These exploratory observations may sup-
port prior work observing CB normalizing effects among PLWH when considering task-based
activity within SN regions (Meade et al., 2019) and rsFC metrics centered on the insula
(Flannery, Riedel, Salo, Hill-Bowen, et al., 2021). As identifying specific brain network inter-
action patterns linked with both CB use and possible CB normalizing effects among PLWH
could help inform clinical practices regarding medicinal CB use among this population,
Neurociencia en red
805
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
future work should further examine aspects of CB use among PLWH and its effects on net-
work rsFC.
Network-Level rsFC: Relation With Error Awareness
While both the HIV-only (HIV+/CB−) and CB-only (HIV−/CB+) groups displayed diminished
error awareness relative to controls, the co-occurring (HIV+/CB+) group’s error awareness was
more similar to that of the controls. These outcomes align with prior observations of a partially
normalizing effect of CB use on HIV-associated brain function alterations and error processing
(Flannery, Riedel, Salo, Hill-Bowen, et al., 2021; Hall et al., 2021; Meade et al., 2019). Como
chronic inflammation in the central nervous system is one mechanism through which HIV
may lead to progressive cognitive declines (Benatti et al., 2016; Boerwinkle & Ances,
2019), it has been hypothesized that the anti-inflammatory properties of CB could offer some
benefits among PLWH (Burstein, 2015; Ellis et al., 2009; Gallily et al., 2018; Watson et al.,
2020). Supporting this notion, CB use has been linked with reduced inflammatory biomarkers
in cerebral spinal fluid and blood (Castro et al., 2019; Ellis et al., 2020; Rizzo et al., 2018), y
PLWH frequently report using CB to relieve somatic complaints, and anxious or depressed
moods (Harris et al., 2014; Towe et al., 2020). dicho eso, continued research is still needed
to better understand the impact of CB use on interacting physical, affective, and neurocogni-
tive symptoms of HIV (Bonnet & Preuss, 2017; Bovasso, 2001; Okafor et al., 2019; Thames
et al., 2016).
Además, we observed that higher SN-DMN rsFC was associated with diminished error
awareness. These outcomes highlight the relevance of network interactions assessed at rest for
error monitoring during tasks. While increased SN-DMN rsFC has not previously been linked
to compromised error awareness, it has been linked to attentional problems in the context of
TDAH, with medication down regulating DMN rsFC with regions comprising the SN (Biskup
et al., 2016; Querne et al., 2017). Given the theorized role of the SN in toggling neurocogni-
tive resources across large-scale brain networks, we hypothesized that SN-DMN rsFC has
implications for task-induced DMN suppression. As task-induced DMN suppression is thought
to support task vigilance and detection of salient stimuli (singh & Fawcett, 2008), it is likely
also vital for error recognition.
Network-Level rsFC: Relation With Error-Related Brain Activity
While research has shown that properties of functional networks identified during rest reflect a
similar functional network architecture during tasks, it remains unclear how the two are related
(Cole y col., 2014). To more fully understand altered network organization among PLWH, nosotros
examined relations between rsFC and task event-induced network responsivity. We did not
observe a relation between measures of network function during rest and those during task.
While it is possible that both alterations in rsFC and task-based responsivity are associated with
a certain condition, en este caso, living with HIV, the two may be unrelated consequences.
Sin embargo, it should be noted that rest and task are distinct cognitive states and functional con-
nectivity and task event responsivity are different quantifications of brain function with distinct
data preparation and analysis techniques (stevens, 2016). De este modo, failure to detect a relation
between specific alterations in both could be influenced by multiple differences in these
two variables and their quantification. Más, while rest and task data were collected during
the same scan session (1.5 hr), a lack of within-subject reliability/stability across scans has
previously been documented (Pannunzi et al., 2017). Whether rsFC alterations, observado
among certain populations, represent a broad-spectrum alteration that also impacts brain pro-
cesses probed during tasks is a potentially important scientific question that could help
Neurociencia en red
806
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
advance understanding of neurobiological consequences of HIV. Future work should continue
to consider how to best examine relations between the degree of regional responsivity to
stimuli/events and functional synchrony between of two or more regions.
Reduced task-induced DMN suppression has been linked with various neurocognitive dis-
orders (Gauffin et al., 2013; Oyegbile et al., 2019; Whitfield-Gabrieli & Vado, 2012), incluido
addiction (Bednarski et al., 2011; Sutherland et al., 2012; R. zhang & Volkow, 2019), TDAH
(Liddle et al., 2011; Peterson et al., 2009), major depressive disorder (Bartova et al., 2015), y
schizophrenia (Zhou y cols., 2016). Además, our own recent work characterized reduced
error-related suppression of the medial prefrontal cortex (mPFC) and posterior cingulate cortex
(PCC; two primary nodes of the DMN) among PLWH (Flannery, Riedel, Salo, Poudel, et al.,
2021). Aquí, we replicated these findings, when adopting a network-level (en contraposición a un
regional) analytic framework, such that PLWH demonstrated reduced error-related DMN sup-
presion. A robust body of work indicates that task-based DMN suppression plays a critical
role in monitoring task stimuli (Hinds et al., 2013) and is related to increasing cognitive
demandas (Allen et al., 2013; Leech et al., 2011) and task performance (Anticevic et al.,
2012; Le et al., 2007). Given our finding that SN-DMN rsFC was both heightened among
PLWH and associated with reduced error awareness, we expected that less error-related
DMN suppression among PLWH would be similarly related to error awareness; sin embargo,
we did not detect an association between DMN suppression and error awareness.
Limitaciones
While we elected to utilize the RAI metric to quantify network-level interactions consistent
with previous drug use–related studies (Lerman et al., 2014; Sutherland et al., 2012), this work
should be considered in light of limitations. Primero, recent work by Moradi et al. (2020) alto-
lighted the impact of mask definition on RAIs. To address mask definition–related variability,
we used two different strategies to define our network masks (atlas-based and ICA-based) y
came to similar conclusions. Segundo, we note that, while past use of and dependence on sub-
stances other than cannabis was permitted across each group to provide a more representative
and generalizable sample, the CB-using groups reported significantly more past month nico-
tine use than non-cannabis-using groups. Además, because of the disproportionate HIV
diagnoses among males in the United States, and challenges recruiting among this population,
our subject groups did not have an equal distribution of self-reported males and females. Nosotros
statistically controlled for both sex and current cigarette smoking status but were unable to rule
out potentially confounding effects that may have still influenced results. Finalmente, while our
work controlled for age effects, we did not have a large enough sample to assess complex
interactive effects of age in the context of HIV and CB use; sin embargo, these interactive trajec-
tories may be of interest in future research.
Conclusions
Our results demonstrate heightened SN-DMN rsFC among PLWH (vs. HIV− participants) eso
was linked with diminished error awareness behavior across all participants, but contrary to
our hypothesis, was not linked with error-related network responsivity. Sin embargo, we did
observe significantly reduced error-related DMN suppression among PLWH. Curiosamente,
our results also displayed a significant HIV × CB interaction on error awareness behavior such
that both the HIV-only (HIV+/CB−) and CB-only (HIV−/CB+) groups exhibited diminished error
awareness relative to controls, whereas the co-occurring (HIV+/CB+) group displayed error
awareness more similar to that of the controls. These findings demonstrate a pattern of dysreg-
ulated network function among PLWH that has been associated with other neurocognitive
Neurociencia en red
807
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
condiciones, further highlighting the importance and ubiquity of this functional network per-
perspectiva. Our results also suggest that such dysregulation may exist during both resting state
and task performance and reflect an inability to disengage irrelevant mental operations, ulti-
mately hindering error awareness. As monitoring errors is vital for everyday functioning and
critical disease management behaviors, we speculate that interventions facilitating DMN sup-
presion (p.ej., mindfulness-based practices, working memory training; Garrison et al., 2015;
Salmi et al., 2018) may be beneficial for PLWH and the challenges they face.
EXPRESIONES DE GRATITUD
We thank the FIU Instructional and Research Computing Center (IRCC, https://ircc.fiu.edu) para
providing access to the HPC computing resources that contributed to the generation of the
research results reported herein.
DATA AND CODE AVAILABILITY
The authors have released all code associated with this manuscript. Code and tabular data are
available on GitHub (https://github.com/Flanneryg3/HIVCB_ProjectCode; Flannery, 2021).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00241.
CONTRIBUCIONES DE AUTOR
Jessica S. Flannery: Conceptualización; Análisis formal; Administración de proyecto; Writing –
original draft. miguel c.. Riedel: Análisis formal; Metodología. Lauren D. Hill-Bowen:
Escritura – revisión & edición. Ranjita Poudel: Administración de proyecto; Escritura – revisión & edición.
Katherine L. Bottenhorn: Escritura – revisión & edición. Taylor Salo: Curación de datos; Formal
análisis; Escritura – revisión & edición. Angela R. Laird: Adquisición de financiación; Escritura – revisión
& edición. Raul Gonzalez: Adquisición de financiación; Administración de proyecto; Escritura – revisión &
edición. Matthew T. Sutherland: Conceptualización; Adquisición de financiación; Project administra-
ción; Supervisión; Escritura – revisión & edición.
INFORMACIÓN DE FINANCIACIÓN
Matthew T. Sutherland, Foundation for the National Institutes of Health (https://dx.doi.org/10
.13039/100000009), Award ID: K01DA037819. Raul Gonzalez, Foundation for the National
Institutes of Health (https://dx.doi.org/10.13039/100000009), Award ID: R01DA033156.
Matthew T. Sutherland, Foundation for the National
Institutes of Health (https://dx.doi
.org/10.13039/100000009), Award ID: U54MD012393. Matthew T. Sutherland, Founda-
tion for the National Institutes of Health (https://dx.doi.org/10.13039/100000009), Award
ID: R01DA041353. Angela R. Laird, Fundación Nacional de Ciencia (https://dx.doi.org/10
.13039/501100008982), Award ID: 1631325.
REFERENCIAS
Abidin, A. Z., D’Souza, A. METRO., Nagarajan, METRO. B., Wang, l., Qiu, X.,
Schifitto, GRAMO., & Wismuller, A. (2018). Alteration of brain network
topology in HIV-associated neurocognitive disorder: A novel
functional connectivity perspective. NeuroImagen: Clinical, 17,
768–777. https://doi.org/10.1016/j.nicl.2017.11.025, PubMed:
29527484
Abrahán, A., Pedregosa, F., Eickenberg, METRO., Gervais, PAG., Mueller,
A., Kossaifi, J., Gramfort, A., Thirion, B., & Varoquaux, GRAMO.
Neurociencia en red
808
Large-scale brain network interactions associated with HIV infection
(2014). Machine learning for neuroimaging with scikit-learn.
Frontiers in Neuroinformatics, 8, 14. https://doi.org/10.3389
/fninf.2014.00014, PubMed: 24600388
Adhikari, B. METRO., Jahanshad, NORTE., Shukla, D., Glahn, D., Blangero, J.,
Reynolds, r., Cox, r., Fieremans, MI., Veraart, J., Nóvikov, D., &
Nichols, t. (2018). Heritability estimates on resting state fMRI
data using ENIGMA analysis pipeline. Pacific Symposium on
B i o c o m p u t i n g , 2 3 , 3 0 7 – 3 1 8 . h t t p s : / / d o i . o rg / 1 0 . 11 4 2
/9789813235533_0029, PubMed: 29218892
Adhikari, B. METRO., Jahanshad, NORTE., Shukla, D., Tornero, J., Grotegerd,
D., Dannlowski, Ud., Kugel, h., Engelen, J., Dietsche, B., krug,
A., Kircher, T., Fieremans, MI., Veraart, J., Nóvikov, D. S., Boedhoe,
PAG. S. w., van der Werf, Y. D., van den heuvel, oh. A., Ipser, J., Uhlmann,
A., … Kochunov, PAG. (2019). A resting state fMRI analysis pipeline
for pooling inference across diverse cohorts: An ENIGMA rs-fMRI
protocol. Brain Imaging and Behavior, 13(5), 1453–1467. https://
doi.org/10.1007/s11682-018-9941-x, PubMed: 30191514
Alexopoulos, GRAMO. S., Hoptman, METRO. J., Kanellopoulos, D., Murphy,
C. F., Lim, k. o., & Gunning, F. METRO. (2012). Functional connec-
tivity in the cognitive control network and the default mode
network in late-life depression. Journal of Affective Disorders,
139(1), 56–65. https://doi.org/10.1016/j.jad.2011.12.002,
PubMed: 22425432
allen, METRO., Smallwood, J., Christensen, J., Gramm, D., Rasmussen,
B., Jensen, C. GRAMO., Roepstorff, A., & Lutz, A. (2013). The balanced
mente: The variability of task-unrelated thoughts predicts error
supervisión. Frontiers in Human Neuroscience, 7, 743. https://
doi.org/10.3389/fnhum.2013.00743, PubMed: 24223545
Altmann, A., Ng, B., Landau, S., Jagust, w., & Greicius, METRO. (2015).
Regional brain hypometabolism is unrelated to regional amyloid
plaque burden. Cerebro, 138, 3734–3746. https://doi.org/10.1093
/brain/awv278, PubMed: 26419799
Anticevic, A., Col, METRO. w., Murray, j. D., Corlett, PAG. r., Wang, X. J.,
& Krystal, j. h. (2012). The role of default network deactivation
in cognition and disease. Tendencias en Ciencias Cognitivas, 16(12),
584–592. https://doi.org/10.1016/j.tics.2012.10.008, PubMed:
23142417
Ashare, R. l., Falcone, METRO., & Lerman, C. (2014). Cognitive function
during nicotine withdrawal: Implications for nicotine depen-
dence treatment. Neuropharmacology, 76(punto. B), 581–591.
https://doi.org/10.1016/j.neuropharm.2013.04.034, PubMed:
23639437
Atkins, D., & Gallop, R.
(2007). Rethinking how family
researchers model infrequent outcomes: A tutorial on count
regression and zero-inflated models. Journal of Family Psychology,
21(4), 726–735. https://doi.org/10.1037/0893-3200.21.4.726,
PubMed: 18179344
Bartova, l., Meyer, B. METRO., Diers, K., Rabl, Ud., Scharinger, C., popovic,
A., Pail, GRAMO., Kalcher, K., Boubela, R. NORTE., Huemer, J., Mandorfer, D.,
Windischberger, C., Sitte, h. h., Kasper, S., Praschak-Rieder, NORTE.,
Moser, MI., Brocke, B., & Pezawas, l. (2015). Reduced default mode
network suppression during a working memory task in remitted
major depression. Journal of Psychiatric Research, 64, 9–18.
https://doi.org/10.1016/j.jpsychires.2015.02.025, PubMed:
25801734
Bassel, C., Rourke, S. B., Halman, METRO. h., & Herrero, METRO. l. (2002).
Working memory performance predicts subjective cognitive
complaints in HIV infection. Neuropsychology, 16(3), 400–410.
https://doi.org/10.1037/0894-4105.16.3.400, PubMed:
12146687
Bednarski, S. r., zhang, S., hong, k. I., Sinha, r., Rounsaville, B. J.,
& li, C. S. (2011). Deficits in default mode network activity pre-
ceding error in cocaine dependent individuals. Drug and Alcohol
Dependence, 119(3), e51–e57. https://doi.org/10.1016/j
.drugalcdep.2011.05.026, PubMed: 21703783
Behzadi, y., Restom, K., Liau, J., & Liu, t. t. (2007). A component
based noise correction method (CompCor) for BOLD and perfu-
sion based fMRI. NeuroImagen, 37(1), 90–101. https://doi.org/10
.1016/j.neuroimage.2007.04.042, PubMed: 17560126
Benatti, C., Blom, j. METRO. C., Rigillo, GRAMO., Alboni, S., Zizzi, F., Torta, r.,
Brunello, NORTE., & Tascedda, F. (2016). Disease-induced neuroin-
flammation and depression. CNS and Neurological Disorders—
Drug Targets, 15(4), 414–433. https://doi.org/10.2174
/1871527315666160321104749, PubMed: 26996176
Benedict, R. h., Schretlen, D., Groninger, l., & Brandt, j. (1998).
Hopkins Verbal Learning Test–Revised: Normative data and anal-
ysis of inter-form and test-retest reliability. The Clinical Neuropsy-
chologist, 12(1), 43–55. https://doi.org/10.1076/clin.12.1.43
.1726
Biskup, C. S., Helmbold, K., Baurmann, D., Klasen, METRO., Gaber, t. J.,
Bubenzer-Busch, S., Königschulte, w., Fink, GRAMO. r., & Zepf, F. D.
(2016). Resting state default mode network connectivity in chil-
dren and adolescents with ADHD after acute tryptophan deple-
ción. Acta Psychiatrica Scandinavica, 134(2), 161–171. https://doi
.org/10.1111/acps.12573, PubMed: 27145324
Blackstone, K., moore, D. J., Heaton, R. K., franklin, D. r., Jr.,
Woods, S. PAG., Clifford, D. B., Minero, A. C., Marra, C. METRO., Gelman,
B. B., McArthur, j. C., Morgello, S., Simpson, D. METRO., Rivera-Mindt,
METRO., Deutsch, r., Ellis, R. J., Hampton Atkinson, J., & Grant, I.
(2012). Diagnosing symptomatic HIV-associated neurocognitive
disorders: Self-report versus performance-based assessment of
everyday functioning. Journal of the International Neuropsycho-
logical Society, 18(1), 79–88. https://doi.org/10.1017
/S135561771100141X, PubMed: 22114912
Boerwinkle, A., & Ances, B. METRO. (2019). Molecular imaging of neu-
roinflammation in HIV. Journal of Neuroimmune Pharmacology,
14(1), 9-15. https://doi.org/10.1007/s11481-018-9823-4,
PubMed: 30515624
Bonavita, S., Sacco, r., Esposito, S., d’Ambrosio, A., Della Corte,
METRO., Corbo, D., Docimo, r., Gallo, A., Lavorgna, l., Cirillo, METRO.,
& Bisecco, A. (2017). Default mode network changes in multiple
sclerosis: A link between depression and cognitive impairment?
European Journal of Neurology, 24(1), 27–36. https://doi.org/10
.1111/ene.13112, PubMed: 27633185
Bonnet, Ud., & Preuss, Ud.. W.. (2017). The cannabis withdrawal syn-
drome: Current insights. Substance Abuse and Rehabilitation, 8,
9–37. https://doi.org/10.2147/SAR.S109576, PubMed:
28490916
Bovasso, GRAMO. B. (2001). Cannabis abuse as a risk factor for depressive
symptoms. Revista americana de psiquiatría, 158(12), 2033–2037.
https://doi.org/10.1176/appi.ajp.158.12.2033, PubMed:
11729021
Brandt, j. (1991). The Hopkins Verbal Learning Test: Desarrollo
of a new memory test with six equivalent forms. The Clinical
Neuropsychologist, 5(2), 125–142. https://doi.org/10.1080
/13854049108403297
Neurociencia en red
809
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
Buckner, R. l., & DiNicola, l. METRO. (2019). The brain’s default net-
trabajar: Updated anatomy, physiology and evolving insights.
Naturaleza Reseñas Neurociencia, 20(10), 593–608. https://doi.org
/10.1038/s41583-019-0212-7, PubMed: 31492945
Burgess, GRAMO. C., Kandala, S., Nolan, D., Laumann, t. o., Fuerza,
j. D., Esperemos, B., Harms, METRO. PAG., Petersen, S. MI., & Respeto,
D. METRO. (2016). Evaluation of denoising strategies to address
motion-correlated artifacts in resting-state functional magnetic
resonance imaging data from the Human Connectome Project.
Brain Connectivity, 6(9), 669–680. https://doi.org/10.1089/brain
.2016.0435, PubMed: 27571276
Burstein, S. (2015). Cannabidiol (CBD) and its analogs: Una revisión de
their effects on inflammation. Bioorganic and Medicinal Chemis-
intentar, 23(7), 1377–1385. https://doi.org/10.1016/j.bmc.2015.01
.059, PubMed: 25703248
carey, C. l., Woods, S. PAG., Rippeth, j. D., González, r., moore,
D. J., Marcotte, t. D., Grant, I., Heaton, R. K., & HNRC Group.
(2004). Initial validation of a screening battery for the detection
of HIV-associated cognitive impairment. The Clinical Neuropsy-
c h o l o g i s t , 1 8 ( 2 ) , 2 3 4 – 2 4 8 . h t t p s : / / d o i . o r g / 1 0 . 1 0 8 0
/13854040490501448, PubMed: 15587671
Castelo, j. METRO. B., sherman, S. J., Courtney, METRO. GRAMO., Melrose, R. J., &
Stern, C. mi. (2006). Altered hippocampal-prefrontal activation in
HIV patients during episodic memory encoding. Neurología,
6 6 ( 11 ) , 1 6 8 8 – 1 6 9 5 . h t t p s : / / d o i . o rg / 1 0 . 1 2 1 2 / 0 1 . w n l
.0000218305.09183.70, PubMed: 16769942
Castro, F. oh. F., silva, j. METRO., Dorneles, GRAMO. PAG., Barros, j. B. S., Ribeiro,
C. B., Noronha, I., Barbosa, GRAMO. r., Souza, l. C. S., Guilarde,
A. o., Pereira, A., Guimaraes, R. F., Oliveira, t. F., Oliveira,
S. mi. F., Peres, A., Romao, PAG. R. T., Pfrimer, I. A. h., & Fonseca,
S. GRAMO. D. (2019). Distinct inflammatory profiles in HIV-infected
individuals under antiretroviral therapy using cannabis, cocaína
or cannabis plus cocaine. AIDS, 33(12), 1831–1842. https://doi
.org/10.1097/QAD.0000000000002296, PubMed: 31490211
Cattie, j. MI., Doyle, K., Weber, MI., Grant, I., Woods, S. PAG., & HNRP
Group. (2012). Planning deficits in HIV-associated neurocogni-
tive disorders: Component processes, cognitive correlates, y
implications for everyday functioning. Journal of Clinical and
Experimental Neuropsychology, 34(9), 906–918. https://doi.org
/10.1080/13803395.2012.692772, PubMed: 22731608
Centers for Disease Control and Prevention. (2020). Estimated HIV
incidence and prevalence in the United States, 2014–2018 (HIV
Surveillance Supplemental Report Vol. 25, No. 1). Retrieved from
https://www.cdc.gov/hiv/library/reports/hiv-surveillance.html
Chang, X., shen, h., Wang, l., Liu, Z., Xin, w., Hu, D., & Miao, D.
(2014). Altered default mode and fronto-parietal network subsys-
tems in patients with schizophrenia and their unaffected siblings.
Brain Research, 1562, 87–99. https://doi.org/10.1016/j.brainres
.2014.03.024, PubMed: 24675026
Chen, GRAMO., taylor, PAG. A., & Cox, R. W.. (2017). Is the statistic value all we
should care about in neuroimaging? NeuroImagen, 147, 952–959.
https://doi.org/10.1016/j.neuroimage.2016.09.066, PubMed:
27729277
Choi, J., jeong, B., Sotavento, S. w., & Go, h. j. (2013). Aberrant devel-
opment of functional connectivity among resting state-related
functional networks in medication-naive ADHD children. PLoS
ONE, 8(12), e83516. https://doi.org/10.1371/journal.pone
.0083516, PubMed: 24386219
Chong, j. S. X., Ng, GRAMO. j. PAG., Sotavento, S. C., & zhou, j. (2017). Salience
network connectivity in the insula is associated with individual
differences in interoceptive accuracy. Brain Structure and Func-
ción, 222(4), 1635–1644. https://doi.org/10.1007/s00429-016
-1297-7, PubMed: 27573028
Col, METRO. w., bassett, D. S., Fuerza, j. D., Más valiente, t. S., & Petersen,
S. mi. (2014). Intrinsic and task-evoked network architectures of
el cerebro humano. Neurona, 83(1), 238–251. https://doi.org/10
.1016/j.neuron.2014.05.014, PubMed: 24991964
Cordero-Grande, l., Christiaens, D., Hutter, J., Precio, A. NORTE., &
amanecer, j. V. (2019). Complex diffusion-weighted image estima-
tion via matrix recovery under general noise models. Neuro-
Image, 200, 391–404. https://doi.org/10.1016/j.neuroimage
.2019.06.039, PubMed: 31226495
Cox, R. W.. (1996). AFNI: Software for analysis and visualization of
functional magnetic resonance neuroimages. Computers and
Biomedical Research, 29(3), 162–173. https://doi.org/10.1006
/cbmr.1996.0014, PubMed: 8812068
Damoiseaux, j. S., Prater, k. MI., Molinero, B. l., & Greicius, METRO. D. (2012).
Functional connectivity tracks clinical deterioration in Alzhei-
mer’s disease. Neurobiology of Aging, 33(4), 828.e19–828.e30.
https://doi.org/10.1016/j.neurobiolaging.2011.06.024, PubMed:
21840627
Dawes, S., Suarez, PAG., casey, C. y., Cherner, METRO., Marcotte, t. D.,
Letendre, S., Grant, I., Heaton, R. K., & HNRC Group. (2008).
Variable patterns of neuropsychological performance in HIV-1
infection. Journal of Clinical and Experimental Neuropsychology,
30(6), 613–626. https://doi.org/10.1080/13803390701565225,
PubMed: 18608689
Di, X., & Biswal, B. B. (2014). Identifying the default mode network
structure using dynamic causal modeling on resting-state functional
magnetic resonance imaging. NeuroImagen, 86, 53–59. https://doi
.org/10.1016/j.neuroimage.2013.07.071, PubMed: 23927904
D’Souza, GRAMO., Matson, PAG. A., grady, C. D., Nahvi, S., Merenstein, D.,
Weber, k. METRO., Greenblatt, r., Burian, PAG., & wilson, t. mi. (2012).
Medicinal and recreational marijuana use among HIV-infected
women in the Women’s Interagency HIV Study ( WIHS) grupo,
1994–2010. Journal of Acquired Immune Deficience Syndromes,
61(5), 618–626. https://doi.org/10.1097/QAI.0b013e318273ab3a,
PubMed: 23011399
Ellis, R. J., Peterson, S. NORTE., li, y., Schrier, r., Iudicello, J., Letendre,
S., morgan, MI., Espiga, B., Grant, I., & Cherner, METRO. (2020). Reciente
cannabis use in HIV is associated with reduced inflammatory
markers in CSF and blood. Neurología: Neuroimmunology and
Neuroinflammation, 7(5), e809. https://doi.org/10.1212/ NXI
.0000000000000809, PubMed: 32554630
Ellis, R. J., Toperoff, w., Vaida, F., van den Brande, GRAMO., González, J.,
Gouaux, B., Bentley, h., & Atkinson, j. h. (2009). Smoked medic-
inal cannabis for neuropathic pain in HIV: A randomized, cross-
over clinical trial. Neuropsicofarmacología, 34(3), 672–680.
https://doi.org/10.1038/npp.2008.120, PubMed: 18688212
Ernst, T., Chang, l., & arnold, S. (2003). Increased glial metabolites
predict increased working memory network activation in HIV
brain injury. NeuroImagen, 19(4), 1686–1693. https://doi.org/10
.1016/s1053-8119(03)00232-5, PubMed: 12948723
Esteban, o., Markiewicz, C. J., Blair, R. w., Moody, C. A., Isik, A. I.,
Erramuzpe, A., Kent, j. D., Goncalves, METRO., DuPre, MI., Snyder, METRO.,
Oya, h., Ghosh, S. S., Wright, J., Durnez, J., Poldrack, R. A., &
Neurociencia en red
810
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
Gorgolewski, k. j. (2018). FMRIPrep: A robust preprocessing
pipeline for functional MRI. bioRxiv. https://doi.org/10.1101
/306951
Falcone, METRO., Wileyto, mi. PAG., Ruparel, K., Gerraty, R. T., LaPrate, l.,
Detre, j. A., Gur, r., Loughead, J., & Lerman, C. (2014). Age-
related differences in working memory deficits during nicotine
withdrawal. Addiction Biology, 19(5), 907–917. https://doi.org
/10.1111/adb.12051, PubMed: 23496760
Primero, METRO. B. (2014). Structured clinical interview for the DSM (SCID). En
R. l. Cautin & S. oh. Lilienfeld (Editores.), The encyclopedia of clinical
psicología. https://doi.org/10.1002/9781118625392.wbecp351
Flannery, j. S. (2021). HIVCB project code, GitHub, https://github
.com/Flanneryg3/HIVCB_ProjectCode
Flannery, j. S., Riedel, METRO., Salo, T., Hill-Bowen, l., Poudel, r.,
Adams, A., Laird, A., González, r., & Sutherland, METRO. (2021).
Interactive effects of HIV infection and cannabis use on insula
subregion functional connectivity. Journal of Neuroimmune
Pharmacology. https://doi.org/10.1007/s11481-021-10005-8,
PubMed: 34427866
Flannery, j. S., Riedel, METRO. C., Salo, T., Poudel, r., Laird, A. r., González,
r., & Sutherland, METRO. t. (2021). HIV infection is linked with reduced
error-related default mode network suppression and poorer medica-
tion management abilities. Progress in Neuro-Psychopharmacology
and Biological Psychiatry, 111, 110398. https://doi.org/10.1016/j
.pnpbp.2021.110398, PubMed: 34224796
fonov, V. S., evans, A. C., McKinstry, R. C., almíli, C. r., & collins,
D. l. (2009). Unbiased nonlinear average age-appropriate brain
templates from birth to adulthood. NeuroImagen, 47(S1), S102.
https://doi.org/10.1016/S1053-8119(09)70884-5
Gallily, r., Yekhtin, Z., & Hanuš, l. oh. (2018). The anti-
inflammatory properties of terpenoids from cannabis. Cannabis
and Cannabinoid Research, 3(1), 282–290. https://doi.org/10.1089
/can.2018.0014, PubMed: 30596146
gao, C., Meng, J., xiao, X., Wang, METRO., williams, A. B., & Wang, h.
(2020). Antiretroviral therapy improves neurocognitive impair-
ment in people living with HIV? A meta-analysis. Internacional
Journal of Nursing Sciences, 7(2), 238–247. https://doi.org/10
.1016/j.ijnss.2020.03.007, PubMed: 32685622
Garrison, k. A., Zeffiro, t. A., Scheinost, D., Constable, R. T., &
Cervecero, j. A. (2015). Meditation leads to reduced default mode
network activity beyond an active task. Cognitivo, Afectivo, y
Neurociencia conductual, 15(3), 712–720. https://doi.org/10
.3758/s13415-015-0358-3, PubMed: 25904238
Gauffin, h., van Ettinger-Veenstra, h., Landtblom, A. METRO., Ulrici, D.,
McAllister, A., Karlsson, T., & Engstrom, METRO. (2013). Impaired lan-
guage function in generalized epilepsy: Inadequate suppression
de
the default mode network. Epilepsy and Behavior, 28(1),
26–35. https://doi.org/10.1016/j.yebeh.2013.04.001, PubMed:
23648277
Gorgolewski, K., Burns, C. D., Madison, C., clark, D., Halchenko,
Y. o., Waskom, METRO. l., & Ghosh, S. S. (2011). Nipype: A flexible,
lightweight and extensible neuroimaging data processing frame-
work in Python. Frontiers in Neuroinformatics, 22(5), 13. https://
doi.org/10.3389/fninf.2011.00013, PubMed: 21897815
Greve, D. NORTE., & pescado, B. (2009). Accurate and robust brain image
alignment using boundary-based registration. NeuroImagen, 48(1),
63–72. https://doi.org/10.1016/j.neuroimage.2009.06.060,
PubMed: 19573611
Hahn, B., ross, t. J., Cual, y., kim, I., Huestis, METRO. A., & piedra, mi. A.
(2007). Nicotine enhances visuospatial attention by deactivating
areas of the resting brain default network. Journal of Neurosci-
ence, 27(13), 3477–3489. https://doi.org/10.1523/JNEUROSCI
.5129-06.2007, PubMed: 17392464
Sala, S. A., Lalee, Z., Campana, R. PAG., Towe, S. l., & Meade, C. S. (2021).
Synergistic effects of HIV and marijuana use on functional brain
network organization. Progress in Neuro-Psychopharmacology
and Biological Psychiatry, 104, 110040. https://doi.org/10.1016
/j.pnpbp.2020.110040, PubMed: 32687963
harris, GRAMO. MI., Dupuis, l., Mugford, GRAMO. J., Johnston, l., Haase, D.,
Página, GRAMO., Haldane, h., harris, NORTE., Midodzi, W.. K., & dow, GRAMO.
(2014). Patterns and correlates of cannabis use among individ-
uals with HIV/AIDS in Maritime Canada. Canadian Journal of
Infectious Diseases and Medical Microbiology, 25(1), e1–e7.
https://doi.org/10.1155/2014/301713, PubMed: 24634690
Heaton, r., Clifford, D., franklin, D., Woods, S., Ake, C., Vaida, F.,
Ellis, r., Letendre, S., Marcotte, T., Atkinson, J., & Rivera-Mindt,
METRO. (2010). HIV-associated neurocognitive disorders persist in the
era of potent antiretroviral therapy: CHARTER Study. Neurología,
7 5 ( 2 3 ) , 2 0 8 7 – 2 0 9 6 . h t t p s : / / d o i . o r g / 1 0 . 1 2 1 2 / W N L
.0b013e318200d727, PubMed: 21135382
Hester, r., Foxe, j. J., Molholm, S., Shpaner, METRO., & Garavan, h.
(2005). Neural mechanisms involved in error processing: A com-
parison of errors made with and without awareness. NeuroImagen,
27(3), 602–608. https://doi.org/10.1016/j.neuroimage.2005.04
.035, PubMed: 16024258
Hester, r., Nandam, l. S., O’Connell, R. GRAMO., Wagner, J., Strudwick,
METRO., Nathan, PAG. J., Mattingley, j. B., & Bellgrove, METRO. A. (2012).
Neurochemical enhancement of conscious error awareness.
Revista de neurociencia, 32(8), 2619–2627. https://doi.org/10
.1523/JNEUROSCI.4052-11.2012, PubMed: 22357846
Hester, r., Nestor, l., & Garavan, h. (2009). Impaired error aware-
ness and anterior cingulate cortex hypoactivity in chronic canna-
bis users. Neuropsicofarmacología, 34(11), 2450–2458.
https://doi.org/10.1038/npp.2009.67, PubMed: 19553917
Hester, r., Simoes-Franklin, C., & Garavan, h. (2007). Post-error
behavior in active cocaine users: Poor awareness of errors in
the presence of intact performance adjustments. Neuropsycho-
pharmacology, 32(9), 1974–1984. https://doi.org/10.1038/sj.npp
.1301326, PubMed: 17268406
Hinds, o., Thompson, t. w., Ghosh, S., Yoo, j. J., Whitfield-
Gabrieli, S., Triantafyllou, C., & Gabrieli, j. D. (2013). Roles of
default-mode network and supplementary motor area in human
vigilance performance: Evidence from real-time fMRI. Diario de
Neurofisiología, 109(5), 1250–1258. https://doi.org/10.1152/jn
.00533.2011, PubMed: 23236006
Hinkin, C., van Gorp, w., Satz, PAG., Marcotte, T., Durvasula, r.,
Wood, S., Campbell, l., & Baluda, METRO. (1996). Actual versus
self-reported cognitive dysfunction in HIV-1 infection:
Memory-metamemory dissociations. Journal of Clinical and
Experimental Neuropsychology, 18(3), 431–443. https://doi.org
/10.1080/01688639608408999, PubMed: 8877626
Høgestøl, mi. A., Nygaard, GRAMO. o., Alnæs, D., Beyer, METRO. K., Westlye,
l. T., & Harbo, h. F. (2019). Symptoms of fatigue and depression
is reflected in altered default mode network connectivity in mul-
tiple sclerosis. PLoS ONE, 14(4), e0210375. https://doi.org/10
.1371/diario.pone.0210375, PubMed: 30933977
Neurociencia en red
811
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
hong, S., & Banks, W.. A. (2015). Role of the immune system in
HIV-associated neuroinflammation and neurocognitive implica-
ciones. Cerebro, Comportamiento, and Immunity, 45, 1–12. https://doi.org
/10.1016/j.bbi.2014.10.008, PubMed: 25449672
Ipser, j. C., Marrón, GRAMO. GRAMO., Bischoff-Grethe, A., Connolly, C. GRAMO., Ellis,
R. J., Heaton, R. K., Grant, I., & Translational Methamphetamine
AIDS Research Center Group. (2015). HIV infection is associated
with attenuated frontostriatal intrinsic connectivity: A preliminary
estudiar. Journal of the International Neuropsychological Society,
21(3), 203–213. https://doi.org/10.1017/S1355617715000156,
PubMed: 25824201
Jenkinson, METRO., Bannister, PAG., Brady, METRO., & Herrero, S. (2002).
Improved optimization for the robust and accurate linear registra-
tion and motion correction of brain images. NeuroImagen, 17(2),
825–841. https://doi.org/10.1006/nimg.2002.1132, PubMed:
12377157
jones, D. T., Knopman, D. S., Gunter, j. l., Graff-Radford, J.,
Vemuri, PAG., Boeve, B. F., Petersen, R. C., Weiner, METRO. w., Jacobo,
C. r., Jr., & Alzheimer’s Disease Neuroimaging Initiative.
(2016). Cascading network failure across the Alzheimer’s disease
spectrum. Cerebro, 139(punto. 2), 547–562. https://doi.org/10.1093
/brain/awv338, PubMed: 26586695
Sotavento, D., Sotavento, J., Sotavento, j. MI., & Jung, Y. C. (2017). Altered functional con-
nectivity in default mode network in Internet gaming disorder: Influ-
ence of childhood ADHD. Progress in Neuro-Psychopharmacology
and Biological Psychiatry, 75, 135–141. https://doi.org/10.1016/j
.pnpbp.2017.02.005, PubMed: 28174127
Sanguijuela, r., Kamourieh, S., beckman, C. F., & Sharp, D. j. (2011).
Fractionating the default mode network: Distinct contributions of
the ventral and dorsal posterior cingulate cortex to cognitive con-
controlar. Revista de neurociencia, 31(9), 3217–3224. https://doi.org
/10.1523/JNEUROSCI.5626-10.2011, PubMed: 21368033
Lerman, C., Gu, h., Loughead, J., Ruparel, K., Cual, y., & piedra,
mi. A. (2014). Large-scale brain network coupling predicts acute
nicotine abstinence effects on craving and cognitive function.
JAMA Psychiatry, 71(5), 523–530. https://doi.org/10.1001
/jamapsychiatry.2013.4091, PubMed: 24622915
li, C., yan, PAG., Bergquist, K., & Sinha, R. (2007). Greater activation
of the “default” brain regions predicts stop signal errors. Neuro-
Image, 38(3), 640–648. https://doi.org/10.1016/j.neuroimage
.2007.07.021, PubMed: 17884586
Liddle, mi. B., hollys, C., Batty, METRO. J., Groom, METRO. J., Totman, j. J.,
Liotti, METRO., Scerif, GRAMO., & Liddle, PAG. F. (2011). Task-related default
mode network modulation and inhibitory control in ADHD:
Effects of motivation and methylphenidate. Journal of Child Psy-
chology and Psychiatry, 52(7), 761–771. https://doi.org/10.1111/j
.1469-7610.2010.02333.X, PubMed: 21073458
Liu, X., Jiang, w., & Yuan, Y. (2018). Aberrant default mode network
underlying the cognitive deficits in the patients with late-onset
depresión. Frontiers in Aging Neuroscience, 10, 310. https://
doi.org/10.3389/fnagi.2018.00310, PubMed: 30337869
Loughead, J., Rayo, r., Wileyto, mi. PAG., Ruparel, K., Sanborn, PAG., Siegel,
S., Gur, R. C., & Lerman, C. (2010). Effects of the α4β2 partial
agonist varenicline on brain activity and working memory in
abstinent smokers. Biological Psychiatry, 67(8), 715–721. https://
doi.org/10.1016/j.biopsych.2010.01.016, PubMed: 20207347
Meade, C. S., Campana, R. PAG., Towe, S. l., Chen, norte. K., Hobkirk, A. l., &
Huettel, S. A. (2019). Synergistic effects of marijuana abuse and
HIV infection on neural activation during a cognitive interference
tarea. Addiction Biology, 24(6), 1235–1244. https://doi.org/10
.1111/adb.12678, PubMed: 30239074
menón, B. (2019). Towards a new model of understanding—The
triple network, psychopathology and the structure of the mind.
Medical Hypotheses, 133, 109385. https://doi.org/10.1016/j
.mehy.2019.109385, PubMed: 31494485
menón, V. (2011). Large-scale brain networks and psychopathol-
ogia: A unifying triple network model. Trends in Cognitive Sci-
ences, 15(10), 483–506. https://doi.org/10.1016/j.tics.2011.08
.003, PubMed: 21908230
menón, v., & Uddin, l. q. (2010). Saliency, switching, atención
and control: A network model of insula function. Brain Structure
and Function, 214(5–6), 655–667. https://doi.org/10.1007
/s00429-010-0262-0, PubMed: 20512370
Minosse, S., Picchi, MI., Di Giuliano, F., Sarmati, l., Teti, MI.,
Pistolese, C. A., Lanzafame, S., Di Cio, F., Guerrisi, METRO., Andreoni,
METRO., Floris, r., Toschi, NORTE., & Garaci, F. (2021). Functional brain
network reorganization in HIV infection. Journal of Neuroimaging,
31(4), 796–808. https://doi.org/10.1111/jon.12861, PubMed:
33900655
Montgomery, l., Bagot, K., Marrón, j. l., & Haeny, A. METRO. (2019). El
association between marijuana use and HIV continuum of care
resultados: A systematic review. Current HIV/AIDS Reports, 16(1),
17–28. https://doi.org/10.1007/s11904-019-00422-z, PubMed:
30671919
Moradi, METRO., Ekhtiari, h., Kuplicki, r., McKinney, B., Stewart, j. l.,
Victor, t. A., Paulus, METRO. PAG., & Tulsa, I. (2020). Evaluating the
resource allocation index as a potential fMRI-based biomarker
for substance use disorder. Drug and Alcohol Dependence,
216, 108211. https://doi.org/10.1016/j.drugalcdep.2020
.108211, PubMed: 32805548
morgan, mi. MI., Iudicello, j. MI., Weber, MI., Duarte, norte. A., Riggs, PAG. K.,
Delano-Wood, l., Ellis, r., Grant, I., Woods, S. PAG., & HNRP
Group. (2012). Synergistic effects of HIV infection and older
age on daily functioning. Journal of Acquired Immune Deficiency
Syndrome, 61(3), 341–348. https://doi.org/10.1097/QAI
.0b013e31826bfc53, PubMed: 22878422
Mothobi, norte. Z., & Brew, B. j. (2012). Neurocognitive dysfunction in
the highly active antiretroviral therapy era. Opinión actual en
Infectious Diseases, 25(1), 4–9. https://doi.org/10.1097/QCO
.0b013e32834ef586, PubMed: 22156897
Muschelli, J., Nebel, METRO. B., Caffo, B. S., Barber, A. D., Pekar, j. J., &
Mostofsky, S. h. (2014). Reduction of motion-related artifacts in
resting state fMRI using aCompCor. NeuroImagen, 96, 22–35.
https://doi.org/10.1016/j.neuroimage.2014.03.028, PubMed:
24657780
Okafor, C., Plankey M. w., li, METRO., Chen, X., Surkan, PAG. J., Shoptaw,
S., Martín, MI., cohen, r., Sacktor, NORTE., & Cocinar, R. l. (2019). también-
ciation of marijuana use with changes in cognitive processing
speed and flexibility for 17 years in HIV-seropositive and
HIV-seronegative men. Substance Use and Misuse, 54(4),
525–537. https://doi.org/10.1080/10826084.2018.1495736,
PubMed: 30700235
Oyegbile, t. o., VanMeter, j. w., Motamedi, GRAMO. K., Campana, W.. l.,
Gaillard, W.. D., & Hermann, B. PAG. (2019). Default mode network
deactivation in pediatric temporal lobe epilepsy: Relationship to
a working memory task and executive function tests. Epilepsy
Neurociencia en red
812
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
and Behavior, 94, 124–130. https://doi.org/10.1016/j.yebeh
.2019.02.031, PubMed: 30909075
Pacek, l. r., Towe, S. l., Hobkirk, A. l., Nash, D., & Goodwin,
R. D. (2018). Frequency of cannabis use and medical cannabis
use among persons living with HIV in the United States: Findings
from a nationally representative sample. AIDS Education and Pre-
vention, 30(2), 169–181. https://doi.org/10.1521/aeap.2018.30.2
.169, PubMed: 29688777
Pannunzi, METRO., Hindriks, r., Bettinardi, R. GRAMO., Wenger, MI., Lisofsky,
NORTE., Martensson, J., mayordomo, o., Filevich, MI., Becker, METRO., Lochstet,
METRO., & Kühn, S. (2017). Resting-state fMRI correlations: De
link-wise unreliability to whole brain stability. NeuroImagen,
157, 250–262. https://doi.org/10.1016/j.neuroimage.2017.06
.006, PubMed: 28599964
Peterson, B. S., Potenza, METRO. NORTE., Wang, Z., Zhu, h., Martín, A.,
Marsh, r., Plessen, k. J., & Yu, S. (2009). An fMRI study of the
effects of psychostimulants on default-mode processing during
Stroop task performance in youths with ADHD. American Journal
of Psychiatry, 166(11), 1286–1294. https://doi.org/10.1176/appi
.ajp.2009.08050724, PubMed: 19755575
Fuerza, j. D., Barnes, k. A., Snyder, A. Z., Schlaggar, B. l., &
Petersen, S. mi. (2012). Spurious but systematic correlations in
functional connectivity MRI networks arise from subject motion.
NeuroImagen, 59(3), 2142–2154. https://doi.org/10.1016/j
.neuroimage.2011.10.018, PubMed: 22019881
Fuerza, j. D., Mitra, A., Laumann, t. o., Snyder, A. Z., Schlaggar,
B. l., & Petersen, S. mi. (2013). Methods to detect, characterize,
and remove motion artifact in resting state fMRI. NeuroImagen,
84, 320–341. https://doi.org/10.1016/j.neuroimage.2013.08
.048, PubMed: 23994314
Querne, l., Caer, S., Le Moing, A. GRAMO., Bourel-Ponchel, MI.,
Delignières, A., Simonnot, A., de Broca, A., Gondry-Jouet, C.,
Boucart, METRO., & Berquin, PAG. (2017). Effects of methylphenidate
on default-mode network/task-positive network synchronization
in children with ADHD. Journal of Attention Disorders, 21(14),
1208–1220. https://doi.org/10.1177/1087054713517542,
PubMed: 24420764
Reese, mi. D., Hacer, j. y., McKay, k. GRAMO., piedra, mi. A., ross, t. J., &
Daughters, S. B. (2019). Triple network resting state connectivity
predicts distress tolerance and is associated with cocaine use.
Journal of Clinical Medicine, 8(12), 2135. https://doi.org/10
.3390/jcm8122135, PubMed: 31817047
Rich, S., Klann, MI., Bryant, v., Richards, v., Wijayabahu, A., Bryant,
K., Mai, v., & Cocinar, R. (2020). A review of potential
microbiome-gut-brain axis mediated neurocognitive conditions
in persons living with HIV. Cerebro, Comportamiento, and Immunity—
Salud, 9, 100168. https://doi.org/10.1016/j.bbih.2020.100168,
PubMed: 34589903
Rizzo, METRO. D., Crawford, R. B., Henriquez, j. MI., Aldhamen, Y. A.,
Gulick, PAG., Amalfitano, A., & Kaminski, norte. mi. (2018). HIV-infected
cannabis users have lower circulating CD16+ monocytes and
IFN-gamma-inducible protein 10 levels compared with nonusing
HIV patients. AIDS, 32(4), 419–429. https://doi.org/10.1097
/QAD.0000000000001704, PubMed: 29194121
Robertson, K., Landay, A., Miyahara, S., Vecchio, A., Maestros,
METRO. C., Marrón, t. T., & Taiwo, B. oh. (2020). Limited correlation
between systemic biomarkers and neurocognitive performance
before and during HIV treatment. Journal of NeuroVirology,
26(1), 107–113. https://doi.org/10.1007/s13365-019-00795-2,
PubMed: 31468473
Salmi, J., Nyberg, l., & Laine, METRO. (2018). Working memory training
mostly engages general-purpose large-scale networks for learn-
En g. Revisiones de neurociencia y biocomportamiento, 93, 108–122.
https://doi.org/10.1016/j.neubiorev.2018.03.019, PubMed:
29574197
Saloner, r., Heaton, R. K., Campbell, l. METRO., Chen, A., franklin, D.,
Jr., Ellis, R. J., Minero, A. C., Marra, C., Clifford, D. B., Gelman, B.,
Sacktor, NORTE., Morgello, S., McCutchan, j. A., Letendre, S., Grant,
I., & Fennema-Notestine, C. (2019). Effects of comorbidity bur-
den and age on brain integrity in HIV. AIDS, 33(7), 1175–1185.
https://doi.org/10.1097/QAD.0000000000002192, PubMed:
30870195
Schilbach, l., Hoffstaedter, F., Müller, v., Cieslik, mi. C., Goya-
Maldonado, r., Trost, S., Sorg, C., Riedl, v., Jardri, r., verano,
I., & Kogler, l. (2016). Transdiagnostic commonalities and dif-
ferences in resting state functional connectivity of the default
mode network in schizophrenia and major depression. Neuro-
Image: Clinical, 10, 326–335. https://doi.org/10.1016/j.nicl
.2015.11.021, PubMed: 26904405
Seeley, W.. w., menón, v., Schatzberg, A. F., Keller, J., guantero,
GRAMO. h., Kenna, h., Reiss, A. l., & Greicius, METRO. D. (2007). Disso-
ciable intrinsic connectivity networks for salience processing and
executive control. Revista de neurociencia, 27(9), 2349–2356.
https://doi.org/10.1523/ JNEUROSCI.5587-06.2007, PubMed:
17329432
Seider, t. r., Gongvatana, A., Woods, A. J., Chen, h., Porges, mi. C.,
Cummings, T., Correia, S., Tashima, K., & cohen, R. A. (2016).
Age exacerbates HIV-associated white matter abnormalities.
Journal of NeuroVirology, 22(2), 201–212. https://doi.org/10
.1007/s13365-015-0386-3, PubMed: 26446690
Shirer, w., Ryali, S., Rykhlevskaia, MI., menón, v., & Greicius, METRO.
(2012). Decoding subject-driven cognitive states with whole-
brain connectivity patterns. Corteza cerebral, 22(1), 158–165.
https://doi.org/10.1093/cercor/bhr099, PubMed: 21616982
Shoaib, METRO., & Bizarro, l. (2005). Deficits in a sustained attention
task following nicotine withdrawal in rats. Psicofarmacología,
178(2–3), 211–222. https://doi.org/10.1007/s00213-004-2004-6,
PubMed: 15338107
singh, k. D., & Fawcett, I. PAG. (2008). Transient and linearly graded
deactivation of the human default-mode network by a visual
detection task. NeuroImagen, 41(1), 100–112. https://doi.org/10
.1016/j.neuroimage.2008.01.051, PubMed: 18375149
Herrero, S. METRO., Fox, PAG. T., Molinero, k. l., Glahn, D. C., Fox, PAG. METRO.,
Mackay, C. MI., Filippini, NORTE., watkins, k. MI., Toro, r., Laird,
A. r., & beckman, C. F. (2009). Correspondence of the brain’s
functional architecture during activation and rest. Actas de
the National Academy of Sciences, 106(31), 13040–13045.
https://doi.org/10.1073/pnas.0905267106, PubMed: 19620724
Sridharan, D., Levitin, D. J., & menón, V. (2008). A critical role
for the right fronto-insular cortex in switching between
central-executive and default-mode networks. Actas de
the National Academy of Sciences, 105(34), 12569–12574.
https://doi.org/10.1073/pnas.0800005105, PubMed: 18723676
stevens, METRO. C. (2016). The contributions of resting state and
task-based functional connectivity studies to our understanding
of adolescent brain network maturation. Neurociencia y
Neurociencia en red
813
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
Revisiones de biocomportamiento, 70, 13–32. https://doi.org/10.1016/j
.neubiorev.2016.07.027, PubMed: 27502750
Sutherland, METRO. T., McHugh, METRO. J., Pariyadath, v., & piedra, mi. A.
(2012). Resting state functional connectivity in addiction:
Lessons learned and a road ahead. NeuroImagen, 62(4),
2281–2295. https://doi.org/10.1016/j.neuroimage.2012.01.117,
PubMed: 22326834
Sutherland, METRO. T., & piedra, mi. A. (2018). Functional neurocircuits
and neuroimaging biomarkers of tobacco use disorder. Trends
in Molecular Medicine, 24(2), 129–143. https://doi.org/10.1016
/j.molmed.2017.12.002, PubMed: 29398401
Sutherland, METRO. T., Xia, l., Yihong, y., & piedra, mi. (2015). Beyond
functional localization: Advancing the understanding of
addiction-related processes by examining brain connectivity. En
S. j. wilson (Ed.), The Wiley Handbook on the Cognitve Neuro-
science of Addicition (1st ed.). John Wiley & Sons. https://doi.org
/10.1002/9781118472415.ch20
Tanabe, J., Nyberg, MI., Martín, l. F., Martín, J., Cordes, D.,
Kronberg, MI., & Tregellas, j. R. (2011). Nicotine effects on default
mode network during resting state. Psicofarmacología, 216(2),
287–295. https://doi.org/10.1007/s00213-011-2221-8, PubMed:
21331518
Thames, A. D., Mahmood, Z., Burggren, A. C., Karimian, A., &
Kuhn, t. PAG. (2016). Combined effects of HIV and marijuana use
on neurocognitive functioning and immune status. AIDS Care,
28(5), 628–632. https://doi.org/10.1080/09540121.2015
.1124983, PubMed: 26694807
tomás, j. B., Brier, METRO. r., Ortega, METRO., Benzinger, t. l., & Ances,
B. METRO. (2015). Weighted brain networks in disease: Centrality and
entropy in human immunodeficiency virus and aging. Neurobi-
ology of Aging, 36(1), 401–412. https://doi.org/10.1016/j
.neurobiolaging.2014.06.019, PubMed: 25034343
Tournier, j. D., Herrero, r., Raffelt, D., Tabbara, r., Dhollander, T.,
Pietsch, METRO., Christiaens, D., Jeurissen, B., Yeh, C. h., & connelly,
A. (2019). MRtrix3: A fast, flexible and open software framework
for medical image processing and visualisation. NeuroImagen, 202,
116137. https://doi.org/10.1016/j.neuroimage.2019.116137,
PubMed: 31473352
Towe, S. l., Meade, C. S., Cloak, C. C., Campana, R. PAG., Baptiste, J., &
Chang, l. (2020). Reciprocal influences of HIV and cannabinoids
on the brain and cognitive function. Journal of Neuroimmune
Pharmacology, 15, 756–779. https://doi.org/10.1007/s11481
-020-09921-y, PubMed: 32445005
Tustison, norte. J., Avants, B. B., Cocinar, PAG. A., Zheng, y., Egan, A.,
Yushkevich, PAG. A., & Gee, j. C. (2010). N4ITK: Improved N3
bias correction. IEEE Transactions on Medical Imaging, 29(6),
1310–1320. https://doi.org/10.1109/ TMI.2010.2046908,
PubMed: 20378467
Valcour, v., Pablo, r., Neuhaus, J., & Shikuma, C. (2011). The effects
of age and HIVon neuropsychological performance. Journal of the
International Neuropsychological Society, 17(1), 190–195. https://
doi.org/10.1017/S1355617710001438, PubMed: 21144102
Van Gorp, w., Satz, PAG., Hinkin, C., Selnes, o., Molinero, MI., McArthur,
J., cohen, B., & Paz, D. (1991). Metacognition in HIV-1 sero-
positive asymptomatic individuals: Self-ratings versus objective
neuropsychological performance. Journal of Clinical and Exper-
imental Neuropsychology, 13(5), 812–819. https://doi.org/10
.1080/01688639108401091, PubMed: 1955533
Vance, D. MI., Farr, k. F., & Struzick, t. (2008). Assessing the clinical
value of cognitive appraisal in adults aging with HIV. Diario de
Gerontological Nursing, 34(1), 36–41. https://doi.org/10.3928
/00989134-20080101-11, PubMed: 18274303
Veraart, J., Fieremans, MI., & Nóvikov, D. S. (2016). Diffusion MRI
noise mapping using random matrix theory. Resonancia magnetica
En medicina, 76(5), 1582–1593. https://doi.org/10.1002/mrm
.26059, PubMed: 26599599
Veraart, J., Nóvikov, D. S., Christiaens, D., Ades-Aron, B., Sijbers, J.,
& Fieremans, mi. (2016). Denoising of diffusion MRI using random
matrix theory. NeuroImagen, 142, 394–406. https://doi.org/10
.1016/j.neuroimage.2016.08.016, PubMed: 27523449
Verfaillie, S. C., Binette, A. PAG., Vachon-Presseau, MI., Tabrizi, S.,
Savard, METRO., Bellec, PAG., Ossenkoppele, r., Scheltens, PAG., van der
Flier, W.. METRO., Breitner, j. C., & Villeneuve, S. (2018). Subjective
cognitive decline is associated with altered default mode net-
work connectivity in individuals with a family history of Alzhei-
mer’s disease. Biological Psychiatry: Cognitive Neuroscience and
Neuroimaging, 3(5), 463–472. https://doi.org/10.1016/j.bpsc
.2017.11.012, PubMed: 29735156
Wang, K., Xu, METRO., Ji, y., zhang, l., Du, X., li, J., luo, P., & li, F.
(2019). Altered social cognition and connectivity of default
mode networks in the co-occurrence of autistic spectrum
disorder and attention deficit hyperactivity disorder. Australian
and New Zealand Journal of Psychiatry, 53(8), 760–771.
https://doi.org/10.1177/0004867419836031, PubMed:
30843728
Wang, y., Risacher, S. l., Oeste, j. D., McDonald, B. C., MaGee, t. r.,
Farlow, METRO. r., gao, S., O’Neill, D. PAG., & Saykin, A. j. (2013).
Altered default mode network connectivity in older adults with
cognitive complaints and amnestic mild cognitive impairment.
Journal of Alzheimer’s Disease, 35(4), 751–760. https://doi.org
/10.3233/JAD-130080, PubMed: 23481685
watson, C., Paolillo E. w., morgan, mi. MI., Umlauf, A., Sundermann,
mi. MI., Ellis, R. J., Letendre, S., Marcotte, t. D., Heaton, R. K., &
Grant, I. (2020). Cannabis exposure is associated with a lower
likelihood of neurocognitive impairment in people living with
HIV. Journal of Acquired Immune Deficiency Syndromes and
Human Retrovirology, 83(1), 56–64. https://doi.org/10.1097
/QAI.0000000000002211, PubMed: 31809361
Wechsler, D. (2008). Wechsler Adult Intelligence Scale–Fourth
edition ( WAIS–IV ) (volumen. 22). NCS Pearson. https://doi.org/10
.1037/t15169-000
Wendelken, l. A., & Valcour, V. (2012). Impact of HIV and aging on
neuropsychological function. Journal of NeuroVirology, 18(4),
256–263. https://doi.org/10.1007/s13365-012-0094-1, PubMed:
22528478
Wesnes, k. A., Edgar, C. J., Kezic, I., Salih, h. METRO., & de Boer, PAG.
(2013). Effects of nicotine withdrawal on cognition in a clinical
trial setting. Psicofarmacología, 229(1), 133–140. https://doi
.org/10.1007/s00213-013-3089-6, PubMed: 23591603
Whitfield-Gabrieli, S., & Vado, j. METRO. (2012). Default mode network
activity and connectivity in psychopathology. Annual Review of
Clinical Psychology, 8, 49–76. https://doi.org/10.1146/annurev
-clinpsy-032511-143049, PubMed: 22224834
williams, METRO. MI., Ipser, j. C., piedra, D. J., Joska, j. A., & Naude,
PAG. j. W.. (2020). Peripheral immune dysregulation in the ART
era of HIV-associated neurocognitive impairments: A systematic
Neurociencia en red
814
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Large-scale brain network interactions associated with HIV infection
revisar. Psychoneuroendocrinology, 118, 104689. https://doi.org
/10.1016/j.psyneuen.2020.104689, PubMed: 32479968
Woods, S. PAG., moore, D. J., Weber, MI., & Grant, I. (2009). Cognitivo
neuropsychology of HIV-associated neurocognitive disorders.
Neuropsychology Review, 19(2), 152–168. https://doi.org/10
.1007/s11065-009-9102-5, PubMed: 19462243
Wu, X., li, r., Fleisher, A. S., Reiman, mi. METRO., Guan, X., zhang, y., Chen,
K., & Yao, l. (2011). Altered default mode network connectivity in
Alzheimer’s disease—A resting functional MRI and Bayesian net-
work study. Mapeo del cerebro humano, 32(11), 1868–1881. https://
doi.org/10.1002/hbm.21153, PubMed: 21259382
Yin, y., Él, X., Xu, METRO., Hou, Z., Song, X., Sui, y., Liu, Z., Jiang, w.,
yue, y., zhang, y., & Liu, Y. (2016). Structural and functional con-
nectivity of default mode network underlying the cognitive
impairment in late-onset depression. Informes Científicos, 6, 37617.
https://doi.org/10.1038/srep37617, PubMed: 27886212
zhang, j. T., Mamá, S. S., yan, C. GRAMO., zhang, S., Liu, l., Wang, l. J.,
Liu, B., Yao, Y. w., Cual, Y. h., & Fang, X. Y. (2017). Altered
coupling of default-mode, executive-control and salience
networks in Internet gaming disorder. European Psychiatry, 45,
114–120. https://doi.org/10.1016/j.eurpsy.2017.06.012,
PubMed: 28756109
zhang, r., & Volkow, norte. D. (2019). Brain default-mode network dys-
function in addiction. NeuroImagen, 200, 313–331. https://doi.org
/10.1016/j.neuroimage.2019.06.036, PubMed: 31229660
zhou, l., PU, w., Wang, J., Liu, h., Wu, GRAMO., Liu, C., Mwansisya,
t. MI., tao, h., Chen, X., Huang, X., Lv, D., Xue, Z., shan, B., &
Liu, z. (2016). Inefficient DMN suppression in schizophrenia
patients with impaired cognitive function but not patients with
preserved cognitive function. Informes Científicos, 6, 21657.
https://doi.org/10.1038/srep21657, PubMed: 26882844
Zhuang, y., Qiu, X., Wang, l., Mamá, P., Mapstone, METRO., Luque, A.,
Weber, METRO., Tivarus, METRO., Molinero, MI., Arduino, R. C., Zhong, J., &
Schifitto, GRAMO. (2017). Combination antiretroviral therapy improves
cognitive performance and functional connectivity in treatment-
naive HIV-infected individuals. Journal of NeuroVirology, 23(5),
704–712. https://doi.org/10.1007/s13365-017-0553-9, PubMed:
28791662
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
6
3
7
9
1
2
0
3
5
9
7
9
norte
mi
norte
_
a
_
0
0
2
4
1
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurociencia en red
815