INVESTIGACIÓN
The missing role of gray matter in studying
brain controllability
Hamidreza Jamalabadi1∗
, Agnieszka Zuberer1,2,3,7∗
Meng Li6,7, Sarah Alizadeh1, Ali Moradi Amani8, Christian Gaser7,9,
Michael Esterman2,3,10, and Martin Walter1,4,5,6,7
, Vinod Jangir Kumar6,
1Department of Psychiatry and Psychotherapy, University of Tübingen, Tübingen, Alemania
2Boston University School of Medicine, Department of Psychiatry, Bostón, MAMÁ, EE.UU
3Boston Attention and Learning Laboratory, VA Boston Healthcare System, Bostón, MAMÁ, EE.UU
4Clinical Affective Neuroimaging Laboratory, Magdeburg, Alemania
5Leibniz Institute for Neurobiology, Magdeburg, Alemania
6Max Planck Institute for Biological Cybernetics, Tübingen, Alemania
7Department of Psychiatry and Psychotherapy, Jena University Hospital, Jena, Alemania
8School of Engineering, RMIT University, Melbourne, Victoria, Australia
9Department of Neurology, Jena University Hospital, Jena, Alemania
10National Center for PTSD, VA Boston Healthcare System, Bostón, MAMÁ, EE.UU
*These authors contributed equally.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
norte
mi
norte
_
a
_
0
0
1
7
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Palabras clave: Network control theory, Gray matter, Brain controllability
ABSTRACTO
Brain controllability properties are normally derived from the white matter fiber tracts in
which the neural substrate of the actual energy consumption, namely the gray matter, tiene
been widely ignored. Aquí, we study the relationship between gray matter volume of regions
across the whole cortex and their respective control properties derived from the structural
architecture of the white matter fiber tracts. The data suggests that the ability of white fiber
tracts to exhibit control at specific nodes not only depends on the connection strength of the
structural connectome but additionally depends on gray matter volume at the host nodes.
Our data indicate that connectivity strength and gray matter volume interact with respect to
the brain’s control properties. Disentangling effects of the regional gray matter volume and
connectivity strength, we found that frontal and sensory areas play crucial roles in
controllability. Together these results suggest that structural and regional properties of the
white matter and gray matter provide complementary information in studying the control
properties of the intrinsic structural and functional architecture of the brain.
RESUMEN DEL AUTOR
Network control theory suggests that the functions of large-scale brain circuits can be
partially described with respect to the ability of brain regions to steer brain activity to
different states. This ability, often quantified in terms of controllability metrics, has normally
been derived from the structural architecture of the white matter fiber tracts. Sin embargo, gray
matter as the substrate that engenders much of the neural processes is widely ignored in this
contexto. In the present work, we study the relationship between regional gray matter volume
and control properties across the whole cortex and provide evidence that control properties
not only depend on the connection strength of the structural connectome but also depend on
sufficient gray matter volume at the host nodes.
un acceso abierto
diario
Citación: Jamalabadi, h., Zuberer, A.,
Kumar, V. J., li, METRO., Alizadeh, S., Amani,
A. METRO., … Walter, METRO. (2021). The missing
role of gray matter in studying brain
controllability. Neurociencia en red,
5(1), 198–210. https://doi.org/10.1162
/netn_a_00174
DOI:
https://doi.org/10.1162/netn_a_00174
Supporting Information:
https://doi.org/10.1162/netn_a_00174
Recibió: 7 Abril 2020
Aceptado: 23 Octubre 2020
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autores correspondientes:
Hamidreza Jamalabadi
hamidreza.jamalabadi@uni-tuebingen.de
Agnieszka Zuberer
azuberer@bu.edu
Editor de manejo:
Olaf Sporns
Derechos de autor: © 2020
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
The missing role of gray matter in studying brain controllability
Network control theory:
A mathematical approach to
combine control and network theory.
Network control theory can be used
to describe dynamics of any system
that involves interconnected units,
and it allows making predictions
about the behavior of the system after
perturbation.
Controllability:
The property of a dynamical system
that quantifies the ability of external
inputs to steer the system dynamics
toward any desired state.
INTRODUCCIÓN
Network control theory, as recently applied to white matter (W.M.) fiber tracts in the human
cerebro, provides a novel mechanistic framework to describe the ease of switching between
different dynamical functional brain states, and the regions that best drive these dynamics
(bassett & despreciar, 2017; Medalla, 2019; Medalla, Pasqualetti, hamilton, Thompson-Schill,
& bassett, 2017). This approach has the potential to inform theories of dynamic cognitive
procesos, clinical neuroscience, neurodegeneration, and brain reserve. Específicamente, hay
evidence that these global brain state transitions are impaired in clinical populations (Braun
et al., 2019; Jeganathan et al., 2018; Kenett, Beaty, & Medalla, 2018) and that such impair-
ments can be traced back to specific driver nodes (Jeganathan et al., 2018; Kenett,
Beaty, et al., 2018; Muldoon et al., 2016; Zoeller et al., 2019). Sin embargo, thus far, estos
control properties have been exclusively derived from WM fiber tracts without the consider-
ation of gray matter (GM) propiedades. Given the importance of GM properties for cognitive
functioning and brain health, and the established interrelationships between white and gray
asunto, it has been suggested that regional gray matter integrity may be a critical contributor
and proxy for network and node controllability (Medalla, Pasqualetti, et al., 2017; Medalla,
Zurn, Sinnott-Armstrong, & bassett, 2017).
Several lines of research suggest that GM may be essential to understanding brain control-
lability. Primero, GM is a proxy for the quantity of neurons and synaptic densities in a particular
región (Lüders, Steinmetz, & Jäncke, 2002), and metabolic energy expenditure is primarily re-
alized through the gray matter cell bodies that scaffold white matter tracts (Zhu et al., 2012). En
some neurodegenerative disorders, region-specific lesions of GM only partially agree with cor-
responding lesions in WM (Agosta et al., 2011; Bodini et al., 2009; Douaud et al., 2007; Raine,
Lencz, Bihrle, LaCasse, & Colletti, 2000; Villain et al., 2008), suggesting that GM reserve and
WM may provide independent additional information with respect to controllability properties
of the structural connectome. Tomados juntos, these studies motivate the hypothesis that the
controllability properties suggested by the WM should be partially related to or even predicted
by GM integrity. críticamente, it has been argued that including GM metrics in control theory will
extend traditional volumetrics into network neuroscience (Medalla, Pasqualetti, et al., 2017).
Sin embargo, to our knowledge the nature of the interdependence of controllability properties
and GM properties has not been addressed empirically yet.
To tackle this issue, we used two independent datasets to investigate whether—and if so,
how—control properties extracted from the structural connectome relate to properties of the
gray matter, eso es, GM volume that engenders other GM metrics such as surface and thickness
(Kong et al., 2015; Winkler et al., 2010). Since previous studies have shown that brain control-
lability can be largely explained by the connectivity strength of the structural connectome, nosotros
also considered whether GM volume could explain additional variance in controllability not
accounted for by white matter connectivity. Initially, we investigated how WM and GM fac-
tors affect brain controllability on a whole-brain level. In a further step, we identified the brain
regions for which controllability was most sensitive to GM and/or WM properties. We discuss
our findings with respect to their potential relevance to cognitive and clinical neuroscience.
METHODS AND MATERIALS
Data Acquisition
The structural and diffusion datasets are from 65 healthy subjects with the age range of 22 a
36 (28 METRO, mean age 29.2), which were taken from the Human Connectome Project (HCP,
Neurociencia en red
199
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
norte
mi
norte
_
a
_
0
0
1
7
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The missing role of gray matter in studying brain controllability
principal investigators: David Van Essen and Kamil Ugurbil; 1U54MH091657; VanEssen
et al., 2012). While HCP offers more than 1,100 subjects, the data in the present study are
limited by the resources necessary for preprocessing. We have tried to lift the potential bias by
including an independent dataset (see the Replication Study section).
Structural images were acquired with the following specification: T1w
MRI data specification.
MPRAGE, TR 2,400 EM, EL 2.14 EM, TI 1,000 EM, flip angle 8 degrees, field of view (FOV)
224 × 224, 256 slices, voxel size 0.7 mm isotropic, bandwidth 210 Hz/Px, IPAT 2, adquisición
tiempo 7:40 mín..
Diffusion-weighted imaging (DWI) data were acquired by using a spin-echo EPI sequence
with TR 5,520 EM, EL 89.5 EM, flip angle 78 degrees, voxel size, 1.25 mm isotropic, 111
slices, multiband factor, 3, echo spacing, 0.78 EM, b-values 1,000, 2,000, y 3,000 s/mm2.
Para más detalles, see Glasser et al. (2013) and Van Essen et al. (2012).
The 3D
Automated anatomical labeling (AAL) mask definitions and native space transformation.
anatomy atlas of the AAL2 was acquired from the Neurofunctional Imaging Group (http://www
.gin.cnrs.fr/en/tools/aal-aal2/; Tzourio-Mazoyer et al., 2002). It contains 120 regiones, cual
include subcortical structures such as thalamus, caudate, putamen, and pallidum. Sin embargo, él
misses the brain stem. The 12-parameter affine transformation (Jenkinson, Bannister, Brady, &
Herrero, 2002; Jenkinson & Herrero, 2001) was computed for each volunteer’s T1 and nondiffusion
image and the MNI space standard brain. The resulting transformation matrix was applied to
the left and right AAL brain regions to transform them into the native structural and diffusion
espacio.
The tissue-type segmentation employed SPM12 unified segmenta-
Structural volume analysis.
tion approach. The process resulted in segmented gray, white, and cerebrospinal fluid (CSF)
volumes.
In the next step, we determined the volume of the brain, gray matter, and under
each AAL atlas region for all subjects. The skull-extracted AC-PC aligned native space NIFTI
structural scans were obtained from the Human Connectome database. In the next step, el
tissue-type segmentation was applied to delineate the gray matter within the brain using the
SPM12 unified segmentation approach (Ashburner & Friston, 2005). This segmentation ap-
proach employs a generative model that combines nonlinear registration, tissue classification,
and bias correction.
The obtained HCP diffusion data were reconstructed using a
Preprocessing and diffusion fit.
SENSE1 algorithm (Sotiropoulos et al., 2013). The DWI data were corrected for motion and
distorsión (andersson, Skare, & Ashburner, 2003; andersson & Sotiropoulos, 2015, 2016).
Además, preprocessing included unringing, denoising, and tensor analysis implemented
in MRtrix (Tournier, Calamante, & connelly, 2012).
The data were reconstructed using the multi-shell multi-tissue constrained spherical decon-
volution (Jeurissen, Tournier, Dhollander, connelly, & Sijbers, 2014). The resulting orientation
distribution function (ODF) was registered to the structural space. The initial tractogram was
generated using mrtrix-tckgen, Resultando en 100 million streamlines within each subject. En el
next step, we applied spherical deconvolution informed filtering of tractograms (SIFT) to reduce
the streamline count to 10 millón. In the final step, the number of streamlines was determined
between AAL brain regions to produce a connectome. The analysis steps are documented in
Neurociencia en red
200
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
norte
mi
norte
_
a
_
0
0
1
7
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The missing role of gray matter in studying brain controllability
more detail at the MRtrix docs (https://mrtrix.readthedocs.io/en/latest/quantitative_structural
_connectivity/structural_connectome.html).
Network Control Framework
Controllability is one of the fundamental concepts in the control theory. The notion of control-
lability of a dynamical system was first introduced in Kalman (1963). Estado (producción) controlla-
bility of a dynamical system is defined as the possibility of driving states (outputs) del sistema
from an arbitrary initial condition to any desired values in finite time by applying appropriate
control signals (Kailath, 1980). The most famous classic method to ensure state controllability
of a dynamical system defined by the noise-free linear discrete-time and time-invariant network
model says that the system
X(k + 1) = Ax(k) + Bu(k)
y(k) = Cx(k) + Du(k)
(1)
(2)
is full state controllable if and only if the Kalman’s controllability matrix [B, AB, . . . , An−1B]
In the system represented in Equations 1 y 2, x ∈ Rn and
has full rank (Kailath, 1980).
u ∈ R p are state and input signals, respectivamente. A, B, C, and D are matrices with appropriate
dimensions, where A and B are called state and input matrices, respectivamente. When applied in
the context of brain controllability, x describes the activity of brain regions. A is an adjacency
matrix that represents the interactions between brain regions, and its elements are often the
strength of the white tracts connecting two areas (see the Statistical Analysis section below for
details). The input matrix B identifies the control nodes in the brain that may be confined to one
or more brain areas, whose activities are denoted by the corresponding elements of x. Mientras
the controllability matrix is a valuable metric to study the overall character of a system, it does
not directly quantify the potential ability of different nodes of the system to act as driver nodes.
To achieve this, a common practice is to use Tr(Wk), which is the trace of the controllability
Gramian Wk = ∑∞
i=0 AiBBT(AT)i when the system is controlled from node k. Referred to as
average controllability (AC), this metric is the most commonly used controllability measure in
the neuroimaging literature (Gu et al., 2015; Medalla, 2019); it is a measure of the average
energy required for node k to steer the brain into all possible output states (see Tang & bassett,
2018, for a formal definition). In addition to AC that quantifies the ability of the nodes to drive
the system into all potential target states, modal controllability (MC) is another commonly used
métrico; it is a measure of the ability of the nodes to push the system toward difficult-to-reach
estados. Formally defined as φk = ∑N
j (A)]v2
kj, MC is a scaled measure of difficulty of
driving the system toward all N modes of A from node k (Pasqualetti, Zampieri, & Bullo, 2014).
j [1 − ξ2
Statistical Analysis
Linear mixed-effects (LME) regression (Baayen, Davidson, & Bates, 2008) allows us to model
the interrelationship among multiple variables and has the ability to accommodate various ex-
perimental designs, including repeated measurements, subject variability, and grouping struc-
turas, in one unified implementation (Boisgontier & Cheval, 2016). en este documento, we model
the interrelationship between brain controllability (AC and MC), GM volume, and connectiv-
ity strength, for which we built multiple LMEs. En particular, we include regional gray matter
(rGM) and total intracranial volume (TIV) in the LMEs.
To predict brain controllability metrics based on structural measures of the brain, we built
a linear mixed-effects regression (Baayen et al., 2008) using a stepwise approach, retaining
an effect only if there was a significant difference between the log-likelihood ratio of the two
Average controllability:
A controllability metric that
quantifies the ability of nodes to steer
the network dynamics toward
easy-to-reach brain states.
Modal controllability:
A controllability metric that
quantifies the ability of nodes to steer
the network dynamics toward
difficult-to-reach states.
Linear mixed-effects model:
A form of linear data modeling that
allows a hierarchical data structure
and dependency between
observations by concurrently
mapping random and fixed effects.
Neurociencia en red
201
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
norte
mi
norte
_
a
_
0
0
1
7
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The missing role of gray matter in studying brain controllability
modelos, based on an analysis of variance (ANOVA, pag < 0.05). Statistical analysis was per-
formed using the lme4 package in R (Bates, Maechler, Bolker, & Walker, 2014). Specifically,
∼
two different models were constructed. The first model is defined by controllability (AC/MC)
TIV + Regions + Nodal degree × rGM + (1|participants), where × denotes the interaction and
(1|participants) assigns participants as a random intercept with a fixed slope. The aim was to
quantify the contribution of regional gray matter and nodal degree in explaining AC after con-
trolling for the regional differences of AC. In a second model, which is defined as AC/MC ∼
TIV + Regions × Nodal degree + Regions × rGM + (1|participants), we investigate the contri-
bution of regional differences of regional gray matter and regional differences of nodal degree
onto AC.
In both models, the volume of the entire brain (TIV) was added as a covariate given its
inter-relationship with GM volume (Lüders et al., 2002). In these models all the variables are
centered around zero within each subject and normalized using z-transformation. Models
were built with the lme4 package in R (Bates et al., 2014).
In these models, the elements Aij of the structural connectivity matrix (i.e., A in Equation
1) represent the number of streamlines between regions i and j. To ensure robustness, we
keep only 10% of the strongest connections using the Brain Connectivity Toolbox (Rubinov &
Sporns, 2010). Within this scheme, ith node degree is estimated by the sum of all elements
of A in the ith row. GM volume is estimated from the unified segmentation approach within
SPM12 (see the section on structural volume analysis, above, for details).
To further test our hypothesis, similar to Lee, Rodrigue, Glahn, Bassett, and
Null models.
Frangou (2020), we built random null models by randomizing the structural connectivity matrix
(i.e., A in Equation 1) and estimated the interrelationship between controllability, gray matter,
and degree distribution as explained in the section on LME formulation and statistical model
comparison, above. Specifically, preserving its degree distribution, we randomized matrix A
1,000 times using the Brain Connectivity Toolbox (Rubinov & Sporns, 2010) and compared the
beta values of rGM in the randomized networks with those obtained in the original network.
RESULTS
Effects of Gray Matter on Brain Controllability
In the first step, we investigated whether we could replicate previously reported findings that
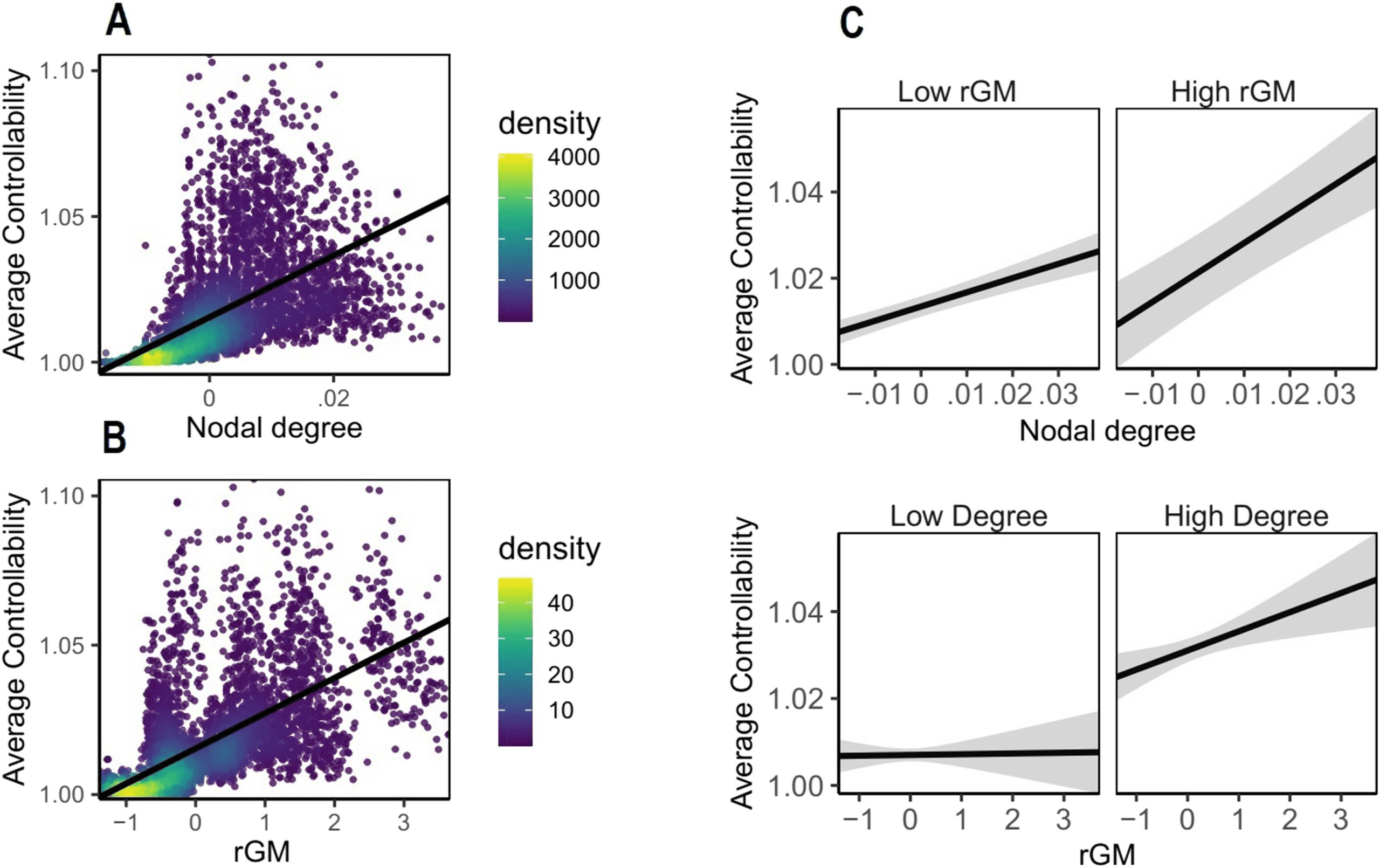
higher nodal degree relates to higher AC (see Figure 1A). We built a linear mixed-effects
model to predict AC based on nodal degree while controlling for TIV and including sub-
jects as a random intercept (for details, see the Supporting Information: Model comparisons,
Table C1). Our results, summarized in Figure 1A, replicate previous findings (Gu et al., 2015),
suggesting that structural connectivity strength quantified in terms of nodal degree across
the whole brain is positively associated with nodal AC. In the second step we investigated
whether, beyond this positive association between degree and AC, rGM explains additional
variance of AC. To this aim, we extended our model by including GM volume as an ad-
ditional predictor to nodal degree. Our results show that rGM and nodal degree are both
critical to explain AC, and their respective sizes of effects are comparable (βdegree = 0.36,
p value < 0.001; βrGM = 0.44, p value < 0.001). Next, we included regions as addi-
tional predictors to further explain AC and to improve the fitness of the model. Our results
show that rGM and AC are significantly positively associated (see Figure 1B) and interact with
nodal degree (β = 0.04, 95% CI [0.01, 0.07], pbonf = 0.01), suggesting that the highest lev-
els of average controllability were best explained with concurrent high rGM and high node
Network Neuroscience
202
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
n
e
n
_
a
_
0
0
1
7
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The missing role of gray matter in studying brain controllability
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
n
e
n
_
a
_
0
0
1
7
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Visualization of interaction effect of nodal degree and rGM in the mixed-effects model
predicting average controllability (AC). This effect was controlled for by the TIV and regional differ-
ences of average controllability. The figure shows that AC is best explained by WM structure and
rGM together. Each dot represents one region from one subject. The density bar shows where the
(A) Association between nodal degree and AC. (B) Association
majority of the data are located.
between rGM and AC. (C) Interaction of rGM and degree on AC, suggesting that the highest levels
of AC are reached when both degree and rGM are high together. For visualization, median split
was used to classify rGM and degree into high and low. In the original model, both GM and nodal
degree were preserved as continuous variables.
degree (see Figure 1C). To verify that the AC cannot not be explained with simpler mod-
els, we compared competing models (see the Supporting Information: Model comparisons,
Table C1), suggesting that the final model outperformed the competing models. Finally, we
used randomized null networks (for details, see the section on null models, above) to inves-
tigate whether rGM would remain a significant factor. Our results show that the contribution
of rGM in the randomized networks is significantly lower than those in the original networks
(p value < 0.001). Taken together, our results stress the interdependence of nodal connectivity
strength and GM volume for brain controllability.
Finally, in a further step, we used the same model to assess the relationship between MC,
rGM, and nodal degree (see the Supporting Information: Modal controllability, Figure B1).
Replicating previously reported findings that MC and nodal degree relationships are negatively
correlated (see Supporting Information: Modal controllability, Figure B1a), we find that rGM
explains a large part of MC variance and that the combination of nodal degree, rGM, and their
interaction is necessary.
Regional Distribution of Average Controllability Based on Gray Matter Volume
Further, we investigated whether this global interdependence of WM and rGM (see the previous
section) differs on a regional level. Given our previous results that MC and AC are strongly
negatively correlated we focus on AC.
Network Neuroscience
203
The missing role of gray matter in studying brain controllability
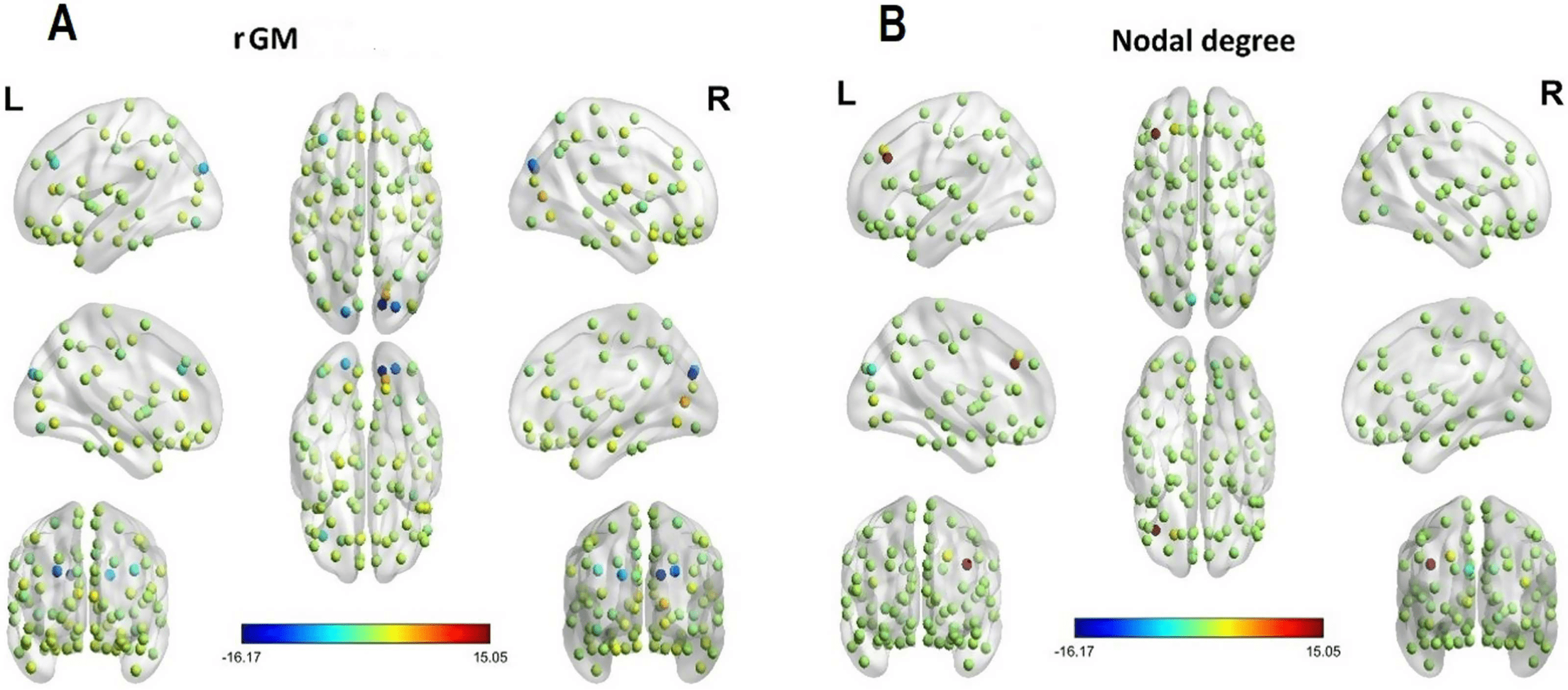
Figure 2. Visualization of interaction effects of mixed-effects model predicting average control-
lability (AC) based on regional GM (A) and regional nodal degree (B). For visualization, colors
represent standardized beta coefficients for effects of rGM and nodal degree, respectively, for each
brain region. Higher values indicate a beneficial effect and lower values indicate an impeding effect
of rGM/nodal degree on AC.
Our results (see the competing models Supporting Information: Model comparisons,
Table C2, and the full outcomes of the winning model in Table S2) show that higher rGM and
nodal degree concomitantly are associated with higher AC (see Figure 2; Table S2). Notably,
the highest AC levels with higher nodal degree were exhibited in the left frontal middle gyrus
(β = 15.11, 95% CI [4.09, 26.13], pbonf = 0.007) and the left superior frontal gyrus (β = 3.01,
95% CI [0.24, 5.78], pbonf = 0.033), which agrees with previous research that also locates
driver nodes for AC in the frontal lobes. Further, higher levels of AC were linked to higher lev-
els of nodal degree in the left calcarine (β = 1.78, 95% CI [0.69, 2.86], pbonf = 0.001). There
were also regions where higher levels of nodal degree exacerbated AC, with the strongest
effects located in the right and left cuneus (right cuneus: β = −1.34, 95% CI [−2.11, −0.57],
< 0.001). When turning
pbonf = 0.001; left cuneus: β = −2.70, 95% CI [−3.33, −2.08], pbonf
to the relation of rGM and AC, higher rGM is associated with higher AC levels in the right
< 0.001), right lingual area (β = 2.98, 95%
calcarine (β = 5.61, 95% CI [4.50, 6.73], pbonf
< 0.001), and the left and right anterior cingulate (left anterior cingu-
CI [2.63, 3.33], pbonf
< 0.001; right anterior cingulate: β = 2.88, 95%
late: β = 3.76, 95% CI [2.61, 4.91], pbonf
CI [2.28, 3.48], pbonf
< 0.001).
There were several regions exhibiting lower AC levels with higher rGM. The strongest effects
< 0.001)
were found in the right cuneus (β = −16.17, 95% CI [−18.46, −13.88], pbonf
and the left frontal middle gyrus (β = −3.34, 95% CI [−6.62, −0.07], pbonf = 0.045). The
finding suggests that although on a whole-brain level nodal degree and rGM are concomitantly
associated with increased AC, some regions, most notably the left frontal middle gyrus, exhibit
higher AC with higher nodal degree, and lower rGM, together (see the Supporting Information,
Table S2).
Replication Study
To investigate whether the results in the effects of Gray Matter on Brain Controllability sec-
tion (complementary effects of rGM and nodal degree and AC and MC) are replicable, we
used data from a cohort of 48 subjects from another publicly available dataset, where we
used a slightly different preprocessing pipeline (for details, see the Supporting Information:
Network Neuroscience
204
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
n
e
n
_
a
_
0
0
1
7
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The missing role of gray matter in studying brain controllability
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
n
e
n
_
a
_
0
0
1
7
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
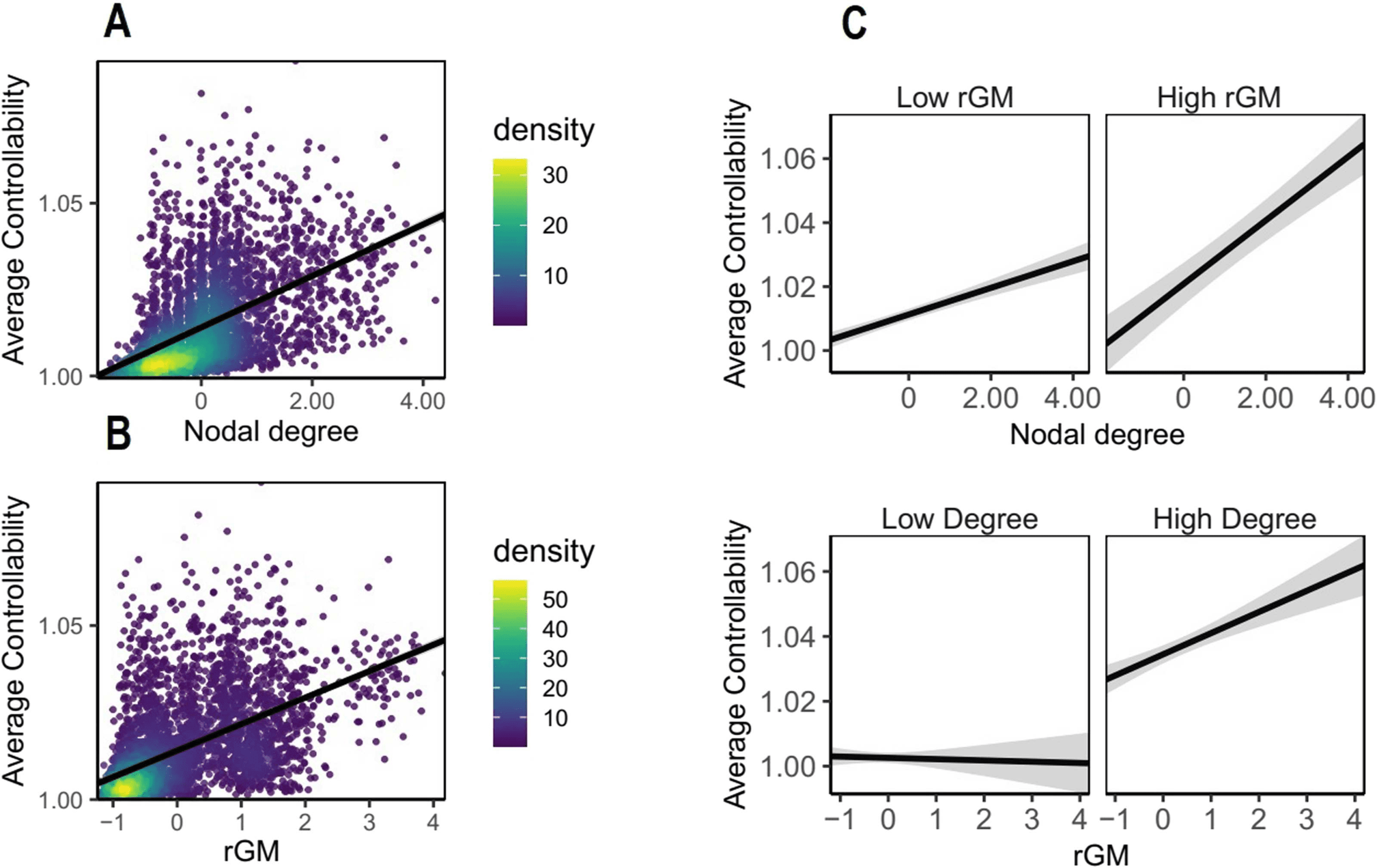
Figure 3. Replication sample. Average controllability (AC) is estimated based on WM structure
but also relates to rGM. Each dot represents data from one region of one subject and density bar
shows where the majority of data are located. (A) Effect of nodal degree on AC. (B) Effect of rGM
on AC. (C) Interaction effect of rGM and nodal degree suggests that the highest levels of AC are
reached when both degree and rGM are high together. For visualization, median split was used to
classify rGM and degree into high and low. In the original model, both GM and nodal degree were
preserved as continuous variables.
Replication study methods). Also in this dataset, nodal degree and rGM increased AC (see
Figure 3; for details see Table S3), while the highest AC levels were achieved when both nodal
degree and higher rGM were high together (β = 0.08, 95% CI [0.04, 0.12], pbonf = 0.01).
Furthermore, rGM and nodal degree both decrease MC, and the lowest values of MC were
achieved only for the lowest levels of rGM and nodal degree (see the Supporting Information:
Modal controllability, Figure B2). Taken together, these results suggest that this association
between rGM and nodal degree is robust and not driven by individual differences in different
datasets.
DISCUSSION
In this work, we investigated how brain volumetrics contribute to global network control prop-
erties derived from the structural connectome composed of the white matter fiber tracts. In line
with Medaglia, Pasqualetti, et al. (2017), we hypothesized that large-scale network dynamics
derived from the structural connectome (here quantified by average and modal brain con-
trollability) would be further explained by structural properties of GM. This work is, to our
knowledge, the first attempt to map the interdependence of both metrics, and we discuss our
findings with respect to their clinical relevance.
We show that, on average, the amount of rGM directly affects the brain’s availability to dy-
namically transition between brain states and to adopt new modes of activity. However, levels
of brain controllability were best explained when combining information from structural prop-
erties of both WM and rGM, suggesting that volumetrics might provide additional information
Network Neuroscience
205
The missing role of gray matter in studying brain controllability
in relating brain controllability to understanding cognition, neurological and neuropsychiatric
disorders, and the concept of brain reserve.
Mediating Role of GM on the Relationship Between WM and Brain Controllability
Our finding that nodal degree is highly predictive of brain controllability agrees with previous
work (Gu et al., 2015; Medaglia, 2019), suggesting that the brain’s ability to traverse into easy
and difficult-to-reach brain states relies on the strength of structural connectivity, which might
reflect the degrees of freedom to steer the transition of brain states. However, our findings sug-
gest that this picture is incomplete. Structural connectivity relies on sufficient support from GM
reserves. The highest effects of AC were reached with enhanced nodal degree within frontal
regions; this supports the rich literature showing that frontal brain networks play a central role
in initiating dynamic reconfigurations during executive cognition. However, increased rGM
within that very region was negatively related to brain controllability. While within clinical
populations reduced rGM is generally related to neuropathology, there is research suggesting
that within healthy subjects, rGM decreases with increases of WM density throughout devel-
opment from adolescence to adulthood. This finding has been related to reduced quantity
of synapses resulting from synaptic pruning (Giorgio et al., 2010), which has been predomi-
nantly found in primary visual (calcarine sulcus) and prefrontal cortex (middle frontal gyrus)
(Huttenlocher, 1979; Huttenlocher & Dabholkar, 1997). In our data, average brain control-
lability was maximal when exactly these regions showed reduced rGM and increased con-
nectivity of the white matter connectome. One could speculate that this finding reflects more
efficient and developmentally advanced brain functioning in a broad range of tasks potentially
related to synchronizing the actions with intentions in a goal-directed way.
Potential Contribution of Sensory Regions to Brain Controllability
On a functional level, we find several key visual areas that stand out with respect to both
average as well as modal controllability. Enhanced rGM in the right cuneus has previously
been reported to predict higher error rates in a response inhibition task in bipolar (Haldane,
Cunningham, Androutsos, & Frangou, 2008) and has also been related to motor response in
functional imaging studies (Booth et al., 2005; Matthews, Simmons, Arce, & Paulus, 2005).
We believe that these findings suggest that the function of those primary visual areas goes
far beyond unimodal information processing and that primary sensory cortices might occupy
more “hub-like” positions in the brain through enhanced long-distance connectivity across
brain-wide communities (Esfahlani, Bertolero, Bassett, & Betzel, 2020). Taken together, we
speculate that sensory regions could be ideal hot spots for brain controllability nodes. Given
their high global interconnectivity, these sensory nodes act potentially as the controllers with
respect to the afferent inputs while the other regions act as controllers for efferent demands.
Linking GM and WM in the Context of Controllability
Cognitive functioning arises from complex reconfigurations across metabolically expensive
large-scale networks, facing a trade-off between wiring cost (topological efficiency) and effi-
cient adaptation patterns between multiple neuronal populations (topological value; Bullmore
& Sporns, 2012). Recent studies have suggested that the behavioral relevance of this trade-off
between topological efficiency and topological value can be described by the brain’s energy
expenditure to exhibit control along large-scale structural networks. The ratio of neuronal
signaling- to nonsignaling-related metabolic energy expenditure has shown opposite direc-
tionalities for white and gray matter (Yu, Herman, Rothman, Agarwal, & Hyder, 2018; Zhu
Network Neuroscience
206
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
n
e
n
_
a
_
0
0
1
7
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The missing role of gray matter in studying brain controllability
et al., 2012). Here, we speculate that energy expenditure could be one of the key factors
linking GM and WM in the framework of controllability analysis. AC relates to the average en-
ergy a brain region needs to exert to steer the brain dynamics into all possible brain states (Gu
et al., 2015; Kenett, Medaglia, et al., 2018; Liu, Slotine, & Barabasi, 2011) and therefore, more
regional gray matter volume is more likely to provide the sufficient energy. In contrast, in ab-
sence of sufficient WM tracts, that is, lower nodal degree, rGM cannot fully force the transitions
since the energy cannot be exerted. This conception must be expressed on a behavioral level,
in that the brain system’s control capacity is especially sensitive to rGM. Indeed, a range of
studies have suggested that rGM but not white matter changes relate to abnormal behavioral
conditions, such as in antisocial personality disorder (Raine et al., 2000), medication-naive
high-functioning children with autism spectrum disorder (Palmen et al., 2005), and alcohol-
dependent individuals (Fein et al., 2002). Closely related, MC is strongest when nodal degree
and rGM are simultaneously low. MC reflects the ability to drive the brain dynamics toward
difficult-to-reach states by changing the modes of activity on the whole-brain level. It is there-
fore conceivable that similar to the relevance of nodal sparsity to enable optimal MC (Gu et al.,
2015), the scarcity of rGM enhances the ability of the host node by targeting only a minimal
set of other nodes.
Limitations: Beyond Linear Full Controllability
Our results in the current study warrant the conclusion that the interplay between gray matter
and controllability is of complex nature. Although these results highlight the potential missing
role of gray matter in studying brain controllability, there are important aspects that remain yet
to be explored. The choice of nonlinear dynamics to define the range of controllability metrics
could have considerable effects on our findings. For instance, it is suggested that importance
of nodal geometry could actually follow opposite trends when nonlinear and control models
are compared (Jiang & Lai, 2019). How the nonlinearity might (re)define the role of rGM
for control is an interesting question. Brain controllability metrics considered in the current
paper are trajectory unspecific. Studies of dynamical functional and structural connectivity
and analysis of structural covariance have reliably shown that brain state trajectories are not
random, but rather follow general rules (see Gu et al., 2017; Tang & Bassett, 2018, for recent
attempts to accommodate trajectory dependence in the broader context of network control
theory). Taken together, we suggest that future work should also include GM in studying brain
controllability and further investigate non-linearity of brain controllability.
ACKNOWLEDGMENTS
HJ was supported by Fortüne grant of Medical Faculty of University of Tübingen (No. 2487-1-
0). AZ was supported by the Swiss National Science Foundation (P2ZHP1_181435). MW was
supported by EU-ERA-Net: Neuromarket, EU-WIDESPREAD: Fat4BBrain, DFG Wa2673/10,
and Neurobiologie motivierten Verhaltens (TPA06) from SFB 779. The authors declare no
conflict of interest.
DATA AVAILABILITY
The data used in the current study are publicly available online. See the Methods section for
detail.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00174.
Network Neuroscience
207
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
n
e
n
_
a
_
0
0
1
7
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The missing role of gray matter in studying brain controllability
AUTHOR CONTRIBUTIONS
Hamidreza Jamalabadi: Conceptualization; Formal analysis; Investigation; Methodology; Proj-
ect administration; Software; Validation; Writing – original draft; Writing – review & editing.
Agnieszka Zuberer: Conceptualization; Formal analysis; Investigation; Methodology; Project
administration; Software; Validation; Visualization; Writing – original draft; Writing – review
& editing. Vinod Jangir Kumar: Data curation; Writing – review & editing. Meng Li: Data
curation; Writing – review & editing. Sarah Alizadeh: Conceptualization; Validation; Writing –
review & editing. Ali Moradi Amani: Methodology; Writing – review & editing. Christian
Gaser: Investigation; Validation; Writing – review & editing. Michael Esterman: Investigation;
Resources; Supervision; Writing – review & editing. Martin Walter: Investigation; Resources;
Supervision; Validation; Writing – review & editing.
FUNDING INFORMATION
Hamidreza Jamalabadi, Fortüne Grant of the Medical Faculty of University of Tübingen, Award
ID: 2487-1-0. Agnieszka Zuberer, Swiss National Science Foundation, Award ID: P2ZHP1_
181435. Martin Walter, ERA-NET NEURON, Award ID: Neuromarket. Martin Walter, EU-
WIDESPREAD, Award ID: Fat4BBrain. Martin Walter, DFG, Award ID: Wa2673/10. Martin
Walter, Neurobiologie motivierten Verhaltens from SFB 779, Award ID: TPA06.
REFERENCES
Agosta, F., Pievani, M., Sala, S., Geroldi, C., Galluzzi, S., Frisoni,
G. B., & Filippi, M. (2011). White matter damage in Alzheimer
disease and its relationship to gray matter atrophy. Radiology,
258(3), 853–863. DOI: https://doi.org/10.1148/radiol.10101284,
PMID: 21177393
Andersson, J. L., Skare, S., & Ashburner, J. (2003). How to correct
susceptibility distortions in spin-echo echo-planar images: Appli-
cation to diffusion tensor imaging. NeuroImage, 20(2), 870–888.
DOI: https://doi.org/10.1016/S1053-8119(03)00336-7
Andersson, J. L., & Sotiropoulos, S. N. (2015). Non-parametric rep-
resentation and prediction of single- and multi-shell diffusion-
weighted MRI data using Gaussian processes. NeuroImage, 122,
166–176. DOI: https://doi.org/10.1016/j.neuroimage.2015.07
.067, PMID: 26236030, PMCID: PMC4627362
Andersson, J. L., & Sotiropoulos, S. N.
(2016). An integrated ap-
proach to correction for off-resonance effects and subject move-
ment in diffusion MR imaging. NeuroImage, 125, 1063–1078.
DOI: https://doi.org/10.1016/j.neuroimage.2015.10.019, PMID:
26481672, PMCID: PMC4692656
Ashburner, J., & Friston, K. J. (2005). Unified segmentation. Neuro-
Image, 26(3), 839–851. DOI: https://doi.org/10.1016/j.neuroimage
.2005.02.018, PMID: 15955494
Baayen, R. H., Davidson, D. J., & Bates, D. M.
(2008). Mixed-
effects modeling with crossed random effects for subjects and
items. Journal of Memory and Language, 59(4), 390–412. DOI:
https://doi.org/10.1016/j.jml.2007.12.005
Bassett, D. S., & Sporns, O. (2017). Network neuroscience. Nature
Neuroscience, 20(3), 353. DOI: https://doi.org/10.1038/nn.4502,
PMID: 28230844, PMCID: PMC5485642
Bates, D., Maechler, M., Bolker, B., & Walker, S. (2014). lme4: Lin-
ear mixed-effects models using Eigen and S4. R package version,
1(7), 1–23. DOI: https://doi.org/10.18637/jss.v067.i01
Bodini, B., Khaleeli, Z., Cercignani, M., Miller, D. H., Thompson,
A. J., & Ciccarelli, O. (2009). Exploring the relationship between
white matter and gray matter damage in early primary progres-
sive multiple sclerosis: An in vivo study with TBSS and VBM. Hu-
man Brain Mapping, 30(9), 2852–2861. DOI: https://doi.org/10
.1002/hbm.20713, PMID: 19172648, PMCID: PMC6871131
Boisgontier, M. P., & Cheval, B. (2016). The anova to mixed model
transition. Neuroscience & Biobehavioral Reviews, 68, 1004–1005.
DOI: https://doi.org/10.1016/j.neubiorev.2016.05.034, PMID:
27241200
Booth, J. R., Burman, D. D., Meyer, J. R., Lei, Z., Trommer, B. L.,
Davenport, N. D., . . . Marsel Mesulam, M.
(2005). Larger
deficits in brain networks for response inhibition than for vi-
sual selective attention in attention deficit hyperactivity disorder
Journal of Child Psychology and Psychiatry, 46(1),
(ADHD).
94–111. DOI: https://doi.org/10.1111/j.1469-7610.2004.00337.x,
PMID: 15660647
.
.
J.
. Chen,
Braun, U., Harneit, A., Pergola, G., Menara, T., Schaefer, A., Betzel,
R. F.,
Brain state stability during
working memory is explained by network control theory, modu-
lated by dopamine D1/D2 receptor function, and diminished in
schizophrenia. arXiv:1906.09290. DOI: https://doi.org/10.1101
/679670
(2019).
Bullmore, E., & Sporns, O. (2012). The economy of brain network
organization. Nature Reviews Neuroscience, 13(5), 336. DOI:
https://doi.org/10.1038/nrn3214, PMID: 22498897
Douaud, G., Smith, S., Jenkinson, M., Behrens, T., Johansen-Berg,
H., Vickers, J., . . . Matthews, P. M.
(2007). Anatomically re-
lated grey and white matter abnormalities in adolescent-onset
schizophrenia. Brain, 130(9), 2375–2386. DOI: https://doi.org
/10.1093/brain/awm184, PMID: 17698497
Esfahlani, F. Z., Bertolero, M. A., Bassett, D. S., & Betzel, R. F.
Space-independent community and hub structure of
(2020).
Network Neuroscience
208
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
n
e
n
_
a
_
0
0
1
7
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The missing role of gray matter in studying brain controllability
functional brain networks. NeuroImage, 116612. DOI: https://
doi.org/10.1016/j.neuroimage.2020.116612, PMID: 32061801
Fein, G., Di Sclafani, V., Cardenas, V., Goldmann, H., Tolou-
Shams, M., & Meyerhoff, D. J. (2002). Cortical gray matter loss
in treatment-naive alcohol dependent individuals. Alcoholism:
Clinical and Experimental Research, 26(4), 558–564. DOI: https://
doi.org/10.1097/00000374-200204000-00017, https://doi.org/10
.1111/j.1530-0277.2002.tb02574.x, PMID: 11981133
Giorgio, A., Watkins, K. E., Chadwick, M., James, S., Winmill, L.,
Douaud, G.,
Longitudinal
changes in grey and white matter during adolescence. Neuro-
Image, 49(1), 94–103. DOI: https://doi.org/10.1016/j.neuroimage
.2009.08.003, PMID: 19679191
. . . Johansen-Berg, H.
(2010).
Glasser, M. F., Sotiropoulos, S. N., Wilson, J. A., Coalson, T. S.,
Fischl, B., Andersson, J. L., . . . Polimeni, J. R.
(2013). The
minimal preprocessing pipelines for the Human Connectome
Project. NeuroImage, 80, 105–124. DOI: https://doi.org/10.1016
/j.neuroimage.2013.04.127,
PMCID:
PMC3720813
23668970,
PMID:
Gu, S., Betzel, R. F., Mattar, M. G., Cieslak, M., Delio, P. R.,
Grafton, S. T., . . . Bassett, D. S. (2017). Optimal trajectories of
brain state transitions. NeuroImage, 148, 305–317. DOI: https://
doi.org/10.1016/j.neuroimage.2017.01.003, PMID: 28088484,
PMCID: PMC5489344
. . . Bassett, D. S.
Gu, S., Pasqualetti, F., Cieslak, M., Telesford, Q. K., Yu, A. B.,
(2015). Controllability of
Kahn, A. E.,
structural brain networks. Nature Communications, 6, 8414.
DOI: https://doi.org/10.1038/ncomms9414, PMID: 26423222,
PMCID: PMC4600713
Haldane, M., Cunningham, G., Androutsos, C., & Frangou, S.
(2008). Structural brain correlates of response inhibition in Bipo-
lar Disorder I. Journal of Psychopharmacology, 22(2), 138–143.
DOI: https://doi.org/10.1177/0269881107082955, PMID:
18308812
Huttenlocher, P. R.
(1979). Synaptic density in human frontal
cortex-developmental changes and effects of aging. Brain Re-
search, 163(2), 195–205. DOI: https://doi.org/10.1016/0006
-8993(79)90349-4
Huttenlocher, P. R., & Dabholkar, A. S.
(1997). Regional differ-
Journal of
ences in synaptogenesis in human cerebral cortex.
Comparative Neurology, 387(2), 167–178. DOI: https://doi.org
/10.1002/(SICI)1096-9861(19971020)387:2<167::AID-CNE1>3
.0.CO;2-z
Jeganathan, J., Perry, A., bassett, D. S., Roberts, GRAMO., mitchell, PAG. B.,
& romper la lanza, METRO.
(2018). Fronto-limbic dysconnectivity leads
to impaired brain network controllability in young people with
bipolar disorder and those at high genetic risk. NeuroImagen:
Clinical, 19, 71–81. DOI: https://doi.org/10.1016/j.nicl.2018.03
.032, PMID: 30035004, PMCID: PMC6051310
Jenkinson, METRO., Bannister, PAG., Brady, METRO., & Herrero, S.
Im-
proved optimization for the robust and accurate linear registra-
tion and motion correction of brain images. NeuroImagen, 17(2),
825–841. DOI: https://doi.org/10.1006/nimg.2002.1132, PMID:
12377157
(2002).
Jenkinson, METRO., & Herrero, S. (2001). A global optimisation method for
robust affine registration of brain images. Medical Image Analy-
hermana, 5(2), 143–156. DOI: https://doi.org/10.1016/S1361-8415(01)
00036-6
Jeurissen, B., Tournier, J.-D., Dhollander, T., connelly, A., & Sijbers,
(2014). Deconvolución esférica restringida por múltiples tejidos para
j.
análisis mejorado de datos de resonancia magnética de difusión multicapa. Neuro-
Image, 103, 411–426. DOI: https://doi.org/10.1016/j.neuroimage
.2014.07.061, PMID: 25109526
Jiang,
J., & Dejar, Y.-C. (2019). Irrelevance of linear controllabil-
ity to nonlinear dynamical networks. Comunicaciones de la naturaleza,
10(1), 1–10. DOI: https://doi.org/10.1038/s41467-019-11822-5,
PMID: 31481693, PMCID: PMC6722065
Kailath, t. (1980). Linear systems (volumen. 156). Prentice Hall.
Kalmán, R. mi. (1963). Mathematical description of linear dynam-
Journal of the Society for Industrial and Applied
ical systems.
Matemáticas, Series A: Control, 1(2), 152–192. DOI: https://
doi.org/10.1137/0301010
Kenett, Y. NORTE., Beaty, R. MI., & Medalla, j. D.
(2018). A compu-
tational network control theory analysis of depression symp-
toms. Personality Neuroscience, 1. DOI: https://doi.org/10.1017
/pen.2018.15, PMID: 30706049, PMCID: PMC6349380
Kenett, Y. NORTE., Medalla,
j. D., Beaty, R. MI., Chen, P., Betzel,
R. F., Thompson-Schill, S. l., & Qiu, j. (2018). Driving the brain
towards creativity and intelligence: A network control the-
ory analysis. Neuropsicología. DOI: https://doi.org/10.1016/j
.neuropsychologia.2018.01.001, PMID: 29307585, PMCID:
PMC6034981
kong, l., Herold, C. J., Zöllner, F., Salat, D. h., Lässer, METRO. METRO.,
Schmid, l. A., . . . Schad, l. R. (2015). Comparison of grey matter
volume and thickness for analysing cortical changes in chronic
schizophrenia: A matter of surface area, grey/white matter inten-
sity contrast, and curvature. Psychiatry Research: Neuroimaging,
231(2), 176–183. DOI: https://doi.org/10.1016/j.pscychresns.2014
.12.004, PMID: 25595222
Sotavento, W.. h., Rodrigue, A., Glahn, D. C., bassett, D. S., & Frangou,
S. (2020). Heritability and cognitive relevance of structural brain
controllability. Corteza cerebral, 30(5), 3044–3054. DOI: https://
doi.org/10.1093/cercor/bhz293, PMID: 31838501, PMCID:
PMC7197079
Liu, Y. y., Slotine, j. J., & Barabasi, A. l.
(2011). Controllabil-
ity of complex networks. Naturaleza, 473(7346), 167–173. DOI:
https://doi.org/10.1038/nature10011, PMID: 21562557
Lüders, MI., Steinmetz, h., & Jäncke, l.
(2002). Brain size and
grey matter volume in the healthy human brain. Neuroreport,
13(17), 2371–2374. DOI: https://doi.org/10.1097/00001756
-200212030-00040, PMID: 12488829
Matthews, S. C., Simmons, A. NORTE., Arce, MI., & Paulus, METRO. PAG.
(2005). Dissociation of inhibition from error processing using
a parametric inhibitory task during functional magnetic reso-
nance imaging. Neuroreport, 16(7), 755–760. DOI: https://doi
.org/10.1097/00001756-200505120-00020, PMID: 15858420
Medalla, j. D.
(2019). Clarifying cognitive control and the con-
trollable connectome. Wiley Interdisciplinary Reviews: Cognitivo
Ciencia, 10(1), e1471. DOI: https://doi.org/10.1002/wcs.1471,
PMID: 29971940, PMCID: PMC6642819
Medalla, j. D., Pasqualetti, F., hamilton, R. h., Thompson-Schill,
S. l., & bassett, D. S. (2017). Brain and cognitive reserve: Trans-
lation via network control theory. Neurociencia & Biobehavioral
Neurociencia en red
209
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
norte
mi
norte
_
a
_
0
0
1
7
4
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The missing role of gray matter in studying brain controllability
Reseñas, 75, 53–64. DOI: https://doi.org/10.1016/j.neubiorev
.2017.01.016, PMID: 28104411, PMCID: PMC5359115
of Imaging Systems and Technology, 22(1), 53–66. DOI: https://
doi.org/10.1002/ima.22005
Medalla, j. D., Zurn, PAG., Sinnott-Armstrong, w., & bassett, D. S.
(2017). Mind control as a guide for the mind. Nature Human Be-
haviour, 1(6). DOI: https://doi.org/10.1038/s41562-017-0119
Muldoon, S. F., Pasqualetti, F., Gu, S., Cieslak, METRO., Grafton, S. T.,
Vettel, j. METRO., & bassett, D. S. (2016). Stimulation-based control
of dynamic brain networks. Biología Computacional PLoS, 12(9),
e1005076. DOI: https://doi.org/10.1371/journal.pcbi.1005076,
PMID: 27611328, PMCID: PMC5017638
Palmen, S. J., Pol, h. mi. h., Kemner, C., Schnack, h. GRAMO., Durston,
S., Lahuis, B. MI., . . . Van Engeland, h. (2005). Increased gray-
matter volume in medication-naive high-functioning children
with autism spectrum disorder. Psychological Medicine, 35(4),
561–570. DOI: https://doi.org/10.1017/S0033291704003496,
PMID: 15856726
Pasqualetti, F., Zampieri, S., & Bullo, F.
(2014). Controllability
IEEE
métrica, limitations and algorithms for complex networks.
Transactions on Control of Network Systems, 1(1), 40–52. DOI:
https://doi.org/10.1109/TCNS.2014.2310254
Raine, A., Lencz, T., Bihrle, S., LaCasse, l., & Colletti, PAG. (2000).
Reduced prefrontal gray matter volume and reduced autonomic
activity in antisocial personality disorder. Archives of General Psy-
chiatry, 57(2), 119–127. DOI: https://doi.org/10.1001/archpsyc
.57.2.119, PMID: 10665614
Rubinov, METRO., & despreciar, oh.
(2010). Complex network measures
of brain connectivity: Uses and interpretations. NeuroImagen,
52(3), 1059–1069. DOI: https://doi.org/10.1016/j.neuroimage
.2009.10.003, PMID: 19819337
.
.
J.,
. Lenglet, C.
Sotiropoulos, S. NORTE., Moeller, S., Jbabdi, S., Xu, J., andersson, j. l.,
im-
Auerbach, mi.
age reconstruction on fiber orientation mapping from multi-
channel diffusion MRI: Reducing the noise floor using SENSE.
Resonancia Magnética en Medicina, 70(6), 1682–1689. DOI:
https://doi.org/10.1002/mrm.24623, PMID: 23401137, PMCID:
PMC3657588
Efectos de
(2013).
Espiga, MI., & bassett, D. S. (2018). Coloquio: Control of dynamics
in brain networks. Reseñas de Física Moderna, 90(3), 031003.
DOI: https://doi.org/10.1103/RevModPhys.90.031003
Tournier, j. D., Calamante, F., & connelly, A. (2012). MRtrix: Dif-
fusion tractography in crossing fiber regions. International Journal
Tzourio-Mazoyer, NORTE., Landeau, B., Papathanassiou, D., Crivello,
(2002). Automated
F., Etard, o., Delcroix, NORTE., . . . Joliot, METRO.
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of
single-subject
cerebro. NeuroImagen, 15(1), 273–289. DOI: https://doi.org/10.1006
/nimg.2001.0978, PMID: 11771995
the MNI MRI
VanEssen, D. C., Ugurbil, K., Auerbach, MI., Respeto, D., Behrens,
T., Bucholz, r.,
(2012). The Human
. Curtiss, S. W..
Connectome Project: A data acquisition perspective. NeuroImagen,
62(4), 2222–2231. DOI: https://doi.org/10.1016/j.neuroimage
.2012.02.018, PMID: 22366334, PMCID: PMC3606888
.
.
Villain, NORTE., Desgranges, B., Viader, F., De La Sayette, v., Mézenge,
F., Landeau, B., . . . Chételat, GRAMO.
(2008). Relationships be-
tween hippocampal atrophy, white matter disruption, and gray
matter hypometabolism in Alzheimer’s disease. Journal of Neu-
roscience, 28(24), 6174–6181. DOI: https://doi.org/10.1523
/JNEUROSCI.1392-08.2008,
PMCID:
PMC2902815
18550759,
PMID:
Winkler, A. METRO., Kochunov, PAG., Blangero, J., Almasy, l., Zilles, K.,
(2010). Cortical thickness or
Fox, PAG. T., . . . Glahn, D. C.
grey matter volume? The importance of selecting the phenotype
for imaging genetics studies. NeuroImagen, 53(3), 1135–1146.
DOI: https://doi.org/10.1016/j.neuroimage.2009.12.028, PMID:
20006715, PMCID: PMC2891595
Yu, y., Herman, PAG., Rothman, D. l., agarwal, D., & Hyder, F.
(2018). Evaluating the gray and white matter energy budgets
Journal of Cerebral Blood Flow
of human brain function.
and Metabolism, 38(8), 1339–1353. DOI: https://doi.org/10.1177
/0271678X17708691, PMID: 28589753, PMCID: PMC6092772
Zhu, X.-H., Qiao, h., Du, F., xiong, P., Liu, X., zhang, X.,
. . . Chen, W..
(2012). Quantitative imaging of energy ex-
penditure in human brain. NeuroImagen, 60(4), 2107–2117.
DOI: https://doi.org/10.1016/j.neuroimage.2012.02.013, PMID:
22487547, PMCID: PMC3325488
(2019).
Zoeller, D., Sandini, C., Schaer, METRO., Eliez, S., bassett, D., & Van De
Ville, D.
Structural control energy of resting-state
functional brain states reveals inefficient brain dynamics in
psychosis vulnerability. bioRxiv:703561. DOI: https://doi.org/10
.1101/703561
Neurociencia en red
210
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
5
1
1
9
8
1
8
8
9
6
8
9
norte
mi
norte
_
a
_
0
0
1
7
4
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3