Revisar
Questions and controversies in the
study of time-varying functional
connectivity in resting fMRI
Daniel J. Lurie
Michael Breakspear
1,∗
2,∗
, Daniel Kessler
7,8
, Shella Keilholz
13
, Danielle S. bassett
, Aaron Kucyi
9
10
, Anthony Randal McIntosh
3,4,5,6
, Richard F. Betzel
, Raphaël Liégeois
, Russell A. Poldrack
14,15
16,18
20
, William Hedley Thompson
, Robyn L. Molinero
21
, Dominik Kraft
22
, Natalia Z. Bielczyk
,
, Muthuraman Muthuraman
Lorenzo Pasquini
Hua Xie
, Adeel Razi
, and Vince D. Calhoun
, Diego Vidaurre
22,30,31,∗
25,26,27
24
29
28
,
Martin A. Lindquist
James M.. Brillar
17
Linda Douw
3
,
,
11,12
16
,
23
,
19
un acceso abierto
diario
Citación: Lurie, D. J., Kessler, D.,
bassett, D. S., Betzel, R. F., romper la lanza,
METRO., Keilholz, S., . . . Calhoun, V. D.
(2020). Questions and controversies in
the study of time-varying functional
connectivity in resting fMRI. Red
Neurociencia, 4(1), 30–69. https://
doi.org/10.1162/netn_a_00116
DOI:
https://doi.org/10.1162/netn_a_00116
Supporting Information:
https://doi.org/10.1162/netn_a_00116
https://doi.org/10.5281/zenodo.3565986
Recibió: 17 Puede 2019
Aceptado: 22 Noviembre 2019
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autores correspondientes:
Daniel J. Lurie
danjlurie@gmail.com
Vince Calhoun
vcalhoun@gsu.edu
Editor de manejo:
Emily Finn
Derechos de autor: © 2019
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
1
3
10
Department of Psychology, Universidad de California, berkeley, berkeley, California, EE.UU
2Departments of Statistics and Psychiatry, University of Michigan, ann-arbor, MI, EE.UU
Department of Bioengineering, School of Engineering and Applied Sciences, Universidad de Pennsylvania,
Filadelfia, Pensilvania, EE.UU
4Department of Physics and Astronomy, College of Arts & Ciencias, Universidad de Pennsylvania, Filadelfia, Pensilvania, EE.UU
5Department of Neurology, Perelman School of Medicine, Universidad de Pennsylvania, Filadelfia, Pensilvania, EE.UU
6Department of Electrical and Systems Engineering, School of Engineering and Applied Sciences, Universidad de Pennsylvania,
Filadelfia, Pensilvania, EE.UU
7University of Newcastle, Callaghan, NSW, Australia
8QIMR Berghofer, Brisbane, Australia
9Department of Biomedical Engineering, Emory University and Georgia Institute of Technology, Atlanta, Georgia, EE.UU
Department of Neurology and Neurological Sciences, Universidad Stanford, Stanford CA, EE.UU
11Institute of Bioengineering, Center for Neuroprosthetics, Ecole Polytechnique Fédérale de Lausanne, Suiza
12Department of Radiology and Medical Informatics, University of Geneva, Suiza
13Department of Biostatistics, Universidad Johns Hopkins, baltimore, Maryland, EE.UU
14Rotman Research Institute – Baycrest Centre, toronto, Canada
15Department of Psychology, universidad de toronto, toronto, Canada
16Department of Psychology, Universidad Stanford, stanford, California, EE.UU
17Brain and Mind Centre, University of Sydney, NSW, Australia
18Department of Clinical Neuroscience, Karolinska Institutet, Stockholm, Suecia
19Stichting Solaris Onderzoek en Ontwikkeling, Nimega, Los países bajos
20Department of Anatomy and Neurosciences, VU University Medical Center, Ámsterdam, Los países bajos
21Department of Psychology, Goethe University Frankfurt, Frankfurt am Main, Alemania
22The Mind Research Network, Albuquerque, NM, EE.UU
Biomedical Statistics and Multimodal Signal Processing Unit, Movement Disorders and Neurostimulation, Department of
Neurología, Focus Program Translational Neuroscience, Johannes-Gutenberg-University Hospital, Mainz, Alemania
24Memory and Aging Center, Department of Neurology, Universidad de California, San Francisco, San Francisco, California, EE.UU
25Monash Institute of Cognitive and Clinical Neurosciences and Monash Biomedical Imaging, Monash
Universidad, Clayton, Australia
26Wellcome Centre for Human Neuroimaging, Institute of Neurology, University College London, Londres, Reino Unido
27Department of Electronic Engineering, NED University of Engineering and Technology, Karachi, Pakistán
28Wellcome Trust Centre for Integrative Neuroimaging, Oxford Centre for Human Brain Activity, Universidad de Oxford,
Reino Unido
Department of Psychiatry and Behavioral Sciences, Universidad Stanford, stanford, California, EE.UU
30Department of Electrical and Computer Engineering, University of New Mexico, Albuquerque, NM, EE.UU
31Tri-institutional Center for Translational Research in Neuroimaging and Data Science (TReNDS), Georgia State,
Georgia Tech, Emory, Atlanta, Georgia, EE.UU
Indicates equal contribution.
29
23
∗
Palabras clave: Conectividad funcional, Brain networks, Brain dynamics, resonancia magnética funcional, Rest, Revisar
ABSTRACTO
The brain is a complex, multiscale dynamical system composed of many interacting regions.
Knowledge of the spatiotemporal organization of these interactions is critical for establishing
a solid understanding of the brain’s functional architecture and the relationship between
neural dynamics and cognition in health and disease. The possibility of studying these
dynamics through careful analysis of neuroimaging data has catalyzed substantial interest in
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
Arousal:
A continuous property of brain and
bodily states that influences
information processing and
behavioral responsivity though
effects on neural signaling.
“Resting” state:
Behavioral state with minimal (or no)
explicit task demands. Participantes
may engage in a wide range of
self-directed cognitive processes.
methods that estimate time-resolved fluctuations in functional connectivity (often referred to
as “dynamic” or time-varying functional connectivity; TVFC). Al mismo tiempo, debates have
emerged regarding the application of TVFC analyses to resting fMRI data, and about the
statistical validity, physiological origins, and cognitive and behavioral relevance of resting
TVFC. These and other unresolved issues complicate interpretation of resting TVFC findings
and limit the insights that can be gained from this promising new research area. This article
brings together scientists with a variety of perspectives on resting TVFC to review the current
literature in light of these issues. We introduce core concepts, define key terms, summarize
controversies and open questions, and present a forward-looking perspective on how resting
TVFC analyses can be rigorously and productively applied to investigate a wide range of
questions in cognitive and systems neuroscience.
TIME-VARYING FUNCTIONAL CONNECTIVITY: AN INTRODUCTION
Even when sitting quietly in a dark room, the brain is active, yielding a constant stream of
thoughts and ideas, along with changes in awareness, arousal, and vigilance. The brain con-
stantly constructs and updates internal models of the world to anticipate and plan future adap-
tive behaviors (Parr, rees, & Friston, 2018), and wakeful rest is no less cognitively rich and
complex than task engagement. The notion that patterns of neuronal activity and interregional
coupling may exhibit the statistical and dynamical fingerprints of these mental wanderings—
even in the absence of an explicit task—accords with the most fundamental observations of our
“stream of consciousness.” While it is relatively straightforward to quantify changes in brain
activity and functional connectivity that are time-locked to perceptual stimuli and externally
cued tasks (cohen, 2018; Gonzalez-Castillo & Bandettini, 2018), detecting and characterizing
changes that arise “spontaneously”—from endogenous and unknown causes and at seemingly
random times—is substantially more difficult. Despite these challenges, studies of intrinsic
brain dynamics and self-directed “resting” cognition provide an important, ecologically valid
perspective on brain function and mental life. A large proportion of our time (hasta 50%) es
spent engaging in cognition and behavior unrelated to the task at hand (Killingsworth & Gilbert,
2010), and emerging evidence suggests that these task-unrelated thoughts and actions may ex-
plain up to twice the variance in neural activity than task-related variables (Musall, Kaufman,
Juavinett, Gluf, & Iglesia, 2019).
Conectividad funcional (FC) analyses of resting fMRI (rfMRI) data allow researchers to
noninvasively estimate patterns of interregional neural interactions. An integral component
of modern neuroimaging research, FC is traditionally calculated over an entire scan or experi-
mental condition (“static” functional connectivity), but recent years have seen rapidly growing
interest in studying time-resolved fluctuations in FC (often referred to as “dynamic” or time-
varying functional connectivity; TVFC; Calhoun, Molinero, Pearlson, & Adali, 2014; hutchison,
Womelsdorf, allen, et al., 2013; ver figura 1). A burgeoning literature now spans studies using
varied imaging modalities (p.ej., resonancia magnética funcional, Sakoglu et al., 2010; EEG, Tagliazucchi, von Wegner,
Morzelewski, Brodbeck, & Laufs, 2012; and MEG, Baker et al., 2014) to investigate fluctuations
in FC during a wide range of cognitive and behavioral states ranging from explicitly cued task
execution (p.ej., Gonzalez-Castillo & Bandettini, 2018) to wakeful rest (p.ej., Allen et al., 2014),
sleep (p.ej., Tagliazucchi & Laufs, 2014), and anesthesia (p.ej., hutchison, Womelsdorf, LISTO,
Everling, & menón, 2013). Interindividual differences in resting TVFC have been associated
with a wide range of cognitive and behavioral traits (Liegeois et al., 2019; Vidaurre, Herrero,
& lana rica, 2017), and emerging evidence suggests that in some cases TVFC may be a more
sensitive marker of these differences than static FC (Jin et al., 2017; Liegeois et al., 2019; Rashid
Neurociencia en red
31
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. Growth of the fMRI TVFC literature. The field of TVFC research has grown rapidly, como
demonstrated by the increasing number of fMRI TVFC papers published each year (as indexed by
PubMed). To account for overall growth in the rate of scientific publishing, the height of the bars has
been normalized by the total number of all papers published in each year. Because of inconsistencies
in the way TVFC analyses are described, these figures likely represent a conservative estimate of the
size of the fMRI TVFC literature, particularly for earlier years. For details on the search terms used
to identify TVFC papers, please see the Supporting Information.
et al., 2016; Vidaurre, Llera, Herrero, & lana rica, 2019). Alterations in TVFC have also been
observed in a growing number of psychiatric and neurological conditions including autism
(de Lacy, Doherty, Rey, Rachakonda, & Calhoun, 2017), TDAH (de Lacy & Calhoun, in press),
depresión (Kaiser et al., 2016), PTSD (Jin et al., 2017), schizophrenia (Sakoglu et al., 2010),
Parkinson’s (Diez-Cirarda et al., 2018), and Alzheimer’s disease (Jones et al., 2012).
Like any emerging research program, resting TVFC research has encountered its share of
growing pains and challenges. Studying the brain at rest has a number of advantages—minimal
demands on study participants, analytic flexibility afforded by the lack of an externally imposed
tarea, the absence of potential performance confounds—and may potentially provide a richer
characterization of brain activity than task studies (Ponce-Alvarez, Él, Hagmann, & decoración,
2015). Sin embargo, while resting TVFC research benefits from the advantages of rfMRI, también
suffers from its pitfalls: the lack of clear benchmarks, the absence of experimental control of
behavioral or cognitive state, and the inability to objectively monitor behavioral task perfor-
mance. Paralleling similar debates from the early days of rfMRI (see Box 1), there is active
debate about the extent to which BOLD TVFC is able to detect transient changes in neural
signaling or cognitive state during rest. A number of important open questions contribute to
Neurociencia en red
32
Questions and controversies in the study of TVFC in resting fMRI
Box 1. A brief history of studying the brain at rest
Studying the brain at rest is not a new idea. Scientists have been interested in the dynamics
of resting cognition at least since the writings of William James in the late 1800s (James,
1890), and much of Hans Berger’s pioneering EEG research in the 1920s was focused on the
properties of intrinsic brain activity (Karbowski, 1990). Following the development of PET and
BOLD fMRI in the 1980s and 1990s, human functional neuroimaging was initially dominated
by task activation paradigms. Sin embargo, researchers quickly began to notice a set of regions
that consistently deactivated in response to external task demands, and that exhibited high
metabolic activity during rest. This set of regions was named the default mode network (DMN)
in a seminal 2001 paper by Raichle et al. (2001). In a complementary line of work, Biswal
et al. estimated BOLD fMRI functional connectivity between primary motor cortex and other
brain areas, independent of any overt task (Biswal, Yetkin, Haughton, & Hyde, 1995). El
resulting spatial patterns of FC mirrored patterns of activation seen when subjects executed a
respuesta motora. These and other findings led to renewed interest in the study of the brain at
descansar, with the hope that better characterizing “resting state” FC networks would reveal core
features of the brain’s functional organization.
Neuroimaging studies of
the brain at rest quickly converged on a set of canoni-
cal FC networks that are consistently observed at rest and correspond with patterns of
task-evoked activation and functional connectivity (Calhoun, Kiehl, & Pearlson, 2008;
Damoiseaux et al., 2006; Smith et al., 2009). While early studies focused on investi-
individual networks (p.ej., DMN; METRO. D. Greicius, Krasnow, Reiss, &
gating the FC of
menón, 2003),
func-
tional organization by mapping FC across the whole brain (p.ej., Yeo et al., 2011). Estos
initial observations have been widely replicated across hundreds of studies using a variety of
analytic methods (p.ej., seed-based functional connectivity, ICA, community detection).
this work eventually expanded into efforts to investigate global
Interindividual differences in resting FC patterns have been associated with a wide range
of phenotypic traits (p.ej., working memory and executive control; Col, Yarkoni, Repovs,
Anticevic, & Más valiente, 2012; Hampson, Driesen, Skudlarski, Sangre, & Constable, 2006) y
clinical conditions (p.ej., psychiatric and neurological disorders; Fox & Greicius, 2010;
METRO. Greicius, 2008), and can be used to predict behavioral performance (p.ej., METRO. D. Rosenberg
et al., 2016) and individual identity (p.ej., Finn et al., 2015).
Despite the success of the resting FC research program in expanding our understanding of
human brain function, it has historically been limited by the use of methods that are unable
to address fundamental motivating questions about inherently dynamic cognitive and neural
procesos. In response to this limitation, the past decade has seen the emergence of new tools
for studying the time-varying properties of the brain at rest.
this lack of consensus: To what extent are estimates of resting BOLD TVFC driven by fluctua-
tions in arousal and cognitive state versus nonneural physiological factors (p.ej., head motion,
cardiovascular and respiratory effects)? What are the most appropriate ways to test observed
estimates of TVFC against “static” null hypotheses? Whereas detecting change-points or fluctu-
ating dependence structure in neuroimaging data is in principle an achievable outcome of sig-
nal analysis—and indeed these are the goals of many TVFC analysis methods—understanding
the putative causes of these changes requires other techniques: online measures of cognitive
and bodily states, insights from pathological conditions, the inversion of generative models,
and causal manipulations such as brain stimulation and administration of pharmacological
agents. It is our goal to summarize the current literature surrounding these and related issues,
and to provide suggestions for future work that may help adjudicate these debates.
While there are indeed real points of
fundamental disagreement among researchers
about various aspects of BOLD TVFC, debates in the literature have at times been needlessly
Neurociencia en red
33
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
muddied by inconsistent or imprecise definitions and operationalizations. Por ejemplo, el
term “metastate” has been variously used to describe (a) a small number of replicable pat-
terns of connectivity that recur across or within individuals (es decir., functional connectivity states;
Brillar, Koyejo, & Poldrack, 2016), (b) subsets of functional connectivity and activity states
that share certain temporal characteristics (Vidaurre et al., 2017), o (C) a specific location in a
second-order state-space (R. l. Miller et al., 2016). As has been previously suggested (Liegeois,
Laumann, Snyder, zhou, & yo, 2017; R. l. Molinero, Abrol, Adali, Levin-Schwarz, & Calhoun,
2018), we believe that progress on resolving these debates requires standardizing our termi-
nology and identifying common frameworks. While intuitive notions of brain dynamics may
seem straightforward, there is currently no consensus about operational definitions for many
key concepts related to TVFC. Establishing appropriate terminology for the phenomenon under
study is particularly important. Although “dynamic functional connectivity” is frequently used
in the literature, different uses and definitions of the term “dynamic” across disciplines can
lead to troublesome ambiguity. Tal como, we have opted here to use the more broadly applica-
ble phrase “time-varying functional connectivity,” where functional connectivity refers to any
of various notions of statistical dependence, most commonly (but not exclusively) correlation
between time series. We define this and other key terms in the glossary presented in Table 1,
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Term
Conectividad funcional (FC)
Static functional connectivity
Statistical stationarity
Mesa 1. Glossary of key terms
Definición
Statistical dependencies among neurophysiological time series derived from regions or networks. Mayoría
often estimated as a correlation coefficient.
An estimate of statistical dependence made under the assumption that the dependence structure does not
vary as a function of time.
A formal definition of certain statistical properties being invariant to a shift in time. En la práctica, stationarity
can only be assessed given multiple realizations of a time series (rather than for a single dataset).
– Strong stationarity: The probability distribution of the time series is invariant under a shift in time.
– Weak stationarity (or second-order stationarity): The mean and covariance of the time series are finite
and invariant under a shift in time. This is the definition most time series models use in practice.
Time-varying functional connectivity (TVFC)
Functional connectivity that varies as a function of time. Also referred to as “dynamic functional
connectivity.”
Functional connectivity state
A transient pattern of whole-brain functional connectivity. Usually identified by analytic techniques that
attempt to model the full repertoire of functional connectivity patterns as being made up of a relatively
small number of FC states (often referred to in shorthand simply as “states”). Some of these low-dimensional
models constrain the brain to be in a single state at a time, whereas others permit each time point to be a
mixture of states.
Activity state
A transient pattern of whole-brain activation, analogous to a functional connectivity state.
Windowed functional connectivity
Dynamical system
Hidden Markov model (HMM)
Functional connectivity estimated over a defined time window that is shorter than the full time series.
Windowing can involve weighting or tapering. “Sliding window” methods can be used to produce time-
resolved estimates of functional connectivity (one for each window).
A system composed of interacting components (neuronas, regiones del cerebro, etc.) whose state evolves forward
in time according to a particular rule (such as a difference or differential equation). Such systems yield
complex behaviors that can be observed via an (often indirect) measurement process.
A statistical model wherein observed data are assumed to be generated from a process that moves among
unobserved states. Fitting an HMM involves estimating (1) the properties of each state, (2) transition prob-
abilities between the states, y (3) which state is active at each time point. For TVFC applications, cada
state might correspond to a distinct pattern of brain activity and functional connectivity, the transition prob-
abilities would explain how the brain moves from one state to another, and the estimates of active states
would give time-resolved estimates of which state was active at each time point.
Neurociencia en red
34
Questions and controversies in the study of TVFC in resting fMRI
while Box 2 provides a brief discussion of the nuances involved in relating TVFC estimates to
the underlying neural phenomena we seek to study and understand.
Box 2. Distinguishing the map from the territory in TVFC research
When studying TVFC (and FC in general), it is critically important to ensure that one dis-
tinguishes between the method (p.ej., functional connectivity operationalized as statistical
dependence between time series) and the target theoretical properties we wish to infer (p.ej.,
interregional neural interactions). Failure to do so commits the logical fallacy of confusing the
map for the territory (Korzybski, 1933), y (to use a recent example from Reid et al., 2019)
“is akin to defining the moon as the photons that hit one’s retina when looking at a particular
location in the sky (a common method for detecting the moon), rather than as a physical ob-
ject with a variety of properties consistent with the laws of physics (theoretical properties of
interés)" (pag. 1751).
In the context of FC, it is relatively straightforward to define the map as estimates of sta-
tistical dependence between neurophysiological time series. This definition can be extended
to the case of TVFC by allowing these estimates to vary over time. Following Reid and col-
ligas, we define the territory as time-varying patterns of causal interaction between neural
entidades (p.ej., neuronas, poblaciones, redes). These interactions have many different prop-
erties we may be interested in, such as their direction (p.ej., A→B, B→A, etc.), directness (es decir.,
mono- or multisynaptic), and timing (es decir., when an interaction between two entities takes
lugar). While most FC methods applied to BOLD fMRI data are limited in the extent to which
they can provide information about the exact structure of the underlying causal graph, ellos
nonetheless constrain the space of possible network configurations (Reid et al., 2019). Estudios
of TVFC extend the FC paradigm by allowing researchers to make inferences about how this
space of possible network configurations changes over time, and how these changes relate to
cognition and behavior.
Given the somewhat controversial nature of FC research (Mehler & cordón, 2018), es
also worth emphasizing that FC estimates (both static and time-varying) can be useful above
and beyond any mechanistic information they may (or may not) provide about interregional
neural interactions and their relationship to cognition. Como se ha mencionado más arriba, patterns of FC
and TVFC are sensitive to individual differences in health and disease, and emerging work
suggests they may have powerful utility as clinical biomarkers (p.ej., as predictors of treatment
respuesta; Drysdale et al., 2017; Etkin et al., 2019; Reggente et al., 2018).
This paper is the result of a collaborative, open-invitation community effort to review the
current resting TVFC literature and to discuss key open questions and outstanding contro-
versies regarding this exciting new domain of research. As a group of scientists with diverse
perspectives on TVFC, we have attempted to reconcile and synthesize our views on contro-
versial issues, and to contextualize them in light of alternative opinions held by others in the
comunidad. While we offer some general suggestions for how researchers might best take ad-
vantage of the TVFC research program, we avoid making specific technical or methodological
recommendations except in cases where they are supported by the empirical literature.
We frame our discussion in terms of three broad questions: (1) Are rfMRI time series statisti-
cally consistent with functional connectivity that truly varies in time? (2) What is the biological
basis of BOLD TVFC (neural or otherwise)? (3) Qué (if any) is the cognitive and behavioral
relevance of resting BOLD TVFC? We begin with a survey of the current landscape of analytic
and modeling approaches for studying BOLD TVFC, and then proceed to address each of the
three questions outlined above. Primero, we review methodological considerations and statistical
challenges for studying TVFC in fMRI. Segundo, we review the literature on the physiological
Neurociencia en red
35
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
basis of BOLD TVFC. Tercero, we provide an in-depth discussion of the cognitive and behavioral
relevance of BOLD TVFC, including evidence both for and against this proposition. Subsequent
sections highlight experimental approaches that may help adjudicate questions about the cog-
nitive relevance of TVFC, and briefly review strategies for cleaning rfMRI data to mitigate the
impact of potential confounds on TVFC analyses. We conclude by suggesting ways that the
TVFC research community can continue to advance this exciting field and help facilitate con-
sensus on controversial issues.
ANALYTIC APPROACHES
Approaches to studying functional connectivity in fMRI data can be considered along a spec-
trum of temporal resolution. On one end, some methods assume that the dependence structure
(conectividad funcional) between regions is constant over an arbitrarily long time window (es decir.,
“static” FC); on the other end are methods that can estimate time-resolved FC at each individ-
ual time point (p.ej., instantaneous and sliding-window approaches). In between are methods
that aim to discover discrete, temporally contiguous functional connectivity states character-
ized by their interregional dependence structure (p.ej., sliding windows + clustering). In these
state-based models, the dependence structure changes only when moving between states.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Another important property of methods used to study TVFC is the extent to which they
consider the temporal ordering of the observed data points. Some approaches directly lever-
age the information in this ordering (p.ej., time-frequency approaches; Chang & guantero, 2010;
Yaesoubi, allen, Molinero, & Calhoun, 2015), while others ignore ordering completely and treat
data points as exchangeable samples (Liu, zhang, Chang, & duyn, 2018; Yaesoubi, Adali, &
Calhoun, 2018). Many common TVFC analysis pipelines have stages that alternately leverage
and neglect temporal ordering. Por ejemplo, one might begin by estimating sliding-window
correlations (calculated using time series with time points ordered as observed), apply k-means
clustering to the resulting time-resolved FC matrices (k-means ignores the temporal ordering
of the windows), and then evaluate state properties such as dwell times and transition proba-
bilities (which again considers the temporal order of time points; Allen et al., 2014).
Beyond differences in temporal resolution and sensitivity to time point ordering, methods
for studying TVFC can be considered as taking one of two broad conceptual approaches to
the challenge of studying brain dynamics. The first approach includes methods that attempt
to estimate changes in FC (and/or identify FC states) directly from the observed BOLD data
(p.ej., sliding windows, Sakoglu et al., 2010; clustering, Calhoun & Adali, 2016; and HMMs,
Vidaurre et al., 2017). The second approach includes methods that explicitly model the neural
processes underlying changes in the observed BOLD data (p.ej., simulations of the brain as a
dynamical system, romper la lanza, 2017; Parque, Friston, Pae, Parque, & Razi, 2018). These approaches
are complementary, and we expect future work on BOLD TVFC to increasingly make use of
these methods in combination. Abajo, we provide illustrative examples of each of the two
approaches, but emphasize that these are not meant as a comprehensive review of all extant
TVFC methods. Bastante, they are intended to provide a general idea of the breadth of available
methodological approaches. Cifra 2 illustrates common workflows for TVFC analyses, mientras
Mesa 2 provides a selection of key papers on BOLD TVFC, including a number of recent
reviews of TVFC methods.
Ejemplo 1: Data-Driven Methods for Estimating TVFC
One family of approaches for investigating time-varying functional connectivity focuses di-
rectly on the observed BOLD signal without explicitly modeling the underlying neural activity.
Neurociencia en red
36
Questions and controversies in the study of TVFC in resting fMRI
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Null model:
A model of the data-generating
process under the null hypothesis;
can generate synthetic data to
compare with observed data.
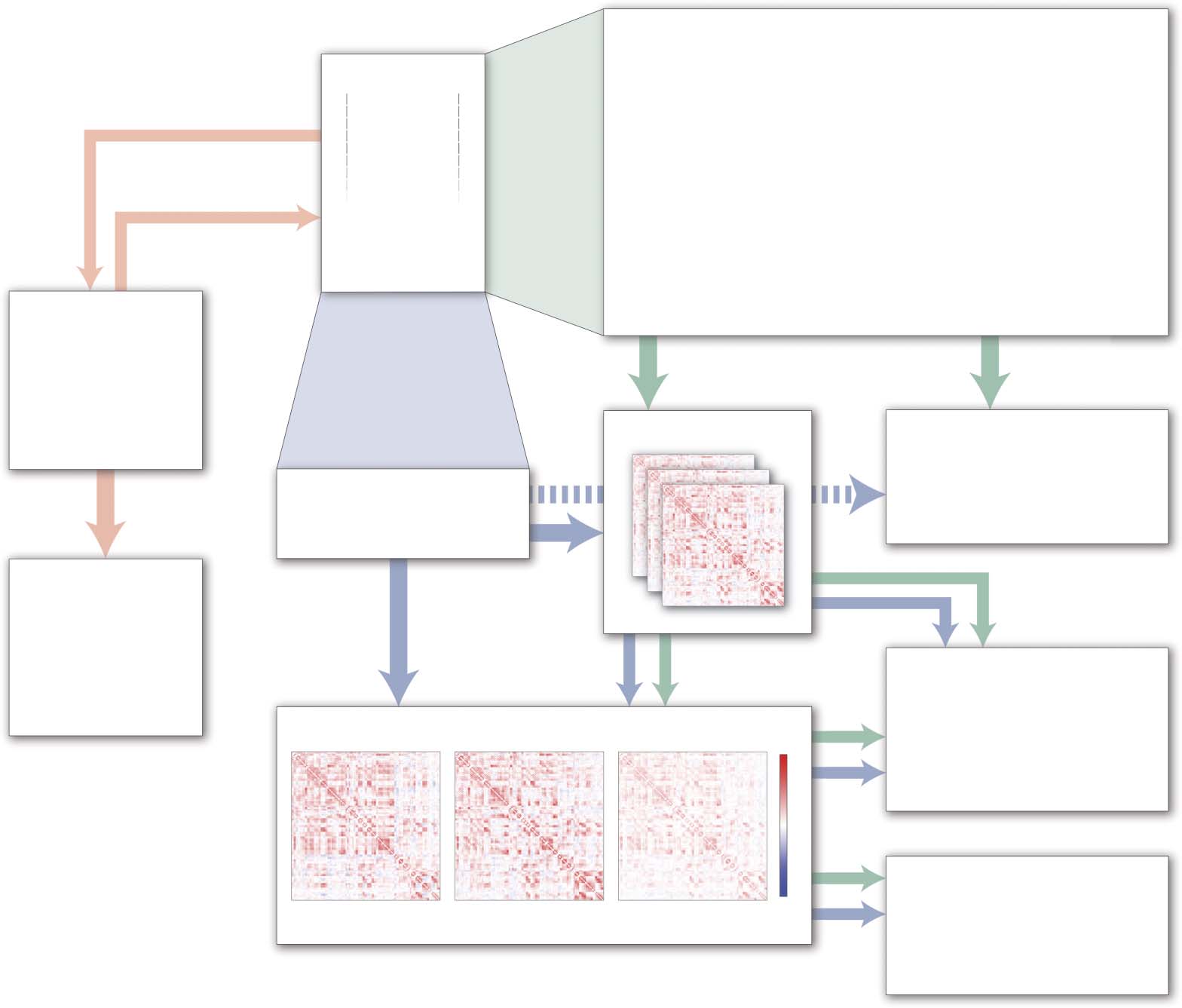
Cifra 2. Schematic illustration of common analysis and modeling approaches for studying TVFC
in fMRI data. Green arrows indicate a typical workflow based on sliding-window correlation, cual
is currently the most common data-driven approach for estimating TVFC. Blue arrows represent the
diversity of alternative data-driven approaches. Some alternative approaches (p.ej., HMM) estimate
functional connectivity states directly from BOLD time series, while others (p.ej., phase synchrony,
a time-frequency method) are more similar to the sliding-window approach. Regardless of how FC
time series or functional connectivity states are estimated, it is possible to calculate a wide range of
measures describing their properties. Por ejemplo, fluctuations in the strength of FC between two
areas can be tested for associations with concurrently measured behavioral variables, while network
measures can be used to describe the properties of whole-brain FC patterns and how they change
con el tiempo. Whether TVFC estimates are considered to constitute bona fide “dynamics” depends on
the specific feature of interest and null model against which they are tested. Orange arrows represent
a computational modeling workflow that fits a dynamic biophysical model to empirical BOLD time
series in order to estimate model parameters and simulate underlying fast timescale neural activity.
These techniques typically approach the observed fMRI data as multivariate time series and
seek to identify the time-resolved dependence structure between them. The most widely used
approach in this class estimates pairwise correlations within a sliding window, resulting in time-
resolved correlation matrices (one per window; Sakoglu et al., 2010). There are many variations
on this theme, including the type of window used (square, Sakoglu et al., 2010; tapered, allen
et al., 2014; or exponentially decaying, Lindquist, Xu, Nebel, & Caffo, 2014), the flexibility of
the window (fixed, Allen et al., 2014; or adaptive, Lindquist et al., 2014; Yaesoubi et al., 2015),
as well as the length of the window (Leonardi & Van De Ville, 2015; Liegeois et al., 2016;
Sakoglu et al., 2010; V. METRO. Vergara, Mayer, Damaraju, & Calhoun, 2017; Brilla & romper la lanza,
Neurociencia en red
37
Questions and controversies in the study of TVFC in resting fMRI
Mesa 2. Key papers on resting BOLD TVFC
A method for evaluating dynamic functional network connectivity and task-modulation: Application to schizophrenia
Sakoglu et al., 2010; https://doi.org/10.1007/s10334-010-0197-8
Time-frequency dynamics of resting-state brain connectivity measured with fMRI
Chang & guantero, 2010; https://doi.org/10.1016/j.neuroimage.2009.12.011
Published almost simultaneously, these two papers were among the first to apply sliding-window and time-frequency analyses to the study of BOLD TVFC.
Tracking whole-brain connectivity dynamics in the resting state
Allen et al., 2014 (published online in 2012); https://doi.org/10.1093/cercor/bhs352
One of the first papers to combine sliding-window analysis and clustering to estimate functional connectivity states and study their dynamics.
Dynamic BOLD functional connectivity in humans and its electrophysiological correlates
Tagliazucchi et al., 2012; https://doi.org/10.3389/fnhum.2012.00339
EEG correlates of time-varying BOLD functional connectivity
Chang et al., 2013; https://doi.org/10.1016/j.neuroimage.2013.01.049
Two of the earliest studies to explore the electrophysiological basis of BOLD TVFC using simultaneous EEG/fMRI.
Resting-state networks show dynamic functional connectivity in awake humans and anesthetized macaques
Hutchison et al., 2013; https://doi.org/10.1002/hbm.22058
One of the first studies to directly investigate the extent to which BOLD TVFC may exist independently of ongoing cognition.
Dynamic functional connectivity: Promise, asuntos, and interpretations
Hutchison et al., 2013; https://doi.org/10.1016/j.neuroimage.2013.05.079
Important early review of BOLD TVFC findings and methods.
Periods of rest in fMRI contain individual spontaneous events which are related to slowly fluctuating spontaneous activity
Petridou et al., 2013; https://doi.org/10.1002/hbm.21513
Time-varying functional network information extracted from brief instances of spontaneous brain activity
Liu and Duyn, 2013; https://doi.org/10.1073/pnas.1216856110
Two early studies suggesting that BOLD FC may be shaped by the dynamics of transient coactivation patterns (CAPs).
Time-resolved resting-state brain networks
Zalesky et al., 2014; https://doi.org/10.1073/pnas.1400181111
Early example of how sliding-window BOLD TVFC can be combined with graph theory analyses to investigate dynamic reorganization of functional brain
networks during rest.
Dynamic functional connectivity of the default mode network tracks daydreaming
Kucyi and Davis, 2014; https://doi.org/10.1016/j.neuroimage.2014.06.044
Early demonstration that resting BOLD TVFC is associated with time-resolved self-reports of ongoing cognition.
The chronnectome: Time-varying connectivity networks as the next frontier in fMRI data discovery
Calhoun et al., 2014; https://doi.org/10.1016/j.neuron.2014.10.015
Review of BOLD TVFC methods, including an in-depth discussion of approaches that seek to estimate functional connectivity states.
Evaluating dynamic bivariate correlations in resting-state fMRI: A comparison study and a new approach
Lindquist et al., 2014; https://doi.org/10.1016/j.neuroimage.2014.06.052
Can sliding-window correlations reveal dynamic functional connectivity in resting-state fMRI?
Hindriks et al., 2016; https://doi.org/10.1016/j.neuroimage.2015.11.055
On spurious and real fluctuations of dynamic functional connectivity during rest
Leonardi and Van De Ville, 2015; https://doi.org/10.1016/j.neuroimage.2014.09.007
Three papers that carefully evaluate the potential pitfalls of sliding-window approaches and emphasize the importance of comparing against null models.
Classification of schizophrenia and bipolar patients using static and dynamic resting-state fMRI brain connectivity
Rashid et al., 2016; https://doi.org/10.1016/j.neuroimage.2016.04.051
One of the first studies to demonstrate the superiority of BOLD TVFC over static FC for classifying individuals based on psychiatric diagnosis.
Neurociencia en red
38
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
The dynamic functional connectome: State-of-the-art and perspectives
Preti et al., 2017; https://doi.org/10.1016/j.neuroimage.2016.12.061
Detailed review of a wide range of methods for studying BOLD TVFC.
Mesa 2. continued
Temporal metastates are associated with differential patterns of time-resolved connectivity, network topology, and attention
Brillo y col., 2016; https://doi.org/10.1073/pnas.1604898113
A TVFC analysis of two large longitudinal single-subject datasets identified replicable temporal metastates with distinct functional network
topologies, time-varying properties, and associations with cognition.
On the stability of BOLD fMRI correlations
Laumann et al., 2017; https://doi.org/10.1093/cercor/bhw265
Influential paper challenging the notion that resting BOLD TVFC is related to ongoing cognition. Argues that resting BOLD is consistent with a stationary
process and that resting TVFC can largely be explained by sampling variability, apparent head motion, and fluctuations in arousal.
Interpreting temporal fluctuations in resting-state functional connectivity MRI
Liegeois et al., 2017; https://doi.org/10.1016/j.neuroimage.2017.09.012
Detailed exploration of which statistical properties are consistent with “dynamic” FC. Includes a detailed review of the concept of statistical stationarity, como
well as an assessment of several common statistical models.
Comparing test-retest reliability of dynamic functional connectivity methods
Choe et al., 2017; https://doi.org/10.1016/j.neuroimage.2017.07.005
Replicability of time-varying connectivity patterns in large resting state fMRI samples
Abrol et al., 2017; https://doi.org/10.1016/j.neuroimage.2017.09.020
Two of the first large, systematic evaluations of the reliability of methods for estimating BOLD TVFC and identifying functional connectivity states.
Brain network dynamics are hierarchically organized in time
Vidaurre et al., 2017; https://doi.org/10.1073/pnas.1705120114
HMM analysis reveals a rich hierarchical temporal structure in the pattern of transitions between FC states, and that individual differences in “meta
state” occupancy are related to cognition.
Dynamic models of large-scale brain activity
romper la lanza, 2017; https://doi.org/10.1038/nn.4497
Accessible review of methods for modeling large-scale brain dynamics. Includes a primer on core concepts from dynamical systems theory.
Neuronal origin of the temporal dynamics of spontaneous BOLD activity correlation
Matsui et al., 2019; https://doi.org/10.1093/cercor/bhy045
Simultaneous recording of calcium imaging and optical hemodynamics reveal a clear neural basis for BOLD TVFC, and that fluctuations
in BOLD TVFC are related to transient neural CAPs.
Simulations to benchmark time-varying connectivity methods for fMRI
Thompson et al., 2018; https://doi.org/10.1371/journal.pcbi.1006196
Recent work using multiple simulation strategies to undertake a systematic evaluation of the sensitivity of common TVFC methods.
Provides an open-source toolbox for simulation and benchmarking.
Putting the “dynamic” back into dynamic functional connectivity
Heitmann and Breakspear, 2018; https://doi.org/10.1162/netn_a_00041
Application of large-scale modeling to investigate which kinds of neural dynamics may give rise to BOLD TVFC. Argues that BOLD TVFC
likely reflects complex nonlinear and nonstationary neural dynamics.
2015). Otro (windowless) methods estimate FC without assuming locality of the neighboring
time points (Yaesoubi et al., 2018), or utilize time-frequency methods to estimate instantaneous
FC using phase synchrony (Chang & guantero, 2010; Pedersen, Omidvarnia, Brilla, & Jackson,
2018; Yaesoubi et al., 2015). Regardless of the particular method used, a common next step
is to assess the potential time-varying properties of the resulting time-resolved FC estimates,
and to explore possible associations with other dynamic phenomena (p.ej., behavioral per-
rendimiento, Verano, Esterman, Riley, & Valera, 2016; Patanaik et al., 2018; or cognitive state,
Verano & davis, 2014). TVFC estimates can also be summarized through the use of descriptive
Neurociencia en red
39
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
Estadísticas (p.ej., variance, Chang & guantero, 2010; Verano, Salomons, & davis, 2013) or meth-
ods that attempt to identify functional connectivity “states.” Methods for identifying states in-
clude sliding windows + clustering (p.ej., Allen et al., 2014), hidden Markov models (HMM;
Shappell, Caffo, Pekar, & Lindquist, 2019; Vidaurre et al., 2017), change-point modeling
(Cribben, Haraldsdottir, Atlas, Apostar, & Lindquist, 2012; Xu & Lindquist, 2015), and win-
dowless dynamic connectivity (Yaesoubi et al., 2018). After identifying states, it is possible
to estimate a variety of parameters such as mean dwell times, transition probabilities, y
graph theoretic measures that describe the observed FC patterns and brain dynamics (p.ej., net-
work modularity; Brilla, Proporcionó, cocineros, Perdido, & romper la lanza, 2014). These parameters can
then be probed for association with measures of inter- or intraindividual differences (p.ej., Beaty
et al., 2018; Marusak et al., 2018; Vidaurre et al., 2017). State-based approaches can differ in
whether they assume smooth transitions between states (Allen et al., 2014; Ou et al., 2015) o
instantaneous reconfigurations (Liu, zhang, et al., 2018; Yaesoubi et al., 2018), their focus on
a particular signal domain (p.ej., frequency, Yaesoubi et al., 2015; tiempo, Allen et al., 2014; o
espacio, S. Mamá, Calhoun, Phlypo, & Adali, 2014), and whether the state definitions are “hard” or
“soft” (es decir., whether each time point exhibits a single state, Allen et al., 2014; or is composed of
a mixture of multiple states, Leonardi, Shirer, Greicius, & Van De Ville, 2014; R. l. Miller et al.,
2016). Temporal network theory, a subfield of graph theory, can also be used to quantify how
functional network properties change over time (Holme & Saramäki, 2012; W.. h. Thompson,
Brantefors, & Fransson, 2017; Yu et al., 2015). En todos los casos, it is critical to benchmark these
Estadísticas (es decir., the TVFC estimates or state-related parameters) against those derived from refer-
ence data that embody a null or alternative hypothesis (p.ej., that FC is “static” and does not in
fact vary over time). We return to the issue of null models in the section on statistical challenges
in studying BOLD TVFC, abajo.
Ejemplo 2: Modeling the Underlying Neuronal Dynamics
In contrast to methods that seek to analyze the observed BOLD signal directly, a second family
of approaches instead aims to model the underlying neural fluctuations and interactions that
give rise to BOLD TVFC. This approach posits that observed BOLD time series are generated by
underlying nonlinear brain dynamics that are then corrupted by measurement noise. Under this
vista, activity in large-scale neural systems is inherently dynamic and exhibits complex phe-
nomena such as partial synchronization, multistable attractor landscapes, and edge-of-chaos
behavior indicative of criticality (cocineros, Perdido, Brilla, & romper la lanza, 2017; decoración, Jirsa,
robinson, romper la lanza, & Friston, 2008; Heitmann & romper la lanza, 2018; Roberts, Boonstra, &
romper la lanza, 2015; Zalesky et al., 2014). These dynamics generate physiological time series
with highly nonlinear structure and can be formally modeled by biophysically derived differ-
ential equations. By combining these equations with models of the observation process (p.ej.,
neurovascular coupling), it is possible to simulate how these underlying dynamics would mani-
fest in the BOLD signal (es decir., after the addition of measurement noise). There are a wide variety
of multiscale models of interconnected pools of neurons, including neural mass and neural
field models (Bojak, Oostendorp, Reid, & Kotter, 2010; romper la lanza, 2017; Deco et al., 2008).
These have been shown to produce neurobiologically plausible behaviors such as general-
ized synchronization, metastability, and multistability (romper la lanza, 2017; Deco et al., 2008;
Golos, Jirsa, & Dauce, 2015; Heitmann & romper la lanza, 2018; Roberts et al., 2019). Exploratory
computational work involves adjusting the model structure and tuning parameters in order
to obtain, through simulation, synthetic BOLD data that exhibits similar dependence structure and
dynamics to empirical observations (p.ej., decoración, Cruzat, & Kringelbach, 2019; Demirtas et al.,
2019; Kashyap & Keilholz, 2019; PAG. Wang y cols., 2019). Model-based approaches need to make
strong assumptions about the processes that generate observed BOLD data (Deco et al., 2008).
Neurociencia en red
40
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
Under these assumptions, it is possible to estimate from observed BOLD data the parameters
of these models, and thus the underlying neural dynamics (including time-varying aspects;
p.ej., Deco et al., 2019; Kashyap & Keilholz, 2019). This process is known as model inversion.
Models can be evaluated using a variety of methods (p.ej., information criteria) that consider
how well they fit observed data while penalizing model complexity. Careful model con-
struction facilitates the testing of specific hypotheses about underlying dynamics, así como
validation of findings from approaches that model the BOLD signal directly (Zalesky et al.,
2014).
A Rich Diversity of Methods for Studying TVFC
There is no single “best” method for studying time-varying functional connectivity; the choice
of analytic strategy should be informed by the available data and the particular questions un-
der investigation. Different approaches provide different (complementary) perspectives on the
datos, and a full understanding of the factors giving rise to TVFC and their relationship to cogni-
tion and behavior will likely necessitate integrating knowledge gained through the application
of a wide variety of methods (see Box 3). Some approaches (p.ej., Ejemplo 1) make minimal
(or no) explicit assumptions about the underlying biology, while others (p.ej., Ejemplo 2) buscar
to model the biophysical parameters directly. Improved biological specificity is often accom-
panied by greater model complexity and more extensive explicit model assumptions. Eso
dicho, methods that directly model the observed BOLD signal can also be highly statistically
articulated (p.ej., HMM) and come with their own assumptions (p.ej., that the data are best
represented by a limited number of states) that are often just as strong as assumptions made by
biophysical models.
Highly articulated “data-driven” models (p.ej., autoregressive models, Rogers, Katwal, morgan,
Asplund, & Sangre, 2010; or HMMs, Vidaurre, Abeysuriya, et al., 2018) may explain the data
very well without recourse to biological assumptions, but do not provide information about the
underlying neuronal dynamics without additional parameterization. As we learn more about
brain physiology and dynamics, additional biologically informed constraints can be added
to restrict the space of possible model solutions and improve the ability of these methods to
accurately describe the neural processes underlying noisy BOLD data. A diferencia de, dynami-
California (nonlinear) systems theory provides an adequately rich parameterization to enable explicit
exploration of how networks of neurons—modeled as coupled oscillators or populations of
spiking neurons—may give rise to the observed BOLD signal. Scientific investigation of TVFC
is likely to be enriched by the application of both approaches, as they have complementary
fortalezas, and the results from one perspective can inform the application of the other. por ejemplo-
amplio, data-driven models of the observed BOLD signal can yield new biological hypotheses,
eso, if confirmed, can then be integrated into richer empirically grounded dynamical models.
Two recent studies on how anatomical features shape static FC provide an excellent example of
how empirical work can inform modeling efforts, y viceversa. PAG. Wang et al. (2019) inferred
a hierarchy of recurrent anatomical connectivity across cortical regions by inverting a large-
scale dynamic circuit model fit to empirically observed resting fMRI data. Complementary
work by Demirtas et al. (2019) used MRI to map anatomical hierarchy as indexed by corti-
cal myelination, and found that incorporating hierarchy information into a biophysical model
of neural dynamics significantly improved the fit to human rfMRI data. Tomados juntos, estos
studies suggest that connectional hierarchy plays a fundamental role in shaping intrinsic neu-
ral dynamics. We expect that future work incorporating characteristics of empirically observed
TVFC into dynamical models will provide similarly important insights into brain organization
and function.
Neurociencia en red
41
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
Box 3. The elusive concept of dynamic functional connectivity
The term “dynamic functional connectivity” has been used to refer to a wide range of ap-
proaches for studying time-varying aspects of brain function. These approaches differ in the
insights they offer into brain dynamics, and it is important to distinguish which inferences can
(and cannot) be drawn from each method. Abajo, we briefly outline how four broad classes
of TVFC methods can be used to expand our understanding of brain function.
Time-resolved estimates of functional connectivity: Empirical estimates of time-resolved
functional connectivity allow scientists to explore how the strength of interregional coupling
varies over time. These estimates form the basis of empirical studies of TVFC. In their most
basic form (es decir., time-resolved correlations), they can provide insight into the trajectories by
which static (“time-averaged”) FC is realized. Time-resolved estimates also allow for fine-
grained evaluations of the relationship between FC and ongoing cognition, as well as how
summary measures (p.ej., variability of FC) may be related to phenotypic traits in health and
enfermedad.
Models of states and transitions: Many empirical studies of TVFC also seek to estimate tran-
sient “brain states” and their transitions. In this paradigm, each state describes a different
pattern of whole-brain activity or functional connectivity. Different models impose varying
constraints on the estimated states, such as whether they manifest in isolation (one state per
time point) or in combination (a mix of states at each time point). The dynamics of these states
(p.ej., time spent in each state, the probability of transitioning between states) can provide a
detailed portrait of how functional relationships reorganize through time. Formal model se-
lection and comparison (p.ej., using information-theoretic criteria) allows for the evaluation
of which models best describe the observed data, and thus permit adjudication of competing
hypotheses about data-generating processes.
Comparison to surrogate (null) datos: Insight into the dynamical properties of a system can
also be achieved by comparing observed data to surrogate data that lack a particular statistical
feature of interest. Por ejemplo, one can generate surrogate “null” time series that have the
same low-order features as empirical data (p.ej., significar, variance, spatiotemporal correlation
estructura) but lack a higher order feature proposed to exist in the real data (p.ej., switching
dinámica). The strength of this approach is that it draws from a rich existing literature on time
series analysis and enables testing of specific hypotheses about the dynamical properties of
an observed time series. Care must be taken to ensure that the tests undertaken are sufficiently
narrow and are interpreted as such. Por ejemplo, claims should be made about the presence
or absence of a particular statistical feature rather than “dynamic” FC in general, as “dynamic”
phenomena can exist under a wide range of conditions.
Modeling of nonlinear brain dynamics: Unlike the three approaches above that begin with
empirically measured BOLD data, it is also possible to instead begin the study of TVFC by
constructing a detailed biophysical model of the underlying processes thought to give rise to
TVFC. With appropriate model fitting and tuning, it is possible to invert the observed data into
a generative model, and then study the complex (fast timescale) dynamical properties of that
model that would normally be obscured by the measurement process. Having established
a model of the dynamical processes underlying the observed data, researchers can under-
take detailed mechanistic investigations of complex neural dynamics and their relationship
to BOLD TVFC.
STATISTICAL CHALLENGES IN STUDYING BOLD TVFC
Before diving into questions about the biological basis and cognitive relevance of resting BOLD
TVFC, we must first ask whether there is statistical evidence for this phenomenon: Does func-
tional connectivity estimated from resting BOLD fMRI actually vary over time? En esta sección,
we discuss the importance of testing TVFC estimates against null models, review the role of
Neurociencia en red
42
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
Sampling variability:
For a given distribution, how much a
computed statistic varies across
muestras (p.ej., time points, scans, o
Participantes).
sampling variability in TVFC estimation, and describe approaches for evaluating and validating
TVFC methods.
The Importance of Testing Against Null Models
Any method designed to estimate TVFC will inevitably return time-resolved estimates of func-
tional connectivity that vary to some degree with time (Lindquist et al., 2014). Researchers must
therefore carefully evaluate whether the observed TVFC estimates significantly deviate from
those that might have been obtained from time series generated by a process that lacks a par-
ticular property of interest (p.ej., state switching, fluctuating FC). It is then possible to compare
empirically observed time series with a suitable surrogate “null” distribution, typically gener-
ated through simulation or nonparametric resampling (romper la lanza, Brammer, bullmore, El,
& williams, 2004; Prichard & Theiler, 1994). Multiple methods have been developed to gen-
erate surrogate data, including methods that represent a null model based on a specific system
(Hindriks et al., 2016), biophysical models that simulate different classes of dynamics in the
cerebro (Heitmann & romper la lanza, 2018), and techniques that are designed to test the properties
of specific methods used to estimate TVFC (Allen et al., 2014; Shakil, Sotavento, & Keilholz, 2016).
When evaluating TVFC through comparison with null models, it is important to carefully
consider both the features of the process used to generate null data, as well as the test statistic
used to evaluate whether observed TVFC estimates deviate from that null. Por ejemplo, Alabama-
though some work has focused on statistical stationarity as a feature of interest (Laumann et al.,
2017), subsequent work (Liegeois et al., 2017; R. l. Miller et al., 2018) has demonstrated that
the space of stationary models includes many processes that exhibit TVFC (p.ej., HMMs with
switching covariance structure). De este modo, statistical stationarity is not necessarily tantamount to
static functional connectivity. En cambio, evidence of nonstationarity does not always imply
the presence of a “meaningful” change and/or trend in the data (Koutsoyiannis, 2011; Lins,
2012). Asimismo, it is important to keep in mind that TVFC estimates that fail to differ signifi-
cantly from a given null do not necessarily equate to “meaningless fluctuations.” Rather, semejante
fluctuations could be consistent with a more restricted space of stationary stochastic models
that may still have scientifically interesting properties (es decir., have heavy spatial and temporal
tails; Cocchi et al., 2017; R. l. Miller et al., 2018; Roberts et al., 2015).
It remains an open question which time series features and null models are most appropri-
ate for evaluating various aspects of TVFC, and as such we refrain from making any specific
recomendaciones. dicho eso, the case of statistical stationarity provides a good example of
the process by which one might assess the properties of null models and time series features,
test for the presence of candidate features in empirical data, and interpret the results of these
analiza. Laumann et al. (2017) proposed testing for the presence of TVFC by evaluating the
multivariate kurtosis of rfMRI time series, with multivariate kurtosis used as a test statistic to
assess the stationarity of the time series, and stationarity used as an index of the extent to
which the time series exhibit “dynamic” fluctuations in FC. The values of multivariate kurtosis
observed by Laumann et al. were insufficient to reject the null of a stationary process, y el
authors interpreted this finding as evidence against the presence of TVFC in rfMRI. Sin embargo,
as mentioned above, subsequent analyses by Liegeois et al. (2017) found that multiple com-
monly used “dynamic” models (p.ej., HMM, autoregressive models) are statistically stationary,
and that this stationarity exists even for models with switching covariance structure (HMM).
Additional work by R. l. Miller et al. (2018) found that time series properties leading to ele-
vated multivariate kurtosis (which Laumann et al. interpreted as evidence of nonstationarity)
are sometimes more consistent with stationary than nonstationary processes. Tomados juntos,
these results suggest that (a) stationary processes are consistent with the presence of TVFC,
Neurociencia en red
43
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
y (b) multivariate kurtosis is likely a poor proxy for statistical stationarity. More generally,
the papers by Laumann, Liegois, and Miller provide an excellent example of how the research
community can work together to begin establishing a consensus on which time series proper-
ties and null models are most appropriate for testing various aspects of TVFC.
The Role of Sampling Variability
Sampling variability is a key consideration for statistical inference. BOLD FC is typically estimated
as the bivariate correlation between two time series, and a peculiar property of correlations of
time series (first discussed over 90 years ago; Bartlett, 1935) is that one can obtain high corre-
lation coefficients even in the absence of a real relationship. Este fenómeno (resultante de
autocorrelation) can largely be summarized as an issue of sampling variability, which refers to
how much a statistic varies across realizations of the data. The lower the sampling variability,
the more precise the subsequent inference (p.ej., confidence intervals and hypothesis tests).
Como ejemplo, consider the sampling variability of the sliding-window approach. Porque
sliding windows (and other TVFC methods) estimate a series of correlations, it can be useful to
think of these values as “repeated samples” of correlations across time. Desde esta perspectiva,
the key question being asked when evaluating TVFC estimates is whether each sample was
drawn from the same distribution (static FC) or from distinct distributions (TVFC). If we choose
a small window size, the correlation coefficient will be based on few data points; this gives
rise to larger sampling variability. De este modo, short window lengths may give rise to signals that
show compellingly “dynamic” changes in correlation across time, even if the FC is actually
static (Hlinka & Hadrava, 2015; Leonardi & Van De Ville, 2015; Lindquist et al., 2014). Este
problem becomes less pronounced as window length increases, but longer windows come at
the cost of reduced sensitivity to transient changes in correlation. Además, if overlapping
windows are used, an autocorrelation (beyond that already present because of the smoothness
of the BOLD signal) is induced in the estimated TVFC values, which can make changes in FC
appear artificially smooth (Lindquist et al., 2014). dicho eso, trabajo reciente (V. Vergara, Abrol,
& Calhoun, in press; V. METRO. Vergara et al., 2017) suggests that the optimal window length to
minimize these concerns may be shorter than the minimum of ∼60 s that has been previously
recommended (Leonardi & Van De Ville, 2015; Brilla & romper la lanza, 2015), and one can
consider the choice of window size to be a tunable filter that can be optimized based on the
question of interest (Lindquist et al., 2014; V. Vergara et al., in press).
Establishing the Sensitivity and Reliability of TVFC Methods
Prior to the use of any new method, it is crucial to systematically evaluate the accuracy and
reliability of its performance. One key metric of algorithmic accuracy is sensitivity, which for
TVFC methods is the ability to accurately recover TVFC from noisy data. As the “ground truth”
of the fluctuating neural interactions underlying TVFC is often unknowable (and perhaps even
undefined), evaluations of sensitivity typically make use of simulated data containing a known
TVFC signal of interest (es decir., a particular pattern of time-varying dependence structure). A
variety of simulation tools are available to help researchers evaluate how TVFC methods per-
form under a range of different data-generating conditions (Erhardt, allen, Wei, Eichele, &
Calhoun, 2012; Sanz Leon et al., 2013; W.. h. Thompson, Richter, Plaven-Sigray, & Fransson,
2018; Welvaert & Rosseel, 2014). Results from sensitivity analyses suggest not only that dif-
ferent TVFC methods have different degrees of sensitivity, but that sensitivity is influenced by
factors such as window length and data quantity (es decir., scan duration; Hindriks et al., 2016;
W.. h. Thompson et al., 2018).
Neurociencia en red
44
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
It is also critical to demonstrate that estimates of BOLD TVFC are reliable enough to serve as
robust markers of ongoing cognition and/or individual differences. Recent work has shown that
whole-brain patterns of TVFC at rest are largely reproducible across individuals (Abrol et al.,
2017; Choe et al., 2017; Vidaurre, Abeysuriya, et al., 2018), even when considering data from
multiple scan sites and heterogeneous populations (Abrol et al., 2017). Complementary work
has shown that individual differences in resting TVFC dynamics show good test-retest reliability
(Choe et al., 2017; Liao, Cao, Xia, & Él, 2017; Vidaurre, Abeysuriya, et al., 2018). Estos
studies satisfy an important prerequisite for continued research into resting TVFC, and future
work should continue to refine our understanding of which factors influence the reliability of
estas medidas (Lehmann, Blanco, Henson, Leva, & Geerligs, 2017). Additional work is also
necessary to assess which properties of TVFC are stable over time within an individual (es decir.,
“trait” characteristics; Geerligs, Rubinov, Leva, & Henson, 2015) and which are modulated by
the particular experimental context or cognitive state.
THE BIOLOGICAL BASIS OF BOLD TVFC
Cross-Modal Comparisons of BOLD TVFC and Direct Measures of Neural Activity
FMRI is unique in its ability to noninvasively measure and localize activity simultaneously
across the entire brain at relatively high spatial resolution. This has made it the modality of
choice for many researchers interested in understanding large-scale brain dynamics (especially
in humans). Sin embargo, the BOLD signal is a noisy, indirect measure of underlying neural activ-
idad, and the sluggish hemodynamic response places a fundamental limit on the temporal resolu-
tion of TVFC estimated from fMRI data. It is well established that the shape of the hemodynamic
response function varies across brain areas (Handwerker, Ollinger, & D'Esposito, 2004) y
individuals (Aguirre, Zarahn, & D'Esposito, 1998), and emerging work suggests that neurovas-
cular coupling may also vary across behavioral and bodily states (Elbau et al., 2018; Lecrux
& Hamel, 2016; Winder, Echagarruga, zhang, & Dibujó, 2017). While these and other fac-
tors can complicate the neurophysiological interpretation of fMRI findings, we do not believe
they preclude the use of BOLD fMRI for studies of time-varying neural interactions. Bastante,
they strongly motivate the need to validate and extend findings from fMRI through comparison
with other modalities.
If we wish to use BOLD TVFC to study temporal fluctuations in interregional neural in-
teractions, it is first necessary to establish a firm neural basis for regional BOLD activity and
conectividad funcional. Intracranial recordings have consistently revealed a positive corre-
lation between the regional BOLD signal and electrophysiological high-frequency broadband
fuerza (∼ 50 − 150 Hz, also sometimes referred to as “high gamma”; Logothetis, Pauls, Augath,
Trinath, & Oeltermann, 2001; k. j. Molinero, Weaver, & Ojemann, 2009; Mukamel et al., 2005;
Nir et al., 2007; Scholvinck, Maier, S.M, duyn, & Leopold, 2010), and patterns of FC estimated
from fluctuations in high-frequency broadband power reliably exhibit similar topography to
intrinsic BOLD FC networks when the two modalities are compared within the same individu-
como (Foster, Rangarajan, Shirer, & Parvizi, 2015; hacker, Snyder, Pahwa, Corbetta, & Leuthardt,
2017; Él, Snyder, Zempel, Smyth, & Raichle, 2008; Verano, Schrouff, et al., 2018). Los estudios han
also observed correspondence between patterns of BOLD FC and interareal correlations in the
band-limited power of a range of lower frequencies (p.ej., delta, theta, alfa, and beta), cual
can be detected using both invasive electrophysiology (Foster et al., 2015; Hacker et al., 2017;
Lu et al., 2007; l. Wang, Saalmann, Pinsk, Arcaro, & Kastner, 2012) and MEG (Baker et al.,
2014; Brookes et al., 2011; Hipp, Hawellek, Corbetta, Siegel, & ángel, 2012; Hipp & Siegel,
Neurociencia en red
45
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
4
1
3
0
1
8
6
6
7
0
9
norte
mi
norte
_
a
_
0
0
1
1
6
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Questions and controversies in the study of TVFC in resting fMRI
2015; Houck et al., 2017). The correspondence observed between BOLD FC and electrophys-
iological FC at these lower frequencies may be specific to particular functional brain networks
(Hacker et al., 2017; Hipp & Siegel, 2015).
Recientemente, multimodal recording approaches have been adopted to directly investigate the
neurophysiological basis of BOLD TVFC (see G. j. Thompson, 2018, para una revisión). These stud-
ies suggest that like static FC, BOLD TVFC may reflect fluctuations of electrophysiological FC
across multiple frequency bands. Simultaneous fMRI and intracranial recordings in rats found
that BOLD TVFC between left and right somatosensory areas tracked changes in FC calculated
from band-limited electrophysiological power, and that these associations exist across several
canonical frequency bands (theta, beta, and gamma; GRAMO. j. Thompson, Merritt, et al., 2013). Pre-
liminary support for these relationships in humans comes from TVFC analyses of simultane-
ously recorded EEG-fMRI data, which have found associations between BOLD TVFC and changes
in power across multiple frequency bands (delta, theta, alfa, beta, and low-gamma; allen,
Damaraju, Eichele, Wu, & Calhoun, 2018; Chang, Liu, Chen, Liu, & duyn, 2013; Tagliazucchi
et al., 2012). Desafortunadamente, because of the poor spatial resolution of EEG, these studies are
unable to speak directly to the electrophysiological basis of spatially specific variations in cou-
pled activity between brain regions. Sin embargo, studies using MEG (which provides improved
spatial localization relative to EEG for many cortical regions) have observed time-varying inter-
regional correlations of band-limited power that have similar spatial topography to BOLD FC
redes (de Pasquale et al., 2010; Vidaurre, Hunt, et al., 2018). The temporal correspondence
of these effects with BOLD TVFC remains uncertain, as MEG cannot be recorded simultane-
ously with fMRI.
Studies have shown that fluctuations in local field potentials at low frequencies directly
comparable to BOLD fluctuations (<1 Hz) also contribute substantially to measures of func-
tional connectivity and TVFC (Grooms et al., 2017; He 2008; Hiltunen 2014; Pan,
Thompson, Magnuson, Jaeger, & Keilholz, 2013; G. J. Thompson, Pan, &
Keilholz, 2014). Notably, these infraslow fluctuations in neural activity have been linked to
quasiperiodic spatiotemporal patterns BOLD that involve coordinated propa-
gation across the brain al.,
2014). In this vein, studies successfully modeled as being driven by tran-
sient periods high-amplitude coactivations (Karahanoglu Van De Ville, 2015;
Tagliazucchi, Siniatchkin, Laufs, Chialvo, 2016). These coactivation are reliable
across individuals (Gutierrez-Barragan, Basson, Panzeri, Gozzi, 2019; Liu Duyn, 2013) and
may preferentially occur at distinct phases global signal (Gutierrez-Barragan
et 2019). addition observing a close correspondence between windowed cal-
culated from optically imaged hemodynamic signals simultaneously recorded calcium
transients, recent work rodents found variation transient was
associated with TVFC, neither calcium nor were
consistent simulations assumed stationary covariance structure (Matsui, Murakami,
& Ohki, 2019).
Overall, it seems possible (even probable) multiple, potentially dissociable neurophys-
iological processes time-varying functional
connectivity, there is good reason believe heterogeneity electrophysiological
frequency bands reported associated static FC not merely
artifactual or due experimental variability. Different electrophysiological activity
likely reflect neurophysiological (Buzsaki Draguhn, 2004), recent
work human intracranial recordings—including within prominent nodes canonical
Network Neuroscience
46
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
>