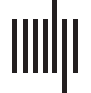INVESTIGACIÓN
Synchronization lag in post stroke: relation to
motor function and structural connectivity
Xin Wang1, Caio Seguin2, Andrew Zalesky2,3, Wan-wa Wong1,
Winnie Chiu-wing Chu4, and Raymond Kai-yu Tong1
1Department of Biomedical Engineering, The Chinese University of Hong Kong, Hong Kong, Porcelana
2Melbourne Neuropsychiatry Centre, Department of Psychiatry, Universidad de Melbourne, Melbourne, Australia
3Department of Biomedical Engineering, Universidad de Melbourne, Melbourne, Australia
4Department of Imaging and Interventional Radiology, The Chinese University of Hong Kong, Hong Kong, Porcelana
Palabras clave: Functional lag, Chronic stroke, Brain navigation, Network efficiency, Motor function
un acceso abierto
diario
ABSTRACTO
Stroke is characterized by delays in the resting-state hemodynamic response, Resultando en
synchronization lag in neural activity between brain regions. Sin embargo, the structural basis
of this lag remains unclear. en este estudio, we used resting-state functional MRI (rs-fMRI) a
characterize synchronization lag profiles between homotopic regions in 15 individuals
(14 machos, 1 femenino) with brain lesions consequent to stroke as well as a group of healthy
comparison individuals. We tested whether the network communication efficiency of each
individual’s structural brain network (conectoma) could explain interindividual and
interregional variation in synchronization lag profiles. Para tal fin, connectomes were
mapped using diffusion MRI data, and communication measures were evaluated under
two schemes: shortest paths and navigation. We found that interindividual variation in
synchronization lags was inversely associated with communication efficiency under both
schemes. Interregional variation in lag was related to navigation efficiency and navigation
distancia, reflecting its dependence on both distance and structural constraints. Además,
severity of motor deficits significantly correlated with average synchronization lag in
stroke. Our results provide a structural basis for the delay of information transfer between
homotopic regions inferred from rs-fMRI and provide insight into the clinical
significance of structural-functional relationships in stroke individuals.
INTRODUCCIÓN
Neuroplasticity refers to the brain’s ability to form new connections throughout life, cual
is particularly evident in response to traumatic events and stroke. Brain lesions are a long-
term consequence of stroke, resulting in deficits in language, motor, or cognition, depending
on the lesion location. Through neuroplastic processes, the lesioned brain can reorganize its
estructura, función, and connections to adapt to such damage (Dimyan & cohen, 2011; Kolb
& Mahoma, 2014). Understanding how brain structure and function reorganize after stroke
is crucial in managing stroke recovery.
Resting-state functional MRI (rs-fMRI) has been widely used to characterize brain activity
in the absence of external stimuli (Smith et al., 2013). Functional connectivity can be inferred
from rs-fMRI data and quantifies the statistical dependence (synchrony) between neural ac-
tivity at distant brain regions. Previous rs-fMRI studies have delineated intrinsic resting-state
networks that can be ascribed to distinct functions, such as vision, executive control, y
prominencia (van den heuvel & Hulshoff Pol, 2010; Yeo et al., 2011). Regions comprising a
Citación: Wang, X., Seguin, C., Brilla,
A., Wong, w., Chu, W.. C., & Tong, R. k.
(2019). Synchronization lag in
poststroke: relation to motor function
and structural connectivity. Red
Neurociencia, 3(4), 1121–1140.
https://doi.org/10.1162/netn_a_00105
DOI:
https://doi.org/10.1162/netn_a_00105
Supporting Information:
https://doi.org/10.1162/netn_a_00105
Recibió: 30 Marzo 2019
Aceptado: 25 Julio 2019
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Raymond Kai-yu Tong
kytong@cuhk.edu.hk
Editor de manejo:
Olaf Sporns
Derechos de autor: © 2019
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
norte
mi
norte
_
a
_
0
0
1
0
5
pag
d
t
/
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Structural basis for functional lag in chronic stroke
common intrinsic network normally exhibit relatively synchronized neural activity. Cómo-
alguna vez, synchronization in blood oxygen level-dependent (BOLD) activity between distant (o
homotopic) regions comprising the same intrinsic network shows inherent lag due to the time
required for signal propagation as well as other neural processes (Mitra, Snyder, hacker, &
Raichle, 2014; Mitra, Snyder, Blazey, & Raichle, 2015). This means that the BOLD activity
measured at one region is most strongly synchronized with a delayed, or temporally shifted
(lagged), version of the BOLD activity at a distant region. These lags in BOLD synchroniza-
tion can exceed a second in the healthy human brain and may reflect neuronal processes, como
opposed to hemodynamic delay (Mitra et al., 2014; Mitra et al., 2018).
Recientemente, Siegel and colleagues suggested that synchronization lag measured at rest could
serve as a potential index of neuroplasticity that indicates behavioral recovery in individuals
with stroke (Siegel, Snyder, Ramsey, Shulman, & Corbetta, 2016). Increased lag in BOLD sig-
nal synchronization following stroke could be due to two possible reasons. Por un lado,
from a local hemodynamic perspective, brain regions share the same vascular supply. Once the
shared vascular supply is interrupted because of the effects of cerebral infarction, the hemody-
namic response of the intact regions could also be affected, thus leading to increased lag in syn-
chronization (Amemiya, Kunimatsu, saito, & Ohtomo, 2014; Lv et al., 2013; Siegel, Shulman,
& Corbetta, 2017). Por otro lado, from a whole-brain perspective, locally damaged tissue
may have a brain-wide impact on the structural and functional reorganization of brain net-
obras. Studies have shown that even if structural damage is focal, the function of regions
far from the locally damaged site can change after stroke (Carretero, Shulman, & Corbetta, 2012;
Crofts et al., 2011), including additional recruitment of contralesional motor areas (Lotze et al.,
2006; Pequeño, Hlustik, noll, Genovese, & Solodkin, 2002) and increased activity in the non-
primary motor areas (Tombari et al., 2004; Wong, chan, Espiga, Meng, & Tong, 2013) cuando
performing motor activities. Por eso, signal propagation between distant regions may be de-
layed as a consequence, leading to increased synchronization lag after stroke. This increase
in lag may be related to structural-functional reorganization, which has not been extensively
investigated in stroke.
Connectomics (Proporcionó, Brilla, & bullmore, 2016) enables quantitative investigation of
structural and functional reorganization in individuals with stroke (Caliandro et al., 2017;
Laney, Adali, McCombe Waller, & Westlake, 2015; Siegel et al., 2018; Silasi & Murphy, 2014).
With this approach, brain connectivity is represented as a graph (network), enabling study of
how synchronization in neural activity (es decir., conectividad funcional) is facilitated by signal
propagation and communication through the brain’s network of white matter pathways. Estos
pathways comprise a sequence of structural connections (white matter fascicles), which can
be mapped using diffusion MRI and fiber tracking techniques (Mori, Crain, Chacko, & van Zijl,
1999; Sarwar, Ramamohanarao, & Brilla, 2019). Signal propagation across the brain’s struc-
tural network is conjectured to follow certain communication schemes (Avena-Koenigsberger,
Miši´c, & despreciar, 2017). A communication scheme dictates the polysynaptic paths that informa-
tion traverses between pairs of regions that are not directly connected by a white matter fiber.
Two widely studied communication schemes are (a) shortest paths (bullmore & despreciar, 2009;
Rubinov & despreciar, 2010) y (b) navigation (Boguñá, Krioukov, & Claffy, 2009; Kleinberg,
2000; Seguin, van den heuvel, & Brilla, 2018). Shortest paths traverse the minimal num-
ber of synapses (es decir., intermediate nodes) from one region to another. In weighted networks,
shortest paths are often determined to minimize the sum of edge weights traversed. El COM-
putation of shortest paths mandates assumptions that might be considered unrealistic in bio-
logical systems, such as individual regions possessing global knowledge of the full network
architecture (Seguin et al., 2018). Navigation relaxes some of these unrealistic assumptions
synchronization lag:
The temporal delay between the
fMRI signals measured at two
distinct brain regions, típicamente
computed using the
cross-correlation function.
navigation:
A network communication model
describing how information is routed
across multi-hop (polysynaptic)
paths in a network.
Neurociencia en red
1122
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
norte
mi
norte
_
a
_
0
0
1
0
5
pag
d
/
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Structural basis for functional lag in chronic stroke
spatially-embedded network:
A network with an inherent spatial
layout and/or geometric attributes,
contrast to networks that are
purely topological.
scale-free network:
A network where the distribution of
the node degree follows a power law,
containing hub nodes with a degree
that substantially exceeds the average
degree in the network.
homotopic regions:
The same regions but in different
hemispheres. Inter-hemispheric
functional connectivity is typically
stronger between mirror (homotopic)
regions of the brain.
and thereby yields a more biologically plausible model of neural communication than short-
est paths. Under navigation, if a region aims to communicate with another region with which
it is not directly connected, it establishes a path by progressing to an intermediate region to
which it is directly connected. If there are multiple intermediate regions to which it is directly
conectado, the region that is closest in distance to the desired destination is chosen. Naviga-
tion performs particularly well in spatially embedded, scale-free networks like the brain. El
feasibility of navigation in brain networks has recently been demonstrated (Allard & serrano,
2018; Seguin et al., 2018).
Compensatory and recovery mechanisms have been extensively described in the litera-
tura, including enhanced axonal growth between areas that are normally not connected in the
vicinity of a lesion and long-distance cortico–spinal axonal sprouting (Buchli & Schwab, 2005;
Dancause et al., 2005; Wieloch & Nikolich, 2006). Alternative paths that are established to
compensate for lesion-induced disruptions in connectivity may traverse longer silent pathways
(latent pathways that are inactive) or synapses that are unmasked or activated (Cramer, 2008;
Murphy & Corbett, 2009), leading to communication with reduced efficiency and longer de-
lays. Sucesivamente, longer delays in communication could potentially manifest as synchronization lag
in resting-state functional connectivity. Finding evidence of putative rerouting of neural infor-
mation flow following stroke can provide new insights into stroke recovery. Además, cualquier
potential rerouting of information could be indexed using graph-theoretic measures like com-
munication efficiency and communication distance. These measures may explain interindivid-
ual or interregional variation in synchronization lag inferred from rs-fMRI, thereby providing
a link between structural connectivity changes and functional lag. We hypothesized that in-
creased synchronization lag would be associated with lower communication efficiency and
longer communication distance in the post-stroke connectome.
En el estudio actual, we aim to investigate the structural basis of homotopic synchroniza-
tion lag in a group of stroke individuals with predominant motor deficits. More specifically, nosotros
first aim to test whether interindividual variation in synchronization lag inferred from rs-fMRI
can be explained by the efficiency with which information can be communicated through
each individual’s post-stroke structural connectome. We then aim to test whether interregional
variation in synchronization lag between homotopic regions associates with measures of re-
gional communication efficiency and distance computed in the structural connectome. Nosotros
thus seek to establish a link between functional alterations and impaired structural communi-
catión. Finalmente, we test whether synchronization lag can predict to severity of motor deficits.
Colectivamente, we aim to develop new insight into recovery mechanisms relating to interregional
communication after brain damage due to stroke.
RESULTADOS
Demographic and Clinical Characteristics
Demographic and clinical characteristics of the stroke individuals are listed in Table 1. El
stroke patients had moderate-to-severe upper-limb impairment (Action Research Arm Test
[ARAT]: 12.62 ± 6.54). More patients had lesions in the right hemisphere (norte = 10) than in
the left hemisphere (norte = 5), with the majority of infarcts in the territory irrigated by the ante-
rior and middle cerebral arteries. Lesions spanned the following regions: putamen (norte = 11),
insula (norte = 10), rolandic operculum (norte = 6), inferior frontal gyrus (norte = 6), and temporal pole
(norte = 4). There was no significant difference (pag = 0.165) in age between the stroke (age =
54 ± 8 años) and control (age = 58 ± 3 años) grupo.
Neurociencia en red
1123
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
norte
mi
norte
_
a
_
0
0
1
0
5
pag
d
t
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Structural basis for functional lag in chronic stroke
Mesa 1. Demographics and Clinical Characteristics of the Participants
Lesion locations
R
l
R
l
R
R
R
l
R
l
R
R
R
l
R
Stroke type Stroke onset (y) ARAT
55–59
60–64
45–49
65–69
65–69
45–49
60–64
55–59
55–59
50–54
40–44
40–44
55–59
50–54
45–49
54 ± 8
Age range Gender Lesion side
METRO
METRO
METRO
METRO
METRO
METRO
METRO
METRO
METRO
METRO
METRO
METRO
METRO
F
METRO
brainstem
PLIC, putamen
MFG, SFG, precentral, supramarginal, SMA
insula, putamen, IFG, temporal pole
insula, ITG, IOG, putamen
ITG, MTG, STG, MOG, angular, supramarginal
insula, putamen, rolandic operculum, IFG
insula, IFG, putamen
insula, IFG, putamen, rolandic operculum, temporal pole
putamen, caudate nucleus
insula, rolandic operculum, IFG, STG, putamen, temporal pole
insula, MTG, STG, putamen, temporal pole, rolandic operculum
insula, rolandic operculum, IFG
insula, rolandic operculum, putamen
insula, putamen
Subject
28
S1
14
S2
19
S3
15
S4
12
S5
4
S6
15
S7
10
S8
8
S9
9
S10
11
S11
3
S12
16
S13
10
S14
12
S15
12 ± 6
Mean ± SD
Nota. y = year; M = male; F = female; R = right hemisphere lesion; L = left hemisphere lesion; IFG = inferior frontal gyrus; IOG =
inferior occipital gyrus; ITG = inferior temporal gyrus; MFG = middle frontal gyrus; MOG = middle occipital gyrus; MTG = middle
temporal gyrus; PLIC = posterior limb of the internal capsule; SFG = superior frontal gyrus; SMA = supplementary motor area;
STG = superior temporal gyrus; H = hemorrhagic stroke; I = ischemic stroke; ARAT: Action Research Arm Test (maximum: 57);
SD = standard deviation.
11
11
1
8
1
0.67
3
5
7
1
5
3
6
3
1
4 ± 3
I
I
I
h
h
h
I
h
I
I
h
h
I
h
h
Increased Synchronization Lag in Stroke
Average synchronization lag across 40 homotopic regions was compared between the stroke
and healthy comparison group. Lag was significantly increased in the stroke group compared
with controls (stroke: mean = 1.07 s, controls: mean = 0.26 s; pag < 0.0001, t = 5.51). The
distribution of synchronization lags (Figure 1) across the 40 homotopic regions suggests a sub-
stantially longer tail in the distribution for the stroke individuals, extending to lags of 10 seconds
for some regional pairs, whereas lag was typically below a second in the healthy comparison
group. Each of the 40 pairs of homotopic regions was tested independently to identify which
pairs were associated with the longest increase in lag compared with controls. Among the 40
pairs of homotopic regions, pallidum (t = 2.80, p = 0.0097, uncorrected) and middle temporal
gyrus (t = 3.32, p = 0.0027, uncorrected) tended to show longer lag than those in the control
group. However, none of these 40 pairs of regions survived control of the false discovery rate
at 5%.
Considering the relatively small sample size of the control group and the scanner differ-
ence, we also tested the lag distribution in two additional healthy comparison groups, includ-
ing individuals participating in the Human Connectome Project (HCP) (Supporting Information
Figure S1). The increased temporal resolution of the HCP data (0.72 s) enabled estimation of
lag with greater precision than the clinical data (2 s). We found that the stroke individuals were
associated with significantly longer lags when compared with the group of age-matched con-
trol (control 2: mean = 0.30 s, p < 0.0001, t = 6.74) and healthy comparison individuals from
the HCP (HCP control: mean = 0.37 s, p < 0.0001, t = 12.46). These findings are summarized
in Table 2. Moreover, we found that synchronization lag did not significantly differ between
the three groups of healthy comparison individuals (F = 0.493; p = 0.611). This suggests that
interscanner variation is an unlikely explanation for the longer synchronization lags found in
the stroke individuals.
Network Neuroscience
1124
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Distribution of synchronization lag in stroke (yellow) and healthy comparison individu-
als (blue, red) across 40 homotopic regions. The distribution incorporates all 40 pairs of regions and
all individuals. Two healthy comparison datasets acquired using different scanners and acquisition
protocols were used here. Lag is binned to a temporal resolution of 0.5 seconds. The inset shows
the distribution without the lag of zero in order to represent the tail of the distribution more clearly.
We next sought to investigate whether stroke-related increases in synchronization lag were
global or circumscribed to specific subnetworks. To this end, each of the 40 regional pairs
were assigned to one of seven intrinsic resting-state subnetworks delineated in previous stud-
ies (Schirner, McIntosh, Jirsa, Deco, & Ritter, 2018; Yeo et al., 2011). The seven subnetworks
were as follows: default mode, executive control, sensorimotor, fronto-parietal, auditory, vi-
sual, and subcortical networks (Supporting Information). For each individual, each of these
seven subnetworks was then classified as either affected or unaffected, according to whether
the subnetwork encompassed a lesion or not, respectively. The set of all affected subnetworks
was called the affected network, and the remaining subnetworks were collectively referred to
as the unaffected network. In this way, an affected and unaffected network was defined for
each individual.
Table 2.
healthy comparison groups
Longer synchronization lag in stroke individuals compared with three independent
Control Group 1
Control Group 2
HCP
Note. HCP = Human Connectome Project.
Healthy Comparison
0.26 ± 0.12
0.30 ± 0.12
0.37 ± 0.17
Stroke
1.07 ± 0.49
Healthy vs. Stroke
t = 5.5064; p = 1.01e − 05
t = 6.7363; p = 1.12e − 07
t = 12.4558; p = 4.11e − 27
Network Neuroscience
1125
Structural basis for functional lag in chronic stroke
ipsilesional hemisphere:
The brain hemisphere that is
on the same side as a lesion.
We computed an average synchronization lag for the affected and unaffected network by
averaging lags across all pairs of homotopic regions (both lesioned and intact regions) com-
prising the respective networks. Average synchronization lag was significantly increased in the
affected network relative to the unaffected network in the stroke individuals (affected: mean =
1.57 s, unaffected: mean = 0.67 s, p = 0.001, t = 3.658).
We confirmed that this between-network difference remained significant when considering
only intact homotopic pairs belonging to the affected network (p = 0.0216, t = 2.43). This
suggests that lesions may impart downstream effects on unaffected regions within the same
network, resulting in lag increases in these unaffected regions. To establish whether this phe-
nomenon was exclusive to the lesioned brain, we performed a control analysis in which the
average synchronization lag for the unaffected and affected networks (defined using the stroke
data) was computed based on the lag derived from the healthy control group. As expected, no
significant difference was found between the two networks in the healthy comparison individu-
als (p > 0.05). This suggests that lag increases are predominantly circumscribed to lesioned re-
gions or regions comprising the same canonical resting-state networks as the lesioned regions.
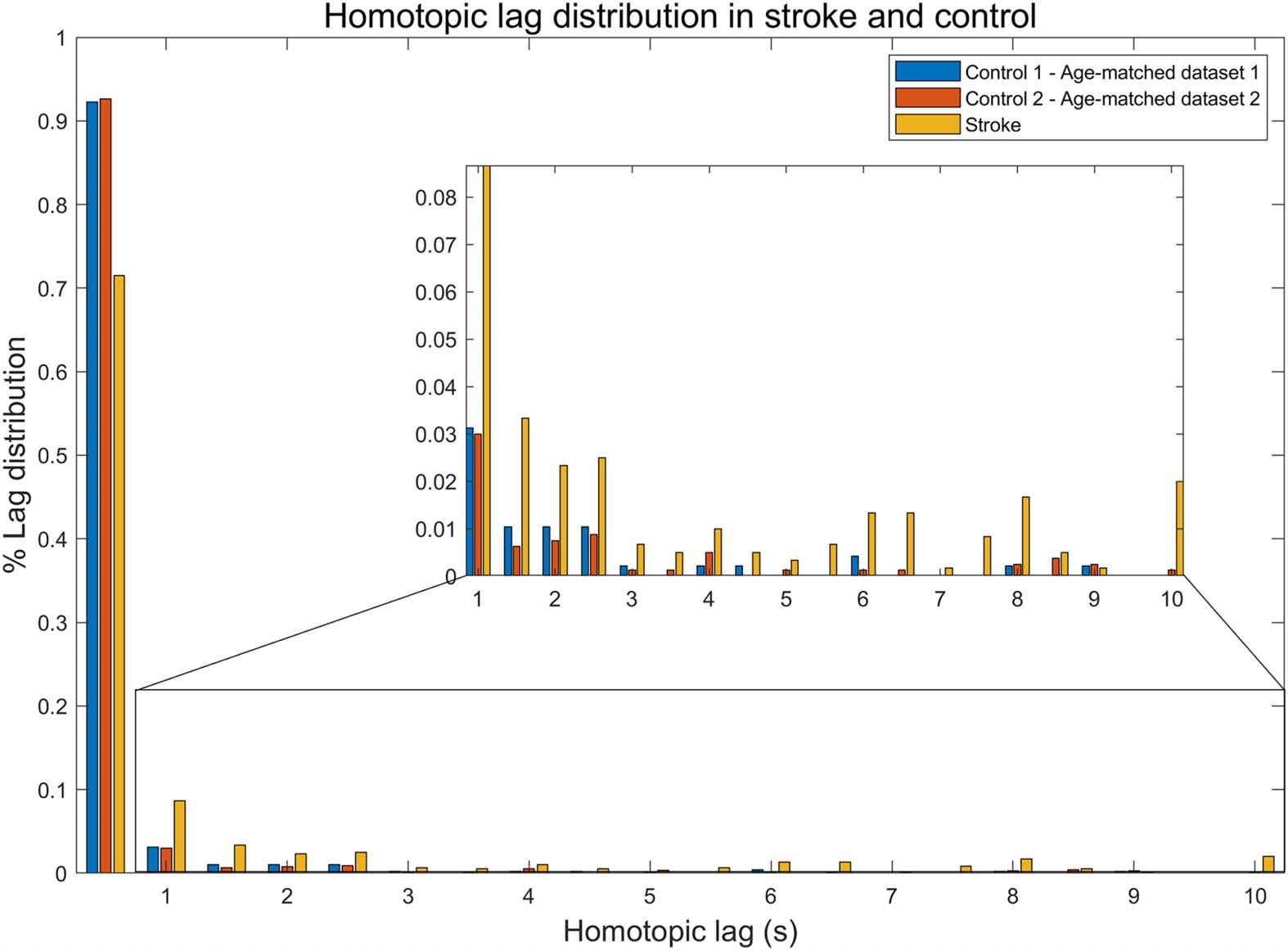
Cifra 2 shows the synchronization lag map and the lesion map of each stroke individuals.
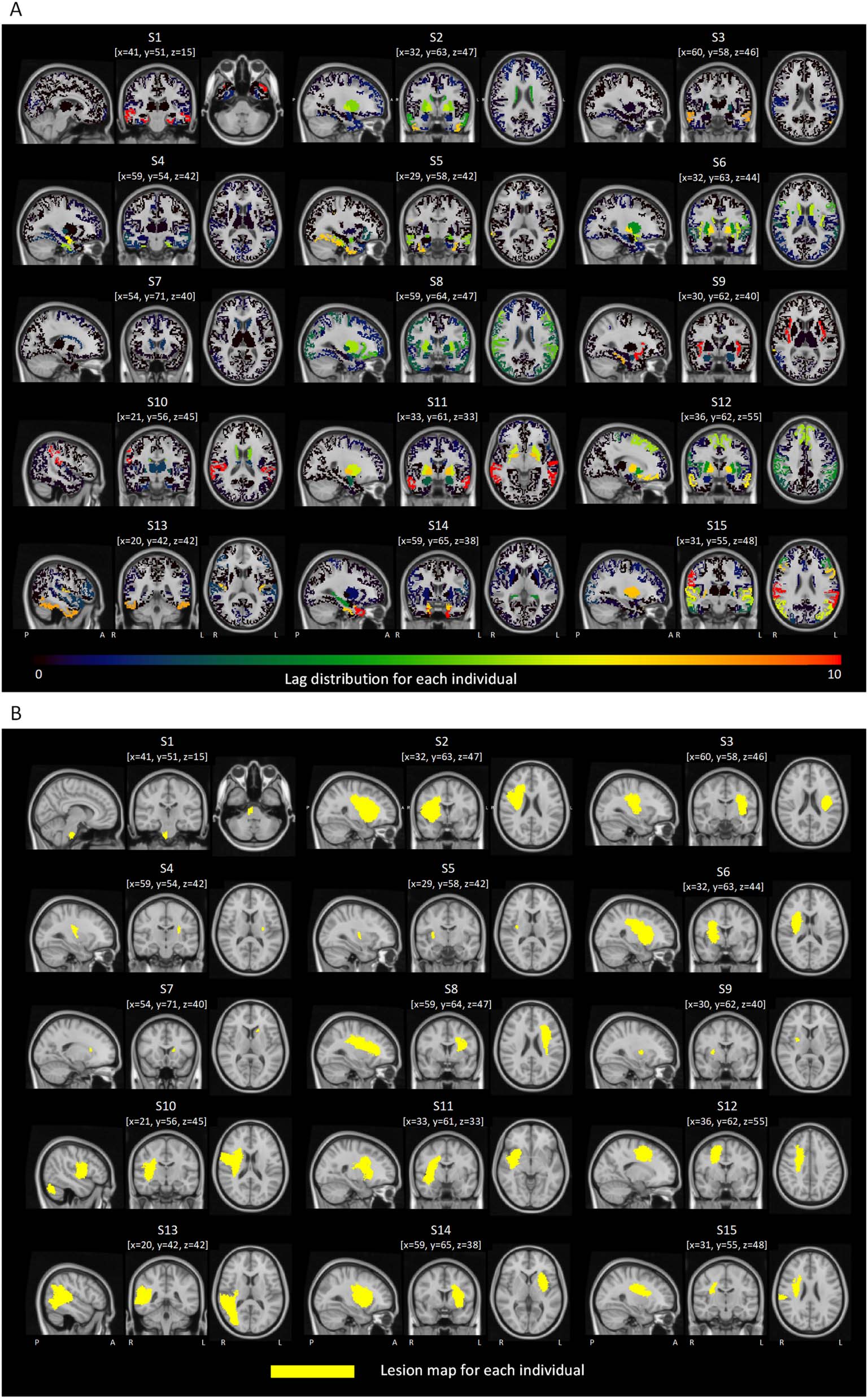
Relation Between Synchronization Lag and Distance to Lesion
Próximo, we investigated whether the lag for each pair of homotopic regions was associated with
the Euclidean distance between the center of mass of the region in the ipsilesional hemi-
sphere and the lesion. Given the presence of multiple distinct lesions in some individuals, el
region-to-lesion distance was computed as the shortest distance between a region and any
lesion. Only the regions in the ipsilesional hemisphere were considered when calculating dis-
tance. For each pair of homotopic regions, lag and the region-to-lesion distances were averaged
across stroke individuals when investigating this relationship. An individual with lesions in the
brainstem was excluded, given that this area was not represented in the regional parcellation
adopted in this study. We found that the distance to the lesion significantly and negatively
correlated with synchronization lag across the 40 homotopic pairs of regions (r = −0.687,
pag < 0.0001; Figure 3). To confirm that this relationship was not driven by certain individuals,
rather than averaging across individuals and then computing the correlation across regions,
we computed an independent correlation across regions for each individual separately. With
this approach, we found that the Euclidean distance to the lesion tended to be correlated
with synchronization lag in 57% of individuals (Supporting Information Table S1). This result
implies that the further a region in the ipsilesional hemisphere is to the lesion, the less the
synchronization lag between corresponding pairs of homotopic regions across hemispheres.
Structural Basis of Synchronization Lag
Communication efficiency explains interindividual variation in lag. We sought to determine a
structural basis for the significant increase in synchronization lag that we found in the stroke
group. To this end, we first evaluated whether the tractography-derived streamline counts
(derived from diffusion MRI data) between homotopic regions related to synchronization lag
(derived from functional MRI data). Lag and streamline counts were averaged across homo-
topic regions for each individual to yield a whole-brain–averaged streamline count and lag for
each individual. We found a negative trend toward an association between streamline count
and lag across the 40 homotopic regions (r = −0.456, p = 0.088; Supporting Information
Figure S2). Although streamline counts do not have an explicit biological correlate, they are
likely to be influenced by axonal myelination, caliber, and end-to-end axonal counts. This
Network Neuroscience
1126
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Maps of lag distribution (A) and lesions (B) for each stroke individual. The same brain
slice is used in the lag and lesion views, where MNI coordinates are denoted with [x,y,z]. In B,
lesions are colored with yellow. Images are shown in MNI152 standard space.
Network Neuroscience
1127
Structural basis for functional lag in chronic stroke
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Scatter plot of the negative association between synchronization lag and the Euclidean
Figure 3.
distance to the lesion. Each data point indicates a homotopic pair of regions. For each pair of ho-
motopic regions, lag was averaged across all stroke individuals and then transformed using a loga-
rithmic remapping. Note that subsecond lags become negative after logarithm transformation. The
distance from each pair of homotopic regions to the lesion was defined as the Euclidean distance
between the center of mass of the region in the ipsilesional hemisphere and the lesion. In the case
of multiple lesions, only the nearest lesion was considered when calculating distance. Distances
were also averaged across stroke individuals. ROI = region of interest.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
suggests that shorter lags tend to be associated with enhanced axonal connectivity between
homotopic regions.
To understand the relation between structural measures and lag in greater detail, graph-
theoretic analysis of two network communication measures was considered. In particular, we
computed network communication efficiency for each stroke individual under two communi-
cation schemes: shortest paths and navigation. Whole-brain communication efficiency under
these two schemes was computed independently for each stroke individual by using their
structural connectome, yielding two efficiency values per individual. For each communica-
tion scheme, lags were averaged across all homotopic regions for each individual to yield a
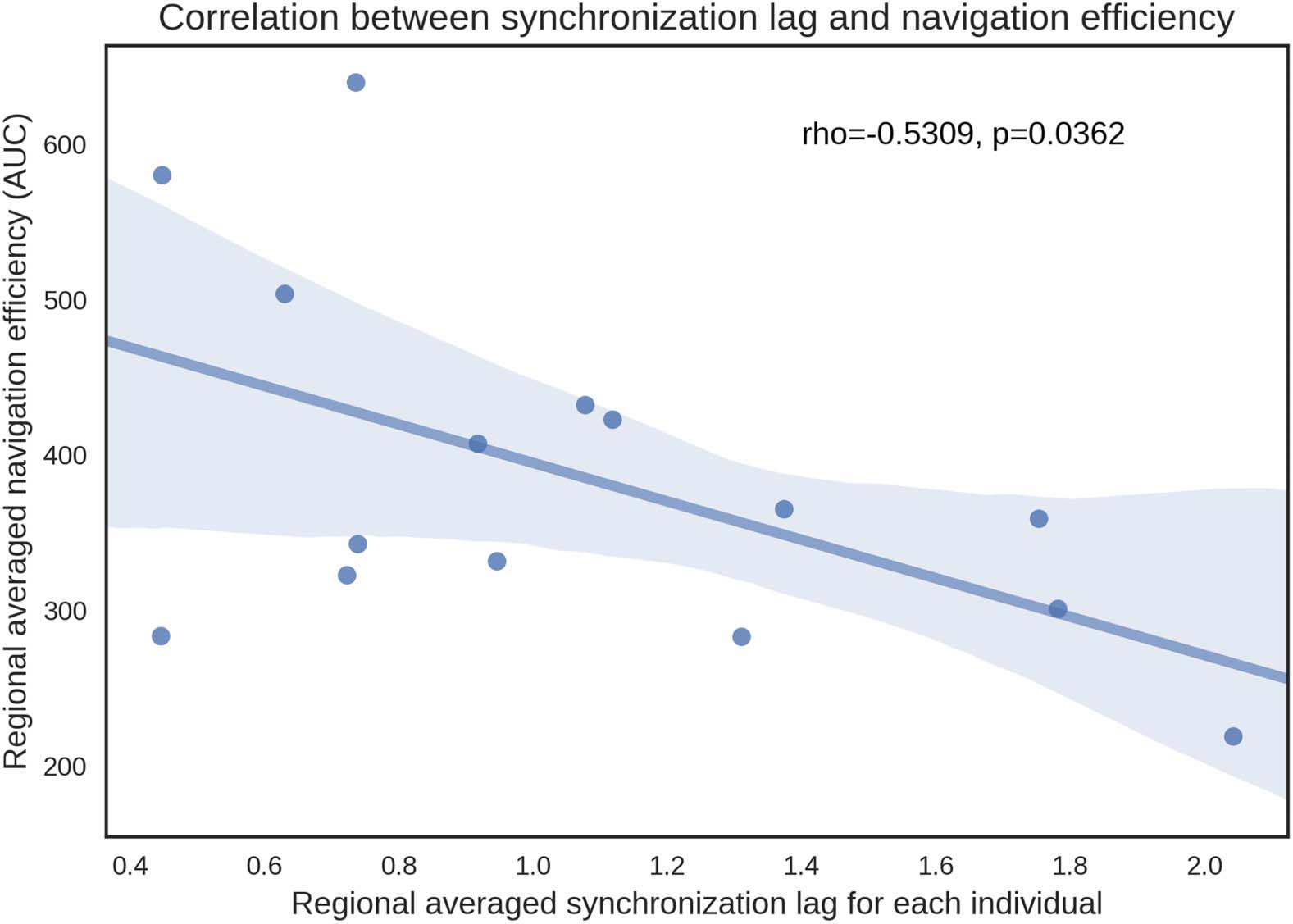
whole-brain–averaged measure of shortest paths and navigation efficiency. Lag was found to
be significantly correlated with both navigation efficiency (r = −0.531, p = 0.0362; Figure 4)
and shortest path efficiency (r = −0.531, p = 0.0415). These associations remained significant
after controlling for the effects of age and gender. To establish whether this correlation was
exclusive to the lesioned brain, we performed the same correlation between efficiency and
lag in healthy controls. No significant correlation was found with either navigation efficiency
(r = −0.093, p = 0.568) or shortest path efficiency (r = −0.093, p = 0.568).
Navigation distance explains interregional variation in lag. We have thus far shown than inter-
individual variation in lag can be explained, to a certain extent, by the efficiency with which
Network Neuroscience
1128
Structural basis for functional lag in chronic stroke
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4.
Scatter plot of the association between whole-brain–averaged synchronization lag and
navigation efficiency. Each data point represents an individual with stroke. Lags were averaged
across all homotopic regions for each individual. Navigation efficiency was summarized using the
area under curve (AUC), computed across structural networks with connection density ranging
between 10% and 50% at increments of 5%.
information can be communicated in each individual’s structural connectome. We next sought
to investigate the structural basis for interregional variance in homotopic lag in the stroke in-
dividuals. Synchronization lag and efficiency for each pair of homotopic regions were aver-
aged across individuals. We found that synchronization lag was negatively correlated with
navigation efficiency across the set of homotopic regions (Spearman rho = -0.3289, p =
0.0388), but not for shortest paths efficiency (Spearman rho = -0.1705, p = 0.2916). This sug-
gests that the increase in lag evident in the stoke individuals may be due to reduced efficiency
of neural signal propagation between homotopic regions.
Following our previous results on the relationship between lag and region-to-lesion dis-
tance, we sought to determine whether homotopic Euclidean distance and synchronization
lag were associated. Indeed, we found a significant correlation between interregional ho-
motopic distances and lag (r = 0.592, p < 0.0001), further suggesting the importance of
Euclidean distance in shaping synchronization delays. However, neural information transfer is
ultimately constrained by underlying white matter pathways, which are not oriented along the
straight lines delineated by the Euclidean distance between brain regions. Navigation distance
combines anatomical (white matter communication pathways) and geometric (influence of
Euclidean distance) constraints into a single information transfer measure (see Methods). In-
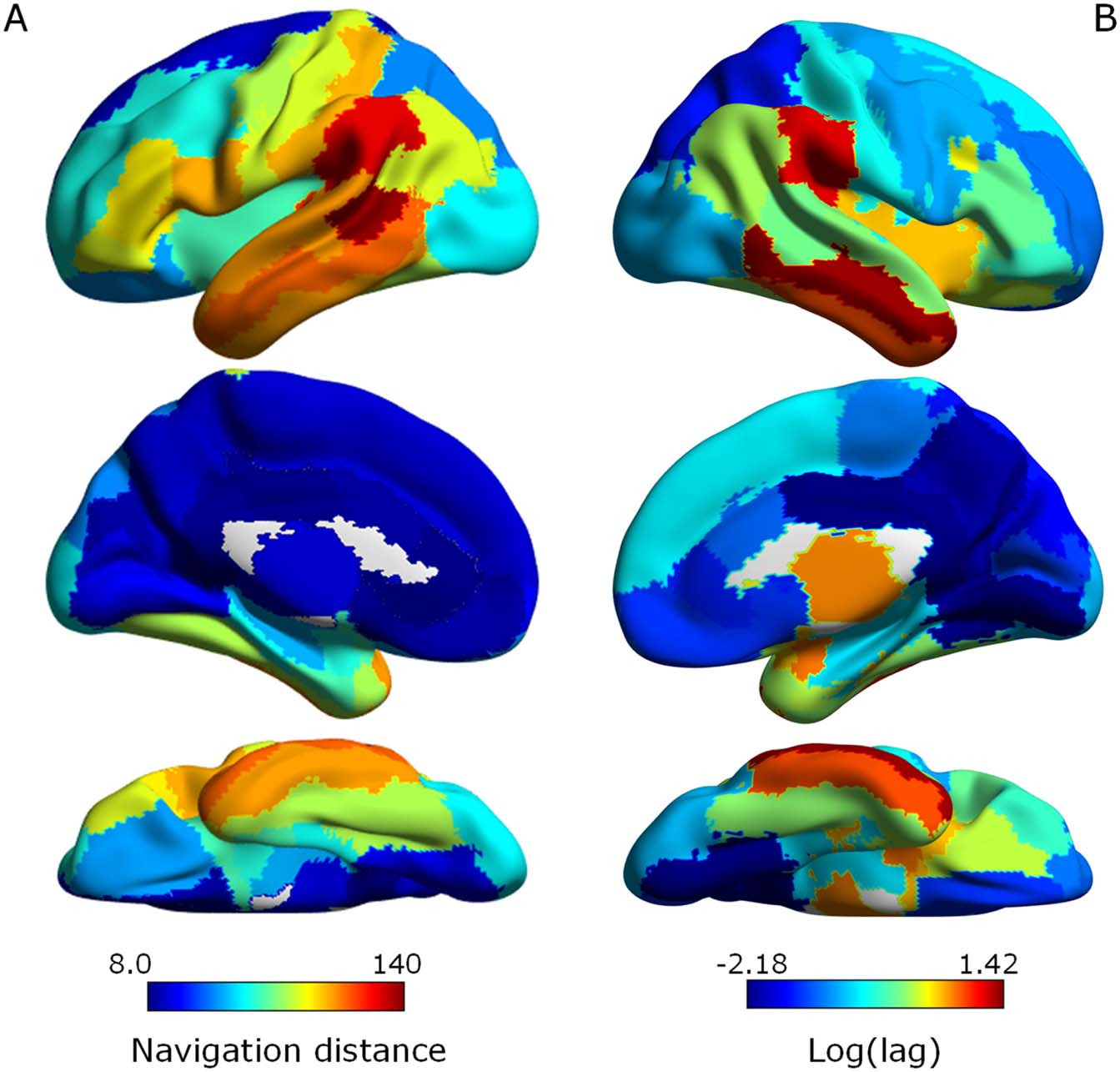
deed, navigation distance was strongly associated with interregional synchronization lag (r =
0.604, p < 0.0001; Figure 5). In addition, while Euclidean distance explained 35.08% of
the variance in lag, combining navigation and Euclidean distances in a general linear model
increased the variance explained to 44.98%. This implies that although Euclidean distance is
navigation distance:
The length of a communication path
between two nodes quantified under
navigation.
Network Neuroscience
1129
Structural basis for functional lag in chronic stroke
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Variation in the efficiency of neural signal propagation between homotopic pairs of
regions in the structural connectome associated with interregional variation in synchronization lag.
(A) Efficiency of neural signal propagation under navigation between homotopic pairs rendered
onto the cortical surface. Efficiency was quantified with the navigation distance. Communication
between regions with longer navigation distances (warm colors) is less efficient. (B) Synchronization
lag rendered onto the cortical surface following logarithmic remapping. Warm colors indicate longer
lags. Note that the two maps tend to have similar distributions.
a significant predictor of lag, synchronization delays are still shaped by underlying network
communication facilitated by the structural connectome.
Next, we tested whether the relationship between synchronization lag and navigation/
Euclidean distances was present in healthy individuals belonging to the age-matched control
group (AMC) and HCP datasets. For both datasets, we found no significant correlation between
homotopic lag and navigation distances (AMC: p = 0.9868; HCP: p = 0.7727), nor between
homotopic lag and Euclidean distance (AMC: p = 0.2515; HCP: p = 0.233). This indicates that
the association between increased lag and decrease communication efficiency is specific to the stroke
cohort, capturing changes in white matter structure and neural signaling resulting from lesions.
Relationship between lag and motor deficit in stroke
Finally, we tested whether interindividual variation in average synchronization lag was asso-
ciated with variation across stroke individuals in motor impairment. ARAT scores quantifying
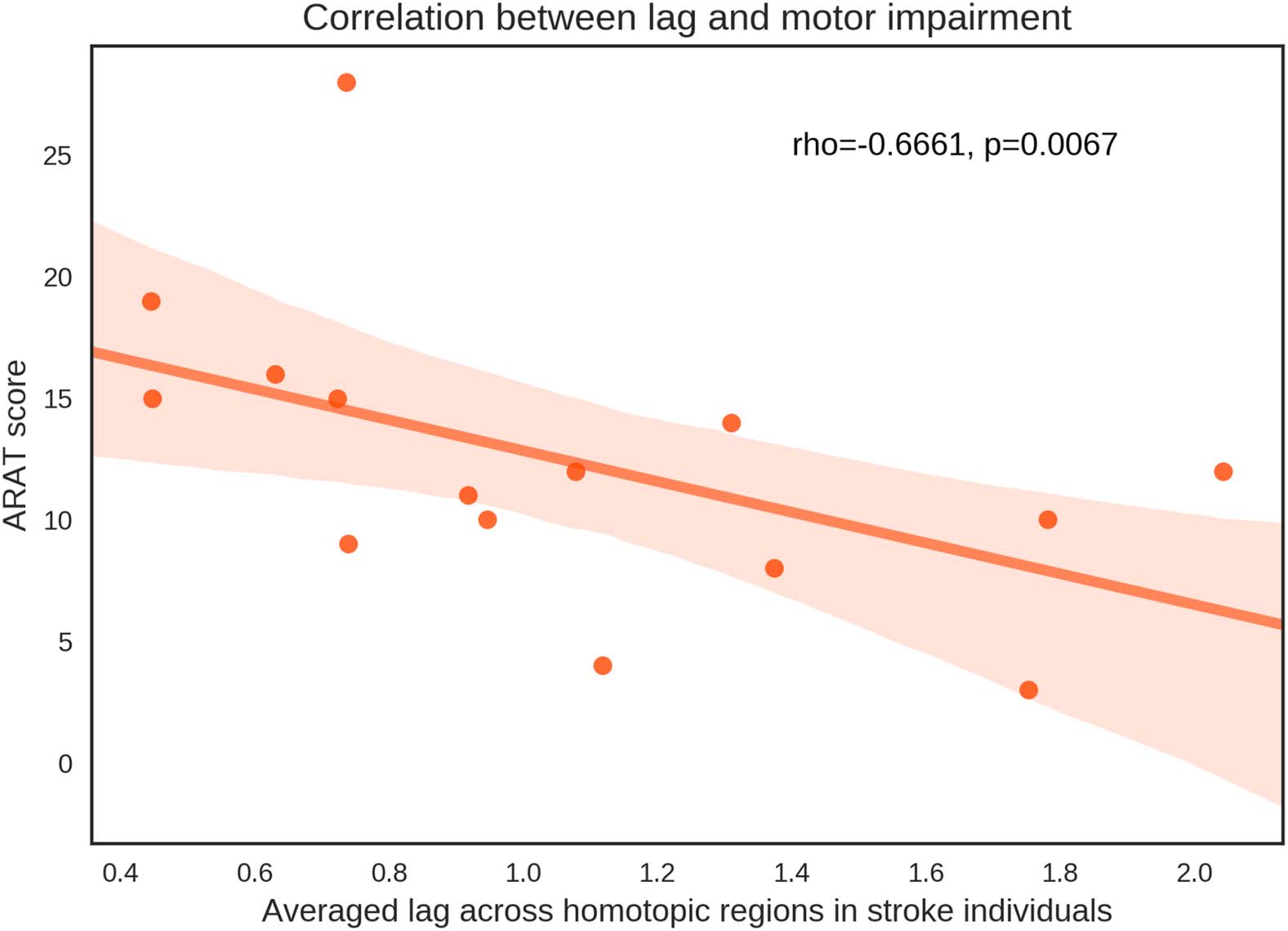
upper-limb motor function were significantly associated with the average lag (r = −0.6661,
p = 0.0067; Figure 6), providing support for the clinical utility of this biomarker. This result
indicates that better upper-limb motor function is associated with shorter lag.
Network Neuroscience
1130
Structural basis for functional lag in chronic stroke
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Scatter plot of the association between whole-brain–averaged synchronization lag and
Figure 6.
motor impairment indicated by Action Research Arm Test (ARAT). Each data point represents an
individual with stroke. Lags were averaged across all homotopic regions for each individual.
We sought to establish whether lag would be a stronger predictor of upper-limb motor
impairment compared with the average functional connectivity strength (Pearson correlation)
between pairs of homotopic regions. In agreement with previous studies (Carter et al., 2010),
although the average functional connectivity was positively correlated with the ARAT scores
(r = 0.5248, p = 0.0447), synchronization lag yielded greater explanatory power and was thus
deemed a more sensitive biomarker.
DISCUSSION
Previous studies suggest that stroke is associated with an increase in synchronization lag be-
tween homotopic regions (Siegel et al., 2017, 2016; Zhao, Lambon Ralph, & Halai, 2018).
In the present study, we replicated this finding and investigated its structural underpinnings
from a graph-theoretic standpoint. We found that both interindividual and interregional vari-
ation in stroke-related increases in synchronization lag between homotopic regions could be
explained in terms of the efficiency of signal propagation in an individual’s structural connec-
tome. Our findings suggest that lesions may obstruct efficient signaling pathways, requiring
recruitment of alternative pathways that traverse longer distances and/or more synapses, result-
ing in increased propagation delays and synchronization lags. In this way, our results provide
a structural basis for the delay of information transfer between homotopic regions in stroke
individuals. Moreover, synchronization lag was found to correlate with interindividual motor
deficits more strongly than functional connectivity, providing support for the clinical utility of
structural and connectomic biomarkers in stroke.
Network Neuroscience
1131
Structural basis for functional lag in chronic stroke
Increased latency in BOLD responses after stroke have been reported in both task-based
(Amemiya, Kunimatsu, Saito, & Ohtomo, 2012; Bonakdarpour, Parrish, & Thompson, 2007;
Roc et al., 2006) and resting-state (Siegel et al., 2017; Zhao et al., 2018) fMRI studies. We
found that synchronization lag was significantly increased in stroke individuals compared
with healthy controls, particularly in canonical functional networks encompassing the lesions.
These results are in line with previous work demonstrating that functional alterations after
stroke extend beyond the area of the lesion (Ovadia-Caro et al., 2013; L. Wang et al., 2010).
Regions in the same functional network communicate and interact as a self-organized coali-
tion to complete similar tasks (Sporns, Chialvo, Kaiser, & Hilgetag, 2004). Once part of the
network is damaged, lesion-induced functional alterations can also be found in anatomically
intact areas that are distant to but connected with the lesion either directly or via a polysynap-
tic path (Carrera & Tononi, 2014; Laganiere, Boes, & Fox, 2016). These functional alterations
may reflect reorganization and/or utilization of redundancy in connectivity to compensate for
the lesion-induced network disruptions (Hurtado et al., 2006). Activations occurring in perile-
sional regions and shifting to other motor areas are commonly observed following stroke with
motor deficit (Frías et al., 2018; Grefkes & Fink, 2011). These newly recruited regions cannot
fully compensate for the impairment of the lesioned area (Ward & Cohen, 2004), prolonging
information transfer and leading to synchronization lag in regions belonging to the same func-
tional network. Accordingly, synchronization lag between homotopic regions increased as a
function of the Euclidean distance between the lesion and ipsilesional region. Regions in close
spatial proximity to the lesion are likely to share the same vascular supply as the lesion area.
Therefore, once a cerebral artery is occluded because of the lesion, vascular supply to sur-
rounding regions may also be comprised. In contrast, regions distant to the lesion are more
likely to be served by alternative vascular networks that are minimally influenced by the lesion.
Linking structural and functional properties of the human connectome is a major goal in
neuroscience (Amico & Goñi, 2018; Diez et al., 2017; Honey et al., 2009; Miši´c et al., 2016;
Seguin, Razi, & Zalesky, 2019). In the present study, we focused on understanding the putative
link between synchronization lag (functional property) and the efficiency of signal propaga-
tion in the connectome (structural property). At both interindividual and interregional levels,
signaling efficiency was negatively associated with synchronization lag in the stroke cohort,
but not in the healthy control group. Following stroke, synchronization lags emerge most
likely because of the obstruction of axonal signaling pathways caused by the lesion and pos-
sibly subsequent recruitment of alternative, less efficient pathways. These pathways could be
redundant pathways performing similar functions, functionally silent pathways, or sprouting of
fibers from the surviving periinfarct neurons (Cramer, 2008; Cramer et al., 2011; Hurtado et al.,
2006; Murphy & Corbett, 2009). Compensatory pathways are not necessarily as efficient in
communicating information as the original pathways disrupted by the lesion (L. Wang et al.,
2010). Our results suggest that graph-theoretic measures of communication efficiency are able
to capture aspects of structure-function reorganization following stroke.
decentralized communication:
A network communication model
that does not mandate global
knowledge of the network topology
to determine communication
paths, contrast to centralized
communication models.
Navigation is a decentralized communicationscheme that has been proposed as a more
biologically plausible model for large-scale neural communication than shortest path routing
(Seguin et al., 2018). At the interregional level, navigation efficiency was significantly associ-
ated with lag, whereas shortest path efficiency was not. This finding indicates that decentralized
models of neural communication may more accurately describe aspects of neural information
processing and neuroplasticity (Avena-Koenigsberger et al., 2017). In addition, the geometric
nature of neural signaling under the navigation model highlights the importance of both spatial
and anatomical constraints to information transfer in the brain.
Network Neuroscience
1132
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
Interestingly, we also found that the Euclidean distance between homotopic regions was a
strong predictor of synchronization lag in stroke. We posit that distant homotopic regions are
more vulnerable to disruption because of their use of longer signaling pathways, resulting in
an increased likelihood of rerouting. Under navigation, communication pathways are identi-
fied based on the distance between regions, while still restricted to the underlying anatomical
connections. Hence, measures of communication under navigation capture the three-way re-
lationship between neural signaling, white matter connectivity, and the spatial positioning of
regions. Indeed, we found that navigation distance showed the strongest association with syn-
chronization lag out of all measures evaluated, and remained significantly correlated to the
lag after controlling for Euclidean distance.
Interhemispheric homotopic functional connectivity is a more accurate predictor of behav-
ioral deficits following stroke than other kinds of functional measures including intrahemi-
spheric connectivity (Carter et al., 2010; Chi et al., 2018; Frías et al., 2018). Consistent with
these studies, we also found a significant correlation between homotopic functional connec-
tivity and post-stroke motor deficits. Interestingly, we found that homotopic lag was more
strongly associated with motor deficits than functional connectivity. Stroke individuals with
better upper-limb motor function tend to have shorter synchronization lag in homotopic re-
gions, possibly suggesting a reorganization of communication between hemispheres facilitat-
ing improved function. This is consistent with other studies, indicating that lesions can change
communication between the cerebral hemispheres (Jones, 2017; Nowak, Grefkes, Ameli, &
Fink, 2009).
Several methodological considerations should be noted. First, image acquisition in the
stroke individuals was performed using a different scanner and acquisition sequence than the
healthy comparison individuals. This may have resulted in systematic bias when comparing
the groups. To assess the impact of this potential bias, we verified our results by using two
independent groups of healthy comparison individuals. Consistent with the primary group of
healthy comparison individuals, the stroke individuals were still found to show significantly
longer synchronization lags when compared with each of these two additional control groups.
Furthermore, synchronization lag did not significantly differ between the healthy comparison
groups, each of which was acquired using a different scanner and acquisition protocol. This
suggests that the estimation of synchronization lag is relatively robust to interscanner differ-
ences. Furthermore, our results replicate previous studies showing a significant difference in
lag between stroke and healthy comparison individuals (Siegel et al., 2017, 2016). Second,
our predominantly male cohort is not representative of the typical sex ratio associated with
stroke incidence. We attempted to recruit a balanced sample initially, but then found that
males participated in the MRI component of the clinical study at higher rates than females,
which resulted in a biased sex ratio. We further note that reanalyzing our data after exclud-
ing the single female participant did not alter the significance of the findings reported in this
study. Heterogeneity of the sample is another consideration in terms of stroke types and the
time since injury. Studies have shown that most functional recovery occurs within the first
3 months after stroke onset (Cramer, 2008; Zeiler & Krakauer, 2013). Given that some of the
chronic stroke individuals studied here were still within the recovery window, the possibility
of further reorganization in functional connectivity is possible, although the pace of recovery
is typically slow (Cramer et al., 2011; Saur et al., 2006) compared with that in the acute
stage. Hence, it is possible that ongoing reorganization of functional connectivity could lead to
further changes in synchronization lag, which would be evident if follow-up scans were to be
acquired. However, synchronization lag was not associated with variation across individuals
in the time elapsed from the stroke event to MRI acquisition in the current analysis.
Network Neuroscience
1133
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
In conclusion, we have proposed and tested a structural explanation for the established
phenomenon of synchronization lag in stroke. In particular, we used advanced graph-theoretic
analyses of network communication schemes on the connectome to evaluate whether the pu-
tative efficiency with which information could be propagated between homotopic regions was
related to synchronization lag. This enabled explanation of synchronization lag based on an
independent modality (diffusion MRI). We found that interindividual variation in synchroniza-
tion lag could be explained to a significant extent by network communication efficiency in the
post-stroke connectome. Furthermore, interregional variation in lag was associated with nav-
igation distance, which is dependent on both Euclidean distance and structural constraints.
Finally, the correlation between synchronization lag and motor deficits provided support for
the clinical utility of this biomarker in stroke recovery.
METHODS
Participants
Fifteen individuals with chronic stroke (14 males and 1 female; mean age: 54 ± 8 years) were
selected from an ongoing clinical motor training trial, which is described in detail elsewhere
(NCT03286309). All individuals who participated in this trial and completed an MRI scan were
included. All individuals suffered from a single cerebrovascular accident prior to recruitment.
The inclusion criteria were (a) sufficient cognition to follow experimental instructions, deter-
mined by a Mini-Mental State Examination (MMSE) score exceeding 21, and (b) hemiparesis
resulting from a single unilateral brain lesion with stroke onset more than 6 months before
recruitment. Exclusion criteria were (a) history of alcohol, drug abuse, or epilepsy, and (b) bi-
lateral infracts, uncontrolled medical problems, and serious cognitive deficits. Motor deficits
were the primary stroke-related symptom in all individuals. Compared with previous studies
of synchronization lag in stroke (Siegel et al., 2017, 2016), our cohort is relatively homoge-
neous with respect to clinical symptoms, in that it is constrained to individuals with motor
deficits. Paretic upper-limb motor functions were assessed by trained clinical assessors who
were blinded to the experiment. Action Research Arm Test (ARAT) was used to evaluate the
paretic upper-limb motor function. Twelve right-handed and age-matched healthy comparison
individuals (6 males and 6 females; mean age: 58 ± 3 years) without any cerebral abnormali-
ties were included to serve as a control group. The control group was sourced from the open
database of Institute of Psychology, Chinese Academy of Sciences (G.-X. Wei, Dong, Yang,
Luo, & Zuo, 2014). All individuals gave informed consent of the study protocol. The study was
approved by the Joint Chinese University of Hong Kong-New Territories East Cluster Clinical
Research Ethics Committee.
Image Acquisition
Stroke individuals were scanned with a 3T Philips MR scanner (Achieva TX, Philips Medical
System, Best, the Netherlands) with an 8-channel head coil. The imaging datasets of healthy
controls were acquired from an open database (https://doi.org/10.15387/fcp_indi.corr.ipcas8),
where scanning was performed with 3T MRI scanner (Siemens Trio Tim, Erlangen, Germany)
with a 12-channel head coil. The following imaging data were acquired in both groups: (a)
high-resolution T1-weighted anatomical images, (b) BOLD fMRI images, and (c) diffusion-
weighted images. The detailed acquisition parameters can be found in the Supporting
Information. The resting-state fMRI acquisition lasted for 8 minutes resulting in 240 brain
volumes for both stroke and control scans.
Network Neuroscience
1134
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
Given that the healthy comparison individuals were scanned on a different scanner to the
stroke individuals, we tested for potential interscanner differences in synchronization lag esti-
mation. To this end, two alternative groups of healthy comparison individuals were analyzed:
(a) 20 age-matched control subjects from an open dataset (D. Wei et al., 2018) and (b) 200
healthy individuals from the HCP (Van Essen et al., 2013). The detailed acquisition parameters
can be found elsewhere (Glasser et al., 2013; Seguin et al., 2018) and in the Supporting Infor-
mation. In total, we therefore evaluated synchronization lags in three independent groups of
healthy comparison individuals. Each of the three groups was acquired using different scanners
and acquisition protocols.
Data Preprocessing and Analysis
The fMRI data were preprocessed using Analysis of Func-
Functional MRI data preprocessing.
tional NeuroImages (AFNI) software (http://afni.nimh.nih.gov/afni). The analysis steps fol-
lowed the recommended analysis procedures for rs-fMRI data (Jo et al., 2013; Jo, Saad, Simmons,
Milbury, & Cox, 2010). The first 10 volumes of each individual’s fMRI data were removed. After
preprocessing, our procedures included despiking of large transients, slice-timing correction,
and motion correction with six-parameter rigid body transformation. Volumes were spatially
filtered by a 4-mm full-width-at-half-maximum isotropic Gaussian kernel. T1 images were seg-
mented into gray matter, white matter, and CSF by using FreeSurfer (Fischl et al., 2002) for each
individual to create corresponding masks. The time series from lateral ventricles and white mat-
ter were then derived for nuisance regression. Other nuisances included motion parameters
and motion parameter time derivatives. Volumes with excessive motion were censored if the
Euclidean norm of the derivatives of the motion parameters exceeded 0.2 by using the function
regress_censor_motion in AFNI (X. Wang, Wong, Sun, Chu, & Tong, 2018). Bandpass filtering
(0.009–0.08 Hz) was also applied. Global signal regression was not used in the processing.
Diffusion data processing and fiber tractography. Diffusion-weighted images were first prepro-
cessed using the FMRIB Software Library, including correction for eddy currents and brain
extraction. Structural networks (connectomes) were then mapped for each individual using
MRtrix3 software (http://www.mrtrix.org/). White matter masks for fiber tracking were derived
from FreeSurfer-based structural segmentations. Diffusion tensors were estimated from the pre-
processed diffusion images using an iteratively reweighted linear least squares estimator. To
avoid the potential gaps between gray and white matter due to registration error, the white
matter boundaries were dilated by 1 voxel. Streamlines were uniformly seeded from the white
matter mask and propagated according to the principal orientation of the estimated diffusion
tensor. Propagation was terminated when streamlines exited the white matter mask into gray
matter or reached a voxel with low fractional anisotropy (FA) (Seguin et al., 2018). Determin-
istic diffusion fiber tracking was performed using the FACT tracking algorithm with 5 × 106
streamlines, 0.5-mm tracking step-size, 400-mm maximum streamline length, and 0.1 FA cutoff
for termination of tracks. We used deterministic tracking rather than probabilistic to conform to
recent connectome mapping recommendations (Maier-Hein et al., 2017; Sarwar et al., 2019).
The connectivity strength between a pair of gray matter regions was defined as the total number
of streamlines with endpoints residing in both regions. For a given parcellation comprising N
gray matter regions, this resulted in an N × N weighted connectivity matrix for each individual.
Measurement of Synchronization Lag
Cortical gray matter was parcellated into N = 80 regions of interest (40 regions per hemisphere)
based on the Desikan/Killiany atlas (Desikan et al., 2006). In keeping with previous studies
Network Neuroscience
1135
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
(Siegel et al., 2017), synchronization lag was only quantified between pairs of homotopic
regions, primarily because functional connectivity was greatest between homotopic regions
in the healthy comparison individuals, thus enabling accurate estimation of cross-correlations.
Furthermore, interhemispheric functional connectivity is typically investigated in stroke since it shows
a relationship between ipsilesional and contralesional hemisphere and has previously been
associated with deficits after stroke (Carter et al., 2010). The synchronization lag in the hemo-
dynamic response between a pair of homotopic regions was derived from the cross-correlation
in regionally-averaged BOLD time series signals (Mitra et al., 2014; Siegel et al., 2016):
Ci,j(τ) = 1
nτ
∑
t
si(t) · sj(t + τ)
(1)
where si and sj are the de-meaned and standard deviation normalized BOLD time series signals
of homotopic regions i and j. Cross-correlation was computed separately for temporal shifts (τ)
between −10 and 10 seconds, corresponding to shifts in TR between −5 and 5. In the above
formula, nτ denotes the number of time points comprising the signal with temporal shift of τ.
The synchronization lag for regions i and j was then defined as,
τ
ij = arg max
τ∈[−10,10]
(cid:2)
(cid:2)
Cij(τ)
(cid:2)
(cid:2)
(2)
In words, the synchronization lag is the temporal shift (lag), denoted with τ, yielding maxi-
mal cross-correlation in BOLD activity. Given the relatively coarse temporal resolution of the
BOLD signal, parabolic interpolation between consecutive time points was used to determine
the exact lag value resulting in maximal cross-correlation. Therefore, each pair of homotopic
regions for each individual was associated with a synchronization lag. In some analyses, lag
was averaged across all homotopic regions to yield an average synchronization lag for each
individual.
Network Communication Measures
We computed network communication efficiency under two communication schemes: short-
est path and navigation. Let W denote a weighted connectivity matrix, where Wij is the
connection weight (streamline count) between nodes i and j. We computed the connection
length matrix L = −log10(W/max(W) + 1), ensuring a monotonic remapping of connec-
tion weights into connection lengths that allows for the computation of communication path
lengths (Goñi et al., 2014; Seguin et al., 2018). Path lengths were defined as Λ
ij = Liu +
· · · + Lvj, where {u . . . v} is the sequence of nodes comprised along the shortest or navigation
paths from node i to node j. The shortest path length Λsp
is the globally minimum communi-
ij
cation cost between nodes i and j. In contrast, navigation path length Λnav
expresses the cost
of communication paths identified using local knowledge of network topology and the spatial
positioning of nodes. Navigation paths from node i to j were identified by progressing to the
neighbor of i closest in Euclidean distance to j. This process was repeated for each new node
until j is reached (successful navigation) or a node is revisited (failed navigation, resulting in
Λnav
ij
ij = ∞).
The communication cost of navigation can be alternatively defined in terms of the Euclidean
distance traversed along navigation paths. We refer to this measure as navigation distance and
ij = diu + · · · + dvj, where dij is the Euclidean distance between noedes i and
define it as Dnav
j, and {u,…,v} is the sequence of node along the navigation path.
Network Neuroscience
1136
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
The network efficiency of the brain was computed based on corresponding path length:
E =
−1
∑i(cid:3)=j (Λ
ij)
N(N−1)
(3)
connection density:
The proportion of actual connected
edges among all possible
connections in a network.
where N was the number of regions in brain parcellation. All the graph-theoretical analy-
ses were computed using the Brain Connectivity Toolbox (https://sites.google.com/site/bctnet/)
(Rubinov & Sporns, 2010).
Statistics
Statistical inference was conducted using MATLAB (The MathWorks). Two-sample, two-sided
t tests were used to test the null hypothesis of equality in lag between the stroke and healthy
comparison group. Each of the 40 pairs of homotopic regions was tested independently as
well. To evaluate the impact of structural communication measures on synchronization lag, the
Pearson correlation coefficient was computed between mean synchronization lag and mea-
sures of the structural network. Network efficiency was summarized using the AUC, computed
across structural networks with connection density ranging between 10% and 50% at 5% in-
tervals. For interregional analysis, all the lag values were transformed using logarithm given
that the lag distribution in homotopic regions was not normally distributed. The relationship
between lag and stroke motor function was tested with the Spearman correlation coefficient.
The statistical significance level was set at p < 0.05. All p values were corrected using the false
discovery rate.
ACKNOWLEDGMENTS
We thank the Research Grants Council of the Hong Kong Special Administrative Region and
CUHK Global Scholarship Programme for Research Excellence.
SUPPORTING INFORMATION
Supporting information for this article is available at https://www.doi.org/10.1162/netn_a_00105.
AUTHOR CONTRIBUTIONS
Xin Wang: Data curation; Formal analysis; Methodology; Validation; Visualization; Writing
– Original Draft; Writing – Review & Editing. Caio Seguin: Data curation; Formal analysis;
Methodology; Writing – Original Draft; Writing – Review & Editing. Andrew Zalesky: Con-
ceptualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodol-
ogy; Supervision; Writing - Original Draft; Writing – Review & Editing. Wan-wa Wong: Data
curation; Formal analysis; Methodology; Writing – Original Draft; Writing – Review &
Editing. Winnie Chiu-wing Chu: Conceptualization; Project administration; Software;
Writing – Original Draft. Raymond Kai-yu Tong: Conceptualization; Funding acquisition;
Project administration; Supervision; Writing - Original Draft.
FUNDING INFORMATION
Raymond Kai-yu Tong, Research Grants Council of the Hong Kong Special Administrative Re-
gion, Award ID: GRF-CUHK 14207617. Xin Wang, CUHK Global Scholarship Programme
for Research Excellence. Andrew Zalesky, Australian National Health and Medical Research
Council (NHMRC) Senior Research Fellowship B, Award ID: 1136649. Caio Seguin, Melbourne
Research Scholarship.
Network Neuroscience
1137
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
REFERENCES
Allard, A., & Serrano, M. A. (2018). Navigable maps of struc-
tural brain networks across species. ArXiv:1801.06079 [Physics,
q-Bio]. Retrieved from http://arxiv.org/abs/1801.06079
Amemiya, S., Kunimatsu, A., Saito, N., & Ohtomo, K. (2012). Im-
paired hemodynamic response in the ischemic brain assessed
with BOLD fMRI. NeuroImage, 61(3), 579–590. https://doi.org/
10.1016/j.neuroimage.2012.04.001
Amemiya, S., Kunimatsu, A., Saito, N., & Ohtomo, K. (2014). Cere-
bral hemodynamic impairment: assessment with resting-state
functional MR imaging. Radiology, 270(2), 548–555. https://doi.
org/10.1148/radiol.13130982
Amico, E., & Goñi, J. (2018). Mapping hybrid functional-structural
connectivity traits in the human connectome. Network Neuro-
science, 2(3), 306–322. https://doi.org/10.1162/netn_a_00049
Avena-Koenigsberger, A., Miši´c, B., & Sporns, O. (2017). Commu-
nication dynamics in complex brain networks. Nature Reviews
Neuroscience, 19, 17.
Boguñá, M., Krioukov, D., & Claffy, K. C. (2009). Navigability of
complex networks. Nature Physics, 5(1), 74–80. https://doi.org/
10.1038/nphys1130
Bonakdarpour, B., Parrish, T. B., & Thompson, C. K.
(2007).
Hemodynamic response function in patients with stroke-induced
aphasia: implications for fMRI data analysis. NeuroImage, 36(2),
322–331. https://doi.org/10.1016/j.neuroimage.2007.02.035
Buchli, A. D., & Schwab, M. E. (2005). Inhibition of Nogo: a key
strategy to increase regeneration, plasticity and functional recov-
ery of the lesioned central nervous system. Annals of Medicine,
37(8), 556–567. https://doi.org/10.1080/07853890500407520
Bullmore, E., & Sporns, O. (2009). Complex brain networks: graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10(3), 186–198. https://doi.org/10.1038/
nrn2575
Caliandro, P., Vecchio, F., Miraglia, F., Reale, G., Della Marca, G.,
La Torre, G., . . . Rossini, P. M. (2017). Small-world characteristics
of cortical connectivity changes in acute stroke. Neurorehabili-
tation and Neural Repair, 31(1), 81–94. https://doi.org/10.1177/
1545968316662525
Carrera, E., & Tononi, G. (2014). Diaschisis: past, present, future.
Brain: A Journal of Neurology, 137(Pt 9), 2408–2422. https://doi.
org/10.1093/brain/awu101
J.,
Carter, A. R., Astafiev, S. V., Lang, C. E., Connor, L. T., Rengachary, J.,
Strube, M.
(2010). Resting interhemi-
spheric functional magnetic resonance imaging connectivity
predicts performance after stroke. Annals of Neurology, 67(3),
365–375. https://doi.org/10.1002/ana.21905
. Corbetta, M.
.
.
Carter, A. R., Shulman, G. L., & Corbetta, M. (2012). Why use
a connectivity-based approach to study stroke and recovery of
function?. NeuroImage, 62(4), 2271–2280. https://doi.org/10.
1016/j.neuroimage.2012.02.070
Chi, N.-F., Ku, H.-L., Chen, D. Y.-T., Tseng, Y.-C., Chen, C.-J.,
Lin, Y.-C., . . . Hu, C.-J. (2018). Cerebral motor functional con-
nectivity at the acute stage: an outcome predictor of ischemic
stroke. Scientific Reports, 8(1). https://doi.org/10.1038/s41598-
018-35192-y
Cramer, S. C.
(2008). Repairing the human brain after stroke:
I. Mechanisms of spontaneous recovery. Annals of Neurology,
63(3), 272–287. https://doi.org/10.1002/ana.21393
Cramer, S. C., Sur, M., Dobkin, B. H., O’Brien, C., Sanger, T. D.,
Trojanowski, J. Q., . . . Vinogradov, S. (2011). Harnessing neuro-
plasticity for clinical applications. Brain: A Journal of Neurology,
134(Pt 6), 1591–1609. https://doi.org/10.1093/brain/awr039
Crofts, J. J., Higham, D. J., Bosnell, R., Jbabdi, S., Matthews, P. M.,
Behrens, T. E. J., & Johansen-Berg, H. (2011). Network analy-
sis detects changes in the contralesional hemisphere following
stroke. NeuroImage, 54(1), 161–169. https://doi.org/10.1016/j.
neuroimage.2010.08.032
Dancause, N., Barbay, S., Frost, S. B., Plautz, E. J., Chen, D.,
Zoubina, E. V., . . . Nudo, R. J. (2005). Extensive cortical rewiring
after brain injury. Journal of Neuroscience, 25(44), 10167–10179.
https://doi.org/10.1523/JNEUROSCI.3256-05.2005
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson, B. C.,
Blacker, D., . . . Killiany, R. J. (2006). An automated labeling
system for subdividing the human cerebral cortex on MRI scans
into gyral based regions of interest. NeuroImage, 31(3), 968–980.
https://doi.org/10.1016/j.neuroimage.2006.01.021
Diez, I., Drijkoningen, D., Stramaglia, S., Bonifazi, P., Marinazzo, D.,
Gooijers, J., . . . Cortes, J. M. (2017). Enhanced prefrontal functional–
structural networks to support postural control deficits after
traumatic brain injury in a pediatric population. Network Neuro-
science, 1(2), 116–142. https://doi.org/10.1162/NETN_a_00007
Dimyan, M. A., & Cohen, L. G. (2011). Neuroplasticity in the con-
text of motor rehabilitation after stroke. Nature Reviews Neurol-
ogy, 7(2), 76–85. https://doi.org/10.1038/nrneurol.2010.200
Fischl, B., Salat, D. H., Busa, E., Albert, M., Dieterich, M.,
Haselgrove, C., . . . Dale, A. M. (2002). Whole brain segmen-
tation: automated labeling of neuroanatomical structures in the
human brain. Neuron, 33(3), 341–355.
Fornito, A., Zalesky, A., & Bullmore, E. T. (2016). Fundamentals of
brain network analysis. Boston: Elsevier/Academic Press.
Frías, I., Starrs, F., Gisiger, T., Minuk, J., Thiel, A., & Paquette, C.
(2018). Interhemispheric connectivity of primary sensory cor-
tex is associated with motor impairment after stroke. Scientific
Reports, 8(1). https://doi.org/10.1038/s41598-018-29751-6
Glasser, M. F., Sotiropoulos, S. N., Wilson, J. A., Coalson, T. S.,
Fischl, B., Andersson, J. L., . . . WU-Minn HCP Consortium
(2013). The minimal preprocessing pipelines for the Human Con-
nectome Project. NeuroImage, 80, 105–124. https://doi.org/10.
1016/j.neuroimage.2013.04.127
Goñi, J., van den Heuvel, M. P., Avena-Koenigsberger, A., Velez de
Mendizabal, N., Betzel, R. F., Griffa, A., . . . Sporns, O. (2014).
Resting-brain functional connectivity predicted by analytic mea-
sures of network communication. Proceedings of the National
Academy of Sciences of the United States of America, 111(2),
833–838. https://doi.org/10.1073/pnas.1315529111
Grefkes, C., & Fink, G. R. (2011). Reorganization of cerebral net-
works after stroke: new insights from neuroimaging with con-
nectivity approaches. Brain: A Journal of Neurology, 134(Pt 5),
1264–1276. https://doi.org/10.1093/brain/awr033
Network Neuroscience
1138
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
Honey, C. J., Sporns, O., Cammoun, L., Gigandet, X., Thiran, J. P.,
Meuli, R., & Hagmann, P. (2009). Predicting human resting-
state functional connectivity from structural connectivity. Pro-
ceedings of the National Academy of Sciences of the United
States of America, 106(6), 2035–2040. https://doi.org/10.1073/
pnas.0811168106
Hurtado, O., Pradillo, J. M., Alonso-Escolano, D., Lorenzo, P.,
Sobrino, T., Castillo, J., . . . Moro, M. A. (2006). Neurore-
pair versus neuroprotection in stroke. Cerebrovascular Diseases
(Basel, Switzerland), 21 Suppl 2, 54–63. https://doi.org/10.1159/
000091704
Jo, H. J., Gotts, S. J., Reynolds, R. C., Bandettini, P. A., Martin, A.,
Cox, R. W., & Saad, Z. S. (2013). Effective preprocessing proce-
dures virtually eliminate distance-dependent motion artifacts in
resting state FMRI. Journal of Applied Mathematics, 2013, 1–9.
https://doi.org/10.1155/2013/935154
Jo, H. J., Saad, Z. S., Simmons, W. K., Milbury, L. A., & Cox, R. W.
(2010). Mapping sources of correlation in resting state fMRI, with
artifact detection and removal. NeuroImage, 52(2), 571–582.
https://doi.org/10.1016/j.neuroimage.2010.04.246
Jones, T. A. (2017). Motor compensation and its effects on neural
reorganization after stroke. Nature Reviews Neuroscience, 18(5),
267–280. https://doi.org/10.1038/nrn.2017.26
Kleinberg, J. M. (2000). Navigation in a small world. Nature, 406, 845.
Kolb, B., & Muhammad, A. (2014). Harnessing the power of neuro-
plasticity for intervention. Frontiers in Human Neuroscience, 8,
377. https://doi.org/10.3389/fnhum.2014.00377
Laganiere, S., Boes, A. D., & Fox, M. D. (2016). Network local-
ization of hemichorea-hemiballismus. Neurology, 86(23), 2187–
2195. https://doi.org/10.1212/WNL.0000000000002741
Laney, J., Adali, T., McCombe Waller, S., & Westlake, K. P. (2015).
Quantifying motor recovery after stroke using independent vec-
tor analysis and graph-theoretical analysis. NeuroImage Clinical,
8, 298–304. https://doi.org/10.1016/j.nicl.2015.04.014
Lotze, M., Markert, J., Sauseng, P., Hoppe, J., Plewnia, C., & Gerloff,
C. (2006). The role of multiple contralesional motor areas for
complex hand movements after internal capsular lesion. Journal
of Neuroscience, 26(22), 6096–6102. https://doi.org/10.1523/
JNEUROSCI.4564-05.2006
Lv, Y., Margulies, D. S., Cameron Craddock, R., Long, X., Winter, B.,
Gierhake, D., . . . Villringer, A. (2013). Identifying the perfusion
deficit in acute stroke with resting-state functional magnetic res-
onance imaging. Annals of Neurology, 73(1), 136–140. https://
doi.org/10.1002/ana.23763
Maier-Hein, K. H., Neher, P. F., Houde,
J.-C., Côté, M.-A.,
Garyfallidis, E., Zhong, J., . . . Descoteaux, M. (2017). The chal-
lenge of mapping the human connectome based on diffusion
tractography. Nature Communications, 8(1). https://doi.org/10.
1038/s41467-017-01285-x
Miši´c, B., Betzel, R. F., de Reus, M. A., van den Heuvel, M. P.,
Berman, M. G., McIntosh, A. R., & Sporns, O. (2016). Network-
level structure-function relationships in human neocortex. Cere-
bral Cortex, 26(7), 3285–3296. https://doi.org/10.1093/cercor/
bhw089
Mitra, A., Kraft, A., Wright, P., Acland, B., Snyder, A. Z., Rosenthal,
Z., . . . Raichle, M. E. (2018). Spontaneous infra-slow brain activ-
ity has unique spatiotemporal dynamics and laminar structure.
Neuron, 98(2), 297–305.e6. https://doi.org/10.1016/j.neuron.
2018.03.015
Mitra, A., Snyder, A. Z., Blazey, T., & Raichle, M. E.
(2015).
Lag threads organize the brain’s intrinsic activity. Proceedings
of the National Academy of Sciences of the United States of
America, 112(17), E2235–E2244. https://doi.org/10.1073/pnas.
1503960112
Mitra, A., Snyder, A. Z., Hacker, C. D., & Raichle, M. E. (2014).
Lag structure in resting-state fMRI. Journal of Neurophysiology,
111(11), 2374–2391. https://doi.org/10.1152/jn.00804.2013
Mori, S., Crain, B. J., Chacko, V. P., & van Zijl, P. C. (1999). Three-
dimensional tracking of axonal projections in the brain by mag-
netic resonance imaging. Annals of Neurology, 45(2), 265–269.
Murphy, T. H., & Corbett, D. (2009). Plasticity during stroke recov-
ery: from synapse to behaviour. Nature Reviews Neuroscience,
10(12), 861–872. https://doi.org/10.1038/nrn2735
Nowak, D. A., Grefkes, C., Ameli, M., & Fink, G. R. (2009). Inter-
hemispheric competition after stroke: brain stimulation to enhance
recovery of function of the affected hand. Neurorehabilitation
and Neural Repair, 23(7), 641–656. https://doi.org/10.1177/
1545968309336661
Ovadia-Caro, S., Villringer, K., Fiebach, J., Jungehulsing, G. J.,
van der Meer, E., Margulies, D. S., & Villringer, A. (2013). Lon-
gitudinal effects of lesions on functional networks after stroke.
Journal of Cerebral Blood Flow and Metabolism, 33(8), 1279–1285.
https://doi.org/10.1038/jcbfm.2013.80
Roc, A. C., Wang, J., Ances, B. M., Liebeskind, D. S., Kasner,
S. E., & Detre, J. A. (2006). Altered hemodynamics and regional
cerebral blood flow in patients with hemodynamically signifi-
cant stenoses. Stroke, 37(2), 382–387. https://doi.org/10.1161/
01.STR.0000198807.31299.43
Rubinov, M., & Sporns, O. (2010). Complex network measures
of brain connectivity: uses and interpretations. NeuroImage,
52(3), 1059–1069. https://doi.org/10.1016/j.neuroimage.2009.
10.003
Sarwar, T., Ramamohanarao, K., & Zalesky, A. (2019). Mapping con-
nectomes with diffusion MRI: deterministic or probabilistic trac-
tography? Magnetic Resonance in Medicine, 81(2), 1368–1384.
https://doi.org/10.1002/mrm.27471
Saur, D., Lange, R., Baumgaertner, A., Schraknepper, V., Willmes,
K., Rijntjes, M., & Weiller, C. (2006). Dynamics of language reor-
ganization after stroke. Brain: A Journal of Neurology, 129(Pt 6),
1371–1384. https://doi.org/10.1093/brain/awl090
Schirner, M., McIntosh, A. R., Jirsa, V., Deco, G., & Ritter, P.
(2018). Inferring multi-scale neural mechanisms with brain net-
work modelling. ELife, 7. https://doi.org/10.7554/eLife.28927
Seguin, C., Razi, A., & Zalesky, A. (2019). Inferring neural signalling
directionality from undirected structural connectomes. BioRxiv.
https://doi.org/10.1101/573071
Seguin, C., van den Heuvel, M. P., & Zalesky, A. (2018). Naviga-
tion of brain networks. Proceedings of the National Academy of
Sciences of the United States of America, 115(24), 6297–6302.
https://doi.org/10.1073/pnas.1801351115
Siegel, J. S., Seitzman, B. A., Ramsey, L. E., Ortega, M., Gordon,
E. M., Dosenbach, N. U. F., . . . Corbetta, M. (2018). Re-
emergence of modular brain networks in stroke recovery. Cortex,
101, 44–59. https://doi.org/10.1016/j.cortex.2017.12.019
Network Neuroscience
1139
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Structural basis for functional lag in chronic stroke
Siegel, J. S., Shulman, G. L., & Corbetta, M. (2017). Measuring
functional connectivity in stroke: Approaches and considera-
tions. Journal of Cerebral Blood Flow and Metabolism, 37(8),
2665–2678. https://doi.org/10.1177/0271678x17709198
Siegel, J. S., Snyder, A. Z., Ramsey, L., Shulman, G. L., & Corbetta,
M. (2016). The effects of hemodynamic lag on functional con-
nectivity and behavior after stroke. Journal of Cerebral Blood
Flow & Metabolism, 36(12), 2162–2176. https://doi.org/10.1177/
0271678x15614846
Silasi, G., & Murphy, T. H. (2014). Stroke and the connectome:
how connectivity guides therapeutic intervention. Neuron, 83(6),
1354–1368. https://doi.org/10.1016/j.neuron.2014.08.052
Small, S. L., Hlustik, P., Noll, D. C., Genovese, C., & Solodkin,
A. (2002). Cerebellar hemispheric activation ipsilateral to the
paretic hand correlates with functional recovery after stroke.
Brain: A Journal of Neurology, 125(Pt 7), 1544–1557.
Smith, S. M., Beckmann, C. F., Andersson, J., Auerbach, E. J.,
Bijsterbosch, J., Douaud, G., . . . Glasser, M. F. (2013). Resting-
state fMRI in the human connectome project. NeuroImage, 80,
144–168. https://doi.org/10.1016/j.neuroimage.2013.05.039
Sporns, O., Chialvo, D. R., Kaiser, M., & Hilgetag, C. C. (2004).
Organization, development and function of complex brain net-
works. Trends in Cognitive Sciences, 8(9), 418–425. https://doi.
org/10.1016/j.tics.2004.07.008
Tombari, D., Loubinoux, I., Pariente, J., Gerdelat, A., Albucher, J.-F.,
Tardy, J., . . . Chollet, F. (2004). A longitudinal fMRI study: in
recovering and then in clinically stable sub-cortical stroke pa-
tients. NeuroImage, 23(3), 827–839. https://doi.org/10.1016/j.
neuroimage.2004.07.058
van den Heuvel, M. P., & Hulshoff Pol, H. E. (2010). Exploring
the brain network: a review on resting-state fMRI functional
connectivity. European Neuropsychopharmacology, 20(8), 519–
534. https://doi.org/10.1016/j.euroneuro.2010.03.008
Van Essen, D. C., Smith, S. M., Barch, D. M., Behrens, T. E. J., Yacoub,
E., Ugurbil, K., & WU-Minn HCP Consortium (2013). The WU-
minn Human Connectome Project: an overview. NeuroImage,
80, 62–79. https://doi.org/10.1016/j.neuroimage.2013.05.041
Wang, L., Yu, C., Chen, H., Qin, W., He, Y., Fan, F., . . . Zhu,
C. (2010). Dynamic functional reorganization of the motor ex-
ecution network after stroke. Brain: A Journal of Neurology,
133(Pt 4), 1224–1238. https://doi.org/10.1093/brain/awq043
Wang, X., Wong, W., Sun, R., Chu, W. C., & Tong, K. Y. (2018).
Differentiated effects of robot hand training with and without
neural guidance on neuroplasticity patterns in chronic stroke.
Frontiers in Neurology, 9. https://doi.org/10.3389/fneur.2018.
00810
Ward, N. S., & Cohen, L. G. (2004). Mechanisms underlying recov-
ery of motor function after stroke. Archives of Neurology, 61(12),
1844–1848. https://doi.org/10.1001/archneur.61.12.1844
Wei, D., Zhuang, K., Ai, L., Chen, Q., Yang, W., Liu, W., . . . Qiu,
J. (2018). Structural and functional brain scans from the cross-
sectional Southwest University adult lifespan dataset. Scientific
Data, 5(180134). https://doi.org/10.1038/sdata.2018.134
Wei, G.-X., Dong, H.-M., Yang, Z., Luo, J., & Zuo, X.-N. (2014). Tai
Chi Chuan optimizes the functional organization of the intrin-
sic human brain architecture in older adults. Frontiers in Aging
Neuroscience, 6, 74. https://doi.org/10.3389/fnagi.2014.00074
Wieloch, T., & Nikolich, K. (2006). Mechanisms of neural plasticity
following brain injury. Current Opinion in Neurobiology, 16(3),
258–264. https://doi.org/10.1016/j.conb.2006.05.011
Wong, W.-W., Chan, S.-T., Tang, K.-W., Meng, F., & Tong,
K.-Y. (2013). Neural correlates of motor impairment during mo-
tor imagery and motor execution in sub-cortical stroke. Brain
Injury, 27(6), 651–663. https://doi.org/10.3109/02699052.2013.
771796
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari,
D., Hollinshead, M., . . . Buckner, R. L. (2011). The organization
of the human cerebral cortex estimated by intrinsic functional
connectivity. Journal of Neurophysiology, 106(3), 1125–1165.
https://doi.org/10.1152/jn.00338.2011
Zeiler, S. R., & Krakauer, J. W. (2013). The interaction between
training and plasticity in the poststroke brain. Current Opinion
in Neurology, 26(6), 609–616. https://doi.org/10.1097/WCO.
0000000000000025
Zhao, Y., Lambon Ralph, M. A., & Halai, A. D. (2018). Relating
resting-state hemodynamic changes to the variable language pro-
files in post-stroke aphasia. NeuroImage: Clinical, 20, 611–619.
https://doi.org/10.1016/j.nicl.2018.08.022
Network Neuroscience
1140
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
1
1
8
6
6
8
8
4
n
e
n
_
a
_
0
0
1
0
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3