INVESTIGACIÓN
Centralized and distributed cognitive task
processing in the human connectome
Enrico Amico1,2, Alex Arenas
3, and Joaquín Goñi1,2,4
1School of Industrial Engineering, Purdue University, West-Lafayette, EN, EE.UU
2Purdue Institute for Integrative Neuroscience, Purdue University, West-Lafayette, EN, EE.UU
3Departament d’Enginyeria Informàtica i Matemàtiques, Universitat Rovira i Virgili, Tarragona, España
4Weldon School of Biomedical Engineering, Purdue University, West-Lafayette, EN, EE.UU
Palabras clave: Brain connectomics, Conectividad funcional, Network science, Information theory,
Cognitive task processing
un acceso abierto
diario
ABSTRACTO
A key question in modern neuroscience is how cognitive changes in a human brain can be
quantified and captured by functional connectivity (FC). A systematic approach to measure
pairwise functional distance at different brain states is lacking. This would provide a
straightforward way to quantify differences in cognitive processing across tasks; también, it would
help in relating these differences in task-based FCs to the underlying structural network. Aquí
proponemos un marco, based on the concept of Jensen-Shannon divergence, to map the
task-rest connectivity distance between tasks and resting-state FC. We show how this
information theoretical measure allows for quantifying connectivity changes in distributed
and centralized processing in functional networks. We study resting state and seven tasks
from the Human Connectome Project dataset to obtain the most distant links across tasks.
We investigate how these changes are associated with different functional brain networks,
and use the proposed measure to infer changes in the information-processing regimes.
Además, we show how the FC distance from resting state is shaped by structural
conectividad, and to what extent this relationship depends on the task. This framework
provides a well-grounded mathematical quantification of connectivity changes associated
with cognitive processing in large-scale brain networks.
RESUMEN DEL AUTOR
A key question in modern neuroscience is how cognitive changes in a human brain can be
quantified and captured by functional connectivity (FC). Here we propose a framework,
based on Jensen-Shannon divergence, to define “connectivity distance” and to infer about
brain network reconfiguration across different tasks with respect to resting state, y para
explore changes in centralized and distributed processing in FCs.
Three functional networks (atención dorsal, frontoparietal and DMN) showed major changes
in distributed processing and minor changes in centralized processing. Changes in
centralized processing depend on the underlying structural connectivity weights and
structural path “hiddenness.”
These findings suggest that the cognitive “switch” between resting state and task states is a
complex interplay between maximally and minimally distant functional connections, y el
underlying structure.
Citación: Amico, MI., Arenas, A., & Goñi,
j. (2019). Centralized and distributed
cognitive task processing in the human
conectoma. Neurociencia en red,
3(2), 455–474. https://doi.org/
10.1162/netn_a_00072
DOI:
https://doi.org/10.1162/netn_a_00072
Supporting Information:
https://doi.org/10.1162/netn_a_00072
https://engineering.purdue.edu/
ComplexityLab
Recibió: 31 Enero 2018
Aceptado: 24 Septiembre 2018
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Joaquín Goñi
jgonicor@purdue.edu
Editor de manejo:
Danielle Bassett
Derechos de autor: © 2018
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
Red:
An ordered pair formed by a set of
nodes and a set of edges (cual
represent connections between
pairs of nodes). Nodes are usually
represented by circles, mientras
edges are represented by lines or
arcs connecting pairs of nodes.
Diffusion-weighted imaging (DWI):
A form of magnetic resonance
imaging (resonancia magnética) technique based on
measuring the random Brownian
motion of water molecules within
a voxel based on sampling 3D
directions. This technique allows for
estimating white matter streamlines
and fiber bundles connecting brain
regiones.
Tractography:
Computational reconstruction
procedure that may be used to
obtener, from DWI data, white
matter streamlines or fiber tracts
connecting different brain regions.
Functional magnetic resonance
imaging (resonancia magnética funcional):
A noninvasive technique that
estimates brain activity by detecting
changes associated with blood flow.
The rationale of this technique relies
on the fact that there is a positive
association between cerebral blood
flow and neuronal activation.
Connectome or structural
conectividad (CAROLINA DEL SUR) matrix:
A network representation of the
physical connections in the brain.
Nodes represent brain regions,
whereas edges represent physical
connections of pairs of brain
regions through the white matter.
Weights typically denote the density
or the integrity of the connections.
INTRODUCCIÓN
The progress in neuroimaging methodologies in recent years, together with the rise of pub-
licly available datasets (VanEssen, Herrero, et al., 2013; VanEssen, Ugurbil, et al., 2012), tiene
boosted research on quantitative analysis of brain connectivity patterns based on network sci-
ence. The intuition of modeling the brain as a network (bassett & bullmore, 2006; bullmore
& despreciar, 2009; Proporcionó, Brilla, & bullmore, 2016; despreciar, 2011) has rapidly expanded into
the scientific area denominated brain connectomics (Proporcionó, Brilla, & romper la lanza, 2015;
Fornito et al., 2016). In brain network models, nodes correspond to gray matter regions (based
on brain atlases or parcellations), while links or edges correspond to structural or functional
connections. Structural connections are estimated from diffusion-weighted imaging (Bihan,
2003; Tournier, Mori, & Leemans, 2011) data by modeling white matter pathways through
tractography algorithms (R. mi. Herrero, Tournier, Calamante, & connelly, 2012, 2015; Tournier,
Calamante, & connelly, 2012). Functional connections represent statistical dependencies be-
tween brain region time series while subjects are either at rest or performing a task during
resonancia magnética funcional (resonancia magnética funcional) sessions (van den heuvel & Hulshoff Pol, 2010). These functional asso-
ciations are usually measured via correlations among fMRI time series to study functional con-
conectividad (FC) en el cerebro humano (Fox & Raichle, 2007; van den heuvel & Hulshoff Pol, 2010).
These recent advances have led the brain connectivity community to start exploring and
quantify differences between resting-state FCs and task-based FCs (Col, bassett, Fuerza, Más valiente,
& Petersen, 2014; Cole y col., 2013; Col & Schneider, 2007; Gonzalez-Castillo et al., 2012;
graton, Laumann, gordon, Esperemos, & Petersen, 2016; Krienen, yo, & Buckner, 2014).
Some of the main lines of research in this direction involved whole-brain network similar-
ity analyses on the intrinsic and task-evoked network architecture of human connectome
(Cole y col., 2014); the mapping of cortical hubs and brain region for adaptive task control
(so-called cognitive control network (Cole y col., 2013; Col & Schneider, 2007); and the in-
vestigation of activity flow from resting-state FCs to infer brain regions that carry diverse cog-
nitive task information (Cole y col., 2013; Ito et al., 2017). Despite all these efforts in trying to
characterize connectivity differences between resting-state and task activity in brain networks,
a systematic analysis on how to measure pairwise (es decir., at the level of FC links) “cognitive dis-
tance” between these different functional states is still lacking. Such a methodology would pro-
vide a straightforward way to quantify differences in cognitive processing across tasks; también, él
would help in relating these local differences in task-based FCs to the underlying structural net-
work architecture, another exciting avenue for the brain connectomics community (Fukushima
et al., 2018; Hermundstad et al., 2013; Hermundstad et al., 2014; Medaglia et al., 2018; Miši´c
et al., 2016; see also our recent work, Amico & Goñi, 2018).
Here we propose a framework, based on Jensen-Shannon (JS) divergencia (Brit & Harremos,
2009; Cover & tomás, 2012), to map the “cognitive distance” between task and resting-state
functional connections. We show how this simple measure allows for quantifying the amount
of changes in distributed and centralized processing in human functional networks.
We use resting-state and seven different task sessions from the Human Connectome Project
(HCP) database to obtain the most JS-distant edges across tasks. We study how these changes
across tasks are associated with different functional brain networks, and use the proposed
measure to infer modifications in the information-processing regimes of these networks. Fur-
thermore, we show how cognitive distance is shaped by the brain structural architecture and
the level of nestedness of axonal pathways, and to what extent this relationship depends on the
tarea. We conclude by discussing the new insights offered by this approach, as well as potential
applications and future directions.
Neurociencia en red
456
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
Jensen-Shannon divergence:
In probability theory and statistics,
the Jensen-Shannon divergence
is a method of measuring the
dissimilarity between two probability
distributions. The square root of the
Jensen-Shannon divergence is a
metric often referred to as Jensen-
Shannon distance.
MÉTODOS
Dataset
The fMRI dataset used in this work is from the publicly available Human Connectome Project
(HCP, http://www.humanconnectome.org/ ), Release Q3. Per HCP protocol, all subjects gave
written informed consent to the Human Connectome Project consortium. Below is the full
description of the acquisition protocol and processing steps.
HCP: Functional Data
We assessed the 100 unrelated subjects (54 hembras, 46 machos, edad media = 29.1 años, DE =
3.7 años) as provided at the HCP 900 subjects data release (VanEssen, Herrero, et al., 2013;
VanEssen, Ugurbil, et al., 2012). This subset of subjects provided by HCP ensures that they
are not family relatives. This criterion was crucial to exclude the need of family-structure co-
variables in our analyses as well as possible identifiability confounds. The fMRI resting-state
runs were acquired in separate sessions on two different days (HCP filenames: rfMRI_REST1
and rfMRI_REST2), with two different acquisitions (left to right or LR, and right to left or RL)
per day (Glasser et al., 2013; S. METRO. Smith et al., 2013). The seven fMRI tasks were gam-
bling (tfMRI_GAMBLING), relational (tfMRI_RELATIONAL), social (tfMRI_SOCIAL), working
memory (tfMRI_WM), motor (tfMRI_MOTOR), idioma (tfMRI_LANGUAGE, including both a
story-listening and an arithmetic task), and emotion (tfMRI_EMOTION). The working memory,
gambling, and motor tasks were acquired on the first day, and the other tasks were acquired
on the second day (Barch et al., 2013; VanEssen, Herrero, et al., 2013). The HCP scanning
protocol was approved by the local Institutional Review Board at Washington University in
Calle. luis. For all sessions, data from both the left-right (LR) and right-left (rl) phase-encoding
runs were averaged to calculate connectivity matrices. This operation was done for all seven
fMRI tasks. Full details on the HCP dataset have been published previously (Glasser et al.,
2013; S. METRO. Smith et al., 2013; VanEssen, Ugurbil, et al., 2012).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
HCP: Structural Data
We used DWI runs from the same 100 unrelated subjects of the HCP 900 subjects data re-
lease (VanEssen, Herrero, et al., 2013; VanEssen, Ugurbil, et al., 2012). The diffusion acquisi-
tion protocol is covered in detail elsewhere (Glasser et al., 2013; Sotiropoulos et al., 2013;
Uurbil et al., 2013). Below we mention the main characteristics. Very high-resolution acqui-
sitions (1.25 mm isotropic) were obtained by using a StejskalTanner (monopolar) (Stejskal &
Tanner, 1965) diffusion-encoding scheme. Sampling in q-space was performed by including
three shells at b = 1, 000, 2, 000, y 3, 000 s/mm2. For each shell corresponding to 90 difusión
gradient directions and five reference volumes ( b = 0) were obtained, with the phase-encoding
direction reversed for each pair (es decir., LR and RL pairs). Directions were optimized within and
across shells (es decir., staggered) to maximize angular coverage using the approach of Caruyer et al.
(2011; http://www-sop.inria.fr/members/Emmanuel.Caruyer/q-space-sampling.php), and form
a total of 270 noncollinear directions for each phase-encoding direction. Correction for EPI
and eddy current-induced distortions in the diffusion data was based on manipulation of the
acquisitions so that a given distortion manifests itself differently in different images (andersson,
Skare, & Ashburner, 2003). To ensure better correspondence between the phase-encoding re-
versed pairs, the whole set of diffusion-weighted (DW) volumes is acquired in six separate
series. These series were grouped into three pairs, and within each pair the two series
contained the same DW directions but with reversed phase-encoding (es decir., a series of Mi
Neurociencia en red
457
Cognitive task processing in the human connectome
DW volumes with RL phase-encoding is followed by a series of Mi volumes with LR phase-
encoding, where i = [1, 2, 3]).
Brain atlas. We employed a cortical parcellation into 360 brain regions as recently proposed
by Glasser et al. (2016). For completeness, 14 subcortical regions were added, as provided
by the HCP release (filename Atlas_ROI2.nii.gz). para hacerlo, this file was converted from NIFTI
to CIFTI format by using the HCP workbench software (Glasser et al., 2013; Marcus et al., 2011;
http://www.humanconnectome.org/software/connectome-workbench.html, command -cifti-
create-label).
HCP Preprocessing: Functional Data
The HCP functional preprocessing pipeline (Glasser et al., 2013; S. METRO. Smith et al., 2013)
was used for the employed dataset. This pipeline included artifact removal, motion correction,
and registration to standard space. Full details on the pipeline can be found in Glasser et al.
(2013) and S. METRO. Smith et al. (2013). The main steps were spatial (minimal) preprocessing,
in both volumetric and grayordinate forms (es decir., where brain locations are stored as surface
vertices; S. METRO. Smith et al., 2013); weak high-pass temporal filtering (>2,000 s full width at
half maximum) applied to both forms, achieving slow drift removal; MELODIC ICA (Jenkinson,
beckman, Behrens, lana rica, & Herrero, 2012) applied to volumetric data; and artifact com-
ponents identified using FIX (Salimi-Khorshidi et al., 2014). Artifacts and motion-related time
courses were regressed out (es decir., the six rigid-body parameter time series, their backwards-
looking temporal derivatives, plus all 12 resulting regressors squared) of both volumetric and
grayordinate data (S. METRO. Smith et al., 2013).
For the resting-state fMRI data, we also added the following steps: global gray matter signal
was regressed out of the voxel time courses (Power et al., 2014); a bandpass first-order But-
terworth filter in forward and reverse directions [0.001 Hz, 0.08 Hz] (Power et al., 2014) era
aplicado (MATLAB functions butter and filtfilt); and the voxel time courses were z-scored and
then averaged per brain region, excluding outlier time points outside of 3 standard deviation
from the mean, using the workbench software (workbench command -cifti-parcellate). For task
fMRI data, we applied the same above-mentioned steps but we opted for a more liberal band-
pass filter [0.001 Hz, 0.25 Hz], since it is still unclear the connection between different tasks
and optimal frequency ranges (Cole y col., 2014).
Pearson correlation coefficients between pairs of nodal time courses were calculated
(MATLAB command corr), resulting in a symmetric connectivity matrix for each fMRI session
of each subject. In this paper we will refer to this matrix as functional connectivity matrix or
functional connectome (FC). As aforementioned, data from both the left-right (LR) y correcto-
izquierda (rl) phase-encoding runs were averaged to calculate individual functional connectomes
in each fMRI session. Functional connectivity matrices were kept in its signed weighted form,
hence neither thresholded nor binarized. Finalmente, the resulting individual functional connec-
tivity matrices were ordered (rows and columns) according to seven functional cortical sub-
redes (FNs) as proposed by Yeo and colleagues (Yeo et al., 2011). para hacerlo, for each brain
region in the Glasser atlas, the FN-membership (as a percentage) to each of the seven functional
redes (sum of the membership vector being equal to 1) was calculated. Finalmente, each brain
region was assigned to the most highly present FN. For completeness, an eighth subnetwork
including the 14 HCP subcortical regions was added (as analogously done in recent papers;
Amico & Goñi, 2018; Amico et al., 2017).
Functional connectome/connectivity
(FC) matrix:
A network representation of the
functional coupling between brain
regiones. Such coupling is usually
measured by quantifying the
statistical dependencies between
time series of brain regions (p.ej.,
pairwise Pearson’s correlation,
mutual information) as obtained by
functional magnetic resonance
imaging (resonancia magnética funcional).
Neurociencia en red
458
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
HCP Preprocessing: Structural Data
The HCP DWI data were processed following the MRtrix3 (Tournier et al., 2011) pautas
(http://mrtrix.readthedocs.io/en/latest/tutorials/hcp_connectome.html). En resumen, we first
generated a tissue-segmented image appropriate for anatomically constrained tractography
(ACT; R. mi. Smith et al., 2012; MRtrix command 5ttgen); we then estimated the multi-shell,
multi-tissue response function (Christiaens et al., 2015; MRtrix command dwi2response
msmt_5tt) and performed the multi-shell, multi-tissue constrained spherical deconvolution
(Jeurissen, Tournier, Dhollander, connelly, & Sijbers, 2014; MRtrix dwi2fod msmt_csd). Después-
wards, we generated the initial tractogram (MRtrix command tckgen, 10 million streamlines,
maximum tract length = 250, FA cutoff = 0.06) and applied the successor of spherical-
deconvolution informed filtering of tractograms (SIFT2; R. mi. Smith et al., 2015) methodology
(MRtrix command tcksift2). Both SIFT (R. mi. Herrero, Tournier, Calamante, & connelly, 2013)
and SIFT2 (R. mi. Smith et al., 2015) methods provide more biologically meaningful estimates
of structural connection density. SIFT2 allows for a more logically direct and computationally
efficient solution to the streamlines connectivity quantification problem: By determining an
appropriate cross-sectional area multiplier for each streamline rather than removing stream-
lines altogether, measures of fiber connectivity are obtained while making use of the complete
streamlines reconstruction (R. mi. Smith et al., 2015). Finalmente, we mapped the SIFT2 outputted
streamlines onto the 374 chosen brain regions (360 from the Glasser et al., 2016, brain atlas
plus 14 subcortical regions; see the Brain Atlas section) to produce a structural connectome
(MRtrix command tck2connectome). Finalmente, a log10 transformation (Fornito et al., 2016) era
applied on the structural connectomes (CAROLINA DEL SUR; es decir., the anatomical networks) to better account
for differences at different magnitudes. En consecuencia, SC values ranged between 0 y 5 en
this dataset.
Jensen-Shannon Distance on Functional Edges
The Jensen-Shannon divergence is a method commonly used to measure dissimilar-
ities between two probability distributions (Brit & Harremos, 2009; Cover & tomás, 2012;
De Domenico, Nicosia, Arenas, & latora, 2015). In the case of two discrete probability distri-
butions P and Q, the Jensen-Shannon divergence (JSD) is defined by
JSD(PAG||q) = 1
2
DKL(PAG||METRO) + 1
2
DKL(q||METRO),
(1)
where M = 1
2
For two discrete probability distributions P and Q, it is defined by
(PAG + q) and DKL is the Kullback-Leibler divergence (Cover & tomás, 2012).
DKL(PAG||q) = − ∑
i
PAG(i) registro
q(i)
PAG(i) .
(2)
For the particular case of measuring the dissimilarity between two probability distributions P
and Q, the Jensen-Shannon divergence is bounded between 0 y 1, given that one uses the
base 2 logarithm:
0 ≤ JSD(PAG||q) ≤ 1.
(3)
It has been shown that the square root of the Jensen-Shannon divergence is a well-defined
distance metric (Endres & Schindelin, 2003; Österreicher &Vajda, 2003), often referred to as
“Jensen-Shannon distance”:
(cid:2)
JSdist(PAG||q) =
JSD(PAG||q).
(4)
459
Neurociencia en red
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
We used the JSdist to map “connectivity distance” between resting-state and task sessions.
Here we assume resting state to be the “cognitive baseline,” and we measured the JSdist link
to link from every task FCs to resting-state FCs. Below follows a detailed description of the
procedimiento (see also the scheme at Figure 1). Primero, for every edge in a functional connectome,
we extracted the corresponding individual values (out of 100 HCP subjects, we picked 50
subjects for resting-state FCs and 50 different subjects for the task FCs). en este estudio, this resulted
in having two vectors with 50 elementos, one for each resting-state edge and one for each
edge in the task FCs whose JS distance is to be evaluated. These vectors represent Pearson’s
correlation distributions of connectivity values across all subjects in the cohort. En segundo lugar, nosotros
transform these two Pearson’s distributions into discrete probability distributions. We sampled
el [−1, 1] Pearson’s range via uniform binning (bin width = 0.2), and counted the likelihood
of occurrence of the connectivity values in each bin. Finalmente, the JSdist between these two
probability distributions was computed for every edge and HCP task considered in this study.
This edgewise functional connectivity distance from resting state can be seen as a task-specific
connectivity distance. Eso es, how far the distribution of values is in a specific task with respect
to the resting-state FC baseline.
The Jensen-Shannon distance as defined above is thought for the general case where the
two distributions P and Q come from different data samples (p.ej., different subjects). This is not
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
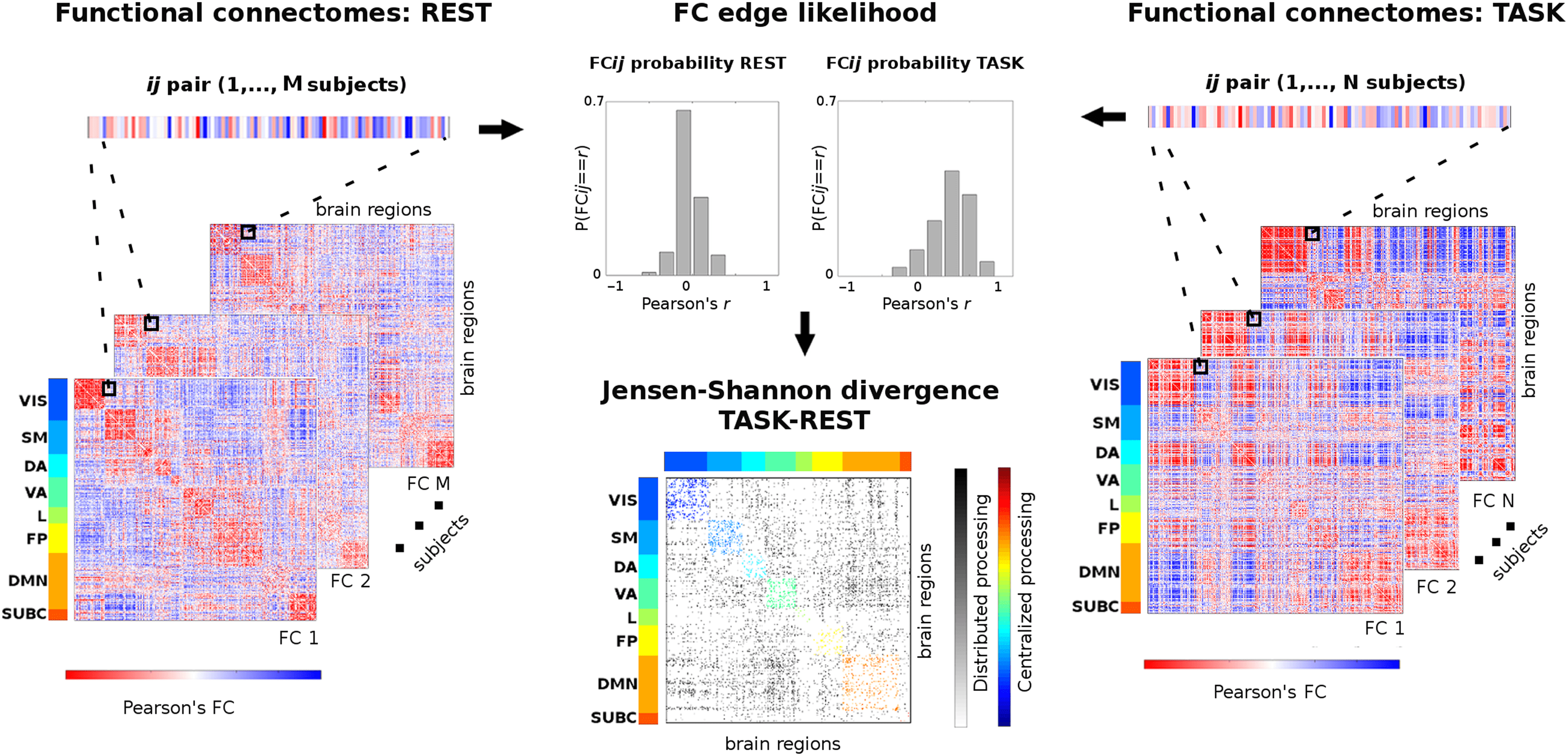
Cifra 1. Workflow scheme for task-rest connectivity distance (unpaired). This scheme summarizes the procedure to measure edgewise dis-
tance from two cohorts of (M and N) functional connectomes (FCs) at rest (izquierda) to a task-based one (bien). Primero, an edge ij is extracted from
the set, for both the resting-state and task-based FCs; these two vectors of M and N connectivity values are then transformed into probability
distributions (center top); finally, the Jensen-Shannon distance for these two edgewise probabilities is computed (center bottom). Iterating this
procedure over all possible ij pairs gives a Jensen-Shannon (JS) matrix of local distance in task FCs with respect to the REST baseline. El
JS matrix is ordered by the seven functional networks (FNs): visual (VIS), somatomotor (SM), atención dorsal (Y), atención ventral (Virginia),
límbico (l), frontoparietal (FP), and default mode network (DMN). An eighth subcortical network (SUBC) is added for completeness. Within-
network most distant edges are color coded according to FNs. Between-network most distant edges are in gray scale. This method allows for
quantifying the changes between centralized (within-network) and distributed (between-network) processing when a specific task is performed
with respect to the resting-state baseline.
Neurociencia en red
460
Cognitive task processing in the human connectome
dist
the most proper assessment in situations (like in the HCP data) where repeated measurements
from same subjects are available, allowing for paired comparisons between FCs. To cover the
necessity of that, we extended the concept of Jensen-Shannon distance by proposing a “paired”
version of JSdist, namely JSpaired
, como sigue. Similarly to JSdist, for every edge in a functional
conectoma, we extract the corresponding individual values. Then we take the element-wise
difference between the two vectors, where subjects are indexed in the same order. Such a
difference vector is then used to obtain distribution P([−2, 2] range, with uniform binning size
de 0.1). We then compare P against a “null” distribution Q, which is probability 1 at the bin
including the 0 valor, y 0 for all other bins. Such Q distribution reflects the expectation
of having no difference between repeated measurements. Deviations of P with respect to Q
on functional edges reflect individual changes (in any direction and/or magnitude) entre
repeated measurements on subjects. Finalmente, we iterate this procedure for every edge and task
to compute JSpaired
.
dist
Centralized and Distributed Processing in Functional Connectomes
The aforementioned procedure produced 374 × 374 (es decir., number of regions in the employed
brain atlas) JSdist matrices per each task. Próximo, we sought to relate the proposed connectivity
distance measure with changes in functional processing across functional networks (FNs). A
hazlo, we first thresholded the JSdist matrices based on the 95th percentile of the entire JSdist
distribution of values across all tasks, to select only the most distant links from resting state (ver
Cifra 2). Próximo, we quantify the amount of change in each of the eight functional FNs (see the
HCP Preprocessing: Functional Data section for details on the chosen FNs) by counting the
number of edges that survived the threshold divided by the total number of edges present in
each FN. We can then formalize changes in centralized processing (CP), for each functional
network k, como
CPk =
(cid:2)JSdist(i, j)
W Nk(i, j) ,
∑
i,j∈W Nk
∑
i,j∈W Nk
(5)
dónde (cid:3)JSdist is the binary version of the JSdist matrix thresholded at the 95 percentile (1 para
surviving edges, 0 elsewhere) for a specific task, and W Nk
is a binary matrix of the same size
como (cid:3)JSdist, con 1 if an edge falls within functional network k, y 0 elsewhere. Similarmente, uno
can quantify changes in distributed processing (DP) como
DPkl =
(cid:2)JSdist(i, j)
BNkl(i, j) ,
∑
i,j∈BNkl
∑
i,j∈BNkl
(6)
where now BNkl
functional networks k, yo, y 0 elsewhere.
is a binary matrix of the same size as (cid:3)JSdist, con 1 if an edge falls between
Por eso, for each one of the eight functional networks considered here (see the HCP Prepro-
cesando: Functional Data section for details), one can obtain one value of CP and seven values
of DP (considering all pairwise FN interactions), for a specific task. These values provide an
estimate of the density of the most connectivity distant functional links across, dentro, y
Neurociencia en red
461
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
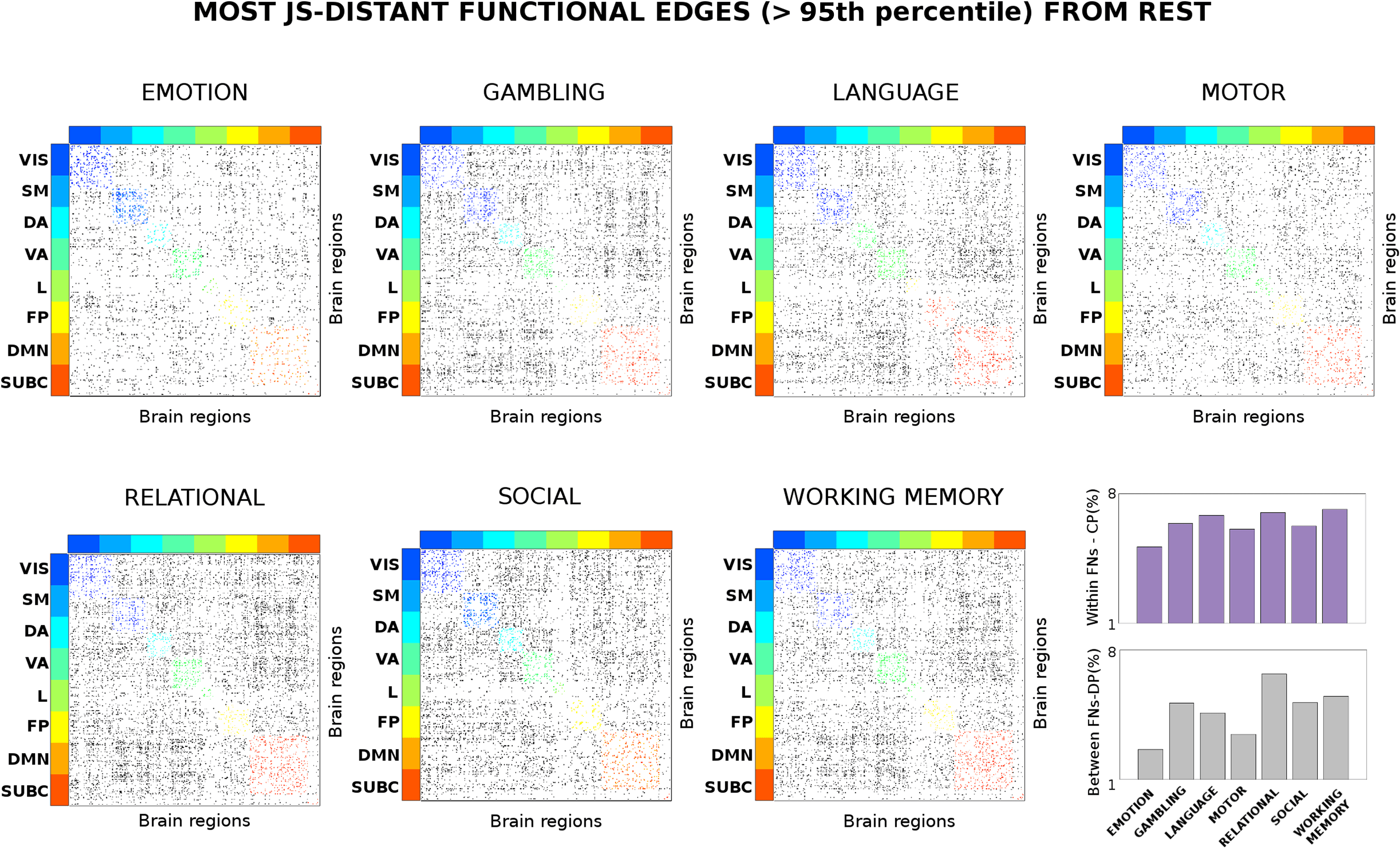
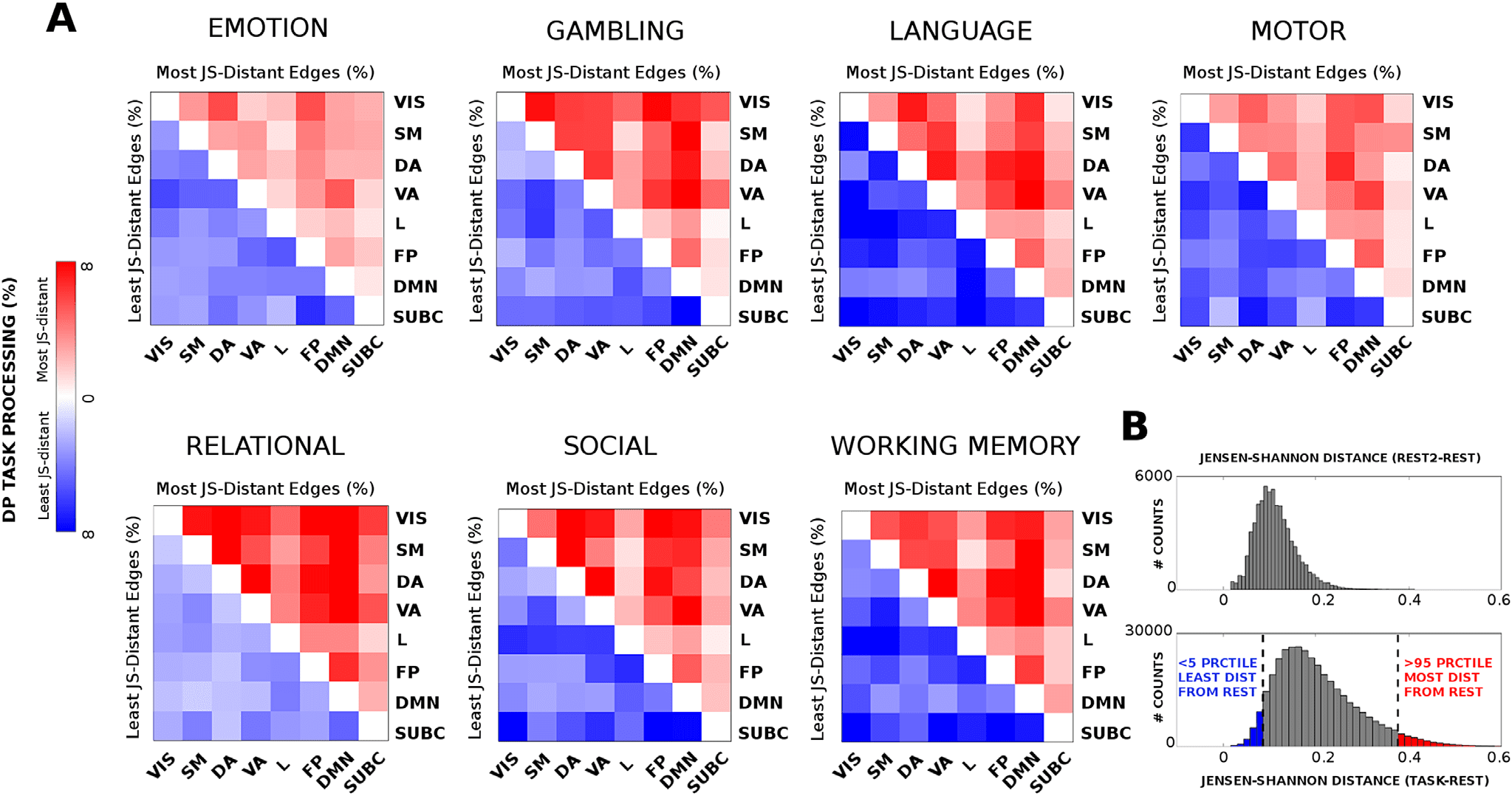
Cifra 2. Connectivity distance across different tasks. Evaluation of the most distant functional links (in terms of Jensen-Shannon [JS] distancia;
see Methods) across seven different task sessions. The JS matrices were thresholded at the 95th percentile of the distribution of JS values across
the seven tasks. The JS matrices were then ordered by seven functional networks (FNs; Yeo et al., 2011); visual (VIS), somatomotor (SM), dorsal
atención (Y), atención ventral (Virginia), límbico (l), frontoparietal (FP), and default mode network (DMN). An eight subcortical network (SUBC)
was added for completeness. The edges surviving the threshold corresponding to within-FN connections are color-coded accordingly. Edges
corresponding to between-FN connections are depicted in gray scale. Note how the connectivity distance depends on the task: en algunos casos
within-FN connectivity is more recruited (es decir., for the emotion task), in other between-FN connections are the most distant (es decir., relational
tarea). The bottom-right bar plots depict the average percentage of within-FN most distant edges, es decir., centralized processing (CP, violet bars)
and the average percentage of between-FN edges, eso es, distributed processing (DP, gray bars) across the different tasks.
between FN connectivity. Eso es, the amount of local changes in distributed and centralized
processing in each FN from baseline, defined as resting-state functional connectivity.
Bandpass Filter Evaluation on Centralized and Distributed Processing Analysis
In order to check whether the different bandpass ranges applied for resting state and task had
an impact on centralized and distributed processing in FCs, we applied the same (liberal,
es decir., [0.001Hz, 0.25Hz]) bandpass filter onto resting-state data. We then evaluated changes
in centralized and distributed processing after the new bandpass, and checked the similarity
with the “standard” bandpass results by computing the cosine similarity between the vectors
defined by centralized and distributed processing coordinates. Cosine similarity is a measure
of similarity between two nonzero vectors of an inner product space that measure the angle
between them. Aquí, each vector represents a point in the state space defined by centralized
and distributed processing.
Neurociencia en red
462
Cognitive task processing in the human connectome
Null Model Evaluation for Connectivity Distance Analysis
To validate the JSdist results in functional connectomes, we tested the same approach on
randomized counterparts (or “surrogates”) of the original data. para hacerlo, we employed the
amplitude adjusted fourier transform (AAFT) surrogates method (Schreiber & Schmitz, 2000)
to obtain data random surrogates. Starting from the 374 fMRI time series (one per brain region
in the atlas; see also the Brain Atlas section), we generated AAFT fMRI time series surrogates as
proposed in (Schreiber & Schmitz, 2000). This method aims to build surrogate time series that
preserve the amplitude distribution and the power spectrum of the original data (Schreiber &
Schmitz, 2000).
For each of the seven HCP tasks and resting state, we computed 100 surrogate versions of
the functional connectivity matrices, and then evaluated the number of nonzero elements in
(cid:3)JSdistsurrogate for each of the 100 realizations. This provided null distributions (one per task) para
the connectivity distance measure, which allowed us to test whether the results obtained on
the original FCs were statistically significant.
Finalmente, in order to test the significance of differences between pairs of (highly structured)
correlation matrices, we also employed a permutation test that preserved intact the correlation
structure of the FC matrices but randomly permuted the task/rest labels, computed the JS dis-
tance on the permuted data, and then evaluated the number of JS edges per task surviving the
95% threshold based on the original data.
Estimation of Functional Connectivity Distance Associations with Structural Connectomes
Próximo, we sought to assess the role of structural connections in the connectivity distance of func-
tional links across all seven tasks. In order to do so, we divided the group-averaged structural
conectividad (CAROLINA DEL SUR) weights (see the HCP Preprocessing: Structural Data section for details on SC
computation) into five different percentile intervals {0−20; 20−40; 40−60; 60−80; 80−100}. Nosotros
then counted the average number of most distant edges (es decir., the nonzero elements of (cid:3)JSdist)
falling in each of the five percentile intervals, for each of the seven HCP tasks. This provides
an estimate on the relationship between structural connections and connectivity distance and
whether it depends on the specific task being performed. We also tested whether centralized
and distributed processing depend on the “nestedness” or “hiddenness” of the structural path-
maneras, as measured by search information (Goñi et al., 2014; Rosvall, Grönlund, Minnhagen,
& Sneppen, 2005; Trusina, Rosvall, & Sneppen, 2005). Search information (SI) quantifies the
accessibility or hiddenness of the shortest path between a source node and a target node within
the network by measuring the amount of knowledge or information in bits needed to access
el camino (Goñi et al., 2014; Rosvall et al., 2005; Trusina et al., 2005; Wirsich et al., 2016) El
more nested the shortest path is between two brain regions ij, the higher its SI value; estafa-
versely, the less hidden or integrated the path, the lower its SI value. Similarly to the exper-
iment performed on SC weights, we again divided the group-average SI range of values into
five different percentile intervals: {0−20; 20−40; 40−60; 60−80; 80−100}. Finalmente, we counted
the average number of most distant edges (es decir., the nonzero elements of (cid:3)JSdist) falling in each
of the five SI percentile intervals, for each of the seven HCP tasks. This provides an estimate on
the relationship between structural “hiddenness” and connectivity distance and its associations
with the specific task being performed. For both SC weights and SI, the significance of the asso-
ciations with centralized and distributed processing was assessed through one-way analysis of
variance (ANOVA; Hogg & Ledolter, 1987; MATLAB command anova1), with “observations”
being centralized and distributed processing values for the seven tasks, and “groups” being the
five percentile intervals described above.
Search information:
A measurement that quantifies the
accessibility or hiddenness of the
shortest path between a source
node and a target node within the
network by measuring the amount
of knowledge or information
(expressed in bits) needed to
access that exact path.
Neurociencia en red
463
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
RESULTADOS
Metric:
In mathematics, a metric or distance
function is a function that defines a
distance between each pair of
elements of a set and satisfies the
properties of nonnegativity, identity
of indiscernibles, symmetry, y
triangle inequality. A set with a
metric is called a metric space.
The dataset used for this study consisted of functional data from the 100 unrelated subjects in
the Q3 release of the HCP (VanEssen, Herrero, et al., 2013; VanEssen, Ugurbil, et al., 2012).
We defined the “connectivity distance” between task FC links and resting-state FC links as the
edgewise Jensen-Shannon distance (JSdist) between resting-state FCs and task FCs (see also
the scheme at Figure 1). This metric quantifies the connectivity distance of a functional link
recruited in a task with respect to its correspondent “usage” in resting state. For each of the
seven HCP tasks (see Methods for details), we computed the corresponding JSdist and JSpaired
matrices, and extracted the most connectivity distant edges (≥ 95 percentile distribution of
JSdist and JSpaired
values across all tasks). Cifra 2 shows the results corresponding to JSdist
(unpaired, all different subjects for resting-state and task FCs), and Figure S1 (Amico, Arenas,
& Goñi, 2019) summarizes the results for JSpaired
(paired, same subjects for resting-state and
dist
task FCs). Please also see the Jensen-Shannon Distance on Functional Edges section for details.
dist
dist
Notablemente, the results obtained are significantly different from the same analyses performed
en 100 realizations of surrogate data built from the fMRI time series considered in this study
(Figure S2 and Table S1, Amico et al., 2019); see the Null Models Evaluation for Connectivity
Distance Analysis section for details). Además, with the only exception of motor versus
emotion for absolute frame displacement, no significant differences were observed in frame-
wise displacement estimates (pag < 0.01, double-sided t test between task pairs). This included
absolute frame displacement (root mean squared, HCP filename Abs_RMS) and relative frame
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
n
e
n
_
a
_
0
0
0
7
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
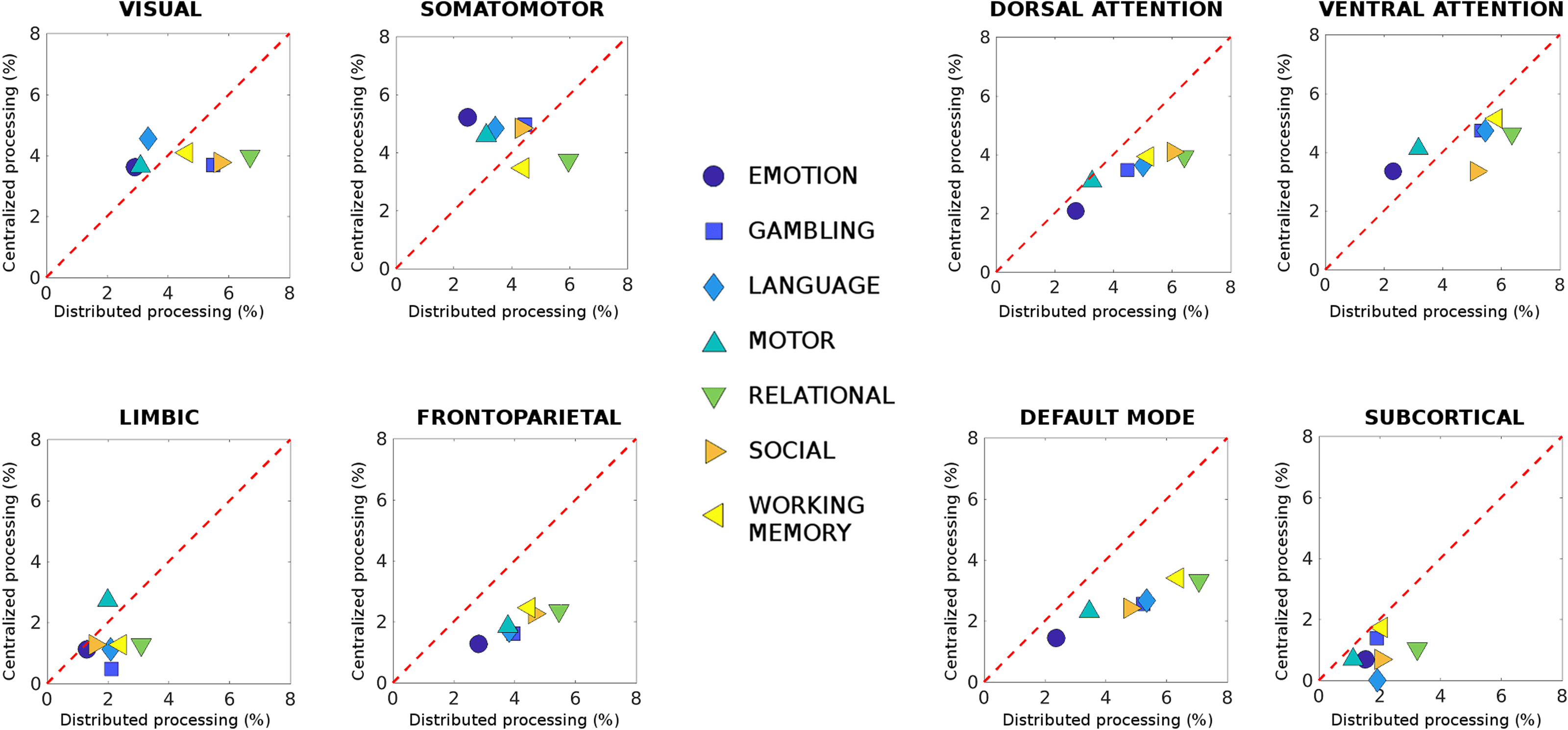
Figure 3. Centralized and distributed task processing in functional connectomes. Each plot shows differences in centralized versus distrib-
uted processing (see Methods) for each of the seven functional networks (FNs; visual, somatomotor, dorsal and ventral attention, limbic,
frontoparietal, and DMN; Yeo et al., 2011) and subcortical network, for all seven different HCP tasks. The difference in centralized processing
with respect to resting state was defined as the number of most Jensen-Shannon (JS) distant edges within-FN divided by the total number of
edges in the FN (reported as percentage). Similarly, deviations from distributed processing in resting state were defined as the number of most
JS-distant edges between FN divided by the total number of between-FN connections. Note how FP and DMN networks deviate from rest
mainly in the amount of distributed processing, that is, between-FN connectivity.
Network Neuroscience
464
Cognitive task processing in the human connectome
displacement (root mean squared, HCP filename Rel_RMS). These findings suggest that head
motion is not biasing rest to task JS distances depicted in Figure 2 and Figure S1 (Amico et al.,
2019).
Interestingly, the level of distance from resting state seems to be associated with the specific
task (Figure 2). For some tasks, the within-functional network links are more distant, that is,
more involved (e.g., for the emotion and motor tasks); in others the between-FN connections
are the most distant ones (i.e., relational or working memory tasks). The dichotomy between
intra-network (i.e., centralized) and inter-network (i.e., distributed) distance led us to quan-
tify the changes in centralized and distributed processing in task FCs (Figure 3; also see the
Centralized and Distributed Processing in Functional Connectomes section).
Note how, for three functional networks—dorsal, frontoparietal, and default mode—there is
a clear demarcation between centralized and distributed processing, for all seven tasks evalu-
ated (Figure 3). This indicates that the functional connections between these networks get more
distant from rest when they are recruited in a task. Furthermore, with the exception of limbic
and subcortical networks, where little difference in centralized and distributed processing can
be observed (Figure 3), in all the other FNs (i.e., visual, somatomotor, and ventral attention)
there is balance between intra- and inter-network processing. This trade-off seems to depend on
the task at hand (slightly more centralized in some, more distributed in others; Figure 3). Note
that these different distributions of centralized and distributed processing across tasks are not
related to the different bandpass applied for rest and task data, since very similar results were
obtained when the same (liberal, i.e., [0.001Hz, 0.25Hz]) bandpass parameters were applied
onto resting-state data; (see Figure S3 in the supporting information, Amico et al., 2019).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
n
e
n
_
a
_
0
0
0
7
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Least and most distant edges per functional network across tasks. (A) Heat maps, for all seven fMRI tasks evaluated, showing the most
(red, upper triangular) and least (blue, lower triangular) distributed processing (DP) values between pairs of functional networks with respect
to REST. (B) Top: distribution of JS-distance values when comparing REST2 session to the baseline REST session. Bottom: distribution of
Jensen-Shannon distance values across the seven tasks evaluated. The tails of the histogram are highlighted in blue (least distant edges, < 5th
percentile) and red (most distant edges, > 95th percentile).
Neurociencia en red
465
Cognitive task processing in the human connectome
When looking at JS-distance differences between functional networks across tasks, tarea-
specific patterns may be observed (see Figure 4A). Por ejemplo, relational tasks exhibit a
whole-brain tendency to get more distant from rest (es decir., higher general distributional process-
En g); emotion or motor tasks are among the least distant from rest; whereas some other tasks
(p.ej., language or social) display a more specialized distributed processing across functional
networks combining most and least distant functional edges. The histogram of the distribution
of JS-distance values across all tasks gives more insights on the cognitive distance task-rest:
there is a general tendency to be different from REST. Sin embargo, some edges (< 5%, Figure 4B,
blue bars) stay almost unchanged with respect to REST configuration; others (> 95%, Figura 4B,
red bars) switch to more distant values, allowing for the cognitive reconfiguration of the sys-
tema. Note that the JS distance does not depend on the baseline (es decir., REST FCs) magnitude of
the correlations, as the average correlation between JS-values and median REST FC correlation
values across tasks is −0.006 ± 0.026. En tono rimbombante, no single edge survived to the 95th per-
centile threshold after permutation testing of the TASK-REST labels (see Methods for details;
Figure S4, Amico et al., 2019). Analogously, less than 1% of REST2-REST edges survived to
that same threshold (Figura 4B). En general, these analyses indicate that the conservative thresh-
old chosen will depict actual task-rest FC changes.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
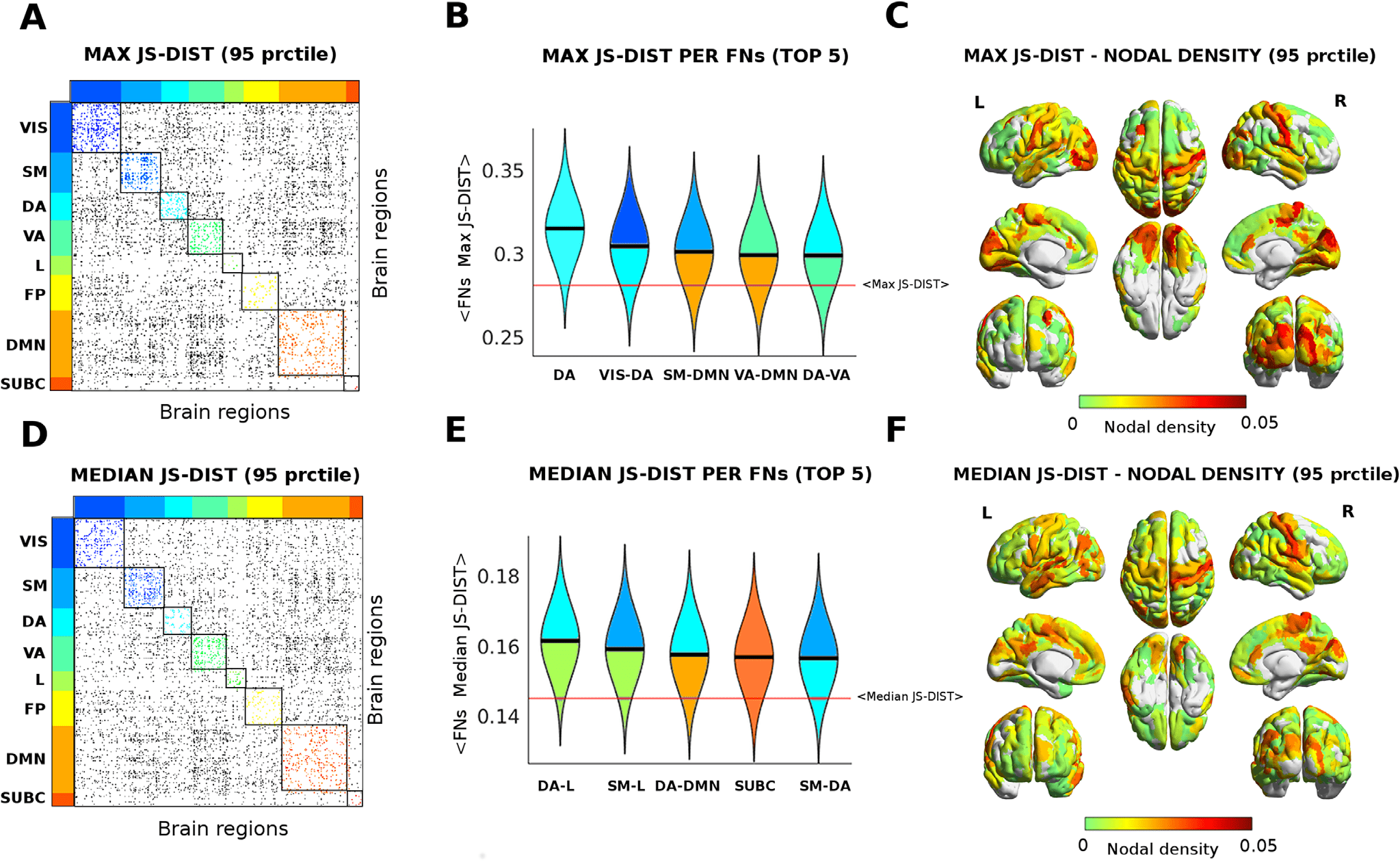
Cifra 5. Functional reconfiguration via Jensen-Shannon distance. (A–D) Edgewise max (A) and median (D) Jensen-Shannon distance across
all tasks (thresholded by 95th percentiles for max and for median). The colored dots depict JS values within FNs; gray dots indicate significant
JS-distant edges between FNs. (B–E) Violin plot of edgewise JS distance (max and median) for the top five FNs and FN interactions. Within-FNs
are color coded accordingly (as in A–D), while between-FNs are color coded using the colors of the two FNs involved. Solid black lines depict
median values of each distribution; solid red lines indicate the whole-brain median value of max and median distributions. (C–F) Brain render
of max and median JS distances as nodal density per region. The strength per brain region computed as sum of JS distance (max and median)
for functional edges above the 95 percentile threshold divided by the total number of brain regions.
Neurociencia en red
466
Cognitive task processing in the human connectome
The results in Figure 4 display the extent to which JS distance is sensitive to different levels
of functional reconfiguration (Kriene et al., 2014; Schultz & Col, 2016; Brillo y col., 2016)
for different functional networks across different tasks. Hence we decided to explore further
on this, and evaluated the edgewise maximum and median JS distance across all tasks. Este
would provide an overall summary of the main edges and functional networks involved in the
cognitive switch across the seven tasks (ver figura 5). Notice that median and max give two
different “flavors” of brain network reconfiguration: The median tells us how far, on average,
regiones del cerebro (and functional networks) change when subjects perform tasks. The max tells us
what are the most distant values, hence the highest achieved functional reconfiguration for a
brain region across the seven different tasks under study.
In order to determine whether changes in task processing are related to the underlying
structural connectivity, we first evaluated the relationship between connectivity distance in
each task and structural connectivity weights (Cifra 6, A1–B1). Curiosamente, a significant
trend arises for all tasks between centralized processing and number of tracts (one-way ANOVA
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
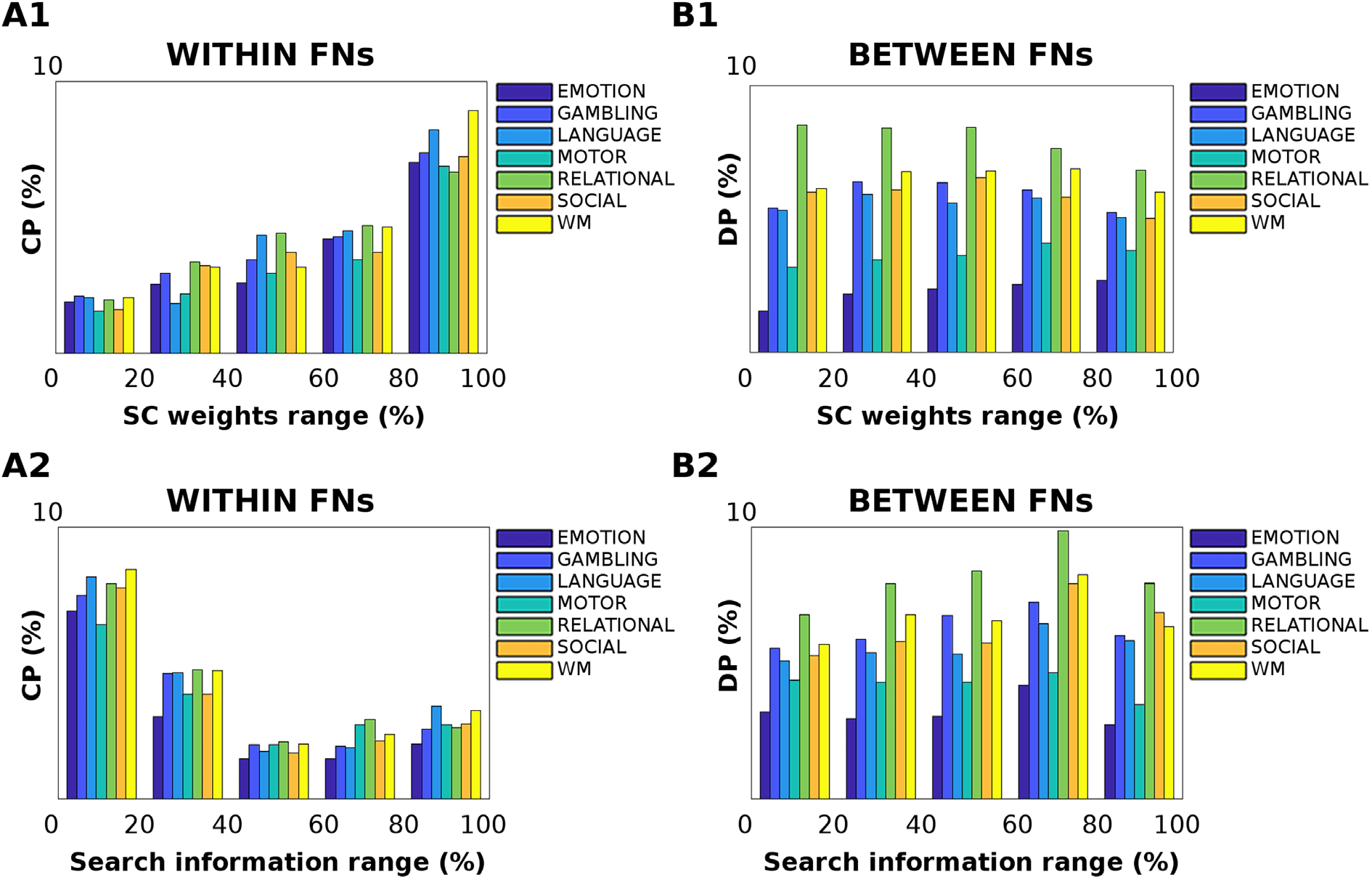
Cifra 6. Effect of structural pathways on centralized and distributed processing changes. (A1–B1) The relationship between the anatomical
connections and Jensen-Shannon distance was evaluated across the seven different tasks. The bar plots show the percentage of centralized
Procesando (CP) within functional networks (FNs, A1) and distributed processing (DP) between FNs (B1), per five different percentile ranges
of structural connectivity weights: 0−20, 20−40, 40−60, 60−80, and 80−100. The percentile range was extracted from the group-averaged
structural connectome. Note how, for within-FN connections (A1), the change in centralized processing significantly correlates with the
−20); conversely, the underlying
strength of structural connections across all tasks (one-way ANOVA F = 163.39, d f = 4, pag = 6.62 · 10
structural connectivity does not play a major role in distributed processing changes (one-way ANOVA F = 1.11, d f = 4, pag = 0.37). (A2–B2)
The effect of structural path accessibility (as measured by search information; see Methods) on centralized and distributed processing was
tested across the seven different tasks, per five different percentile intervals of search information: 0−20, 20−40, 40−60, 60−80, and 80−100.
The percentile range was extracted from the group-averaged search information matrix. Notablemente, changes in centralized processing (A2) son
−18); conversely, No
significantly associated with low values of search information (one-way ANOVA F = 131.75, d f = 4, pag = 1.41 · 10
significant association between SI and distributed processing changes was found (one-way ANOVA F = 1.85, d f = 4, pag = 0.14).
Neurociencia en red
467
Cognitive task processing in the human connectome
−20, Cifra 6, A1). Eso es, the more structurally connected
F = 163.39, d f = 4, pag = 6.62 · 10
two regions are within a functional network, the higher the number of centralized processing
edges recruited in a task. Notice how this trend is general and independent from the task, albeit
the magnitude of this linear association between structure and function depends on the task at
mano (Cifra 6, A1). Por otro lado, when looking at changes in distributed processing, eso
es, for edges involved in between-functional network connectivity, no significant associations
with structural connectivity were observed (one-way ANOVA F = 1.11, d f = 4, pag = 0.37,
Cifra 6, B1).
We then dug deeper into the relationship between task processing and structural connec-
tome by evaluating the level of hiddenness or accessibility of a structural path, as measured by
search information (Goñi et al., 2014; see also the Estimation of Functional Connectivity Dis-
tance Associations with Structural Connectomes section for details), and testing its association
with changes in cognitive task processing (Cifra 6, A2–B2). Curiosamente, the hiddenness of
structural paths appears to be inversely related to changes in centralized processing (one-way
−18, Cifra 6, A2). Eso es, the more “isolated” the
ANOVA F = 131.75, d f = 4, pag = 1.41 · 10
structural pathway between two brain regions within a functional network, the higher will be
its recruitment in a task. The more nested or integrated the path, the less distant the centralized
processing with respect to resting state (Cifra 6, A2). No significant associations were found
when looking at changes in distributed processing versus search information range of values
(one-way ANOVA F = 1.85, d f = 4, pag = 0.14, ver figura 6, B2).
DISCUSIÓN
Cognitive brain network mapping (Cole y col., 2013; Col & Schneider, 2007; Finn et al., 2017;
Kriene et al., 2014), or the analysis of brain network features underlying task performance
y control cognitivo (Cole y col., 2014; Finn et al., 2017; Gratton et al., 2016; Khambhati,
Medalla, Karuza, Thompson-Schill, & bassett, 2018; Tavor et al., 2016), is a recent and ex-
citing new line of investigation in brain connectomics. While the general intrinsic common
architecture between resting-state and task-based functional patterns has been explored (Col
et al., 2014), still very little is known about task connectivity distances and their associa-
tions with information processing (Col, Ito, bassett, & Schultz, 2016; Col & Schneider, 2007;
Ito et al., 2017). Además, an even more intricate question relates to the relationship be-
tween the different task-based FC scenarios and the underlying structural connectivity (Amico
& Goñi, 2018; Fukushima et al., 2018; Hermundstad et al., 2013; Hermundstad et al., 2014;
Miši´c et al., 2016).
Here we addressed these questions by proposing a novel methodology in neuroscience,
rooted to the concept of Jensen-Shannon divergence (Brit & Harremos, 2009; De Domenico
et al., 2015), to measure task-based pairwise functional distance with respect to the “cognitive
baseline” defined by resting-state FCs (Cifra 1). This framework may also be seen from a
multilayer perspective, with the ground layer being resting-state FCs and top layers defined
by the multiple task-based different connectivity scenarios. The distance defined here can be
thought as interlayer coupling, or as the amount of cognitive processing necessary to make the
“cognitive switch” from the resting-state ground layer to the top task-based functional layers.
The JS divergence has several advantages: it is a nonparametric test, does not assume any form
of distribution, and allows for quantifying fine-grained changes between two distributions. Como
shown in Figure S5 (Amico et al., 2019), there were a large number of functional edges for
which REST or TASK FC distributions did not pass a normality test.
Neurociencia en red
468
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
The work presented here complements the aforementioned recent studies on cognitive map-
ping, where the resting-state scaffolding was usually used to infer or also predict task changes
in connectivity (Cole y col., 2014; Cole y col., 2016; Ito et al., 2017; Tavor et al., 2016). Aquí
we evaluate and investigate the pairwise distance task-rest, and use it to map specific changes
dictated by the task at hand. This adds up to previous studies in that it improves our under-
standing of how edge specific is the cognitive switch, and its level of recruitment (in terms of
“connectivity distance”), as well as in terms of centralized and distributed processing changes
in functional networks (Cifra 2).
We exploited this new concept of connectivity distance to infer about the level of recruit-
ment of an edge or of a functional network (Cifra 2). Notablemente, the connectivity distant patterns
present in the original data were significantly different from the ones obtained by surrogate data
built from the original fMRI time series (Figure S2 and Table S1, Amico et al., 2019) or from the
ones obtained by randomly shuffling rest-task FCs. En efecto, the more distant a functional net-
work is in a specific task, the more different its recruitment with respect to resting state, and hence,
the more changes in cognitive information processing that functional subsystem will undergo.
This intuition led us to explore the concept of centralized and distributed processing in large-
scale functional networks, which we defined as the difference between intra- (es decir., centralized)
and inter- (es decir., distributed) network connectivity (Cifra 3). Curiosamente, three functional net-
obras (atención dorsal, frontoparietal, and DMN) showed major changes in distributed pro-
cessing and very minor changes in centralized processing, for all seven tasks evaluated with
respect to resting state (Cifra 3). This is in line with recent findings showing that frontoparietal
and attentional areas appear to be the more flexible for cognitive control and task performance
(Cole y col., 2013; Col & Schneider, 2007; Kriene et al., 2014; Brillo y col., 2016).
The fact that these networks and the DMN, which is well known to play a major role in
resting state (Greicius, Krasnow, Reiss, & menón, 2003; Raichle, 2015; Raichle et al., 2001),
change mainly in terms of intercommunication when transitioning to task is also noteworthy.
This finding goes along with the concept of integration of information between neural sub-
sistemas (Tononi, despreciar, & Edelman, 1994) and also with our recent findings on the associa-
tion between FP-DMN disconnectivity and degradation in arousal and levels of consciousness
(Amico et al., 2017). Possibly, the more demanding the task, the more the cross-talk between
FP, DMN, attentional networks, and the rest of the brain might be needed to achieve the
proper amount of cognitive processing or “brain network reconfiguration” (Kriene et al., 2014;
Schultz & Col, 2016).
We further investigated on the concept of brain network reconfiguration across tasks, por
evaluating most and least distant functional edges between functional networks (Cifra 4).
Curiosamente, some tasks seem to require extremely distant interactions between FNs (p.ej.,
relational), and other tasks require specific subsets of FN interactions (p.ej., idioma, working
memory). The investigation of the max and median nodal JSdist centrality (Cifra 5) entre tareas
added more information on the complex scenario depicted in Figure 4. Dorsal and occipital
regions seem to be the ones that are generally more distant from rest when engaged in a task
(Cifra 5, C.A). Por otro lado, somatomotor and dorsal areas seem to be the ones that
achieve the largest reconfiguration (es decir., maximal JSdist from resting state) across the seven
tasks evaluated (Cifra 5, D-F).
Tomados juntos, these findings suggest that the cognitive “switch” between resting-state and
task states is more than a general shift in terms of functional links, but rather a complex interplay
between maximally distant and minimally distant functional connections (Cifra 4, Cifra 5). Este
Neurociencia en red
469
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
is in line with recent studies investigating the complex reconfiguration of brain networks during
tareas (Bassett y cols., 2013; Khambhati et al., 2018; Kriene et al., 2014; Brillo y col., 2016).
Another major question relates to how these changes in cognitive processing are shaped or
determined by the underlying structural architecture of a human brain. Very few studies so far
have tried to elucidate the relationship between cognitive changes and axonal pathways, either
for localized cortical subsystems (p.ej., fusiform gyrus; Saygin et al., 2012) or for a specific task
(p.ej., visual stimuli; Osher et al., 2015), or at the whole-brain level (Hermundstad et al., 2013;
Hermundstad et al., 2014; Miši´c et al., 2016). In a recent work we tackled this problem from
a whole-brain network perspective, by means of an ICA-based approach to extract the main
“hybrid” functional-structural connectivity features sensitive to cognitive changes across seven
different tasks (Amico & Goñi, 2018).
Here we took this investigation one step further by assessing functional connectivity dis-
tance associations with respect to the underlying structural connectivity weights (Cifra 6).
Curiosamente, for changes in centralized processing, the relationship with structural connectivity
is linear (Cifra 6, A1). Eso es, when the cognitive processing involves mainly within-network
conectividad, the higher the fiber strength is between two regions, the more distant they will
ser. Sin embargo, this relationship is not present when looking at distributed processing link-to-
link effects (Cifra 6, B1). Previous findings reported that between-network connectivity links
can play a key role in the creation of more complex cognitive regimes (Fukushima et al., 2018;
Hermundstad et al., 2013; Hermundstad et al., 2014; Miši´c et al., 2016). The cross-talk be-
tween functional networks might bring the brain network up to a more integrated level, allow-
ing for a more dynamic and distributed cognitive processing, that ultimately deviates far from
the static underlying boundaries given by the structural fiber tracts.
To test this hypothesis, we evaluated the relationship between changes in distributed and
centralized processing and structural path “hiddenness” or accessibility, as measured by search
información (Goñi et al., 2014; see also the Estimation of Functional Connectivity Distance
Associations with Structural Connectomes section). Notablemente, when looking at centralized pro-
cessing deviations from rest, these two quantities appear to be inversely related (Cifra 6, A2,
B2). The less integrated the path is between two regions within a functional network, the higher
the value in centralized processing, the more integrated the structural pathways, and the less
centralized activity (Cifra 6, A2).
These findings corroborate the hypothesis on the integration segregation in the human brain
(Tononi et al., 1994). They are also in line with our findings on the importance of cross-talking
between functional networks for task changes (Amico & Goñi, 2018), which can be summa-
rized as follows: For a human brain to make a cognitive switch, a delicate interplay between
centralized and distributed processing is necessary. The centralized activity within functional
subsystems is shaped by brain structure. Además, the more isolated the shortest path con-
necting two centralized brain regions is, the higher the level of task processing (Cifra 4).
Sin embargo, in order to achieve proper cognitive complexity for the task at hand, an appropriate
level of distributed processing and subsequent integration between these subsystems is needed:
The level of cross-talking and structural integration will depend on the specific task at hand
(Cifra 2, Cifra 4) and on the functional subnetwork involved (Cifra 3), with no significant
function-structure associations (Cifra 4).
This study has several limitations. The framework presented here is based on the quan-
tification of distances between estimations of functional connectivity data. The accuracy and
representativity of the estimate JS distance will be subject to the quantity and quality of the fMRI
Neurociencia en red
470
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
data as well as the processing steps. Further studies should explore how different aspects of the
data and subsequent processing may have an impact in the JS-distance quantifications, incluir-
ing number of subjects, duration of the REST and TASK fMRI sessions, spatiotemporal limita-
tions on fMRI data, and motion regressors included, among others. Analogously, our findings
associating SC properties (weights and search information on SC shortest paths) with JS dis-
tance may also be, hasta cierto punto, sensitive to diffusion tractography user-defined free param-
eters. The effect of different brain atlases (here we used the one proposed by Glasser et al.
(2016)) and functional network organization (here we used the one proposed by Yeo et al.
(2011)) on the centralized and processing changes should be explored. Because of the way it
is defined, the measure does not allow for individualized connectivity distance patterns. Cómo-
alguna vez, future studies could explore to what extent individual FCs can be predicted by group-wise
changes in centralized and distributed processing, or use the information on the most distant
link at the group level to select the most meaningful pairwise connectivity for the task at hand.
Another potentially promising application of this methodology may be related to measuring JS
divergence at the single-subject level with dynamic functional connectivity. En este caso, el
different FCs would be given by the FC snapshots obtained from sliding the FC window along
the same subject acquisition during resting state and a specific task (as opposed to the slices
being subjects as depicted in Figure 1 of this paper).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
It is also important to remark on the importance of introducing a well-defined distance met-
ric (es decir., JSdist) in the functional connectome domain, as we propose in this work. This can open
several promising new lines of research in brain connectomics in terms of topological analysis
of the connectivity domain, and of the definition of metrizable spaces where to map cogni-
tive changes in brain functional networks. Finalmente, the simplicity of the methodology allows
for its applicability in the clinical domain, where it could be employed to measure connectiv-
ity distance between “healthy” and “diseased” populations (p.ej., Alzheimer’s, schizophrenia,
coma), or to determine task-rest distance in situations where the cognitive switch is damaged
or disrupted (p.ej., in autism or similar neurological disorders).
En conclusión, we have reported a new methodology that aims at capturing the functional
differences between different tasks when compared with resting state. The methodology based
on the Jensen-Shannon distance is promising, and has been proved to discern between cen-
tralized and distributed activity across brain areas for different tasks. These results pave the
way to the usage of this framework in other experiments, and to the development of a new
information theoretical framework for the analysis of functional and structural connectomes.
EXPRESIONES DE GRATITUD
Data were provided [en parte] by the Human Connectome Project, WU-Minn Consortium (prin-
cipal investigators: David Van Essen and Kamil Ugurbil; 1U54MH091657), funded by the 16
NIH Institutes and Centers that support the NIH Blueprint for Neuroscience Research; and by
the McDonnell Center for Systems Neuroscience at Washington University.
SUPPORTING INFORMATION
The code used for computing Jensen-Shannon distance in functional connectomes will be made
available on the CONNplexity lab website (https://engineering.purdue.edu/ConnplexityLab).
CONTRIBUCIONES DE AUTOR
Enrico Amico: Conceptualización; Curación de datos; Análisis formal; Investigación; Writing –
original draft; Escritura – revisión & edición. Alex Arenas: Conceptualización; Metodología;
Neurociencia en red
471
Cognitive task processing in the human connectome
Supervisión; Escritura – revisión & edición. Joaquín Goñi: Conceptualización; Investigación;
Metodología; Administración de proyecto; Supervisión; Validación; Escritura – borrador original;
Escritura – revisión & edición.
INFORMACIÓN DE FINANCIACIÓN
Joaquín Goñi, Institutos Nacionales de Salud (http://dx.doi.org/10.13039/100000002), Award
ID: R01EB022574. Joaquín Goñi, Institutos Nacionales de Salud (http://dx.doi.org/10.13039/
100000002), Award ID: R01MH108467. Joaquín Goñi, Indiana Clinical and Translational
Instituto de Ciencias (http://dx.doi.org/10.13039/100006975), Award ID: UL1TR001108. Alex
Arenas, Ministerio de Economía y Competitividad (http://dx.doi.org/10.13039/501100003329),
Award ID: FIS2015-71582-C2-1. Alex Arenas, Institució Catalana de Recerca i Estudis Avançats
(http://dx.doi.org/10.13039/501100003741). Alex Arenas, James S. McDonnell Foundation,
(http://dx.doi.org/10.13039/100000913), Award ID: 220020325.
REFERENCIAS
Amico, MI., Arenas, A., & Goñi, j. (2019). Supporting information
for “Centralized and distributed cognitive task processing in the
human connectome.” Network Neuroscience, 3(2), 455–474.
https://doi.org/10.1162/netn_a_00072
Amico, MI., & Goñi, j. (2018). Mapping hybrid functional-structural
connectivity traits in the human connectome. Network Neuro-
ciencia, 2(3), 306–322. https://doi.org/10.1162/netn_a_00049
Amico, MI., Marinazzo, D., Di Perri, C., Heine, l., Annen, J., Martial,
C., . . . Goi, j. (2017). Mapping the functional connectome traits
of levels of consciousness. NeuroImagen, 148, 201–211. https://
doi.org/10.1016/j.neuroimage.2017.01.020
andersson, j. l. r., Skare, S., & Ashburner, j. (2003). How to correct
susceptibility distortions in spin-echo echo-planar images: Appli-
cation to diffusion tensor imaging. NeuroImagen, 20(2), 870–888.
https://doi.org/10.1016/S1053-8119(03)00336-7
Respeto, D. METRO., Burgess, GRAMO. C., Harms, METRO. PAG., Petersen, S. MI.,
Schlaggar, B. l., Corbetta, METRO., . . . VanEssen, D. C. (2013). Func-
tion in the human connectome: Task-fMRI and individual differ-
ences in behavior. NeuroImagen, 80, 169–189. https://doi.org/10.
1016/j.neuroimage.2013.05.033
bassett, D. S., & bullmore, mi. (2006). Small-world brain networks.
The Neuroscientist, 12(6), 512–523.
bassett, D. S., Wymbs, norte. F., Rombach, METRO. PAG., Portero, METRO. A., Mucha,
PAG. J., & Grafton, S. t. (2013). Task-based core-periphery organ-
ization of human brain dynamics. Biología Computacional PLoS,
9(9), e1003171.
Bihan, D. l. (2003). Looking into the functional architecture of the
brain with diffusion MRI. Naturaleza Reseñas Neurociencia, 4(6), 469.
Brit, J., & Harremos, PAG. (2009). Properties of classical and quantum
Jensen-Shannon divergence. Physical Review A, 79(5), 052311.
https://doi.org/10.1103/PhysRevA.79.052311
bullmore, MI., & despreciar, oh. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Naturaleza
Reseñas Neurociencia, 10(3), 186–198. https://doi.org/10.1038/
nrn2575
Caruyer, MI., cheng, J., Lenglet, C., Sapiro, GRAMO., Jiang, T., & Deriche,
R. (2011). Optimal design of multiple Q-shells experiments for
diffusion MRI. https://hal.inria.fr/inria-00617663/documento
Christiaens, D., Reisert, METRO., Dhollander, T., Sunaert, S., Suetens,
PAG., & Maes, F. (2015). Global tractography of multi-shell diffusion-
weighted imaging data using a multi-tissue model. Neuro-
Image, 123, 89–101. https://doi.org/10.1016/j.neuroimage.2015.
08.008
Col, METRO. w., bassett, D. S., Fuerza, j. D., Más valiente, t. S., & Petersen,
S. mi. (2014). Intrinsic and task-evoked network architectures of
el cerebro humano. Neurona, 83(1), 238–251. https://doi.org/10.
1016/j.neuron.2014.05.014
Col, METRO. w., Ito, T., bassett, D. S., & Schultz, D. h. (2016). Activity
flow over resting-state networks shapes cognitive task activations.
Neurociencia de la naturaleza, 19(12), 1718.
Col, METRO. w., Reynolds, j. r., Fuerza, j. D., Repovs, GRAMO., Anticevic,
A., & Más valiente, t. S. (2013). Multi-task connectivity reveals flexi-
ble hubs for adaptive task control. Neurociencia de la naturaleza, 16(9),
1348. https://doi.org/10.1038/nn.3470
Col, METRO. w., & Schneider, W.. (2007). The cognitive control net-
trabajar: Integrated cortical regions with dissociable functions. Neuro-
Image, 37(1), 343–360. https://doi.org/10.1016/j.neuroimage.
2007.03.071
Cover, t. METRO., & tomás, j. A. (2012). Elements of information
theory. Hoboken, Nueva Jersey: John Wiley & Sons. (Google-Books-ID:
VWq5GG6ycxMC)
De Domenico, METRO. D., Nicosia, v., Arenas, A., & latora, V. (2015).
Structural reducibility of multilayer networks. Nature Communi-
cations, 6, 6864. https://doi.org/10.1038/ncomms7864
Endres, D. METRO., & Schindelin, j. mi. (2003). A new metric for probabil-
ity distributions. Transacciones IEEE sobre teoría de la información, 49(7),
1858–1860.
Finn, mi. S., Scheinost, D., Finn, D. METRO., shen, X., Papademetris, X.,
& Constable, R. t. (2017). Can brain state be manipulated to
emphasize individual differences in functional connectivity?
NeuroImagen, 160, 140–151.
Proporcionó, A., Brilla, A., & romper la lanza, METRO. (2015). The connectomics
of brain disorders. Naturaleza Reseñas Neurociencia, 16(3), 159–172.
https://doi.org/10.1038/nrn3901
Proporcionó, A., Brilla, A., & bullmore, mi. (2016). Fundamentals of
brain network analysis. Londres: Prensa académica.
Neurociencia en red
472
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
Fox, METRO. D., & Raichle, METRO. mi. (2007). Spontaneous fluctuations
in brain activity observed with functional magnetic resonance
imaging. Naturaleza Reseñas Neurociencia, 8(9), 700–711. https://
doi.org/10.1038/nrn2201
Fukushima, METRO., Betzel, R. F., Él, y., van den heuvel, METRO. PAG., Zuo,
X.-N., & despreciar, oh. (2018). Structure–function relationships dur-
ing segregated and integrated network states of human brain
conectividad funcional. Brain Structure and Function, 223(3),
1091–1106.
vidrio, METRO. F., carbonero, t. S., robinson, mi. C., hacker, C. D.,
Harwell, J., Yacoub, MI., . . . VanEssen, D. C. (2016). A multi-
modal parcellation of human cerebral cortex. Naturaleza, 536(7615),
171–178. https://doi.org/10.1038/nature18933
vidrio, METRO. F., Sotiropoulos, S. NORTE., wilson, j. A., carbonero, t. S.,
pescado, B., andersson, j. l., . . .
Jenkinson, METRO. (2013). El
minimal preprocessing pipelines for the Human Connectome
Proyecto. NeuroImagen, 80, 105–124. https://doi.org/10.1016/
j.neuroimage.2013.04.127
Goñi,
J., van den heuvel, METRO. PAG., Avena-Koenigsberger, A.,
de Mendizabal, norte. v., Betzel, R. F., Griffa, A., . . . despreciar, oh.
(2014). Resting-brain functional connectivity predicted by ana-
lytic measures of network communication. Actas de la
Academia Nacional de Ciencias, 111(2), 833–838. https://doi.org/
10.1073/pnas.1315529111
Gonzalez-Castillo, J., Saad, z. S., Handwerker, D. A., Inati, S. J.,
Brenowitz, NORTE., & Bandettini, PAG. A. (2012). Whole-brain, tiempo-
locked activation with simple tasks revealed using massive av-
eraging and model-free analysis. Actas del Nacional
Academia de Ciencias, 109(14), 5487–5492.
graton, C., Laumann, t. o., gordon, mi. METRO., Esperemos, B., &
Petersen, S. mi. (2016). Evidence for two independent factors that
modify brain networks to meet task goals. Informes celulares, 17(5),
1276–1288.
Greicius, METRO. D., Krasnow, B., Reiss, A. l., & menón, V. (2003).
Functional connectivity in the resting brain: A network analy-
sis of the default mode hypothesis. Actas del Nacional
Academia de Ciencias, 100(1), 253–258. https://doi.org/10.1073/
pnas.0135058100
Hermundstad, A. METRO., bassett, D. S., Marrón, k. S., Aminoff, mi. METRO.,
Clewett, D., Hombre libre, S., . . . Carlson, j. METRO. (2013). Structural
foundations of resting-state and task-based functional connectiv-
ity in the human brain. Actas de la Academia Nacional de
Ciencias, 110(15), 6169–6174.
Hermundstad, A. METRO., Marrón, k. S., bassett, D. S., Aminoff, mi. METRO.,
Frithsen, A., Johnson, A., . . . Carlson, j. METRO. (2014). Structurally
constrained relationships between cognitive states in the human
cerebro. Biología Computacional PLoS, 10(5), e1003591.
Hogg, R. v., & Ledolter, j. (1987). Engineering statistics. Macmillan.
Ito, T., Kulkarni, k. r., Schultz, D. h., Mill, R. D., Chen, R. h.,
Solomyak, l. I., & Col, METRO. W.. (2017). Cognitive task informa-
tion is transferred between brain regions via resting-state net-
work topology. Comunicaciones de la naturaleza, 8(1), 1027. https://doi.
org/10.1038/s41467-017-01000-w
Jenkinson, METRO., beckman, C. F., Behrens, t. mi. J., lana rica, METRO. w.,
& Herrero, S. METRO. (2012). FSL. NeuroImagen, 62(2), 782–790. https://
doi.org/10.1016/j.neuroimage.2011.09.015
Jeurissen, B., Tournier,
j.
J.-D., Dhollander, T., connelly, A., &
Sijbers,
(2014). Multi-tissue constrained spherical decon-
volution for improved analysis of multi-shell diffusion MRI
datos. NeuroImagen, 103, 411–426. https://doi.org/10.1016/
j.neuroimage.2014.07.061
Khambhati, A. NORTE., Medalla,
j. D., Karuza, mi. A., Thompson-
Schill, S. l., & bassett, D. S. (2018). Subgraphs of functional
brain networks identify dynamical constraints of cognitive con-
controlar. PLOS Computational Biology, 14(7), 1–33. https://doi.org/
10.1371/journal.pcbi.1006234
Krienen, F. METRO., yo, B. T., & Buckner, R. l. (2014). Reconfigurable
task-dependent functional coupling modes cluster around a core
functional architecture. Philosophical Transactions of the Royal
Society of London B: Ciencias Biologicas, 369(1653), 20130526.
marco, D., Harwell, J., Olsen, T., Hodge, METRO., vidrio, METRO., Previo,
Informatics and data mining
F., . . . VanEssen, D.
the Human Connectome Project.
tools and strategies for
Frontiers in Neuroinformatics, 5. https://doi.org/10.3389/fninf.
2011.00004
(2011).
Medalla, j. D., Huang, w., Karuza, mi. A., Kelkar, A., Thompson-
Schill, S. l., Ribeiro, A., & bassett, D. S. (2018). Funcional
alignment with anatomical networks is associated with cognitive
flexibility. Nature Human Behaviour, 2(2), 156.
Miši´c, B., Betzel, R. F., De Reus, METRO. A., van den heuvel, METRO. PAG.,
Berman, METRO. GRAMO., McIntosh, A. r., & despreciar, oh. (2016). Red-
level structure-function relationships in human neocortex. Cere-
bral Cortex, 26(7), 3285–3296.
Osher, D. MI., sajonia, R. r., Koldewyn, K., Gabrieli, j. D., Kanwisher,
NORTE., & Saygin, z. METRO. (2015). Structural connectivity fingerprints
predict cortical selectivity for multiple visual categories across
corteza. Corteza cerebral, 26(4), 1668–1683.
Österreicher, F., & Vajda, I. (2003). A new class of metric diver-
gences on probability spaces and its applicability in statistics.
Annals of the Institute of Statistical Mathematics, 55(3), 639–653.
Fuerza, j. D., Mitra, A., Laumann, t. o., Snyder, A. Z., Schlaggar,
B. l., & Petersen, S. mi. (2014). Methods to detect, characterize,
and remove motion artifact in resting state fMRI. NeuroImagen, 84,
320–341. https://doi.org/10.1016/j.neuroimage.2013.08.048
Raichle, METRO. mi. (2015). The brain’s default mode network. Annual Re-
view of Neuroscience, 38(1), 433–447. https://doi.org/10.1146/
annurev-neuro-071013-014030
Raichle, METRO. MI., MacLeod, A. METRO., Snyder, A. Z., Powers, W.. J.,
Gusnard, D. A., & Shulman, GRAMO. l. (2001). A default mode
of brain function. Proceedings of the National Academy of Sci-
ences, 98(2), 676–682. https://doi.org/10.1073/pnas.98.2.676
Rosvall, METRO., Grönlund, A., Minnhagen, PAG., & Sneppen, k.
(2005).
Searchability of networks. Physical Review E, 72(4), 046117.
Salimi-Khorshidi, GRAMO., Douaud, GRAMO., beckman, C. F., vidrio, METRO. F.,
Griffanti, l., & Herrero, S. METRO. (2014). Automatic denoising of func-
tional MRI data: Combining independent component analysis
and hierarchical fusion of classifiers. NeuroImagen, 90, 449–468.
https://doi.org/10.1016/j.neuroimage.2013.11.046
Saygin, z. METRO., Osher, D. MI., Koldewyn, K., Reynolds, GRAMO., Gabrieli,
j. D., & sajonia, R. R. (2012). Anatomical connectivity patterns pre-
dict face selectivity in the fusiform gyrus. Neurociencia de la naturaleza,
15(2), 321.
Neurociencia en red
473
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cognitive task processing in the human connectome
Schreiber, T., & Schmitz, A. (2000). Surrogate time series. Physica
D: Nonlinear Phenomena, 142(3), 346–382. https://doi.org/10.
1016/S0167-2789(00)00043-9
Schultz, D. h., & Col, METRO. W..
(2016). Higher intelligence is
associated with less task-related brain network reconfiguration.
Revista de neurociencia, 36(33), 8551–8561.
Brillar, j. METRO., bisset, PAG. GRAMO., Campana, PAG. T., Koyejo, o., balseros, j. h.,
Gorgolewski, k. J., . . . Poldrack, R. A. (2016). The dynamics
of functional brain networks: Integrated network states during
desempeño de tareas cognitivas. Neurona, 92(2), 544–554.
Herrero, R. MI., Tournier, J.-D., Calamante, F., & connelly, A. (2012).
Anatomically constrained tractography: Improved diffusion MRI
streamlines tractography through effective use of anatomical
información. NeuroImagen, 62(3), 1924–1938. https://doi.org/10.
1016/j.neuroimage.2012.06.005
Herrero, R. MI., Tournier,
J.-D., Calamante, F., & connelly, A.
(2013). SIFT: Spherical-deconvolution informed filtering of
tractograms. NeuroImagen, 67, 298–312. https://doi.org/10.1016/
j.neuroimage.2012.11.049
Herrero, R. MI., Tournier,
J.-D., Calamante, F., & connelly, A.
(2015). SIFT2: Enabling dense quantitative assessment of
brain white matter connectivity using streamlines tractography.
NeuroImagen, 119, 338–351. https://doi.org/10.1016/j.neuroimage.
2015.06.092
Herrero, S. METRO., beckman, C. F., andersson, J., Auerbach, mi. J.,
Bijsterbosch, J., Douaud, GRAMO., . . . vidrio, METRO. F. (2013). descansando-
state fMRI in the Human Connectome Project. NeuroImagen, 80,
144–168. https://doi.org/10.1016/j.neuroimage.2013.05.039
Sotiropoulos, S. NORTE., Jbabdi, S., Xu, J., andersson, j. l., Moeller,
S., Auerbach, mi. J., . . . Behrens, t. mi. j. (2013). Avances en
diffusion MRI acquisition and processing in the Human Con-
nectome Project. NeuroImagen, 80, 125–143. https://doi.org/10.
1016/j.neuroimage.2013.05.057
despreciar, oh. (2011). The human connectome: A complex network.
Annals of the New York Academy of Sciences, 1224(1), 109–125.
https://doi.org/10.1111/j.1749-6632.2010.05888.x
Stejskal, mi. o., & Tanner, j. mi. (1965). Spin diffusion measurements:
Spin echoes in the presence of a time-dependent field gradient.
Journal of Chemical Physics, 42(1), 288–292. https://doi.org/10.
1063/1.1695690
Tavor, I., jones, oh. PAG., Mars, R. B., Herrero, S. METRO., Behrens, t. MI., &
Jbabdi, S. (2016). Task-free MRI predicts individual differences
in brain activity during task performance. Ciencia, 352(6282),
216–220. https://doi.org/10.1126/science.aad8127
Tononi, GRAMO., despreciar, o., & Edelman, GRAMO. METRO. (1994). A measure for brain
complejidad: Relating functional segregation and integration in the
sistema nervioso. procedimientos de la Academia Nacional de Ciencias,
91(11), 5033–5037. http://www.pnas.org/content/91/11/5033
Tournier, J.-D., Calamante, F., & connelly, A. (2012). MRtrix: Diffu-
sion tractography in crossing fiber regions. International Journal
of Imaging Systems and Technology, 22(1), 53–66. https://doi.
org/10.1002/ima.22005
Tournier, J.-D., Mori, S., & Leemans, A. (2011). Diffusion tensor
imaging and beyond. Resonancia Magnética en Medicina, 65(6),
1532–1556. https://doi.org/10.1002/mrm.22924
Trusina, A., Rosvall, METRO., & Sneppen, k. (2005). Comunicación
boundaries in networks. Physical Review Letters, 94(23), 238701.
Uurbil, K., Xu, J., Auerbach, mi. J., Moeller, S., Vu, A. T., Duarte-
Carvajalino, j. METRO., . . . Yacoub, mi. (2013). Pushing spatial and
temporal resolution for functional and diffusion MRI in the Hu-
man Connectome Project. NeuroImagen, 80, 80–104. https://doi.
org/10.1016/j.neuroimage.2013.05.012
van den heuvel, METRO. PAG., & Hulshoff Pol, h. mi. (2010). Exploring the
brain network: A review on resting-state fMRI functional connec-
actividad. European Neuropsychopharmacology, 20(8), 519–534.
https://doi.org/10.1016/j.euroneuro.2010.03.008
VanEssen, D. C., Herrero, S. METRO., Respeto, D. METRO., Behrens, t. mi. J.,
Yacoub, MI., & Ugurbil, k. (2013). The WU-Minn Human Con-
nectome Project: An overview. NeuroImagen, 80, 62–79. https://
doi.org/10.1016/j.neuroimage.2013.05.041
VanEssen, D. C., Ugurbil, K., Auerbach, MI., Respeto, D., Behrens,
t. mi. J., Bucholz, r., . . . Wu-Minn HCP Consortium (2012).
The Human Connectome Project: A data acquisition perspec-
tivo. NeuroImagen, 62(4), 2222–2231. https://doi.org/10.1016/
j.neuroimage.2012.02.018
Wirsich, J., Perry, A., Ridley, B., Proix, T., Golos, METRO., Bénar, C.,
. . . Guye, METRO. (2016). Whole-brain analytic measures of network
communication reveal increased structure-function correlation
in right temporal lobe epilepsy. NeuroImagen: Clinical, 11, 707–718.
yo, B. t. T., Krienen, F. METRO., Sepulcro, J., Jabonoso, METRO. r., Ejército,
D., Hollinshead, METRO., .
. . Buckner, R. l. (2011). The orga-
nization of the human cerebral cortex estimated by intrinsic
Revista de neurofisiología, 106(3),
conectividad funcional.
1125–1165. https://doi.org/10.1152/jn.00338.2011
Neurociencia en red
474
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
3
2
4
5
5
1
0
9
2
6
0
0
norte
mi
norte
_
a
_
0
0
0
7
2
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3