INVESTIGACIÓN
Enhanced prefrontal functional–structural
networks to support postural control deficits after
traumatic brain injury in a pediatric population
Ibai Diez1, David Drijkoningen2, Sebastiano Stramaglia3,4, Paolo Bonifazi1,7,
Daniele Marinazzo5, Jolien Gooijers2, Stephan P. Swinnen2,6, and Jesus M. Cortes1,7,8
1Biocruces Health Research Institute, Cruces University Hospital, Barakaldo, España.
2KU Lovaina, Movement Control and Neuroplasticity Research Group, Group Biomedical Sciences, Leuve, Bélgica.
3Dipartimento di Fisica, Universita degli Studi di Bari and INFN, Barí, Italia.
4Basque Center for Applied Mathematics (BCAM), Bilbao, España.
5Department of Data Analysis, Faculty of Psychological and Pedagogical Sciences, University of Ghent, Ghent, Bélgica.
6KU Lovaina, Leuven Research Institute for Neuroscience & Disease (LIND), Lovaina, Bélgica.
7Ikerbasque: The Basque Foundation for Science, Bilbao, España.
8Department of Cell Biology and Histology, University of the Basque Country, Leioa, España.
Palabras clave: Traumatic brain injury, Corteza prefrontal, Network reorganization, Resting state,
Functional networks, Structural networks, Brain hierarchical atlas
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
ABSTRACTO
Traumatic brain injury (TBI) affects structural connectivity, triggering the reorganization of
structural–functional circuits in a manner that remains poorly understood. We focus here on
brain network reorganization in relation to postural control deficits after TBI. We enrolled
young participants who had suffered moderate to severe TBI, comparing them to young,
typically developing control participants. TBI patients (but not controls) recruited prefrontal
regions to interact with two separated networks: (1) a subcortical network, including parts of
the motor network, basal ganglia, cerebellum, hippocampus, amygdala, posterior cingulate
gyrus, and precuneus; y (2) a task-positive network, involving regions of the dorsal
attention system, together with dorsolateral and ventrolateral prefrontal regions. Nosotros también
found that the increased prefrontal connectivity in TBI patients was correlated with some
postural control indices, such as the amount of body sway, whereby patients with worse
balance increased their connectivity in frontal regions more strongly. The increased
prefrontal connectivity found in TBI patients may provide the structural scaffolding for
stronger cognitive control of certain behavioral functions, consistent with the observations
that various motor tasks are performed less automatically following TBI and that more
cognitive control is associated with such actions.
RESUMEN DEL AUTOR
Using a new hierarchical atlas whose modules are relevant for both structure and function,
we found increased structural and functional connectivity in prefrontal regions in TBI patients
as compared to controls, in addition to a general pattern of overall decreased connectivity
across the TBI brain. Although this increased prefrontal connectivity reflected interactions
between brain areas when participants were at rest, the enhanced connectivity was found
to be negatively correlated with active behavior such as postural control performance.
Thus our findings, obtained when the brain was at rest, potentially reflect how TBI patients
orchestrate task-related activations to support behavior in everyday life. En particular, nuestro
findings of enhanced connectivity in TBI might help these patients overcome deficits in
cerebellar and subcortical connections, in addition to compensating for deficits when
un acceso abierto
diario
Citación: Diez, I., Drijkoningen, D.,
Stramaglia, S., Bonifazi, PAG., Marinazzo,
D., Gooijers, J., Swinnen, S. PAG., &
Cortes, j. METRO. (2017). Enhanced
prefrontal functional–structural
networks to support postural control
deficits after traumatic brain injury in a
pediatric population. Red
Neurociencia, 1(2), 116–142.
https://doi.org/10.1162/netn_a_00007
DOI:
https://doi.org/10.1162/netn_a_00007
Supporting Information:
Recibió: 15 Septiembre 2016
Aceptado: 28 Enero 2017
Conflicto de intereses: Los autores tienen
declaró que no hay intereses en competencia
existir.
Autor correspondiente:
Jesus M. Cortes
jesus.m.cortes@gmail.com
Editor de manejo:
Olaf Sporns
Derechos de autor: © 2017
Instituto de Tecnología de Massachusetts
Publicado bajo Creative Commons
Atribución 4.0 Internacional
(CC POR 4.0) licencia
La prensa del MIT
Enhanced prefrontal functional–structural networks
interacting with the task-positive network. Por eso, it appears that greater cognitive control is
exerted over certain actions in order to overcome deficits in their automatic processing.
INTRODUCCIÓN
Traumatic brain injury (TBI) involves brain tissue damage resulting from an external mechanical
fuerza, such as rapid head acceleration/deceleration or impact. On a neural level, TBI generally
disrupts functional and structural large-scale brain networks (es decir., the networks of white matter
tracts connecting different brain regions), while on a behavioral level, TBI often triggers var-
ious deficits, including cognitive impairments, motor problems, emotional sequelae, y entonces
adelante, that can persist for years postinjury (Caeyenberghs, Leemans, De Decker, et al., 2012;
Ham & Sharp, 2012; D. h. Herrero & Meaney, 2000). We focus here on deficits in balance
control after TBI, which can last from months to several years after the traumatic impact in
both adults (Guskiewicz, Riemann, Perrin, & Nashner, 1997; McCulloch, Buxton, Hackney,
& Lowers, 2010) and children (Drijkoningen, Caeyenberghs, Vander Linden, et al., 2015;
Drijkoningen, Leunissen, et al., 2015; Katz-Leurer, Rotem, Lewitus, Keren, & Meyer, 2008),
which is psychosocially important because it increases the risk of falling, and thus affects
the patient’s independence (McCulloch, Buxton, Hackney, & Lowers, 2010; Wade, Canning,
Fowler, Felmingham, & Baguley, 1997).
Over the past decades, imaging techniques such as diffusion-weighted imaging (DWI)
and functional magnetic resonance imaging (resonancia magnética funcional) have increased our understanding of the
physiopathology of TBI. En particular, recent advances in MRI techniques have allowed for an
analysis of the injured brain and for correlation of the damages there with behavior from a
network perspective—that is, for exploring the structural and functional connectivity of neu-
ronal networks in vivo (Barbey et al., 2015; Bonnelle et al., 2011; Caeyenberghs et al., 2014;
Caeyenberghs, Leemans, De Decker, et al., 2012; Fagerholm, Hellyer, Scott, Sanguijuela, & Sharp,
2015; Ham & Sharp, 2012; Mäki-Marttunen, Diez, Cortes, Chialvo, & Villarreal, 2013;
Sharp et al., 2011; Sharp, Scott, & Sanguijuela, 2014). Por ejemplo, diffusion-weighted results
have revealed reduced structural connectivity and reduced network efficiency in TBI
patients in relation to poorer cognitive functioning (Bonnelle et al., 2011; Caeyenberghs et al.,
2014; Fagerholm et al., 2015; Kim y cols., 2014) and poorer pediatric balance control
(Caeyenberghs, Leemans, De Decker, et al., 2012). Además, with respect to resting-state
functional connectivity—that is, looking at regional BOLD interactions when the brain is at
(Bonnelle et al., 2012;
rest—multiple studies have reported TBI-induced alterations
Bonnelle et al., 2011; Hillary et al., 2011;
Sharp et al., 2014;
Tarapore et al., 2013), even in cases of mild TBI (Mayer, Mannell, Abadejo, Gasparovic, & yo,
It has been shown, por ejemplo, that TBI patients have increased
2011; Zhou y cols., 2012).
functional connectivity within the default mode network (DMN), as compared to healthy
controls (Hillary et al., 2011; Palacios et al., 2013; Sharp et al., 2011), which possibly acts as
a compensatory mechanism for the loss of structural connections (es decir., axonal injury). Impor-
tantly, TBI-induced changes in resting-state functional connectivity seem to predict the devel-
opment of attention impairments (Bonnelle et al., 2011). Finalmente, combining the information
from structural and functional networks has resulted in better prediction of task-switching
performance in TBI (Caeyenberghs, Leemans, Leunissen, Michiels, & Swinnen, 2013).
Sharp et al., 2011;
Although there is now sufficient evidence that TBI damages the large-scale and emerg-
ing properties of brain structural and functional networks, and that the degree of network
impairment is correlated with behavioral and cognitive deficits (Drijkoningen, Caeyenberghs,
Large-scale functional brain
redes:
Brain networks obtained by
quantifying statistical dependency
between the BOLD signals
corresponding to different brain
regiones, built from functional
magnetic resonance imaging.
Large-scale structural brain networks:
Brain networks obtained by counting
the number of streamlines
connecting different brain regions,
built from diffusion-weighted
imaging.
Resting state:
A brain condition in which the
subject is not asked to perform any
goal-oriented task, and therefore, como
close as possible to doing nothing.
Default mode network:
A brain network that is active when
the patient is at rest, composed of
parts of prefrontal cortex, posterior
cingulate cortex, inferior parietal
lobule, lateral temporal cortex,
hippocampus, and precuneus. Este
network is well-known to alter for
most brain pathologies.
Neurociencia en red
117
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
Leunissen, et al., 2015; Palacios et al., 2013; Zhou y cols., 2012),
the precise pattern of
structural–functional circuit reorganization after TBI is still poorly characterized. Aquí, nosotros
used a novel brain atlas Diez, Bonifazi, et al., 2015 to probe the working hypothesis that when
structural networks are damaged and reorganized as a result of TBI, there is an associated
reorganization of the corresponding functional networks, y viceversa, thus emphasizing
the strong mutual relationship between brain structure and function (Damoiseaux & Greicius,
in addition to pre-
2009; Diez, Bonifazi, et al., 2015; Parque & Friston, 2013). Además,
vious work correlating white matter microstructural
information with balance perfor-
mance postinjury (Caeyenberghs et al., 2010; Caeyenberghs, Leemans, Heitger, et al., 2012;
Drijkoningen, Caeyenberghs, Leunissen, et al., 2015), we aimed here to assess whether this
structure–function network reorganization is in any manner related to critical behavior, semejante
as the postural control deficits after TBI.
MATERIALES Y MÉTODOS
Participantes
The study included a total of 41 young subjects, 14 of whom had incurred a TBI (edad: 13.14 ±
3.25 años; six males and eight females), junto con 27 healthy control subjects who had
developed normally (edad: 15.04 ± 2.26 años; 12 machos, 15 hembras). Group differences for
age and sex yielded, respectivamente, pag = 0.1306 (after a t-test) and p = 0.9226 (after a chi-
squared test), so the TBI and control groups were age- and sex-matched. The TBI patients
had suffered moderate to severe head injury, as defined by the Mayo classification system for
injury severity. This system classifies patients according to the length of posttraumatic amnesia,
loss of consciousness duration, lowest Glasgow Coma Scale score in the first 24 h, and MRI or
computed tomography images, as assessed by a specialized clinical neurologist. Demographic
data for all patients are given in Table 1. Independently of the specific lesions observed during
the acute scan (columna 2 en mesa 2), at the time of the study all 14 TBI patients had diffuse
axonal injury, and all of them had no severe focal lesions (columna 3 en mesa 2). The TBI
Postural control deficits:
Deficits in balance while trying to
maintain a posture against gravity.
Diffuse axonal injury:
A traumatic brain injury class
characterized by widespread lesions
(in comparison to focal lesions) en
white matter tracts.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
ID
T01
T02
T03
T04
T05
T06
T07
T08
T09
T10
T11
T12
T13
Age
8.6
18.1
9.3
16.5
14.2
13.4
19.0
15.6
13.9
8.5
11.4
13.3
16.0
Mesa 1. Demographic data of TBI patients.
Gender (y)
Cause of Injury
Age at Injury (y)
Time Since Injury (y)
GCS/Coma Duration
METRO
F
F
F
F
METRO
F
METRO
METRO
F
METRO
METRO
F
TA
TA
TA
TA
TA
TA
Caer
TA
TA
TA
Sports injury
TA
N/A
7.9
15.6
7.9
7.2
7.7
12.5
12.5
12.5
13.5
7.7
9.8
12.1
N/A
0.7
2.5
1.4
9.3
6.5
0.8
6.5
3.2
0.3
0.8
1.5
1.2
N/A
C: 5 días
C: 5 días
C: 2 semanas
N/A
N/A
N/A
N/A
C: 10 días
GCS: 3
N/A
N/A
N/A
N/A
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
13.8
T14
M = male, F = female; TA = traffic accident, C = coma, GCS = Glasgow Coma Scale score, N/A = Information not
disponible
Object Impact
10.8
N/A
3.0
F
Neurociencia en red
118
Enhanced prefrontal functional–structural networks
Mesa 2. Clinical data of the TBI patients.
ID
T01
T02
Acute MRI Scan Within 24 h After Injury Lesion
Location/Pathology
Subdural hematoma R FL/PL/TL; cortical contusion
R FL/PL; DAI in R FL
Subdural hematoma/hemorrhagic contusion TL/FL;
injuries R FL, thalamus, R cerebral peduncle, l
mesencephalon; cortical and subcortical hemorrhagic
areas in PL/TL
T03
DAI in L TL/FL, R TL/FL/PL
T04
T05
Epidural hematoma R FL/TL; shift midline
NA
T06
Hemorrhagic contusion L TL; brain edema
T07
T08
T09
T10
T11
T12
T13
Subdural hematoma L FL/TL/PL
DAI R TL, internal capsule, supra-orbital R FL, L FL WM
(anterior corona radiata), L middle cerebellar peduncle
DAI FL, TL, L OL (hemorrhagic injury), cerebellum,
CC, external capsule, R globus pallidus, L thalamus,
R cerebral peduncle, R mesencephalon
NA
Contusion L FL/TL; Enlarged, asymmetric ventricle
(temporal horn)
DAI in genu and splenium CC, L FL
NA
T14
Hemorrhagic contusion L FL, atrophy L FL
MRI Scan Analyzed in This Study Lesion Location/Pathology
Hemosiderin deposits: R semiovale center and CC
Small injuries surrounding drain trajectory in RH
(superior frontal gyrus, head nucleus caudatus, crus
anterius of internal capsule, thalamus, and pons)
Contusion: R anterior temporal pole and R orbitofrontal
corteza; Injuries and atrophy in CC (body and splenium);
Atrophy of R pons; Hemosiderin deposits in L cerebellar
hemisferio, R nucleus lentiformis, L/R FL, L/R PL and R PL
Injuries in R medial frontal gyrus.
Atrophy of the cerebellum; Injuries at the level of L FL,
premotor cortex, L/R medial frontal gyrus, cingulate gyrus,
orbitofrontal cortex (L > R); Contusion anterior temporal
pole (R > L); Hemosiderin deposits in CC, L thalamus,
striatum (R > L)
Hemosiderin deposits: several spread out over L/R PL, R
cerebellum, L superior frontal gyrus. Hemociderosis as a
remnant of subdural hemorrhage
Hemosiderin deposits R cerebellar vermis
Atrophy cerebellum; Contusion R FL WM
Hemosiderin deposits: L FL, periventricular WM, body
and genu CC, L thalamus, R external capsule, anterior
TL (L > R), L/R cerebellum; limited atrophy cerebellum
Enlarged fourth ventricle, atrophy of cerebellar vermis,
contusion R cerebellar vermis, hypotrophy of middle
cerebellar peduncle and L pons; contusion L TL;
Hemosiderin deposits R FL, L TL, Vermis
Hemosiderin deposit: splenium CC
Hemosiderin deposits L FL, genu CC
Mild atrophy in cerebellum and cerebrum, más
pronounced atrophy in frontal cortices, enlarged ventricles;
contusion L/R anterior temporal pole and L/R orbitofrontal
corteza. Hemosiderin deposits in cerebellum, R FL
Contusion: L anterior middle frontal gyrus and
L anterior superior frontal gyrus
WM = white matter, RH = right hemisphere, LH = left hemisphere, FL = frontal lobe, TL = temporal lobe, PL = parietal lobe, OL =
occipital lobe, CC = corpus callosum; R = right, L = left.
patients’ mean age at the time of injury was 10 ± 3.45 años, and the average time interval
between the injury and the present MRI was 3.5 ± 3.5 años. Exclusion criteria were based on
preexisting developmental disorders, central neurological disorders, intellectual disabilities,
Neurociencia en red
119
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
Center-of-pressure trajectory:
Spatial trajectory of the application
point of the ground reaction force,
used in postural control studies.
and musculoskeletal disease. Additional exclusion criteria were having an abbreviated injury
score above 2 for the upper or lower limbs, indicating seriously impaired limb function. El
demographic and clinical descriptors of the TBI group are given in Tables 1 y 2.
The study was approved by the ethics committee for biomedical research at KU Leuven, y
the patients were all recruited from several rehabilitation centers in Belgium (principal investi-
gator, Stephan Swinnen). Written informed consent was obtained from either the participants
themselves or the patients’ first-degree relatives, according to the Declaration of Helsinki.
Balance Tests
Balance control was assessed using three protocols from the EquiTest System (NeuroCom
Internacional, Clackamas, Oregón).
This test measures static postural control while subjects are
Sensory organization test (SOT)
standing as still as possible, barefoot, on a movable platform (forceplate) under four sensory
condiciones: (1) eyes open, fixed platform; (2) eyes closed, fixed platform; (3) eyes open with
the platform tilting in response to body sway to prevent the ankles from bending (reduced
somatosensory feedback); y (4) eyes closed, tilting platform. To familiarize the subject with
the test and avoid any initial effect of surprise on the sensory manipulations, we included one
practice trial for each condition prior to completing the actual measurements. Después, cada
condition was repeated three times in a randomized order. Each trial lasted 20 s. We used
an established protocol that had been applied in earlier studies to assess balance control in
both young and older healthy adults: calculating the center-of-pressure (COP) trajectory from
the forceplate recordings (100 Hz) (Van Impe, Coxon, Goble, Doumas, & Swinnen, 2012). A
mean SOT balance score was acquired for each condition from the three trials, a excepción de
trials in which the subject fell. We evaluated the behavioral outcome through the inverse path
length (iPL) of the COP trajectory in order to acquire a SOT balance index in which higher
scores reflected better balance control (and less body sway).
This is a more dynamic test of balance control that involves goal-
Limits-of-stability test (LOS)
directed postural adjustments, in which subjects intentionally displace their center of gravity
(COG) in different directions without stepping, falling, or lifting their heel or toes. At the begin-
ning of each trial, the COG (provided by the Equitest forceplate) was positioned in the center,
as indicated by a representation on a screen in front of the subject. On presentation of a
visual cue and by leaning over in the right direction, the subject had to move the COG
from the center toward one of the radial targets presented on the screen as quickly and accu-
rately as possible. The following eight target directions were assessed: frente, right front, bien,
right back, atrás, left back, izquierda, and left front. After two practice trials, each direction was
assessed once in a random order. The trial was interrupted and repeated if the subject fell
or took a step, and that trial was not analyzed. Directional control (corriente continua) was computed as
the outcome measure reflecting dynamic balance control. Específicamente, the DC (expressed
as a percentage) was calculated as the difference between on-target (in the target direction)
and off-target (extraneous) movimiento, divided by the amount of on-target movement, as fol-
lows: (amount of on-target movement – amount of off-target movement)/(amount of on-target
movimiento) × 100%. Higher scores reflect better DC, and only a straight line toward the target
would result in a score of 100%, with no off-target movements. Finalmente, to produce a single
measure to be correlated with the imaging results, DC scores were averaged across the eight
target directions for further analysis.
Neurociencia en red
120
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
Like the LOS, this is a dynamic test of balance control mea-
Rhythmic weight shift test (RWS)
suring the ability to move the COG rhythmically from right to left, or forward and backward,
between two target positions. Each direction (backward–forward, left–right) was performed at
three different speeds: slow (a pace of 3 s between each target), medio (a pace of 2 s), y
fast (a pace of 1 s). Each combination of speed and direction (a total of six combinations) era
performed in a separate trial of six movement repetitions that were preceded by four practice
repetitions. The trial was interrupted and repeated if the subject fell or took a step. The DC
was calculated as above (similar to LOS), and the DC scores were averaged across directions
and velocities for further analysis.
En resumen, postural control was evaluated by means of three different score indexes: uno
measuring static postural control (iPL-SOT), and two measuring dynamical postural control
(DC-LOS and DC-RWS). The three indexes were used as behavioral outcome to correlate with
the imaging results.
Imaging
MRI acquisition MRI scanning was performed on a Siemens 3-tesla Magnetom Trio MRI scan-
ner with a 12-channel matrix head coil.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
A high-resolution T1 image was acquired with a 3D magnetization-prepared
Anatomical data.
rapid-acquisition gradient echo (MPRAGE): repetition time [TR] = 2,300 EM, tiempo de eco [EL] =
2.98 EM, voxel size = 1 × 1 × 1.1 mm3, slice thickness = 1.1 mm, field of view [FOV] =
256 × 240 mm2, 160 contiguous sagittal slices covering the entire brain and brainstem.
A DTI diffusion-weighted single-shot spin-echo echoplanar
Diffusion tensor imaging (DTI).
imaging sequence was acquired with the following parameters: TR = 8,000 EM, TE = 91 EM,
voxel size = 2.2 × 2.2 × 2.2 mm3, slice thickness = 2.2 mm, FOV = 212 × 212 mm2,
60 contiguous sagittal slices covering the entire brain and brainstem. A diffusion gradient was
applied along 64 noncollinear directions with a b value of 1,000 s/mm2. Además, one set
of images was acquired with no diffusion weighting (b = 0 s/mm2).
Resting-state fMRI time series were acquired over a 10-min ses-
Resting-state functional data.
sion using the following parameters: 200 whole-brain gradient-echo echoplanar images with
TR/TE = 3,000/30 EM, FOV = 230 × 230 mm2, voxel size = 2.5 × 2.5 × 3.1 mm3, 80 ×
80 matrix, slice thickness = 2.8 mm, y 50 sagittal slices, interleaved in descending order.
MRI preprocessing
Diffusion tensor imaging. We applied DTI preprocessing similar to that in previous work
(Alonso-Montes et al., 2015; Amor et al., 2015; Diez, Bonifazi, et al., 2015) using FSL
(FMRIB Software Library, versión 5.0) and the Diffusion Toolkit. Primero, an eddy current cor-
rection was applied in order to overcome the artifacts produced by variation in the direction
of the gradient fields of the MR scanner, together with the artifacts produced by head move-
mentos. To ensure that group differences were not due to differences in motion, the average
En
motion of each subject was used as a covariate of noninterest in the statistical analyses.
particular, the motion of the subject in the scanner was extracted from the transformation
applied by the eddy current correction step from every volume to the reference volume (el
first one, the b = 0 volumen). Próximo, using the corrected data, a local fitting of the diffusion tensor
Neurociencia en red
121
Enhanced prefrontal functional–structural networks
was applied in order to compute the diffusion tensor model for each voxel. Próximo, a fiber assign-
ment by continuous tracking algorithm was applied (Mori, Crain, Chacko, & van Zijl, 1999).
We then computed the transformation from the Montreal Neurological Institute (MNI) espacio
to the individual-subject diffusion space and projected a high-resolution functional partition
a este ultimo, composed of 2,514 regiones de interés (ROI), hereafter named simply regions,
and generated after applying spatially constrained clustering to the functional data using in
Craddock, James, Holtzheimer, Hu, and Mayberg (2012). This allowed for building 2,514 ×
2,514 structural connectivity (CAROLINA DEL SUR) matrices, each per subject, by counting the number of white
matter streamlines connecting all region pairs within the entire 2,514-region dataset. De este modo, el
element matrix (i, j) of SC was given by the streamline number between regions i and j. SC is
a symmetric matrix, in which the connectivity from i to j is equal to that from j to i. Finalmente, nosotros
made the SC matrices binary for the analysis, considering only two possible values: 0 cuando
no streamlines existed between i and j, y 1 when any nonzero number existed between the
two regions i and j.
Resting-state fMRI. We applied resting-state fMRI preprocessing similar to that used in
previous work (Alonso-Montes et al., 2015; Amor et al., 2015; Diez, Bonifazi, et al., 2015;
Diez, Erramuzpe, et al., 2015; Mäki-Marttunen et al., 2013), by using FSL and AFNI (http://
afni.nimh.nih.gov/afni/). Primero, slice-time correction was applied to the fMRI dataset;
next, each volume was aligned to the middle volume to correct for head movement artifacts.
All voxels were then spatially smoothed with a 6-mm full-width-at-half-maximum isotropic
Gaussian kernel, and after intensity normalization, a bandpass filter was applied between 0.01
y 0.08 Hz (Cordes et al., 2001), followed by the removal of linear and quadratic trends. Nosotros
next regressed out the movement time courses, the average cerebrospinal fluid (CSF) señal,
the average white matter signal, and the average global signal. Finalmente, the functional data were
spatially normalized to the MNI152 brain template, with a voxel size of 3 × 3 × 3 mm3. En
addition to head motion correction, we performed scrubbing, by which means all time points
with framewise displacement greater than 0.5 were interpolated by a cubic spline (Yan et al.,
2013). Además, to remove the effect of head movement in the group comparison analysis,
we also used global frame displacement as a covariate of noninterest.
The initial 2,514 regions were
Clustering ROIs into modules by using a new hierarchical brain atlas
grouped into modules using a recently published atlas (Diez, Bonifazi, et al., 2015), en el cual
modules are regions that are functionally coherent (es decir., the dynamics of the voxels belonging
to one module are very similar) y, at the same time, structurally wired (es decir., the voxels
belonging to a given module are interconnected by white matter fibers). Some existing atlases
are purely anatomical or structural (Desikan et al., 2006; Eickhoff et al., 2005; Lancaster et al.,
2000; Tzourio-Mazoyer et al., 2002), and others are purely functional, such as those achieved
after data-driven methods (Craddock et al., 2012). Although obtaining suitable brain partitions
(or atlases) has been studied intensively (Craddock et al., 2013), a lo mejor de nuestro conocimiento
we were the first to propose a brain partition that accounts for modules that are relevant to
both structure and function (Diez, Bonifazi, et al., 2015), which we now implemented in the
present project.
Although full details are given in Diez, Bonifazi, et al. (2015), here we will briefly summa-
rize the hierarchical clustering approach, cual, when applied to a combination of functional
and structural datasets, resulted in a hierarchical tree, or dendrogram, in which nodes were
progressively merged together into M different modules following a nested hierarchy of “simi-
larity” (which reflects both the correlations from the functional data and the numbers of white
Neurociencia en red
122
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
matter streamlines from the structural data). De este modo, cutting the tree at a certain level led to pool-
ing of the initial 2,514 ROIs into a finite number of modules, 1 ≤M≤ 2, 514 (in principle, un
arbitrary value for M could be obtained by varying the depth of the cut). De este modo, to provide some
examples, the highest dendrogram level, m = 1, corresponded to all 2,514 regions belonging
to a single module, coincident with the entire brain, whereas the lowest level, m = 2,514,
corresponded to 2,514 separate modules, all of them composed of a single region.
Diez, Bonifazi, et al. (2015) also showed that the hierarchical brain partition with M = 20
modules was optimal in terms of cross-modularity, an index simultaneously accounting for
three features: (1) the modularity of the structural partition, (2) the modularity of the functional
partition, y (3) similarity between the structural and functional modules. The MATLAB code
to calculate the cross-modality index between structural and functional connectivity matrices
can be downloaded from www.nitrc.org/projects/biocr_hcatlas/.
To compute cross-modularity, we first assessed modularity simply to account for the qual-
ity of the brain partition; eso es, a partition with high modularity has modules highly isolated
from each other—for instance, by maximizing the fraction of intramodule to intermodule con-
nections with respect to randomization.
En particular, we applied Newman’s algorithm to
address modularity (Hombre nuevo, 2004). In addition to modularity, cross-modularity made use of
similarity between the structural and functional modules, which was approached by calculat-
ing Sorensen’s index, a normalized quantity equal to twice the number of common connections
in the two modules, divided by the total number of connections in the two modules.
The entire hierarchical brain partition can be downloaded from www.nitrc.org/
projects/biocr_hcatlas/.
Statistical Analyses
Behavioral data We compared our three balance control scores—iPL-SOT, DC-LOS, y
DC-RWS—between the TBI and healthy control groups by using a two-tailed t-test.
Imaging data
From the modules defined in the hierarchical atlas,
Group differences in structural networks.
structural networks (SN) were assessed by counting all the connections (es decir., streamlines) comenzar-
ing from one module and ending in a different one. Notice that modules can be defined at
any level of the hierarchical tree. We then calculated the module’s connectivity degree (el
total number of connections reaching a module, cual, because SC is a symmetric matrix,
coincides with the total number of connections leaving it).
Próximo, we applied a two-sample t-test using age and average head motion as covariates of
noninterest, to search for significant differences (pag < 0.05). In particular, to test whether the
means of two groups differed, we performed the hypothesis test using a general linear model,
where Y contained the data and X the experimental design variables and confounds. Using
the appropriate contrast (searching for mean group differences while removing the confound
variables), we computed a two-sample unpaired t-test.
To assess the significance of the structural differences, we applied a permutation test by
performing 1,000 random subject-label permutations. We then generated the probability dis-
tribution for these values, which constituted the null hypothesis, since all dependencies were
removed by the shuffling procedure. All regions with p > 0.05 were discarded.
Neurociencia en red
123
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
As a final remark, although the original SC matrices of size 2,514 × 2,514 were binarized,
at the module level we worked with weighted degrees for the group comparison analysis.
To determine group
Group differences in resting-state brain dynamics within individual regions.
differences in the resting-state brain dynamics within each of the M = 20 modules, we first
obtained the time series of the first principal component for each module, chosen as a repre-
sentative for the entire module. Próximo, we compared four different descriptors extracted from
these time series: variance (2nd standardized moment; quantifies fluctuation size), skewness
(3rd standardized moment; identifies extreme brain dynamics in the resting state [Amor et al.,
2015], which measures how much asymmetry a distribution has with respect to its mean),
kurtosis (4th standardized moment; measures the long-tail effect on the data distribution), y
the number of points resulting from the point process analysis (PPA; measured by counting the
number of amplitude peaks in the BOLD signal [Tagliazucchi, Balenzuela, Fraiman, & Chialvo,
2012], and in particular, counting the points with values greater than the mean value of the
time series plus 1 Dakota del Sur). These descriptors were subjected to a two-sample t-test with age and
head motion as covariates in order to evaluate the differences between the TBI and control
Participantes (pag < 0.05).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
n
e
n
_
a
_
0
0
0
0
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
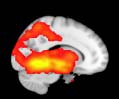
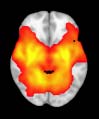
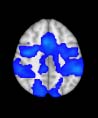
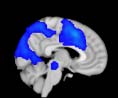
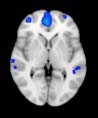
Group differences in functional networks. Motivated by an earlier study (S. M. Smith et al.,
2009), functional networks (FN) were assessed by quantifying the interactions between each
of the M = 20 modules and the rest of the brain (Figure S1). First, within each of the M = 20
modules, we applied a principal component analysis (PCA) so as to reduce the dimensionality
of the data, resulting in 20 components for each of the M = 20 modules. Next we applied an
independent component analysis (ICA), to obtain C = 20 independent time-series components
associated with each of the M = 20 modules. Finally, we applied a general linear model, to
quantify the contribution of each brain voxel to each component (i.e., component’s spatial
map). Then we clustered all of the spatial maps by applying the k-means clustering algorithm,
using the spatial correlation between observations as the similarity measure (Bishop, 2006);
thus, two maps belonged to the same cluster if they showed high spatial correlation. After
applying k-means, the 820 observations per module (41 subjects with C = 20 independent
components each) were grouped into five clusters, which we named the five most represen-
tative clusters (MRCs). Here, the number 5 was chosen after careful inspection, to guarantee
good discrimination between the different clusters. The k-means analysis, in addition to re-
turning the five MRCs, also provided a label for each of the 820 observations—1, 2, 3, 4, or
5—indicating the MRC to which the observation belonged.
As a result of PCA + ICA, we obtained 20 spatial maps per module and subject. Each spatial
map—that is, an observation—was assigned to one of the five MRCs, and this was done for
each module. We took all the spatial maps, per MRC and module, and performed a t-test
comparing the TBI group and the healthy controls.
To correct for multiple comparisons due to a voxel-by-voxel analysis, a statistically sig-
nificant cluster-level family-wise error (FWE) was applied. In particular, a Monte Carlo sim-
ulation (3dClustSim; AFNI, http://afni.nimh.nih.gov) was performed with 10,000
iterations to estimate the probability of false-positive clusters with p < 0.05, corrected with
FWE. We used the new version of the 3dClustSim program (included in AFNI) software, which
corrects for a bug detected by Eklund, Nichols, & Knutsson (2016). After correcting for multi-
ple comparisons, three classes of activity maps for each region were calculated: (1) the average
Network Neuroscience
124
Enhanced prefrontal functional–structural networks
FN in control participants (corresponding to the contrast [1 0 0 0], where the last two zeros
correspond to the movement and age variables); (2) the average FN in TBI patients (contrast
[0 1 0 0]); and (3) the differences between the average FNs of control and TBI participants
(by applying the different contrasts [1 –1 0 0] and [–1 1 0 0], we calculated, respectively,
control > TBI connectivity and TBI > control connectivity).
Note that all of the TBI patients in the MRI session used for this study had diffuse axonal
injury, with no severe focal lesions or regional atrophy, which justified pooling all of the TBI
patients into the same group to be compared with a group of healthy controls.
Relationship between the behavioral and imaging data We used a general linear model that in-
cluded the age and average frame displacement as covariates of noninterest to estimate the
relationship between postural control and the variance/kurtosis/skewness/PPA in every voxel.
Próximo, we used a t-test to assess the association between the postural control scores (iPL-SOT,
DC-LOS, and DC-RWS) and the different fMRI measures, using 3dClustSim with a cluster-based
FWE multiple-comparison correction. Within this region, we used the mask of TBI > control
structural connectivity and correlated the variance/kurtosis/skewness/PPA of voxels within this
region with the three behavioral scores. To assess the association between the postural control
variable and the fMRI variables, we took the region that survived the multiple comparison cor-
rection and plotted the correlation of the corrected variance (es decir., the variance after removing
the effects of age and head motion) and the postural control variable. We performed both
Pearson and Spearman correlational analyses, since the latter are less affected by the presence
of outliers.
RESULTADOS
TBI-Induced Alterations in Postural Control Performance
Alterations in postural control performance were measured through three different tests (es decir.,
SOT, LOS, RWS). Primero, the SOT showed that TBI patients had a smaller inverse path length
in the COP trajectory than did control participants (iPL-SOT: control, 106.72 ± 8.41; TBI,
88.63 ± 28.80; pag = 0.0041; t= 3.054), which reflects that TBI patients had worse balance
(more body sway) than the controls. Similarmente, the RWS test showed that TBI patients performed
more poorly than controls (DC-RWS: control, 83.77 ± 7.01; TBI, 77.11 ± 9.82; pag = 0.0174;
t= 2.4868), confirming poorer dynamic balance among TBI patients. A diferencia de, the LOS test
did not show significant group differences as measured by the dynamic control index (DC-LOS:
control, 83.80 ± 6.75; TBI, 83.46 ± 5.89; pag = 0.8731; t= 0.1608).
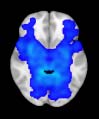
TBI-Induced Alterations in Structural Networks
Alterations in structural networks were assessed by calculating the connectivity degree for each
module in the hierarchical atlas from the intermodule connectivity matrix and performing a
group comparison (after correcting for multiple comparisons by performing random subject-
label permutations). At the level of M = 20 modules (Mesa 3), control participants showed
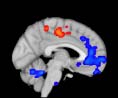
a greater number of connections reaching another module than did TBI patients (Cifra 1).
This suggests that a global decrease in connectivity is associated with TBI. More specifically,
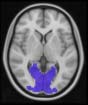
significant differences in connectivity degree were evident within Module 14 ( pag = 0.01, t=
2.64), which included parts of the hippocampus and parahippocampal gyrus, amygdala,
putamen, insula, ventral diencephalon, temporal gyrus, and temporal pole, and Module 20
(pag = 0.003, t= 3.13), which included parts of the cerebellum and parahippocampal gyrus.
See Table 4 for full statistical details.
Neurociencia en red
125
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
Mesa 3. Anatomical description of
(Diez, Bonifazi, et al., 2015) and available to download at www.nitrc.org/projects/biocr_hcatlas/.
the M = 20 modules (along with volumes)
in the hierarchical atlas published recently
Module
(Volumen)
Module 1
(7.26 cm3)
Module 2
(104.36 cm3)
Module 3
(221.18 cm3)
Module 4
(91.48 cm3)
Anatomical Description
Posterior cingulate: posterior area of the cingulate gyrus or callosal convolution. Located above the corpus
callosum, it goes from the frontal lobe back to the temporal uncus and up to the splenium. It belongs to
the default mode network.
Putamen: a round structure located at the base of the telencephalon. It is also one of the basal ganglia
estructuras.
cingulado anterior: anterior frontal region of the cingulate gyrus, initiated above the rostrum of the corpus
callosum.
Rostral pars of middle frontal gyrus: anterior inferior end of the middle frontal gyrus.
Superior parietal gyrus: parietal gyrus located posterior to the postcentral gyrus.
Supramarginal gyrus: region in the parietal lobe encircling the posterior extreme of the Sylvian fissure.
Insula: triangular area of cerebral cortex forming the medial wall of the Sylvian fissure.
Inferior parietal gyrus: parietal gyrus located behind the postcentral gyrus and below the superior parietal
gyrus.
Precentral gyrus: frontal gyrus that defines the anterior boundary of the fissure of Rolando with a mainly
motor function.
Superior frontal gyrus: antero-superior parasagittal frontal gyrus, located anterior to the precentral gyrus.
Paracentral lobule: medial gyrus that connects the pre- and postcentral gyrus.
Precentral gyrus (cf. Module 2)
Postcentral gyrus: Parietal gyrus located between the fissure of Rolando and the postcentral sulcus, cual
has a mainly sensory function.
Precuneus: square brain lobule located before the parieto-occipital sulcus and behind the paracentral
lobule at the medial surface of the brain hemisphere.
Superior frontal gyrus (cf. Module 2).
Superior parietal gyrus (cf. Module 2)
Superior temporal gyrus: temporal gyrus at the lateral surface of the temporal lobe. It is located below
the Sylvian fissure and above the superior temporal sulcus. It belongs to the temporal neocortex.
Supramarginal gyrus (cf. Module 2).
Insula (cf. Module 2)
Cuneus: occipital gyrus between the parieto-occipital sulcus and the calcarine sulcus at the medial surface
of the occipital lobe.
Lateral occipital sulcus: external lateral surface of the occipital lobe close to the occipital lobe, dividing
the external occipital gyrus.
Lingual gyrus: occipital extension of the parahippocampal gyrus at the medial surface of the occipital
lobe.
Pericalcarine cortex: occipital area encircling the calcarine sulcus with a function associated to visual
percepción.
Precuneus (cf. Module 3)
Table continued on next page
Neurociencia en red
126
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
Module
(Volumen)
Module 5
(37.02 cm3)
Module 6
(159.33 cm3)
Module 7
(22.30 cm3)
Module 8
(3.29 cm3)
Module 9
(163.67 cm3)
Mesa 3. continued from previous page.
Anatomical Description
Medial frontal gyrus: frontal gyrus at the lateral surface below the superior frontal gyrus.
Precentral gyrus (cf. Module 2)
Rostral pars of middle frontal gyrus (cf. Module 2)
Cerebellum: posterior part of the rombencephalon made up of the two hemispheres and the central ver-
mal. It is located below the occipital lobe.
Fusiform gyrus: temporal gyrus in the inferior surface between the inferior temporal gyrus and the parahip-
pocampal gyrus. It has two areas, the medial occipito-temporal gyrus and the lateral occipito-temporal
gyrus.
Inferior temporal gyrus: inferior gyrus located in the lateral surface of the temporal lobe, below the infe-
rior temporal sulcus.
Lateral occipital sulcus (cf. Module 4)
Superior parietal gyrus (cf. Module 2)
Thalamus: middle symmetrical structure of the diencephalon with multiple afferent and efferent connec-
ciones, situated around the third ventricle.
Caudate nucleus (symmetrical structure): one of the basic structures of the basal ganglia belonging to the
corpus striatum. It is located at the lateral surface of the lateral ventricles surrounding the thalamus.
Putamen (cf. Module 2)
Pallidum: symmetrical structure within the basal ganglia. Medial diencephalic region of the lenticular
núcleo.
Accumbens nucleus: symmetrical structure located in the ventral region of the corpus striatum, por lo tanto
belonging to the basal ganglia.
Caudate nucleus (cf. Module 7)
Putamen (cf. Module 2)
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
Cerebellum (cf. Module 6)
Caudal middle frontal: frontal gyrus on the lateral surface located below and lateral to the superior frontal
gyrus. This region refers to its most caudal part.
Cingulate isthmus: intersection narrowing between the cingulate and the hippocampal gyrus. It is located
behind and below the splenium of the corpus callosum.
Posterior cingulate (cf. Module 1)
Precuneus (cf. Module 3)
Inferior parietal gyrus (cf. Module 2)
Rostral pars of middle frontal gyrus (cf. Module 2)
Superior frontal gyrus (cf. Module 2)
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Module 10
(103.55 cm3)
cingulado anterior (cf. Module 2)
Inferior parietal gyrus (cf. Module 2)
Orbital gyrus: frontobasal gyrus lateral located to the straight gyrus.
Pars opercularis: opercular part of the inferior frontal gyrus.
Table continued on next page
Neurociencia en red
127
Enhanced prefrontal functional–structural networks
Module
(Volumen)
Module 11
(31.08 cm3)
Module 12
(33.24 cm3)
Module 13
(24.46 cm3)
Module 14
(92.75 cm3)
Mesa 3. continued from previous page.
Anatomical Description
Pars orbitalis: orbital part of the inferior frontal gyrus.
Pars triangularis: inferior part of the inferior frontal gyrus.
cingulado anterior (cf. Module 2)
Rostral pars of middle frontal gyrus (cf. Module 2)
Superior frontal gyrus (cf. Module 2)
Caudate nucleus (cf. Module 7)
Accumbens nucleus (cf. Module 7)
Lateral frontal orbital gyrus: external orbital gyrus, located frontobasal and lateral to the medial or-
bitofrontal gyrus.
Orbital gyrus (cf. Module 10)
cingulado anterior (cf. Module 10)
Inferior parietal gyrus (cf. Module 2)
Inferior temporal gyrus (cf. Module 6)
Lateral frontal orbital gyrus (cf. Module 11)
Pars orbitalis (cf. Module 10)
Pars triangularis (cf. Module 10)
Rostral pars of middle frontal gyrus (cf. Module 2)
Superior frontal gyrus (cf. Module 2)
Caudate nucleus and anterior cingulate (cf. Modules 7 y 2)
Middle frontal gyrus: caudal part of the middle frontal gyrus.
Pars opercularis (cf. Module 10)
Precentral gyrus (cf. Module 2)
Superior frontal gyrus (cf. Module 2)
Thalamus (cf. Module 7)
Hippocampus: symmetrical grey matter structure, located in the medial-temporal region, at the base of
the temporal horn.
Amygdala: grey nuclei located in the temporal uncus, above the temporal ventricular horn. It belongs to
the rhinencephalon.
Putamen (cf. Module 2)
Ventral diencephalon: multiple structures containing the hypothalamus, mammillary tubercle, subthala-
mic nucleus, substantia nigra, red nucleus, geniculate body, optic tract and cerebral peduncles.
Banks of the superior temporal sulcus: Temporal lobe structure between the superior temporal gyrus and
the middle temporal gyrus.
Parahippocampal gyrus: convolution located below the hippocampal sulcus in the temporal mesial
región.
Superior temporal gyrus (cf. Module 3)
Insula (cf. Module 2)
Table continued on next page
Neurociencia en red
128
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
Module
(Volumen)
Anatomical Description
Mesa 3. continued from previous page.
Module 15
(42.96 cm3)
Module 16
(65.58 cm3)
Module 17
(5.29 cm3)
Module 18
(74.39 cm3)
Middle temporal gyrus: gyrus located on the lateral surface of the temporal lobe between the inferior and
superior temporal sulcus.
Temporal pole: anterior end of the temporal lobe.
Thalamus (cf. Module 7)
Putamen (cf. Module 2)
Pallidum (cf. Module 7)
Brainstem: it consists of three parts, the myelencephalon, pons (metencephalon) and midbrain (mesen-
cephalon). It is the main communication route between the brain, spinal cord, and peripheral nerves.
Hippocampus (cf. region14)
Amygdala (cf. Module 14)
Accumbens nucleus (cf. Module 7)
Ventral diencephalon (cf. Module 14)
Orbital gyrus (cf. Module 10)
Insula (cf. Module 2)
Cerebellum (cf. Module 6)
Banks of the superior temporal sulcus (cf. Module 14)
Inferior parietal gyrus (cf. Module 2)
Cingulate isthmus (cf. Module 9)
Middle temporal gyrus (cf. Module 14)
Precuneus (cf. Module 3)
Superior temporal gyrus (cf. Module 3)
Banks of the superior temporal sulcus (cf. Module 14)
Middle temporal gyrus (cf. Module 14)
Hippocampus (cf. Module 14)
Amygdala (cf. Module 14)
Entorhinal cortex: area in the medial-temporal lobe located between the hippocampus and temporal
neocortex.
Fusiform gyrus (cf. Module 6)
Inferior temporal gyrus (cf. Module 6)
Middle temporal gyrus (cf. Module 14)
Parahippocampal gyrus (cf. Module 14)
Temporal pole (cf. Module 14)
Module 19
(28.54 cm3)
Module 20
(34.91 cm3 )
Cerebellum (cf. Module 6)
Brainstem (cf. Module 15)
Cerebellum (cf. Module 6)
Parahippocampal gyrus (cf. Module 14)
Neurociencia en red
129
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
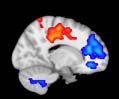
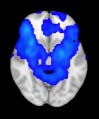
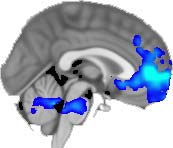
TBI alterations in structural networks revealed by diffusion tensor imaging
a
Brain Hierarchical Atlas
https://www.nitrc.org/projects/biocr_hcatlas/
M=1
Degree for inter-module connectivity
control > TBI
TBI > control
2.2
3.4
2.2
t-statistic
3.4
M=20
M=120
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
b
Control > TBI connectivity (M=20)
TBI > control connectivity (M=120)
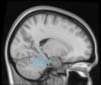
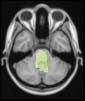
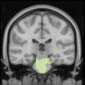
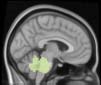
module 14
module 11
module 20
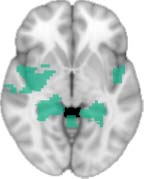
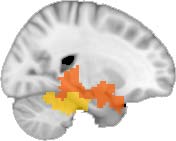
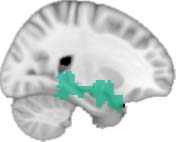
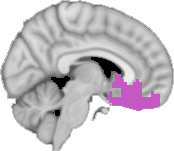
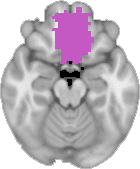
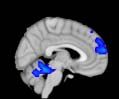
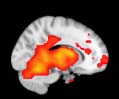
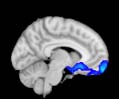
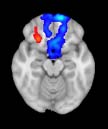
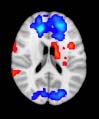
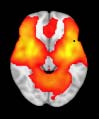
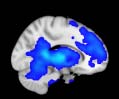
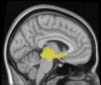
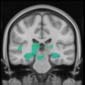
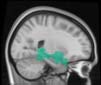
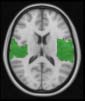
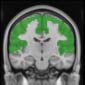
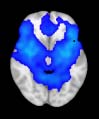
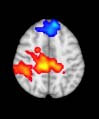
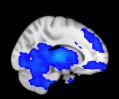
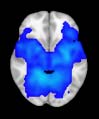
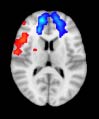
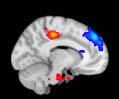
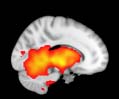
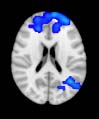
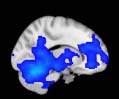
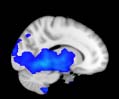
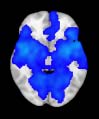
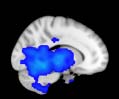
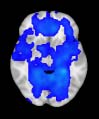
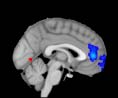
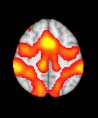
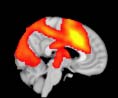
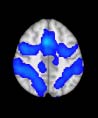
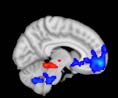
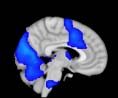
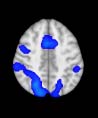
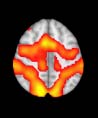
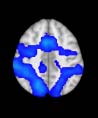
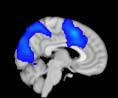
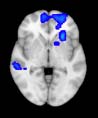
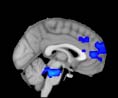
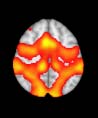
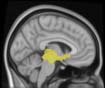
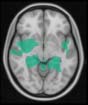
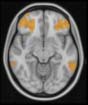
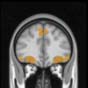
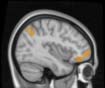
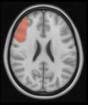
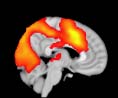
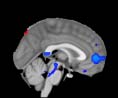
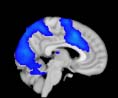
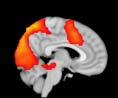
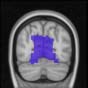
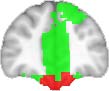
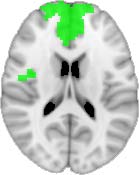
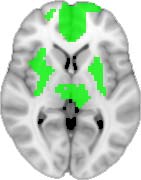
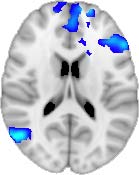
Cifra 1. TBI-induced alterations to structural networks revealed by diffusion tensor imaging.
(a) Hierarchical tree or dendrogram defining a hierarchal brain partition (Diez, Bonifazi, et al., 2015)
in which three different levels of the tree have been emphasized: m = 1, where all brain regions
belong to a single module; m = 20, the optimal brain partition (see Materials and Methods); y
m = 120, the level at which structural connectivity was higher in TBI patients than in controls.
Group differences were calculated on module degree maps derived from the intermodule connec-
tivity matrix and after a two-sample t-test with age and head motion as covariates of noninterest
(pag < 0.05). Multiple comparison corrections were achieved by applying subject-label permuta-
tions, thereby building the a null-hypothesis distribution, since all correlations were removed by
this shuffling. Greater connectivity in controls than in TBI patients (red scale) was found at M = 20,
and at M = 120, TBI > control connectivity was also found (blue scale). Brain maps represent val-
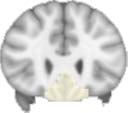
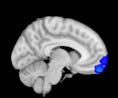
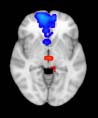
ues of the t-statistic. (b) At M = 20 (left graph), significant control > TBI connectivity was evident in
Module 14 (including parts of the hippocampus and parahippocampal gyrus, amygdala, putamen,
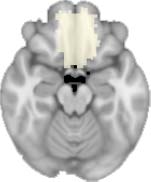
insula, ventral diencephalon, temporal gyrus, and temporal pole) and Module 20 (including parts of
the cerebellum and parahippocampal gyrus). At M = 120 (right graph), TBI > control connectivity
was found within Module 11, including parts of the rectus and superior and inferior frontal orbital
gyri. The module colors are just indicative and coincide with the colors used in Diez, Bonifazi, et al.
(2015), where we first published the hierarchical brain atlas.
At the level of M = 20 in the hierarchical tree, the intermodule connectivity degree was
higher in controls than in patients, indicating that one must go down the hierarchical tree
to find a representation with a higher spatial scale (es decir., the number M of modules increases
lower down on the tree), where TBI connectivity might be higher than control connectivity.
Proceeding in this way, at the level of M = 120 modules we found higher connectivity values
for TBI patients than for controls within Module 11 of the hierarchical atlas (pag = 0.009, t=
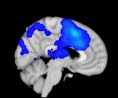
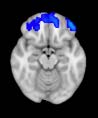
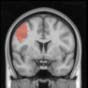
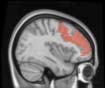
2.75). This module includes parts of the caudate nucleus, nucleus accumbens, lateral frontal
orbital gyrus, orbital gyrus, and anterior cingulate gyrus. De este modo, whereas at the level of M = 20
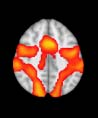
TBI reduced participants’ connectivity relative to controls, at the level of M = 120 modules
(es decir., at a higher spatial scale), prefrontal regions showed an increase in connectivity for TBI as
compared to controls.
Neurociencia en red
130
Enhanced prefrontal functional–structural networks
Mesa 4. TBI versus control differences with respect to structural networks revealed by diffusion tensor imaging (cf. Cifra 1).
At the level of M = 20 modules
Module
Effect Size (Hedges)
t-Statistic
p-Value
Confidence Intervals
14
20
At the level of M = 120 modules
Module 120
2.6351
3.1276
0.0120
0.0033
0.8511
1.0101
0.1539, 0.2990
1.5152, 1.6821
Module 20
t-Statistic
p-Value
Effect Size (Hedges)
Confidence Intervals
1
25
45
70
72
79
84
85
87
88
106
114
117
118
119
11
12
13
6
6
14
14
14
14
14
18
19
20
20
20
–2.7511
2.6874
2.2003
2.6289
2.4432
2.2327
2.7039
2.4728
2.8865
3.4186
2.4281
2.7931
3.3141
2.5526
2.2096
0.0090
0.0105
0.0338
0.0122
0.0192
0.0314
0.0101
0.0179
0.0063
0.0015
0.0199
0.0080
0.0020
0.0147
0.0331
–0.8885
0.8679
0.7106
0.8491
0.7891
0.7211
0.8733
0.7986
0.9323
1.1041
0.7842
0.9021
1.0703
0.8244
0.7136
–1.5544, –0.1883
0.1694, 1.5329
0.0244, 1.3694
0.1521, 1.5131
0.0969, 1.4507
0.0341, 1.3802
0.1743, 1.5384
0.1057, 1.4606
0.2282, 1.6002
0.3840, 1.7815
0.0924, 1.4456
0.2007, 1.5686
0.3535, 1.7457
0.1295, 1.4874
0.0272, 1.3725
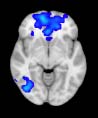
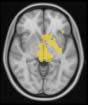
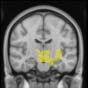
TBI-Induced Alterations in Resting-State Brain Dynamics Within Individual Modules
Alterations in the resting-state brain dynamics within each of the M = 20 modules were as-
sessed by calculating differences in the time series of the first principal component extracted
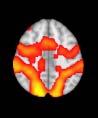
from each module (Cifra 2). The explained data variance across modules varied from 28% a
54%, with a mean value of 38%. En particular, differences emerged with respect to the second
moment (es decir., variance), the third moment (skewness), the fourth moment (kurtosis), y el
number of time-series points that had a value above the mean value of the time series plus 1,
times the standard deviation. After repeating the same procedure for all M = 20 modules of
the hierarchical atlas, significant differences emerged only within Module 11 ( pag = 0.01, t=
–2.55; explicando 48.55% of the data variance of the first principal component), revealing that
the variance of brain dynamics was higher in the TBI than in the control group. Full statistical
details are given in Table 5.
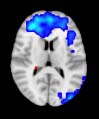
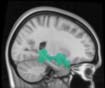
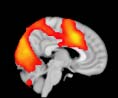
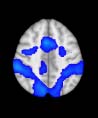
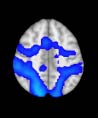
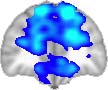
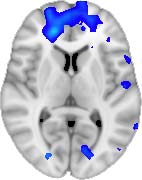
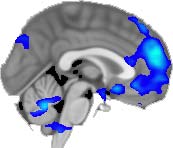
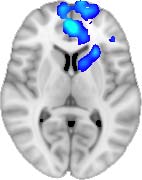
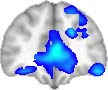
TBI-Induced Alterations in Functional Networks
Functional networks were addressed by quantifying the interaction of each of the M = 20
modules with the rest of the brain. Within each module, we first obtained C = 20 compo-
nents (after PCA followed by ICA), and next we performed spatial regression of the C = 20
components to all the brain voxels, in this way obtaining C = 20 spatial maps for each of the
modules. We grouped all 820 of the observations (41 subjects, each with C = 20 independiente
componentes) per each module into the five MRCs.
After this procedure, it was possible to obtain the same MRC from different modules. En
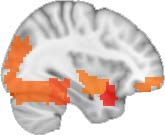
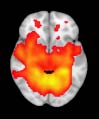
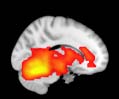
particular, Cifra 3 shows the results associated with one of the MRCs, obtained from the
Neurociencia en red
131
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
a
yo
1
1
mi
tu
d
oh
metro
b
)
mi
C
norte
a
i
r
a
v
(
t
r
q
s
40
35
30
25
20
15
10
5
0
time series of the
first component
significar + 1 Dakota del Sur
significar
*
Control
TBI
Control TBI
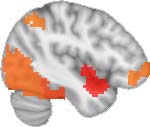
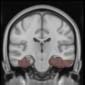
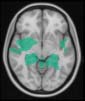
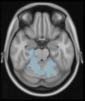
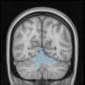
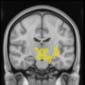
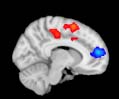
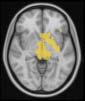
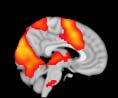
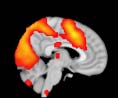
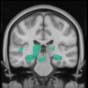
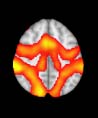
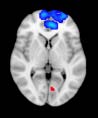
Cifra 2. TBI-induced alterations to brain dynamics within individual modules revealed by
resting-state fMRI. For each of the M = 20 modules in the hierarchical atlas, we extracted the
the vari-
time series of the first principal component and calculated four different descriptors:
ance, skewness, kurtosis, and number of points after the point process analysis (PPA; Materials and
Métodos). (a) Only Module 11 showed differences between the TBI and control groups with respect
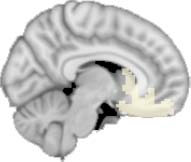
to the variance of the time series of the first principal component. The dashed lines represent the
mean value of the time series, and the solid lines represent the threshold used for the PPA, aquí
equal to the mean + 1 Dakota del Sur. (b) For Module 11, the variance of the first component (plotted here as
its square root—i.e., the standard deviation) differed between TBI and control subjects. In particu-
lar, the fact that the variance was higher in TBI (rojo) than in controls (azul) showed compensation
rather than a deficit. The color of Module 11 (magenta) is just indicative and coincides with the
color used in Diez, Bonifazi, et al. (2015), where we first published the hierarchical brain atlas.
following modules: Module 3 (including parts of the sensory–motor and auditory networks),
Modules 14 y 15 (including parts of the thalamus, hippocampus, amygdala, putamen,
ventral diencephalon, and insula), Module 18 (including parts of the hippocampus and
entorhinal cortex, fusiform gyrus, inferior and middle temporal gyrus, and parahippocampal
Mesa 5. TBI versus control differences with respect to brain dynamics within individual modules
revealed by resting-state fMRI (cf. Cifra 2).
Module
11 (variance)
t-Statistic
–2.5512
p-Value
0.0148
Effect Size (Hedges)
–0.8240
Confidence Intervals
–1.4870, –0.2000
Neurociencia en red
132
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
A subcortical functional network (FN) was found to interact with the modules 3,14,15,18,19 y 20
3
mi
yo
tu
d
oh
metro
4
1
mi
yo
tu
d
oh
metro
5
1
mi
yo
tu
d
oh
metro
8
1
mi
yo
tu
d
oh
metro
9
1
mi
yo
tu
d
oh
metro
0
2
mi
yo
tu
d
oh
metro
2
10
2
10
2
2
4
4
(t-statistic)
2
10
2
10
2
2
4
4
(t-statistic)
Cifra 3. Prefrontal recruitment into a subcortical functional network (FN). Significant brain maps
show different contrasts: columna 1, “FN in control,” with a red bar and corresponding to the contrast
[1 0 0 0] (see Materials and Methods); columna 2, “FN in TBI,” with a blue bar and corresponding
to the contrast [0 1 0 0]; and column 3, “differences,” corresponding to two contrasts, [1 –1 0 0]
y [–1 1 0 0], represented in red (control > TBI activation) and blue (TBI > control activation), re-
spectively. TBI patients (but not control participants) recruited the prefrontal part of the brain when
interacting with a subcortical network (colored in blue at the “differences” column). En todos los casos,
the bar scale represents the strength of significance, measured by the t-statistic values. Contrasts
[1 0 0 0] y [0 1 0 0] define the “subcortical network” (which corresponds to one most represen-
tative cluster including parts of the cerebellum, basal ganglia, thalamus, amygdala, and temporal
poles). This network resulted from the interactions of Module 3 (including parts of the sensory–motor
and auditory networks), Modules 14 y 15 (including parts of the thalamus, hippocampus, amyg-
dala, putamen, ventral diencephalon, and insula), Module 18 (including parts of the hippocampus
and entorhinal cortex, fusiform gyrus, inferior and middle temporal gyrus, and parahippocampal
gyrus), Module 19 (including parts of the cerebellum and brainstem), and Module 20 (incluido
parts of the cerebellum and parahippocampal gyrus). The module colors are just indicative and co-
incide with the colors used in Diez, Bonifazi, et al. (2015), where we first published the hierarchical
brain atlas.
gyrus), Module 19 (including parts of the cerebellum and brainstem), and Module 20 (incluido
parts of the cerebellum and parahippocampal gyrus).
The anatomical representation of this MRC (obtained from Modules 3, 14, 15, 18, 19, y
20) revealed a subcortical network (ver figura 3, “FN in control” and “FN in TBI” columns),
consisting of part of the motor network, basal ganglia, cerebellum, thalamus, parahippocam-
pus, hippocampus, precuneus, amygdala, insula, caudate nucleus, putamen, and pallidum.
Curiosamente, the TBI > control connectivity comparison (obtained with the contrast [–1 1 0
0]) revealed one cluster in the frontal lobe (Cifra 3, column “differences,” colored in blue),
a region including part of the middle frontal and superior orbital gyri, rectus, olfactory lobe,
frontal medial orbital, precuneus, and anterior cingulate gyrus. En otras palabras, the subcortical
network illustrated in Figure 3 with the labels “FN in control” and “FN in TBI” recruited the
prefrontal brain in TBI patients but not in control participants.
A different MRC resembled the task-positive network (Cifra 4, “FN in control” and “FN
in TBI” columns). En particular, this MRC consisted of parts of the cerebellum, lingual gyrus,
fusiform gyrus, inferior occipital gyrus, calcarine sulcus, cuneus, precuneus, superior temporal
Neurociencia en red
133
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
The task-positive functional network (FN) was found to interact with modules 1,4,5,12,14 y 15
1
mi
yo
tu
d
oh
metro
4
mi
yo
tu
d
oh
metro
5
mi
yo
tu
d
oh
metro
2
1
mi
yo
tu
d
oh
metro
4
1
mi
yo
tu
d
oh
metro
5
1
mi
yo
tu
d
oh
metro
2
10 2
10
2
2
4
4
(t-statistic)
2
10 2
10
2
2
4
4
(t-statistic)
Cifra 4. Prefrontal recruitment into the task-positive functional network (FN). These are depicted
como en la figura 3, but the most representative cluster now resembles the task-positive network (see the
labels “FN in control” and “FN in TBI”), which is now resulting from Module 1 (posterior cingu-
late cortex), Module 4 (medial visual cortex), Module 5 (medial frontal gyrus), Module 12 (inferior
parietal and temporal gyrus, lateral frontal orbital gyrus, rostral pars of middle frontal gyrus, y
pars orbitalis and triangularis), and Modules 14 y 15 (subcortical structures). Similar to what is
como se muestra en la figura 3, now TBI patients recruited the prefrontal part of the brain in interaction with the
task-positive network (colored in blue in the “differences” column).
pole, superior motor area, and insula. The MRC resulted from the functional interactions
between Module 1 (including part of the posterior cingulate gyrus), Module 4 (the medial
visual network), Module 5 (including parts of the medial frontal and precentral gyri and rostral
pars of the middle frontal gyrus), Module 12 (including parts of the inferior parietal gyrus,
inferior temporal gyrus, lateral frontal orbital gyrus, pars orbitalis, pars triangularis, rostral pars
of middle frontal gyrus, superior frontal gyrus, caudate nucleus, and anterior cingulate gyrus),
and Modules 14 y 15 (including parts of the thalamus, hippocampus, amygdala, putamen,
ventral diencephalon, and insula).
Although both the control and TBI groups revealed similar task-positive networks (ver
Cifra 4), the TBI connectivity > control connectivity contrast (Cifra 4, colored blue in the
“differences” column) revealed a network in the frontal brain—more specifically, a network
including parts of the frontal medial orbital, anterior cingulate, precuneus, superior frontal,
and angular gyrus. De este modo, like the subcortical network represented in Figure 3, the task-positive
network recruited the prefrontal cortex in TBI patients but not in control participants.
Brain Regions Showing Increased Connectivity in TBI for Both Functional and Structural Networks
Because we observed increased connectivity in TBI patients relative to controls for both func-
tional and structural networks, we decided to take a closer look at these overlapping findings by
superimposing the regions (Cifra 5). With regard to the analysis performed for the structural
redes, a higher degree of connectivity in TBI patients was found in a small subnetwork,
composed of a hub (Figure 5a, plotted in red) connected to other regions (Figure 5a, areas in
verde). The region’s hub belonged to Module 11 in the hierarchical atlas and connected to
Neurociencia en red
134
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
a
(Brain regions with TBI > control connectivity based on structural networks)
t
oh
S
–
l
PAG
i
mi
r
oh
C
s
z
2
1
0
−1
−2
−3
−4
−5
c=−0.865625
(p=0.00013)
Hub (región 11)
Areas structurally
connected to the hub
−1000
1000
3000
5000
7000
Corrected variance
b (Brain regions with TBI > control connectivity based on functional networks)
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
From the interactions with
a subcortical network
From the interactions with
the task-positive network
Cifra 5. Common regions where TBI > control connectivity resulted from both the structural
(a) Structural network compensation. TBI > control structural
and functional network analyses.
connectivity occurred within a subnetwork consisting of a hub (colored in red) connected to other
regiones (colored in green); the hub includes the orbitofrontal and rectus regions and belongs to
Module 11 in the M = 20 hierarchical atlas. All of the following regions are connected to the hub:
frontal superior regions, anterior cingulate gyrus, thalamus, striatum, insula, amygdala, hippocam-
pus and parahippocampus, olfactory cortex, and cerebellum. The corrected variance of the first
principal component of Module 11 was also correlated with postural control measures; here this is
represented by the iPL-SOT score. (b) Functional network compensation. TBI > control functional
conectividad (azul) occurred when interacting with subcortical structures (including the superior
frontal gyrus, superior medial frontal and middle frontal gyri, and anterior cingulate) and the task-
positive network (including the anterior cingulate, the medial frontal and middle orbital gyri, el
superior frontal medial gyrus, and the rectus). For both situations, the spatial maps represent the
functional results of averaging all of the spatial maps with contrast [–1 1 0 0] En figura 3 (subcortical
network) y figura 4 (task-positive network). De este modo, TBI patients (but not control participants) en-
corporated prefrontal parts of the brain into both the subcortical and task-positive networks.
superior frontal regions, anterior cingulate, thalamus, striatum, insula, amygdala, hippocam-
pus and parahippocampus, olfactory lobe, and cerebellum.
With regard to the analysis performed for the functional networks (Figura 5b), two regions
showed increased connectivity in TBI relative to controls: one region interacting with a sub-
cortical network (including superior frontal gyrus, superior medial frontal gyrus and middle
frontal gyrus, and anterior cingulate) and another region interacting with the task-positive
Neurociencia en red
135
Enhanced prefrontal functional–structural networks
network (including anterior cingulate gyrus, frontal medial gyrus, middle orbital gyrus, su-
perior frontal medial gyrus, and rectus).
By looking at both Figures 5a and 5b, we can see an overlap between the maps of increased
structural connectivity (Figure 5a) and the maps corresponding to increased functional con-
conectividad (Figura 5b), which occurred for both the subcortical and task-positive networks.
Relation Between Postural Control and Prefrontal Dynamics at Rest
We found within Module 11 of the hierarchical atlas increased connectivity in both structural
and functional networks for TBI patients in comparison to control participants. Correlational
analyses revealed that the prefrontal activation dynamics of Module 11 at rest (represented by
the corrected variance of the voxel fMRI dynamics) was correlated with the inverse path-length
score of the static SOT (iPL-SOT), giving a Pearson correlation of r = –0.86 (pag = 0.00013) y
a Spearman correlation of s = –0.78 (pag = 0.0026). These results suggest that better balance
performance is associated with decreased dynamic activation in Module 11. Neither DC-RWS
nor DC-LOS was significantly correlated with any of the fMRI measures within Module 11.
DISCUSIÓN
Here we have provided the first evidence that TBI-induced alterations in functional and struc-
tural networks show overlapping results. With respect to both neuronal networks, TBI pa-
tients demonstrated increased prefrontal connectivity, relative to controls. Además, estos
TBI-induced network alterations were associated with changes in balance performance.
TBI-Induced Alterations in Structural Networks
In agreement with previous studies (Gentry, Godersky, & Thompson, 1988; Hulkower, Poliak,
Rosenbaum, Zimmerman, & Lipton, 2013; Zappalà, Thiebaut de Schotten, & Eslinger, 2012),
we found that TBI patients showed reduced structural connectivity (es decir., a smaller degree of
conectividad) for most brain areas, as compared to healthy participants. Más precisamente, nosotros
found a strong decrease in connectivity degree in motor areas, brainstem, cingulate gyrus,
cerebellum, and the temporal poles, areas that are typically associated with the performance
of motor skills and balance control. En efecto, decreased subcortical connectivity, En particular
in the brainstem and cerebellum, was recently associated with postural impairments in TBI
patients (Drijkoningen, Leunissen, et al., 2015), suggesting a possible diffuse pathology across
subcortical structures.
Although for most brain areas we found a lower connectivity degree in TBI patients relative
to controls, we also found a higher connectivity degree in TBI relative to controls exclusively
in the prefrontal cortex. The latter finding, together with the observation that TBI patients have
poorer performance in postural control, may suggest that patients have developed a mechanism
for stronger cognitive control of such motor actions.
TBI-Induced Alterations in Functional Networks
Our approach, by focusing on interactions between modules defined by the hierarchical at-
las while the brain was at rest, revealed that TBI patients incorporated the prefrontal cortex
with a subcortical network. This possibly suggests a mechanism to compensate for TBI-induced
subcortical–cortical axonal disruptions, confirmed by the results of our analysis of structural
redes, showing decreased white matter connectivity between cortical and subcortical path-
maneras. This disconnection is also consistent with gray matter deficits reported in the frontal
Neurociencia en red
136
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
and temporal cortices and cingulate gyrus, as well as within subcortical structures including
the cerebellum (Gale, Baxter, Roundy, & Johnson, 2005; Zappalà et al., 2012).
We also found that TBI patients incorporated the prefrontal cortex with a task-positive net-
trabajar (Fox et al., 2005) employed during the performance of attention-demanding tasks. Este
suggests more cognitive control and less automatic movement in TBI patients than in control
Participantes.
TBI-Induced Alterations in Both Structural and Functional Networks and Their Association
With Behavior
In many studies of brain networks, changes in functional or in structural network connectivity
have often been associated with TBI, yet very few studies have addressed combined effects of
changes in both structural and functional connections. Here we have shown that prefrontal
brain areas in TBI patients increase their structural and functional connectivity as compared to
control participants, and that the resting dynamics of the areas where connectivity increases
are negatively correlated with postural control performance. This may refer to a compensatory
plasticity mechanism that suggests a different mode of balance control—namely, increased
controlled processing or less automatic processing of balance movements. De este modo, this mode
does not represent a successful compensation, whereby increased functional and structural
connectivities would lead to increased balance performance, but rather a mandatory change
in performance mode that is necessary for accomplishing balance tasks.
Previous work has shown increased prefrontal functional connectivity in patients after
TBI (Gooijers et al., 2016; Hillary, Genova, Chiaravalloti, Rypma, & DeLuca, 2006; Rasmussen
et al., 2008), but as far as we know, we have provided the first evidence that when structural
networks are damaged as a result of brain pathology, an associated reorganization of the corre-
sponding functional networks is also established, y viceversa. Además, the prefrontal cor-
tex is the most appropriate locus from which this network reorganization can be orchestrated.
It is well-known that prefrontal areas do not operate in isolation. En particular, Ha sido
widely reported that interactions between the frontal cortex and the basal ganglia play a key
role in movement control (Alexander, Crutcher, &DeLong, 1990; a, 2006; Coxon et al.,
2010; Coxon, Van Impe, Wenderoth, & Swinnen, 2012; Hikosaka & Isoda, 2010; Mink, 1996).
De este modo, the fronto-striato-thalamic circuit, which enables frontal lobe regions to communicate
with the basal ganglia, is involved in a rich spectrum of different functions: motor and oculo-
motor circuits, funciones ejecutivas, social behavior, and motivational states (ver, p.ej., Franco,
Scheres, & sherman, 2007). Además, there is evidence that the reduced connectivity we
have reported in the fronto-striato-thalamic circuit is correlated with reduced subcortical gray
matter volume and task performance after TBI (Leunissen et al., 2014a, 2014b).
Además, it has been shown that white matter connectivity and subcortical gray matter
volume continue to decrease for up to 4 years postinjury (Eierud et al., 2014; Farbota et al.,
2012), which may lead to a reorganization of the prefrontal brain regions to compensate
for the damage to the fronto-striato-thalamic circuit. This potential response to the insult is
in agreement with our findings and with previous results (Leunissen et al., 2014a, 2014b;
Palacios et al., 2013).
Methodological Issues and Current Limitations
We are aware that the clinical population we studied is small (norte = 14) and highly hetero-
geneous, with their time since injury varying from 4 months to 10 años, and their ages ranging
Neurociencia en red
137
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
de 8 a 19 años. Sin embargo, patients with moderate to severe TBI in a pediatric population
are challenging to recruit. Although we recruited additional patients, these patients had focal
brain lesions, and thus their inclusion in our sample would have further increased sample
heterogeneity. This motivated us to limit our cohort to N = 14 patients, all with diffuse brain
injury. Trabajo futuro, involving larger subject cohorts and/or more homogeneous samples, will
be needed to fully address these limitations.
the global signal. Aquí,
Functional connectivity matrices depend crucially on the specific steps used in the pre-
processing pipelines. One step that severely affects connectivity matrices is regressing (or not
regressing) afuera
in agreement with previous work (alonso-
Montes et al., 2015; Amor et al., 2015; Diez, Bonifazi, et al., 2015; Diez, Erramuzpe, et al.,
2015; Mäki-Marttunen et al., 2013; Marinazzo et al., 2014; Stramaglia, Angelini, Cortes, &
Marinazzo, 2015; Stramaglia et al., 2016), we regressed from each individual time series
the global signal, which is well-known to produce more negative correlations in functional
connectivity matrices (Murphy, Hijo, Handwerker, jones, & Bandettini, 2009; Saad et al., 2012).
After we repeated the entire analysis without regressing out the global signal, Cifra 3 no lo hizo
cambiar, but the results for the task-positive network differed from those shown in Figure 4. En
particular, the significance of prefrontal regions’ interaction with the task-positive network did
not survive correction for multiple comparisons (but did appear in the uncorrected data).
To perform group comparison between the SC matrices, we assessed differences in the
module degree statistics, which allowed for localizing brain regions that were connected dif-
ferently in the two groups, but beyond these node-degree group differences, alternative net-
work statistics (based on measures that could go more deeply into the network topology) might
identify further group differences (Brilla, Proporcionó, & bullmore, 2010). Future work should
take this into consideration.
Recent work has suggested that tractography algorithms might produce false-positive con-
nectivity increases in the pathology of TBI (Squarcina, Bertoldo, Ham, Heckemann, & Sharp,
2012), mainly due to the existence of smaller fractional anisotropy y values in different tracts
after TBI, which ultimately might translate into inaccurate tractography (es decir., counting more
streamlines than really exist). We performed a group comparison (FWE, 3dClustSim) of frac-
tional anisotropy values between our groups and found that fractional anisotropy values in
prefrontal brain areas were not significant smaller in the TBI group than in controls (resultados
not shown), thus corroborating that the increased connectivity we found in TBI patients as
compared to controls was not a consequence of this limitation.
The main motivation to use the brain hierarchical atlas was to examine how our different
modules are meaningful for both structure and function. The analysis based on PCA + ICA
postprocessing (Figure S1), only used for Figures 3 y 4, is not specific to the brain atlas we
calculated, but is based on a functional strategy to extract information beyond the average
activity within a region (es decir., the first principal component). Por lo tanto, this strategy will be
generally valid for any other brain partition.
EXPRESIONES DE GRATITUD
I.D. undertook a 2-month lab rotation to visit the laboratories of S.P.S. and D.M. that was
funded by the Health Department of the Basque Government. J.M.C. acknowledges finan-
cial support from Ikerbasque: The Basque Foundation for Science; Grant DPI2016-79874-R
from the Ministerio Economia, Industria y Competitividad (España); and FEDER and Euskampus
Neurociencia en red
138
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
at UPV/EHU. S.P.S. acknowledges financial support from Bizkaia Talent and the European
Commission through COFUND with the research project Brain Aura Mathematical Simulation
(BRAhMS; Grant AYD-000-285). S.P.S. was also supported by FWO Vlaanderen (Levenslijn
G.A114.11 and G.0708.14), the Research Fund of KU Leuven (C16/15/070), and the Inter-
university Attraction Poles program of the Belgian federal government (Belspo, P7/11).
CONTRIBUCIONES DE AUTOR
D.D., J.G., S.P.S. recruited patients and acquired brain imaging and behavioral data; I.D. pre-
processed brain imaging; I.D., J.M.C made the figures; I.D., S.S., P.B., D.M. S.P.S., J.M.C.
developed the methods; D.M., S.P.S., J.M.C. designed the research; S.P.S., J.M.C have equal
last-author contribution; all the authors wrote the manuscript and agreed in its submission.
REFERENCIAS
Alexander, GRAMO. MI., Crutcher, METRO. D., & DeLong, METRO. R. (1990). Basal
ganglia–thalamocortical circuits: Parallel substrates for motor,
oculomotor, “prefrontal” and “limbic” functions. Progress in
Brain Research, 85, 119–146.
Alonso-Montes, C., Diez, I., Remaki, l., Escudero, I., Mateos, B.,
Rosseel, y., . . . Cortes, j. METRO. (2015). Lagged and instantaneous
dynamical influences related to brain structural connectivity.
Fronteras en psicología, 6, 1024.
doi:10.3389/fpsyg.2015.
01024
Amor, t. A., ruso, r., Diez,
. . . Chialvo, D. R.
I., Bharath, PAG., Zirovich, METRO.,
Extreme brain
Stramaglia, S.,
events: Higher-order statistics of brain resting activity and its
relation with structural connectivity. Europhysics Letters, 111(6),
68007. doi:10.1209/0295-5075/111/68007
(2015).
a, A. R. (2006). Cortical and subcortical contributions to stop
signal response inhibition: Role of the subthalamic nucleus.
Revista de neurociencia, 26(9), 2424–2433. doi:10.1523/
JNEUROSCI.4682-05.2006
Barbey, A. K., Belli, A., logan, A., Frotar, r., Zamroziewicz, METRO.,
& Operskalski, j. t. (2015). Network topology and dynamics in
traumatic brain injury. Current Opinion in Behavioral Sciences,
4, 92–102. doi:10.1016/j.cobeha.2015.04.002
obispo, C. METRO.
(2006). Pattern recognition and machine learning.
Nueva York, Nueva York: Saltador.
Bonnelle, v., Ham, t. MI., Sanguijuela, r., Kinnunen, k. METRO., Mehta, METRO. A.,
madera verde, R. J., & Sharp, D. j.
(2012). Salience network
integrity predicts default mode network function after traumatic
brain injury. procedimientos de la Academia Nacional de Ciencias,
109(12), 4690–4695. doi:10.1073/pnas.1113455109
Bonnelle, v., Sanguijuela, r., Kinnunen, k. METRO., Ham, t. MI., beckman,
C. F., De Boissezon, X., . . . Sharp, D. j. (2011). Default mode
network connectivity predicts sustained attention deficits after
traumatic brain injury. Revista de neurociencia, 31(38), 13442–
13451. doi:10.1523/JNEUROSCI.1163-11.2011
Caeyenberghs, K., Leemans, A., De Decker, C., Heitger, METRO.,
Drijkoningen, D., Linden, C. v., . . . Swinnen, S. PAG. (2012). Cerebro
connectivity and postural control in young traumatic brain injury
patients: A diffusion MRI based network analysis. NeuroImagen:
Clinical, 1(1), 106–115. doi:10.1016/j.nicl.2012.09.011
Caeyenberghs, K., Leemans, A., Geurts, METRO., Taymans, T., Linden,
C. v., Smits-Engelsman, B. C. METRO., . . . Swinnen, S. PAG.
(2010).
Brain-behavior relationships in young traumatic brain injury
patients: DTI metrics are highly correlated with postural control.
Mapeo del cerebro humano, 31(7), 992–1002. doi:10.1002/hbm.
20911
Caeyenberghs, K., Leemans, A., Heitger, METRO. h., Leunissen,
I.,
Dhollander, T., Sunaert, S., . . . Swinnen, S. PAG.
(2012). Graph
analysis of functional brain networks for cognitive control of
action in traumatic brain injury. Cerebro, 135(punto. 4), 1293–1307.
doi:10.1093/brain/aws048
Caeyenberghs, K., Leemans, A., Leunissen, I., Gooijers, J., Michiels,
K., Sunaert, S., & Swinnen, S. PAG. (2014). Altered structural net-
works and executive deficits in traumatic brain injury patients.
Brain Structure and Function, 219(1), 193–209. doi:10.1007/
s00429-012-0494-2
Caeyenberghs, K., Leemans, A., Leunissen,
I., Michiels, K., &
(2013). Topological correlations of structural
Swinnen, S. PAG.
and functional networks in patients with traumatic brain injury.
Frontiers in Human Neuroscience, 7, 726. doi:10.3389/fnhum.
2013.00726
Cordes, D., Haughton, V. METRO., Arfanakis, K., Carew, j. D., Turski,
PAG. A., Moritz, C. h., . . . Meyerand, METRO. mi. (2001). Frequencies
contributing to functional connectivity in the cerebral cortex in
“resting-state” data. AJNR. American Journal of Neuroradiology,
22(7), 1326–1333.
Coxon, j. PAG., Goble, D. J., Van Impe, A., De Vos, J., Wenderoth, NORTE.,
& Swinnen, S. PAG. (2010). Reduced basal ganglia function when
elderly switch between coordinated movement patterns. Cere-
bral Cortex, 20(10), 2368–2379. doi:10.1093/cercor/bhp306
Coxon, j. PAG., Van Impe, A., Wenderoth, NORTE., & Swinnen, S. PAG. (2012).
Aging and inhibitory control of action: Cortico-subthalamic
Diario de
connection strength predicts stopping performance.
Neurociencia, 32(24), 8401–8412. doi:10.1523/JNEUROSCI.
6360-11.2012
Neurociencia en red
139
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
Craddock, R. C., James, GRAMO. A., Holtzheimer, PAG. MI., Hu, X. PAG., &
Mayberg, h. S. (2012). A whole brain fMRI atlas generated via
spatially constrained spectral clustering. Mapeo del cerebro humano,
33(8), 1914–1928. doi:10.1002/hbm.21333
Craddock, R. C., Jbabdi, S., yan, C-G., Vogelstein, j. T., Castellanos,
F. X., Di Martino, A., . . . Milham, METRO. PAG. (2013). Imaging human
connectomes at the macroscale. Nature Methods, 10(6), 524–
539. doi:10.1038/nmeth.2482
Damoiseaux, j. S., & Greicius, METRO. D.
(2009). Greater than the
its parts: A review of studies combining structural
sum of
Cerebro
connectivity and resting-state functional connectivity.
Structure and Function, 213(6), 525–533. doi:10.1007/s00429-
009-0208-6
Desikan, R. S., Ségonne, F., pescado, B., quinn, B. T., Dickerson,
B. C., Blacker, D., . . . Killiany, R. j.
(2006). An automated
labeling system for subdividing the human cerebral cortex on
MRI scans into gyral based regions of interest. NeuroImagen,
31(3), 968–980. doi:10.1016/j.neuroimage.2006.01.021
Diez, I., Bonifazi, PAG., Escudero, I., Mateos, B., Muñoz, METRO. A.,
Stramaglia, S., & Cortes, j. METRO.
(2015). A novel brain partition
highlights the modular skeleton shared by structure and func-
ción. Scientific Reports, 5, 10532. doi:10.1038/srep10532
Diez, I., Erramuzpe, A., Escudero, I., Mateos, B., Cabrera, A.,
Marinazzo, D., . . . Alzheimer’s Disease Neuroimaging Initiative.
(2015). Information flow between resting-state networks. Cerebro
Connectivity, 5(9), 554–564. doi:10.1089/brain.2014.0337
Drijkoningen, D., Caeyenberghs, K., Leunissen, I., Vander Linden,
C., Leemans, A., Sunaert, S., . . . Swinnen, S. PAG. (2015). Capacitación-
induced improvements in postural control are accompanied by
alterations in cerebellar white matter in brain injured patients.
NeuroImagen: Clinical, 7, 240–251. doi:10.1016/j.nicl.2014.12.
006
Drijkoningen, D., Caeyenberghs, K., Vander Linden, C., Van Herpe,
K., Duysens, J., & Swinnen, S. PAG. (2015). Associations between
muscle strength asymmetry and impairments in gait and posture
in young brain-injured patients. Journal of Neurotrauma, 32(17),
1324–1332. doi:10.1089/neu.2014.3787
Drijkoningen, D., Leunissen, I., Caeyenberghs, K., Hoogkamer, w.,
Sunaert, S., Duysens, J., & Swinnen, S. PAG.
(2015). Regional
volumes in brain stem and cerebellum are associated with pos-
tural impairments in young brain-injured patients. Human Brain
Cartografía, 36(12), 4897–4909. doi:10.1002/hbm.22958
Eickhoff, S. B., Esteban, k. MI., Mohlberg, h., Grefkes, C., Fink,
GRAMO. r., Amunts, K., & Zilles, k. (2005). A new SPM toolbox for
combining probabilistic cytoarchitectonic maps and functional
imaging data. NeuroImagen, 25(4), 1325–1335. doi:10.1016/
j.neuroimage.2004.12.034
Eierud, C., Craddock, R. C., Fletcher, S., Aulakh, METRO., King-Casas,
B., Kuehl, D., & LaConte, S. METRO. (2014). Neuroimaging after mild
traumatic brain injury: Review and meta-analysis. NeuroImagen.
Clinical, 4, 283–294. doi:10.1016/j.nicl.2013.12.009
Eklund, A., Nichols, t. MI., & Knutsson, h. (2016). Cluster failure:
Why fMRI inferences for spatial extent have inflated false-positive
tarifas. procedimientos de la Academia Nacional de Ciencias, 113(28),
7900–7905. doi:10.1073/pnas.1602413113
Fagerholm, mi. D., Hellyer, PAG. J., Scott, GRAMO., Sanguijuela, r., & Sharp, D. j.
(2015). Disconnection of network hubs and cognitive impair-
ment after traumatic brain injury. Cerebro, 138(punto 6), 1696–1709.
doi:10.1093/brain/awv075
Farbota, k. D. METRO., Sodhi, A., Bendlin, B. B., McLaren, D. GRAMO.,
(2012). Longitudinal
Xu, GRAMO., Rowley, h. A., & Johnson, S. C.
volumetric changes following traumatic brain injury: A tensor-
based morphometry study. Journal of the International Neuro-
psychological
1006–1018. doi:10.1017/
S1355617712000835
Sociedad,
18(6),
Fox, METRO. D., Snyder, A. Z., Vincent, j. l., Corbetta, METRO., VanEssen,
D. C., & Raichle, METRO. mi. (2005). From the cover: The human brain
is intrinsically organized into dynamic, anticorrelated functional
redes. procedimientos de la Academia Nacional de Ciencias,
102(27), 9673–9678. doi:10.1073/pnas.0504136102
Franco, METRO. J., Scheres, A., & sherman, S. j.
(2007). Understand-
ing decision-making deficits in neurological conditions: Insights
from models of natural action selection. Philosophical Trans-
actions of the Royal Society B: Ciencias Biologicas, 362(1485),
1641–1654. doi:10.1098/rstb.2007.2058
Gale, S. D., Baxter, l., Roundy, NORTE., & Johnson, S. C. (2005). Trau-
matic brain injury and grey matter concentration: A preliminary
Journal of Neurology, nuevo-
voxel based morphometry study.
rosurgery, and Psychiatry, 76(7), 984–988. doi:10.1136/jnnp.
2004.036210
Gentry, l. r., Godersky, j. C., & Thompson, B. (1988). Imágenes por resonancia magnética
of head trauma: Review of the distribution and radiopathologic
features of traumatic lesions. American Journal of Roentgenol-
ogia, 150(3), 663–672. doi:10.2214/ajr.150.3.663
Gooijers, J., Beets, I. A. METRO., Albouy, GRAMO., Beeckmans, K., Michiels,
K., Sunaert, S., & Swinnen, S. PAG. (2016). Movement preparation
and execution: Differential functional activation patterns after
traumatic brain injury. Cerebro, 139(9), 2469–2485. doi:10.1093/
brain/aww177
Guskiewicz, k. METRO., Riemann, B. l., Perrin, D. h., & Nashner, l. METRO.
(1997). Alternative approaches to the assessment of mild head
injury in athletes. Medicine and Science in Sports and Exercise,
29(7, Supl.), S213–221.
Ham, t. MI., & Sharp, D. j. (2012). How can investigation of network
function inform rehabilitation after traumatic brain injury? Canalla-
rent Opinion in Neurology, 25(6), 662–669. doi:10.1097/WCO.
0b013e328359488f
Hikosaka, o., & Isoda, METRO.
(2010). Switching from automatic to
controlled behavior: Cortico-basal ganglia mechanisms. Trends
in Cognitive Sciences, 14(4), 154–161. doi:10.1016/j.tics.2010.
01.006
Hillary, F. GRAMO., Genova, h. METRO., Chiaravalloti, norte. D., Rypma, B., &
DeLuca, j.
(2006). Prefrontal modulation of working memory
performance in brain injury and disease. Mapeo del cerebro humano,
27(11), 837–847. doi:10.1002/hbm.20226
Hillary, F. GRAMO., Slocomb, J., Hills, mi. C., Fitzpatrick, norte. METRO., Medalla,
j. D., Wang, J., . . . Wylie, GRAMO. R.
(2011). Changes in resting
connectivity during recovery from severe traumatic brain injury.
International Journal of Psychophysiology, 82(1), 115–123. doi:
10.1016/j.ijpsycho.2011.03.011
Hulkower, METRO. B., Poliak, D. B., Rosenbaum, S. B., Zimmerman,
METRO. MI., & Lipton, METRO. l.
(2013). A decade of DTI in traumatic
brain injury: 10 years and 100 articles later. American Journal of
Neuroradiology, 34(11), 2064–2074. doi:10.3174/ajnr.A3395
Neurociencia en red
140
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
Katz-Leurer, METRO., Rotem, h., Lewitus, h., Keren, o., & Meyer, S.
(2008). Relationship between balance abilities and gait charac-
teristics in children with post-traumatic brain injury. Brain Injury,
22(2), 153–159. doi:10.1080/02699050801895399
kim, J., parker, D., Whyte, J., Hart, T., Pluta, J., Ingalhalikar, METRO., . . .
Verma, R.
(2014). Disrupted structural connectome is associ-
ated with both psychometric and real-world neuropsychological
Journal of the
impairment in diffuse traumatic brain injury.
International Neuropsychological Society, 20(9), 887–896.
doi:10.1017/S1355617714000812
Lancaster, j. l., Wolforff, METRO. GRAMO., Parsons, l. METRO., Liotti, METRO., Freitas,
C. S., Rainey, l., . . . Fox, PAG. t.
(2000). Automated Talairach
atlas labels for functional brain mapping. Human Brain Map-
ping, 10(3), 120–131.
Leunissen, I., Coxon, j. PAG., Caeyenberghs, K., Michiels, K., Sunaert,
S., & Swinnen, S. PAG. (2014a). Subcortical volume analysis in trau-
matic brain injury: The importance of the fronto-striato-thalamic
circuit in task switching. Corteza, 51, 67–81.
doi:10.1016/
j.cortex.2013.10.009
Leunissen, I., Coxon, j. PAG., Caeyenberghs, K., Michiels, K., Sunaert,
S., & Swinnen, S. PAG. (2014b). Task switching in traumatic brain
injury relates to cortico-subcortical integrity. Human Brain Map-
ping, 35(5), 2459–2469. doi:10.1002/hbm.22341
I., Cortes,
Mäki-Marttunen, v., Diez,
j. METRO., Chialvo, D. r., &
Villarreal, METRO.
(2013). Disruption of transfer entropy and inter-
hemispheric brain functional connectivity in patients with dis-
order of consciousness. Frontiers in Neuroinformatics, 7, 24.
doi:10.3389/fninf.2013.00024
Marinazzo, D., Pellicoro, METRO., Wu, GRAMO., Angelini, l., Cortés, j. METRO.,
& Stramaglia, S.
Information transfer and criticality in
the Ising model on the human connectome. PLoS ONE, 9(4),
e93616. doi:10.1371/diario.pone.0093616
(2014).
Mayer, A. r., Mannell, METRO. v., Abadejo, J., Gasparovic, C., & yo, R. A.
(2011). Functional connectivity in mild traumatic brain injury.
Mapeo del cerebro humano, 32(11), 1825–1835. doi:10.1002/hbm.
21151
McCulloch, k. l., Buxton, MI., Hackney, J., & Lowers, S.
(2010).
Balance, atención, and dual-task performance during walking
after brain injury: Associations with falls history. Diario de
Head Trauma Rehabilitation, 25(3), 155–163. doi:10.1097/HTR.
0b013e3181dc82e7
Mink, j. W.. (1996). The basal ganglia: Focused selection and inhi-
bition of competing motor programs. Progress in Neurobiology,
50(4), 381–425.
Mori, S., Crain, B. J., Chacko, V. PAG., & van Zijl, PAG. C. (1999). Three-
dimensional tracking of axonal projections in the brain by mag-
netic resonance imaging. Annals of Neurology, 45(2), 265–269.
Murphy, K., Hijo, R. METRO., Handwerker, D. A., jones, t. B., &
Bandettini, PAG. A. (2009). The impact of global signal regres-
sion on resting state correlations: Are anti-correlated net-
works introduced? NeuroImagen, 44(3), 893–905. doi:10.1016/
j.neuroimage.2008.09.036
Hombre nuevo, METRO. mi. j. (2004). Fast algorithm for detecting community
structure in networks. Physical Review E, 69(6). doi:10.1103/
PhysRevE.69.066133
Palacios, mi. METRO., Sala-Llonch, r., Junque, C., Roig, T., Tormos, j. METRO.,
Bargallo, NORTE., & Vendrell, PAG. (2013). Resting-state functional mag-
netic resonance imaging activity and connectivity and cognitive
outcome in traumatic brain injury. JAMA Neurology, 70(7), 845–
851. doi:10.1001/jamaneurol.2013.38
Parque, H.-J., & Friston, k.
(2013). Structural and functional brain
redes: From connections to cognition. Ciencia, 342(6158),
1238411–1238411. doi:10.1126/science.1238411
Rasmussen, I.-A., Xu, J., Antonsen, I. K., brunner, J., Skandsen, T.,
(2008). Simple dual tasking
Axelson, D. MI., . . . Håberg, A.
recruits prefrontal cortices in chronic severe traumatic brain in-
jury patients, but not in controls. Journal of Neurotrauma, 25(9),
1057–1070. doi:10.1089/neu.2008.0520
Saad, z. S., Gotts, S. J., Murphy, K., Chen, GRAMO., Jo, h. J., Martín, A., &
(2012). Trouble at rest: How correlation patterns
Cox, R. W..
and group differences become distorted after global signal re-
gression. Brain Connectivity, 2(1), 25–32. doi:10.1089/cerebro.
2012.0080
Sharp, D. J., beckman, C. F., madera verde, r., Kinnunen, k. METRO.,
Bonnelle, v., De Boissezon, X., . . . Sanguijuela, R.
(2011). Default
mode network functional and structural connectivity after trau-
matic brain injury. Cerebro, 134(punto 8), 2233–2247. doi:10.1093/
brain/awr175
Sharp, D. J., Scott, GRAMO., & Sanguijuela, R.
(2014). Network dysfunction
after traumatic brain injury. Reseñas de naturaleza: Neurología, 10(3),
156–166. doi:10.1038/nrneurol.2014.15
Herrero, D. h., & Meaney, D. F. (2000). Axonal damage in traumatic
brain injury. The Neuroscientist, 6(6), 483–495. doi:10.1177/
107385840000600611
Herrero, S. METRO., Fox, PAG. T., Molinero, k. l., Glahn, D. C., Fox, PAG. METRO.,
Mackay, C. MI., . . . beckman, C. F.
(2009). Correspondence
of the brain’s functional architecture during activation and rest.
procedimientos de la Academia Nacional de Ciencias, 106(31),
13040–13045. doi:10.1073/pnas.0905267106
Squarcina, l., Bertoldo, A., Ham, t. MI., Heckemann, r., & Sharp,
D. j.
(2012). A robust method for investigating thalamic white
matter tracts after traumatic brain injury. NeuroImagen, 63(2),
779–788. doi:10.1016/j.neuroimage.2012.07.016
Stramaglia, S., Angelini, l., Cortes, j. METRO., & Marinazzo, D. (2015).
Synergy, redundancy and unnormalized Granger causality.
En
Proceedings of the Annual International Conference of the IEEE
Engineering in Medicine and Biology Society 2015 (páginas. 4037–
4040). Piscataway, Nueva Jersey: IEEE Press. doi:10.1109/EMBC.2015.
7319280
Stramaglia, S., Angelini, l., Wu, GRAMO., Cortes, j. METRO., Faes, l., &
(2016). Synergetic and redundant information
Marinazzo, D.
flow detected by unnormalized Granger causality: Application
to resting state fMRI. IEEE Transactions on Biomedical Engineer-
En g, 63, 2518–2524. doi:10.1109/TBME.2016.2559578
Tagliazucchi, MI., Balenzuela, PAG., Fraiman, D., & Chialvo, D. R.
(2012). Criticality in large-scale brain fmri dynamics unveiled
by a novel point process analysis. Frontiers in Physiology, 3, 15.
doi:10.3389/fphys.2012.00015
Tarapore, PAG. MI., Findlay, A. METRO., Lahue, S. C., Sotavento, h., Honma,
(2013). Resting state
S. METRO., Mizuiri, D., . . . Mukherjee, PAG.
magnetoencephalography functional connectivity in traumatic
brain injury. Journal of Neurosurgery, 118(6), 1306–1316. doi:
10.3171/2013.3.JNS12398
Neurociencia en red
141
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Enhanced prefrontal functional–structural networks
Tzourio-Mazoyer, NORTE., Landeau, B., Papathanassiou, D., Crivello, F.,
Etard, o., Delcroix, NORTE., . . . Joliot, METRO.
(2002). Automated ana-
tomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
NeuroImagen, 15(1), 273–289. doi:10.1006/nimg.2001.0978
Van Impe, A., Coxon,
J., Doumas, METRO., &
j. PAG., Goble, D.
Swinnen, S. PAG. (2012). White matter fractional anisotropy predicts
balance performance in older adults. Neurobiology of Aging,
33(9), 1900–1912. doi:10.1016/j.neurobiolaging.2011.06.013
Wade, l. D., Canning, C. GRAMO., Fowler, v., Felmingham, k. l., &
Baguley, I. j. (1997). Changes in postural sway and performance
of functional tasks during rehabilitation after traumatic brain
injury. Archives of Physical Medicine and Rehabilitation, 78(10),
1107–1111.
yan, C.-G., Cheung, B., Kelly, C., Colcombe, S., Craddock, R. C.,
(2013). A comprehensive
Di Martino, A., . . . Milham, METRO. PAG.
assessment of regional variation in the impact of head micro-
movements on functional connectomics. NeuroImagen, 76, 183–
201. doi:10.1016/j.neuroimage.2013.03.004
Brilla, A., Proporcionó, A., & bullmore, mi. t. (2010). Network-based
statistic: Identifying differences in brain networks. NeuroImagen,
53(4), 1197–1207. doi:10.1016/j.neuroimage.2010.06.041
Zappalà, GRAMO., Thiebaut de Schotten, METRO., & Eslinger, PAG. j.
(2012).
Traumatic brain injury and the frontal lobes: What can we
gain with diffusion tensor imaging? Corteza, 48(2), 156–165. doi:
10.1016/j.cortex.2011.06.020
zhou, y., Milham, METRO. PAG., Lui, Y. w., Miles, l., Reaume,
J.,
Sodickson, D. K., . . . Ge, Y. (2012). Default-mode network dis-
ruption in mild traumatic brain injury. Radiology, 265(3), 882–
892. doi:10.1148/radiol.12120748
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
/
1
2
1
1
6
1
0
9
1
9
1
8
norte
mi
norte
_
a
_
0
0
0
0
7
pag
d
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Neurociencia en red
142