Communicated by Stephen José Hanson
Bridging the Gap Between Neurons and Cognition Through
Assemblies of Neurons
Christos H. Papadimitrio
christos@columbia.edu
Columbia University, Nueva York, Nueva York 10027, U.S.A.
Angela D. Friederici
friederici@cbs.mpg.de
Max Planck Institute for Human Cognitive and Brain Sciences, Departamento
of Neuropsychology, D-04303 Leipzig, Alemania
During recent decades, our understanding of the brain has advanced
dramatically at both the cellular and molecular levels and at the cogni-
tive neurofunctional level; sin embargo, a huge gap remains between the
microlevel of physiology and the macrolevel of cognition. We propose
that computational models based on assemblies of neurons can serve
as a blueprint for bridging these two scales. We discuss recently devel-
oped computational models of assemblies that have been demonstrated
to mediate higher cognitive functions such as the processing of simple
oraciones, to be realistically realizable by neural activity, and to possess
general computational power.
1 Introducción
Over the past decades, neuroscientific research at the molecular and cellular
levels has dramatically increased our understanding of the physiology and
function of neurons, as well as their interconnectivity. At the other end of the
escala, cognitive neuroscience has advanced considerably our understand-
ing of the functional neuroanatomy of higher cognitive functions such as
memory and language. The gap between the microlevel of physiology and
the macrolevel of functional neuroanatomy, sin embargo, remains huge. Este
is due not only to the difference in granularity but also to current method-
ological difficulties in experimental measurements at the levels in between.
There is another gap separating the two scales: cognitive imaging stud-
ies are for the most part carried out on human subjects, while neuron-level
neuroscientific experiments typically involve laboratory animals. Sin embargo,
comparative anatomical and electrophysiological studies across different
species provide compelling support for the idea that the computations re-
alized in local cortical networks enabling perceptual, motor and cognitive
functions are remarkably similar (Balaban et al., 2010; douglas & Martín,
Computación neuronal 34, 291–306 (2022)
https://doi.org/10.1162/neco_a_01463
© 2021 Instituto de Tecnología de Massachusetts
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
292
C. Papadimitriou and A. Friederici
2004; Markov & Kennedy, 2013; bullmore & despreciar, 2009). Modeling hu-
man cognitive function based on findings from animal studies is not unjus-
tified (harris, 2005; Friederici & Cantante, 2015); ver, Por ejemplo, the direct
comparison of event-related brain potentials to processing auditory stimuli
between human and nonhuman primates (Milne et al., 2016). It has been ar-
gued (Schomers, Garagnani, & Pulvermüller, 2017) that differences in cog-
nitive abilities between humans and animals, such as language, may be
attributed mainly to differences in connectivity between distal areas.
The need for bridging the gap between these two established approaches
to understanding the brain has been articulated recently by Richard Axel
(2018), who states in an interview, “[necesitamos] a logic for the transformation
of neuronal activity into thought and action.”
What kind of computational model, mediating between the single-cell
level and the cognition level, would fit the bill? Several genres of computa-
tional models, operating between the two scales, have been developed over
the past decades, and in a recent review (Kriegeskorte & douglas, 2018, Higo-
ura 3) they are categorized into three kinds according to the twin criteria of
cognitive fidelity and biological fidelity: (1) models of cognition (Bayesian
and graphic models, reinforcement learning models, and cognitive pro-
gramos, among many others) are good at expressing cognitive functions but
lack in biological plausibility; (2) models from computational neuroscience
can implement, through biologically faithful model neurons—and their cir-
cuits and dynamics—only relatively low-level cognitive functions, como
working memory; y (3), between these two in both axes, deep neural net-
works are capable of certain aspects of intelligent behavior but have been
broadly recognized to violate important biological constraints, such as lo-
cal sparsity, recurrence, lack of supervision, as well as information propaga-
tion from postsynaptic to presynaptic loci (but see Lillicrap, Santoro, Marris,
Akerman, & Hinton, 2020, for a recent argument about the biological plau-
sibility of deep nets). We believe that none of these families of models
promises to bridge the gap—that is, to explicate “the transformation of neu-
ronal activity into thought and action” (Axel 2018).
2 Cognitive Modeling by Assemblies
D. oh. Hebb (1949) proposed that there are persistent and densely intercon-
nected populations of neurons whose near-synchronous firing is cotermi-
nous with the subject thinking of an object, concepto, or word; he called
such populations assemblies. Assemblies of neurons were sought by neu-
roscientists for more than five decades after Hebb’s prediction, until they
were observed experimentally in the animal brain (harris, Csicsvari, Hi-
rase, Dragoi, & Buzsáki, 2003). Novel neuroscientific methods have since
been developed (Carillo-Reid, Han Yang Akrouh, & Yuste, 2019) for de-
tecting, measuring, and manipulating assemblies. Results reported in Ison,
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Bridging Neurons and Cognition Through Assemblies of Neurons
293
Quiroga, and Fried (2015) and elsewhere suggest that in the human medial
−3 (es decir., un
temporal lobe (MTL), a typical assembly has sparsity roughly 10
assembly comprises about one in every 1000 excitatory neurons in the spe-
cific brain area), and consists of roughly 104 neuronas, or a small multiple
of this number. There is a growing consensus (harris, 2005; Buszáki, 2010,
Buszáki, 2019; Eichenbaum, 2018) that assemblies play an important role
in the way the brain works. Since their scale falls squarely between the two
extreme scales of single neurons and cognition discussed above, assemblies
hold definite promise of bridging the gap.
En efecto, computational models of cognition based on assemblies have
been proposed recently. In the 1990s, researchers seeking an explanation
for the continuing firing—after the stimulus was no longer present—of neu-
rons in the prefrontal cortex of animals engaged in a working memory task
(Amit, Brunel, & Tsodyks, 1994; Amit, Fusi, & Yakovlev, 1997; see also the
review by Durstewitz, Seamans, & Sejnowski, 2000) hypothesized that such
a neuron must be part of an assembly encoding the stimulus, an assembly
that was formed by the stimulus through recurrent excitation and Hebbian
plasticity. In two recent papers (Kunze, Peterson, Haueisen, & Knösche,
2017; Kunze, Haueisen, & Knösche, 2019), the analysis of the dynamics
of canonical microcircuits of interacting populations of excitatory and in-
hibitory neurons in cortex reveals three basic modes of these systems, todo
three corresponding to cognitive behaviors: gating (choosing one among
several synaptic input streams), memory (creating a stable assembly), y
priming (lowering a perceptual threshold). The authors also demonstrate
how such microcircuits can facilitate cognitive functions and, En particular,
language understanding (much more on language below).
How are new assemblies created? A picture emerges from the above and
other related works. Synaptic input from an extant assembly—perhaps a
primitive assembly representing a stimulus at the hippocampus (Quiroga,
2016) or in the olfactory bulb (Franks et al., 2011)—causes a population of
neurons in a downstream area to fire. Inhibitory reaction in the downstream
area helps select a few of the neurons in the downstream area to keep fir-
En g. Próximo, continuing afferent synaptic input, combined with the recurrent
excitation from the firing neurons in the downstream area, results in the
firing of a slightly different set of neurons, and then yet another. With con-
tinuing excitation from the upstream assembly, eventually a stable set of
excitatory neurons will be selected in the downstream area, and these will
form the new assembly. The resulting assembly has strong internal synap-
tic connectivity for two reasons, Primero, the way its excitatory neurons were
selected favors neurons with synapses from within the set; furthermore, el
repeated firing increases synaptic strengths through Hebbian plasticity. El
new assembly also has strong synaptic connectivity from the parent assem-
bly, again due to Hebbian plasticity: if the parent assembly fires again, el
newly formed assembly will follow suit.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
294
C. Papadimitriou and A. Friederici
En tono rimbombante, this process can now be repeated. The new assembly, si
excited repeatedly, can create another assembly, in an area further down-
stream, in the exact same manner. This seems to be an important behav-
ior of assemblies, which can be called projection. Projection creates a new
“copy” of an assembly, in a different brain area, that will henceforth fire
every time the parent assembly does, providing a mechanism whereby
memories formed at the hippocampus can be propagated further in the
corteza, thus mediating cognition, reasoning, planificación, idioma, y entonces
adelante.
In a recent paper, Pokorny et al. (2020) verified through the simulation
of a circuit of realistic model spiking neurons with spike timing-dependent
plasticity (STDP) that indeed an assembly established in one brain area
can create a projection in another brain area and that this new assembly
will henceforth behave as described above. Además, the simultaneous
presentation of two parent assemblies in the upstream area will modify,
through plasticity, the corresponding two assemblies in the downstream
área, so they overlap more. Such an increase in overlap, recording the affin-
idad, or association, of the two world objects represented by the two assem-
blies, had been recently noted in experiments in humans (Ison et al., 2015).
By presenting the image of a person known to an individuum (decir, a sibling)
at a particular place (decir, the pyramids) resulted in a neuron that thereto-
fore only responded to images of the pyramids to also respond to images of
the sibling. Presumably this means that two extant assemblies representing
the person and the place were modified in response to the presentation of
the combined image to increase their overlap.
Besides projection and association, a third behavior of assemblies, esta vez
related to language processing, has been recently captured by neurorealis-
tic simulations (Müller, Papadimitrio, Maass, & Legenstein„ 2020). Es
broadly accepted that words such as cat are encoded in a largely perma-
nent manner in a subarea of the left MTL containing the medial temporal
gyrus (MTG) and usually referred to as lexicon. The results in Müller et al.
(2020) suggest that the assembly representing the word cat in the lexicon
can be bound to the syntactic category subject in the context of the sentence,
“The cat chased the dog.” This binding can be accomplished through a so-
phisticated variant of projection, involving the creation of a newly formed
projected copy of the word cat without changing substantially the repre-
sentation of the word in the lexicon. En efecto, in a recent MRI experiment
(Frankland & verde, 2015) two different subareas of the superior tempo-
ral gyrus (STG) were found to exhibit activity in response to the word cat
in the sentences (1) “The cat chased the dog,” where cat is the subject. y
(2) “The dog chased the cat,” where cat is the object of the sentence. Nosotros
hypothesize that this activity reflects the binding operation.
Note that the category subject can be encoded in a sentence by dif-
ferent means depending on the language and sentence type. It could be
encoded by word position in a sentence (the first noun is the subject in
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Bridging Neurons and Cognition Through Assemblies of Neurons
295
English or French), or by case marking in a language like German or Russian
(the nominative case indicates subject). In a passive sentence, the subject can
be marked by the preposition by (“by the man” where the thematic role of
the actor is indicated by the preposition). As already mentioned, brain data
and analysis by Frankland and Greene (2015) suggest that there may be two
distinct brain subareas in the STG to which new assemblies are created for
the subject and the object (or agent and patient) of a sentence during syn-
tactic processing.
Now let us consider how this binding of words to syntactic roles can
be accomplished. Consider two brain regions, one containing the lexicon
(stable representations of words) and another area (possibly located at the
posterior part of the superior temporal lobe, STL) corresponding to the syn-
tactic role of subject, and suppose that there is synaptic connectivity from
the lexicon area to the subject area, and back. If the cat assembly fires (pre-
sumably because the auditory or visual stimulus cat was presented) y
after the syntactic role of “cat” as subject has been decided (a process dis-
cussed below), then synaptic input will excite neurons in the subject area, como
in the beginning of the projection operation. But now, because of the back-
ward connectivity, from subject to lexicon, there will be afferent excitation
to the “cat” assembly. Eventually the projection in the subject area will sta-
bilize. When this happens—and the simulation experiments in Müller et al.
(2020) show that it does happen—the new assembly in the subject area will
have strong synaptic connectivity to and from the “cat” assembly in the
lexicon area: “cat” has been bound to “subject” through a new assembly
created in the subject area. This behavior of assemblies, which can be called
binding or reciprocal projection, was verified in Müller et al. (2020) through a
realistic model of spiking neurons.
In another recent paper, Papadimitrio, Vempala, Mitropolsky, collins,
and Maass (2020) proposed that assemblies are capable of an even broader
repertoire of operations: the three operations project, associate, and bind
that we have seen, plus two more: pattern completion and merge. Patrón
completion entails an assembly firing in response to a few of its neurons (decir,
10% a 20%) firing; this is a consequence of the dense synaptic connectivity
within the assembly. Finalmente, merge is an extension of binding: two assem-
blies, in two different brain areas A and B with synaptic connectivity to a
third area C and back, create, through the firing of their parent assemblies, a
new assembly in area C, which has strong synaptic connectivity to and from
the two original assemblies. Linguists have long believed that human brains
must be capable of such a merge operation, whereby two words (or other
syntactic elements) are combined to form a more complex syntactic element
(Chomsky, 1995). By iterating such merges, arbitrarily large syntactic trees
can be formed, capturing the hierarchical structure of language. De hecho, él
has been verified experimentally (Zaccarella & Friederici, 2015; Zaccarella,
Meyer, Makuuchi, & Friederici, 2017) that on completion of a phrase or a
oración, there is neural activity in the posterior portion of Broca’s area, a
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
296
C. Papadimitriou and A. Friederici
brain area believed to be implicated in syntactic analysis; such activity could
very well be the result of a merge assembly operation.
At the neurophysiological level, one could ask why these particular five
assembly operations—projection, association, pattern completion, binding,
and merge—represent realistic behaviors of neurons and synapses. The ar-
gument is three-fold. Primero. there is mathematical proof in a simplified and
analytically tractable abstract model of spiking neurons in cortex (Papadim-
itriou & Vempala, 2019) that these operations can indeed be simulated by
stylized neurons and synapses. Segundo, as noted above, all of these oper-
ations have been corroborated through simulations in neurorealistic envi-
ambientes (Müller et al., 2020; Pokorny et al. 2020) and by simulations of the
abstract model presented in Papadimitriou et al., 2020). Finalmente, these oper-
ations are consistent with, and appear to provide an explanation for, several
experimental results mentioned above—Frankland and Greene 2015), Ison
et al. (2015), and Zaccarella and Friederici (2015), among many others.
In terms of computation, these five operations, complemented with three
more—inhibition and disinhibition of an area, firing (causing a particular as-
sembly to fire), and readout (identifying the assembly that fired in a par-
ticular area)—comprise a repertoire of operations on assemblies called the
Assembly Calculus. Papadimitriou et al. (2020) proved that the Assembly
Calculus is a full-fledged computational framework capable of arbitrary
computations, akin to programming languages and the Turing machine
(Turing, 1937). Such expressive power is an important desideratum for
a framework hypothesized to underlie general cognitive functions. (Ver
sección 4 for more about the computational power of the Assembly
Calculus.) As we mentioned above, there is evidence (through both math-
ematical proof and simulations) that the primitives of the Assembly Calcu-
lus can be “compiled down” to the level of neurons and synapses, in very
much the same way that a program written in a programming language like
Python and Matlab can be translated into machine code and executed on a
computer. Además, it has been shown that certain simplified cognitive
phenomena related to language processing can be expressed as programs
in the Assembly Calculus. En general, we believe that the Assembly Calcu-
lus appears quite well positioned to bridge the gap between neurons and
cognition.
3 Language Processing through Assemblies
The Assembly Calculus can help explicate the ways in which neuronal ac-
tivity results in high-level cognitive phenomena. Here we focus on one im-
portant high-level cognitive phenomenon: the processing of language.
Why language? Primero, language is well described in its functional sub-
parts by linguistic theory (Berwick & Chomsky, 2015). Segundo, there are a
number of functional and structural experiments investigating and defin-
ing the neuroanatomical basis of language (Friederici, 2011; Fedorenko,
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Bridging Neurons and Cognition Through Assemblies of Neurons
297
Hsieh, Nieto-Castanon, Whitfield-Gabrieli, & Kanwisher, 2010; Pallier,
Devauchelle, & Dehaene, 2011). Tercero, it is understood that language is
grounded on general neurophysiological principles (Friederici & Cantante,
2015).
The neural basis of language has been described as a fronto-temporal
network in the left hemisphere involving Broca’s area in the inferior frontal
gyrus (Brodmann areas (BA) 44 y 45), the medial temporal lobe (MTL)
and middle temporal gyrus (MTG), and the superior temporal gyrus (STG).
These regions are structurally connected by white matter fiber tracts run-
ning dorsally and ventrally between frontal and temporal brain regions.
These fiber tracts connect the language-relevant brain regions functionally
during language processing (see Figure 1A and Friederici, 2011, for a re-
vista). Por ejemplo, the important dorsal fiber called the arcuate fasciculus
connects the STG with the posterior portion of Broca’s area (BA 44), and it is
known to have, in humans, larger fiber density in the left hemisphere than
in the right (Nucifora, Verma, Melhem, Gur, & Gur, 2005); furthermore, es
degree of maturation in children is correlated with accuracy and the speed
of sentence processing (Skeide, Brauer, & Friederici, 2016).
En años recientes, new insights have been gained into the workings of this
network. It is known that soon after the onset of a sentence (read or heard)
and once word forms have been identified, neural activity in the MTL sig-
nals that the representations of those words contained in the sentence are
“looked up” in the lexicon, which is presumed to consist of a large set of
assemblies in the MTL (Xu & Sudhof, 2013; Breitenstein et al., 2005) and re-
lated assemblies in the MTG (Lau, Phillips, & Poeppel, 2008). The assembly
representing each word in the lexicon besides semantic information pre-
sumably encodes information about each word related to its use in lan-
guage, such as its phonological form (cat) and its syntactic category (el
word cat is a noun), which determines the corresponding roles it can play
in the sentence (in the case of noun, subject or object, among others). El
STG appears to assign the role of each word in the sentence under consid-
eration. In a sentence such as “The dog chased the cat,” dog is identified as
the subject, and cat as the object of the sentence, while chased is identified as
the verb. It has been proposed that in the STG, the word dog affects neural
activity in different subareas depending on its syntactic function in the two
sentences “the dog chased the cat” and “the cat chased the dog” (Frankland
& Verde, 2015), implying that new representations of the words are created
during processing. At this processing stage, words are bound to syntactic
roles such as subject, object, and verb. The details of this process at the neu-
rophysiological level need to be investigated in future studies. As a next
step, activity in the subareas of Broca’s area BA 44 and BA 45 suggests that
the buildup of phrases (syntactically licensed parts of the sentence, como
a verb phrase consisting of a verb and a determiner/noun phrase “chased
the cat”; ver Figura 1B) are supported by BA 44. Finalmente, representaciones
of the whole sentence, and presumably its meaning, are constructed in the
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
298
C. Papadimitriou and A. Friederici
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
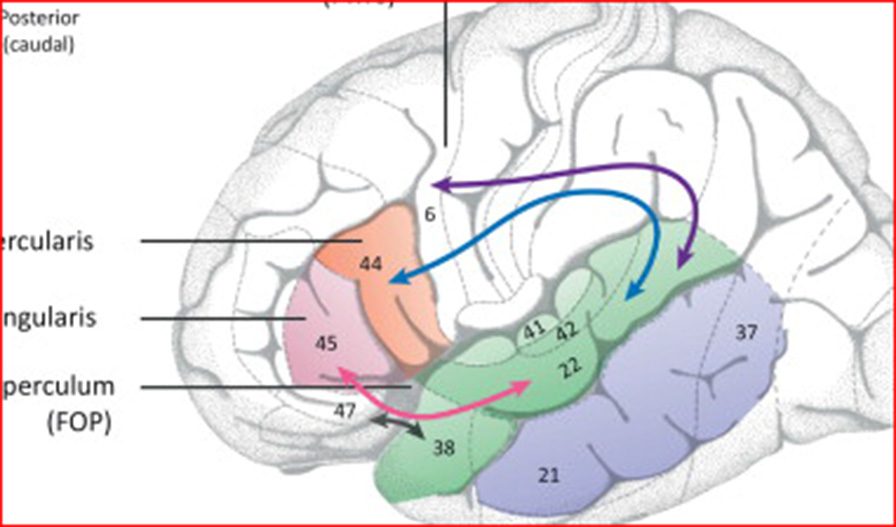
Cifra 1: Syntax of language mediated by assemblies. (A) Brain areas impli-
cated in language processing in the left hemisphere. Medial temporal lobe
(MTL) is located in the medial part of the temporal cortex behind the middle
temporal gyrus (MTG, purple) that is believed to contain the lexicon, representar-
sentations of all words used in language. The superior temporal gyrus (STG,
verde) is believed to be implicated in syntactic manipulation of words; the pars
opercularis of Broca’s area (BA 44, naranja) and the pars triangularis (BA 45, mamá-
genda) are believed to be implicated in the building of more complex syntactic
elementos, such as phrases and sentences. Finalmente, the neural fibers connect the
STG and frontal cortex. Two of them (azul, purple) are located dorsally: uno
of them targets the premotor cortex (BA 6, azul) and the other the posterior
portion of Broca’s area (BA 44, purple). Two other fiber tracts (rojo, negro) son
located ventrally, with the red one targeting the anterior portion of Broca’s area
(BA 45). BA stands for Brodmann area according to the cytoarchitectonic classi-
fication of Brodmann (1909). (B) The syntax tree is a tree-like structure capturing
the underlying structure, or syntax, of a sentence. The top node represents the
whole sentence (S), while the next lower node represents the verb phrase (vicepresidente)—
the part of the sentence describing what the subject does or is. Two nodes rep-
resent the two noun phrases (notario público) corresponding to the subject (izquierda) y el
object (bien) of the sentence. Each of these noun phrases consists of the deter-
miner the and a noun. Identifying the syntax tree of a sentence (or “parsing the
Bridging Neurons and Cognition Through Assemblies of Neurons
299
interplay between the temporal cortex and Broca’s area under the support
of BA 45.
This narrative identifies rather convincingly a very specific brain net-
work for the syntactic analysis of language, cual, at least according to the
“syntax first” school of thought in language processing, is the first step to
comprensión. Sin embargo, this network cannot be static. The words in the
sentence must somehow be “copied” from the lexicon in MTL to the in-
ferior frontal gyrus to determine their syntactic role in the sentence, entonces
bound to the appropriate subarea of STG corresponding to this role. También,
the representations of phrases and sentences in BA 44 and BA 45 must be
created through communication with the representations of their subparts
and presumably remain in communication with them after the new repre-
sentations are formed. Such communication between brain regions is pos-
sible via the brain’s white matter connections: the long-range connections
between the language-related regions in the frontal and temporal cortex
(Catani, jones, & Ffytche, 2005; Friederici, 2009; Saur et al., 2008), también
as short-range connections within the frontal cortex (Makuuchi, Bahlmann,
anwander, & Friederici, 2009) and the temporal cortex (Upadhyay et al..
sentence”) is considered the first important step toward comprehending the
oración. (C) A proposed dynamic architecture involving assemblies and as-
sembly operations in the left temporal and frontal lobes for performing syntac-
tic analysis of language. The colors of the domains of this architecture (cajas)
match the colors of areas in panel A. White circles in these boxes are assem-
blies in the corresponding areas. Starting from below, assemblies representing
the presented (through audio or text) words are identified in the lexicon. Su
syntactic category (verb, noun) and their syntactic role (verb, sujeto, object)
are determined first. For verbs, this syntactic role is often readily available in
the word’s representation; distinguishing between object and subject is subtle
and relies on word order (Inglés, Francés) or case (Russian, Alemán). Once the
syntactic roles have been determined, the three assemblies representing, en esto
ejemplo, CAT, CHASED, and DOG, are projected through the bind assembly
operación (red arrows), each to the appropriate subarea of the STG, creating
three new assemblies there. Próximo, a merge operation creates an assembly in BA
44 representing the verb phrase. Finalmente, a second merge operation joins the sub-
ject with the verb phrase to create the sentence. These operations create a more
rudimentary parse tree than panel B, which encodes the syntactic analysis of the
oración. (D) The Parser in Mitropolsky et al. (in press) implements a different,
nongrammatical formalism of syntax called a dependency graph. For the same
oración, the Parser would generate the dependency graph shown, en el cual
pairs of words are connected by directed edges (in red), each labeled by the
kind of dependency connecting the two words (the labels used here are “verb,"
“subject,” “object,” and “determiner”). The root dependency always points to
the principal verb of the sentence.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
300
C. Papadimitriou and A. Friederici
2008). But precisely how is this communication between cell populations
in different brain areas established and carried out? The second author of
this view predicted (Friederici, 2017, published almost three years before
Papadimitriou et al., 2020) “that, for language, there are what I call mirror
cell ensembles through which two distinct brain regions communicate with
each other.”
Assemblies and assembly operations provide precisely the apparatus
that is needed here (this apparatus is illustrated in Figure 1C). An assembly
representing a word in the MTL can be copied via the projection, or binding,
operation to the STG and bound to the appropriate subarea corresponding
to its grammatical role in the sentence: sujeto, verb, or object. The impor-
tant step of the identification of these grammatical roles, which appears to
be happening in the inferior frontal gyrus together with the STG, needs fur-
ther investigation (see Figure 1C for a brief discussion). An assembly repre-
senting the verb phrase can be formed by a merge operation of the verb and
the object assemblies in BA 44. Finalmente, another merge of the subject assem-
bly and the verb phrase assembly will form a new assembly representing
the whole sentence. This completes the syntactic analysis of the sentence.
Along the way, this process actually builds a “syntactic scaffold” of the sen-
tence, a simplified syntactic tree of the sentence with three leaves and two
nodos internos (the structure defined by the blue double arrows in Figure
1C), which can be used as a platform for understanding the precise meaning
of the sentence, as well as for, Por ejemplo, further processing, repetition,
and memorization.
In a recent paper, Mitropolsky, collins, and Papadimitriou (in press) de-
scribe a program written in an enhanced version of the Assembly Calculus
that successfully parses (es decir., identifies the syntactic structure of) reasonably
complex English sentences such as, “This morning, the ferocious dog from
the next house briefly chased your cat.” The program, called Parser, obras
exclusively by the spiking of artificial neurons through the commands of
the Assembly Calculus, with two bespoke enhancements: (1) there are com-
mands for inhibiting and disinhibiting neural fibers (synapse bundles con-
necting different brain areas), instead of whole areas only, y (2) it is as-
sumed that each word assembly in the lexicon is equipped with inhibitory
neurons that, upon the firing of the assembly, inhibit or disinhibit the ap-
propriate fibers. This latter capability enables the Parser to correctly identify
the syntactic role of words, based exclusively on the word’s part of speech
encoded in the (dis)inhibition actions, and the current state of the system.
Besides the lexicon, specific brain areas are employed, named, Por ejemplo,
“verb,” “subject,” and “adverb,” where the words in the sentence are pro-
jected, while the inhibition or disinhibition of fibers prepares each word for
projection to the correct area. The output of the Parser is not a syntactic tree
as described in the previous paragraph and Figure 1C, but a different ob-
ject also capturing the syntactic structure of the sentence, called dependency
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Bridging Neurons and Cognition Through Assemblies of Neurons
301
graph. Obviamente, other languages will require different uses of this capa-
habilidad; in Mitropolsky et al. (in press), a toy Russian parser is also described
and implemented.
The Parser of Mitropolsky et al. (in press) is an important step toward
demonstrating that the Assembly Calculus can bridge the gap between the
cognitive and the neuronal level in the domain of language, y especialmente
syntactic analysis. But it does not come close to covering all of language.
One important aspect of syntax is recursion and center embedding—the abil-
ity of sentences to embed other sentences as in, “The dog who chased the
cat who scratched the baby is back.” Many consider center embedding to be
the hallmark of language and the Parser does not currently handle this fea-
tura. A brain area can support many assemblies, and presumably the verb
área, Por ejemplo, can contain assemblies corresponding to the verbs of all
sentences in the embedding hierarchy simultaneously; sin embargo, el preciso
implementation of center embedding is left in Mitropolsky et al. (in press)
as an open challenge.
4 Conclusion and Open Questions
We have presented some evidence that assemblies of neurons can be a use-
ful conceptual, even mathematical and computational, tool for bridging the
forbidding gap between neurons and cognition. But of course, even in the
language domain where some progress is being made, we still do not know
the answers to most important questions. World languages rely on a stun-
ning diversity of features to encode a word’s grammatical role. How can
the same brain circuit adapt to a specific encoding, from among all these
very different versions, in the infant brain? Recent brain data indicate that
the language-specific cues, and the resulting processing demands, lead to
language-specific modulation of the fiber tracts connecting the different
brain areas (Goucha, anwander, & Friederici, 2015). Beyond syntax, how is
the meaning of a sentence—its semantics—unraveled in the human brain?
And how are sentences combined to form stories and discourses?
The use of the Assembly Calculus for tackling language processing raises
a theoretical question. Many decades ago, Noam Chomsky (1956) propuesto
his influential hierarchy of languages for the purpose of characterizing nat-
ural language processing problems: regular, context-free, context-sensitive,
and recursively enumerable languages. De estos, the last is far too strong,
containing uncomputable problems. The question arises,1 Is the Assembly
Calculus computationally powerful enough to encompass the first three
levels of the Chomsky hierarchy? The Assembly Calculus is known to be
capable of simulating arbitrary computations with O(s−1) espacio (see the SI
1
Many thanks to an anonymous reviewer for bringing this up.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
302
C. Papadimitriou and A. Friederici
section of Papadimitriou et al. 2020), where s is the sparsity of assemblies
−3. Por lo tanto, what is known about the complex-
believed to be about 10
ity of these three levels (Papadimitrio, 1995) implies that regular, contexto-
gratis, and context-sensitive features of language can be handled through
the Assembly Calculus for sentences containing at least a few dozen
palabras.
At a more general level, let us revisit briefly the hypothesis that opera-
tions on assemblies underlie cognitive processes. According to all evidence,
it is highly unlikely that the brain works in the pristine mathematical, si
highly random, manner of the assembly operations, as described in Pa-
padimitriou et al. (2020) and sketched here. The interesting question may
rather be, Are these operations useful abstractions of the actual neural pro-
cesses underlying cognition, including the processing of language? Más
work is needed to pursue this question.
Finalmente, can assembly-based mechanisms such as the Assembly Calculus
be demonstrated through simulations to be capable of implementing fur-
ther cognitive functions beyond language (a domain in which very partial
progress has already been made)? Two cognitive tasks that come to mind as
interesting candidates are planning and reasoning. Extending the method-
ology developed for handling language to these and other realms of cogni-
tion seems both interesting and challenging.
Expresiones de gratitud
We are grateful to Thomas R. Knösche for valuable feedback on a draft of
this view and to Stephen Jose´Hanson for a number of constructive com-
mentos. The research of C. h. PAG. was supported by NSF Awards CCF1763970
and CCF1910700, by a research contract with Softbank, and by a grant
from Columbia’s Center for AI Technology. The research of A. F. was sup-
ported by the Deutsche Forschungsgemeinschaft, grants FR519/22-1 within
SPP2041.
Referencias
Amit, D. J., Brunel, NORTE., & Tsodyks, METRO. V. (1994). Correlations of cortical Hebbian
reverberations: Theory versus experiment. Revista de neurociencia, 14, 6435–6445.
10.1523/JNEUROSCI.14-11-06435.1994, PubMed: 7965048
Amit, D. J., Fusi, S., & Yakovlev, V. (1997). Paradigmatic working memory (attractor)
cell in IT cortex. Computación neuronal, 9, 1071–1092. 10.1162/neco.1997.9.5.1071,
PubMed: 9188192
Axel, R. (2018). Richard Axel. Neurona, 99(6), 1110–1112.
Balaban, MI., Edelman, S., Grillner, S., Grodzinski, Ud., jarvis, mi. D., Aquél, j. h., . . .
Pipa, GRAMO., (2010). Evolution of dynamic coordination. In C. Von der Malsburg,
W.. A. Phelps, & W.. Cantante (Editores.), Dynamic coordination in the brain. From neurons
to mind (páginas. 59–82). Cambridge, MAMÁ: MIT Press and FIAS.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Bridging Neurons and Cognition Through Assemblies of Neurons
303
Berwick, R. C., & Chomsky, norte. (2015). Why only us: Language and evolution. Leva-
puente, MAMÁ: CON prensa.
Breitenstein, C., Jansen, A., Deppe, METRO., Foerster, A. F., verano, J., Wolbers, T., &
Knecht, S. (2005). Hippocampus activity differentiates good from poor learners
of a novel lexicon. NeuroImagen, 25(3), 958–968. 10.1016/j.neuroimage.2004.12.019,
PubMed: 15808996
Brodmann, k. (1909). Beiträge zur histologischen Lokalisation der Grosshirnrinde:
VI. Die Cortexgliederung des Menschen. Journal für Psychologie und Neurologie,
10, 231–246.
bullmore, MI., & despreciar, oh. (2009). Complex brain networks: Graph theoretical analy-
sis of structural and functional systems. Naturaleza Reseñas Neurociencia, 10, 186–198.
10.1038/nrn2575, PubMed: 19190637
Buzsáki, GRAMO. (2010). Neural syntax: Cell assemblies, synapsembles, and readers. nuevo-
ron, 68, 362–385.
Buzsáki, GRAMO. (2019). The brain from inside out. Oxford: prensa de la Universidad de Oxford.
Carrillo-Reid, l., Han S., Yang W., Akrouh, A., & Yuste, R. (2019). Controlling visu-
ally guided behavior by holographic recalling of cortical ensembles. Cell, 178(2),
447–457.
Catani, METRO., jones, D. K., & Ffytche, D.H. (2005). Perisylvian language net-
works of the human brain. Annals of Neurology, 57(1), 8-dieciséis. 10.1002/ana.20319,
PubMed: 15597383
Chomsky, norte. (1956). Three models for the description of language. IRE Trans. Inform.
Teoría, 2, 113–124. 10.1109/TIT.1956.1056813
Chomsky, norte. (1995). The minimalist program. Cambridge, MAMÁ: CON prensa.
douglas, R. J., & Martín, k. A. C. (2004). Neuronal circuits of the neocor-
tex. Annual Reviews Neuroscience, 27, 419–451. 10.1146/annurev.neuro.27.070203
.144152
Durstewitz, D., Seamans, j. K., & Sejnowski, t. j. (2000). Neurocomputational mod-
els of working memory. Neurociencia de la naturaleza, 3(11), 1184–1191. 10.1038/81460,
PubMed: 11127836
Eichenbaum, h. (2018). Barlow versus Hebb: When is it time to abandon the notion
of feature detectors and adopt the cell assembly as the unit of cognition? Neuro-
science Letters, 680, 88–93. 10.1016/j.neulet.2017.04.006, PubMed: 28389238
Fedorenko, MI., Hsieh, PAG. J., Nieto-Castanon, A., Whitfield-Gabrieli, S., & Kanwisher,
norte. (2010). New method for fMRI investigations of language: Defining ROIs
functionally in individual subjects. Revista de neurofisiología, 104(2), 1177–1194.
10.1152/jn.00032.2010, PubMed: 20410363
Frankland, S. METRO., & verde, j. D. (2015). An architecture for encoding sentence
meaning in left mid-superior temporal cortex. In Proceedings of the National
Academy of Sciences of the United States of America, 112(37), 11732–11737. 10.1073/
pnas.1421236112
Franks, k. METRO., ruso, METRO. J., Sosulski, D. l., Mulligan, A. A., Siegelbaum, S. A., &
Axel, R. (2011). Recurrent circuitry dynamically shapes the activation of piriform
corteza. Neurona 72, 49–56 (2011). 10.1016/j.neuron.2011.08.020, PubMed: 21982368
Friederici, A. D. (2009). Pathways to language: Fiber tracts in the human brain. Trends
in Cognitive Sciences, 13(4), 175–181. 10.1016/j.tics.2009.01.001, PubMed: 19223226
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
304
C. Papadimitriou and A. Friederici
Friederici, A. D. (2011). The brain basis of language processing: From structure
to function. Physiological Reviews, 91, 1357–1392. 10.1152/physrev.00006.2011,
PubMed: 22013214
Friederici, A. D. (2017). Language in our brain. The origins of a uniquely human capacity.
Cambridge, MAMÁ: CON prensa.
Friederici, A. D., & Cantante, W.. (2015). Grounding language processing on basic neu-
rophysiological principles. Tendencias en Ciencias Cognitivas, 19(6), 329–338. 10.1016/
j.tics.2015.03.012, PubMed: 25890885
Goucha, T., anwander, A., & Friederici, A. D. (2015). How language shapes the brain:
Cross-linguistic differences in structural connectivity. Poster presented at the 45th
Annual Meeting of the Society for Neuroscience, chicago.
harris, k. D. (2005). Neural signatures of cell assembly organization. Reseñas de naturaleza
Neurociencia, 6(5), 399–407. 10.1038/nrn1669, PubMed: 15861182
harris, K., Csicsvari, J., Hirase, Dragoi, GRAMO., & Buzsáki, GRAMO. (2003). Organization of
cell assemblies in the hippocampus. Naturaleza, 424, 552–556. 10.1038/nature01834,
PubMed: 12891358
Hebb, D. oh. (1949). The organisation of behavior. Nueva York: wiley.
Ison, METRO. J., Quiroga, q. r., & Frito, I. (2015). Rapid encoding of new memories
by individual neurons in the human brain. Neurona, 87(1), 220–230. 10.1016/
j.neuron.2015.06.016, PubMed: 26139375
Kriegeskorte, NORTE., & douglas, PAG. k. (2018). Cognitive computational neuroscience. Na-
ture Neuroscience, 21, 1148–1160. 10.1038/s41593-018-0210-5, PubMed: 30127428
Kunze, T., Haueisen, J., & Knösche, t. R. (2019). Emergence of cognitive prim-
ing and structure building from the hierarchical interaction of canonical micro-
circuit models. Cibernética biológica, 113, 273–291. 10.1007/s00422-019-00792-y,
PubMed: 30767085
Kunze T., Peterson A. D. h., Haueisen J., & Knösche T. R. (2017). A model of individ-
ualized canonical microcircuits supporting cognitive operations. PLOS One, 12,
e0188003. 10.1371/diario.pone.0188003, PubMed: 29200435
Lau, mi. F., Phillips, C., & Poeppel, D. (2008). A cortical network for semantics:
(De)constructing the N400. Naturaleza Reseñas Neurociencia, 9 (12), 920–933. 10.1038/
nrn2532, PubMed: 19020511
Lillicrap, t. PAG., Santoro, A., Marris, l., Akerman, C. J., & Hinton, GRAMO. (2020). Back-
propagation and the Brain. Naturaleza Reseñas Neurociencia, 21, 335–346. 10.1038/
s41583-020-0277-3, PubMed: 32303713
Makuuchi, METRO., Bahlmann, J., anwander, A., & Friederici, A. D. (2009). Segregating
the core computational faculty of human language from working memory. En
Proceedings of the National Academy of Sciences of the USA, 106, 8362–8367. 10.1073/
pnas.0810928106, PubMed: 19416819
Markov, norte. T., & Kennedy, h.
(2013). The importance of being hierarchi-
California. Opinión actual en neurobiología, 23, 187–194. 10.1016/j.conb.2012.12.008,
PubMed: 23339864
milne, A. MI., Mueller,
j. l., Männel, C., Attaheri, A., Friederici, A. D., &
Petkov, C. I. (2016). Evolutionary origins of non-adjacent sequence process-
ing in primate brain potentials. Informes Científicos, 6: 36259. 10.1038/srep36259,
PubMed: 27827366
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Bridging Neurons and Cognition Through Assemblies of Neurons
305
Mitropolsky, D., collins, METRO., & Papadimitrio, C. h. (in press). A biologically plau-
sible parser. Transacciones de la Asociación de Lingüística Computacional. arXiv:
abs/2108.02189.
Müller M. GRAMO., Papadimitrio, C. h., Maass, w., & Legenstein, R. (2020). A model for
structured information representation in neural networks of the brain. eNeuro,
7(3). 10.1523/ENEURO.0533-19.20207.
Nucifora, PAG. GRAMO, Verma, r., Melhem, mi. r., Gur, R. MI., & Gur, R. C. (2005). Leftward
asymmetry in relative fiber density of the arcuate fasciculus. NeuroReport, 16(8),
791–794. 10.1097/00001756-200505310-00002
Pallier, C., Devauchelle, A.-D., & Dehaene, S. (2011). Cortical representation of the
constituent structure of sentences. In Proceedings of the National Academy of Sciences
of the United States of America, 108(6), 2522–2527. 10.1073/pnas.1018711108
Papadimitrio, C. h. (1995). Computational complexity. Reading, MAMÁ: Addison-
Wesley.
Papadimitrio, C. h., & Vempala, S. S. (2019). Random projection in the brain and
computation with assemblies of neurons. 10th Innovations in Theoretical Computer
Ciencia. Waderman, Alemania: Leibniz International Proceedings in Informatics,
Schloss Dagstuhl.
Papadimitrio, C. h., Vempala, S. S., Mitropolsky D., collins, METRO. J., & Maass, W..
(2020). In Proceedings of the National Academy of Sciences of the United States of Amer-
ica, 117(25), 14464–14472.
Pokorny, C., Ison, METRO. J., Rao, A., Legenstein, r., Papadimitrio, C. h., & Maass,
W.. (2020). STDP forms associations between memory traces in networks
of spiking neurons. Corteza cerebral, 30(3), 952–968. 10.1093/cercor/bhz140,
PubMed: 31403679
Quiroga, R. q. (2016). Neuronal codes for visual perception and memory. Neuropsy-
chologia, 83, 227–241. 10.1016/j.neuropsychologia.2015.12.016
Saur, D., Kreher, B. w., Schnell, S., Kümmerer, D., Kellmeyer, PAG., Vry, M.-S., . . .
Weiller, C. (2008). Ventral and dorsal pathways for language. En procedimientos de
the National Academy of Sciences, 105(46), 18035–18040. 10.1073/pnas.0805234105
Schomers, METRO. r., Garagnani, METRO., & Pulvermüller, F. (2017). Neurocomputational
consequences of evolutionary connectivity changes in perisylvian language cor-
tex. Revista de neurociencia, 37, 3045–3055. 10.1523/JNEUROSCI.2693-16.2017,
PubMed: 28193685
Skeide, METRO. A., Brauer, J., Friederici, A. D. (2016). Brain functional and structural
predictors of language performance. Corteza cerebral, 26(5), 2127–2139. 10.1093/
cercor/bhv042, PubMed: 25770126
Turing, A. METRO. (1937). On computable numbers, with an application to the Entschei-
dungsproblem. In Proceedings of the London Mathematical Society, 42, 230–265.
10.1112/plms/s2-42.1.230
Upadhyay, J., Silver, A., Knaus, t. A., Lindgren, k. A., Ducros, METRO., kim, D.-S.,
& Tager-Flusberg, h.
(2008). Effective and structural connectivity in the
human auditory cortex. Revista de neurociencia, 28(13), 3341–3349. 10.1523/
JNEUROSCI.4434-07.2008, PubMed: 18367601
Xu, w., & Sudhof, t. C. (2013). A neural circuit for memory specificity and generaliza-
ción. Ciencia, 339(6125), 1290–1295. 10.1126/science.1229534, PubMed: 23493706
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
306
C. Papadimitriou and A. Friederici
Zaccarella, MI., & Friederici, A. D. (2015). Merge in the human brain: a sub-region
based functional investigation in the left pars opercularis. Fronteras en psicología,
6, 1818. 10.3389/fpsyg.2015.01818, PubMed: 26640453
Zaccarella, MI., Meyer, l., Makuuchi, METRO., & Friederici, A. D. (2017). Building by syn-
tax: The neural basis of minimal linguistic structures. Corteza cerebral, 27(1), 411–
421. 26464476
Received April 26, 2021; accepted September 2, 2021.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
mi
d
tu
norte
mi
C
oh
a
r
t
i
C
mi
–
pag
d
/
yo
F
/
/
/
/
3
4
2
2
9
1
2
0
0
6
7
9
9
norte
mi
C
oh
_
a
_
0
1
4
6
3
pag
d
.
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3