Early Electrophysiological Markers of
Navigational Affordances in Scenes
Assaf Harel1, Jeffery D. Nador1, Michael F. Bonner2, and Russell A. Epstein3
Abstracto
■ Scene perception and spatial navigation are interdependent
cognitive functions, and there is increasing evidence that corti-
cal areas that process perceptual scene properties also carry
information about the potential for navigation in the environ-
mento (navigational affordances). Sin embargo, the temporal stages
by which visual information is transformed into navigationally
relevant information are not yet known. Nosotros planteamos la hipótesis de que
navigational affordances are encoded during perceptual pro-
cessing and therefore should modulate early visually evoked
ERPs, especially the scene-selective P2 component. To test this
idea, we recorded ERPs from participants while they passively
viewed computer-generated room scenes matched in visual
complejidad. By simply changing the number of doors (0 doors,
1 door, 2 doors, 3 doors), we were able to systematically vary
the number of pathways that afford movement in the local
ambiente, while keeping the overall size and shape of the
environment constant. We found that rooms with 0 doors
evoked a higher P2 response than rooms with three doors, estafa-
sistent with prior research reporting higher P2 amplitude to
closed relative to open scenes. Además, we found P2 ampli-
tude scaled linearly with the number of doors in the scenes.
Navigability effects on the ERP waveform were also observed
in a multivariate analysis, which showed significant decoding
of the number of doors and their location at earlier time win-
dows. Juntos, our results suggest that navigational affor-
dances are represented in the early stages of scene perception.
This complements research showing that the occipital place
area automatically encodes the structure of navigable space
and strengthens the link between scene perception and
navigation. ■
INTRODUCCIÓN
How do we find our way in the environment? What allows
us to successfully move about in the world without getting
lost? Navigation, the act of finding one’s way to a given
destination, is a multisensory process requiring the inte-
gration of multiple types of sensory information (visual,
vestibular, proprioceptive) about the environment,
including information about direction, distancia, and loca-
ción (Ekstrom, Spiers, Bohbot, & Rosenbaum, 2018). En
humanos, the most prominent sensory modality guiding
navigation in the environment is vision (Ekstrom, 2015).
The particular advantage that vision confers to navigation
is that it allows observers to recognize their surroundings
remotely, even before they embark on any movement in
their environment. A este respecto, visual scene perception,
eso es, the recognition of one’s surroundings, can be con-
sidered as an essential part of navigation, serving as a first
vital stage in a cascade of processes, which ultimately
culminate in successful navigation ( Julian, Keinath,
Marchette, & Epstein, 2018). Under this conceptualiza-
ción, scene perception serves as the gate to navigation,
demonstrating more broadly how visual processing
(es decir., scene recognition) is critical for and cannot be
1Wright State University, Dayton, OH, 2Johns Hopkins Univer-
sity, baltimore, Maryland, 3Universidad de Pennsylvania, Filadelfia
© 2022 Instituto de Tecnología de Massachusetts
dissociated from action, planificación, and memory systems
(es decir., navigation).
Support for the link between scene recognition and spa-
tial navigation1 comes from neuroimaging. Several studies
using fMRI reported that scene-selective regions sensitive
to visual scene properties, such as category ( Walther,
Chai, Caddigan, Arroyo, & Fei-Fei, 2011; Walther, Caddigan,
Fei-Fei, & Arroyo, 2009) and spatial expanse (Harel, Kravitz,
& Panadero, 2013; Kravitz, Peng, & Panadero, 2011), also carry
pertinent information for navigation (Persichetti & Dilks,
2018; Bonner & Epstein, 2017; Kamps, Julian, Kubilius,
Kanwisher, & Dilks, 2016). One region in particular that
has been suggested to be involved in representing visually
guided navigation information is the occipital place area
(OPA: Dilks, Julian, Paunov, & Kanwisher, 2013; Hasson,
Harel, Exacción, & Malach, 2003; Nakamura et al., 2000).
OPA activity is sensitive to various forms of navigation-
relevant information, including egocentric position and
heading (es decir., “viewpoint”: Epstein, Higgins, Jablonski, &
Feiler, 2007; Epstein, Higgins, & Thompson-Schill, 2005)
“sense” (left–right: Dilks, Julian, Kubilius, Spelke, &
Kanwisher, 2011), egocentric distance (proximal-distal:
Persichetti & Dilks, 2016), first-person perspective motion
through scenes (Kamps, Lall, & Dilks, 2016), and number
of local elements in a scene, which can be used for obstacle
avoidance (Kamps, Julian, et al., 2016). En tono rimbombante, OPA
response patterns have also been reported to specifically
Revista de neurociencia cognitiva 34:3, páginas. 397–410
https://doi.org/10.1162/jocn_a_01810
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
oh
C
norte
_
a
_
0
1
8
1
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
index the spatial structure of navigational affordances (es decir.,
potential paths for movement in a scene), as operational-
ized by the number and locations of paths in the environ-
mento (Bonner & Epstein, 2017).
Although there is clear evidence that scene-selective cor-
tical regions carry navigation-related information, it is still
not clear how incoming visual information is transformed
into navigation-relevant information that could potentially
be used to guide movement in space. A recent TMS study
demonstrated that OPA plays a causal role in transforming
perceptual inputs into spatial memories linked to environ-
mental boundaries ( Julian, ryan, hamilton, & Epstein,
2016), but this study used a continuous theta-burst proto-
col that lacked the temporal resolution to probe the tem-
poral sequence of these processes. Information about
timing is essential for determining the extent to which
the extraction of navigation-related information from a
scene is indeed a visually driven perceptual process.
One alternative, por ejemplo, is that OPA sensitivity to nav-
igational affordances might reflect recurrent feedback
from posterior parietal cortex (Kravitz, Saleem, Panadero, &
Mishkin, 2011) rather than early sensory processing.2
Determining the nature of scene affordances processing
thus requires a method with high temporal resolution,
such as magnetoencephalography (MEG) and EEG,
which can establish the temporal dynamics of scene per-
ception for navigation. Yet another advantage of EEG is
that it can establish the processes involved in the extrac-
tion of navigationally relevant information: Various ERPs
have been shown to index multiple cognitive processes
(p.ej., attentional allocation, semantic categorization,
memory encoding; para una revisión, see the work of Luck,
2014) and thus be used to determine the mechanisms
underlying a certain task or experimental manipulation.
M/EEG studies have recently started to uncover the
time course of scene perception, by examining how differ-
ent scene properties across a variety of complexity levels
get processed over time. At a categorical level, the process-
ing of scenes can be distinguished from the processing of
other complex visual categories by 220 msec poststimulus
onset (Harel, Groen, Kravitz, Deouell, & Panadero, 2016; Sato
et al., 1999). Específicamente, the amplitude of the posterior
P2 ERP component (peaking around 220 msec after stim-
ulus onset) is higher in response to scenes than to faces
and common objects. The P2 component has thus been
suggested to index scene-selective processing (analogous
to the face-selective N170 ERP component), particularly
the processing of high-level global scene information
(Harel et al., 2016, 2020; Hansen, Noesen, Nador, & Harel,
2018). En efecto, P2 amplitude not only distinguishes
between scenes and other visual categories but is also sen-
sitive to various global scene properties (GSPs), como
spatial expanse and naturalness: It is higher to closed than
open scenes, can distinguish natural from man-made
escenas (Harel et al., 2016, 2020; Hansen et al., 2018),
and notably, it is not modulated by local texture informa-
ción, in contrast to earlier visually evoked components
(Harel et al., 2020). P2 amplitude is also diagnostic of
scene naturalness and spatial expanse at the level of
individual scene images, with its response variance to indi-
vidual scenes significantly explained by both summary
image statistics (approximating naturalness and spatial
expanse) and subjective behavioral ratings (Harel et al.,
2016; see also the work of Cichy, Khosla, Pantazis, & Oliva,
2017). Studies using single-trial decoding approaches
revealed that scene naturalness and spatial expanse (como
well as basic-level scene category) can also be decoded
from the neural signals even earlier, within the first
100 msec of processing (Henriksson, Mur, & Kriegeskorte,
2019; Lowe, Rajsic, Ferber, & Walther, 2018; see also the
works of Groen, Ghebreab, Lamme, & Scholte, 2016;
Groen, Ghebreab, Prins, Lamme, & Scholte, 2013).3 El
latency of both time windows suggests that low- también
as high-level diagnostic scene information is extracted at
the early perceptual stages of processing, supporting rapid
pre-attentive scene categorization (Hansen et al., 2018;
Groen et al., 2016; verde & Oliva, 2009; Rousselet,
Joubert, & Fabre-Thorpe, 2005).
Although the above M/ EEG studies cannot directly
establish OPA as the generator of the spatial expanse sig-
nals indexed by the P2 component, they are nevertheless
invaluable in providing a framework for thinking about the
visual system’s time course for extracting information
about scene navigability. If the processing of ecological,
navigability information occurs at the perceptual stages
of processing, this should be reflected in a modulation
of visually evoked activity during the first 250 mseg
(primarily the scene-selective P2 component) by naviga-
tionally relevant information, suggesting that scene per-
ception and navigation are indeed intrinsically linked.
Alternativamente, the extraction of navigability information
from the scene might reflect postperceptual processes
(p.ej., stimulus evaluation, Toma de decisiones, action plan-
y), which would result in a late effect of navigability,
and hence not impact the P2. One indication that the more
probable alternative of the two is former rather than the
latter is the ubiquity of spatial expanse—the extent to
which a scene depicts an enclosed or an open space—in
modulating early scene-evoked neural responses. El
effect of spatial expanse can be observed during early
stages of visual processing (Henriksson et al., 2019;
Hansen et al., 2018), across a variety of stimulus sets and
physical image properties (p.ej., with both line drawings
and photographs; grayscale as well as color images; con
both artificial and naturalistic scene images: Harel et al.,
2016, 2020; Hansen et al., 2018; Lowe et al., 2018; Cichy
et al., 2017) and across various task contexts (Hansen
et al., 2018; Lowe et al., 2018), indicating its centrality as
a source of information for scene perception. One reason
why spatial expanse may prove to be important for scene
perception is because of its potential link with navigability.
The spatial expanse of a scene conveys information not
only about the structure and geometry of the scene but
also about its function, a saber, the conceivable
398
Revista de neurociencia cognitiva
Volumen 34, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
oh
C
norte
_
a
_
0
1
8
1
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
possibilities for movement in space. And because the spa-
tial expanse of a scene, or rather its openness, is perceived
by humans as a continuous dimension (zhang, Houpt, &
Harel, 2019), closed and open scenes can thus be thought
of as two ends on an ease-of-navigation (navigability)
continuum, with closed environments posing more con-
straints on navigation than open ones. De este modo, it may be
argued that the degree of openness or enclosure also con-
fer constraints on navigability. It therefore stands to reason
that early neural responses to spatial expanse index infor-
mation about scene structure not only with regard to its
openness, but also as it defines navigable space. En efecto,
ERP amplitudes produced by spatial expanse variation
are very slightly, en todo caso, modulated by task demands
(Hansen et al., 2018), in line with the notion that naviga-
tional affordances are extracted automatically (Bonner &
Epstein, 2017).
The present work sought to establish the time course of
the extraction of navigational affordances and, específicamente,
to explicitly test the idea that early scene-evoked activity
represents the potential for navigation in a scene.
Although previous studies have uncovered the temporal
dynamics of spatial layout processing, they did not explic-
itly establish the relation between these neural signatures
and the extraction of navigational affordances. We argued
that if scene perception involves the extraction of func-
tional information for navigation, then the structure of
navigable space (es decir., navigational affordances) should be
encoded relatively early in processing and thus manifest in
early visually evoked ERPs. We hypothesized that the nav-
igational affordances of the visual environment would be
resolved at the P2 time window and, específicamente, that the
amplitude of the P2 component would capture the ease of
navigation in the environment given that the P2 distin-
guishes between open and closed scenes, (which can be
thought of as offering more or less navigability, respetar-
activamente). To test this hypothesis, we recorded ERPs from
participants while they passively viewed computer-
generated room scenes matched in visual complexity
(used in a previous fMRI study of navigability, see the work
of Bonner & Epstein, 2017). The rooms varied in the
number of pathways that afford movement in the local
ambiente: By simply changing the number of doors
(0 doors, 1 door, 2 doors, 3 doors) in the room, nosotros
were able to systematically control the number of move-
ment paths in the scene, while keeping the overall size
and shape of the environment constant. If encoding
navigational affordances engages stimulus-driven percep-
tual processes, then sensitivity to number of movement
paths in the environment should emerge within the first
250 msec of processing. Específicamente, neural activity during
the P2 time window should be modulated by the number
of doors in the scenes, with increasing P2 amplitude as a
function of constraints on navigability (es decir., decreasing
number of doors). Alternativamente, if manipulating of num-
ber of doors in the scene does not result in early pertur-
bations of neural activity and is observed later, then that
would lend support to the idea that encoding navigation
information reflects postperceptual processing rather
than stimulus-driven visual processing. Complementing
the hypothesis-driven univariate ERP analysis of P2 ampli-
tude, we also conducted a more data-driven, multivariate
analysis to assess the decoding of navigability information
con el tiempo, how early it emerges, and to what extent it rep-
resents the extraction of information about the local posi-
tion of navigability cues, or whether that information is
extracted in a position-invariant fashion.
MÉTODOS
Participantes
Thirty-six Wright State University students (21 women;
mean age: 20 años) participated in the study. Participantes
signed an informed written consent form according to the
guidelines of the institutional review board of Wright State
University and were compensated monetarily or with
course credit. All participants had normal or corrected-
to-normal visual acuity and no history of neurological dis-
ease, and all but one were right-handed. Six participants
were excluded from final analyses because of excessive
EEG artifacts.
Estímulos
The stimuli comprised 144 computer-generated images of
simple rectangular rooms, used in a previous neuroimag-
ing study (Bonner & Epstein, 2017). Every room scene
contained either a door or a painting on each of its three
visible walls, yielding eight navigability conditions accord-
ing to the number of door elements present combined
with the walls on which they appeared: 0 doors; 1 door
izquierda, bien, or center; 2 doors left–center, left–right, o
center–right; y 3 doors (ver figura 1 for examples of
estímulos). For each navigability condition, 18 unique rooms
were created by applying different textures to the walls,
making a total of 144 unique exemplars. The stimuli were
presented using Presentation software (Neurobehavioral
Sistemas, Cª, www.neurobs.com). Images were displayed
in 8-bit color at the center of a Dell LCD monitor (1920 ×
1080 píxeles) at a viewing distance of about 150 cm, sub-
tending 12 × 14 degrees of visual angle.
Experimental Design and Procedure
Participants viewed the 144 individual scene stimuli 8
veces (eight blocks), each block containing all 144 estímulos
(a total of 1152 ensayos). Scene stimuli were pseudoran-
domized within individual blocks and across the eight
blocks to prevent direct repetition of any stimulus, naviga-
bility condition, or texture. Stimuli were presented for
500 msec with a randomly jittered interstimulus interval
que van desde 1000 a 2000 mseg. Los participantes actuaron.
a fixation cross task, in which they were required to
Harel et al.
399
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
oh
C
norte
_
a
_
0
1
8
1
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. Examples of room
images used in the experiment.
The rooms varied in the ease
of navigability they afford,
operationalized by the number
of doors in the scene. Por
changing the number of doors
in the room (0 door, 1 door,
2 doors, 3 doors), we were able
to systematically control the
number of movement paths
in the scene, while keeping
the overall size and shape
of the environment constant.
Depicted here are eight scenes
(out of a total 144 escenas)
spanning all four navigability
condiciones. Note that the 1-door
and 2-doors conditions included
three variations in door
ubicación. The rooms also varied
in their spatial layout and
texture, yielding 18 distinto
room variants.
report whether the horizontal or vertical bar of the central
fixation cross (1-degree visual angle) lengthened on each
trial by a factor of 25%. Changes in the fixation cross were
randomized across trials and, hence, were independent
from the actual content of the underlying image, essen-
tially requiring the participants to pay very little, if any,
attention to the background images while completing
this task. This same task has been employed in previous
EEG studies of scene processing using naturalistic real-
world stimuli (Hansen et al., 2018; Harel et al., 2016), como
well as computer-generated room-like stimuli (Harel
et al., 2020). We verified that the task conditions were
indeed independent from the stimulus conditions by
conducting a Task × Condition ANOVA on participants’
accuracy scores (see Results section).
EEG Recording
Analog EEG signals were recorded using 64 Ag-AgCl pin-
type active electrodes (Biosemi ActiveTwo) mounted on
an elastic cap (Electro-Cap International, Cª) according
to the extended 10–20 system. EOGs were recorded from
two additional pairs of pupil-aligned electrodes: One pair
was placed on the skin over the right and left temporal
zygomatic bones; the other was placed over the nasal
zygomatic and frontal bones. Analog EEG data from all
electrodes were referenced to the common mode signal
electrode placed between electrodes PO3 and PO4.
Impedance of all channels was measured before the start
of each recording session, to ensure that all fell below
50 KOhms, and data for each electrode were inspected
to ensure no “bridging” artifacts were present.
In postprocessing, data were rereferenced to an elec-
trode placed on the tip of the nose. Both EEG and EOG
were sampled at 512 Hz with a resolution of 24 bits with
an active input range of −262 μV to +262 μV per bit.
The digitized EEG was saved and processed off-line.
Data Processing
The data were preprocessed using Brain Vision Analyzer 2
(Cerebro Productos GmbH). The raw data were first band-pass
filtered from 0.3 a 80 Hz (24 dB), with a second-order But-
terworth filter, and referenced to the tip of the nose. Eye
movements were corrected in the scalp electrode data
using an automated, restricted infomax ocular correction
independent component analysis (for details see the work
of Jung et al., 1998), effectively removing components
heavily correlated with HEOG and VEOG artifacts follow-
ing a meaned slope algorithm. Del 64 independiente
componentes (one per scalp electrode) calculated for each
partícipe, 14 ± 4 were removed on average. Remaining
artifacts exceeding ±100 μV in amplitude or containing a
change of over 100 μV in a period of 50 msec were
rejected. The preprocessed data were then segmented
into epochs ranging from −200 msec before to 800 mseg
after stimulus onset for all conditions. Participants’ data
were entirely excluded from analyses if fewer than 80%
of epochs could be retained following artifact removal.
Six participants’ data were thus excluded, and for the
remaining 30, an average of 1056 ± 63 (equivalent to
91% ± 6%) epochs were retained for all analyses.
ERP Univariate Analysis
The peaks of the P1, N1, and P2 were determined for each
individual participant, by automatically detecting the max-
imal amplitude in predetermined time windows in each
experimental condition (most positive peak between 80
y 130 mseg, most negative peak between 130 y
400
Revista de neurociencia cognitiva
Volumen 34, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
oh
C
norte
_
a
_
0
1
8
1
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
200 mseg, and most positive peak between 200 y
320 mseg, respectivamente). The mean latencies of the three
components across conditions were consistent with previ-
ous research (P1: 121 mseg [SEM = 3]; N1: 167 mseg
[SEM = 3]; P2: 230 mseg [SEM = 3]). Analyses were
restricted to posterior lateral sites (averaged across P7,
P5, P9, and PO7 for the left hemisphere, and across P8,
P6, P10, and PO8 for the right hemisphere), where maxi-
mal scene effects were previously observed (Harel et al.,
2016, 2020; Hansen et al., 2018). Mean peak amplitudes
(across participants) were analyzed using a two-way
within-subject ANOVA, with Hemisphere (izquierda, bien) y
Navigability (number of doors) as independent factors.
Because numbers of doors varied parametrically, tenemos
also conducted a linear trend analysis to assess whether
the amplitude of the ERP components scales with decreas-
ing number of doors, implemented as a two-way ANOVA
estructura (for further details on this analysis, see the
work of Pinhas, Tzelgov, & Ganor-Stern, 2012).
ERP Multivariate Analysis
Representational similarity analyses (RSA) were con-
ducted on the segmented EEG data, across all 64 canales
(but not external electrodes), for all artifact-free segments.
Data were exported from Brain Vision Analyzer and pro-
cessed in MATLAB ( Version 2016a, MathWorks) as four-
dimensional matrices (Channel × Condition × Segment ×
Time Point). Subject-level ERP data were submitted to
RSAs against two-model representational dissimilarity
matrices (RDMs), pertaining to the two-alternative
hypotheses: location-specific processing, in which the spa-
tial layout of the doors influenced processing (Figura 4A,
left column); and location-invariant processing, en el cual
only the “global” differences (es decir., between the total num-
ber of doors) influenced processing (Figura 4A, right col-
umn). Experimental conditions were coded as binary
triplets, with ones and zeros corresponding to doors
and paintings, respectivamente. Each digit within a triplet cor-
responded to one of the three possible locations (izquierda,
bien, or center). So, Por ejemplo, condition 1–0–0 would
correspond to a single door on the left, whereas 0–0–1
would correspond to a single door on the right. De este modo, el
identity and location of each door or painting was retained
in these condition codes. Codes were then entered in
ascending order as the rows and columns of a confusion
matrix to construct both model and neural RDMs. Tal como,
each cell corresponded to one pairing of stimulus condi-
ciones. For the location-specific model RDM, values in each
cell were derived from the number and location of doors
(and paintings) in its component row and column stimulus
condiciones (Bonner & Epstein, 2017). Mathematically,
these values were obtained by subtracting the similarity
between stimulus conditions from the maximum possible
semejanza. Similarity was computed as the sum of the dot
product of the condition codes corresponding to the row
and column of the cell, yielding a value between 0
(maximally dissimilar) y 3 (maximally similar). Differ-
ences between conditions in a given cell could therefore
be denoted by the difference between maximal similarity
and the calculated similarity. Mientras tanto, cell values in
the location-invariant model RDM were derived only from
the total number of doors (and paintings) present in each
pair of conditions and not their locations. Mathematically,
they were coded as the difference between sums of the row
and column stimulus conditions. Because the neural RDMs
were constructed using Pearson correlation coefficients of
determination ranging from 0 a 1 (see below), we divided
the cell values in the model RDMs (que van desde 0 a 3) por
3 to achieve common metrics for both. La resultante
model RDMs are presented in Figure 4.
Neural RDMs were constructed for each subject by first
calculating the Pearson correlation coefficient of determi-
nation between the ERP amplitudes of all possible pairs of
conditions across electrodes, separately at each time
punto. As these correlation coefficients denote pairwise
similarities between conditions, differences between con-
ditions were represented by taking the complement of the
coefficient of determination (1 − r2). In order to deter-
mine the level of agreement between the neural and
model RDMs for each subject, we took the Spearman cor-
relation between the two matrices (effectively comparing
the obtained pattern of activation across electrodes in the
neural RDMs to those predicted by each model RDM).
Finalmente, the Spearman correlation coefficients were sub-
mitted to cluster analyses (with a cluster induction param-
eter corresponding to a Type I error rate of .01) across
subjects to correct for multiple comparisons (Benjamini
& Hochberg, 2000).
RESULTADOS
Behavioral Performance: Verification of
Task Independence
We conducted a Task (horizontal change in fixation cross,
vertical change in fixation cross) × Stimulus (8 diferente
door conditions, see above) ANOVA on participants’ accu-
racy scores to verify that the orthogonal fixation task was
indeed independent of stimulus conditions. We found no
significant main effects of either factor (Tarea: F(1, 29) =
2.19, pag = .15, ηp
2 = .07; Stimulus: F(7, 203) = 1.85, pag =
.08, ηp
2 = .06), and notably, there was no interaction
between them, F(7, 203) = 1.02, pag = .42, ηp
2 = .03. En general,
this supports the orthogonality of the fixation cross task
to the stimulus (es decir., door) condiciones.
Univariate Analysis
To examine the extent to which navigational affordances,
operationalized as the number of doors in a room (es decir., el
number of pathways that afford movement in the local
ambiente), are encoded by early neuromarkers of
scene perception, we compared the amplitude of the early
Harel et al.
401
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
oh
C
norte
_
a
_
0
1
8
1
0
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
visually evoked ERP components (P1, N1, and P2) en
response to the different room conditions (0 doors,
1 door, 2 doors, 3 doors). We conducted a two-way
repeated-measures ANOVA on the amplitude of the indi-
vidually defined peaks of each of the ERP components,
with Hemisphere (izquierda, bien), and Number of Doors
(0 doors, 1 door, 2 doors, 3 doors) as independent
factores. The significant results of these analyses are
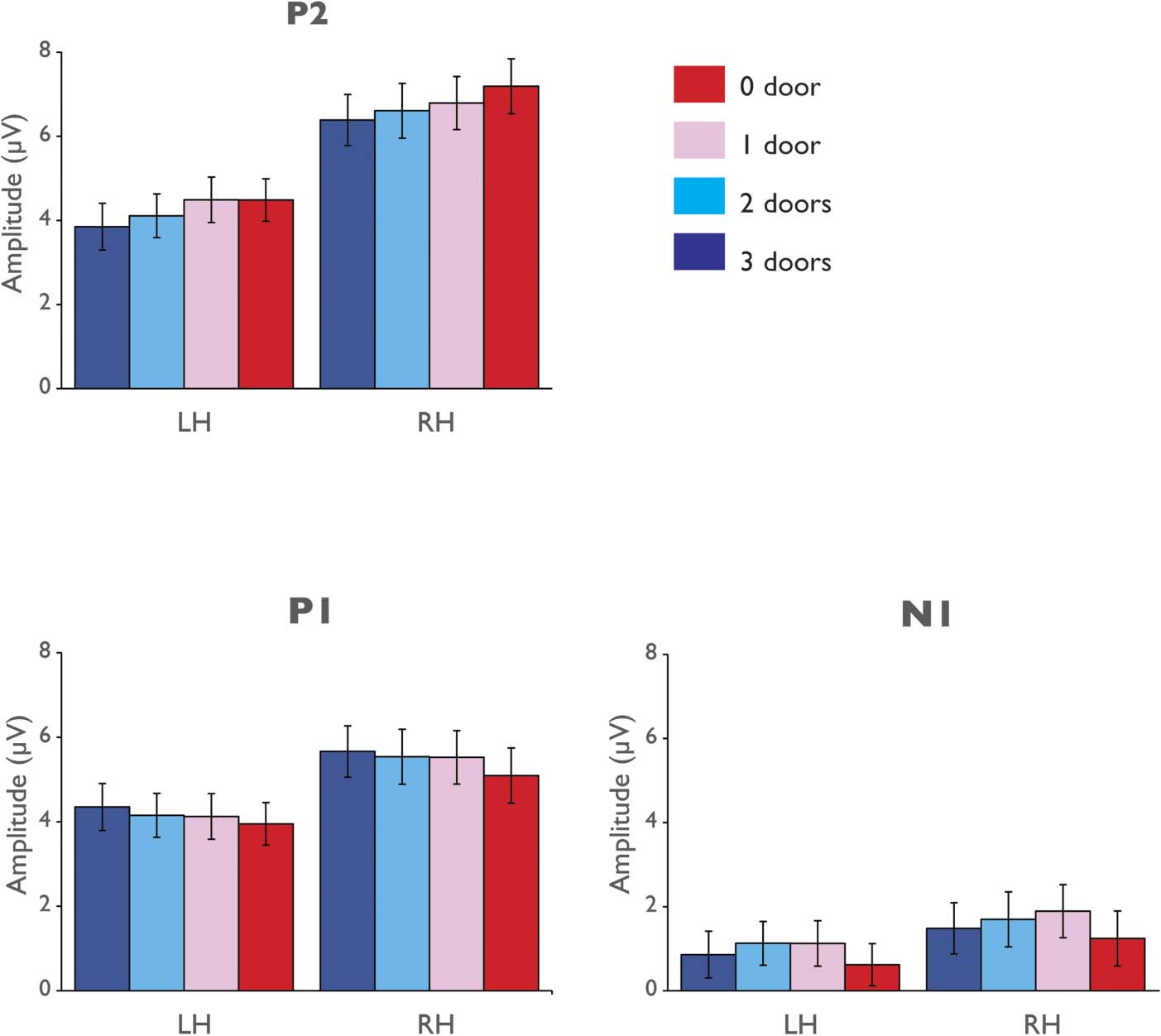
reported in Figure 2, and the grand-average waveforms
are depicted in Figure 3.
P2 Component
We found that the P2 amplitude is sensitive to the number
of doors contained in the room scenes, expressed by a sig-
nificant main effect of Number of Doors, F(3, 87) = 4.29,
pag = .007, ηp
2 = .13. To assess the source of the main effect
of Number of Doors, we have conducted two follow-up
analiza: linear trend analysis and pairwise post hoc com-
parisons. The former analysis showed that P2 amplitude
scales with the number of doors present in the scene,
manifesting in a significant linear trend, F(1, 29) = 7.58,
pag = .01, ηp
2 = .21. Post hoc comparisons revealed a signif-
icantly higher P2 amplitude for the 0-doors condition
(m = 5.83 mV, SEM = 0.51) compared to the 3-doors
condición (m = 5.12 mV, SEM = 0.42) , t(29) = 2.45,
pag = .009, and a higher amplitude to the 1-door (m =
5.64 mV, SEM = 0.47) relative to the 2-door condition
(m = 5.36 mV, SEM = 0.45), t(29) = 2.09, pag = .02.
A main effect of Hemisphere was also observed, F(1,
29) = 18.90, pag = .001, ηp
2 = .40, with higher amplitude
in the right hemisphere (m = 6.74 mV, SEM = 0.62)
compared with the left hemisphere (m = 4.23, SEM =
0.42). The effect of Number of Doors, sin embargo, was not
found to significantly differ across hemispheres (interac-
tion of Hemisphere with Number of Doors, F(3, 87) =
1.36, pag = .26, ηp
2 = .05; as standard ERP practice, nosotros
depict both hemispheres in our figures).
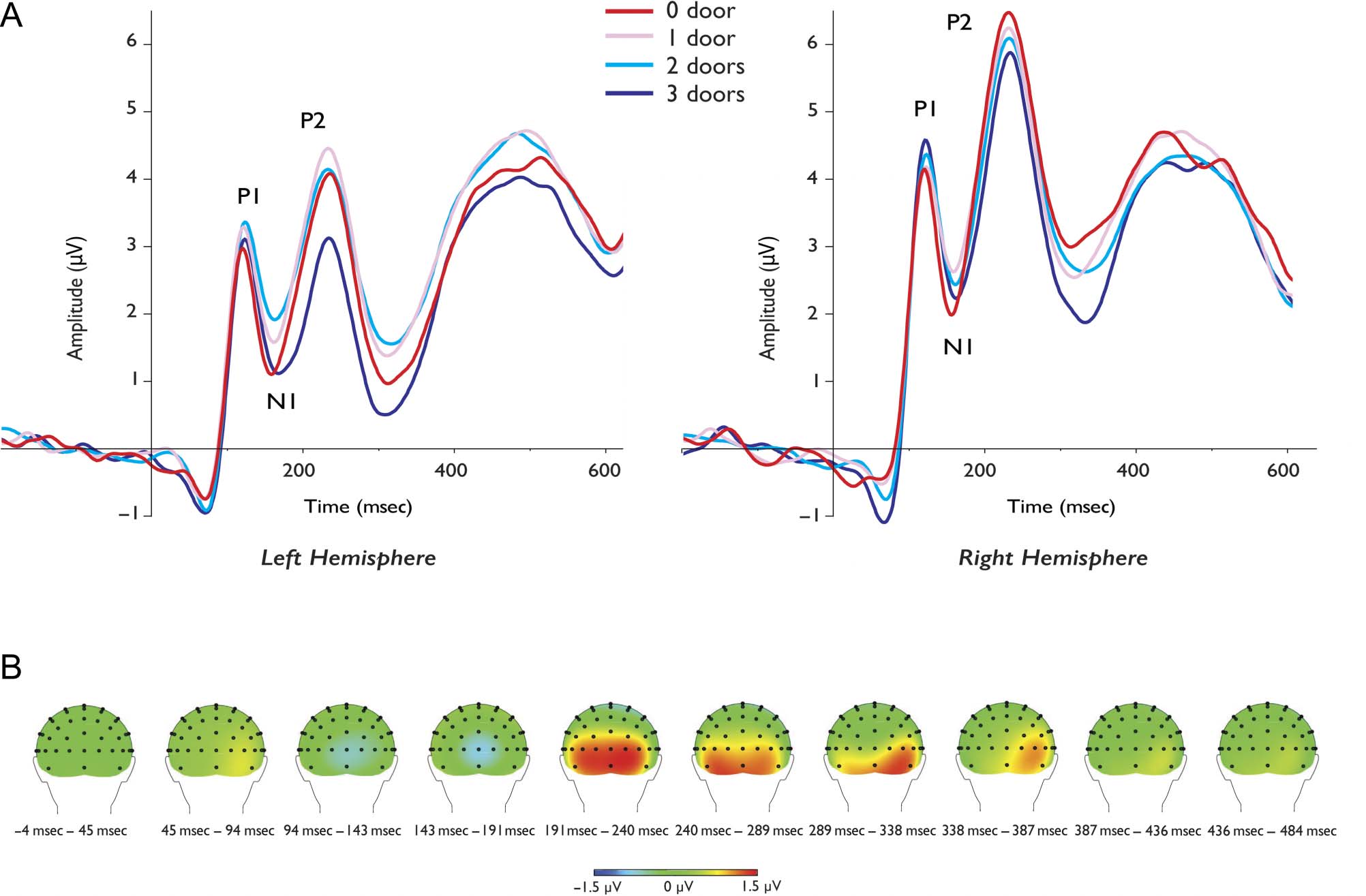
Notablemente, the navigational affordances effect persisted
beyond the P2 time window. Figure 3B depicts the dif-
ference ERP waveform contrasting the 3-doors and the
0-doors conditions across the whole scalp using current
source density (CSD) topographical maps. As can be
seen, the signal depicting difference in perceiving min-
imally navigable scenes to maximally navigable scenes
was present from around 200 msec to 350 msec post-
stimulus onset.
N1 Component
An analysis of the N1 component revealed the effect of
Number of Doors did not reach significance, F(3, 87) =
3.02, pag = .06, ηp
2 = .09. No significant linear trend was
noted as a function of number of doors, F(1, 29) < 1.00,
ηp
2 = .02, and the difference between the no-doors condi-
tion and the three doors condition was not found to be
significant (planned t test: t(29) = 0.85, p = .20). No
significant effects of hemisphere, F(1, 29) = 1.30, p =
Figure 2. Grand average ERP
univariate analysis results. Top
row: mean P2 peak amplitudes
in response to the four
navigability conditions (from
least navigable to most
navigable: 0 door, 1 door,
2 doors, 3 doors) presented
separately for the left and right
hemisphere. Bottom row: mean
P1 and N1 peak amplitudes for
the navigability conditions
presented separately for the left
and right hemispheres. Error
bars indicate between-subjects
SE; all data are plotted for the
posterior lateral sites. P2
amplitude showed a significant
linear increase in magnitude
of response as a function of
number of doors, with less
doors resulting in higher
amplitude. Notably, no such
increase was observed on
the amplitude of the P1 or
N1 components (see text
for details).
402
Journal of Cognitive Neuroscience
Volume 34, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
o
c
n
_
a
_
0
1
8
1
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
o
c
n
_
a
_
0
1
8
1
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. (A) Group-averaged waveforms for the four navigability conditions (from least navigable to most navigable: 0 door, 1 door, 2 doors,
3 doors) presented separately for the left and right hemisphere at posterior lateral sites. (B) Scalp CSD topographical maps depicting the difference
between the 3-doors and the 0-door conditions show a persistent effect of navigability.
2 = .04, or a Hemisphere × Number of Doors inter-
.26, ηp
action, F(3, 87) < 1.00, ηp
2 = .01, were observed.
P1 Component
Analysis of the amplitude of the P1 component showed
the effects of Number of Doors, and the Number of Doors
x Hemisphere did not reach significance, F(3, 87) = 2.56,
p = .08, ηp
2 = .03, respec-
tively. No significant main effect of Hemisphere was
observed, F(1, 29) = 3.94, p = .06, ηp
2 = .08; F(3, 87) = 1.03, p = .37, ηp
2 = .12.
We also performed a secondary analysis investigating
the extent to which the specific location of the door, which
might provide local information about potential move-
ment path, could have an effect on the early visually
evoked ERPs.4 We conducted two separate two-way ANO-
VAs with Hemisphere and Door Location as independent
variables: one ANOVA for the single-door condition and
another one for the two-doors conditions. For the
single-door condition, we were not able to find any signif-
icant effects of Door Location or Door Location x Hemi-
sphere on either the P1, N1, or P2 peak amplitudes (all
ps > .30). A significant effect of Hemisphere was found
on the amplitude of the P2 and P1 components (P2: F(1,
2 = .40; P1: F(1, 29) = 4.58, pag =
2 = .13), with higher amplitude in the right than in
29) = 18.52, pag = .001, ηp
.04, ηp
the left hemispheres.
For the two-doors condition, a significant main effect of
Door Location was found on the amplitude of the P2 com-
ponent, F(2, 58) = 3.70, pag = .04, ηp
2 = .11, and the interac-
tion between Door Location and Hemisphere was not
found to be significant, F(2, 58) < 1.00, ηp
2 = .01. Post
hoc comparisons showed significantly ( p < .05,
Bonferroni-corrected) lower amplitude to rooms in which
the doors were located in the center and right positions
compared to scenes with doors on the left and right, albeit
the latter were not significantly different than scenes with
left- and center-positioned doors.5 No significant effects of
Door Location or Door Location x Hemisphere were
observed on the N1 amplitude (all ps > .78). For the P1
component, we found a significant main effect of Door
Location, F(2, 58) = 6.00, pag = .006, ηp
2 = .17, mientras que el
interaction between Door Location and Hemisphere was
not found to be significant, F(2, 58) < 1.00, ηp
2 = .00. Lastly,
a main effect of Hemisphere was found for both the P1 and
P2 components (P1: F(1, 29) = 4.21, p = .05, ηp
2 = .13; P2:
F(1, 29) = 17.48, p = .001, ηp
2 = .37), reflecting a right hemi-
sphere advantage (see main analysis reported above).
Harel et al.
403
In summary, we did not observe consistent effects of the
specific location of the door/s in the scene. This may stem
from our univariate approach lacking the sensitivity to
detect what might be subtle differences in the visual input
(for similar results in fMRI, see the work of Bonner &
Epstein, 2017). To address this possibility, we conducted
a multivariate analysis, which allowed us to examine (a)
the extent to which both position-dependent and
position-invariant information can be extracted from the
EEG signal, and (2) the different time windows during
which the signals may be observed.
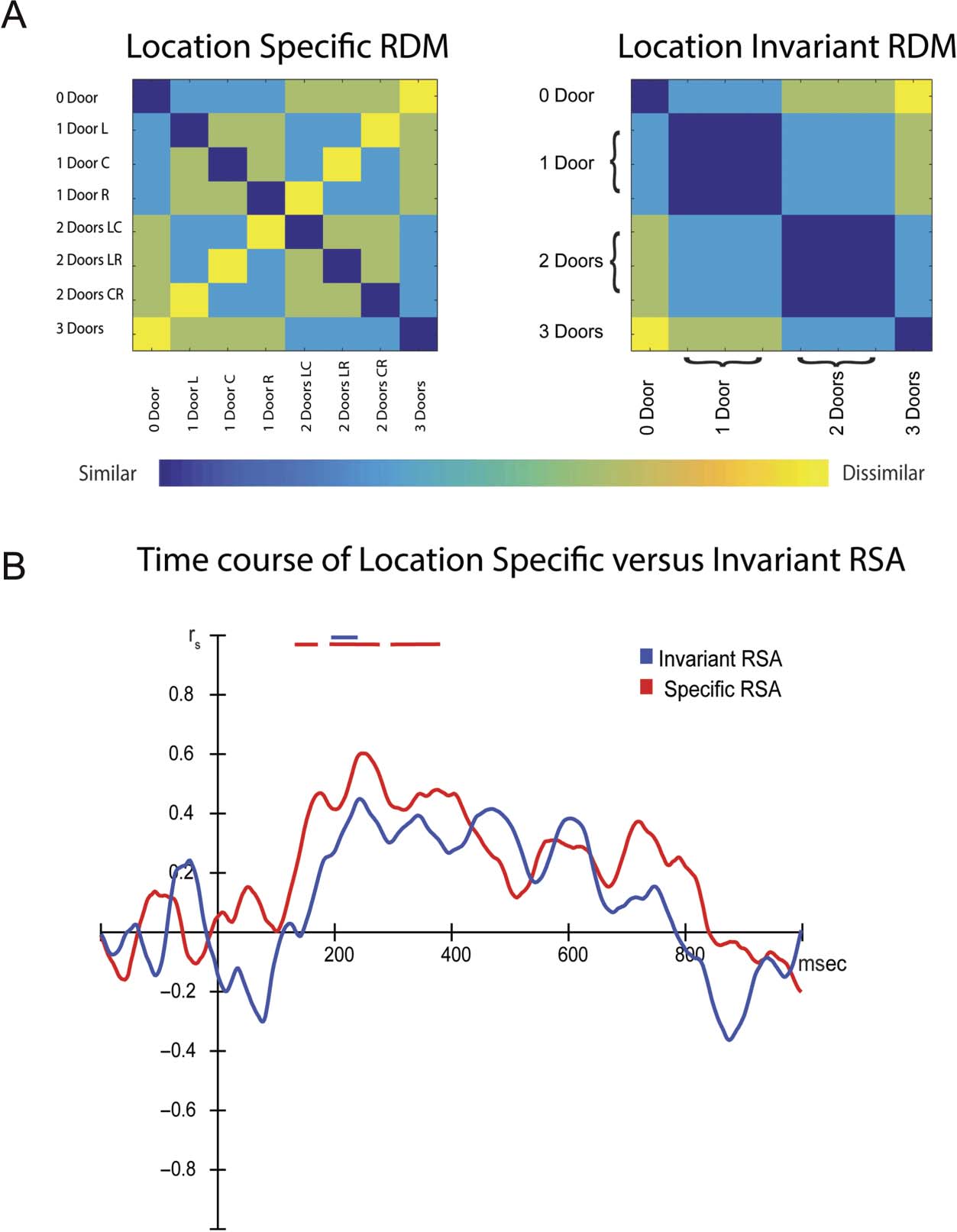
Multivariate Analysis
To determine the time course of location-specific and
location-invariant navigability-related processing, RSAs
were conducted by comparing the obtained neural RDMs
from each participant at each time point with two-model
RDMs, one corresponding to each hypothesis (see
Methods section and Figure 4A). The pattern of “activa-
tion” across electrodes within their averaged ERP
waveforms was correlated with differences between con-
ditions predicted by either or both of the two-model
RDM. This was followed by a cluster analysis across sub-
jects, allowing us to determine the time intervals in which
the data were best explained by one of the predicted
models. We found significant Spearman correlations
(Figure 4B) between the neural RDMs and the location-
specific RDM at several time windows: from 134–170 msec
(cluster-corrected values of p = .035, two-tailed), 193–
275 msec (cluster-corrected values of p = .0024, two-tailed),
and 295–380 msec (equivalent to all cluster-corrected
values of p = .004, two-tailed). Notably, the cluster analy-
ses also uncovered significant Spearman correlations
between the neural RDMs and location-invariant RDM,
from 196 to 237 msec (Figure 4B). Interestingly, this time
window corresponds with the P2 time window. Together,
the multivariate analyses suggest that local featural differ-
ences between stimuli (i.e., the number and location of
doors) in each navigability condition are processed as
early as 134 msec after stimulus onset, and more global
featural differences (0 doors, 1 door, 2 doors, 3 doors)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
o
c
n
_
a
_
0
1
8
1
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Results of the
multivariate analysis. (A) The
two-model RDM used for
the analyses, pertaining to the
two-alternative hypotheses:
location-specific processing,
in which the local featural
differences between conditions
(i.e., between doors and
paintings) influenced
processing (left column); and
location-invariant processing,
in which only the “global”
differences (i.e., between the
total number of doors)
influenced processing (right
column). (B) Time course of
decoding (measured using
Spearman correlation
coefficients), comparing the
obtained pattern of activation to
that predicted by the two
models: location-specific (blue)
and location-invariant (red).
Solid lines indicate significant
decoding ( p < .01, corrected:
see text for details).
404
Journal of Cognitive Neuroscience
Volume 34, Number 3
are most likely processed after local featural differences,
no earlier than 196 msec following stimulus onset.6
DISCUSSION
The current study provides novel evidence that the brain
codes information about the potential for navigation in the
scene as early as 200 msec after stimulus onset, and that
this coding involves both a global, position-invariant signal
about the overall navigability of the space and local infor-
mation regarding the positions of navigational pathways. A
standard univariate ERP analysis revealed that the ampli-
tude of the scene-selective P2 ERP component was higher
in response to images of rooms with no doors compared
to rooms with three doors, analogous to the higher P2
amplitude in response to closed relative to open scenes
reported previously (Harel et al., 2016, 2020; Hansen
et al., 2018). Furthermore, P2 peak amplitude scaled
linearly with the constraints on navigation: The more con-
straints on navigation (i.e., less doors in a room), the
higher was its amplitude. And although the effect of navi-
gability was most pronounced on the P2 ERP component,
the difference in amplitude between no-doors and three-
doors condition continued beyond the P2 time window,
lasting for additional 200 msec. Complementing these
findings, a multivariate analysis revealed that the P2 time
window contains significant information about naviga-
tional affordances: It is the first time period in which
position-invariant navigability information is extracted, in
addition to information regarding the location of the diag-
nostic feature (a specific door), which emerges earlier and
persists at this time window as well. Together, these find-
ings suggest that diagnostic information about the poten-
tial for navigation in a scene is present at the early stages of
visual processing and extracted as early as 220 msec after
stimulus onset.
Based on the current findings, we suggest that perceiv-
ing visual environments and navigating through them
should not necessarily be considered as two separate pro-
cesses, but rather as two points on a single continuum,
scene perception being the first step in a sequence of stages
that support navigation. At the neural level, this outlook
has two spatiotemporal corollaries. First, scene-selective
cortex should not only be engaged in the extraction of
scene-diagnostic features but should also carry informa-
tion about the potential for navigation in the scene.
Second, navigability-related neural activity should be
observed in the early stages of visual scene processing.
The current ERP study joins the original fMRI study, which
used the current scene stimuli (Bonner & Epstein, 2017)
to establish these two points. Bonner and Epstein (2017)
showed that information about the potential for move-
ment in the scenes is, indeed, represented in scene-
selective OPA.7 Our study shows that this same information
is represented as early as 200 msec poststimulus onset,
the same latency during which global properties of the
scene are extracted (Harel et al., 2016, 2020; Hansen
et al., 2018). Because the two studies use the exact scene
stimuli, they form a crucial link in connecting the spatial
and temporal aspects of perceptual processing of the
potential for action in scenes. Furthermore, in a follow-
up study, Bonner and Epstein (2018) reanalyzed their
imaging data and showed using deep convolutional net-
works that the affordance properties of scenes could be
represented through just a few stages of purely feedfor-
ward hierarchical computations, implying that computa-
tions of navigational affordances in OPA could be
achieved rapidly, in line with the current findings.
Given the limited temporal precision of fMRI, the func-
tional nature of the observed OPA activation cannot be
determined unequivocally; whereas OPA activity could
indeed reflect stimulus-driven processing of navigability
information available in the scene, it could also potentially
reflect recurrent feedback from posterior parietal cortex as
part of the occipito-parietal circuit (Kravitz, Saleem, et al.,
2011). Our current results suggest the former, rather than
the latter, alternative is more probable, as we show that
by 200 msec, sufficient evidence has accumulated for
determining the potential for navigation in the scene.
Moreover, the fact that the navigability effect on the
EEG manifests without any apparent need for encoding
or planning movement (see below) strengthens the
notion that navigational affordances are encoded manda-
torily, as originally suggested by Bonner and Epstein
(2017). It is important to note, however, that despite
the similarities between the studies, we cannot unequiv-
ocally conclude that OPA is the neural generator of the
observed effects of navigational affordances on the P2
component. We have not performed source localization
in the current study, as the relationship between ERP gen-
erator locations and scalp electrodes is complex (Nunez &
Srinivasan, 2006), and oftentimes the answers to which
cortical area generates a certain ERP effect could vary as
a function of the mathematical solution favored by the
researcher (for a comprehensive discussion, see the work
of Luck, 2014, Online Chapter 14; for an empirical demon-
stration of the limits of localization, see the work of
Petrov, 2012). Future research combining ERP and fMRI
(e.g., simultaneously recording ERPs in an MRI scanner;
see the work of Sadeh, Podlipsky, Zhdanov, & Yovel,
2010) is needed to determine the relationship between
OPA activity and the P2 sensitivity to navigationally rele-
vant information. Although the current data suggest that
navigational affordances are encoded rapidly, it is still an
open question just how rapid “rapid” is. Some recent
studies show earlier encoding of navigable space, specifi-
cally, around 100–120 msec poststimulus onset. A study
combining MEG and fMRI showed significant scene
boundary encoding (i.e., sensitivity to navigation-
constraining large-scale geometrical boundaries) in OPA
with corresponding MEG response patterns emerging as
early as 65 msec and peaking at about 100 msec poststim-
ulus onset (Henriksson et al., 2019). In a similar vein, dif-
ferential processing of closed and open scenes (which
Harel et al.
405
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
o
c
n
_
a
_
0
1
8
1
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
confer distinct navigational affordances, see below) has
been reported to manifest not only at the P2 time window
but also earlier, around 120 msec poststimulus onset
(Lowe et al., 2018). Findings from our multivariate analy-
ses support these studies demonstrating early coding of
spatial geometry. Information about the direction of
potential movement (i.e., specific door position) was
found to be represented as early as 140 msec poststimulus
onset, suggesting early encoding of navigational affor-
dances. Notably, position-invariant information, which is
perhaps more related to spatial expanse was found later,
at the P2 time window, between 200 and 240 msec. Con-
sequently, we suggest that spatial affordances are picked
up at different points in times, each characterized by sep-
arate neural representations: Location-specific affordance
information is encoded as early as 134 msec, persists until
170 msec, and re-emerges around 200 msec. This is
followed by more global scene representations that are
affected by the affordances of the space, namely, when
there are more affordances (i.e., less constraints on move-
ment overall), the global space becomes larger and more
“open.” This second stage reflects global processing of the
scene, entailing the integration of features across the
entire scene (for discussion, see the work of Harel et al.,
2020). As we note above, this later time window is coinci-
dent with the univariate results highlighting the sensitivity
of the P2 component to navigation-related information
(although the multivariate analyses detect the onset of this
effect slightly earlier than the peak analyses, this is to be
expected, given that the P2 only peaks around 230 msec).
Our focus on the P2 component, suggesting it is the
electrophysiological marker of navigational affordances,
follows our previous work highlighting the P2 as an index
of the processing of high-level global scene information.
The posterior P2 is the first ERP component to show evi-
dence of scene-selectivity, with higher amplitude to
scenes compared with faces and objects (Harel et al.,
2016), and the only visually evoked component to be mod-
ulated by scene inversion, as would be expected if global
scene information is indeed extracted during this period
(Harel & Al Zoubi, 2019). Furthermore, P2 amplitude is
sensitive to GSPs, such as spatial expanse (closed/open)
and naturalness (man-made/natural), and these effects
are automatic, evident across a variety of stimulus presen-
tation conditions, and are largely unperturbed by manipu-
lations of local texture (Harel et al., 2016, 2020; Hansen
et al., 2018). Together, these studies suggest that the
scene-selective P2 and the P2 time window in general
(approx. 200–250 msec) is indicative of and essential for
the processing of the global spatial structure of scenes
(see also the work of Kaiser, Häberle, & Cichy, 2020;
Kaiser, Turini, & Cichy, 2019; Cichy et al., 2017). Spatial
structure is used here broadly as an umbrella term to cap-
ture related concepts, such as scene layout, expanse, and
boundary. Notably, spatial structure is one of the key cat-
egories of GSPs proposed by Greene and Oliva (2009) to
be central for rapid scene categorization. According to
Greene and Oliva, rapid scene categorization is not pri-
marily mediated through objects and parts, but rather
through global, ecological properties that describe spatial
and functional aspects of scene space (such as navigability
or mean depth). Based on Greene and Oliva’s theory, we
hypothesized that closed and open scenes not only
describe the large-scale geometry of the scenes (i.e., its
spatial structure) but may also capture the functional prop-
erty of navigability. As such, closed and open scenes rep-
resent two ends of a continuum wherein enclosed spaces
posit more constraints on movement than open spaces.
And because closed scenes consistently evoke a stronger
neural response than open scenes (Harel et al., 2020;
Hansen et al., 2018; Lowe et al., 2018), we predicted that
scenes conferring more constraints on navigability should
evoke a higher P2 amplitude than scenes that confer less
such constraints. This prediction bore out, not only in its
dichotomous form (0-doors vs. 3-doors) but also as a con-
tinuous, parametric effect of navigability, evident in a lin-
ear decrease of P2 amplitude as a function of the number
of doors, as well as in a significant decoding of number of
doors independent of their local location around the P2
time window, 200–240 msec poststimulus onset. It is still
an open question to what extent the P2 exclusively indexes
global scene information, or whether it also incorporates
the processing of local scene information. Specifically, in
spite of the studies described above, we still found some
modulation of the P2 amplitude by door location (at least
for the two-door condition, see secondary univariate anal-
yses above), which may suggest it is not entirely indepen-
dent of local diagnostic information. The link, however,
between P2 amplitude and local information is not
straightforward, as no modulation was found for the single
door condition, and the modulation of the two-door con-
dition was not systematic and, furthermore, was not
related to hemisphere, as one might have expected given
a retinotopic mapping of the visual field. Future research
will be needed to elucidate the relationship between local
and global scene processing and how these processes
map onto the P2 amplitude.
Our study reveals another interesting characteristic of
the P2. We found that the P2 navigability effect was later-
alized to the right hemisphere; that is, the effect of the
number of doors on P2 amplitude was more pronounced
in the right than in the left hemisphere. This interaction
resonates and converges with previous ERP studies of
P2, which report laterality effects on the P2 amplitude,
with overall higher amplitude in the right hemisphere,
as well as specific GSPs being more discriminable in the
right hemisphere. Specifically, both scene naturalness
and spatial expanse were reported to have a greater effect
in the right hemisphere (Harel et al., 2016, 2020; Hansen
et al., 2018). The finding that the right hemisphere is more
involved than the left in the processing of global scene
information extends previous research on local/global
processing and hemispheric asymmetries and is in line
with the long-lived proposal that the right hemisphere
406
Journal of Cognitive Neuroscience
Volume 34, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
o
c
n
_
a
_
0
1
8
1
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
specializes in global processing (Brederoo, Nieuwenstein,
Lorist, & Cornelissen, 2017; Flevaris & Robertson, 2016;
but see the work of Wiesmann, Friederici, Singer, &
Steinbeis, 2020). Thus, the right hemisphere advantage
for GSP effects further supports our proposal that P2
indexes the processing of the spatial structure of the scene
via the processing of global scene information.
Our findings point to the putative connection between
spatial structure and navigability, which, arguably, may
serve as the mechanism by which perceptual information
is transformed into action-relevant information. Given the
close link between spatial structure and navigability, our
findings raise two intriguing questions. First, are navigabil-
ity and spatial structure (or spatial expanse, in the case of
closed vs. open scenes) one and the same thing? Can the
two constructs be used synonymously, or are they two
independent dimensions? Second, given that they are
independent, does the neural encoding of navigational
affordances reflect intermediate levels of representation
(i.e., GSPs), low-level image statistics (Groen, Silson, &
Baker, 2017) or rather the extraction of higher-level eco-
logical scene properties (for discussion, see the work of
Malcolm, Groen, & Baker, 2016)? Our current design can-
not directly address these questions, as we only used
closed indoor scenes (rooms). However, future research
will be able to shed further light on this issue. To test
the independence of the two dimensions, one would have
to vary the amount of movement a scene affords, in both
closed and open scenes. If spatial expanse and navigability
are independent, then varying navigability should have a
similar effect on the neural responses to open scenes (this
could be manipulated, e.g., by adding an increasing num-
ber of obstacles like boulders, or by varying the number of
paths in an open field). In addition, an alternative, more
naturalistic approach could use a large set of real-world
scene images (instead of highly controlled artificial
scenes), in which people would rank these scenes on both
spatial expanse (Zhang et al., 2019) and navigability
(Bonner & Epstein, 2017, Experiment 2) and then in a sep-
arate electrophysiological study integrate these rankings
with the neural response patterns to determine the rela-
tive contribution of each dimension and other image prop-
erties to the neural representations using computational
model-based approaches (e.g., Lescroart & Gallant,
2019; Bonner & Epstein, 2018; Cichy, Kholsa, Pantazis, &
Oliva, 2017; Lescroart, Stansbury, & Gallant, 2015).
At a broader theoretical level, our results support a tight
link between perception and action, a hallmark of senso-
rimotor and embodied cognition theories (e.g., Jelić, Tieri,
De Matteis, Babiloni, & Vecchiato, 2016; Wilson, 2002;
Clark, 1999). One aspect of these theories in the context
of navigation is the constant need for visual updating when
one explores an environment to minimize prediction error
(Kaplan & Friston, 2018; Hassabis & Maguire, 2009; Kurby
& Zacks, 2008; Zacks, Speer, Swallow, Braver, & Reynolds,
2007). The idea is that exploring the environment requires
continuously modeling the potential outcomes of the
intended action, with affordances serving this function
by constraining visual perception to reflect experience-
dependent, observer-relevant information (Sestito, Flach,
& Harel, 2018). At the neural level, this should translate to
dynamic changes in continuous activity in early visual
areas as one walks around and explores their surround-
ings, even before execution of action (e.g., movement;
Jelić et al., 2016). In line with this idea, a recent study using
mobile brain/body imaging technology in which partici-
pants actively walked in a highly engaging immersive
virtual environment demonstrated that environmental
affordances are extracted and encoded throughout the
entirety of the act of exploring one’s surroundings, starting
from early perceptual stages (P1-N1 complex) all the way
to motor planning and execution (Djebbara, Fich, Petrini,
& Gramann, 2019). Notably, our study adds several key
observations to these findings. First, ambulatory, active,
and continuous exploration of the environment is not
necessary for observing electrophysiological responses
representing the extraction of navigational affordances.
Participants in our study were stationary, sitting in front
of a computer screen, and watched briefly presented,
minimalistic room images. The fact that one can observe
similar electrophysiological responses as a function of nav-
igability even without any movement or the presence of an
immersive environment implies that scene affordances are
extracted across multiple contexts and task demands (for a
similar finding with spatial expanse, see the work of
Hansen et al., 2018). Arguably, a lifetime of experience
navigating in the world results in automatic activation of
sensorimotor scene representations when presented with
visual environments, even if these environments are
sparse, minimalistic scenes deprived of rich detail (for
the role of experience in perceiving novel scene affor-
dances, see the work of Sestito, Harel, Nador, & Flach,
2018). The idea that navigational affordances are pro-
cessed mandatorily is also supported by the finding that
navigability effects are evident even without any explicit
task-context or relevant task demands, as participants in
our study were not required to either move about the envi-
ronment, imagine themselves moving about it, or make
any explicit judgments regarding its potential for naviga-
tion. The fact that we found navigability-based modulations
although no movement in space was performed, nor was
movement in space directly relevant to the task, supports
the conclusion that extracting navigational affordances is
rapid, mandatory, and task-independent. Furthermore,
the similar patterns of results between the two studies sug-
gest that our current laboratory-based findings are likely to
generalize to real-world, realistic settings and thus expand
their validity and utility for future research.
In summary, this study demonstrates that navigational
affordances are extracted at the early, perceptual stages of
visual scene processing, suggesting a close link between
scene perception and navigation. Information about the
potential for navigation in the scene is extracted rapidly
and automatically, without any explicit task or movement
Harel et al.
407
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
o
c
n
_
a
_
0
1
8
1
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
requirements. Complementing prior neuroimaging stud-
ies showing OPA encodes the structure of navigable
space, the current work establishes the temporal dynam-
ics of processing navigational affordances. Significant nav-
igability information is present in two time windows: an
early time window sensitive to the specific position of
navigability diagnostic stimulus features, and a later
one, which incorporates both position-specific and
position-invariant information. The later time window
overlaps with the univariate P2 ERP component, reflect-
ing global processing of scene structure. Finally, the cur-
rent findings are in line with sensorimotor accounts of
perception, suggesting that perceiving visual environ-
ments and navigating through them should not necessar-
ily be considered as two separate processes, but rather as
two integrated processes.
Reprint requests should be sent to Assaf Harel, Department of
Psychology, Wright State University, 335 Fawcett Hall, 3640 Col.
Glenn Highway, Dayton, Ohio 45435, or via e-mail: assaf
.harel@wright.edu.
4. The reason this analysis was considered secondary is that
we did not expect to find door location effects in the univariate
analysis based on Bonner and Epstein’s (2017) fMRI study. No
robust effects of door location were observed in that study in
the univariate response magnitude analysis, while multivariate
analysis of response patterns did not show sufficient sensitivity
to report such effects.
5. Note that whereas in the 1-door condition, the comparison
between conditions is relatively straightforward with door loca-
tion being the diagnostic cue for movement, in the 2-doors con-
ditions, this is less obvious, as the “odd one out” is not the
door, but rather its absence—a single painting. This difficulty
in interpretation is further exacerbated in the case of no signif-
icant interactions with Hemisphere, meaning that painting loca-
tion is not the consistent source of the effect.
6.
It should be noted, however, that significant clusters do not
contain contiguous intervals of significant Spearman correlation
coefficients; rather, clusters denote the intervals within which
there is a conditional probability of p that the interval contains
at least one significant coefficient, given the Type 1 error rate
(cluster induction parameter) for evaluating the coefficients
independently.
7. Notably, Bonner and Epstein (2017) found OPA sensitivity
to navigability information not only using current stimuli, which
are computer-generated, but also using naturalistic images of
indoor scenes.
Diversity in Citation Practices
REFERENCES
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender identi-
fication of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance.
Notes
1. The link between scene recognition and navigation reso-
nates the putative link between perception and action pro-
posed by sensorimotor accounts of perception, which posit that
the potential for action in the environment, also known as affor-
dances, is conveyed by the visual stimulus itself (e.g., Gibson,
1979).
2. This is in fact an inherent limitation of fMRI because of its
low temporal resolution. For a more general discussion on the
inferential challenges of fMRI, see the work of Ghuman and
Martin (2019).
3. The extent to which these early signatures reflect the
extraction of local image statistics or more global scene proper-
ties is still an open question. For instance, scene inversion
effects are only observed around 220–250 msec poststimulus
onset (Kaiser, Häberle, & Cichy, 2020; Harel & Al Zoubi,
2019), consistent with the idea that global scene structure infor-
mation is extracted later than 100 msec.
Benjamini, Y., & Hochberg, Y. (2000). On the adaptive
control of the false discovery rate in multiple testing with
independent statistics. Journal of Educational and
Behavioral Statistics, 25, 60–83. https://doi.org/10.3102
/10769986025001060
Bonner, M. F., & Epstein, R. A. (2017). Coding of navigational
affordances in the human visual system. Proceedings of the
National Academy of Sciences, U.S.A., 114, 4793–4798.
https://doi.org/10.1073/pnas.1618228114, PubMed: 28416669
Bonner, M. F., & Epstein, R. A. (2018). Computational
mechanisms underlying cortical responses to the affordance
properties of visual scenes. PLoS Computational Biology,
14, e1006111. https://doi.org/10.1371/journal.pcbi.1006111,
PubMed: 29684011
Brederoo, S. G., Nieuwenstein, M. R., Lorist, M. M., &
Cornelissen, F. W. (2017). Hemispheric specialization for
global and local processing: A direct comparison of linguistic
and non-linguistic stimuli. Brain and Cognition, 119, 10–16.
https://doi.org/10.1016/j.bandc.2017.09.005, PubMed:
28923763
Cichy, R. M., Khosla, A., Pantazis, D., & Oliva, A. (2017).
Dynamics of scene representations in the human brain
revealed by magnetoencephalography and deep neural
networks. Neuroimage, 153, 346–358. https://doi.org/10.1016
/j.neuroimage.2016.03.063, PubMed: 27039703
Clark, A. (1999). An embodied cognitive science? Trends in
Cognitive Sciences, 3, 345–351. https://doi.org/10.1016/S1364
-6613(99)01361-3
Dilks, D. D., Julian, J. B., Kubilius, J., Spelke, E. S., & Kanwisher,
N. (2011). Mirror-image sensitivity and invariance in object
and scene processing pathways. Journal of Neuroscience, 31,
11305–11312. https://doi.org/10.1523/JNEUROSCI.1935-11
.2011, PubMed: 21813690
Dilks, D. D., Julian, J. B., Paunov, A. M., & Kanwisher, N. (2013).
The occipital place area is causally and selectively involved in
scene perception. Journal of Neuroscience, 33, 1331–1336.
https://doi.org/10.1523/JNEUROSCI.4081-12.2013, PubMed:
23345209
408
Journal of Cognitive Neuroscience
Volume 34, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
o
c
n
_
a
_
0
1
8
1
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Djebbara, Z., Fich, L. B., Petrini, L., & Gramann, K. (2019).
Sensorimotor brain dynamics reflect architectural
affordances. Proceedings of the National Academy of
Sciences, U.S.A., 116, 14769–14778. https://doi.org/10.1073
/pnas.1900648116, PubMed: 31189596
Ekstrom, A. D. (2015). Why vision is important to how we
navigate. Hippocampus, 25, 731–735. https://doi.org/10.1002
/hipo.22449, PubMed: 25800632
Ekstrom, A. D., Spiers, H. J., Bohbot, V. D., & Rosenbaum, R. S.
(2018). Human spatial navigation. Princeton University
Press. https://doi.org/10.2307/j.ctvc773wg
Epstein, R. A., Higgins, J. S., Jablonski, K., & Feiler, A. M. (2007).
Visual scene processing in familiar and unfamiliar
environments. Journal of Neurophysiology, 97, 3670–3683.
https://doi.org/10.1152/jn.00003.2007, PubMed: 17376855
Epstein, R. A., Higgins, J. S., & Thompson-Schill, S. L. (2005).
Learning places from views: Variation in scene processing as a
function of experience and navigational ability. Journal of
Cognitive Neuroscience, 17, 73–83. https://doi.org/10.1162
/0898929052879987, PubMed: 15701240
Flevaris, A. V., & Robertson, L. C. (2016). Spatial frequency
selection and integration of global and local information in
visual processing: A selective review and tribute to Shlomo
Bentin. Neuropsychologia, 83, 192–200. https://doi.org/10
.1016/j.neuropsychologia.2015.10.024, PubMed: 26485158
Ghuman, A. S., & Martin, A. (2019). Dynamic neural
representations: An inferential challenge for fMRI. Trends in
Cognitive Sciences, 23, 534–536. https://doi.org/10.1016/j.tics
.2019.04.004, PubMed: 31103440
Gibson, J. J. (1979). The ecological approach to visual
perception. Boston, MA: Houghton, Mifflin and Company.
Greene, M. R., & Oliva, A. (2009). The briefest of glances the
time course of natural scene understanding. Psychological
Science, 20, 464–472. https://doi.org/10.1111/j.1467-9280
.2009.02316.x, PubMed: 19399976
Groen, I. I. A., Ghebreab, S., Lamme, V. A. F., & Scholte, H. S.
(2016). The time course of natural scene perception with
reduced attention. Journal of Neurophysiology, 115, 931–946.
https://doi.org/10.1152/jn.00896.2015, PubMed: 26609116
Groen, I. I. A., Ghebreab, S., Prins, H., Lamme, V. A., & Scholte,
H. S. (2013). From image statistics to scene gist: Evoked
neural activity reveals transition from low-level natural image
structure to scene category. Journal of Neuroscience, 33,
18814–18824. https://doi.org/10.1523/JNEUROSCI.3128-13
.2013, PubMed: 24285888
Groen, I. I. A., Silson, E. H., & Baker, C. I. (2017). Contributions
of low- and high-level properties to neural processing of
visual scenes in the human brain. Philosophical
Transactions of the Royal Society of London, Series B:
Biological Sciences, 372, 20160102. https://doi.org/10.1098
/rstb.2016.0102, PubMed: 28044013
Hansen, N. E., Noesen, B. T., Nador, J. D., & Harel, A. (2018).
The influence of behavioral relevance on the processing of
global scene properties: An ERP study. Neuropsychologia,
114, 168–180. https://doi.org/10.1016/j.neuropsychologia
.2018.04.040, PubMed: 29729276
Harel, A., & Al Zoubi, H. (2019). Early electrophysiological
correlates of scene perception are sensitive to inversion.
Journal of Vision, 19, 190. https://doi.org/10.1167/19.10.190
Harel, A., Groen, I. I. A., Kravitz, D. J., Deouell, L. Y., &
Baker, C. I. (2016). The temporal dynamics of scene
processing: A multifaceted EEG investigation. eNeuro, 3,
ENEURO-0139-16.2016. https://doi.org/10.1523/ENEURO
.0139-16.2016, PubMed: 27699208
Harel, A., Mzozoyana, M. W., Al Zoubi, H., Nador, J. D., Noesen,
B. T., Lowe, M. X., et al. (2020). Artificially-generated scenes
demonstrate the importance of global scene properties for
scene perception. Neuropsychologia, 141, 107434. https://
doi.org/10.1016/j.neuropsychologia.2020.107434, PubMed:
32179102
Hassabis, D., & Maguire, E. A. (2009). The construction system
of the brain. Philosophical Transactions of the Royal Society
of London, Series B: Biological Sciences, 364, 1263–1271.
https://doi.org/10.1098/rstb.2008.0296, PubMed: 19528007
Hasson, U., Harel, M., Levy, I., & Malach, R. (2003). Large-scale
mirror-symmetry organization of human occipito-temporal
object areas. Neuron, 37, 1027–1041. https://doi.org/10.1016
/S0896-6273(03)00144-2, PubMed: 12670430
Henriksson, L., Mur, M., & Kriegeskorte, N. (2019). Rapid
invariant encoding of scene layout in human OPA. Neuron,
103, 161–171. https://doi.org/10.1016/j.neuron.2019.04.014,
PubMed: 31097360
Jelić, A., Tieri, G., Me Matteis, F., Babiloni, F., & Vecchiato, G.
(2016). The enactive approach to architectural experience: A
neurophysiological perspective on embodiment, motivation,
and affordances. Frontiers in Psychology, 7, 481. https://doi
.org/10.3389/fpsyg.2016.00481, PubMed: 27065937
Julian, J. B., Keinath, A. T., Marchette, S. A., & Epstein, R. A.
(2018). The neurocognitive basis of spatial reorientation.
Current Biology, 28, R1059–R1073. https://doi.org/10.1016/j
.cub.2018.04.057, PubMed: 30205055
Julian, J. B., Ryan, J., Hamilton, R. H., & Epstein, R. A. (2016).
The occipital place area is causally involved in representing
environmental boundaries during navigation. Current
Biology, 26, 1104–1109. https://doi.org/10.1016/j.cub.2016.02
.066, PubMed: 27020742
Jung, T.-P., Humphries, C., Lee, T.-W., Makeig, S., McKeown,
M. J., Iragui, V., et al. (1998). Extended ICA removes artifacts
from electroencephalographic recordings. Advances in
Neural Information Processing Systems, 10, 894–900.
Kaiser, D., Häberle, G., & Cichy, R. M. (2020). Cortical
sensitivity to natural scene structure. Human Brain
Mapping, 41, 1286–1295. https://doi.org/10.1002/hbm.24875,
PubMed: 31758632
Kaiser, D., Turini, J., & Cichy, R. M. (2019). A neural mechanism
for contextualizing fragmented inputs during naturalistic
vision. eLife, 8, e48182. https://doi.org/10.7554/eLife.48182,
PubMed: 31596234
Kamps, F. S., Julian, J. B., Kubilius, J., Kanwisher, N., & Dilks,
D. D. (2016). The occipital place area represents the local
elements of scenes. Neuroimage, 132, 417–424. https://doi
.org/10.1016/j.neuroimage.2016.02.062, PubMed: 26931815
Kamps, F. S., Lall, V., & Dilks, D. D. (2016). The occipital place
area represents first-person perspective motion information
through scenes. Cortex, 83, 17–26. https://doi.org/10.1016/j
.cortex.2016.06.022, PubMed: 27474914
Kaplan, R., & Friston, K. J. (2018). Planning and navigation as
active inference. Biological Cybernetics, 112, 323–343. https://
doi.org/10.1007/s00422-018-0753-2, PubMed: 29572721
Kravitz, D. J., Peng, C. S., & Baker, C. I. (2011). Real-world scene
representations in high-level visual cortex: It’s the spaces
more than the places. Journal of Neuroscience, 31,
7322–7333. https://doi.org/10.1523/JNEUROSCI.4588-10
.2011, PubMed: 21593316
Kravitz, D. J., Saleem, K. S., Baker, C. I., & Mishkin, M. (2011). A
new neural framework for visuospatial processing. Nature
Reviews Neuroscience, 12, 217–230. https://doi.org/10.1038
/nrn3008, PubMed: 21415848
Harel, A., Kravitz, D. J., & Baker, C. I. (2013). Deconstructing
Kurby, C. A., & Zacks, J. M. (2008). Segmentation in the
visual scenes in cortex: Gradients of object and spatial layout
information. Cerebral Cortex, 23, 947–957. https://doi.org/10
.1093/cercor/bhs091, PubMed: 22473894
perception and memory of events. Trends in Cognitive
Sciences, 12, 72–79. https://doi.org/10.1016/j.tics.2007.11.004,
PubMed: 18178125
Harel et al.
409
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
o
c
n
_
a
_
0
1
8
1
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lescroart, M. D., & Gallant, J. L. (2019). Human scene-selective
areas represent 3D configurations of surfaces. Neuron, 101,
178–192. https://doi.org/10.1016/j.neuron.2018.11.004,
PubMed: 30497771
Lescroart, M. D., Stansbury, D. E., & Gallant, J. L. (2015). Fourier
power, subjective distance, and object categories all provide
plausible models of BOLD responses in scene-selective
visual areas. Frontiers in Computational Neuroscience, 9,
135. https://doi.org/10.3389/fncom.2015.00135, PubMed:
26594164
Lowe, M. X., Rajsic, J., Ferber, S., & Walther, D. B. (2018).
Discriminating scene categories from brain activity within 100
milliseconds. Cortex, 106, 275–287. https://doi.org/10.1016/j
.cortex.2018.06.006, PubMed: 30037637
Luck, S. J. (2014). An introduction to the event-related
potential technique (2nd ed.). MIT Press.
Malcolm, G. L., Groen, I. I. A., & Baker, C. I. (2016). Making
sense of real-world scenes. Trends in Cognitive Sciences, 20,
843–856. https://doi.org/10.1016/j.tics.2016.09.003, PubMed:
27769727
Nakamura, K., Kawashima, R., Sato, N., Nakamura, A., Sugiura,
M., Kato, T., et al. (2000). Functional delineation of the
human occipito-temporal areas related to face and scene
processing: A PET study. Brain, 123, 1903–1912. https://doi
.org/10.1093/brain/123.9.1903, PubMed: 10960054
Nunez, P. L., & Srinivasan, R. (2006). Electric fields of the brain:
The neurophysics of EEG. Oxford University Press. https://doi
.org/10.1093/acprof:oso/9780195050387.001.0001
Persichetti, A. S., & Dilks, D. D. (2016). Perceived egocentric
distance sensitivity and invariance across scene-selective
cortex. Cortex, 77, 155–163. https://doi.org/10.1016/j.cortex
.2016.02.006, PubMed: 26963085
Persichetti, A. S., & Dilks, D. D. (2018). Dissociable neural
systems for recognizing places and navigating through them.
Journal of Neuroscience, 38, 10295–10304. https://doi.org/10
.1523/JNEUROSCI.1200-18.2018, PubMed: 30348675
Petrov, Y. (2012). Harmony: EEG/MEG linear inverse source
reconstruction in the anatomical basis of spherical
harmonics. PLoS One, 7, e44439. https://doi.org/10.1371
/journal.pone.0044439, PubMed: 23071497
Pinhas, M., Tzelgov, J., & Ganor-Stern, D. (2012). Estimating
linear effects in ANOVA designs: The easy way. Behavior
Research Methods, 44, 788–794. https://doi.org/10.3758
/s13428-011-0172-y, PubMed: 22101656
Rousselet, G., Joubert, O., & Fabre-Thorpe, M. (2005). How
long to get to the “gist” of real-world natural scenes?
Visual Cognition, 12, 852–877. https://doi.org/10.1080
/13506280444000553
Sadeh, B., Podlipsky, I., Zhdanov, A., & Yovel, G. (2010).
Event-related potential and functional MRI measures of
face-selectivity are highly correlated: A simultaneous
ERP-fMRI investigation. Human Brain Mapping, 31,
1490–1501. https://doi.org/10.1002/hbm.20952, PubMed:
20127870
Sato, N., Nakamura, K., Nakamura, A., Sugiura, M., Ito, K.,
Fukuda, H., et al. (1999). Different time course between
scene processing and face processing: A MEG study.
NeuroReport, 10, 3633–3637. https://doi.org/10.1097
/00001756-199911260-00031, PubMed: 10619657
Sestito, M., Flach, J., & Harel, A. (2018). Grasping the world
from a cockpit: Perspectives on embodied neural
mechanisms underlying human performance and
ergonomics in aviation context. Theoretical Issues in
Ergonomics Science, 19, 692–711. https://doi.org/10.1080
/1463922X.2018.1474504
Sestito, M., Harel, A., Nador, J., & Flach, J. (2018). Investigating
neural sensorimotor mechanisms underlying flight expertise
in pilots: Preliminary data from an EEG study. Frontiers in
Human Neuroscience, 12, 489. https://doi.org/10.3389
/fnhum.2018.00489, PubMed: 30618676
Walther, D. B., Caddigan, E., Fei-Fei, L., & Beck, D. M. (2009).
Natural scene categories revealed in distributed patterns of
activity in the human brain. Journal of Neuroscience, 29,
10573–10581. https://doi.org/10.1523/JNEUROSCI.0559-09
.2009, PubMed: 19710310
Walther, D. B., Chai, B., Caddigan, E., Beck, D. M., & Fei-Fei, L.
(2011). Simple line drawings suffice for functional MRI
decoding of natural scene categories. Proceedings of the
National Academy of Sciences, U.S.A., 108, 9661–9666.
https://doi.org/10.1073/pnas.1015666108, PubMed: 21593417
Wiesmann, C. G., Friederici, A. D., Singer, T., & Steinbeis, N.
(2020). Two systems for thinking about others’ thoughts in
the developing brain. Proceedings of the National Academy
of Sciences, U.S.A., 117, 6928–6935. https://doi.org/10.1073
/pnas.1916725117, PubMed: 32152111
Wilson, M. (2002). Six views of embodied cognition.
Psychonomic Bulletin & Review, 9, 625–636. https://doi.org
/10.3758/BF03196322, PubMed: 12613670
Zacks, J. M., Speer, N. K., Swallow, K. M., Braver, T. S., &
Reynolds, J. R. (2007). Event perception: A mind–brain
perspective. Psychological Bulletin, 133, 273–293. https://doi
.org/10.1037/0033-2909.133.2.273, PubMed: 17338600
Zhang, H., Houpt, J. W., & Harel, A. (2019). Establishing
reference scales for scene naturalness and openness.
Behavior Research Methods, 51, 1179–1186. https://doi.org
/10.3758/s13428-018-1053-4, PubMed: 29845553
410
Journal of Cognitive Neuroscience
Volume 34, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
3
9
7
1
9
8
5
0
1
9
/
j
o
c
n
_
a
_
0
1
8
1
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3