Competencia sesgada por el valor en el
Auditory System of the Brain
Andy J. kim , Laurent Grégoire, and Brian A. anderson
Abstracto
■ Attentional capture by previously reward-associated stimuli has
predominantly been measured in the visual domain. Recientemente,
behavioral studies of value-driven attention have demonstrated
involuntary attentional capture by previously reward-associated
sounds, emulating behavioral findings within the visual domain
and suggesting a common mechanism of attentional capture by
value across sensory modalities. Sin embargo, the neural correlates
of the modulatory role of learned value on the processing of
auditory information has not been examined. Aquí, we conducted
a neuroimaging study on human participants using a previously
established behavioral paradigm that measures value-driven
attention in an auditory target identification task. We replicate
behavioral findings of both voluntary prioritization and involun-
tary attentional capture by previously reward-associated sounds.
When task-relevant, the selective processing of high-value sounds
is supported by reduced activation in the dorsal attention net-
work of the visual system (FEF, intraparietal sulcus, right middle
frontal gyrus), implicating cross-modal processes of biased com-
petition. When task-irrelevant, in contrast, high-value sounds
evoke elevated activation in posterior parietal cortex and are
represented with greater fidelity in the auditory cortex. Nuestro
findings reveal two distinct mechanisms of prioritizing reward-
related auditory signals, with voluntary and involuntary modes of
orienting that are differently manifested in biased competition. ■
INTRODUCCIÓN
Attention can be biased toward features that align with
task goals (p.ej., Folk, Remington, & Johnston, 1992;
lobo, Cave, & Franzel, 1989), physically salient stimuli
(p.ej., Theeuwes, 1991, 1992), and stimuli that have pre-
viously been prioritized by attention, now commonly
referred to as selection history (p.ej., Awh, Belopolsky, &
Theeuwes, 2012). Within the context of selection history,
reward-associated stimuli receive elevated attentional pri-
ority (p.ej., Hickey, Chelazzi, & Theeuwes, 2010; Della
Libera & Chelazzi, 2006), and such attentional biases can
persist even when previously reward-associated stimuli are
nonsalient and task-irrelevant as has been demonstrated
using the value-driven attentional capture ( VDAC) para-
digm (anderson, Laurent, & Yantis, 2011). The influence
of reward history on attention has primarily been investi-
gated in the visual domain (see works of Anderson, 2016a,
2019, for reviews), and a mechanistic understanding of
learning-dependent attentional biases in other sensory
systems is limited.
Cross-modal designs have investigated interactions
across multiple sensory networks in attentional processing,
commonly to understand how bimodal stimulation is
processed and integrated (p.ej., Stormer, McDonald, &
Hillyard, 2009; McDonald, Teder-Salejarvi, Di Russo, &
Hillyard, 2005; McDonald, Teder-Salejarvi, & Hillyard,
Texas A&Universidad M
© 2021 Instituto de Tecnología de Massachusetts
2000). Behavioral evidence suggests that reward associa-
tions can influence how competition between sensory
modalities is resolved (Sanz, Vuilleumier, & Bourgeois,
2018; anderson, 2016b). Sin embargo, recent evidence sug-
gests that the reward value of visual stimuli can dominate
over that of auditory stimuli when in direct competition
(cheng, Saglam, Andre, & Pooresmaeili, 2020), highlight-
ing the importance of measuring attentional biases specif-
ically in the auditory domain to uncover the full breadth of
the underlying mechanisms.
In a task using only auditory stimuli, attention has been
shown to be preferentially allocated to task-relevant audi-
tory streams as a function of reward history (Asutay &
Västfjäll, 2016). We have expanded on these initial find-
ings by showing that previously reward-associated but
currently task-irrelevant auditory stimuli interfere with
auditory target identification (kim, Sotavento, & anderson,
2021), demonstrating involuntary attentional capture by
previously reward-associated sounds. This result provides
a parallel to demonstrations of VDAC in the visual domain
(anderson, 2016a; Anderson et al., 2011) using exclusively
auditory stimuli. Such converging behavioral evidence
suggests a common principle of involuntary attentional
prioritization of previously reward-associated stimuli
across sensory modalities. Sin embargo, the neural mecha-
nisms supporting such value-based prioritization in the
auditory system have not been clarified.
The neurobiology of value-driven attention has been
widely investigated in the visual domain, consistently
Revista de neurociencia cognitiva 34:1, páginas. 180–191
https://doi.org/10.1162/jocn_a_01785
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
oh
C
norte
_
a
_
0
1
7
8
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
revealing a value-driven attention network of regions in
the brain in which high-value stimuli evoke elevated re-
sponses, including the early and ventral visual cortex, posición-
terior parietal cortex, and caudate tail (see the work of
anderson, 2019, para una revisión). Además, value has
been shown to modulate the amplitude of stimulus-
evoked activity and tune neuronal population profiles in
favor of more valuable stimuli within the spatially selective
areas of the early visual cortex (Itthipuripat, Vo, sprague,
& serencias, 2019; serencias & Saproo, 2010; serencias,
2008). In electrophysiological studies, reward-associated
sounds have been shown to produce an elevated N1 re-
sponse over auditory cortex (Folyi & Wentura, 2019; Folyi,
Liesefeld, & Wentura, 2016), suggesting a potential paral-
lel to these findings in the visual domain, although the
stimulus specificity of this response and the contribution
of other brain regions to it remain unexplored. The influ-
ence of reward on auditory processing has been robustly
examined in nonhuman species (see works of Irvine,
2018; Kraus & White-Schwoch, 2015, for reviews). Reward
has been shown to influence neural responses in the
auditory cortex as a function of value in ferrets (David,
Fritz, & Shamma, 2012) and nonhuman primates (Brosch,
Selezneva, & Scheich, 2011). Además, an fMRI study
on rhesus macaques demonstrated that such reward-
associated activity in the auditory cortex interacts with
neural structures that are associated with dopaminergic
(nucleus accumbens) and cholinergic (nucleus basalis)
pathways (Wikman, Rinne, & Petkov, 2019). Interest-
ingly, in humans, listening to music has consistently been
shown to engage neural networks of reward via the dopa-
minergic system, further supporting the role of projec-
tions between the limbic system and the auditory
cortex in representing the value of sounds (p.ej., Ferreri
et al., 2019; Gold, Pearce, Mas-Herrero, Dagher, & Zatorre,
2019; Salimpoor, Zald, Zatorre, Dagher, & McIntosh, 2015;
Salimpoor et al., 2013). Sin embargo, these investigations
have been limited to the role of reward in auditory per-
ception or evaluation, and the modulatory influence of re-
ward on the cognitive processing of auditory stimuli (p.ej.,
control atencional) in humans is yet unclear.
en este estudio, we conducted an fMRI experiment using
our previously established behavioral paradigm (kim
et al., 2021) to elucidate the neural correlates of value-
driven auditory attention. An advantage of this paradigm,
unlike paradigms frequently used to investigate value-
driven attention in the visual domain (p.ej., anderson &
Halpern, 2017; anderson, 2016a; Anderson et al., 2011),
is that robust effects of reward on attention are evident in
response to both task-relevant and task-irrelevant stimuli,
permitting measurement of reward’s influence on both
voluntary and involuntary attention. The neural correlates
of the processing of different sounds as a function of
learned value was probed both as a function of the am-
plitude of stimulus-evoked response as well as the infor-
mation contained within the pattern of activation as
revealed by multivariate pattern analysis (MVPA).
MÉTODOS
Participantes
In our previous behavioral study, a sample size of 35 par-
ticipants found significant effects of response time (RT)
in both the training and test phases (Kim y cols., 2021). A
replicate these results, we proposed to again recruit 35
participants for this study. This sample size is similar to
y, in most cases, exceeds prior fMRI studies of VDAC
(p.ej., kim & anderson, 2019b, 2020a, 2020b; Itthipuripat
et al., 2019; anderson, 2017; Barbaro, Peelen, & Hickey,
2017; Anderson et al., 2016; anderson, Laurent, & Yantis,
2014; Hickey et al., 2010). Thirty-nine participants were
recruited from the university community. All participants
were English-speaking and reported normal or corrected-
to-normal visual acuity and normal color vision. Sin embargo,
two participants did not meet the safety eligibility criteria
for neuroimaging and two participants voluntarily with-
drew from the study before completion. De este modo, 35 partícipe-
ipants were fully scanned and completed the experiment.
Following data exclusion procedures (see Data Analysis
and Exclusion Criteria section), we analyzed 31 complete
behavioral and neuroimaging data sets (13 women, siglos
18–35 [m = 23.1 años, DE = 4.0 años]).
Ethics Statement
All procedures were approved by the university institu-
tional review board and were conducted in accordance
with the principles expressed in the Declaration of
Helsinki. Written informed consent was obtained for
cada participante.
Experimental Procedure
Participants were scheduled for an initial in-laboratory
visit for 1 hr, and each eligible participant underwent
fMRI in a single 1.5-hr session at the scan center on the
following day. During their initial appointment, participar-
pants came into the laboratory for consenting, MRI safety
screening, and screening for adequate performance on
the behavioral task. During the initial in-laboratory ses-
sión, the experiment began with a brief hearing test in
which participants indicated when they perceived five
tones of 300–700 Hz (sin waveform, increments of 100 Hz),
which were presented at intervals that randomly varied
entre 3000 y 11,000 mseg (increments of 2000 mseg).
Each tone was played to each ear separately, in random
orden, and volume was adjusted if needed until the par-
ticipant was 100% correct in identifying the tones. El
computer volume was originally set to ∼56 dB, and all
participants were 100% accurate in the hearing test with-
out adjustment, resulting in the original intensity being
retained for the entire experiment in all cases. Entonces,
participants first completed the test phase task once
(to become familiar with the task procedure without in-
terfering with prior learning) and then the training phase
kim, Grégoire, and Anderson
181
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
oh
C
norte
_
a
_
0
1
7
8
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
2 times to establish learning of the stimulus–reward asso-
ciations (kim & anderson, 2020a, 2020b). During the fMRI
session, participants completed two runs of the training
phase, three runs of the test phase, an anatomical scan,
and an addition run of the training phase (to mitigate pos-
sible extinction effects) and two runs of the test phase.
Participants were compensated the total monetary reward
accumulated at the end of the last training phase or the
combined amount of $10/hr spent in the initial appoint- ment session and $20/hr spent in the fMRI session, cual-
ever amount was higher.
dimly lit room. Participants also wore Beyerdynamic DT
770 Pro 250Ω professional studio headphones to listen to
all sounds.
For the fMRI portion of the experiment, stimulus pre-
sentation was controlled by an Invivo SensaVue display
sistema. The eye-to-screen distance was approximately
125 cm. Key responses were entered using two Cedrus
Lumina two-button response pads. Output sounds were
sent to a Pyle PCA1 stereo power amplifier connected to
Sensimetrics Model S14 fMRI earphones (Sensimetrics
Corporation).
Auditory Stimuli
All auditory stimuli were recorded using a Spark SL con-
denser microphone (Baltic Latvian Universal Electronics
LLC.), with an Arrow audio interface (Universal Audio
Cª), en un 2017 MacBook Pro (Apple Inc.). The recordings
were sampled and modified using the built-in functions
on the Logic Pro X software (Apple Inc.). All recorded
samples of the numbers and letters were cut to begin
at the same time, compressed to make the sound inten-
sity equal, and condensed to be 300 msec in duration to
ensure acoustic similarities across all stimuli. En tono rimbombante,
all letter-to-value assignments were counterbalanced
across participants so that acoustical differences between
stimuli would not bias any of our statistical comparisons.
The auditory stimuli were the same as those used in Kim
et al.’s study (2021).
Apparatus
During the initial in-laboratory visit, all tasks were com-
pleted on a Dell OptiPlex 7040 computer equipped with
MATLAB software (The Mathworks, Cª) and Psychophysics
Toolbox extensions (Brainard, 1997). Stimuli were pre-
sented on a Dell P2717H monitor. The participants viewed
the monitor from a distance of approximately 70 cm in a
Experimental Design
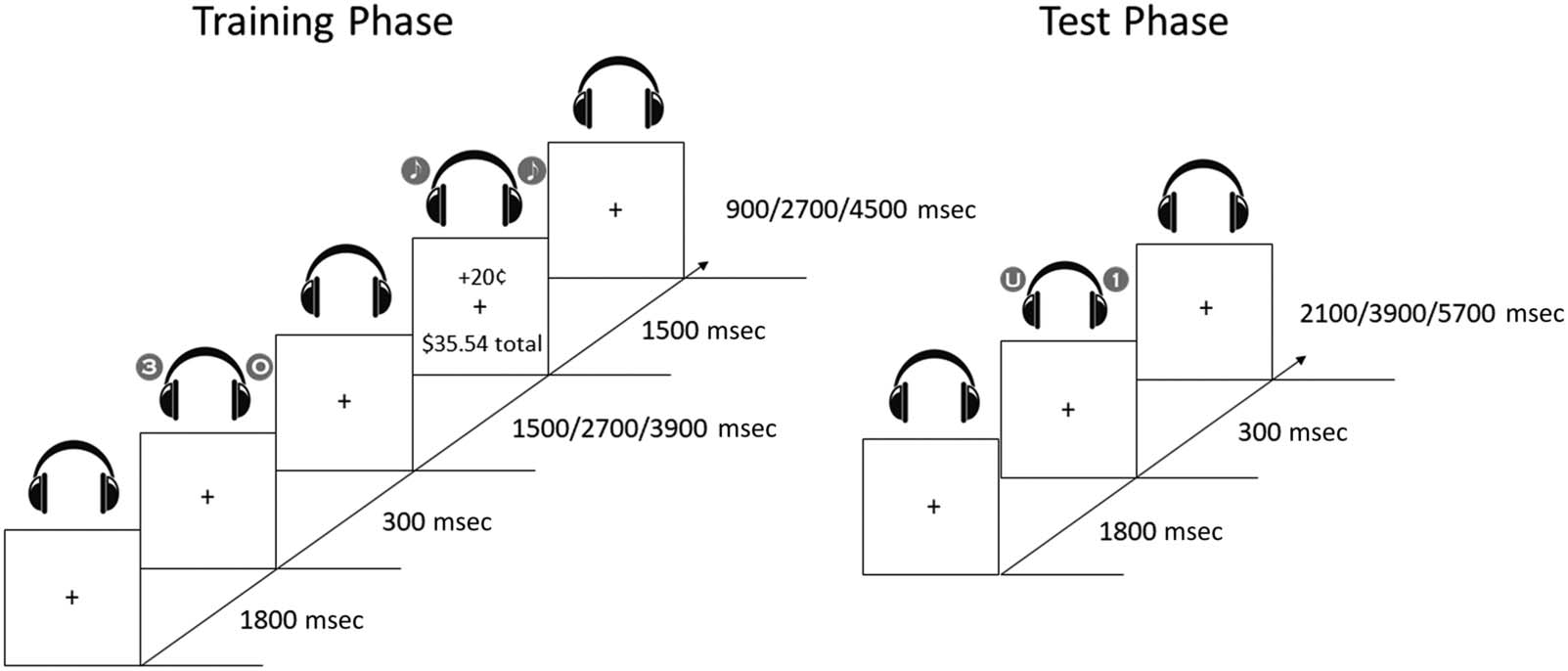
Training Phase
Each run of the training phase consisted of 72 ensayos. Cada
trial began with a fixation display (1800 mseg), seguido
by the target/distractor (300 mseg), an ISI, auditory/visual
comentario (1500 mseg), and an intertrial interval (ITI; ver
Cifra 1). Throughout each trial, a fixation cross (0.7° ×
0.7° visual angle) was presented at the center of the
pantalla. During the presentation of the target/distractor,
participants would simultaneously hear a spoken letter
played to one ear and a spoken number played to the
other ear. The possible letters were U, I, and O, y
the possible numbers were 1, 2, 3, y 4 (Participantes
were informed of these possibilities beforehand). Estos
letters and numbers were chosen based on their phonetics
(not rhyming and similar intonation) y su cerca
proximity on the keyboard. The possible letter–number
combinations and what side they were presented on the
earphones were fully counterbalanced, and the order of
trials was randomized in each run. Los participantes fueron
instructed to listen for the letter they heard and press
the respective key on the keyboard. They were told
that correct responses could result in monetary reward,
but no information was given about reward–letter
Cifra 1. Sequence of trial events in the training and test phases. In both phases, a spoken letter and a spoken number were played simultaneously,
one to each ear. In the training phase, participants responded to the letter they heard and were presented with monetary feedback. In the test phase,
participants responded to the number they heard while trying to ignore the same letters that had served as targets during training.
182
Revista de neurociencia cognitiva
Volumen 34, Número 1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
oh
C
norte
_
a
_
0
1
7
8
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
contingencies. We also specified to participants that they
would receive the total monetary reward attained through-
out the task or the base rate ($10/hr spent in the initial ap- pointment session and $20/hr spent in the fMRI session),
whichever was higher. Además, participants were in-
formed that if they did not complete the full experiment,
they would be paid at the base rate regardless of task earn-
ings. The ISI lasted for 1500, 2700, o 3900 mseg (equally
a menudo, order randomized). Próximo, participants were given
feedback based on what key they pressed. If the partici-
pant did not respond before the end of the ISI, they were
presented with the words “Too Slow” and their accumu-
lated total earnings, and if they pressed the wrong key,
they were presented with the words “Incorrect” and their
accumulated total earnings (no sound was presented
during such feedback). Para cada participante, each letter
was associated with high (20 cents), bajo (4 cents), or no
premio (0 cents). The letter-to-value mapping was coun-
terbalanced across participants. For correct responses,
participants were shown their corresponding reward earn-
ings and their accumulated total earnings, in addition to an
audible cue for 500 mseg (sine waveform, high reward =
650 Hz, low reward = 500 HZ, no reward = 350 Hz). El
visual feedback remained on the screen for the entire
duration of the feedback (1500 mseg) whereas the audible
cue was followed by silence for the remainder of the feed-
back period. We included the auditory feedback to help
ensure that participants robustly processed the feedback,
because it was possible to perform the task without actu-
ally looking at or otherwise processing the visual display.
Por último, the ITI lasted for 900, 2700, o 4500 mseg (expo-
nentially distributed, with the shorter time lengths being
more frequent). The fixation cross disappeared for the
last 200 msec of the ITI to indicate to the participant that
the next trial was about to begin. The auditory stimuli
and timing of trial events exactly matched the training
phase of Kim et al.’s study (2021).
Test Phase
Each run of the test phase consisted of 72 ensayos. Cada prueba
began with a fixation display (1800 mseg), followed by the
target/distractor (300 mseg), and an ITI (ver figura 1).
Throughout each trial, a fixation cross (0.7° × 0.7° visual
angle) was presented at the center of the screen. During
the presentation of the target/distractor, Participantes
would again simultaneously hear a letter and a number
(design identical to the training phase). Sin embargo, partícipe-
ipants were now instructed to listen for the number they
heard and press the respective number key on the key-
board, with the letters now serving as value-associated
but task-irrelevant distractors. Por último, the ITI lasted for
2100, 3900, o 5700 mseg (exponentially distributed, con
the shorter time lengths being more frequent). The fixa-
tion cross again disappeared for the last 200 msec of the
ITI to indicate to the participant that the next trial was
about to begin. The auditory stimuli and timing of trial
events exactly matched the test phase of Kim et al.’s study
(2021). The trial sequence (including order of trials, ISIs,
and ITIs) was fully randomized and newly created for
each run for each participant (for both training and test
phases) to ensure variability across participants.
MRI Data Acquisition
Images were acquired using a Siemens 3-Tesla MAGNETOM
Verio scanner with a 32-channel head coil. High-resolution
whole-brain anatomical images were acquired using a T1-
weighted magnetization prepared rapid gradient echo
pulse sequence (150 coronal slices, voxel size = 1 mm iso-
tropic, repetition time = 7.9 mseg, echo time = 3.65 mseg,
8° flip angle). Whole-brain functional images were
acquired using a T2*-weighted EPI multiband pulse se-
quence (56 axial slices, repetition time = 600 mseg, echo
time = 29 mseg, 52° flip angle, image matrix = 96 × 96,
campo de visión = 240 mm, slice thickness = 2.5 mm with
no gap). Each EPI pulse sequence began with dummy
pulses to allow the magnetic resonance signal to reach
steady state and concluded with an additional 6-sec blank
epoch.
MRI Data Analyses
Preprocesamiento
All preprocessing was conducted using the AFNI software
package (Cox, 1996). Each EPI run for every participant
was motion corrected using 3dvolreg, utilizing the first
image following the anatomical scan as a reference. El
anatomical image was skull-stripped using 3dskullstrip
and nonlinearly registered to the Talairach brain (Talairach
& Tournoux, 1988) using auto_warp.py. EPI images were
coregistered to the corresponding anatomical image for
each participant using align_epi_anat.py, and the EPI then
converted to percent signal change normalized to the
mean of each run. Por último, EPI images were nonlinearly
warped to the Talairach brain by applying the warp param-
eters from the anatomical image using 3dNwarpApply
and then spatially smoothed to a resulting 5-mm FWHM
using 3dBlurToFWHM.
Statistical Analyses
All statistical analyses were performed using the AFNI
software package (Cox, 1996). To determine whether
stimulus-evoked responses were elevated or suppressed
by value, we used a general linear model (GLM) acercarse
to analyze the training and test phase data. The GLM for
the training phase included the following regressors of
interés: (1) high-value target letter on left side, (2) bajo-
value target letter on left side, (3) no-value target letter
on left side, (4) high-value target letter on right side, (5)
low-value target letter on right side, (6) no-value target let-
ter on right side, (7) high-reward feedback, (8) low-reward
comentario, y (9) no-reward feedback. The GLM for the
kim, Grégoire, and Anderson
183
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
oh
C
norte
_
a
_
0
1
7
8
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
test phase included the following regressors of interest:
(1) high-value distractor on left side, (2) low-value distrac-
tor on left side, (3) no-value distractor on left side, (4)
high-value distractor on right side, (5) low-value distractor
on right side, y (6) no-value distractor on right side.
Each of these regressors was modeled using 16 finite
impulse response functions, beginning at the onset of the
respective stimulus (kim & anderson, 2020a, 2020b); eso
es, target/distractor regressors were time-locked to the
target/distractor and feedback regressors to the feedback
display/sound. Six degrees of head motion and drift in the
scanner signal were modeled using nuisance regressors.
The peak beta value for each condition from 3 a 6 sec post
stimulus presentation was extracted (p.ej., kim & anderson,
2020a, 2020b). Incorrect trials were not excluded from
analysis because there were too few from which to derive
a modeled response and participants were still exposed to
the same stimulus input, the processing of which was of
primary interest.
Given that no significant behavioral differences were
found between low- and no-value conditions in RT, rep-
licating prior results using this paradigm (Kim y cols.,
2021), we averaged the peak beta values for these two
condiciones (henceforth referred to as lesser-value) en
both the training and test phase data for ANOVAs, semejante
that the conditions used in these analyses correspond to
the behavioral effects of reward evident in this paradigm.
In the training and test phase, using the AFNI program
3dANOVA3, a three-way ANOVA was conducted compar-
ing the peak response on trials with Value-Associated
Target/Distractor (high vs. lesser) and Side (left vs. bien)
as fixed effects and Participant as a random effect. Multiple
comparison corrections were implemented using the
AFNI program 3dClustSim, with the smoothness of the
data estimated using the auto-correlation function method
via the AFNI program 3dFWHMx (clusterwise α < .05,
voxelwise p < .005). Of interest for both the training and
test phase was the ANOVA contrast on the effect of high-
value target/distractor versus lesser-value target/distractor.
Next, we more specifically probed for effects of reward
on the strength of stimulus-evoked activity in the test
phase within the frontoparietal attention network (FPN)
and the auditory cortex using an ROI approach. ROIs
were identified from the training phase ANOVA, which
provided independent data. The ROI for the FPN was
taken from the main contrast on the effect of high-value
target versus lesser-value targets and included four
clusters identified in the parietal cortex and the FEFs
given their overlap with regions previously identified in
the value-driven attention network (Anderson, 2019;
see Figure 3). The ROI for the auditory cortex was also
identified from the GLM of the training phase. In this
case, we calculated the average response for each of
the six target conditions in the GLM. We then averaged
over the six conditions and determined the ROI based
on the intersection of the functional activation and the
Talairach Atlas’ definition of auditory cortex (see Figure 4).
We used the AFNI program 3dmaskave to extract peak
beta values from each of the six conditions (Reward ×
Side) in the test phase and submitted this to the same
ANOVA as the voxelwise whole-brain analysis.
Lastly, to determine whether information about the
high-value distractor is more robustly represented than
information about lesser-value distractors in the auditory
cortex after they become task-irrelevant, we conducted
two MVPAs. For this purpose, the aforementioned GLMs
for the training and test phase were separated by run
and the peak beta value for each condition was calcu-
lated in the same way, providing one beta-weight esti-
mate per condition per run (Anderson, 2017). In our first
MVPA, we confirmed whether different target sounds
were reliably associated with different patterns of activa-
tion during the training phase. Beta-weights for high-,
low-, and no-value targets in the training phase for each
side were extracted from the contralateral ROI in the
mask created for the auditory cortex. These beta-weights
were then standardized (z scored), separately for each
run and participant, and subjected to MVPA using the lin-
ear support vector machine classifier (fitscvm) in
MATLAB. Linear support vector machine (SVM) was per-
formed using leave-one-run-out cross-validation, such
that the SVM was iteratively trained on the beta-weights
from n − 1 of the runs and tested on the left out run for
each participant resulting in three classification accura-
cies (as in Anderson, 2017; Xu et al., 2017) for each hemi-
sphere (high- vs. low-value, high- vs. no-value, low- vs.
no-value). These accuracies were averaged over the two
hemispheres to generate the mean classification accuracy
per participant, which were then averaged across partic-
ipants to compute a grand mean. The probability of the
observed grand mean classification accuracy under the
null hypothesis was determined using a randomization
procedure in which a distribution of mean classification
accuracy was computed under conditions in which the
training labels were randomly shuffled for each partici-
pant in 10,000 iterations (Anderson, 2017; Xu et al.,
2017); the order of trials in the random condition assign-
ment was matched with the original sequence, but the six
conditions (high-value target on the right, low-value tar-
get on the right, no-value target on the right, high-value
target on the left, low-value target on the left, and no-
value target on the left) were permuted for each run.
Next, we used a similar MVPA approach to investigate
whether the high-value distractor in the test phase pro-
duced a more robust pattern of activation in the auditory
cortex compared to lesser-value stimuli. To the degree
that information about the distractor is represented in
spite of its task-irrelevance, the pattern of activation that
it evokes should be more similar to the pattern evoked
by the same stimulus during the training phase when it
is actually attended. In addition to the acquired beta-
weights for the high-, low-, and no-value targets in the
training phase, beta-weights for the high-, low-, and no-
value distractors in the test phase were similarly
184
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
o
c
n
_
a
_
0
1
7
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
extracted. We utilized a similar MVPA and randomization
test approach as for the training phase. However, in this
case, linear SVM was performed by training the classifier
on the beta-weights for the four training-phase runs and
predicting on the four test-phase runs. In this case,
higher classification accuracy means that a distractor
sound was processed more similarly to when it was
task-relevant and attended. As with our ANOVAs, to
acquire classification accuracies with respect to lesser-
value distractors in order to match our behavior results
but in this case avoid biases in classification accuracy
because of averaging of conditional beta values, we
classified high- versus low-value and high- versus no-value
distractors separately and subsequently averaged the results
of the two for each participant to obtain an overall
estimate of high- versus lesser-value discriminability.
Classification accuracies between conditions were directly
compared using the same randomization approach. A
randomization procedure using random sign flipping on
the resulting classification accuracies (either above/below
0.5 or the difference score when comparing two classifica-
tion accuracies) was also performed on all MVPA results,
which achieved the same conclusions with respect to
statistical significance and is therefore not reported.
Data Analysis and Exclusion Criteria
Behavior results are presented from only the fMRI ses-
sion and not the in-laboratory session the day before
such that the behavioral and fMRI data correspond to
the same trials.1 RT was measured from the onset of
the target/distractor. Only correct trials were included
in the RT analyses. RTs more than 2.5 SDs above and be-
low the mean for a given condition for a given participant
were trimmed (Kim et al., 2021; Kim & Anderson, 2020a,
2020b). In addition, we excluded two participants’ data
whose mean accuracy or RT exceeded 2.5 SDs below or
above the group mean as outliers (see the work of
Anderson, 2016b). Furthermore, two participants made
significant head movements during their neuroimaging
scan and 25.8% and 6.4% of images, respectively, would
have needed to be censored because of excessive motion
exceeding half the width of a voxel during a single dy-
namic (in comparison to an average of 0.2% of images
among all other participants); these participants were
also excluded from the final analysis. In the end, 31 com-
plete behavior and neuroimaging data sets were submit-
ted to final analyses.
Data and Code Availability Statement
The full data set, including the raw MRI data, are available
upon reasonable request made to the corresponding au-
thor and will be provided under the provision that the
data be used strictly for academic research purposes
and not be shared with others without the expressed
written approval of the corresponding author. Data shar-
ing for this article complies with the requirements of the
funding agencies and the stipulations of the university
institutional review board approvals.
RESULTS
Behavioral
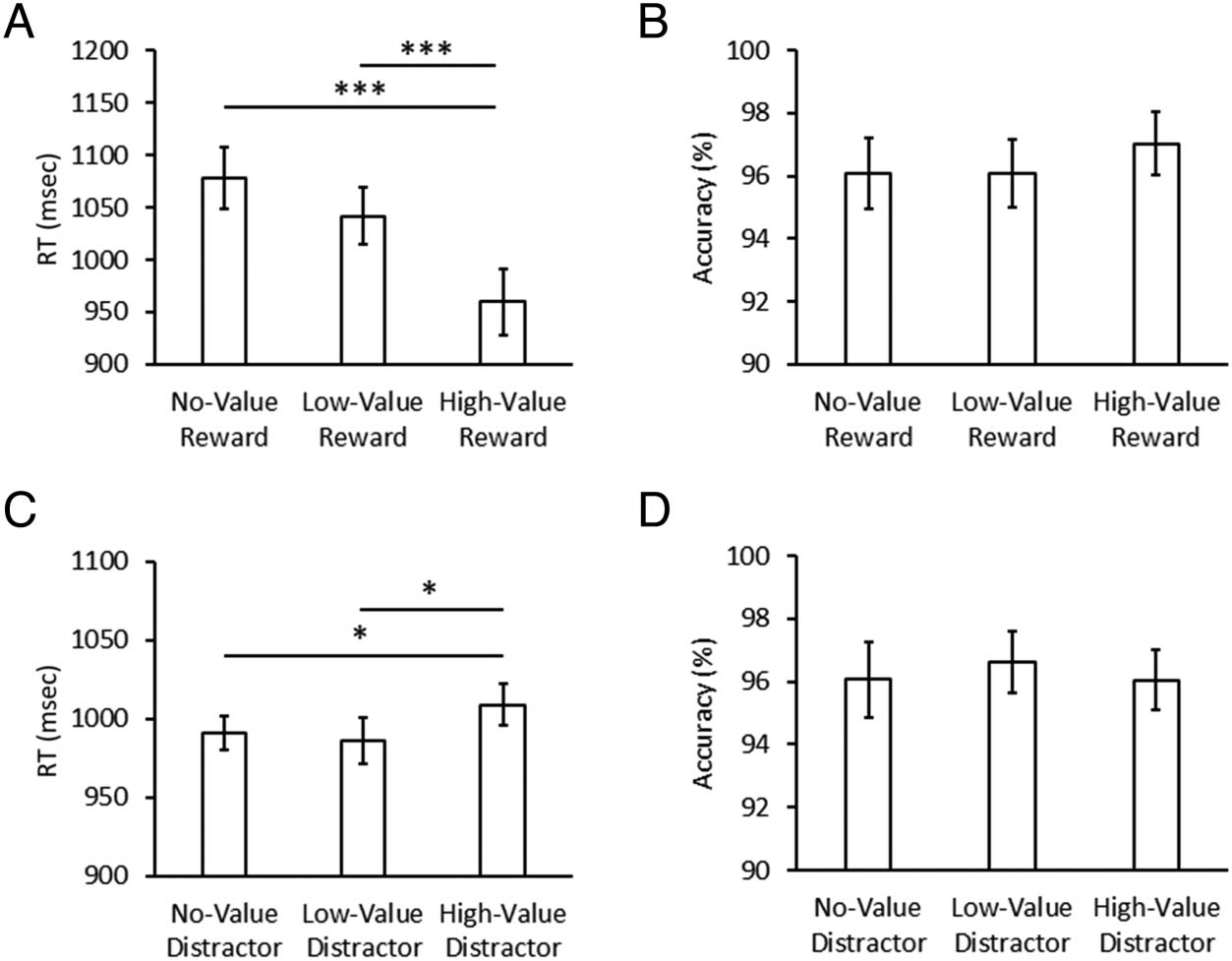
In the training phase, a repeated-measures ANOVA re-
vealed that RTs significantly differed among the three tar-
get conditions, F(2, 60) = 17.73, p < .001, η2
p = .371.
Post hoc comparisons revealed that participants were
Figure 2. Behavior results. RT
(A) and accuracy (B) in the
training phase and RT (C) and
accuracy (D) in the test phase.
Data are broken down by trials
based on target-reward
contingencies in the training
phase and by learned reward–
distractor associations in the
test phase. Error bars depict
within-subject confidence
intervals calculated using the
Cousineau method (Cousineau,
2005) with a Morey correction
(Morey, 2008). *p < .05,
***p < .001.
Kim, Grégoire, and Anderson
185
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
o
c
n
_
a
_
0
1
7
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
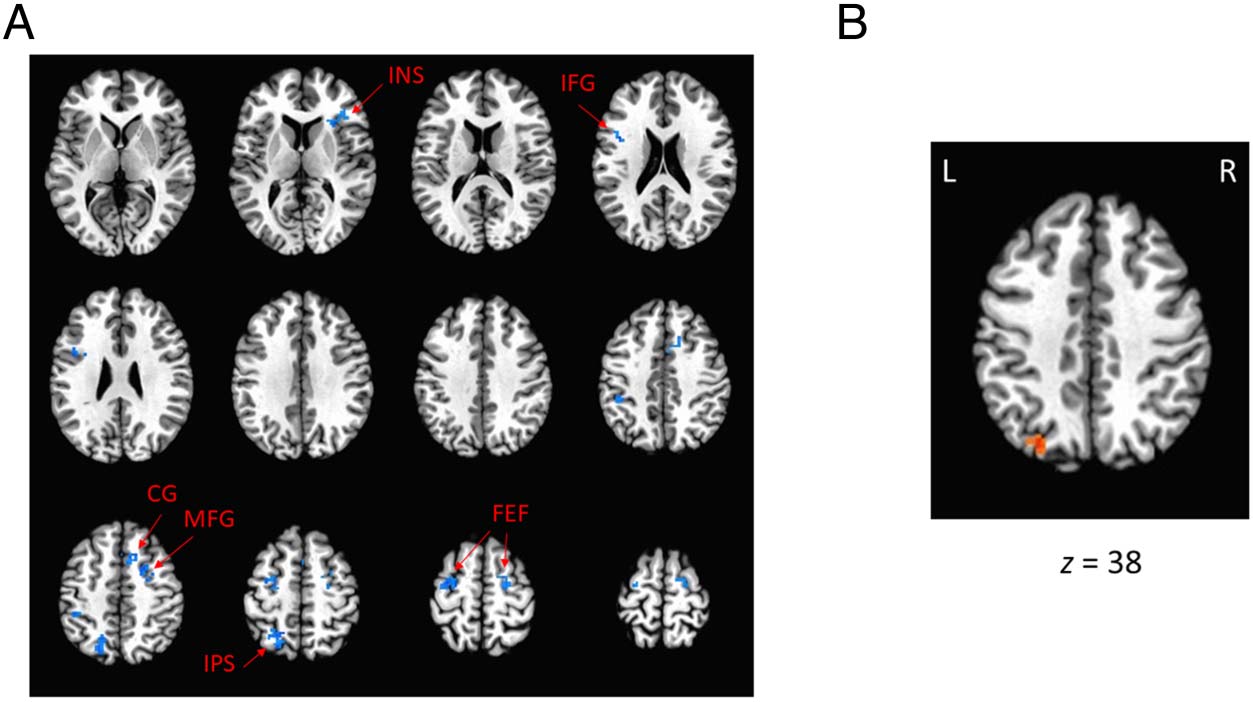
Figure 3. ANOVA contrast on
the effect of high value versus
lesser value (low- and no-value;
clusterwise: a < .05, voxelwise:
p < .005). (A) Montage of
significant clusters in the
training phase indicating
reduced activation in response
to high-value targets. (B)
Significant cluster in the test
phase indicating increased
activation in the posterior
parietal cortex in response
to high-value distractors. All
significant clusters are overlaid
on an image of the Talairach
brain.
significantly faster to report high-value targets compared
to both unrewarded targets, t(30) = 5.42, p < .001, dz =
0.973, and low-value targets, t(30) = 4.02, p < .001, dz =
0.720, but only a marginally significant difference was
found comparing low-value and unrewarded targets,
t(30) = 1.96, p = .060 (see Figure 2A). Accuracy did not
significantly differ among the three target conditions,
F(2, 60) = 1.11, p = 0.338 (see Figure 2B).
In the test phase, a repeated-measures ANOVA revealed
that RTs differed significantly among the three distractor
conditions, F(2, 60) = 3.69, p = .031, η2
p = .110. Post hoc
comparisons revealed that RTs were significantly slower
on high-value distractor trials compared to both no-value
distractor trials, t(30) = 2.40, p = .023, dz = 0.432, and
low-value distractor trials, t(30) = 2.24, p = .032, dz =
0.404, but no significant differences were found compar-
ing low-value and no-value distractor trials, t(30) = 0.58,
p = .564 (see Figure 2C). Accuracy did not significantly
differ among the three distractor conditions, F(2, 60) =
0.40, p = 670 (see Figure 2D).
Neuroimaging
GLM
From our ANOVA contrast on the effect of high-value ver-
sus lesser-value (low- and no-value) in the training phase,
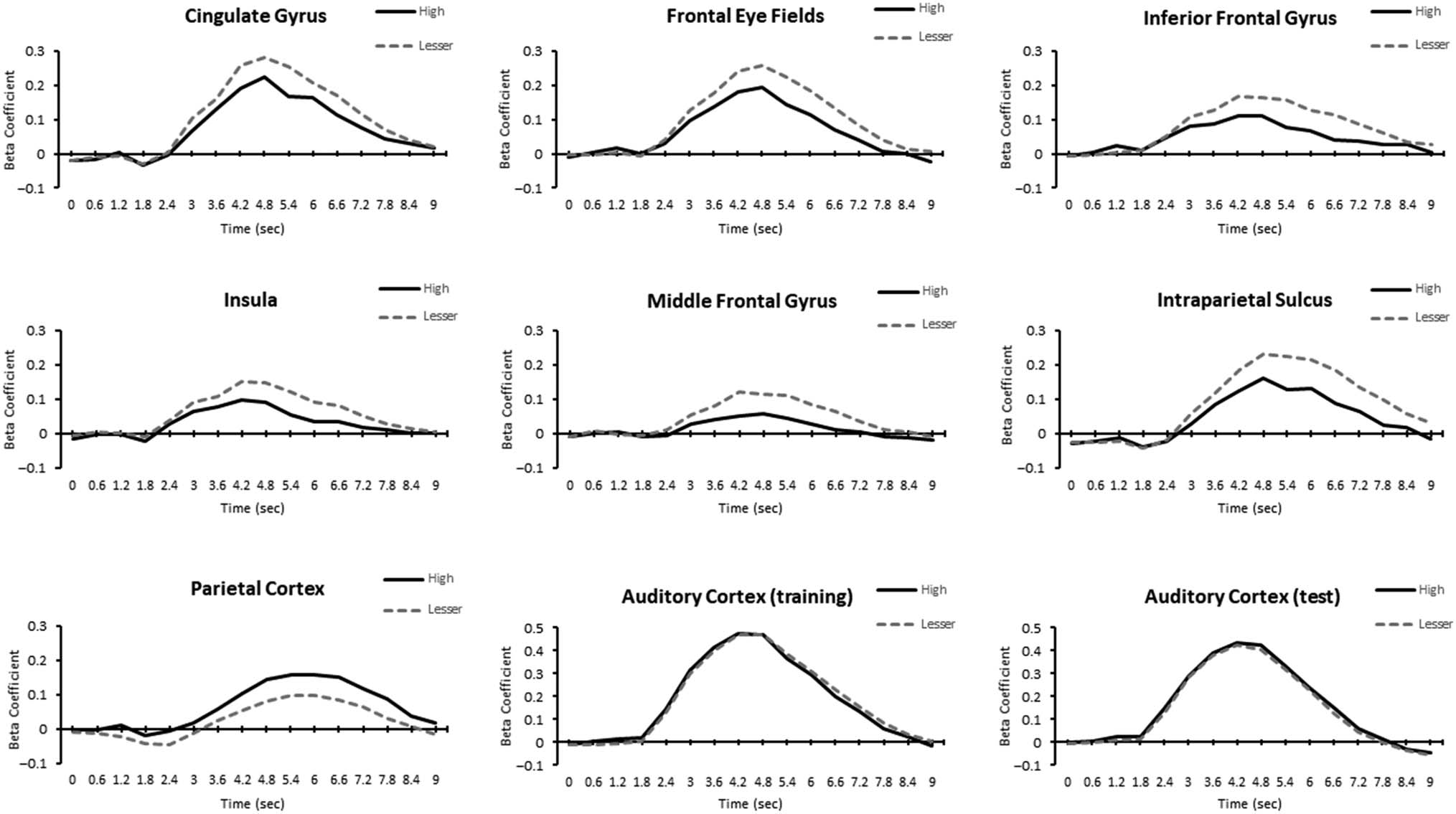
Figure 4. Time course of BOLD responses in each region in the training phase (CG, FEF, IFG, INS, MFG, IPS) and the test phase (parietal cortex) for
which a significant difference between the high-value and lesser-value stimuli emerged, in addition to the auditory cortex.
186
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
o
c
n
_
a
_
0
1
7
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Regions of the Brain Demonstrating Decreased Activation by High-Value Targets during the Training Phase and Increased
Activation to High-Value Distractors during the Test Phase
Region
Training Phase
Right CG
Right FEF
Right anterior INS
Right MFG
Left IPS
Left FEF
Left inferior frontal gyrus
Left parietal cortex
Test Phase
x
−13.8
−21.2
−28.8
−18.8
16.2
31.2
38.8
41.2
y
−16.2
8.8
−18.8
−1.2
61.2
8.8
−6.2
38.8
z
46.2
56.2
11.2
43.8
48.8
56.2
21.2
43.8
Volume (ml)
0.688
0.688
0.406
0.359
1.047
0.875
0.469
0.328
Left posterior parietal cortex
26.2
73.8
36.2
0.422
x, y, and z refer to the Talairach coordinates of the peak voxel of the cluster.
we identified significantly less neural activity by the high-
value target in the insula (INS), inferior frontal gyrus (IFG),
middle frontal gyrus (MFG), cingulate gyrus (CG), intra-
parietal sulcus (IPS), and FEFs (see Figure 3A, Figure 4,
and Table 1). From our ANOVA contrast on the effect of
high-value distractor versus lesser-value distractors (low-
and no-value) in the test phase, we identified elevated
neural activity by the high-value distractor in the posterior
parietal cortex (see Figure 3B, Figure 4, and Table 1). To
specifically probe for an effect of reward on the magnitude
of stimulus-evoked activity within the FPN and auditory
cortex during the test phase that might have been too sub-
tle to detect at the whole-brain level, we first created
masks for these regions of interest (see Methods section;
see also Figure 5). For the FPN, a 2 × 2 ANOVA over factors
Reward (high- vs. lesser-value) and Side (left vs. right)
found no main effects or interaction: main effect of Side,
F(1, 30) = 3.08, p = .089, other Fs < 1.03, ps >.31. Para el
auditory cortex, lo mismo 2 × 2 ANOVA similarly found no
main effects or interaction, Fs < 1.93, ps > .175.
MVPA
We first established that the differently valued stimuli in
the training phase produced reliably different patterns of
activation in the auditory cortex. Our MVPA significantly
classified each pair of conditions (alto- vs. low-value, ac-
curacy = 55%, alto- vs. no-value, accuracy = 56%, bajo-
vs. no-value, accuracy = 57%, ps < .014). Then, we
trained classifiers on these patterns, when the stimuli
were task-relevant and attended, to determine whether
and to what degree this pattern was maintained when
the same stimuli were presented as task-irrelevant dis-
tractors. Classification was significantly above chance in
distinguishing high-value distractors from lesser-value
distractors (accuracy = 53%, p = .003), but not for distin-
guishing between low-value and no-value distractors
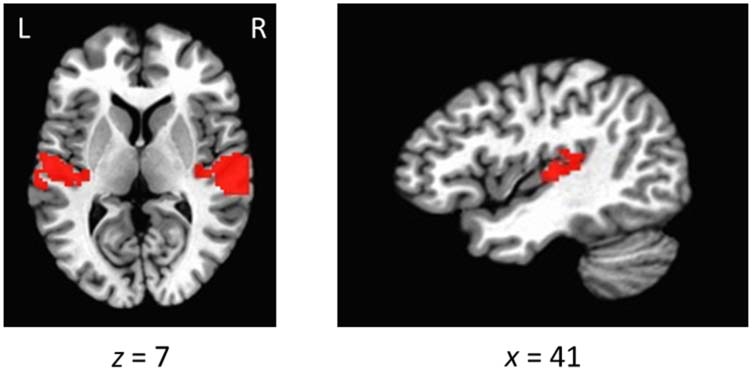
Figure 5. Mask of the auditory cortex for quantification and MVPAs.
Voxels are overlaid on an image of the Talairach brain.
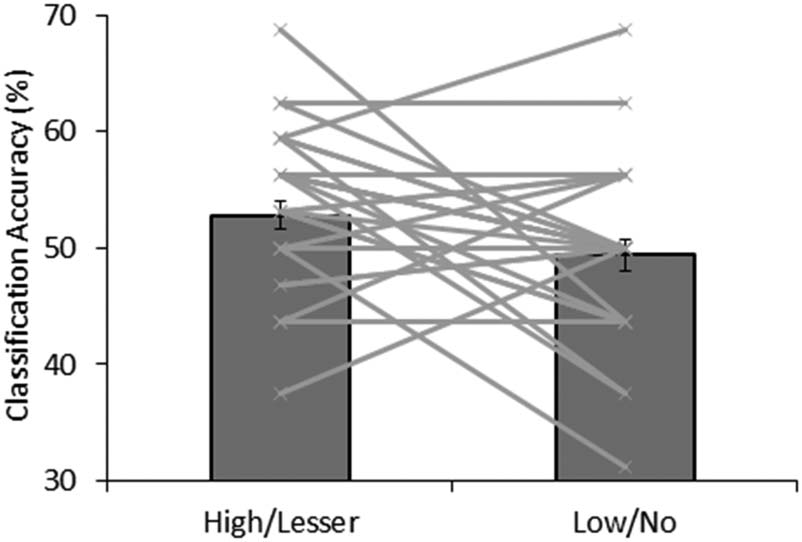
Figure 6. Classification accuracies from MVPA distinguishing high-value
from lesser-value distractors and low-value from no-value distractors.
Each line reflects a single participant connecting the classification
accuracies for that participant across the two conditions.
Kim, Grégoire, and Anderson
187
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
o
c
n
_
a
_
0
1
7
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
(accuracy = 49%, p = .479). The difference between
these two accuracies was also significant, p = .031 (see
Figure 6).
DISCUSSION
RTs were significantly faster in the training phase when
the target was associated with high value, demonstrating
a voluntary attentional bias driven by the motivational ef-
fects of reward. In addition, RTs were significantly slower
in the test phase when the same stimulus was presented
as a task-irrelevant distractor, demonstrating an involun-
tary attentional bias driven by learned value or VDAC.
Thus, we replicated behavioral evidence of two forms
of value-based attentional biases in the auditory domain
(Kim et al., 2021). Our fMRI data permitted an assess-
ment of the neural correlates of each of these two influ-
ences on the control of attention, with implications for
our understanding of mechanisms of value-driven atten-
tion in the auditory domain and the extent to which vol-
untary and involuntary modes of value-based orienting
are distinct.
In the test phase, we found elevated stimulus-evoked
activity in the posterior parietal cortex by the high-value
distractor, consistent with studies of VDAC using visual
stimuli (e.g., Kim & Anderson, 2020b; Anderson, 2017;
Hickey & Peelen, 2015; Anderson et al., 2014; Qi, Zeng,
Ding, & Li, 2013). The posterior parietal cortex plays a
broad role in attentional selection and is a core region
of the dorsal frontoparietal network, which is thought
to selectively bias the representation of task-relevant or
otherwise pertinent stimuli (see works of Corbetta,
Patel, & Shulman, 2008; Behrmann, Geng, & Shomstein,
2004; Corbetta & Shulman, 2002, for reviews). In con-
trast, we did not find evidence for priority signals in the
caudate tail, which plays a central role in value-driven at-
tention in the visual domain (e.g., Kim & Anderson,
2020a, 2020b; Anderson, 2017, 2019; Anderson et al.,
2014, 2016; Kim & Hikosaka, 2013; Yamamoto, Kim, &
Hikosaka, 2013; Yamamoto, Monosov, Yasuda, &
Hikosaka, 2012; Orban, Van Essen, & Vanduffel, 2004),
or any other region of the brain implicated in VDAC by vi-
sual stimuli, arguing against a supramodal network sup-
porting value-based attention.
In the training phase, in contrast, we found reduced
priority signals by the high-value target in the FEFs, IPS,
MFG, IFG, CG, and INS. That is, the representation of
high-value sounds demonstrated a relative suppression
of neural activity within the dorsal FPN typically implicated
in the processing of visual information (Corbetta et al.,
2008; Corbetta & Shulman, 2002), without any apparent
increases in the auditory system. Such a finding echoes
an earlier behavioral demonstration of increased interfer-
ence from previously reward-associated sounds in a visual
task (Anderson, 2016b), suggesting that behavioral
impairments in such cross-modal designs may at least
in part reflect reduced priority to sensory input outside
of the auditory system. Targeted ROI analyses of the test
phase data provide no evidence for the maintenance of
this pattern of reduced priority signals into the test
phase. It is also possible that, in at least some regions
identified, particularly those that have been linked to
affective information processing such as the INS (e.g.,
Berntson et al., 2011; Winston, Gottfried, Kilner, &
Dolan, 2005; Norris, Chen, Zhu, Small, & Cacioppo,
2004), the observed difference in activation is the result
of elevated processing of lesser-value stimuli, possibly
because of these stimuli being represented as compara-
tively aversive. The relative nature of the BOLD response
is necessarily ambiguous with respect to the distinction
between selective enhancement versus suppression of
an evoked response.
Collectively, these contrasting results indicate diver-
gent mechanisms for modulating attentional priority de-
pending on the relationship between value and task
goals. Whereas both the high-value target in the training
phase and high-value distractor in the test phase received
elevated attentional priority, neural activity was generally
suppressed in relative terms when processing task-
relevant reward cues and elevated when processing
task-irrelevant, previously reward-associated distractors.
Our results offer neural evidence against the idea that
VDAC merely reflects a persistence of motivated atten-
tional processes, which would have predicted a similar
pattern of stimulus-evoked activity across phases. Such
a finding is broadly consistent with behavioral evidence
for an independent role for (implicit) associative learning
and target history effects in the control of attention (e.g.,
Grégoire, Kim, & Anderson, 2021; Kim & Anderson,
2019a, 2021; Anderson & Britton, 2019; Anderson,
Chiu, DiBartolo, & Leal, 2017; see also the work of Kim
& Anderson, 2019b) as well as inhibitory accounts of the
selective processing of a target (e.g., Gaspelin & Luck,
2018a, 2018b, 2018c, 2019; Gaspelin, Leonard, & Luck,
2015, 2017; Moher, Lakshmanan, Egeth, & Ewen, 2014).
In the visual domain, stimulus-specific information per-
taining to previously reward-associated stimuli has been
demonstrated in early sensory cortices (Itthipuripat et al.,
2019; Serences & Saproo, 2010). In this study, we provide
a parallel demonstration of this phenomenon in the
auditory system, suggesting that early sensory enhance-
ment of reward-associated signals reflects a modality-
general process at play across multiple sensory systems.
When task-irrelevant, only information pertaining to the
identity of previously high-value stimuli could be de-
coded from the other distractor conditions, suggesting
that stimulus-specific information about high-value
sounds was maintained across phases of the experiment
whereas this was not the case for lesser-value sounds,
which were more effectively ignored. Future research
could explore the nature of this stimulus-specific infor-
mation enhancement in more detail, isolating feature-
specific components (e.g., frequency, as with color in
the visual system, or location) and complex identity
188
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
o
c
n
_
a
_
0
1
7
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
information (with parallel to visual objects) to isolate dif-
ferent stages of information processing.
In probing the neural mechanisms of reward’s influ-
ence on attention in the auditory domain, several
broader principles emerge. Value-driven auditory atten-
tion, like value-driven attention in the visual domain, is
reflected in biased competition within sensory systems,
consistent with an early-stage influence. The posterior
parietal cortex seems to play a role in value-driven atten-
tion that spans sensory modalities, but beyond this, our
results do not suggest a widespread supramodal network
of value-based attentional prioritization, as could be sug-
gested from common behavioral influences of reward
across vision and audition and the neural correlates of
goal-directed and stimulus-driven orienting across modal-
ities throughout the dorsal and ventral attention net-
works (see works of Macaluso, 2010; Macaluso &
Driver, 2005, for reviews). Our findings also highlight a
notable distinction between reward’s influence on moti-
vated attention and involuntary attentional capture in the
case of audition, with motivated attention reflecting the
selective suppression of information in other sensory
systems and learning-dependent prioritization reflected
in priority signals in the parietal cortex along with
stimulus-specific sensory enhancement. In this respect,
our findings provide neural evidence for a distinction be-
tween value-driven attentional processes and the persev-
eration of motivated attention, which has been a topic of
controversy in the attention literature (e.g., Kim &
Anderson, 2019a; Anderson, 2016a).
Acknowledgments
This study was supported by grants from the National Institutes
of Health (R01-DA046410) to B. A. A. We thank David S. Lee for
assistance in creating the auditory stimuli.
The corresponding author has transitioned to a new position.
Reprint requests should be sent to Andy J. Kim, Department of
Gerontology, University of Southern California, 3715 McClintock
Ave. Los Angeles, CA 90089, or or via e-mail: andyk@usc.edu.
Author Contributions
Andy J. Kim: Conceptualization; Data curation; Formal
analysis; Writing—Original draft. Laurent Grégoire: Data
curation; Writing—Review & editing. Brian A. Anderson:
Conceptualization; Formal analysis; Writing—Review &
editing.
Funding Information
Brian A. Anderson, National Institute on Drug Abuse
(https://dx.doi.org/10.13039/100000026), grant number:
R01-DA046410.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/ W = .108, and W/ W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/ W = .102, and W/ W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
Note
1. The main effect of target value was also significant for RT in
the in-laboratory portion, demonstrating a significant effect of
learning.
REFERENCES
Anderson, B. A. (2016a). The attention habit: How reward
learning shapes attentional selection. Annals of the New York
Academy of Sciences, 1369, 24–39. https://doi.org/10.1111
/nyas.12957, PubMed: 26595376
Anderson, B. A. (2016b). Value-driven attentional capture in the
auditory domain. Attention, Perception, & Psychophysics, 78,
242–250. https://doi.org/10.3758/s13414-015-1001-7, PubMed:
26494380
Anderson, B. A. (2017). Reward processing in the value-driven
attention network: Reward signals tracking cue identity and
location. Social Cognitive and Affective Neuroscience, 12,
461–467. https://doi.org/10.1093/scan/nsw141, PubMed:
27677944
Anderson, B. A. (2019). Neurobiology of value-driven attention.
Current Opinion in Psychology, 29, 27–33. https://doi.org/10
.1016/j.copsyc.2018.11.004, PubMed: 30472540
Anderson, B. A., & Britton, M. K. (2019). Selection history in
context: Evidence for the role of reinforcement learning in
biasing attention. Attention, Perception, and Psychophysics,
81, 2666–2672. https://doi.org/10.3758/s13414-019-01817-1,
PubMed: 31309530
Anderson, B. A., Chiu, M., DiBartolo, M. M., & Leal, S. L. (2017).
On the distinction between value-driven attention and
selection history: Evidence from individuals with depressive
symptoms. Psychonomic Bulletin and Review, 24, 1636–1642.
https://doi.org/10.3758/s13423-017-1240-9, PubMed: 28210998
Anderson, B. A., & Halpern, M. (2017). On the value-dependence
of value-driven attentional capture. Attention, Perception, &
Psychophysics, 79, 1001–1011. https://doi.org/10.3758/s13414
-017-1289-6, PubMed: 28176215
Anderson, B. A., Kuwabara, H., Wong, D. F., Gean, E. G.,
Rahmim, A., Brasic, J. R., et al. (2016). The role of dopamine
in value-based attentional orienting. Current Biology, 26,
550–555. https://doi.org/10.1016/j.cub.2015.12.062, PubMed:
26877079
Anderson, B. A., Laurent, P. A., & Yantis, S. (2011). Value-driven
attentional capture. Proceedings of the National Academy of
Sciences, U.S.A., 108, 10367–10371. https://doi.org/10.1073
/pnas.1104047108, PubMed: 21646524
Anderson, B. A., Laurent, P. A., & Yantis, S. (2014). Value-driven
attentional priority signals in human basal ganglia and visual
cortex. Brain Research, 1587, 88–96. https://doi.org/10.1016/j
.brainres.2014.08.062, PubMed: 25171805
Kim, Grégoire, and Anderson
189
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
o
c
n
_
a
_
0
1
7
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Asutay, E., & Västfjäll, D. (2016). Auditory attentional selection
is biased by reward cues. Scientific Reports, 6, 36989. https://
doi.org/10.1038/srep36989, PubMed: 27841363
Awh, E., Belopolsky, A. V., & Theeuwes, J. (2012). Top–down
versus bottom–up attentional control: A failed theoretical
dichotomy. Trends in Cognitive Sciences, 16, 437–443.
https://doi.org/10.1016/j.tics.2012.06.010, PubMed: 22795563
of auditory processing. Biological Psychology, 114, 23–32.
https://doi.org/10.1016/j.biopsycho.2015.12.001, PubMed:
26678665
Folyi, T., & Wentura, D. (2019). Involuntary sensory enhancement
of gain-and loss-associated tones: A general relevance principle.
International Journal of Psychophysiology, 138, 11–26. https://
doi.org/10.1016/j.ijpsycho.2019.01.007, PubMed: 30685230
Barbaro, L., Peelen, M. V., & Hickey, C. (2017). Valence, not
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2015). Direct
utility, underlies reward-driven prioritization in human vision.
Journal of Neuroscience, 37, 10438–10450. https://doi.org/10
.1523/JNEUROSCI.1128-17.2017, PubMed: 28951452
evidence for active suppression of salient-but-irrelevant
sensory inputs. Psychological Science, 26, 1740–1750. https://
doi.org/10.1177/0956797615597913, PubMed: 26420441
Behrmann, M., Geng, J. J., & Shomstein, S. (2004). Parietal
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2017). Suppression of
cortex and attention. Current Opinion in Neurobiology, 14,
212–217. https://doi.org/10.1016/j.conb.2004.03.012, PubMed:
15082327
Berntson, G. G., Norman, G. J., Bechara, A., Bruss, J., Tranel, D.,
& Cacioppo, J. T. (2011). The insula and evaluative processes.
Psychological Science, 22, 80–86. https://doi.org/10.1177
/0956797610391097, PubMed: 21148459
Brainard, D. H. (1997). The Psychophysics toolbox. Spatial
Vision, 10, 433–436. https://doi.org/10.1163/156856897X00357,
PubMed: 9176952
Brosch, M., Selezneva, E., & Scheich, H. (2011). Representation
of reward feedback in primate auditory cortex. Frontiers in
Systems Neuroscience, 5, 5. https://doi.org/10.3389/fnsys
.2011.00005, PubMed: 21369350
Cheng, F. P.-H., Saglam, A., Andre, S., & Pooresmaeili, A. (2020).
Cross-modal integration of reward value during oculomotor
planning. eNeuro, 7. https://doi.org/10.1523/ENEURO.0381
-19.2020, PubMed: 31996392
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The reorienting
system of the human brain: From environment to theory of
mind. Neuron, 58, 306–324. https://doi.org/10.1016/j.neuron
.2008.04.017, PubMed: 18466742
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neuroscience, 3, 201–215. https://doi.org/10.1038/nrn755,
PubMed: 11994752
Cousineau, D. (2005). Confidence intervals in within-subject
designs: A simpler solution to Loftus and Masson’s method.
Tutorials in Quantitative Methods for Psychology, 1, 42–45.
https://doi.org/10.20982/tqmp.01.1.p042
Cox, R. W. (1996). AFNI: software for analysis and visualization
of functional magnetic resonance neuroimages. Computers
and Biomedical Research: An International Journal, 29,
162–173. https://doi.org/10.1006/cbmr.1996.0014, PubMed:
8812068
David, S. V., Fritz, J. B., & Shamma, S. A. (2012). Task reward
structure shapes rapid receptive field plasticity in auditory
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 109, 2144–2149. https://doi.org/10.1073/pnas
.1117717109, PubMed: 22308415
Della Libera, C., & Chelazzi, L. (2006). Visual selective attention
and the effects of monetary rewards. Psychological Science,
17, 222–227. https://doi.org/10.1111/j.1467-9280.2006.01689.x,
PubMed: 16507062
Ferreri, L., Mas-Herrero, E., Zatorre, R. J., Ripollés, P., Gomez-
Andres, A., Alicart, H., et al. (2019). Dopamine modulates the
reward experiences elicited by music. Proceedings of the
National Academy of Sciences, U.S.A., 116, 3793–3798.
https://doi.org/10.1073/pnas.1811878116, PubMed: 30670642
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992).
Involuntary covert orienting is contingent on attentional
control settings. Journal of Experimental Psychology: Human
Perception and Performance, 18, 1030–1044. https://doi.org
/10.1037/0096-1523.18.4.1030, PubMed: 1431742
Folyi, T., Liesefeld, H. R., & Wentura, D. (2016). Attentional
enhancement for positive and negative tones at an early stage
overt attentional capture by salient-but-irrelevant color
singletons. Attention, Perception, & Psychophysics, 79,
45–62. https://doi.org/10.3758/s13414-016-1209-1, PubMed:
27804032
Gaspelin, N., & Luck, S. J. (2018a). Combined electrophysiological
and behavioral evidence for the suppression of salient
distractors. Journal of Cognitive Neuroscience, 30, 1265–1280.
https://doi.org/10.1162/jocn_a_01279, PubMed: 29762104
Gaspelin, N., & Luck, S. J. (2018b). Distinguishing among
potential mechanisms of singleton suppression. Journal
of Experimental Psychology: Human Perception and
Performance, 44, 626. https://doi.org/10.1037/xhp0000484,
PubMed: 29035072
Gaspelin, N., & Luck, S. J. (2018c). The role of inhibition in
avoiding distraction by salient stimuli. Trends in Cognitive
Sciences, 22, 79–92. https://doi.org/10.1016/j.tics.2017.11.001,
PubMed: 29191511
Gaspelin, N., & Luck, S. J. (2019). Inhibition as a potential
resolution to the attentional capture debate. Current
Opinion in Psychology, 29, 12. https://doi.org/10.1016/j
.copsyc.2018.10.013, PubMed: 30415087
Gold, B. P., Pearce, M. T., Mas-Herrero, E., Dagher, A., &
Zatorre, R. J. (2019). Predictability and uncertainty in the
pleasure of music: A reward for learning? Journal of
Neuroscience, 39, 9397–9409. https://doi.org/10.1523
/JNEUROSCI.0428-19.2019, PubMed: 31636112
Gre(cid:1)goire, L., Kim, H., & Anderson, B. A. (2021). Punishment-
modulated attentional capture is context-specific. Motivation
Science, 7, 165–175. https://doi.org/10.1037/mot0000211
Hickey, C., Chelazzi, L., & Theeuwes, J. (2010). Reward guides
vision when it’s your thing: Trait reward-seeking in reward-
mediated visual priming. PLoS One, 5, e14087. https://doi.org
/10.1371/journal.pone.0014087, PubMed: 21124893
Hickey, C., & Peelen, M. V. (2015). Neural mechanisms of
incentive salience in naturalistic human vision. Neuron,
85, 512–518. https://doi.org/10.1016/j.neuron.2014.12.049,
PubMed: 25654257
Irvine, D. R. F. (2018). Plasticity in the auditory system. Hearing
Research, 362, 61–73. https://doi.org/10.1016/j.heares.2017
.10.011, PubMed: 29126650
Itthipuripat, S., Vo, V. A., Sprague, T. C., & Serences, J. T.
(2019). Value-driven attentional capture enhances distractor
representations in early visual cortex. PLoS Biology, 17,
e3000186. https://doi.org/10.1371/journal.pbio.3000186,
PubMed: 31398186
Kim, H., & Anderson, B. A. (2019a). Dissociable components of
experience-driven attention. Current Biology, 29, 841–845.
https://doi.org/10.1016/j.cub.2019.01.030, PubMed: 30773366
Kim, H., & Anderson, B. A. (2019b). Dissociable neural
mechanisms underlie value-driven and selection-driven
attentional capture. Brain Research, 1708, 109–115. https://
doi.org/10.1016/j.brainres.2018.11.026, PubMed: 30468726
Kim, A. J., & Anderson, B. A. (2020a). Arousal-biased
competition explains reduced distraction by reward cues
under threat. eNeuro, 7. https://doi.org/10.1523/ENEURO
.0099-20.2020, PubMed: 32601095
190
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
o
c
n
_
a
_
0
1
7
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Kim, A. J., & Anderson, B. A. (2020b). Neural correlates of
attentional capture by stimuli previously associated with
social reward. Cognitive Neuroscience, 11, 5–15. https://doi
.org/10.1080/17588928.2019.1585338, PubMed: 30784353
Kim, H., & Anderson, B. A. (2021). How does the attention
system learn from aversive outcomes? Emotion, 21, 898–903.
https://doi.org/10.1037/emo0000757, PubMed: 32718173
Kim, H. F., & Hikosaka, O. (2013). Distinct basal ganglia circuits
controlling behaviors guided by flexible and stable values.
Neuron, 79, 1001–1010. https://doi.org/10.1016/j.neuron
.2013.06.044, PubMed: 23954031
Kim, A. J., Lee, D. S., & Anderson, B. A. (2021). Previously
reward-associated sounds interfere with goal-directed
auditory processing. Quarterly Journal of Experimental
Psychology, 74, 1257–1263. https://doi.org/10.1177
/1747021821990033, PubMed: 33438522
Kraus, N., & White-Schwoch, T. (2015). Unraveling the biology
of auditory learning: A cognitive–sensorimotor–reward
framework. Trends in Cognitive Sciences, 19, 642–654.
https://doi.org/10.1016/j.tics.2015.08.017, PubMed: 26454481
Macaluso, E. (2010). Orienting of spatial attention and the
interplay between the senses. Cortex, 46, 282–297. https://doi
.org/10.1016/j.cortex.2009.05.010, PubMed: 19540475
Macaluso, E., & Driver, J. (2005). Multisensory spatial
interactions: A window onto functional integration in the
human brain. Trends in Neurosciences, 28, 264–271. https://
doi.org/10.1016/j.tins.2005.03.008, PubMed: 15866201
McDonald, J. J., Teder-Salejarvi, W. A., Di Russo, F., & Hillyard,
S. A. (2005). Neural basis of auditory-induced shifts in visual
time-order perception. Nature Neuroscience, 8, 1197–1202.
https://doi.org/10.1038/nn1512, PubMed: 16056224
McDonald, J. J., Teder-Salejarvi, W. A., & Hillyard, S. A. (2000).
Involuntary orienting to sound improves visual perception.
Nature, 407, 906–908. https://doi.org/10.1038/35038085,
PubMed: 11057669
Moher, J., Lakshmanan, B. M., Egeth, H. E., & Ewen, J. B.
(2014). Inhibition drives early feature-based attention.
Psychological Science, 25, 315–324. https://doi.org/10.1177
/0956797613511257, PubMed: 24390823
Morey, R. D. (2008). Confidence intervals from normalized data:
A correction to Cousineau (2005). Tutorials in Quantitative
Methods for Psychology, 4, 61–64. https://doi.org/10.20982
/tqmp.04.2.p061
Norris, C. J., Chen, E. E., Zhu, D. C., Small, S. L., & Cacioppo, J. T.
(2004). The interaction of social and emotional processes in
the brain. Journal of Cognitive Neuroscience, 16, 1818–1829.
https://doi.org/10.1162/0898929042947847, PubMed: 15701231
Orban, G. A., Van Essen, D., & Vanduffel, W. (2004).
Comparative mapping of higher visual areas in monkeys and
humans. Trends in Cognitive Sciences, 8, 315–324. https://
doi.org/10.1016/j.tics.2004.05.009, PubMed: 15242691
Qi, S., Zeng, Q., Ding, C., & Li, H. (2013). Neural correlates
of reward-driven attentional capture in visual search. Brain
Research, 1532, 32–43. https://doi.org/10.1016/j.brainres
.2013.07.044, PubMed: 23916733
Salimpoor, V. N., van den Bosch, I., Kovacevic, N., McIntosh, A. R.,
Dagher, A., & Zatorre, R. J. (2013). Interactions between the
nucleus accumbens and auditory cortices predict music
reward value. Science, 340, 216–219. https://doi.org/10.1126
/science.1231059, PubMed: 23580531
Salimpoor, V. N., Zald, D. H., Zatorre, R. J., Dagher, A., &
McIntosh, A. R. (2015). Predictions and the brain: How
musical sounds become rewarding. Trends in Cognitive
Sciences, 19, 86–91. https://doi.org/10.1016/j.tics.2014.12.001,
PubMed: 25534332
Sanz, L. R., Vuilleumier, P., & Bourgeois, A. (2018). Cross-modal
integration during value-driven attentional capture.
Neuropsychologia, 120, 105–112. https://doi.org/10.1016/j
.neuropsychologia.2018.10.014, PubMed: 30342964
Serences, J. T. (2008). Value-based modulations in human visual
cortex. Neuron, 60, 1169–1181. https://doi.org/10.1016/j
.neuron.2008.10.051, PubMed: 19109919
Serences, J. T., & Saproo, S. (2010). Population response
profiles in early visual cortex are biased in favor of
more valuable stimuli. Journal of Neurophysiology,
104, 76–87. https://doi.org/10.1152/jn.01090.2009, PubMed:
20410360
Stormer, V. S., McDonald, J. J., & Hillyard, S. A. (2009). Cross-
modal cuing of attention alters appearance and early cortical
processing of visual stimuli. Proceedings of the National
Academy of Sciences, U.S.A., 106, 22456–22461. https://doi
.org/10.1073/pnas.0907573106, PubMed: 20007778
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic atlas
of the human brain: 3-dimensional proportional system: An
approach to cerebral imaging. Stuttgart, New York: Georg
Thieme.
Theeuwes, J. (1991). Categorization and identification of
simultaneous targets. Acta Psychologica, 76, 73–86. https://
doi.org/10.1016/0001-6918(91)90054-4, PubMed: 1862725
Theeuwes, J. (1992). Perceptual selectivity for color and form.
Perception & Psychophysics, 51, 599–606. https://doi.org/10
.3758/BF03211656, PubMed: 1620571
Wikman, P., Rinne, T., & Petkov, C. I. (2019). Reward cues
readily direct monkeys’ auditory performance resulting in
broad auditory cortex modulation and interaction with sites
along cholinergic and dopaminergic pathways. Scientific
Reports, 9, 1–17. https://doi.org/10.1038/s41598-019-38833-y,
PubMed: 30816142
Winston, J. S., Gottfried, J. A., Kilner, J. M., & Dolan, R. J. (2005).
Integrated neural representations of odor intensity and
affective valence in human amygdala. Journal of Neuroscience,
25, 8903–8907. https://doi.org/10.1523/JNEUROSCI.1569-05
.2005, PubMed: 16192380
Wolfe, J. M., Cave, K. R., & Franzel, S. L. (1989). Guided search:
An alternative to the feature integration model for visual
search. Journal of Experimental Psychology: Human
Perception and Performance, 15, 419. https://doi.org/10
.1037/0096-1523.15.3.419
Xu, K. Z., Anderson, B. A., Emeric, E. E., Sali, A. W., Stuphorn,
V., Yantis, S., et al. (2017). Neural basis of cognitive control
over movement inhibition: Human fMRI and primate
electrophysiology evidence. Neuron, 96, 1447–1458.
https://doi.org/10.1016/j.neuron.2017.11.010, PubMed:
29224723
Yamamoto, S., Kim, H. F., & Hikosaka, O. (2013). Reward value-
contingent changes of visual responses in the primate
caudate tail associated with a visuomotor skill. Journal of
Neuroscience, 33, 11227–11238. https://doi.org/10.1523
/JNEUROSCI.0318-13.2013, PubMed: 23825426
Yamamoto, S., Monosov, I. E., Yasuda, M., & Hikosaka, O.
(2012). What and where information in the caudate tail
guides saccades to visual objects. Journal of Neuroscience,
32, 11005–11016. https://doi.org/10.1523/JNEUROSCI.0828
-12.2012, PubMed: 22875934
Kim, Grégoire, and Anderson
191
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
8
0
2
0
0
7
6
8
5
/
j
o
c
n
_
a
_
0
1
7
8
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3