Using Primary Reinforcement to Enhance Translatability
of a Human Affect and Decision-Making
Judgment Bias Task
Vikki Neville1
, Peter Dayan2,3*, Iain D. Gilchrist1*,
Elizabeth S. Paul1*, and Michael Mendl1*
Abstracto
■ Good translatability of behavioral measures of affect (emo-
ción) between human and nonhuman animals is core to compar-
ative studies. The judgment bias ( JB) tarea, which measures
“optimistic” and “pessimistic” decision-making under ambiguity
as indicators of positive and negative affective valence, ha sido
used in both human and nonhuman animals. Sin embargo, one key
disparity between human and nonhuman studies is that the for-
mer typically use secondary reinforcers (p.ej., dinero) mientras que el
latter typically use primary reinforcers (p.ej., alimento). To address this
deficiency and shed further light on JB as a measure of affect, nosotros
developed a novel version of a JB task for humans using primary
reinforcers. Data on decision-making and reported affective state
during the JB task were analyzed using computational modeling.
En general, participants grasped the task well, and as anticipated,
their reported affective valence correlated with trial-by-trial varia-
tion in offered volume of juice. Además, previous findings from
monetary versions of the task were replicated: More positive pre-
diction errors were associated with more positive affective va-
lence, a higher lapse rate was associated with lower affective
arousal, and affective arousal decreased as a function of number
of trials completed. There was no evidence that more positive va-
lence was associated with greater “optimism,” but instead, allá
was evidence that affective valence influenced the participants’
decision stochasticity, whereas affective arousal tended to influ-
ence their propensity for errors. This novel version of the JB task
provides a useful tool for investigation of the links between pri-
mary reward and punisher experience, afectar, and decision-
haciendo, especially from a comparative perspective. ■
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
oh
C
norte
_
a
_
0
1
7
7
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INTRODUCCIÓN
An important goal in cognitive neuroscience, psychophar-
macology, and affective science is the development of
translational tasks that can be used to assess affective
(emotional) estados. Por ejemplo, the development of novel
treatments for affective disorders in humans depends on
the use of animal models of affect (Rupniak, 2003).
Because we cannot simply ask animals to report their af-
fective state, proxy indicators such as tests of anhedonia
(Van der Harst & Spruijt, 2007) or cognitive bias (Mendl,
Burman, parker, & Pablo, 2009) are often designed to sim-
ulate and assess behavioral characteristics observed in
humans experiencing affective states such as depression
or anxiety (American Psychiatric Association, 2013;
williams, Mathews, & MacLeod, 1996; Wright & Bower,
1992; MacLeod, Mathews, & Tata, 1986). The judgment
bias task has been demonstrated to provide a measure of
affective valence (positivity or negativity) in a range of non-
human animals with relatively “optimistic” (risk-seeking)
decision-making being associated with environmental
1University of Bristol, 2Max Planck Institute for Biological
Cibernética, 3University of Tübingen
*These authors contributed equally.
© 2021 Instituto de Tecnología de Massachusetts
or pharmacological manipulations designed to induce
positive affect and relatively “pessimistic” (risk-averse)
decision-making being associated with environmental or
pharmacological manipulations designed to induce more
negative affect (Lagisz et al., 2020; Neville, Nakagawa, et al.,
2020). Since its conception in 2004 by Harding, Pablo,
and Mendl (2004), several studies have used judgment
bias in nonhuman animals as a translational tool to inves-
tigate human affective disorders and pharmacological
treatments for such disorders (Hales, robinson, &
Houghton, 2016; anderson, Munafò, & robinson, 2013;
Papciak, Popik, Fuchs, & Rygula, 2013; Enkel et al., 2010).
Conducting judgment bias studies with humans, OMS
can report how they feel, may help to provide a better
insight into how performance in judgment bias tasks
reflects affective state. It might also help to elucidate the
putative adaptive function of affect-modulated decision-
haciendo, específicamente, leading to an understanding of
precisely how and why rewarding experiences might lead
to positive affect and then to “optimistic” decision-making,
y viceversa. Sin embargo, human judgment bias studies to
date have painted a mixed picture. Whereas some studies
have found that “pessimism” correlates with subjective
reports of more negative affect (Positive and Negative
Affect Schedule [Paul et al., 2010], State–Trait Anxiety
Revista de neurociencia cognitiva 33:12, páginas. 2523–2535
https://doi.org/10.1162/jocn_a_01776
Inventario [Aylward, Hales, robinson, & robinson, 2020;
anderson, Hardcastle, Munafò, & robinson, 2012], Arroyo
Depression Inventory [Daniel-Watanabe, McLaughlin,
Gormley, & robinson, in press], Visual Analog Scale for
ansiedad [Anderson et al., 2012]), other studies have found
no relationship between reported affect and judgment
inclinación (affect grid and Positive and Negative Affect Schedule
[Iigaya et al., 2016], State–Trait Anxiety Inventory [Daniel-
Watanabe et al., in press]) or even that “optimistic” decision-
making is associated with more negative reported affect
(affect grid [Neville, Dayán, Gilchrist, Pablo, & Mendl, 2021]).
There are several differences between the human and
nonhuman animal judgment bias studies that may have
led to these inconsistencies; humans are required to
learn the task in a shorter period (es decir., within an hour-
long session as opposed to over several days for most
nonhuman animals), humans do not live under the highly
controlled conditions typical of laboratory animals, y
there may be social factors influencing human decision-
haciendo (p.ej., wanting to perform well to satisfy the ex-
perimenter). Sin embargo, one difference that might be of
particular significance is that judgment bias studies in
nonhuman animals are typically conducted using primary
reinforcers such as food or electric shocks, whereas all
judgment bias testing in humans has used secondary re-
inforcers such as monetary gain or loss.
Many studies have identified differences between the
neural processing of primary and second reinforcers
(Sescousse, Caldú, Segura, & Dreher, 2013; delgado,
Jou, & Phelps, 2011; Arroyo, Locke, Savine, Jimura, &
Más valiente, 2010; Grimm & Ver, 2000). Por ejemplo, Tiene
been demonstrated that primary rewards are more
strongly represented in evolutionary older brain regions,
such as the anterior insula, whereas secondary rewards
are more strongly represented in evolutionary newer
regiones del cerebro, such as the anterior OFC (Sescousse
et al., 2013; Delgado et al., 2011). Given these differ-
ences, it is important to develop tasks for humans that
utilize primary reinforcers and hence more closely resem-
ble animal tasks and to investigate whether they yield
similar findings to the typical secondary reinforcer hu-
man task. The use of primary reinforcers in this way is
particularly pertinent to our understanding of judgment
bias from a functional and evolutionary perspective, given
that affect is hypothesized both to reflect ongoing and
prior experience of rewards and punishers and also to
mediate the way this experience guides decision-making
(Bach & Dayán, 2017; marshall, Trimmer, houston,
& McNamara, 2013; Nettle & Bateson, 2012; Mendl,
Burman, & Pablo, 2010). Además, primary reinforcers
may tap into fundamental affective processes and pathol-
ogies that underlie affective disorders more reliably than
secondary reinforcers, given their more direct relevance
to our ability to survive and reproduce. Como consecuencia, a
better understand judgment bias as a measure of affect,
we need a task for humans in which we administer primary
reinforcers and in which we can obtain a fine-scale picture
of sequential effects that might be influencing decision-
making and affective state.
Para tal fin, we aimed to develop a translation of the au-
tomated rat judgment bias task (Neville, Rey, et al., 2020;
Jones et al., 2018) for humans that uses primary rein-
forcers: apple juice and cold salty tea (Pauli et al., 2015).
Using this novel variant of the judgment bias task along-
side computational modeling, we additionally aimed to
elucidate the extent to which latent processes underlying
decision-making might relate to subjective experiences of
afectar, premio, and punishment within the task. We hy-
pothesized that more positive reported affect would be
associated with model parameters characterizing biases
toward the “optimistic” response and that, consistente con
previous research (Neville et al., 2021; Rutledge, Skandali,
Dayán, & Dolan, 2014), the absolute favorability (es decir., aver-
age earning rate) of within-task experience would inform
decision-making and the relative favorability (es decir., premio
prediction error) of within-task experience would inform
reported affective valence.
MÉTODOS
Participantes
Thirty-three people from the Bristol Veterinary School
community participated in the study. All participants
provided written informed consent, and the study was
approved by the Faculty of Science Research Ethics
Committee at the University of Bristol. This sample size
was based on a previous human judgment study (39 par-
ticipants, across two conditions [Neville et al., 2021]) y un
previous study using primary reinforcers as part of a con-
ditioning paradigm (29 Participantes [Pauli et al., 2015]).
The inclusion criteria were that the participant enjoyed
apple juice; was not allergic to apple juice, salt, or black
tea; was not hypertensive or had any medical condition
that meant that he or she must limit his or her salt intake;
and was over the age of 18 años. Participants were asked
to abstain from drinking anything (except water) or eating
in the hour before the study. To cover their time and
expenses, participants were paid £7 for the hour-long
session. To encourage full engagement with the study,
participants were informed that the top three ranking
participants in terms of accuracy would receive an addi-
tional £7 bonus.
Apparatus
The task was conducted on a laptop (Dell Latitude), cual
was connected to two syringe pumps (SPLG100, Mundo
Precision Instruments) that were set to pump liquid at a
tasa de 2 mL per minute. Sterilized food-grade PVC tubing
(outer diameter: 11 mm, inner diameter: 8 mm) eran
attached to syringes (Becton Dickinson; 50-mL Plastipak)
that were driven by the syringe pumps. This tubing was
connected via a tube connector to smaller sterilized
2524
Revista de neurociencia cognitiva
Volumen 33, Número 12
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
oh
C
norte
_
a
_
0
1
7
7
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
food-grade PVC tubing (outer diameter: 6 mm, inner
diameter: 4 mm) and held in place in front of the partici-
pant using a retort clamp and stand, the height of which
could be adjusted by the participant at the start of the ex-
perimento. The participants placed the end of these tubes
in their mouth. Individuals made responses on a keyboard
connected to the laptop. The code for the task was written
in MATLAB (MathWorks, Cª) using the PsychToolBox
extensions (Kleiner et al., 2007; Brainard, 1997).
Procedimiento
The methodology was adapted from that of the human
monetary judgment bias task (Neville et al., 2021), cual
itself was a translation of a rodent judgment bias task
(Jones et al., 2018). On each trial of the task, Participantes
were instructed to press and hold the enter key before
being presented with a fixation cross for 1000 mseg
followed by a random dot kinematogram (RDK) dis-
played for 2000 mseg, which across trials varied in direc-
tion of motion (leftward or rightward) and coherence
(proportion of dots moving in a coherent direction:
0.01, 0.02, 0.16, o 0.32). Participants had two options
when the RDK was presented: to release the key before
the end of the 2-sec RDK presentation (“leave”) or con-
tinue to hold the key for 2 segundo (“stay”). The outcome as-
sociated with either response depended on the stage of
training and the direction of the RDK, with one direction
being favorable and requiring a “stay” response to gain
reward and the other being unfavorable and requiring a
“leave” response to avoid punishment (either leftward or
rightward, counterbalanced across participants).
Participants were provided with written instructions
about the rules of the task and were then provided with
practice trials that were composed of two blocks of 48
ensayos. The aim of the first practice block was to introduce
the participants to the task and train them on the correct
responses to the RDK. In this first practice block, la palabra
“correct” was shown on screen for correct responses (es decir.,
those where “stay” was executed when the RDK direction
was favorable, and “leave” was executed when the RDK
direction was unfavorable), and “incorrect” was shown
on screen for incorrect responses. The direction of motion
of the RDK was very easy to detect (coherence = 0.32) en
all trials, and the direction of motion was leftward on 50%
of trials and rightward on the remaining 50%.
The aim of the second practice block was to acquaint
participants with the delivery and taste of the apple juice
(Apple Juice from Concentrate, Morrisons) and salty tea.
Salty tea was prepared each morning as per Pauli et al.
(2015) with two black tea bags (Bettys & Taylors of
Harrogate; Yorkshire Tea) y 29 g of salt per liter of
boiling water, which was chilled before data collection.
We opted to use salty tea as opposed to electric shocks
as we considered this to be a milder punisher (hence less
of an ethical concern) and so that the modality of the
punisher was the same as the reward (es decir., both gustatory).
En tono rimbombante, liquid reinforcers such as juice and salty tea
have successfully been used in fMRI studies (Pauli et al.,
2015; Metereau & Dreher, 2013; kim, Shimojo, &
O’Doherty, 2011), hence their use in our task paves the
way for future investigations of the neural underpinnings
of judgment bias in humans. In this block, the direction of
the RDK was easy to detect (coherence = 0.16) en 50% de
ensayos, of which half were leftward and half were rightward,
and moderately difficult to detect (coherence = 0.02) en
the remaining 50%, of which half were leftward and half
were rightward. The volume of juice received for “stay”
responses when the direction of the RDK was favorable
era 0.457 mL, and likewise, the volume of salty tea
received for “stay” responses when the RDK was unfavor-
able was 0.457 mL. While juice was delivered and also for
3000 msec after delivery, the words “Juice delivered” were
displayed on screen, whereas during salty tea delivery and
para 3000 msec after this, the words “Salty tea delivered”
were displayed on screen. The potential volume of juice
on each trial (es decir., “0.457 mL”) was shown above an
orange-colored bar with a height proportional to the
potential juice volume, similarly the volume of salty tea
(es decir., “0.457 mL”) was displayed below a brown bar posi-
tioned directly below the orange-colored bar with a height
proportional to the potential tea volume. These bars were
shown before the instruction to press and hold the enter
key. The participant received nothing for making the
“leave” response, and the words “Nothing delivered” were
displayed on screen for 3000 mseg. Across all blocks, el
directions and coherence levels of the RDKs were ran-
domized across trials before the start of the study so that
the order of these was identical for all participants.
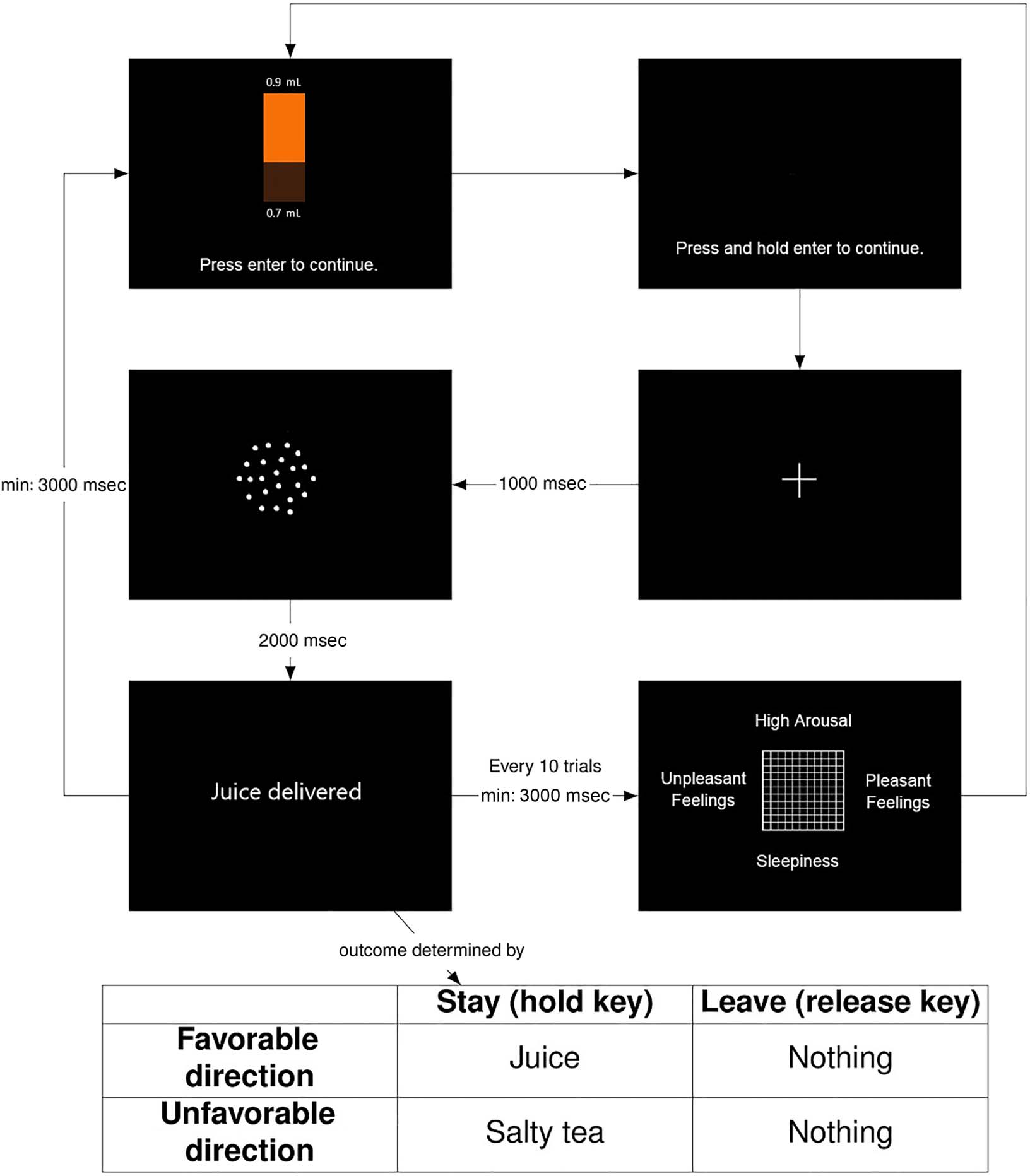
The test session (ver figura 1) comprised 90 trials on
which the RDK moved leftward and 90 trials on which it
moved rightward. For each direction, 30 trials had coher-
ence levels of low (0.01), moderate (0.02), and high
(0.16). RDKs with low and moderate coherence levels
are the ambiguous probe cues, whereas those with a high
coherence level are the reference cues. Por eso, each pos-
sible stimulus was shown on 16.7% of trials. The potential
volume of juice fluctuated across trials according to a
noisy sine wave with a mean volume of 0.457 mL and a
desviación estándar de 0.216 mL, ranging from a minimum
de 0.021 mL to a maximum of 0.781 mL. As a result of
ethical concerns about the effect of large quantities of
salty tea, the potential volume of salty tea remained at
0.457 mL throughout testing. As in the second practice
block, the potential volume of juice and salty tea was dis-
played on screen both as text and graphically using two
colored bars with heights proportional to the volume of
juice and salty tea. Before the first trial and every subse-
quent 10 ensayos, participants were asked to report how
they were feeling using a computerized affect grid
(Killgore, 1998). To complete the affect grid, Participantes
had to move a cross that was initially central in the grid to
the location that best described their current affective
state using the arrow keys on a keyboard. Horizontal
Neville et al.
2525
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
oh
C
norte
_
a
_
0
1
7
7
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
oh
C
norte
_
a
_
0
1
7
7
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. Structure of the human primary reward and punisher judgment bias test session: (1) Participants are shown the potential outcomes of the
“stay” response ( juice volume: orange bar; tea volume: brown bar) and then must press “enter”; (2) participants are instructed to press and hold the
enter key; (3) participants are shown a fixation cross for 1000 mseg; (4) participants are presented with an RDK for 2000 mseg, during which they
must either continue holding the enter key (“stay”) or release the enter key (“go”); (5) participants receive the reward or punishment and are shown
the outcome of their action (which is also determined by the true direction of the RDK); y (6) after at least 3000 mseg, either the next trial starts or
the participant is asked to complete an affect grid (after every 10 ensayos).
movements represented changes in affective valence,
with movements to the right reporting a more positively
valenced affect. Vertical movements represented arousal,
with upward movement reporting higher levels of arousal.
Análisis de los datos
Two participants were excluded from data analysis because
of poor performance at the reference cues, which we
defined as not making the correct response significantly
above chance across both reference cues according to a
one-tailed binomial test (Participant 12, pag = .18; Participant
29, pag = .99).
We conducted both a model-dependent and model-
agnostic analysis of the data. The aim of the model-
dependent analysis was to investigate the latent processes
involved in decision-making and how these might be
modulated by different aspects of reward and punisher
experiencia. Judgment bias RT data (“stay”: 2 segundo; “leave”:
<2 sec) were fitted to the partially observable Markov
decision process (POMDP) model described in full by
Neville et al. (2021). Briefly, we consider that participants
transition through a 2-D state space which they
2526
Journal of Cognitive Neuroscience
Volume 33, Number 12
accumulate
evidence about true direction the
presented stimulus as informed by their observations
(Dimension 1) across discretized duration trial
(Dimension 2). The probability on given trial an
individual will opt for safe “leave” response, and
the speed with they do so, depend their
transitions and value occu-
pying each state.
In model, movement space
and values are determined number parame-
ters including those characterizing psychometric
function, namely, σ, reflects ability the
participant discriminate between stimuli, λref
and λamb, lapse rates reference and
ambiguous respectively; characterizing
the hazard function (probability “timeout,” i.e., time
elapsed> 2 segundo, given “timeout” has not already occurred)
to account for the possibility of making the “stay” response
by default because of inaccurate timekeeping (’ and ζ);
and an inverse temperature parameter (B) to reflect deci-
sion stochasticity. Además, we included a set of param-
eters that allowed for biases toward or away from the
“optimistic” response. This set of parameters was com-
posed of a baseline bias parameter (βδ
0) capturing overall
tendencies for risk-seeking or risk aversion, así como el
overall dislike for the salty tea and enjoyment of the apple
juice, and parameters that encompassed the potential
effect of the average earning rate (βδ
(cid:1)R ), most recent
outcome (βδ
wPE ), y
squared weighted prediction error (βδ
wPE2 ) on bias, también
as a parameter that allowed for decision-making to vary as
a function of the number of trials completed (βn) a
account for fatigue. The average earning rate reflects the
learnt value of the test session from previous juice and
salty tea intake and updates according to a Rescorla–
Wagner learning model, with learning rate α(cid:1)R (following
Neville et al., 2021; Guitarte-Masip, Beierholm, Dolan,
Duzel, & Dayán, 2011), whereas the weighted reward
prediction error is the difference between the model-
predicted outcome and the actual outcome across trials
weighted such that the influence of past prediction errors
attenuates over trials, with forgetting factor γwPE (following
Neville et al., 2021; Rutledge et al., 2014).
oh ), weighted prediction error (βδ
As the task used food rewards, we included a parameter
(κ) that captured the potential effect of satiation as juice
intake increased. Específicamente, the subjective worth (Rn)
depended on the offered juice volume (in milliliters) en
trial n via an exponent that varied as a function of total
juice intake,
PAG
n−1
i¼0 Ri:
!
Xn−1
1−κ
Ri
i¼0
R0
norte
¼ R
norte
An exponent was chosen to reflect the nonlinearity of util-
ity functions and, more specifically, to capture the typical
concavity of utility functions; increases in reward gain lead
to diminishing marginal increases in the subjective value
of those gains (Hsee & Rottenstreich, 2004; kahneman
& Tverski, 1979). As the offered volume of juice is always
lower than 1, a negative value of κ would represent
adherence with the law of diminishing marginal utility,
whereas a value of greater than zero would lead to the
opposite (es decir., a steep increase in the subjective value of
the juice with additional consumption).
Models were fitted to the RT data using maximum like-
lihood, with multiple starting values (including values
found to provide the best fit for the core model as starting
valores) because of nonconvexity. Parameters that charac-
terized decision-making in the absence of biases (es decir., B,
λamb, λref σ, ', and ζ) were included in all models to ac-
count for their potential influence on behavior. Nosotros entonces
added parameters in a stepwise manner, first assessing
whether the parameter that characterized constant biases
in decision-making improved model parsimony and then
those that characterized within-task variation in decision-
haciendo. We then assessed whether the addition of single
parameters to the best-fitting model would further improve
the model fit. Models were compared using Bayesian infor-
mation criterion (BIC) valores, and we additionally com-
pared the final set of models using the Akaike information
criterion (AIC). A stepwise model-fitting procedure was
employed as fitting all possible models was not feasible
because of the large number of possible models and the
computational intensiveness of model fitting. Model fitting
was carried out using the computational facilities of the
Advanced Computing Research Centre at the University
of Bristol.
The parameter estimates from the most parsimonious
model were analyzed using permutation tests to assess
whether they varied significantly from zero, where this
was meaningful (es decir., for βδ
0 and κ).
The aim of the model-agnostic analysis was to investigate
the relationship, primero, between primary reward and
punisher experience and reported affect and, segundo,
between the parameters characterizing decision-making
and reported affect. These analyses were conducted in R
(R Core Team, 2015) using the nlme package (Pinheiro,
Bates, DebRoy, Sarkar, & R Core Team, 2017). Likelihood
ratio tests were used to assess whether the difference in
model deviance was significant after removal of a parame-
ter from a model.
The reported valence (affect grid x coordinate) and re-
ported arousal (affect grid y coordinate) were analyzed
using general linear mixed models (GLMMs) en el cual
both the intercept and slope were allowed to vary among
Participantes, with the predictor variables for which a ran-
dom slope was included determined using BIC compari-
son (es decir., comparing a model with the variable included in
the random effects structure to the model without the
variable included in the random effects structure). El
predictor variables were those encompassing reward
and punisher experience, included those derived from
the best-fitting POMDP model: the most recent volume
Neville et al.
2527
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
oh
C
norte
_
a
_
0
1
7
7
6
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
of offered juice, weighted reward prediction error (wPE),
squared weighted reward prediction error (wPE2), el
previous outcome O, total juice consumed, the number
of trials completed, and the average earning rate (cid:1)R.
Because of correlations between wPE, wPE2, and O, y
also between total juice consumed, number of trials com-
pleted, y (cid:1)R, these predictor variables were not included
in the same GLMM but instead were included separately,
and each model was then compared using their BIC
valores. The GLMMs that provided the best fit were
analyzed further.
Además, the parameter estimates from the POMDP
model were analyzed using a general linear model with
the mean reported arousal and mean reported valence
as fixed effects, except ζ and ’, in which the mean timeout
probability that was jointly determined by these two pa-
rameters was instead used for a more intuitive interpre-
tation of the results, as in Neville et al. (2021). The values
of B were log-transformed because of the presence of ex-
treme outliers exerting undue influence on the model;
the GLMM was also run after removal of these outliers.
RESULTADOS
Judgment Bias
The most parsimonious model of judgment bias RT, ac-
cording to the BIC values, included the following core pa-
rameters: B, λamb, λref σ, ', and ζ, which characterized
decision-making in the absence of biases, in addition to
βδ
0, characterizing baseline biases, and κ, characterizing
the rate of satiation (ver tabla 1). This model fit the ob-
served data well (ver figura 2). The AIC values indicated
that inclusion of βδ
wPE outperformed the BIC best
modelo. Sin embargo, permutation tests revealed that the es-
timates of these parameters did not differ significantly
from zero (βδ
(cid:1)R : mean = −0.13, SE = 0.11, pag = .28;
βδ
wPE: mean = 0.005, SE = 0.01, pag = .72). Por eso, allá
was no strong evidence for the inclusion of these param-
eters, and we selected the BIC best model as our final
model for further analysis.
(cid:1)R or βδ
Decision-Making and Reported Affect
Visual inspection of the parameter estimates revealed
several potential outliers. These were formally defined
as estimates that were either above the third quartile or
below the first quartile by more than 1.5 times the inter-
quartile range (es decir., below Q1–1.5 × IQR or above Q3 +
1.5 × IQR). All analyses were conducted both before and
after the removal of these outliers. The presented results
are from the analyses including the outliers, excepto
where the removal of outliers changed the results quali-
tatively, in which case both results are presented.
The parameter characterizing satiation was significantly
lower than zero (κ: mean = −5.84, SE = 4.23, pag < .001),
reflecting the fact that participants valued juice less with
Table 1. Comparison of POMDP Models
Model
wPE
σ, λamb, λref, ζ, ’, B
σ, λamb, λref, ζ, ’, B, βδ
0
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’’’’’’’’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’’’’’’’’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
σ, λamb, λref, ζ, ’, B, βδ
0
σ, λamb, λref, ζ, ’, B, βδ
0
0, βδ
(cid:1)R
0, βδ
(cid:1)R, α(cid:1)R
0, βδ
0, βδ
0, βδ
0, βδ
0, βδ
0, βn
0, κ
0, κ, βδ
(cid:1)R
0, κ, βδ
0, κ, βδ
; κ, βδ
O
; κ, βn
O
wPE
wPE2
wPE, γwPE
wPE2
wPE2 , γwPE
AIC
BIC
15024.49
15656.70
13652.29
14389.86
13600.18
14443.12
13560.34
14508.65
13646.15
14489.09
13704.29
14652.60
13662.85
14505.80
13590.04
14538.35
13676.61
14519.55
13528.49
14371.43
12641.70
13484.64
12639.33
13587.64
12600.02
13548.33
12991.77
13940.08
12700.81
13649.12
12700.61
13648.92
Values in bold highlight the minimum value for each comparison
method.
increasing juice consumption. Overall, the subjective
value of the juice depreciated across the test session and,
for much of the test session, remained less appetitive
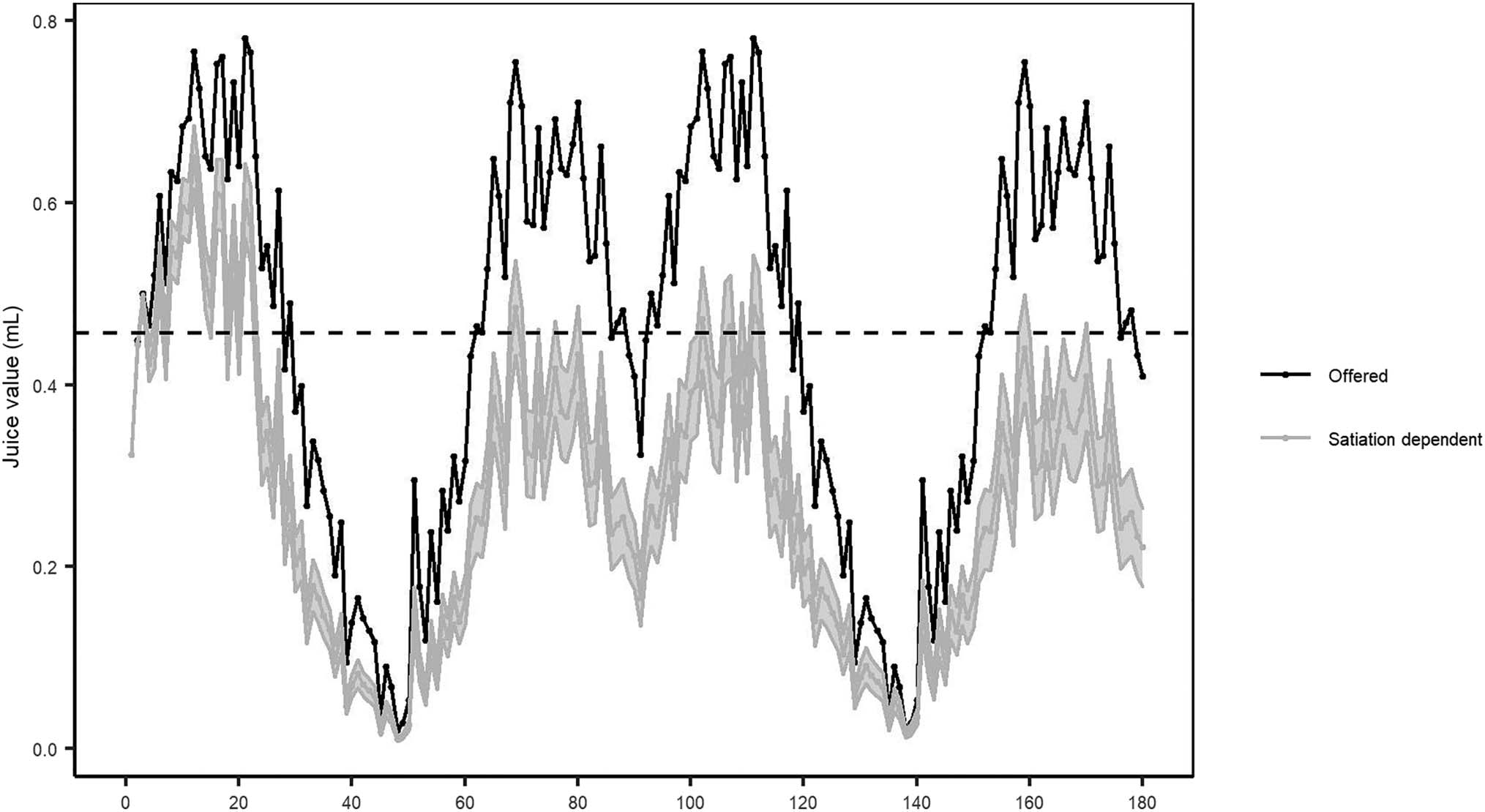
than the salty tea was aversive (Figure 3). This decrease
in the subjective value of the juice exerted a strong influ-
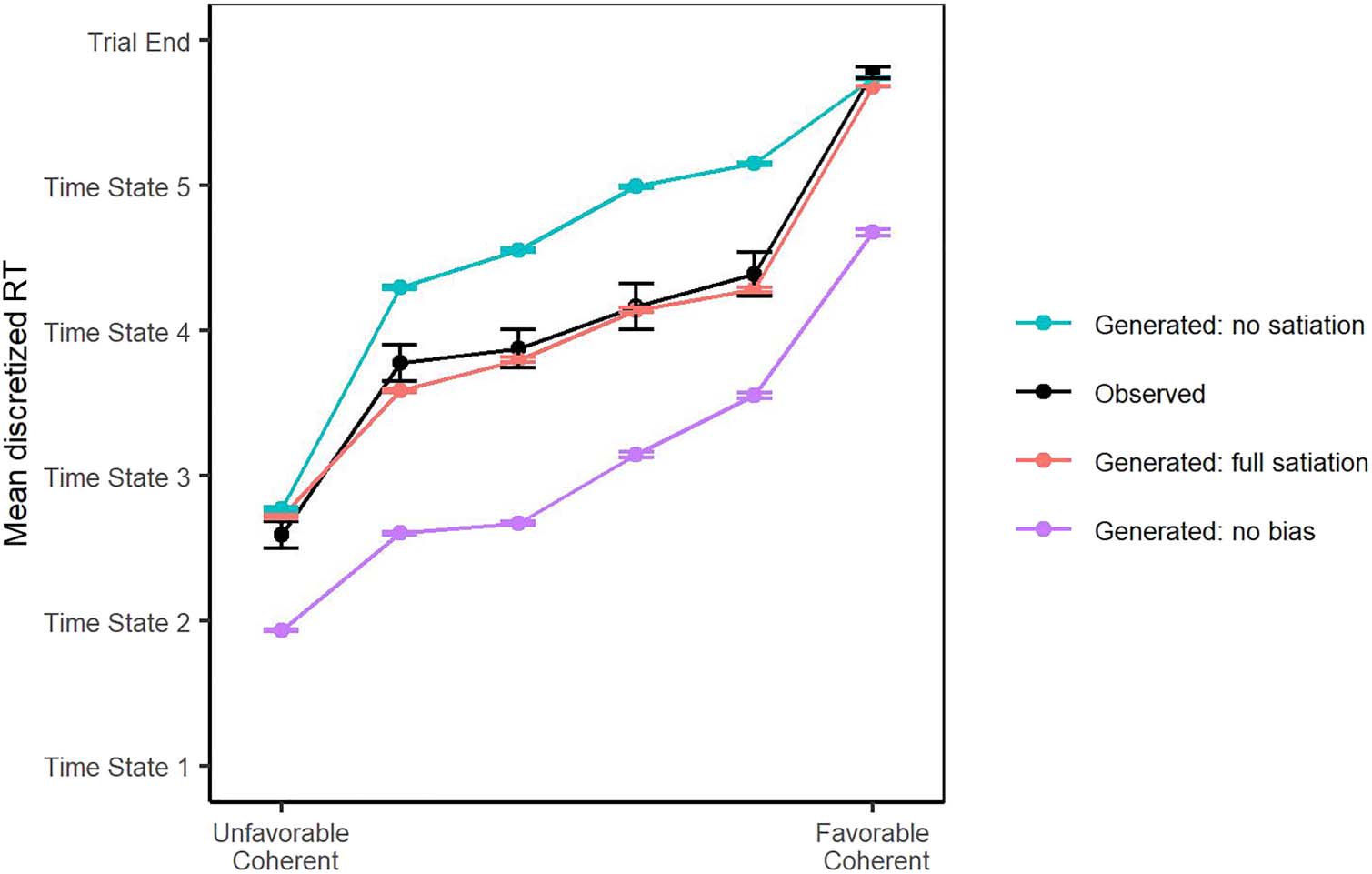
ence on behavior; participants were more risk-averse
than would be expected if there was no effect of satiation
(Figure 4). The effect of satiation was also reflected in
the observed decrease in RTs and proportion of “stay”
responses made as the number of trials increased
(Figure 5). The estimates of κ were not associated with
either reported valence (beta weight = −5.72, SE =
4.64, likelihood ratio test [LRT] = 2.03, p = .16) or arousal
(beta weight = 0.98, SE = 4.64, LRT = 0.075, p = .79).
The bias parameter was significantly greater than zero
(βδ
0: mean = 0.05, SE < 0.01, p < .001), reflecting the par-
ticipants’ overall bias toward persisting with the “stay”
response for a longer duration on a trial than would
otherwise be anticipated (Figure 4), but this parameter
was not predicted by reported affective valence (beta
weight < 0.01, SE < 0.01, LRT < 0.01, p = .95) or arousal
(beta weight < 0.01, SE < 0.01, LRT = 0.55, p = .46). Thus,
although the value of the juice depreciated considerably
over trials (Figure 3), participants occasionally continued
to risk making the “stay” response, which was driven by
this fixed bias. There was no evidence that the extent of
this bias, or the speed of satiation, was influenced by
affective state.
2528
Journal of Cognitive Neuroscience
Volume 33, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
o
c
n
_
a
_
0
1
7
7
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
o
c
n
_
a
_
0
1
7
7
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
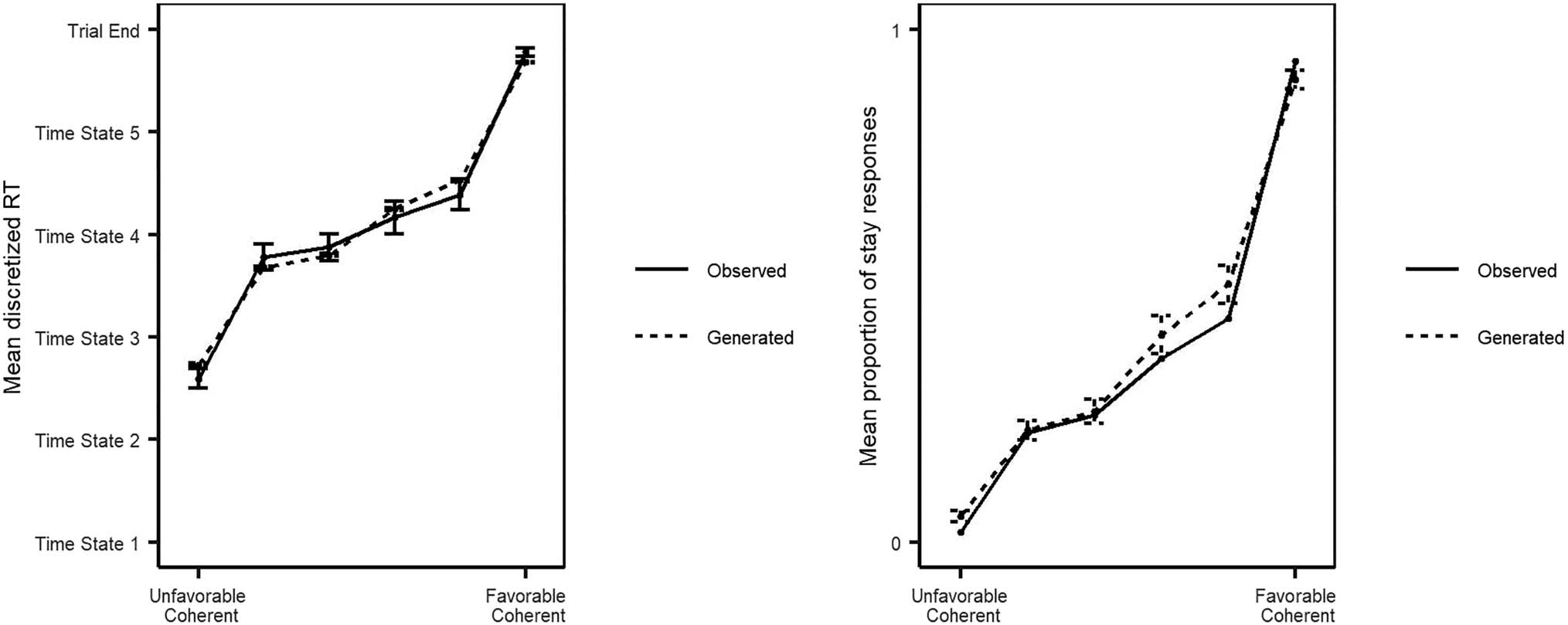
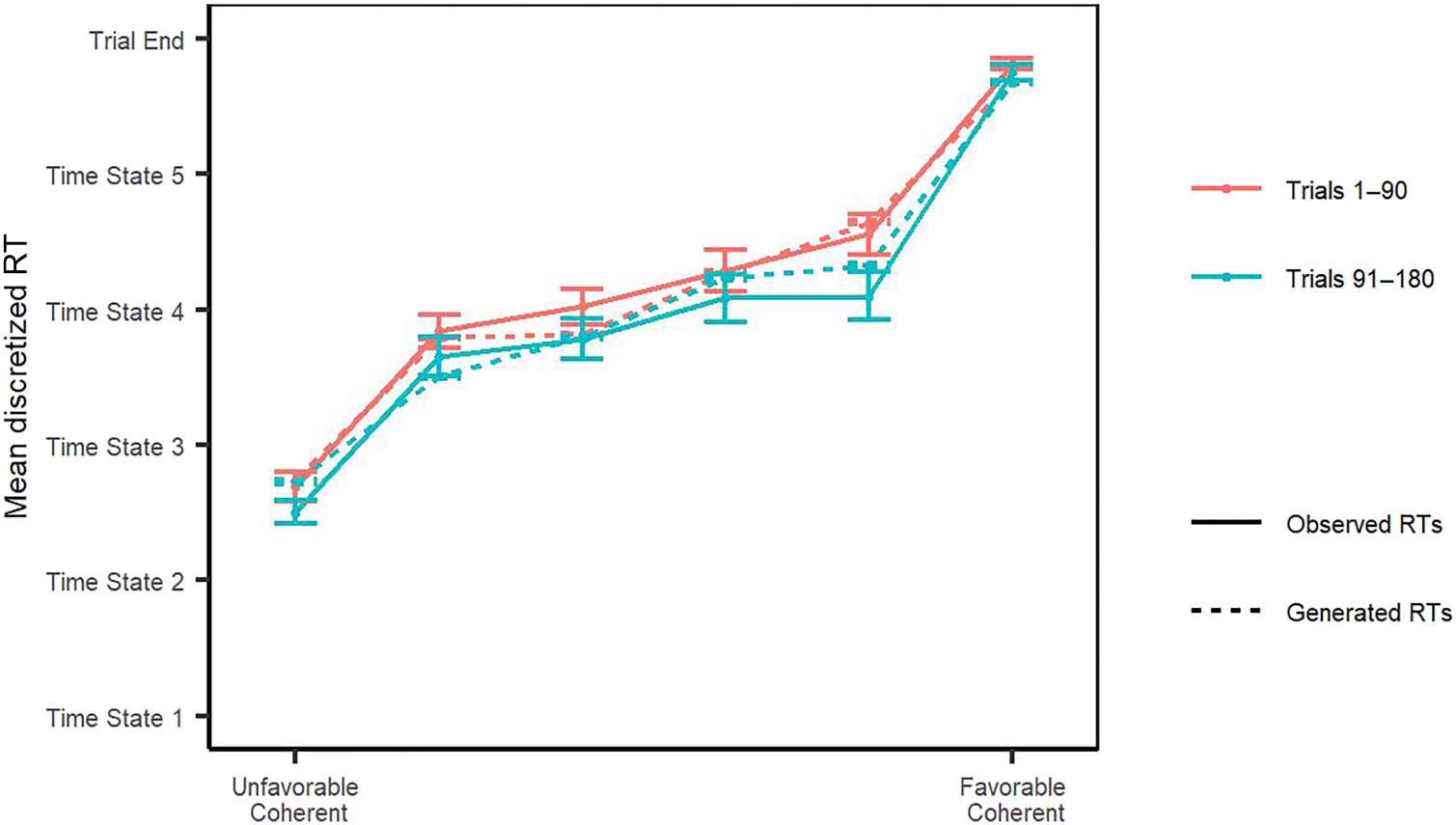
Figure 2. Observed versus generated RT data. The mean discretized RT and mean proportion of stay responses for each stimulus level for both the
model-generated and observed judgment bias data. Error bars represent 1 SE.
Participants who reported lower arousal tended to
make more errors to the ambiguous stimulus; the associ-
ation between the lapse rate and reported arousal was
marginally nonsignificant (λamb: beta weight = −0.02,
SE = 0.01, LRT = 3.77, p = .052). The reference lapse rate
did not depend on reported arousal (λref: beta weight <
0.01, SE < 0.01, LRT = 2.04, p = .15). Neither λamb (beta
weight < 0.01, SE = 0.01, LRT < 0.01, p = .93) nor λref
(beta weight < 0.01, SE < 0.01, LRT = 2.13, p = .15)
depended on reported valence. The slope parameter, σ,
did not significantly depend on reported affective valence
(beta weight = 0.04, SE = 0.04, LRT = 1.10, p = .30) or
arousal (beta weight = 0.03, SE = 0.04, LRT = 0.78, p =
.38). Likewise, the mean timeout probability (determined
by ζ and ’) was not significantly associated with reported
affective valence (beta weight < 0.01, SE = 0.03, LRT <
0.01, p = .95) or arousal (beta weight = 0.01, SE = 0.03,
LRT = 0.41, p = .52).
Figure 3. The value of juice across trials; the black line reflects the offered value, and the gray line reflects the mean satiation-dependent value across
participants as determined by the POMDP model. The dashed line shows the threatened volume of salty tea. The shaded error bars represent 1 SE.
Neville et al.
2529
Figure 4. Observed versus
generated RT data where there
is full satiation or is no satiation,
and the generated RT data
where there is no bias. The
mean discretized RT for each
stimulus level for both the
model-generated (with κ = 0,
βδ
0 = 0, and cumulative juice
intake equal to the maximum
juice intake for the participant)
and observed judgment bias
data. Error bars represent 1 SE.
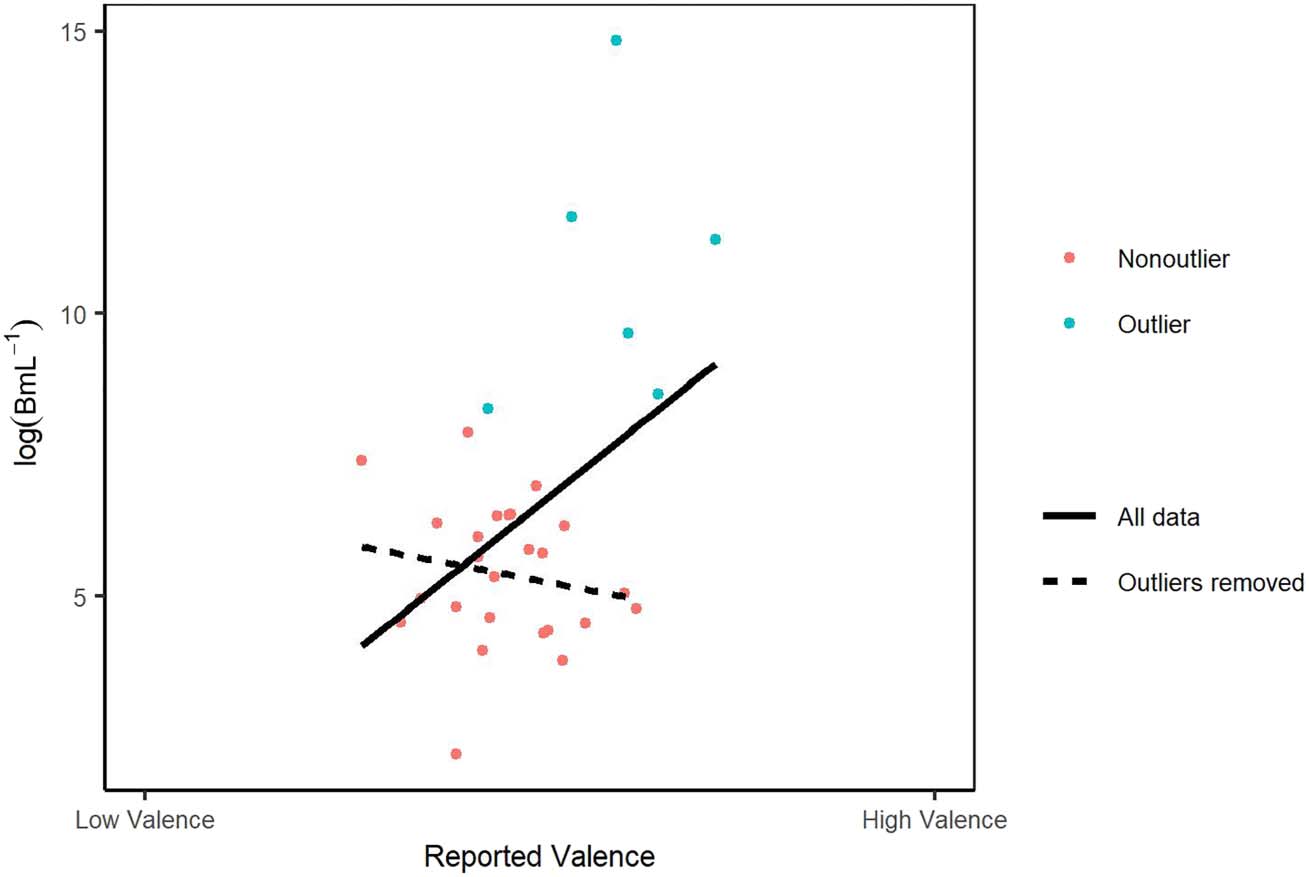
Participants who reported more positive valence had
significantly higher inverse temperature parameter values
(B: beta weight = 1.05, SE = 0.48, LRT = 4.92, p = .027;
Figure 6), and there was no significant relationship
between affective arousal and B (beta weight = 0.22,
SE = 0.48, LRT = 0.23, p = .63). However, after outlier
removal, no significant effect of valence (beta weight =
−0.21, SE = 0.30, LRT = 0.52, p = .47) or arousal (beta
weight = −0.46, SE = 0.28, LRT = 2.97, p = .085) on
inverse temperature was observed (Figure 6). The median
value of the inverse temperature parameters was 336.4297
with an interquartile range of 1224.997, and any value over
3172.95 was deemed to be an outlier. The six outliers iden-
tified, whose removal altered the results qualitatively, had
values of 4099.67, 5309.31, 82248.43, 15654.76, 121346.67,
and 2779806.37 (Figure 6).
Reported Affect and Reward and
Punisher Experience
The best-fitting GLMM of reported valence included the
potential outcome as solely a fixed effect (ΔBIC = 12.27
vs. inclusion as a random linear effect), number of trials
completed with a slope coefficient that varied among
participants (ΔBIC = 26.91 vs. solely a fixed effect; as
opposed to the average earning rate − ΔBIC = 26.75,
and total juice consumed − ΔBIC = 1.10), and weighted
prediction error as solely a fixed effect (ΔBIC = 14.98 vs.
Figure 5. Observed versus
generated RT and decision data.
The mean discretized RT
and mean proportion of
stay responses for each
stimulus level for both the
model-generated and observed
judgment bias data split by trials
in the first and second half of
the test session. Error bars
represent 1 SE.
2530
Journal of Cognitive Neuroscience
Volume 33, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
o
c
n
_
a
_
0
1
7
7
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. The relationship
between the log-transformed
inverse temperature parameter
(with unit mL−1) and reported
affective valence with regression
lines for the data including
(solid line) and excluding
(dashed line) outliers.
inclusion as a random linear effect; as opposed to the
squared weighted prediction error − ΔBIC = 54.25, or
previous outcome ΔBIC = 38.14). Reported valence
was positively correlated with both the prediction error
(beta weight = 40.70, SE = 5.34, LRT = 52.14, p <
.001) and potential outcome (beta weight = 14.30,
SE = 4.24, LRT = 11.33, p < .001). The random slope
coefficients for trial did not differ significantly from zero
according to a permutation test (mean = −0.03, SE =
0.12, p = .81).
The best-fitting GLMM of reported arousal included
potential outcome as solely a fixed effect (ΔBIC =
15.34 vs. inclusion as a random linear effect), number
of trials completed with a slope coefficient that varied
among participants (ΔBIC = 71.70 vs. solely a fixed
effect; as opposed to the average earning rate − ΔBIC =
83.80, or total juice consumed − ΔBIC = 0.95), and
squared prediction error as solely a fixed effect (ΔBIC =
18.37 vs. inclusion as a random linear effect; as opposed
to the weighted prediction error − ΔBIC = 1.11, or pre-
vious outcome − ΔBIC = 1.12). A higher potential out-
come tended to be associated with greater reported
arousal (beta weight = 7.06, SE = 4.66, LRT = 2.89,
p = .09). The squared weight prediction error had no
significant effect of affective arousal (beta weight =
4.96, SE = 4.66, LRT = 1.14, p = .26). The random slope
coefficients for trial were overall significantly lower than
zero (mean = −0.52, SE = 0.17, p = .004), suggesting a
decrease in reported arousal as the test session progressed.
DISCUSSION
In this study, we developed a human judgment bias task
that closely mirrored nonhuman animal versions of the
task by using primary, rather than secondary, reinforcers.
We did this to increase the translatability of the task and
to allow more effective use of results from the human
task to shed light on judgment bias as a measure of affect
in nonhuman animals. To achieve this, we modified a hu-
man judgment bias task, which was itself originally de-
signed as a translation of a rodent judgment bias task,
by replacing monetary gain with apple juice and mone-
tary loss with salty tea. We then sought to investigate
the relationship between prior experience, affect, and
decision-making using computational modeling. The task
itself was largely successful, with more than 90% of
participants performing better than chance in their
responses to the reference cues to which they had been
trained. Furthermore, the data produced were highly
amenable to computational modeling.
Our initial analyses demonstrated that the favorability
of the potential outcome of participants’ responses mod-
ulated self-reported affective valence, with a greater vol-
ume of offered juice leading to more positive affective
valence. There was also a tendency for a greater volume
of offered juice to lead to greater reported affective
arousal. This supports the definition of “emotion” (i.e.,
short-term affect) as a state elicited by (anticipated)
rewards or punishers (Mendl & Paul, 2020; Rolls, 2013).
It also indicates that participants were sensitive to the
fluctuating volume of potential apple juice and were,
indeed, perceiving larger volumes of apple juice as more
rewarding. This demonstrates that the manipulation of
reward experience worked well, having induced the
anticipated shifts in affective states. The finding that
positive affective valence was related to recently experienc-
ing rewards that were greater than anticipated (positive
reward prediction errors) aligns with the results of a num-
ber of other studies (Neville et al., 2021; Otto & Eichstaedt,
2018; Rutledge et al., 2014).
Our results provide evidence for a complex relation-
ship between affect and decision-making that could only
Neville et al.
2531
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
o
c
n
_
a
_
0
1
7
7
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
be teased apart through use of computational modeling.
Contrary to our expectations, there was no evidence in
this study that an “optimistic” bias was directly associated
with more positive reported affect. This conflicts with
some (but not all) previous findings from judgment bias
studies in humans and with the meta-analytic findings in
animal studies that relatively positive affective states are
associated with more “optimistic” decision-making
(Lagisz et al., 2020; Neville, Nakagawa, et al., 2020).
Importantly, the meta-analyses identified a small-to-
moderate effect size with high heterogeneity, using
manipulations including those designed to induce
substantial shifts in affective valence (e.g., enriched vs.
barren housing; administration of pharmacological
substances; restraint stress). It is also possible that the
primary reinforcers used in these animal studies may
have been far more salient to their nonhuman subjects
than the juice/salty tea was for the humans in this study
(e.g., animals were food restricted; animals were
maintained on a nonvaried diet). Given this, it is perhaps
not entirely unsurprising that no significant relationship was
found between the parameter estimates encompassing
“optimism” and affective valence in our nonclinical popula-
tion of human participants. These findings, first, raise the
question of whether judgment bias can be sufficiently
sensitive to detect more subtle variation in baseline affect
and, second, highlights that contextual differences in how
humans and animals experience the tasks may present
significant hurdles to the development of truly translational
and directly comparable tests, even given the explicit
attempt.
We did find that, when looking at the data as a whole,
the inverse temperature parameter was associated with
reported valence; lower decision stochasticity was associ-
ated with more positive reported affect. This result is
consistent with findings that depression, a clinically neg-
atively valenced affective state, has been associated with
more stochastic decision-making (Harlé, Guo, Zhang,
Paulus, & Yu, 2017; Huys, Pizzagalli, Bogdan, & Dayan,
2013). However, this result appeared to be driven by a
subset of participants whose data revealed large outlying
estimates of the inverse temperature parameter and who
tended to report affective valence at the more positive
end of the scale. The cause of the outlying values is
unclear; speculatively, it could reflect a trait characterized
by heightened sensitivity to rewards and punishers and
overall more positive affect. It could also reflect that
the model fits particularly well to these participants
(given that the inverse temperature parameter can also
capture model misfit), although the reasons for this are
not apparent. Nonetheless, the results of our study imply
a relationship between affective valence and the cogni-
tive processes underlying judgment bias. The potential
link between decision stochasticity and affect requires
further investigation.
We also identified a weak relationship between affec-
tive arousal and the cognitive processes underlying
judgment bias. In particular, a higher ambiguous stimulus
lapse rate tended to be associated with lower arousal,
which corroborates results from the monetary version
of the task (Neville et al., 2021) and likely reflects that
higher arousal may lead to higher engagement with the
task and result in better performance. It would therefore
be useful to assess whether lapse rates, extracted using
computational modeling, could provide a measure of
arousal in judgment bias tasks for nonhuman animals.
Our results also highlighted the influence of reward
and punisher experience on both affect and decision-
making. First, the total juice consumed by the participant
was the largest experiential contributor to their decision-
making. The value of the juice decreased as a function of
increasing juice intake, which is consistent with partici-
pants becoming satiated. In accordance with the findings
of the monetary version of the task (Neville et al., 2021),
we also found decreasing arousal as more trials had been
completed, which may indicate that participants found
the task to be tiring. Greater consideration should thus
be given to the potential for satiation and potential fa-
tigue (and individual differences that lead to variation
in how rapidly an individual becomes sated) when con-
ducting nonhuman animal judgment bias tasks.
The degree of satiation indicated in our human partic-
ipants was arguably extreme; the absolute subjective value
(i.e., subjective magnitude) of the juice remained largely
below the value of the salty tea after approximately 20
trials. Yet, despite this, and even after accounting for errors
that were characterized by a lapse rate, participants con-
tinued to make the “optimistic” response more often than
would be anticipated; they exhibited an overall “optimis-
tic” bias. The precise nature of this bias is unclear. One pos-
sibility is that this bias, particularly at the favorable and
near-favorable cue, reflects the intrinsic reward associated
with making accurate (correct) responses. Similarly, it may
reflect that the feedback provided by the “stay” response
was itself rewarding, given that the “leave” response pro-
vides no feedback about the correct action. This would
be in line with studies that have shown that being correct
is in itself rewarding (Satterthwaite et al., 2012; Han,
Huettel, Raposo, Adcock, & Dobbins, 2010). An alternative
is that the “gamble” of the “stay” may have elicited a reward-
ing frisson of excitement, particularly given the repetitive
and likely dull nature of the task. Studies have shown that
risky decision-making can induce excitement and that this
increases under boredom (Kılıç, van Tilburg, & Igou, 2020;
Binde, 2013; Mercer & Eastwood, 2010). This further high-
lights issues in developing truly translational versions of the
judgment bias task; nonhuman animals may not experi-
ence an intrinsic reward associated with being correct,
and judgment bias tasks for nonhuman animals have been
suggested to be enriching for them (Krakenberg et al.,
2021; Mallien et al., 2016).
The extent to which recent outcomes were better or
worse than anticipated was found to determine self-reported
affect. This is in agreement with previous studies
2532
Journal of Cognitive Neuroscience
Volume 33, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
o
c
n
_
a
_
0
1
7
7
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
demonstrating that prediction error is a key determinant
of subjectively experienced affective valence in humans
(Neville et al., 2021; Otto & Eichstaedt, 2018; Rutledge
et al., 2014), at least in relation to short-term rewarding
experiences.
As outlined above, several findings were consistent
between this task and the monetary version from which
it was adapted: A higher lapse rate is associated with
lower affective arousal, more positive prediction errors
are associated with more positive reported affective
valence, and reported affective arousal decreases as a func-
tion of number of trials completed. However, there were
also a couple of inconsistencies. First, we found no
evidence for a relationship between time estimation and
affective valence, and second, we found no effect of the
average earning rate or previous outcome on judgment
bias (Neville et al., 2021). It is unclear whether this might
stem from differences in the processing of primary and
secondary rewards, or other factors such as satiation
having an overriding effect on behavior. This therefore
warrants further examination.
In summary, we have developed a novel version of the
judgment bias task for humans that uses primary as
opposed to secondary reinforcers and, as a result, is
more comparable to versions of the task designed for
nonhuman animals. We identified a relationship between
decision stochasticity and affective valence. Specifically,
we generally observed higher decision stochasticity in
participants who reported more negatively valenced
affect, although only when a cluster of outlying partici-
pants were included. We also found that lower reported
affective arousal tended to be associated with a greater
propensity for errors. The decision-making processes
underlying judgment bias on this task are hence linked
to both reported affective valence and arousal. In addi-
tion, we confirmed some, but not all, previous findings
that had been observed in a monetary version of the task,
such as the positive association between prediction
errors and reported valence. We conclude that our novel
version of the judgment bias task provides a means to
investigate the relationship between affect and decision-
making in greater depth and in a manner that is more
comparable to animal versions of the task.
Reprint requests should be sent to Vikki Neville, Bristol
Veterinary School, University of Bristol, Langford BS40 5DU,
United Kingdom, or via e-mail: vikki.neville@bristol.ac.uk.
Author Contributions
Vikki Neville: Conceptualization; Data curation; Formal analy-
sis; Investigation; Methodology; Writing—Original draft;
Writing—Review & editing. Peter Dayan: Conceptualization;
Formal analysis; Methodology; Supervision; Writing—
Original draft; Writing—Review & editing. Iain D. Gilchrist:
Conceptualization; Methodology; Supervision; Writing—
Review & editing. Elizabeth S. Paul: Conceptualization;
Methodology; Supervision; Writing—Review & editing.
Michael Mendl: Conceptualization; Methodology;
Supervision; Writing—Original draft; Writing—Review &
editing.
Funding Information
Vikki Neville, Biotechnology and Biological Sciences
R e s e a r c h C o u n c i l ( h t t p s : / / d x . d o i . o r g / 1 0 . 1 3 0 3 9
/501100000268), grant number: BB/ M009122/1. Peter
Dayan, Alexander von Humboldt-Stiftung (https://dx.doi
.org/10.13039/100005156). Peter Dayan, Max-Planck-
Gesellschaft (https://dx.doi.org/10.13039/501100004189).
All authors, Biotechnology and Biological Sciences Research
Council (https://dx.doi.org/10.13039/501100000268),
grant number: BB/T002654/1.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the pro-
portions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/ W = .108, and W/ W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/ W = .102, and W/ W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
REFERENCES
American Psychiatric Association. (2013). Diagnostic and
statistical manual of mental disorders (5th ed.).
Washington, DC: American Psychiatric Association. https://
doi.org/10.1176/appi.books.9780890425596
Anderson, M. H., Hardcastle, C., Munafò, M. R., & Robinson,
E. S. J. (2012). Evaluation of a novel translational task for
assessing emotional biases in different species. Cognitive,
Affective & Behavioral Neuroscience, 12, 373–381. https://
doi.org/10.3758/s13415-011-0076-4, PubMed: 22183974
Anderson, M. H., Munafò, M. R., & Robinson, E. S. J. (2013).
Investigating the psychopharmacology of cognitive affective
bias in rats using an affective tone discrimination task.
Psychopharmacology, 226, 601–613. https://doi.org/10.1007
/s00213-012-2932-5, PubMed: 23239131
Aylward, J., Hales, C., Robinson, E., & Robinson, O. J. (2020).
Translating a rodent measure of negative bias into humans:
The impact of induced anxiety and unmedicated mood and
anxiety disorders. Psychological Medicine, 50, 237–246.
https://doi.org/10.1017/S0033291718004117, PubMed:
30683161
Bach, D. R., & Dayan, P. (2017). Algorithms for survival: A
comparative perspective on emotions. Nature Reviews
Neuroscience, 18, 311–319. https://doi.org/10.1038/nrn.2017
.35, PubMed: 28360419
Neville et al.
2533
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
o
c
n
_
a
_
0
1
7
7
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Beck, S. M., Locke, H. S., Savine, A. C., Jimura, K., & Braver, T. S.
(2010). Primary and secondary rewards differentially
modulate neural activity dynamics during working memory.
PLoS One, 5, e9251. https://doi.org/10.1371/journal.pone
.0009251, PubMed: 20169080
Binde, P. (2013). Why people gamble: A model with five
motivational dimensions. International Gambling Studies,
13, 81–97. https://doi.org/10.1080/14459795.2012.712150
Brainard, D. H. (1997). The Psychophysics toolbox. Spatial
Vision, 10, 433–436. https://doi.org/10.1163/156856897X00357,
PubMed: 9176952
Daniel-Watanabe, L., McLaughlin, M., Gormley, S., & Robinson,
O. J. (in press). Association between a directly translated
cognitive measure of negative bias and self-reported
psychiatric symptoms. Biological Psychiatry: Cognitive
Neuroscience and Neuroimaging, 1–9. https://doi.org/10
.1016/j.bpsc.2020.02.010
Delgado, M. R., Jou, R. L., & Phelps, E. A. (2011). Neural
systems underlying aversive conditioning in humans with
primary and secondary reinforcers. Frontiers in Neuroscience,
5, 71. https://doi.org/10.3389/fnins.2011.00071, PubMed:
21637321
Enkel, T., Gholizadeh, D., von Bohlen und Halbach, O., Sanchis-
Segura, C., Hurlemann, R., Spanagel, R., et al. (2010).
Ambiguous-cue interpretation is biased under stress- and
depression-like states in rats. Neuropsychopharmacology,
35, 1008–1015. https://doi.org/10.1038/npp.2009.204,
PubMed: 20043002
Grimm, J. W., & See, R. E. (2000). Dissociation of primary
and secondary reward-relevant limbic nuclei in an animal
model of relapse. Neuropsychopharmacology, 22, 473–479.
https://doi.org/10.1016/S0893-133X(99)00157-8, PubMed:
10731622
Guitart-Masip, M., Beierholm, U. R., Dolan, R., Duzel, E., &
Dayan, P. (2011). Vigor in the face of fluctuating rates of
reward: An experimental examination. Journal of Cognitive
Neuroscience, 23, 3933–3938. https://doi.org/10.1162/jocn_a
_00090, PubMed: 21736459
Hales, C. A., Robinson, E. S. J., & Houghton, C. J. (2016).
Diffusion modelling reveals the decision making processes
underlying negative judgement bias in rats. PLoS One, 11,
e0152592. https://doi.org/10.1371/journal.pone.0152592,
PubMed: 27023442
Han, S., Huettel, S. A., Raposo, A., Adcock, R. A., & Dobbins, I. G.
(2010). Functional significance of striatal responses during
episodic decisions: Recovery or goal attainment? Journal of
Neuroscience, 30, 4767–4775. https://doi.org/10.1523
/JNEUROSCI.3077-09.2010, PubMed: 20357127
Harding, E. J., Paul, E. S., & Mendl, M. (2004). Animal behaviour:
Cognitive bias and affective state. Nature, 427, 312. https://doi
.org/10.1038/427312a, PubMed: 14737158
Harlé, K. M., Guo, D., Zhang, S., Paulus, M. P., & Yu, A. J. (2017).
Anhedonia and anxiety underlying depressive symptomatology
have distinct effects on reward-based decision-making. PLoS
One, 12, e0186473. https://doi.org/10.1371/journal.pone
.0186473, PubMed: 29059254
Hsee, C. K., & Rottenstreich, Y. (2004). Music, pandas, and
muggers: On the affective psychology of value. Journal of
Experimental Psychology: General, 133, 23–30. https://doi
.org/10.1037/0096-3445.133.1.23, PubMed: 14979749
Huys, Q. J., Pizzagalli, D. A., Bogdan, R., & Dayan, P. (2013).
Mapping anhedonia onto reinforcement learning: A
behavioural meta-analysis. Biology of Mood & Anxiety
Disorders, 3, 12. https://doi.org/10.1186/2045-5380-3-12,
PubMed: 23782813
Iigaya, K., Jolivald, A., Jitkrittum, W., Gilchrist, I. D., Dayan, P.,
Paul, E., et al. (2016). Cognitive bias in ambiguity judgements:
Using computational models to dissect the effects of mild
mood manipulation in humans. PLoS One, 11, e0165840.
https://doi.org/10.1371/journal.pone.0165840, PubMed:
27829041
Jones, S., Neville, V., Higgs, L., Paul, E. S., Dayan, P., Robinson,
E. S. J., et al. (2018). Assessing animal affect: An automated
and self-initiated judgement bias task based on natural
investigative behaviour. Scientific Reports, 8, 12400. https://
doi.org/10.1038/s41598-018-30571-x, PubMed: 30120315
Kahneman, D., & Tversky, A. (1979). Prospect theory: An
analysis of decision under risk. Econometrica, 47, 263–292.
https://doi.org/10.2307/1914185
Kılıç, A., van Tilburg, W. A. P., & Igou, E. R. (2020). Risk-taking
increases under boredom. Journal of Behavioral Decision
Making, 33, 257–269. https://doi.org/10.1002/bdm.2160
Killgore, W. D. S. (1998). The Affect Grid: A moderately valid,
nonspecific measure of pleasure and arousal. Psychological
Reports, 83, 639–642. https://doi.org/10.2466/pr0.1998.83.2
.639, PubMed: 9819938
Kim, H., Shimojo, S., & O’Doherty, J. P. (2011). Overlapping
responses for the expectation of juice and money rewards in
human ventromedial prefrontal cortex. Cerebral Cortex, 21,
769–776. https://doi.org/10.1093/cercor/bhq145, PubMed:
20732900
Kleiner, M., Brainard, D., Pelli, D., Ingling, A., Murray, R., &
Broussard, C. (2007). What’s new in Psychtoolbox-3?
Perception, 36, 1–16.
Krakenberg, V., Wewer, M., Palme, R., Kaiser, S., Sachser, N., &
Richter, S. H. (2021). Regular touchscreen training affects
faecal corticosterone metabolites and anxiety-like behaviour
in mice. Behavioural Brain Research, 401, 113080. https://
doi.org/10.1016/j.bbr.2020.113080, PubMed: 33358914
Lagisz, M., Zidar, J., Nakagawa, S., Neville, V., Sorato, E., Paul, E. S.,
et al. (2020). Optimism, pessimism and judgement bias in
animals: A systematic review and meta-analysis. Neuroscience
& Biobehavioral Reviews, 118, 3–17. https://doi.org/10.1016/j
.neubiorev.2020.07.012, PubMed: 32682742
MacLeod, C., Mathews, A., & Tata, P. (1986). Attentional bias in
emotional disorders. Journal of Abnormal Psychology, 95,
15–20. https://doi.org/10.1037/0021-843X.95.1.15, PubMed:
3700842
Mallien, A. S., Palme, R., Richetto, J., Muzzillo, C., Richter, S. H.,
Vogt, M. A., et al. (2016). Daily exposure to a touchscreen-
paradigm and associated food restriction evokes an increase
in adrenocortical and neural activity in mice. Hormones and
Behavior, 81, 97–105. https://doi.org/10.1016/j.yhbeh.2016
.03.009, PubMed: 27059527
Marshall, J. A. R., Trimmer, P. C., Houston, A. I., & McNamara,
J. M. (2013). On evolutionary explanations of cognitive biases.
Trends in Ecology & Evolution, 28, 469–473. https://doi.org
/10.1016/j.tree.2013.05.013, PubMed: 23790393
Mendl, M., Burman, O. H. P., Parker, R. M. A., & Paul, E. S.
(2009). Cognitive bias as an indicator of animal emotion and
welfare: Emerging evidence and underlying mechanisms.
Applied Animal Behaviour Science, 118, 161–181. https://doi
.org/10.1016/j.applanim.2009.02.023
Mendl, M., Burman, O. H. P., & Paul, E. S. (2010). An integrative
and functional framework for the study of animal emotion
and mood. Proceedings of the Royal Society of London,
Series B, Biological Sciences, 277, 2895–2904. https://doi.org
/10.1098/rspb.2010.0303, PubMed: 20685706
Mendl, M., & Paul, E. S. (2020). Animal affect and decision-
making. Neuroscience and Biobehavioral Reviews, 112,
144–163. https://doi.org/10.1016/j.neubiorev.2020.01.025,
PubMed: 31991192
Mercer, K. B., & Eastwood, J. D. (2010). Is boredom associated
with problem gambling behaviour? It depends on what you
mean by ‘boredom.’ International Gambling Studies, 10,
91–104. https://doi.org/10.1080/14459791003754414
2534
Journal of Cognitive Neuroscience
Volume 33, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
o
c
n
_
a
_
0
1
7
7
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Metereau, E., & Dreher, J.-C. (2013). Cerebral correlates of
salient prediction error for different rewards and punishments.
Cerebral Cortex, 23, 477–487. https://doi.org/10.1093/cercor
/bhs037, PubMed: 22368086
Nettle, D., & Bateson, M. (2012). The evolutionary origins of
mood and its disorders. Current Biology, 22, R712–R721.
https://doi.org/10.1016/j.cub.2012.06.020, PubMed: 22975002
Neville, V., Dayan, P., Gilchrist, I. D., Paul, E. S., & Mendl, M.
(2021). Dissecting the links between reward and loss, decision-
making, and self-reported affect using a computational
approach. PLoS Computational Biology, 17, e1008555. https://
doi.org/10.1371/journal.pcbi.1008555, PubMed: 33417595
Neville, V., King, J., Gilchrist, I. D., Dayan, P., Paul, E. S., &
Mendl, M. (2020). Reward and punisher experience alter
rodent decision-making in a judgement bias task. Scientific
Reports, 10, 11839. https://doi.org/10.1038/s41598-020-68737-1,
PubMed: 32678247
Neville, V., Nakagawa, S., Zidar, J., Paul, E. S., Lagisz, M.,
Bateson, M., et al. (2020). Pharmacological manipulations
of judgement bias: A systematic review and meta-analysis.
Neuroscience & Biobehavioral Reviews, 108, 269–286.
https://doi.org/10.1016/j.neubiorev.2019.11.008, PubMed:
31747552
Otto, A. R., & Eichstaedt, J. C. (2018). Real-world unexpected
outcomes predict city-level mood states and risk-taking
behavior. PLoS One, 13, e0206923. https://doi.org/10.1371
/journal.pone.0206923, PubMed: 30485304
Papciak, J., Popik, P., Fuchs, E., & Rygula, R. (2013). Chronic
psychosocial stress makes rats more ‘pessimistic’ in the
ambiguous-cue interpretation paradigm. Behavioural Brain
Research, 256, 305–310. https://doi.org/10.1016/j.bbr.2013.08
.036, PubMed: 23993861
Paul, E. S., Cuthill, I., Kuroso, G., Norton, V., Woodgate, J., &
Mendl, M. (2010). Mood and the speed of decisions about
anticipated resources and hazards. Evolution and Human
Behavior, 32, 21–28. https://doi.org/10.1016/j.evolhumbehav
.2010.07.005
Pauli, W. M., Larsen, T., Collette, S., Tyszka, J. M., Seymour, B.,
& O’Doherty, J. P. (2015). Distinct contributions of
ventromedial and dorsolateral subregions of the human
substantia nigra to appetitive and aversive learning. Journal
of Neuroscience, 35, 14220–14233. https://doi.org/10.1523
/JNEUROSCI.2277-15.2015, PubMed: 26490862
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., & R Core Team.
(2017). nlme: Linear and nonlinear mixed effects models
(R package version 3.1–131). Retrieved from https://CRAN.R
-project.org/package=nlme.
R Core Team. (2015). R: A language and environment for
statistical computing. Vienna: R Foundation for Statistical
Computing Retrieved from https://www.r-project.org.
Rolls, E. T. (2013). What are emotional states, and why do we
have them? Emotion Review, 5, 241–247. https://doi.org/10
.1177/1754073913477514
Rupniak, N. M. J. (2003). Animal models of depression:
Challenges from a drug development perspective.
Behavioural Pharmacology, 14, 385–390. https://doi.org/10
.1097/01.fbp.0000087738.21047.91, PubMed: 14501252
Rutledge, R. B., Skandali, N., Dayan, P., & Dolan, R. J. (2014). A
computational and neural model of momentary subjective
well-being. Proceedings of the National Academy of
Sciences, U.S.A., 111, 12252–12257. https://doi.org/10.1073
/pnas.1407535111, PubMed: 25092308
Satterthwaite, T. D., Ruparel, K., Loughead, J., Elliott, M. A.,
Gerraty, R. T., Calkins, M. E., et al. (2012). Being right is
its own reward: Load and performance related ventral
striatum activation to correct responses during a working
memory task in youth. Neuroimage, 61, 723–729.
https://doi.org/10.1016/j.neuroimage.2012.03.060, PubMed:
22484308
Sescousse, G., Caldú, X., Segura, B., & Dreher, J.-C. (2013).
Processing of primary and secondary rewards: A quantitative
meta-analysis and review of human functional neuroimaging
studies. Neuroscience & Biobehavioral Reviews, 37,
681–696. https://doi.org/10.1016/j.neubiorev.2013.02.002,
PubMed: 23415703
Van der Harst, J. E., & Spruijt, B. M. (2007). Tools to measure
and improve animal welfare: Reward-related behaviour.
Animal Welfare, 16(Suppl. 1), 67–73.
Williams, J. M. G., Mathews, A., & MacLeod, C. (1996). The
emotional Stroop task and psychopathology. Psychological
Bulletin, 120, 3–24. https://doi.org/10.1037/0033-2909.120.1.3,
PubMed: 8711015
Wright, W. F., & Bower, G. H. (1992). Mood effects on
subjective probability assessment. Organizational Behavior
and Human Decision Processes, 52, 276–291. https://doi.org
/10.1016/0749-5978(92)90039-A
Neville et al.
2535
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
2
2
5
2
3
1
9
7
0
8
5
3
/
/
j
o
c
n
_
a
_
0
1
7
7
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3