Tracking the Contents of Spatial Working Memory
during an Acute Bout of Aerobic Exercise
Jordan Garrett
, Tom Bullock , and Barry Giesbrecht
Abstracto
■ Recent studies have reported enhanced visual responses
during acute bouts of physical exercise, suggesting that sensory
systems may become more sensitive during active exploration of
the environment. This raises the possibility that exercise may also
modulate brain activity associated with other cognitive functions,
like visual working memory, that rely on patterns of activity that
persist beyond the initial sensory evoked response. Aquí, nosotros
investigated whether the neural coding of an object location held
in memory is modulated by an acute bout of aerobic exercise.
Participants performed a spatial change detection task while
seated on a stationary bike at rest and during low-intensity cycling
(∼50 watts/50 RPM). Brain activity was measured with EEG. Un
inverted encoding modeling technique was employed to estimate
location-selective channel response functions from topographical
patterns of alpha-band (8–12 Hz) actividad. There was strong
evidence of robust spatially selective responses during stimulus
presentation and retention periods both at rest and during exercise.
During retention, the spatial selectivity of these responses de-
creased in the exercise condition relative to rest. A temporal gener-
alization analysis indicated that models trained on one time period
could be used to reconstruct the remembered locations at other
time periods, sin embargo, generalization was degraded during exer-
cise. Juntos, these results demonstrate that it is possible to recon-
struct the contents of working memory at rest and during exercise,
but that exercise can result in degraded responses, which contrasts
with the enhancements observed in early sensory processing. ■
INTRODUCCIÓN
Nonhuman animals and invertebrates show robust re-
sponse gain in sensory processing areas during locomotion
when compared to rest (Kaneko, Fu, & Stryker, 2017; Fu
et al., 2014; Ayaz, Saleem, Schölvinck, & Carandini, 2013;
Polack, Friedman, & Golshani, 2013; Keller, Bonhoeffer, &
Hübener, 2012; Maimon, Straw, & Dickinson, 2010; Niell &
Stryker, 2010). More recently, EEG studies suggest that a
similar enhancement in human sensory responses may oc-
cur during bouts of acute physical exercise (Cao & Händel,
2019; Bullock, eliot, serencias, & Giesbrecht, 2017; Bullock,
Cecotti, & Giesbrecht, 2015). If sensory processes are
impacted during bouts of acute aerobic exercise, then this
raises the possibility that higher-order cognitive functions
relying on input from these sensory areas may also be
affected. Aquí, we test whether neural representations of
object locations stored in visual working memory ( W.M.)
in humans are modulated during a bout of acute exercise.
WM is an essential cognitive process that is critical for
maintaining and manipulating information. Serving as the
core interface between multiple cognitive systems (p.ej.,
aprendiendo, atención, percepción, long-term memory), este
process provides a platform for guiding goal-directed be-
haviors. Previous research has produced mixed evidence
regarding modulations in WM during exercise. Por ejemplo,
meta-analytic results indicate that information processing
Universidad de California, Santa Bárbara
© 2021 Instituto de Tecnología de Massachusetts
velocidad (es decir., reaction time) in WM tasks is enhanced during
cycling, while accuracy is diminished (McMorris, Sproule,
Tornero, & Hale, 2011). McMorris et al. proposed that this
pattern does not reflect a speed-accuracy tradeoff, pero
rather is because of increased peripheral and serum levels of
neurotransmisores, cual, Sucesivamente, engenders greater levels
of neural noise in some cortical regions while facilitating
activity in others. Contrary to findings that suggest WM is
impaired during exercise, there is evidence that moderate
intensity exercise boosts the functioning of WM overall
(Martins, Kavussanu, Willoughby, & Ring, 2013). Fur-
thermore, it has been shown that walking at a preferred
speed compared to a seated rest condition can result in en-
hanced performance under high memory loads (Schaefer,
Lövdén, Wieckhorst, & Lindenberger, 2010). Finalmente, estafa-
current exercise also has been found to have no impact
on WM (Lambourne, Audiffren, & Tomporowski, 2010).
Given the multimodal nature of WM (p.ej., verbal, visuospa-
tial, auditory), the heterogeneity of these findings may stem
from exercise differentially affecting each domain. En efecto,
there is evidence for a greater improvement of visuospatial
WM compared to verbal-auditory WM as a result of exercise
(Roig, Nordbrandt, Geertsen, & Nielsen, 2013). Regardless
of the mixed behavioral outcomes, none of these studies
provide insight into whether the underlying neural corre-
lates of WM representations are modulated during exercise.
To investigate whether WM representations are modu-
lated during exercise, in this study, participants performed
Revista de neurociencia cognitiva 33:7, páginas. 1271–1286
https://doi.org/10.1162/jocn_a_01714
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
a visuospatial change detection task at rest and during a
bout of low-intensity cycling exercise while EEG was re-
corded at the scalp. Each trial of this task involved remem-
bering the location of a single memorandum presented at
a pseudorandomized location on the circumference of an
imaginary circle centered on fixation. After a delay period
(1750 mseg), a test stimulus was presented at the same
location or at a different location and the participant indi-
cated whether the location of the test stimulus changed
relative to the location of the memorandum presented
at the beginning of the trial. Previous studies have shown
that patterns of EEG alpha power (∼8–12 Hz) measured
at the scalp covary with the locations of attended and
remembered stimuli (MacLean, Bullock, & Giesbrecht,
2019; Rihs, Michel, & Thut, 2007; Thut, Nietzel, Brandt,
& Pascual Leone, 2006; Sauseng et al., 2005), implicating
a key role for this oscillation in spatial attention and WM.
Aquí, alpha power was modeled using a computational
técnica, known as an inverted encoding model (IEM),
to estimate the location-selective representations of the
remembered locations from the unique patterns of brain
activity recorded during this task. The IEM technique has
been applied to fMRI BOLD activity in visual and parietal
corteza (sprague, Saproo, & serencias, 2015; Brouwer &
Heeger, 2009, 2011, 2013; serencias & Saproo, 2012) como
well as in scalp-recorded EEG (Samaha, sprague, &
Postle, 2016; Garcia, Srinivasan, & serencias, 2013) a
recover feature- or location-selective information from
the patterns coded in brain activity. Previous studies have
applied this technique to patterns of scalp-recorded
oscillatory activity in the alpha frequency band to success-
fully track the locations of items that are stored and main-
tained in WM with high temporal precision (MacLean et al.,
2019; Sutterer, Foster, serencias, Vogel, & Awh, 2019;
Foster, Sutterer, serencias, Vogel, & Awh, 2016). In addi-
ción, previous studies not only demonstrate that EEG is
well suited to recording brain activity from physically
active human participants (Cheron et al., 2016; Bullock
et al., 2015), but also that the IEM technique can be
applied effectively to the EEG steady-state visually
evoked response recorded during cycling (Bullock et al.,
2017). Aquí, the IEM technique was used to reconstruct
spatially selective response profiles from topographical
patterns of alpha-band activity recorded at rest and during
exercise. Replicating previous work, the IEM technique
Mesa 1. Demographic Information
Demographics
Measure
Age (años)
Height (in.)
Weight (lb)
Mean ± SD
21.85 ± 0.55
67.79 ± 0.63
152.67 ± 4.47
revealed evidence for location-specific reconstructions of
item locations held in WM coded in alpha activity at rest.
En tono rimbombante, evidence for location-specific information
was also observed during exercise for much of the retention
período. Direct comparisons of the IEM reconstructions
between rest and exercise revealed evidence for degraded
representations during exercise, but only late in the
retention interval.
MÉTODOS
Participantes
Thirty-four (norte = 34, 17 women; ver tabla 1) adult student
volunteers from the University of California, Santa Bárbara,
community took part in the study in exchange for financial
compensación ($20/hr). All participants completed the
Physical Activity Readiness Questionnaire (National
Academy of Sports Medicine) to determine their eligibility
to participate in aerobic activity. Informed consent was
provided before the study began. All participants reported
normal or corrected-to-normal vision. The procedures de-
tailed below were approved by the University of California,
Santa Bárbara, Human Subjects Committee and the US
Army Human Research Protection Office.
Visual Stimuli
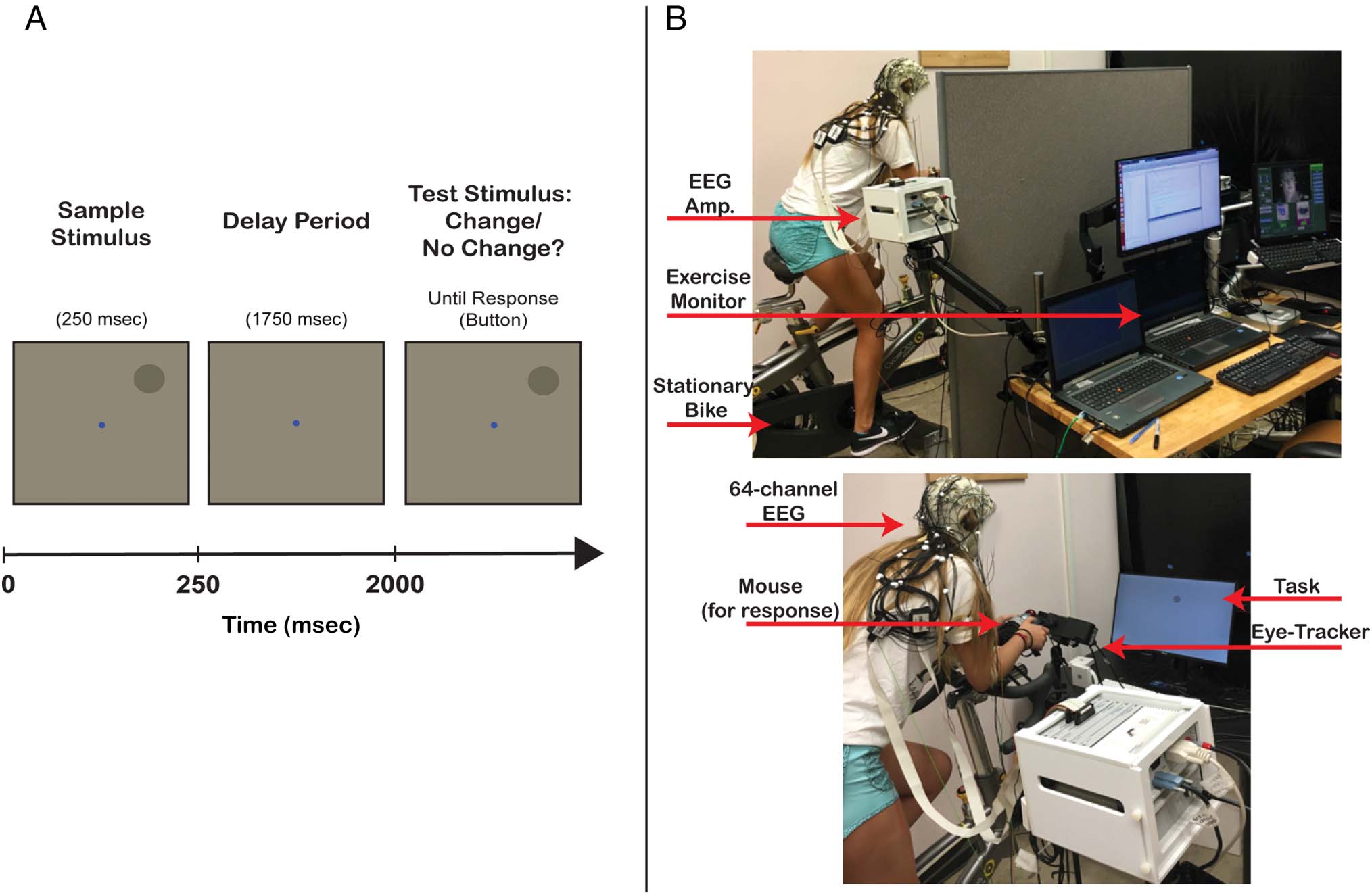
Participants performed a delayed spatial change detection
tarea (Figura 1A) to measure WM performance (Foster
et al., 2016; zhang & Luck, 2008; Wilken & Mamá, 2004). A gray
target circle (subtending 1.6° visual angle) served as the sam-
ple stimulus and appeared centered on a point in an ima-
ginary circle circumventing 4° from a blue fixation dot
(subtending 0.2° visual angle). The sample stimulus was pre-
sented within one of eight equally spaced 45° location bins
relative to fixation (0°, 45°, 90°, 135°, 180°, 225°, 270°, 315°),
with stimulus location jittered randomly within each bin be-
tween +1° and 44°. The task was presented on a 28-in. mon-
itor (ASUS VG278Q, 1920 × 1080) via custom scripts that
used functions from Psychophysics ToolBox for MATLAB
(Brainard, 1997). The viewing distance was ∼100 cm.
Eye-Tracking
Gaze-contingent eye-tracking was employed to ensure
participants remained fixated throughout the trial period,
and to minimize contamination from ocular artifacts in the
EEG signal. Además, this ensured a stable projection
of the visual display on the retina. The eye-tracker (Eyelink
1000, SR Research Ltd.) was positioned 50–70 cm from
both eyes, and binocular tracking sampling at 500 Hz
was enabled. Pupil area and gaze position were collected
throughout the trial period in both conditions. Pupil area
was normalized using the following equation: (x − xmin)/
(xmax − xmin), where x is the area for a given time point.
1272
Revista de neurociencia cognitiva
Volumen 33, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. (A) Delayed spatial change detection task. (B) Experimental setup. Nota, the task was completed in a dark room for optimal eye-tracking
actuación.
Stationary Bike
The stationary bike was a CycleOps 400 Pro Indoor Cycle
(Saris Cycling Group). T2 + Profile Design Aero Bars
(Profile Design) were attached to the handlebars, y un
Logitech Trackball Mouse (Logitech) was fixed to the end
of the bars (Figura 1B). The addition of the aero bars served
two important purposes. Primero, the participant was able to
lean their elbows onto the bars leaving their hands free to
respond to the task. Segundo, the bars stabilized the partic-
ipant and helped reduce head and body movement, cual
is a critical factor for reducing noise during EEG recording.
To minimize discomfort, the bike saddle and handlebar po-
sitions were carefully adjusted for each participant. Heart
rate was tracked using a CycleOps wireless heart rate mon-
itor, whereas pedaling resistance and cadence were set and
recorded through Trainer Road software (Trainer Road).
EEG
EEG data were recorded using a Brain Products ActiCHamp
sistema (Brain Vision LLC) consisting of 64 active electrodes
arranged in an actiCAP elastic cap and placed in accordance
to the 10–20 System. The TP9 and TP10 electrodes were
adhered directly to the right and left mastoids. Connections
were established between electrodes and the scalp using
SuperVisc gel (Brain Products), which is especially viscous,
thus mitigating the potential for loss of signal because of
gel dispersion as well as the potential for electrodes
bridging because of increased sweating during exercise. En
the beginning of each investigation, all impedances were
reduced to below 15 kΩ. Data were sampled at 1000 Hz
and referenced off-line to the average mastoid signal.
Procedimiento
Participants were informed of the study’s structure and the
intensity at which they would be required to exercise.
They then completed a brief set of practice trials while
cycling to ensure that they were capable of completing
the task and to assess the stability of the eye-tracker.
Participants were also familiarized with the Rating of
Perceived Exertion (RPE) escala (Borg, 1970). RPE is a
subjective rating of the intensity of physical sensations
experienced during physical activity; the scale ranges from
6 (no exertion) a 20 (maximal exertion).
Prior to mounting the stationary bike, the wireless heart
rate monitor and EEG cap were placed on the participant.
Once on the bike, the seat position was carefully adjusted
to maximize participant comfort. When ready, Participantes
Garrett, Bullock, and Giesbrecht
1273
initiated the spatial change detection task. Each trial began
with the fixation dot in the center of the screen, junto con
a green dot (subtending 0.4° visual angle) representing the
location of the participant’s gaze. The participant aligned
their gaze dot with the fixation dot and pressed the mouse
button with their right thumb to start the trial. The fixation
dot immediately turned gray to indicate that the trial was
underway. The sample stimulus was then presented for
250 mseg (with onset jittered randomly between 600
y 1500 msec after trial initiation). Stimulus offset was
followed by a 1750-msec retention interval, where the fix-
ation cross exclusively remained on screen. During the fix-
ación, stimulus presentation, and retention periods,
participants were instructed to maintain their gaze at the
center of the screen and covertly shift attention to the po-
sition of the sample stimulus and remember its location. Si
gaze position deviated from fixation >2.4° or eye-blinks
occurred during these periods, the trial was aborted and
the message “Broken Fixation!” appeared on the screen.
Aborted trials were appended to the end of the trial se-
quence, to ensure that a complete set of trials free from
blinks and other eye movements was obtained. En el
end of the retention period, a test stimulus (identical in
size and color to the sample stimulus) appeared, either
in the same location as the sample stimulus (50% of trials)
or in a location shifted 20° clockwise or anticlockwise from
the sample location (50% of trials). Participants were re-
quired to indicate whether the test stimulus appeared at
the same location or different location as the sample stim-
ulus by pressing either the left or right mouse button,
respectivamente.
Participants completed this task in both rest and exer-
cise conditions (counterbalanced 640 trials per exercise
condición; 10 blocks of 64 ensayos) while seated on a sta-
tionary bike. In the exercise condition, they engaged in
low-intensity cycling with a resistance of 50 watts of power
and at a pedaling cadence of 50 RPM. These resistance
and cadence levels were based on the intensity and ca-
dence used in a previous study from our lab (Bullock
et al., 2017). To ensure that participants maintained this
cadence, they were instructed to pedal in time to a met-
ronome set at 100 beats per minute (equaling 50 RPM).
Cadence was continuously monitored throughout exer-
cise. In the resting condition, the pedals were removed
and replaced with a box positioned under each foot.
Using these boxes, participants tapped their feet to a
metronome set at the same frequency as described in
the exercise condition, totaling 50 taps per foot per min-
ute (equivalent to cycling cadence of 50 RPM). This manip-
ulation was intended to attenuate possible dual-task
differences between conditions that may confound mod-
ulations in WM. To mitigate any possible exercise-induced
arousal carryover effects, participants who completed
the exercise condition prior to the rest condition were
required to sit quietly until their heart rate returned to
dentro 10% of resting activity before beginning the resting
condición.
Excluding warm-up and cool-down time, each condition
took ∼50 min to complete. The warm-up consisted of ei-
ther cycling with the aforementioned cadence/resistance
or foot tapping for 3 mín.. Prior to and following warm-
arriba, the RPE scale was displayed to the participant, OMS
then verbally reported their current level of exertion to
the experimenter. Exertion ratings were also collected
after every two blocks. Each experimental session took
∼3.5 hr, including instrumentation time.
Biases in Eye Position
Considering that the tolerance threshold for deviations
from fixation is more lenient than what is typical for stud-
ies of visual WM and attention (es decir., > 1°; Luck, 2014), es
possible that gaze position toward stimulus location may
differ between rest and exercise conditions. Such an effect
may be the source of differences in spatial selectivity be-
tween rest and exercise. To rule out this possibility, ojo
position bias was quantified by calculating the distance be-
tween fixation and stimulus location for each trial using the
eye-tracking data. Distances were baseline corrected to
the mean of the 200-msec prestimulus period. Porque
distance was computed relative to the stimulus location,
more negative values would represent greater deviations
toward the stimulus location. For ease of interpretation,
the absolute value of the average baselined gaze position
was computed and plotted in Figure 10.
EEG Preprocessing
Custom scripts in MATLAB ( Version 2019a, El
MathWorks Inc.) and functions from the EEGLAB toolbox
(Delorme & Makeig, 2004) were used for off-line process-
ing of the EEG data. The continuous data were referenced
to the average mastoid signal and then high- and low-pass
filtered between 4 y 30 Hz, respectivamente (EEGLAB func-
tion pop_eegfiltnew). The data were then resampled at
250 Hz (EEGLAB function pop_resample), to reduce com-
putation time and memory demands, and epoched be-
tween −100 and 2500 sec around the onset of the
stimulus. Trials that were aborted because of eye move-
ments and trials where incorrect responses were made
were excluded from any analysis. Noisy electrodes were re-
moved via visual inspection (mean electrodes removed
[mean ± SEM] = 1 ± 0.39). Electrodes that were excluded
from one condition were also excluded from the other
condition for each participant in order to avoid introduc-
ing bias when comparing EEG results across conditions.
Trials exceeding ± 150 μV in remaining electrodes were
then excluded (mean trials excluded overall: 4.99 ±
0.79; descansar: 4.71 ± 1.28, exercise: 5.26 ± 0.95). For comput-
ing the degree of alpha lateralization (see Alpha
Lateralization section below) in topographical patterns
of activity at the scalp, noisy electrodes were interpolated
to facilitate averaging across participants.
1274
Revista de neurociencia cognitiva
Volumen 33, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Spectral Decomposition
Epoched data were filtered using a third-order Butterworth
bandpass filter (MATLAB function butter) entre 8 y
12 Hz. A Hilbert transformation (MATLAB function hilbert)
was then applied to the filtered signal in order to extract a
measure of instantaneous amplitude and phase. To avoid
edge artifacts, all subsequent EEG analyses were then
focused on time points between −500 and 2000 mseg
(de 500 msec prestimulus onset to the end of the reten-
tion period). Prior to modeling, total power was calculated
as the square of the absolute value of the Hilbert trans-
formed complex values. Total power reflects continuous
oscillatory activity independent of its phase relationship
with stimulus onset.
Alpha Lateralization
Numerous studies have reported alpha power to be greatest
over posterior electrodes ipsilateral to the cued location
when compared to contralateral electrodes—indicating that
spatial attention/memory alters the topographical distri-
bution of alpha (MacLean et al., 2019; Kelly, Lalor, Reilly,
& Foxe, 2006; Thut et al., 2006; Sauseng et al., 2005;
Worden, Foxe, Wang, & Simpson, 2000). Además, exer-
cise has been shown to modulate power across parietal-
occipital electrode sites for a range of frequencies (Ciria
et al., 2019; Ciria, Perakakis, Luque-Casado, & Sanabria,
2018). De este modo, the degree to which exercise influenced the
systematic changes in alpha power topography was deter-
mined by normalizing (es decir., dividing) the difference in alpha
power at contralateral and ipsilateral parietal/occipital
electrodes sites (P5/6, P7/8, PO7/8) by the sum of power at
contralateral and ipsilateral sites. Normalized alpha power at
contralateral and ipsilateral sites was then averaged by
condition for the stimulus (0–250 msec) and retention
(500–2000 msec) time periods.
P1 ERP
Possible differences in spatial selectivity between condi-
tions may be driven by modulations in the early visual
evoked response. Considering this, the impact of exercise
on the P1 ERP component was examined. Raw EEG data
were first referenced to the average mastoid signal, y
then high/low-pass filtered at 1 y 30 Hz, respectivamente.
Nota, this high-pass filter was applied to minimize the
amount of sweat and movement related artifacts (p.ej., cy-
cling cadence was ∼0.83 Hz). Afterward the data were
epoched again between −500 and 2500 mseg (de 500 mseg
prestimulus onset to the end of the retention period).
Trials exceeding ± 150 μV measured at scalp electrodes
de interés (P1/2, P3/4, P5/6, PO7/O8, POz/Oz, O1/2) eran
excluded. Three participants retained fewer than half of
their trials after applying this rejection criterion, de este modo
they were excluded only from subsequent ERP analyses.
Nota, we did not exclude these participants in IEM
analyses because they did not yield the same amount
of artifact-rejected trials when using a more aggressive
high-pass filter (4 Hz), and the focus of the IEM is on WM
processes rather than sensory-evoked activity. Artifact-free
ensayos (mean overall: 494.47 ± 14.91; descansar: 514.10 ± 76.46; ex-
ercise: 474.84 ± 88.12) were baseline corrected between
−100 and 0 mseg.
IEM
Spatially selective neural population “channel” response
functions/profiles (CRFs) were estimated based on the dis-
tribution patterns of total alpha power across the scalp
(Foster et al., 2016). Primero, the model was trained to estimate
the extent to which the linear combination of a priori canon-
ical channel responses (es decir., set of basis functions) captura
the underlying structure of the observed data (topographi-
cal distribution of induced alpha power), yielding a set of
regression weights. Próximo, these weights were tested on ob-
served data that were excluded during training in order to
estimate the channel response. The parameters of these
channel response estimates were then used to quantify
the spatially selective response. This method has been
successfully used to reconstruct feature- y ubicación-
selective responses from human fMRI data (Ester, sprague,
& serencias, 2015; Brouwer & Heeger, 2009, 2011, 2013;
serencias & Saproo, 2012; Naselaris, kay, Nishimoto, &
Gallant, 2011) and EEG recorded at the scalp (MacLean
et al., 2019; Sutterer et al., 2019; Bullock et al., 2017;
Foster et al., 2016; Samaha et al., 2016; Garcia et al., 2013).
The IEM was computed for each participant separately
using total alpha power. Within each location bin, ensayos
were then randomly subdivided into three samples.
Note that because trial-based artifact rejection can result
in an uneven number of trials per condition, it was neces-
sary to ensure that any comparisons between conditions
were not influenced by unequal trial counts. Before enter-
ing the data into the IEM, the minimum number of trials
per location bin (norte) was calculated across both conditions
for each participant. To ensure equal numbers of trials
from each location bin were entered into the model, n−1
trials were randomly selected from each bin. After trials
were randomly subdivided into samples, these samples
were then averaged. De este modo, each condition included
24 samples of averaged trials (8 location bins × 3 muestras
of averaged trials). To ensure the outcome of the model
and subsequent analyses were not influenced by an idio-
syncratic selection of trials, this process was repeated
10 veces, with a randomized selection of trials entered into
the IEM for each of the iterations. For each iteration, un
independent IEM was computed for each time point
over the course of the trial (250-Hz EEG sampling rate ×
2.5 sec = 625 time points) to model the temporal dynamics
of the location-selective response.
For each iteration (and time point), the independent
IEMs were cross-validated using a k-fold scheme, dónde
k = 4. The averaged trials were randomly grouped into
Garrett, Bullock, and Giesbrecht
1275
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
four folds, with each fold having one averaged trial per
location bin. Training was performed using 3/4 folds.
En tono rimbombante, the IEM was trained on equivalent numbers
of trials from both rest and exercise conditions to estimate
a fixed encoding model. This training scheme mitigates
the possibility that differences in spatial selectivity
between rest and exercise are merely a reflection of differ-
ences in the signal-to-noise ratio between conditions
(jardinero & Liu, 2019; sprague, Boynton, & serencias,
2019; Liu, Cable, & jardinero, 2018; Sprague et al., 2018).
For each participant and each of the 10 iterations, IEMs
were computed using the following algorithm. Let m
represent the number of EEG electrodes in each data set
(mean electrodes = 63 ± 0.38; equal across rest and exer-
cise conditions within each participant), n1 represents the
number of trials in the training set (three folds of eight
averaged trials), and n2 represents the number of trials
in the testing set (one fold of eight averaged trials). Let j
be the number of hypothetical location-selective channels
(C1, j × n1), composed of half-sinusoidal functions raised
to the seventh power as the basis set. Aquí, the basis set
was composed of 8 equally spaced locations (es decir. j = 8). B1
(m × n1) represents the training set and B2 (m × n2) el
test set. A standard implementation of the general linear
model was then used to estimate the weight matrix (W.,
m × j ) using the basis set (C1). More specifically, usando
the general linear model:
B1 ¼ WC1
(1)
Entonces, the ordinary least-squares estimate of W can be
computed as:
(cid:3)−1
^W ¼ B1CT
(cid:1)
1 C1CT
(2)
1
Using the estimated weight matrix ( ^W, Ecuación 2) y
the test data (B2), the channel responses C2 ( j × n2) poder
be estimated by:
(cid:4)
(cid:5)
^C2 ¼ ^W
T ^W
− ^W
t
B2
(3)
After ^C2 was solved for each location bin, the CRF on each
average trial was then circularly shifted to a common
stimulus-centered reference frame (degrees of offset from
channel’s designated location bin), and the centered
response functions were averaged across channels. El
model was then repeated for each time point. The final
centered CRF was computed by averaging over the 10
iterations at each time point.
IEM Generalization
To examine the temporal generalization of patterns of
activity underlying spatially selective responses, IEMs were
trained at each point in time, and then tested on every
other point in time (ensuring independence of training
and test sets). To reduce computation time (y el
number of statistical comparisons), the data were down
sampled to 25 Hz prior to training and testing.
Quantifying Spatially Selective Representations
Estimated channel responses were folded around 0° channel
offset and transformed from (−135°, −90°, −45°, 0°, 45°, 90°,
135°, 180°) en (0°, 45°, 90°, 135°, 180°) by averaging the
response at corresponding offsets (± 45°, 90°, and 135°; 0°
and 180° were not averaged) for quantification. Slope was
then computed (MATLAB function polyfit) as the linear
regression weight of total alpha power across offset, y
served as an index for the amount of spatial selectivity in pat-
terns of alpha activity underlying channel responses. Larger
slope values indicate greater spatial selectivity.
Además, the IEM procedure was carried out with
randomly permuted location bin labels for 250 iterations.
In theory, this should generate flat channel response pro-
files devoid of spatial information. Slopes of these corre-
sponding channel responses (es decir., permuted slopes) eran
then calculated for each iteration, which served as our null
distribution for the statistical analyses of “real” slope values.
Hypothesis Testing
All statistical inference relied on computing Bayes factors
(BFs) using functions from the BayesFactor toolbox for R
(Morey, Rounder, & Jamil, 2015), which employs a Cauchy
previo. A BF between 1 y 3 indicates “anecdotal” evidence
for the alternative hypothesis, entre 3 y 10 indicates
“moderate” evidence, entre 10 y 30 indicates
“strong” evidence, and greater than 30 indicates “very
strong” evidence (Kruschke & Liddell, 2018; Dienes,
2016; Wetzels et al., 2011; Kass & Raftery, 1995). BFs <
1, on the other hand, indicate varying degrees of evidence
in favor of the null hypothesis (0.33–1 = anecdotal,
0.1–0.33 = moderate, 0.033–0.1 = strong, 0.01–0.033 =
very strong, < 0.01 = extreme). To determine if there
was evidence indicating nonzero slopes, which would be
expected if the patterns of alpha contained any spatial
information, one-sample BF t tests were computed using
the real location labels from each trial at each time point
(“real” BFs). To test for evidence indicating differences
between rest and exercise conditions, paired BF t tests
were used instead. The one-sample and paired-samples
BF t tests were conducted for each iteration and time point
using the permuted location labels, to generate a per-
muted distribution of BFs (“permuted” BFs) for subse-
quent nonparametric comparisons.
Bayesian inference is more conservative than frequen-
tist inference, and much less likely to result in false confi-
dence (Gelman & Tuerlinckx, 2000). Despite this, the
number of statistical tests conducted overall may still be
of concern, such that it may be possible to observe large
BFs by chance alone. Thus, a cluster-based correction
procedure (Cohen, 2014) was performed to protect against
spuriously large BFs. For each iteration of permuted slopes,
we calculated the maximum cluster size of contiguous time
points where BF ≥ 3, resulting in a null distribution of
maximum cluster sizes. For both real and permuted BFs,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
o
c
n
_
a
_
0
1
7
1
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
1276
Journal of Cognitive Neuroscience
Volume 33, Number 7
only time points displaying at least moderate evidence
(BF ≥ 3) in favor of the alternative hypothesis were consid-
ered for cluster-based correction. Then, the size of the real
BF clusters was calculated and compared to the null distribu-
tion of cluster sizes. If a cluster was larger than 95% of the
null distribution of maximum cluster sizes, it was considered
to be unlikely due to chance alone.
The above statistical routine was also applied to the gen-
eralization matrix. To reduce computation time, the per-
mutation procedure for the generalization analysis was
computed for 100 iterations rather than 250 iterations.
RESULTS
Exercise Physiology
Average heart rate (Table 2) was greater during exercise
(105.78 ± 3.06) as compared to rest (78.44 ± 2.66; BF >
1,000). A similar relationship was observed for mean RPE
puntuaciones (BF > 1,000): descansar (6.59 ± 0.11), exercise (8.65 ±
0.25). Notablemente, the average RPE score for the exercise
condition was within the range of 9–10 on the RPE scale,
which corresponds to “very light” intensity exercise. En
addition, normalized pupil area was larger in the exercise
condition ~1560 to 2000 msec post stimulus onset (BF 2
[3−14], es decir., BF ranged between 3 y 14).
Comportamiento
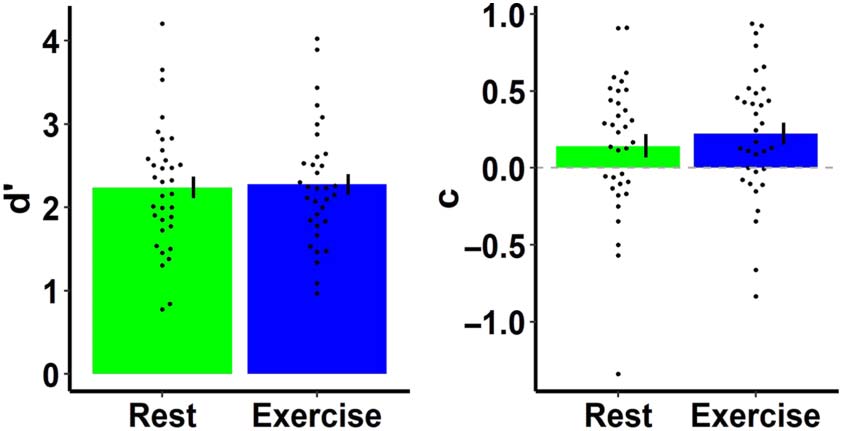
0) y
Task performance was measured using sensitivity (d
response criterion (C) (Cifra 2) from signal detection the-
ory (see Swets, 1961, para una revisión). Hits were defined as ac-
curately detecting a difference between the location of
sample and test stimuli. There was evidence in favor of the
null hypothesis for no difference between rest and exercise
0 (descansar: 2.23 ± 0.13, exercise:
conditions in the measures d
2.27 ± 0.12; novio = 0.2) and c (descansar: 0.15 ± 0.08, exercise:
0.23 ± 0.07; novio = 0.56).
Topographical Distribution of Alpha Power across
the Scalp
Prior to modeling the contribution of alpha activity to WM
representaciones, the topographical distribution of power
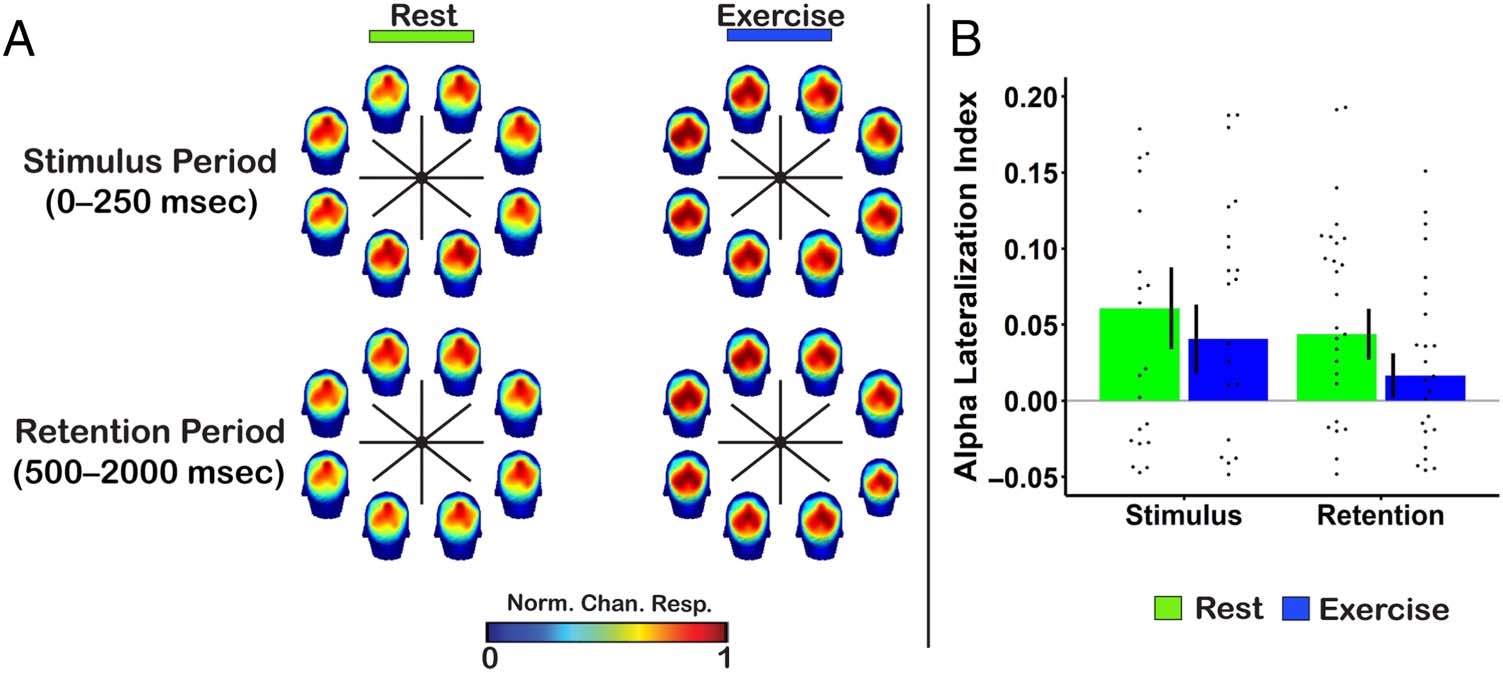
across the scalp was examined. Cualitativamente, alpha power
increased over posterior electrodes ipsilateral to the pre-
sented stimulus location in the resting condition, a finding
consistent with prior work (Figura 3A; MacLean et al.,
Mesa 2. Means and Standard Errors for the Physiological and
Subjective Report Data
Cifra 2. Working Memory task performance as measured by sensitivity
0 ) and response criterion (C) in both conditions. Points represent each
(d
individual’s data, and error bars represent ±1 SEM.
2019; Kelly y col., 2006; Thut et al., 2006; Sauseng et al.,
2005; Worden et al., 2000). More importantly, a similar pat-
tern was present in the exercise condition. When analyzing
the degree of alpha lateralization, there was moderate
evidence in favor of the null hypothesis for no difference
between conditions during both the stimulus (0–250 msec;
descansar: 0.06 ± 0.03, exercise: 0.04 ± 0.02; BF ¼ 0:22) y
retention (500–2000 msec; descansar: 0.04 ± 0.02, exercise:
0.02 ± 0.01; BF ¼ 0:37) periods (Figura 3B).
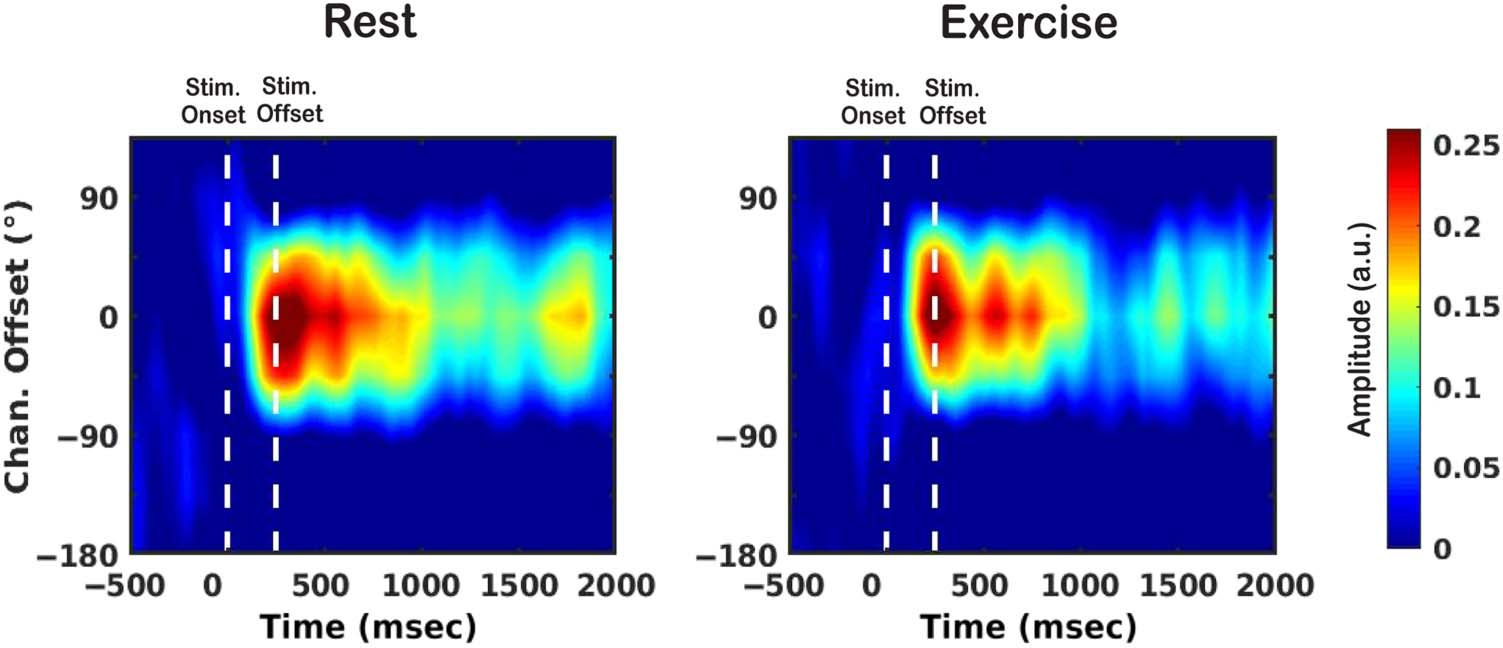
Reconstructing Representations of Stimulus Location
The IEM analysis using alpha band activity yielded evi-
dence for spatially selective responses in both conditions
(Cifra 4). Reconstructed response profiles reached peak
amplitude ∼200 msec after stimulus onset, which is consis-
tent with previous studies using the IEM approach to track
locations maintained in WM (MacLean et al., 2019; Foster
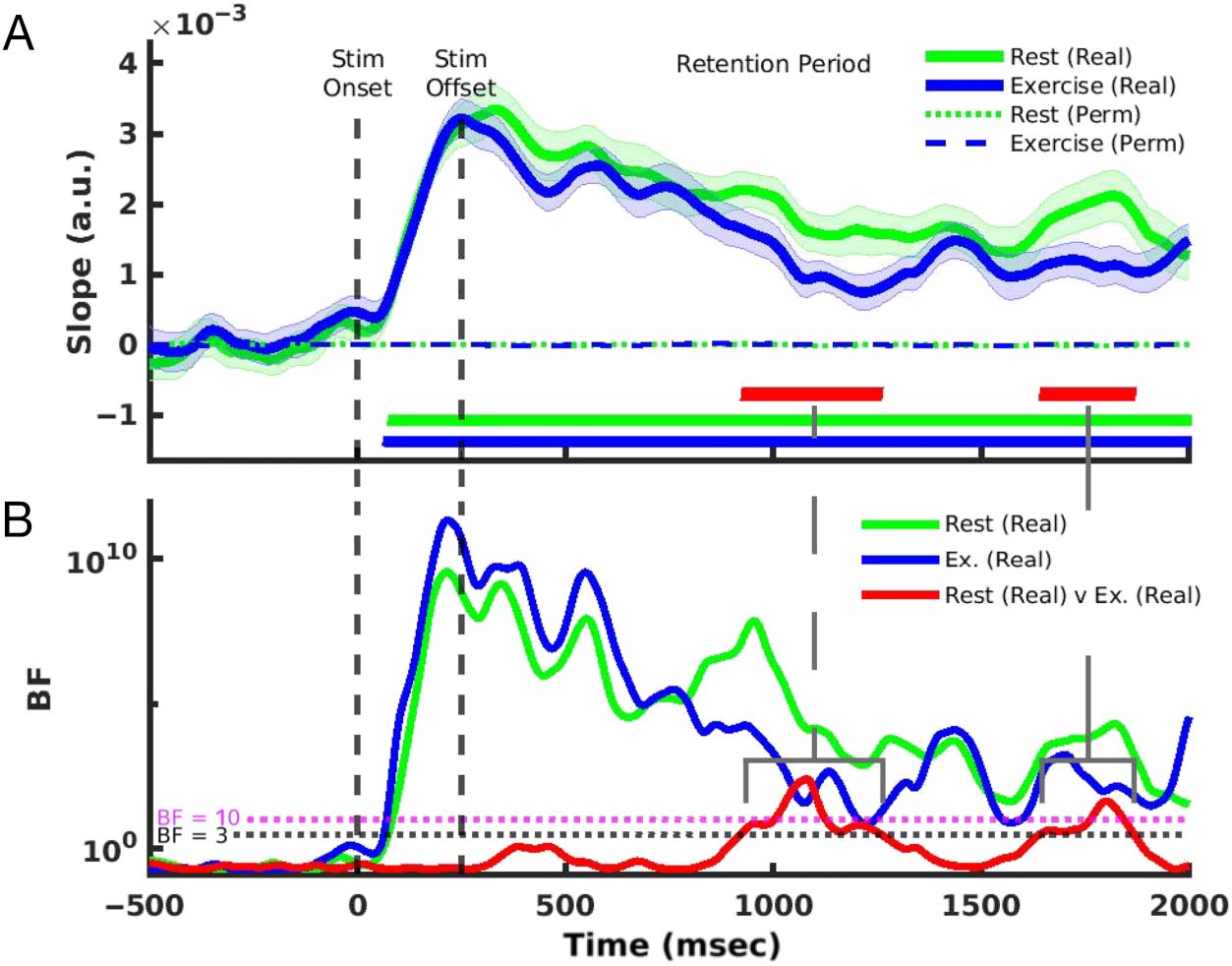
et al., 2016). Comparing real slopes to their permuted null
distributions revealed there was at least moderate evi-
dence for differences throughout the encoding and reten-
tion periods in both conditions (Figure 5A; descansar: BF 2 [3,
3.55 × 109], exercise: BF 2 [3, 2.23 × 1011]), confirming
that mental representations of remembered locations can
be successfully reconstructed both at rest and during exer-
cise. Sin embargo, there was evidence for decreases in
slope between ∼926–1255 msec (BF 2 [3, 238]) y
∼1643–1864 msec (BF 2 [3, 41]) post stimulus onset during
exercise compared to rest. These results suggest that,
although there was evidence for a location-selective repre-
sentation in alpha activity during exercise, this represen-
tation was degraded during late stages of retention
(Figure 5B).
Exercise Physiology
Exercise Modulates Processes Underlying
Spatial Selectivity
Condition Heart Rate (BPM)
RPE
Cadence (RPM)
Rest
78.44 ± 2.66
6.59 ± 0.11
–
Exercise
105.78 ± 306
8.65 ± 0.25
54.77 ± 0.65
Models trained on patterns of activity within a specific time
period that can recover spatially selective responses when
tested on activity from a differing time period exhibit gen-
eralización (MacLean et al., 2019; van Moorselaar et al.,
Garrett, Bullock, and Giesbrecht
1277
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 3. (A) Topographical
distribution of alpha (8–12 Hz)
power across the scalp during
encoding and retention
periods, normalized across
parietal/parieto-occipital
electrodes within each location
bin. The location of each head
plot reflects the corresponding
sample stimulus location
bin. Nota, a time frame of
500–2000 msec was used for
the retention period to avoid
including stimulus evoked
actividad. (B) Alpha lateralization
during the stimulus and
retention periods as a function
of exercise condition. Points
represent each individual’s data,
and error bars represent ±1 SEM.
2018; Rey & Dehaene, 2014). Successful generalization
suggests that a stable unitary code underlies a cognitive
process or processes throughout time. A diferencia de, failure
of models to generalize in this fashion implies that the
trained/tested patterns of activity represent different
codes. Nota, a lack of generalization is not an indication
that neural activity during that time period does not sup-
port representations of location in WM, given that recon-
struction was successful from ∼100 msec onwards when
training and testing within time points.
When testing the fixed encoding model on activity in the
resting condition, there was evidence for temporal gener-
alization throughout the entire trial period poststimulus
onset (Cifra 6 “Test: Rest”). Strong generalization
throughout time indicates that spatial selectivity at rest is
supported by a stable unitary code. There was also evi-
dence for temporal generalization during exercise, pero
was degraded relative to a permuted null distribution
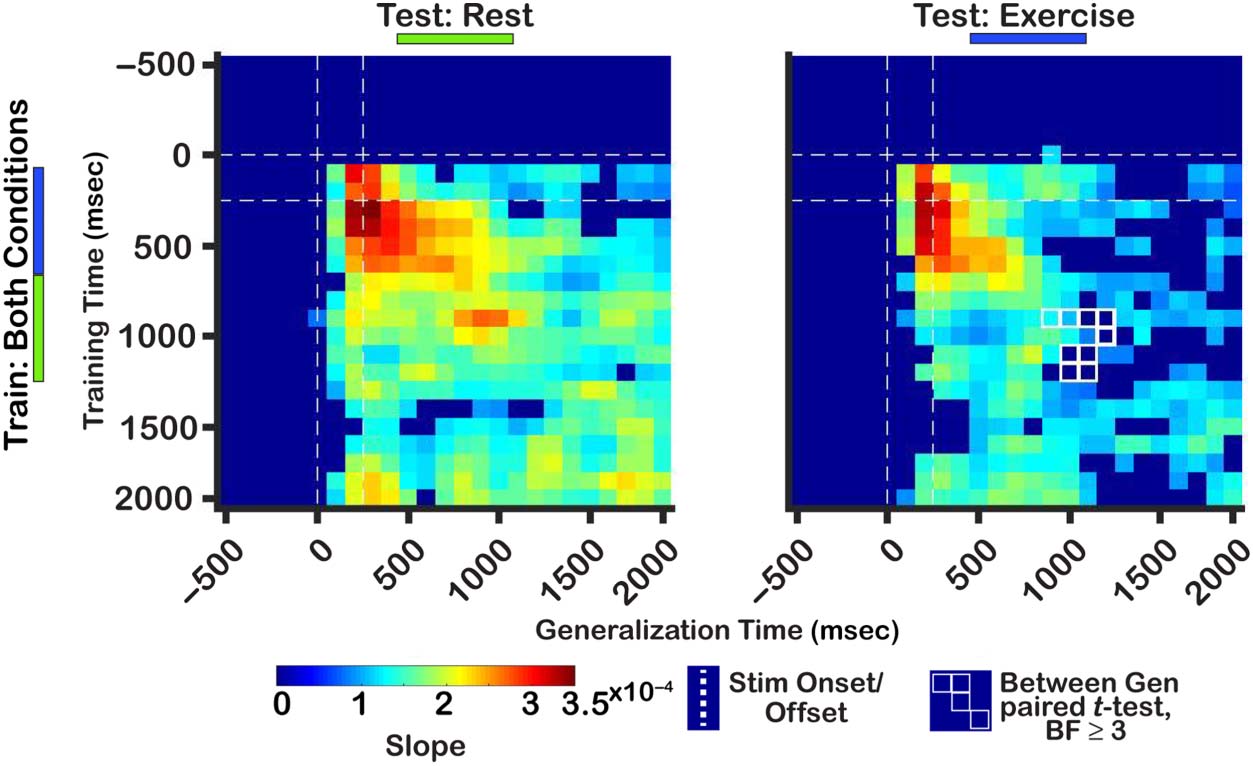
(Cifra 6 “Test: Exercise”). Comparing the degree of gen-
eralization between conditions, there was evidence for
greater generalization when training on activity between
∼900 and 1200 msec and testing on activity within the
same time period in the resting condition (BF 2 [3.89,
148.12]). Tomados juntos, these results imply that similar
unitary codes underlie the representation of locations in
WM during both rest and exercise, but that the stability
of this code over time is degraded during exercise.
Control Analyses
Differences in Reconstruction Are Not Caused by
Differences in Spectral Activity
A series of control analyses were performed on spectral
activity to determine any confounding factors in our
observed differences of selectivity between conditions.
One such analysis was the comparison of total spectral
power between the rest and exercise conditions. Spectral
power across a range of frequency bands has been shown
to be modulated during exercise (p.ej., Bullock et al., 2017;
Kubitz & Mott, 1996). De este modo, it is possible that the observed
differences in spatial selectivity and generalization may
Cifra 4. Estimated channel
responses as a function of
stimulus-centered location
offset (−180°, −135°, −90°,
−45°, 0°, 45°, 90°, 135°),
reconstructed over time from
patterns of alpha-band activity.
Plotted CRFs are baseline
corrected relative to the average
amplitude across channels in a
prestimulus window of −500
a 0 mseg.
1278
Revista de neurociencia cognitiva
Volumen 33, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 5. (A) Spatially selective
CRF slopes. Horizontal green
and blue bars indicate time
points with at least moderate
evidence for real rest and
exercise slope estimates
being different from zero,
respectivamente. Horizontal red bars
indicate time points with at
least moderate evidence for a
difference between real rest
and exercise slopes. Cluster
correction was applied to all
comparisons, and clusters
shown are those whose size
exceeded 95% of a permuted
null distribution. (B) BFs for
comparisons made at each
time point. Shaded error bars
represent ± 1 SEM.
reflect exercised-induced fluctuations in power that are
independent of WM processes. EEG data were high-pass
filtered at 4 Hz, and spectral power was computed for post-
stimulus neural activity (0–2000 msec) using a fast Fourier
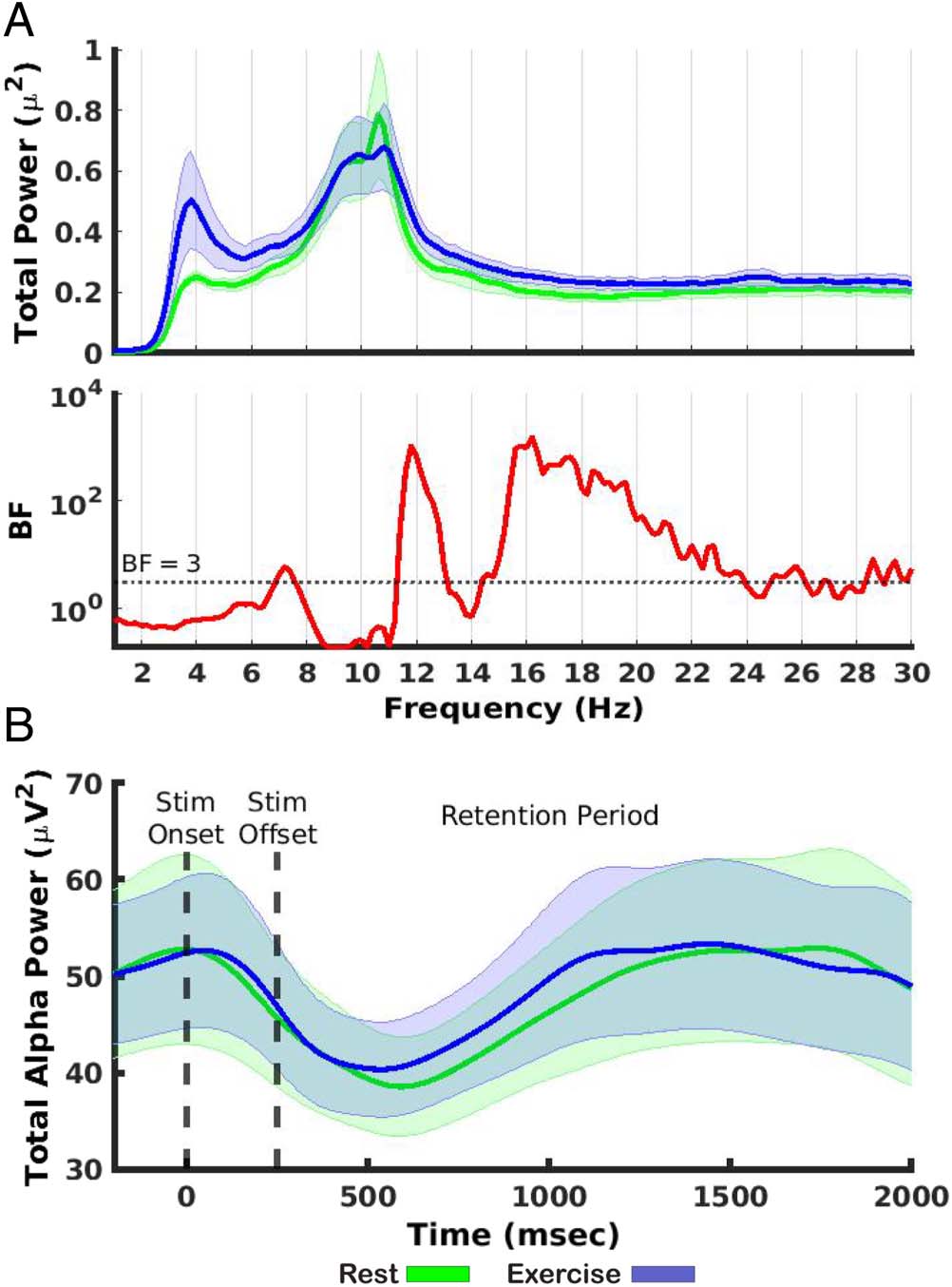
transform (MATLAB function fft). There was at least moder-
ate evidence for a difference in power between the follow-
ing frequency ranges: 7–7.4 Hz (BF 2 [4.11, 5.81]), 11.4–
13 Hz (BF 2 [5.85, 1.04 × 103]), and ∼14–30 Hz (BF 2
3.12, 1.51 × 103; Figure 7A).
Considering there was evidence for a difference be-
tween conditions in the upper range of alpha total power
averaged over the trial period (∼11 to 12 Hz), it is possible
that fluctuations in alpha power over time may be driving
decreased selectivity in the exercise condition. To assess
this possibility, the time-course of mean total alpha power
was compared between both conditions. There was no ev-
idence for differences; if anything, there was evidence in
favor of the null hypothesis (BF 2 [0.18, 0.71]; Figure 7B).
En tono rimbombante, this suggests that decreased spatial selectivity
during exercise is a product of alterations in the topo-
graphical distribution of alpha power over time rather than
dramatic changes in global alpha power.
Degraded Reconstructions Are Not Due to Location
Information Being Carried by Other Frequencies
during Exercise
Previous research has shown that topographical patterns
of oscillatory activity outside the alpha frequency range
do not track the contents of spatial WM (Foster et al.,
Cifra 6. IEM generalizations.
Only contiguous time points
with at least moderate evidence
for slope estimates being
different from zero (grupo
corregido) are shown. All other
time points are in dark blue.
The y-axis represents time
points trained on, mientras que el
x-axis is time points tested on.
Successful generalization to
the right of the main diagonal
indicates forward temporal
generalización, mientras que el
opposite direction indicates
backward temporal
generalización.
Garrett, Bullock, and Giesbrecht
1279
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 7. Control analyses on
spectral activity conducted
to assess the validity of IEM
findings. (A) Top: Total power
averaged across time for all
frequency bands between
4 y 30 Hz. Peak response
occurred over the alpha band
range. Bottom: BF values
comparing power at each
frequency between rest and
exercise. (B) Alpha power over
tiempo. Dashed lines indicate
stimulus onset (0 mseg) y
stimulus offset (250 mseg).
Shaded error bars represent
± 1 SEM.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
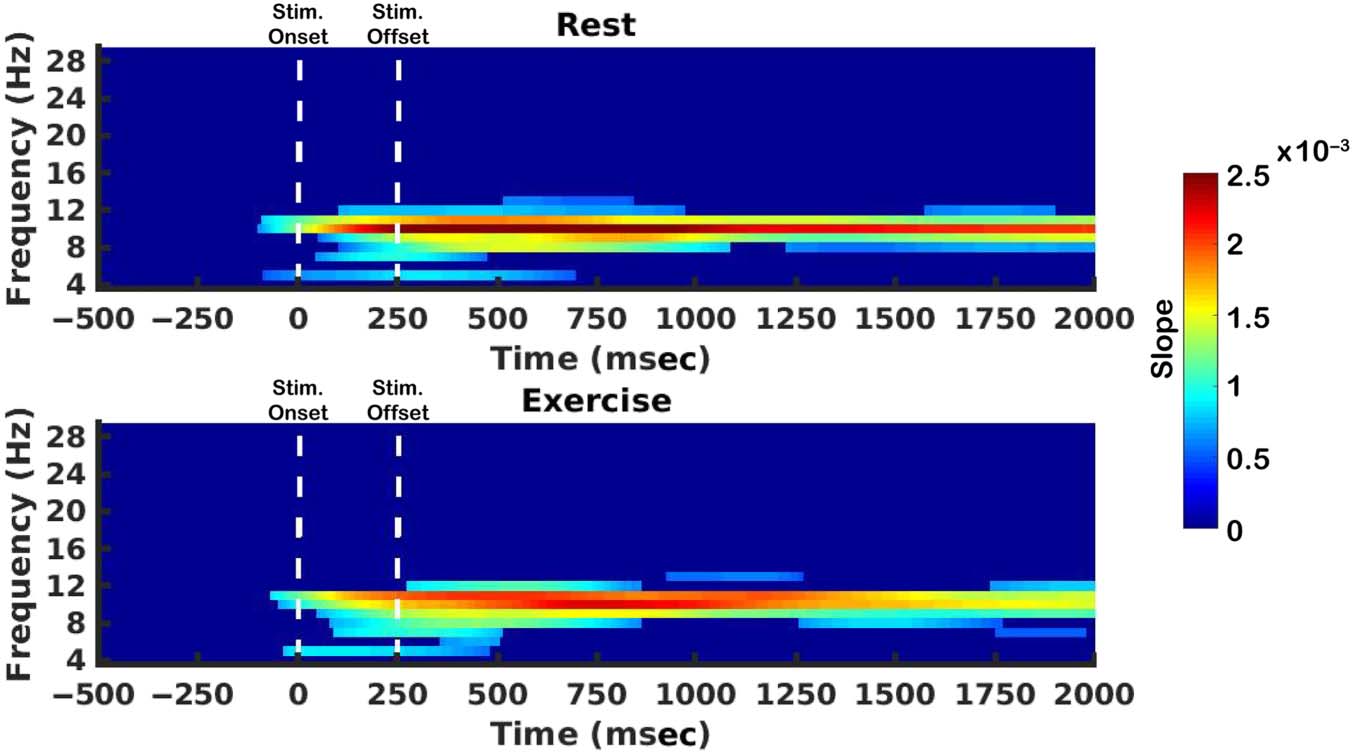
2016). Todavía, it is possible that these frequency bands may be
recruited during a bout of exercise. This would imply that
decreases in slope amplitude in the exercise condition re-
flect a possible change between frequency bands that sup-
port representations. Considering this, the IEM routine
was applied to total power across a broad range of frequen-
cíes (4–30 Hz in 1-Hz increments; Cifra 8). When testing
for nonzero slopes in the computed single frequency
CRFs, at least moderate evidence in favor of the alternative
hypothesis was only observed for activity within the alpha
frequency band range for both rest (BF 2 [3, 1.86 × 106])
and exercise (BF 2 [3, 2.15 × 107]). Evidence in favor of
differences in single frequency slopes between conditions
did not survive cluster-based corrections. De este modo, decreases
in alpha slope amplitude in the retention period during
exercise (Figure 5A) do not reflect the recruitment of
Cifra 8. The IEM routine was
applied to frequencies within
the range of 4–30 Hz at 1-Hz
increments. Shown here are the
slopes of reconstructed CRFs
with at least moderate evidence
for being different from zero
(cluster corrected). Those that
were not different are colored
in dark blue.
1280
Revista de neurociencia cognitiva
Volumen 33, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
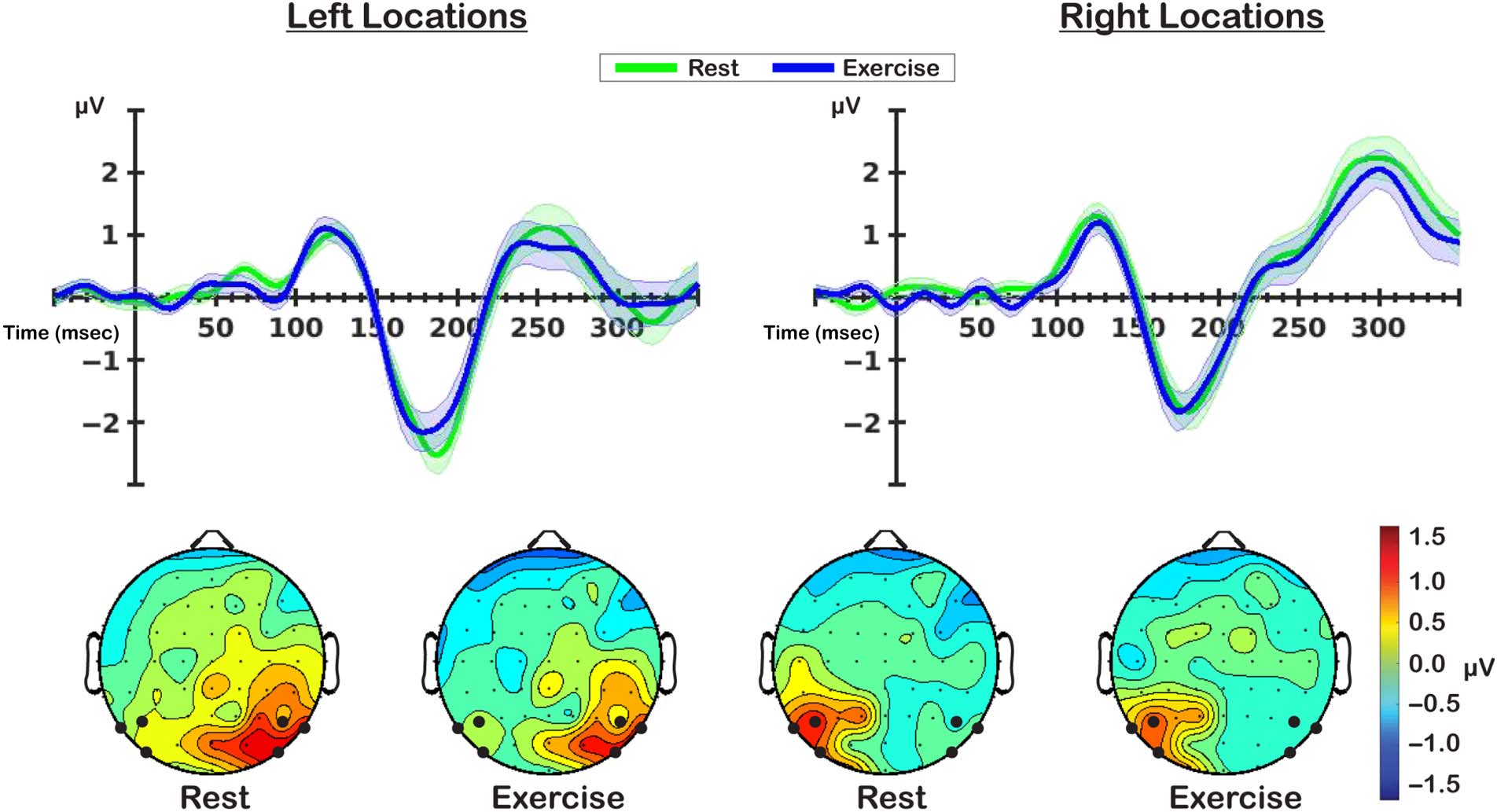
Cifra 9. P1 analyses. Top plots depict P1 components for memoranda presented at left and right locations in the display. Bottom plots show
topographical distribution of mean P1 amplitude between 116 y 136 mseg. Electrodes colored in black represent the ones used for computation of
lateralized difference ERPs.
non-alpha frequency bands to support representations of
location specific information.
Visual Evoked Responses
Para cada ubicación, we calculated the difference in activity
between parieto-occipital electrodes (PO7/O8, P5/6, P7/8)
ipsilateral to the presented stimulus locations and those
that were contralateral. Entonces, difference waves were aver-
aged for locations on either side of the vertical meridian,
yielding two P1 components: one for left locations and the
other for right locations (Cifra 9). P1 mean amplitude was
calculated by finding the peak latency of the positive going
component between 100 y 150 msec post stimulus onset,
and subsequently calculating mean amplitude ±10 msec
around this peak. There was moderate evidence in favor
of the null hypothesis of no difference between rest and
exercise P1 mean amplitudes for left (descansar: 1.19 ± 0.13,
exercise: 1.3 ± 0.15; novio = 0.26) and right locations (descansar:
1.49 ± 0.17, exercise: 1.44 ± 0.15; novio = 0.19). Estos resultados
suggest that the observed differences in spatial selectivity
are not due to modulations of the visual evoked response.
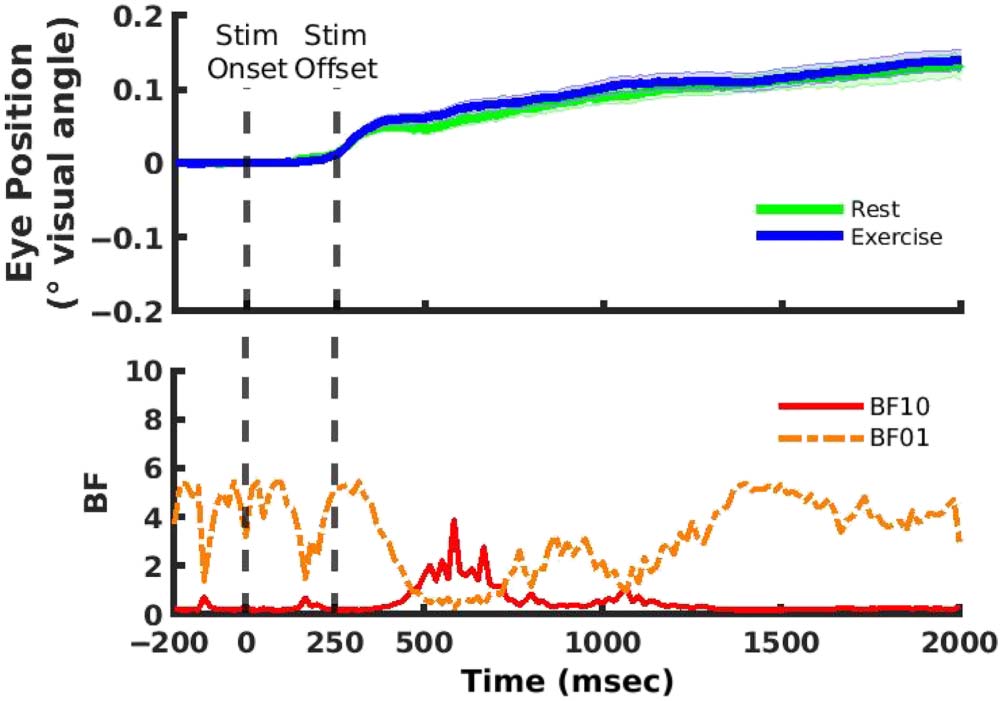
Cifra 10. Top: Eye position
inclinación (es decir., baseline corrected
distance of gaze from stimulus
location in units of degrees of
visual angle) for both the rest
and exercise conditions. Bottom:
BF values for comparisons
between rest and exercise. Uno
time point at 538 msec showing
at least moderate evidence in
favor of the alternative
hypothesis did not survive
cluster correction. Shaded error
bars represent ± 1 SEM.
Garrett, Bullock, and Giesbrecht
1281
Eye Position
To assess whether the difference in the spatially selective
responses derived from alpha band activity is contaminated
by differential eye movements toward remembered loca-
tions in the rest and exercise conditions, we compared
eye position throughout the stimulus and retention pe-
riods. There was a small increase in eye position bias
throughout the trial period in both conditions, but this bias
toward the stimulus location did not exceed 0.15° in either
condición. Además, point-wise comparisons did not
reveal evidence for differences that survived cluster correc-
ción (Cifra 10). If the cluster correction was not applied,
there was a single time point that indicated moderate evi-
dencia (novio = 3.86) for a difference between rest and exer-
cise conditions at ∼538 msec poststimulus onset, pero esto
difference was small (0.02°), occurred during a period when
the slopes of the spatial reconstructions were declining, y
did not occur at the same time point when evidence for dif-
ferences between the slope of the spatial reconstructions in
exercise and rest were observed. A diferencia de, había
moderate evidence in favor of the null hypothesis of no dif-
ference in eye position bias between rest and exercise at
multiple time points throughout the trial period (BF 2 [3,
5.44]). Por lo tanto, eye position biases were likely not the
cause of decreased spatial selectivity during exercise.
DISCUSIÓN
Evidence from human, nonhuman animal, and inverte-
brate studies has demonstrated that early sensory visual
processes are modulated during bouts of acute physical
actividad (Kaneko et al., 2017; Bullock et al., 2015, 2017;
Fu et al., 2014; Ayaz et al., 2013; Pontifex & Hillman,
2007). The goal of this study was to determine whether
higher-order cognitive operations that depend on these
sensory responses are also impacted during physical activ-
idad. EEG was recorded from human participants while they
engaged in a spatial change detection task at rest and dur-
ing a bout of cycling exercise. The IEM technique was then
applied to activity in the alpha band in order to reconstruct
spatially selective response profiles for item locations
stored in WM. There were two key results. Primero, in addition
to replicating previous work demonstrating that topo-
graphically specific patterns of alpha band activity track
the contents of WM at rest (MacLean et al., 2019; Foster
et al., 2016), the present results also demonstrate that it
is possible to reconstruct spatially selective response pro-
files during item encoding and retention while partici-
pants are engaged in a bout of physical activity. Segundo,
although there was evidence for a spatially selective
response during exercise, our results also indicate that
the quality of this reconstructed location information is
degraded relative to the rest condition, but only during
the late stages of the retention period.
The finding that topographic patterns of total alpha
activity track the specific position of a behaviorally relevant
stimulus both during encoding and retention replicates a
number of studies in the literature indicating that the neu-
ral populations that give rise to alpha oscillations in human
EEG code information in WM in a location-selective man-
ner (Sutterer, Polyn, & Woodman, 2021; MacLean et al.,
2019; Sutterer et al., 2019; Foster et al., 2016). The results
reported here also show that these location-selective
codes can be reconstructed from patterns of neural activity
acquired during exercise. críticamente, the slopes of the
location-selective profiles were greater than zero through-
out the trial, indicating that the location-selective WM rep-
resentations coded in alpha band activity are robust to
changes in global behavioral state.
Despite the robust location-selective reconstructions
during encoding and retention observed here, había
also clear evidence during the late stages of retention
(> 900 mseg) that the selectivity of these reconstructions
was degraded during exercise relative to rest. One possible
explanation for the degraded reconstructions of spatially
selective response profiles is that WM representations
coded in the alpha band are more fragile later in the reten-
tion period and vulnerable to decay during exercise.
Another possible explanation, that is not mutually exclu-
sive from the previous explanation, is that the rest and ex-
ercise conditions may differ in their attentional demands.
The similar level of behavioral performance in the two
tasks is inconsistent with this interpretation; sin embargo,
the behavioral task was very easy and may thus not have
been sensitive to more subtle differences in attentional de-
mands between the two conditions. Although we took
measures to equate the conditions by requiring partici-
pants to cycle to the beat of a metronome in the exercise
condition and to tap their feet to the metronome in the
rest condition, it is still possible that cycling to the beat
of a metronome requires greater attentional control than
foot tapping and interferes with attention-based rehearsal
of locations in WM (Postle, Awh, En Jonia, Herrero, &
D'Esposito, 2010; Awh et al., 1999, Awh & En Jonia, 1998).
En tono rimbombante, previous work has shown that shifts in atten-
tion from memoranda in WM toward an opposing task
disrupt spatial selectivity (van Moorselaar et al., 2018).
The notion that cycling can impact resource allocation in
a concurrent cognitive task is supported by previous work
demonstrating modulation of various sensory and cognitive
ERP components (Bullock et al., 2015; Pontifex & Hillman,
2007; Grego et al., 2004; Yagi, Coburn, Estes, & Arruda,
1999). Future work that manipulates levels of dual-task
interference between conditions (p.ej., higher levels
during rest) and employs a more complex WM task is
necessary to elucidate the cause of decreased spatial selec-
tivity during exercise.
Given that topographic patterns of alpha activity also
track with attended locations in spatial attention tasks
(Samaha et al., 2016; Rihs et al., 2007; Thut et al., 2006;
Sauseng et al., 2005), it is reasonable to question whether
the location-selective response profiles observed here at
rest and during exercise represent WM activity or covert
1282
Revista de neurociencia cognitiva
Volumen 33, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
oh
C
norte
_
a
_
0
1
7
1
4
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
spatial attention to the location of the memoranda. Allá
is some evidence against a solely attention-based interpre-
tation of the present results. Por ejemplo, in covert atten-
tion tasks that have revealed spatially specific responses to
attended locations, alpha power tends to ramp-up in am-
plitude prior to the attended stimulus over ipsilateral sites
(Banerjee, Snyder, Molholm, & Foxe, 2011; Rihs et al.,
2007). Aquí, sin embargo, after the initial reduction in alpha
power driven by the evoked response, total alpha was rel-
atively constant up to the presentation of the test stimulus.
Además, the presence of sustained delay period activ-
ity in the absence of a stimulus is a classic indicator of WM
(Sreenivasan & D'Esposito, 2019; Vogel, McCollough, &
Machizawa, 2005; Vogel & Machizawa, 2004). Though
the quality of reconstructions degraded over time, el
slopes of profiles were greater than zero throughout the
entire trial period. A diferencia de, the profiles of reconstruc-
tions observed in spatial attention tasks increase during
periods prior to target onset (Samaha et al., 2016). It is im-
portant to note that we are not arguing that spatial atten-
tion is not involved, rather we are arguing that the pattern
of results is unlikely to be driven by covert attention alone.
Además, when considering the present findings together
with those in the literature providing evidence for the
strong connection between spatial WM and spatial atten-
ción, our results are consistent with the notion that spatial
attention facilitates the coding and maintenance of spatial
representations held in WM (Oberauer, 2019; Postle et al.,
2010; Awh et al., 1999, Awh & En Jonia, 1998).
Previous studies have reported evidence for exercise-
induced enhancements of visual processing (Bullock
et al., 2015, 2017). Aquí, sin embargo, there was no difference
in P1 mean amplitude or location selectivity between the
two conditions during this period, indicating that the initial
sensory coding was similar in the two conditions. Estos
contrasting findings between our studies may be because
of the considerable differences in visual stimulation, tarea
demandas, or the nature of the data submitted to an
IEM. Por ejemplo, in Bullock et al. (2015), Participantes
performed an oddball task with large stimuli presented at
fixation. Bullock et al. (2017) required participants to judge
orientation changes occurring in large, high contrast, cen-
trally presented flickering grating stimuli. Además,
they estimated orientation-selective stimulus reconstruc-
tions using 15-Hz steady-state stimulus evoked activity.
Aquí, participants were required to remember the location
of a small, gray item presented in the periphery and
location-selective reconstructions are based on induced
alpha band activity. The discrepancy between the results
reported here and the exercise-induced enhancements
observed in previous work from our laboratory (Bullock
et al., 2015, 2017) and by others (Pesce, Capranica,
Tessitore, & Figura, 2003) may also reflect a difference in
the effect of global physiological states on sensory evoked
responses to attended stimuli (es decir., as in the previous
trabajar) and in activity that persists well beyond the sensory
evoked response, such as WM retention-related activity.
Patterns of activity coding for spatial locations displayed
robust generalization throughout time when testing on ac-
tivity in the resting condition, indicating the presence of a
stable unitary code. This pattern of generalization is consis-
tent with previous research showing the presence of a rapid
selection process that supports locations held in WM when
external visual input is continuous (MacLean et al., 2019).
Although generalization when testing on activity in the ex-
ercise condition was not as robust relative to the resting
condición, a stable unitary code was also found to support
spatial selectivity when participants are in a physically active
estado. En tono rimbombante, the successful reconstruction of spatially
selective responses when applying the fixed IEM to activity
throughout time from each condition separately suggests a
common unitary code underlies the maintenance of loca-
tions in WM in both physiological states. In later stages of
the retention period, this unitary code is degraded when
one is concurrently engaged in exercise.
As with all simultaneous neuroimaging and exercise stud-
es, EMG and sweat artifacts are potentially confounding fac-
tors in our results. The contribution of EMG artifacts was
minimized by stabilizing each participant’s position on the
bike using aero bars, and coaching them to ensure they lim-
ited upper body movement during cycling or foot tapping.
EMG typically occurs at higher frequencies (> 30 Hz),
whereas sweat artifacts are low frequency (< 1 Hz) oscilla-
tions (Thompson, Steffert, Ros, Leach, & Gruzelier, 2008).
Because the IEM analyses presented here were based on
alpha power (8–12 Hz), it is unlikely that our results were
seriously contaminated by these artifacts.
In summary, we used the IEM technique to investigate
how spatial selectivity for locations held in WM is modulated
during an acute bout of aerobic exercise when compared
to rest. Reconstruction of location-selective representations
was successful during both rest and exercise, but stimulus
representations were degraded at specific time points dur-
ing the retention period as a function of exercise. Evidence
also suggested that WM representations were supported by
a unitary code during both rest and exercise. The current
study is the first to demonstrate that representations of
items stored in WM can be reconstructed during a bout of
physical exercise and provides novel insight into both the
modulation and composition of these representations dur-
ing exercise when compared to rest. Future research will
focus on uncovering how exercise-induced degradation of
stimulus representations in WM impacts behavior in the
human.
Reprint requests should be sent to Jordan Garrett, Psychological
& Brain Sciences, University of California Santa Barbara, 93106-
9010, or via e-mail: jordan.garrett@psych.ucsb.edu.
Funding Information
This work was supported by the Institute for Collaborative
Biotechnologies through cooperative agreement W911NF-
Garrett, Bullock, and Giesbrecht
1283
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
o
c
n
_
a
_
0
1
7
1
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
19-2-0026 with the U.S. Army Research Office. The content
of the information does not necessarily reflect the position
or policy of the government and no official endorsement
should be inferred.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the pro-
portions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/W = .108, and W/W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/W = .102, and W/W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
REFERENCES
Ayaz, A., Saleem, A. B., Schölvinck, M. L., & Carandini, M.
(2013). Locomotion controls spatial integration in mouse
visual cortex. Current Biology, 23, 890–894. DOI: https://
doi.org/10.1016/j.cub.2013.04.012, PMID: 23664971,
PMCID: PMC3661981
Awh, E. & Jonides, J. (1998). Spatial working memory and spatial
selective attention. In R. Parasuraman (Ed.), The attentive
brain (pp. 353–380). Cambridge, MA: MIT Press.
Awh, E., Jonides, J., Smith, E. E., Buxton, R. B., Frank, L. R.,
Love, T., et al., (1999). Rehearsal in spatial working memory:
Evidence from neuroimaging. Psychological Science, 10,
433–437. DOI: https://doi.org/10.1111/1467-9280.00182
Banerjee, S., Snyder, A. C., Molholm, S., & Foxe, J. J. (2011).
Oscillatory alpha-band mechanisms and the deployment
of spatial attention to anticipated auditory and visual target
locations: Supramodal or sensory-specific control mechanisms?
Journal of Neuroscience, 31, 9923–9932. DOI: https://doi.org
/10.1523/JNEUROSCI.4660-10.2011, PMID: 21734284, PMCID:
PMC3343376
Borg, G. (1970). Perceived exertion as an indicator of somatic
stress. Scandinavian Journal of Rehabilitation Medicine, 2,
92–98. DOI: https://doi.org/10.1037/t58166-000, PMID:
5523831
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision,
10, 433–436. DOI: https://doi.org/10.1163/156856897X00357,
PMID: 9176952
Brouwer, G. J., & Heeger, D. J. (2009). Decoding and reconstructing
color from responses in human visual cortex. Journal of
Neuroscience, 29, 13992–14003. DOI: https://doi.org/10
.1523/JNEUROSCI.3577-09.2009, PMID: 19890009, PMCID:
PMC2799419
Brouwer, G. J., & Heeger, D. J. (2011). Cross-orientation
suppression in human visual cortex. Journal of Neurophysiology,
106, 2108–2119. DOI: https://doi.org/10.1152/jn.00540.2011,
PMID: 21775720, PMCID: PMC3214101
Brouwer, G. J., & Heeger, D. J. (2013). Categorical clustering of
the neural representation of color. Journal of Neuroscience,
33, 15454–15465. DOI: https://doi.org/10.1523/JNEUROSCI
.2472-13.2013, PMID: 24068814, PMCID: PMC3782623
Bullock, T., Cecotti, H., & Giesbrecht, B. (2015). Multiple
stages of information processing are modulated during
acute bouts of exercise. Neuroscience, 307, 138–150. DOI:
https://doi.org/10.1016/j.neuroscience.2015.08.046, PMID:
26318337
Bullock, T., Elliott, J. C., Serences, J. T., & Giesbrecht, B. (2017).
Acute exercise modulates feature-selective responses in human
cortex. Journal of Cognitive Neuroscience, 29, 605–618. DOI:
https://doi.org/10.1162/jocn_a_01082, PMID: 27897672
Cao, L., & Händel, B. (2019). Walking enhances peripheral
visual processing in humans. PLoS Biology, 17, e3000511.
DOI: https://doi.org/10.1371/journal.pbio.3000511, PMID:
31603894, PMCID: PMC6808500
Cheron, G., Petit, G., Cheron, J., Leroy, A., Cebolla, A., Cevallos,
C., et al. (2016). Brain oscillations in sport: Toward EEG
biomarkers of performance. Frontiers in Psychology, 7, 246.
DOI: https://doi.org/10.3389/fpsyg.2016.00246
Ciria, L. F., Luque-Casado, A., Sanabria, D., Holgado, D., Ivanov,
P. C., & Perakakis, P. (2019). Oscillatory brain activity during
acute exercise: Tonic and transient neural response to an
oddball task. Psychophysiology, 56, e13326. DOI: https://doi.org
/10.1111/psyp.13326, PMID: 30637763, PMCID: PMC7311047
Ciria, L. F., Perakakis, P., Luque-Casado, A., & Sanabria, D.
(2018). Physical exercise increases overall brain oscillatory
activity but does not influence inhibitory control in young
adults. Neuroimage, 181, 203–210. DOI: https://doi.org/10
.1016/j.neuroimage.2018.07.009, PMID: 29981904
Cohen, M. X. (2014). Analyzing neural time series data: theory
and practice. Cambridge, MA: MIT Press. DOI: https://doi.org
/10.7551/mitpress/9609.001.0001
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://www.sccn.ucsd.edu/eeglab/.
DOI: https://doi.org/10.1016/j.jneumeth.2003.10.009, PMID:
15102499
Dienes, Z. (2016). How Bayes factors change scientific practice.
Journal of Mathematical Psychology, 72, 78–89. DOI:
https://doi.org/10.1016/j.jmp.2015.10.003
Ester, E. F., Sprague, T. C., & Serences, J. T. (2015). Parietal
and frontal cortex encode stimulus-specific mnemonic
representations during visual working memory. Neuron, 87,
893–905. DOI: https://doi.org/10.1016/j.neuron.2015.07.013,
PMID: 26257053, PMCID: PMC4545683
Foster, J. J., Sutterer, D. W., Serences, J. T., Vogel, E. K., & Awh, E.
(2016). The topography of alpha-band activity tracks the content
of spatial working memory. Journal of Neurophysiology, 115,
168–177. DOI: https://doi.org/10.1152/jn.00860.2015, PMID:
26467522, PMCID: PMC4760461
Fu, Y., Tucciarone, J. M., Espinosa, J. S., Sheng, N., Darcy, D. P.,
Nicoll, R. A., et al. (2014). A cortical circuit for gain control by
behavioral state. Cell, 156, 1139–1152. DOI: https://doi.org
/10.1016/j.cell.2014.01.050, PMID: 24630718, PMCID:
PMC4041382
Garcia, J. O., Srinivasan, R., & Serences, J. T. (2013). Near-real-
time feature-selective modulations in human cortex. Current
Biology, 23, 515–522. DOI: https://doi.org/10.1016/j.cub.2013
.02.013, PMID: 23477721, PMCID: PMC3608396
Gardner, J. L., & Liu, T. (2019). Inverted encoding models
reconstruct an arbitrary model response, not the stimulus.
eNeuro, 6. DOI: https://doi.org/10.1523/ENEURO.0363-18
.2019, PMID: 30923743, PMCID: PMC6437661
Gelman, A., & Tuerlinckx, F. (2000). Type S error rates classical
and Bayesian single and multiple comparison procedures.
Computational Statistics, 15, 373–390. DOI: https://doi.org
/10.1007/s001800000040
1284
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
o
c
n
_
a
_
0
1
7
1
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Grego, F., Vallier, J. M., Collardeau, M., Bermon, S., Ferrari, P.,
Candito, M., et al. (2004). Effects of long duration exercise on
cognitive function, blood glucose, and counterregulatory
hormones in male cyclists. Neuroscience Letters, 364, 76–80.
DOI: https://doi.org/10.1016/j.neulet.2004.03.085, PMID:
15196681
Morey, R. D., Rouder, J. N., & Jamil, T. (2015). Package
‘BayesFactor’ (R Package Version 0.9.12-2).
Naselaris, T., Kay, K. N., Nishimoto, S., & Gallant, J. L. (2011).
Encoding and decoding in fMRI. Neuroimage, 56, 400–410.
DOI: https://doi.org/10.1016/j.neuroimage.2010.07.073,
PMID: 20691790, PMCID: PMC3037423
Kaneko, M., Fu, Y., & Stryker, M. P. (2017). Locomotion induces
Niell, C. M., & Stryker, M. P. (2010). Modulation of visual responses
stimulus-specific response enhancement in adult visual
cortex. Journal of Neuroscience, 37, 3532–3543. DOI:
https://doi.org/10.1523/JNEUROSCI.3760-16.2017, PMID:
28258167, PMCID: PMC5373133
by behavioral state in mouse visual cortex. Neuron, 65,
472–479. DOI: https://doi.org/10.1016/j.neuron.2010.01.033,
PMID: 20188652, PMCID: PMC3184003
Oberauer, K. (2019). Working memory and attention—A
Kass, R. E., & Raftery, A. E. (1995). Bayes factors. Journal of the
American Statistical Association, 90, 773–795. DOI: https://
doi.org/10.1080/01621459.1995.10476572
conceptual analysis and review. Journal of Cognition, 2, 36.
DOI: https://doi.org/10.5334/joc.58, PMID: 31517246,
PMCID: PMC6688548
Keller, G. B., Bonhoeffer, T., & Hübener, M. (2012). Sensorimotor
Pesce, B. R., Capranica, L., Tessitore, A., & Figura, F. (2003).
mismatch signals in primary visual cortex of the behaving
mouse. Neuron, 74, 809–815. DOI: https://doi.org/10.1016/j
.neuron.2012.03.040, PMID: 22681686
Kelly, S. P., Lalor, E. C., Reilly, R. B., & Foxe, J. J. (2006). Increases in
alpha oscillatory power reflect an active retinotopic mechanism
for distracter suppression during sustained visuospatial
attention. Journal of Neurophysiology, 95, 3844–3851. DOI:
https://doi.org/10.1152/jn.01234.2005, PMID: 16571739
King, J. R., & Dehaene, S. (2014). Characterizing the dynamics
of mental representations: The temporal generalization
method. Trends in Cognitive Sciences, 18, 203–210. DOI:
https://doi.org/10.1016/j.tics.2014.01.002, PMID: 24593982,
PMCID: PMC5635958
Kruschke, J. K., & Liddell, T. M. (2018). The Bayesian new
statistics: Hypothesis testing, estimation, meta-analysis, and
power analysis from a Bayesian perspective. Psychonomic
Bulletin & Review, 25, 178–206. DOI: https://doi.org/10.3758
/s13423-016-1221-4, PMID: 28176294
Kubitz, K. A., & Mott, A. A. (1996). EEG power spectral densities
during and after cycle ergometer exercise. Research
Quarterly for Exercise and Sport, 67, 91–96. DOI: https://doi
.org/10.1080/02701367.1996.10607929, PMID: 8735998
Lambourne, K., Audiffren, M., & Tomporowski, P. D. (2010). Effects
of acute exercise on sensory and executive processing tasks.
Medicine and Science in Sports and Exercise, 42, 1396–1402.
DOI: https://doi.org/10.1249/MSS.0b013e3181cbee11, PMID:
20019631
Liu, T., Cable, D., & Gardner, J. L. (2018). Inverted encoding
models of human population response conflate noise and
neural tuning width. Journal of Neuroscience, 38, 398–408.
DOI: https://doi.org/10.1523/JNEUROSCI.2453-17.2017, PMID:
29167406, PMCID: PMC5761616
Luck, S. J. (2014). An introduction to the event-related potential
technique. Cambridge, MA: MIT Press.
MacLean, M. H., Bullock, T., & Giesbrecht, B. (2019). Dual
process coding of recalled locations in human oscillatory
brain activity. Journal of Neuroscience, 39, 6737–6750. DOI:
https://doi.org/10.1523/JNEUROSCI.0059-19.2019, PMID:
31300523, PMCID: PMC6703892
Maimon, G., Straw, A. D., & Dickinson, M. H. (2010). Active
flight increases the gain of visual motion processing in
Drosophila. Nature Neuroscience, 13, 393–399. DOI: https://
doi.org/10.1038/nn.2492, PMID: 20154683
Martins, Q. A., Kavussanu, M., Willoughby, A., & Ring, C. (2013).
Moderate intensity exercise facilitates working memory.
Psychology of Sport and Exercise, 14, 323–328. DOI: https://
doi.org/10.1016/J.PSYCHSPORT.2012.11.010
McMorris, T., Sproule, J., Turner, A., & Hale, B. J. (2011).
Acute, intermediate intensity exercise, and speed and
accuracy in working memory tasks: A meta-analytical
comparison of effects. Physiology and Behavior, 102,
421–428. DOI: https://doi.org/10.1016/j.physbeh.2010.12
.007, PMID: 21163278
Focusing of visual attention under submaximal physical load.
International Journal of Sport and Exercise Psychology,
1, 275–295. DOI: https://doi.org/10.1080/1612197X.2003
.9671719
Polack, P.-O., Friedman, J., & Golshani, P. (2013). Cellular
mechanisms of brain state-dependent gain modulation in
visual cortex. Nature Neuroscience, 16, 1331–1339. DOI:
https://doi.org/10.1038/nn.3464, PMID: 23872595, PMCID:
PMC3786578
Pontifex, M. B., & Hillman, C. H. (2007). Neuroelectric and
behavioral indices of interference control during acute
cycling. Clinical Neurophysiology, 118, 570–580. DOI:
https://doi.org/10.1016/j.clinph.2006.09.029, PMID: 17095295
Postle, B. R., Awh, E., Jonides, J., Smith, E. E., & D’Esposito, M.
(2010). The where and how of attention-based rehearsal
in spatial working memory. Cognitive Brain Research, 20,
194–205. DOI: https://doi.org/10.1016/j.cogbrainres.2004
.02.008, PMID: 15183391
Rihs, T. A., Michel, C. M., & Thut, G. (2007). Mechanisms of
selective inhibition in visual spatial attention are indexed
by α-band EEG synchronization. European Journal of
Neuroscience, 25, 603–610. DOI: https://doi.org/10.1111/j
.1460-9568.2007.05278.x, PMID: 17284203
Roig, M., Nordbrandt, S., Geertsen, S. S., & Nielsen, J. B. (2013).
The effects of cardiovascular exercise on human memory: A
review with meta-analysis. Neuroscience and Biobehavioral
Reviews, 37, 1645–1666. DOI: https://doi.org/10.1016/j
.neubiorev.2013.06.012, PMID: 23806438
Samaha, J., Sprague, T. C., & Postle, B. R. (2016). Decoding and
reconstructing the focus of spatial attention from the
topography of alpha-band oscillations. Journal of Cognitive
Neuroscience, 28, 1090–1097. DOI: https://doi.org/10.1162
/jocn_a_00955, PMID: 27003790, PMCID: PMC5074376
Sauseng, P., Klimesch, W., Stadler, W., Schabus, M.,
Doppelmayr, M., Hanslmayr, S., et al. (2005). A shift of visual
spatial attention is selectively associated with human EEG
alpha activity. European Journal of Neuroscience, 22,
2917–2926. DOI: https://doi.org/10.1111/j.1460-9568.2005
.04482.x, PMID: 16324126
Schaefer, S., Lövdén, M., Wieckhorst, B., & Lindenberger, U.
(2010). Cognitive performance is improved while walking:
Differences in cognitive–sensorimotor couplings between
children and young adults. European Journal of Developmental
Psychology, 7, 371–389. DOI: https://doi.org/10.1080
/17405620802535666
Serences, J. T., & Saproo, S. (2012). Computational advances
towards linking BOLD and behavior. Neuropsychologia, 50,
435–446. DOI: https://doi.org/10.1016/j.neuropsychologia
.2011.07.013, PMID: 21840553, PMCID: PMC3384549
Sprague, T. C., Adam, K. C. S., Foster, J. J., Rahmati, M.,
Sutterer, D. W., & Vo, V. A. (2018). Inverted encoding models
assay population-level stimulus representations, not single-
unit neural tuning. eNeuro, 5. DOI: https://doi.org/10.1523
Garrett, Bullock, and Giesbrecht
1285
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
o
c
n
_
a
_
0
1
7
1
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
/ENEURO.0098-18.2018, PMID: 29876523, PMCID:
PMC5987635
Sprague, T. C., Boynton, G. M., & Serences, J. T. (2019). The
importance of considering model choices when interpreting
results in computational neuroimaging. eNeuro, 6. DOI:
https://doi.org/10.1523/ENEURO.0196-19.2019, PMID:
31772033, PMCID: PMC6924997
Sprague, T. C., Saproo, S., & Serences, J. T. (2015). Visual
attention mitigates information loss in small- and large-scale
neural codes. Trends in Cognitive Sciences, 19, 215–226.
DOI: https://doi.org/10.1016/j.tics.2015.02.005, PMID:
25769502, PMCID: PMC4532299
Sreenivasan, K. K., & D’Esposito, M. (2019). The what, where
and how of delay activity. Nature Reviews Neuroscience, 20,
466–481. DOI: https://doi.org/10.1038/s41583-019-0176-7,
PMID: 31086326
Sutterer, D. W., Foster, J. J., Serences, J. T., Vogel, E. K., & Awh, E.
(2019). Alpha-band oscillations track the retrieval of precise
spatial representations from long-term memory. Journal of
Neurophysiology, 122, 539–551. DOI: https://doi.org/10.1152
/jn.00268.2019, PMID: 31188708, PMCID: PMC6734397
Sutterer, D. W., Polyn, S., & Woodman, G. F. (2021). Alpha-band
activity tracks a 2-dimensional spotlight of attention
during spatial working memory maintenance. Journal of
Neurophysiology, 125, 957–971. DOI: https://doi.org/10.1152
/jn.00582.2020, PMID: 33534657
Swets, J. A. (1961). Detection theory and psychophysics: A
review. Psychometrika, 26, 49–63. DOI: https://doi.org/10
.1007/BF02289684
Thompson, T., Steffert, T., Ros, T., Leach, J., & Gruzelier, J.
(2008). EEG applications for sport and performance.
Methods, 45, 279–288. DOI: https://doi.org/10.1016/j.ymeth
.2008.07.006, PMID: 18682293
Thut, G., Nietzel, A., Brandt, S. A., & Pascual-Leone, A. (2006).
α-Band electroencephalographic activity over occipital cortex
indexes visuospatial attention bias and predicts visual target
detection. Journal of Neuroscience, 26, 9494–9502. DOI:
https://doi.org/10.1523/JNEUROSCI.0875-06.2006, PMID:
16971533, PMCID: PMC6674607
van Moorselaar, D., Foster, J. J., Sutterer, D. W., Theeuwes, J.,
Olivers, C. N., & Awh, E. (2018). Spatially selective alpha
oscillations reveal moment-by-moment trade-offs between
working memory and attention. Journal of Cognitive
Neuroscience, 30, 256–266. DOI: https://doi.org/10.1162
/jocn_a_01198, PMID: 29040014, PMCID: PMC6022736
Vogel, E. K., & Machizawa, M. G. (2004). Neural activity predicts
individual differences in visual working memory capacity.
Nature, 428, 748–751. DOI: https://doi.org/10.1038
/nature02447, PMID: 15085132
Vogel, E. K., McCollough, A. W., & Machizawa, M. G. (2005).
Neural measures reveal individual differences in controlling
access to working memory. Nature, 438, 500–503. DOI:
https://doi.org/10.1038/nature04171, PMID: 16306992
Wetzels, R., Matzke, D., Lee, M. D., Rouder, J. N., Iverson, G. J.,
& Wagenmakers, E. J. (2011). Statistical evidence in
experimental psychology: An empirical comparison using 855
t tests. Perspectives on Psychological Science, 6, 291–298.
https://doi.org/10.117/1745691611406923. DOI: https://doi
.org/10.1177/1745691611406923, PMID: 26168519
Wilken, P., & Ma, W. J. (2004). A detection theory account of
change detection. Journal of Vision, 4, 1120–1135. DOI:
https://doi.org/10.1167/4.12.11, PMID: 15669916
Worden, M. S., Foxe, J. J., Wang, N., & Simpson, G. V. (2000).
Anticipatory biasing of visuospatial attention indexed by
retinotopically specific alpha-band electroencephalography
increases over occipital cortex. Journal of Neuroscience, 20,
RC63. DOI: https://doi.org/10.1523/jneurosci.20-06-j0002
.2000, PMID: 10704517, PMCID: PMC6772495
Yagi, Y., Coburn, K. L., Estes, K. M., & Arruda, J. E. (1999).
Effects of aerobic exercise and gender on visual and auditory
P300, reaction time, and accuracy. European Journal of
Applied Physiology and Occupational Physiology, 80,
402–408. DOI: https://doi.org/10.1007/s004210050611,
PMID: 10502073
Zhang, W., & Luck, S. J. (2008). Discrete fixed-resolution
representations in visual working memory. Nature, 453,
233–235. DOI: https://doi.org/10.1038/nature06860, PMID:
18385672, PMCID: PMC2588137
1286
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
7
1
1
9
2
1
2
9
7
/
/
j
o
c
n
_
a
_
0
1
7
1
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3