Creative Connections: Computational Semantic
Distance Captures Individual Creativity and
Resting-State Functional Connectivity
William Orwig1
, Ibai Diez1, Patrizia Vannini1,2, Roger Beaty3
, and Jorge Sepulcre1
Abstracto
■ Recent studies of creative cognition have revealed interac-
tions between functional brain networks involved in the gener-
ation of novel ideas; sin embargo, the neural basis of creativity is
highly complex and presents a great challenge in the field of
cognitive neuroscience, partly because of ambiguity around
how to assess creativity. We applied a novel computational
method of verbal creativity assessment—semantic distance—
and performed weighted degree functional connectivity analy-
ses to explore how individual differences in assembly of
resting-state networks are associated with this objective creativ-
ity assessment. To measure creative performance, a sample of
healthy adults (norte = 175) completed a battery of divergent think-
En g (DT) tareas, in which they were asked to think of unusual
uses for everyday objects. Computational semantic models were
applied to calculate the semantic distance between objects and
responses to obtain an objective measure of DT performance.
All participants underwent resting-state imaging, from which
we computed voxel-wise connectivity matrices between all gray
matter voxels. A linear regression analysis was applied between
DT and weighted degree of the connectivity matrices. Our anal-
ysis revealed a significant connectivity decrease in the visual-
temporal and parietal regions, in relation to increased levels of
DT. Link-level analyses showed higher local connectivity within
visual regions was associated with lower DT, whereas projec-
tions from the precuneus to the right inferior occipital and
temporal cortex were positively associated with DT. Nuestros resultados
demonstrate differential patterns of resting-state connectivity
associated with individual creative thinking ability, extending
past work using a new application to automatically assess crea-
tivity via semantic distance. ■
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
oh
C
norte
_
a
_
0
1
6
5
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INTRODUCCIÓN
Creative thinking is essential to all human progress and in-
innovación. Recent studies of functional neuroimaging and
network neuroscience have revealed interaction between
large-scale brain networks associated with creative cogni-
ción. The neuroscience of creativity seeks to disentangle
these complex brain processes that facilitate the generation
of novel ideas. Creativity has been defined as the produc-
tion of novel and useful ideas to solve problems (Guilford,
1967), and it is often assessed with tasks of divergent think-
En g (DT), which require the production of multiple solu-
tions to open-ended problems. A classic test of DT, el
Alternative Uses Task (AUT), prompts participants to gen-
erate alternative uses for a common object. To assess the
creative quality of ideas, traditional approaches to DT as-
sessment have largely relied on human raters, a subjective
and labor-intensive procedure. Recent developments in
creativity assessment have sought to standardize and
automate creativity assessment by applying computational
measures of semantic distance (Beaty & Johnson, 2020;
1Massachusetts General Hospital and Harvard Medical School,
2Brigham and Women’s Hospital and Harvard Medical School,
3Universidad Estatal de Pensilvania
© 2020 Instituto de Tecnología de Massachusetts
Dumas, Organisciak, & Doherty, 2020; Kenett & Faust,
2019; Heinen & Johnson, 2018; Prabhakaran, Verde, &
Gray, 2014). The application of semantic distance to creativ-
ity assessment is based on the associative theory of
creativity (Kenett & Faust, 2019; Mednick, 1962), cual
characterizes creative thought as a novel and useful recom-
bination of semantic knowledge; de este modo, the integration of
more semantically distant concepts is considered more
creative. This study applies semantic distance models to au-
tomatically and objectively assess DT. Past work has largely
focused on human ratings of creativity, so it is unknown
whether similar neural mechanisms underlie objective as-
sessments of creative thinking that do not rely on the sub-
jective judgments of human raters. Given high correlations
reported between human ratings and semantic distance
(Beaty & Johnson, 2020; Dumas et al., 2020), we expect
to find similar (though perhaps nonredundant) neural pat-
terns for human ratings and semantic distance measures of
DT. We measure semantic distance via a latent semantic dis-
tance factor—a combination of computational semantic
models that show a high correspondence to human ratings
of novelty and creativity (Beaty & Johnson, 2020).
Automated assessments of creativity via semantic distance
represents a new direction in the field of creativity research,
permitting an analysis of the extent to which neural
Revista de neurociencia cognitiva 33:3, páginas. 499–509
https://doi.org/10.1162/jocn_a_01658
correlates of creativity overlap when creativity is assessed
by humans versus machines.
Advances in neuroimaging techniques have enabled re-
searchers to study functional networks involved in creative
cognition. One approach that has been employed is func-
tional connectivity magnetic resonance imaging, cual
allows researchers to measure dynamic interactions be-
tween brain regions. Resting-state functional connectivity
magnetic resonance imaging studies have begun to disso-
ciate large-scale functional networks underlying cognitive
and attentional control processes relevant for creative cog-
nition (Beaty, Benedek, Silvia, & Schacter, 2016). Among
the most well studied of these functional networks, el
Default Mode Network (DMN)—composed of a set of
midline and posterior inferior parietal regions—shows
increased activation in the absence of an externally pre-
sented stimulus (Raichle et al., 2001). DMN activity is
associated with self-generated thought, such as mind
wandering and imagination (Andrews-Hanna, Smallwood,
& Spreng, 2014; Buckner, Andrews-Hanna, & Schacter,
2008). It has been suggested that activity in the DMN
contributes to the generation of candidate ideas, mientras
executive control networks exert top–down monitoring to
meet specific task goals or constraints (Beaty et al., 2016;
Beaty, Silvia, Nusbaum, Jauk, & Benedek, 2014). Task-based
approaches have helped to uncover the dynamic relation-
ship of these large-scale networks during creative task per-
rendimiento (Beaty, Kenett, et al., 2018; Shi et al., 2018). En
addition, resting-state studies have shown that temporal
variability of DMN connectivity correlates with DT and fre-
quency of transitions between functional connectivity-states
are associated with creative ability (Feng et al., 2019; Sol
et al., 2019; Liu et al., 2018; Gao et al., 2017; Le et al., 2017;
Takeuchi et al., 2012). Seed-based connectivity analyses in
resting-state fMRI data have revealed greater connectivity
between inferior pFC and DMN to be associated with idea
generación (Vartanian et al., 2018; Wei et al., 2014). El
objective of the present work was not necessarily to replicate
previous findings of the interaction between default mode
and executive control networks; we do not expect to find
consistent patterns of such network connectivity previously
reported in task-based studies. Bastante, we apply novel
semantic distance measures and data-driven graph theory
methods to resting-state data, exploring the overlap between
humano- and computationally derived creativity metrics.
Illuminating the specific contributions of DMN connec-
tivity to the production of novel ideas continues to be a
topic of great interest in the neuroscience of creativity
literature. Internal attention requires the disengagement
from immediate sensory information and may reflect a
more abstract cognition associated with creativity.
Internally directed cognition, which involves the shielding
of internal processes from external stimuli, has been associ-
ated with extended deactivation of occipital and parietal
regiones (Benedek et al., 2016). A recent series of neuroim-
aging studies have focused on the role of internally directed
attention in creativity and mental imagery (Fink & Benedek,
2019; Benedek, 2018; Fink et al., 2014). Attenuation of visual
input has also been associated with creative insight (Salvi
& Bowden, 2016; Salvi, Bricolo, Franconeri, Kounios, &
Beeman, 2015). Además, occipital and parietal regions,
such as the lateral occipital cortex and the inferior parie-
tal lobule, have been critically involved in the integration
of sensory perception toward multisensory and associa-
tion cortices (Diez et al., 2019; Sepulcro, 2014; Sepulcro,
Jabonoso, yo, Liu, & Johnson, 2012). Creative individuals
may exhibit a higher degree of internally directed cognition
at rest, reflected in lower connectivity within primary visual
systems and increased connectivity to areas of multimodal
integración.
Applications of graph theory in neuroscience have
emerged as an effective approach to studying functional
networks of the brain (van den heuvel & Hulshoff Pol,
2010). Network analysis offers insight into the structural
and functional organization of human brain networks
(despreciar, Chialvo, Kaiser, & Hilgetag, 2004), with brain
regions represented as nodes and the relationships
between nodes (es decir., conectividad) represented as edges
(Sepulcro, Jabonoso, & Goñi, 2014; bullmore & bassett,
2011; bullmore & despreciar, 2009). Individual variability in
functional network organization provides a lens to study
the relationship between brain morphology and cognitive
capacidad. Several network metrics have been developed to
quantify different properties of network architecture. Uno
such measure is the degree of a node, the number of edges
that connects it to the rest of the network (Sepulcre et al.,
2012; bullmore & bassett, 2011; Buckner et al., 2009).
Weighted degree ( WD) provides a measure of centrality,
quantifying the importance of each node within the
whole-brain architecture. Además, link-level analyses
characterize the strength of specific links between discrete
regiones del cerebro. Although graph theory approaches have
been employed in previous studies of creativity—for
ejemplo, using regions of interest applied to task-based
fMRI data and DT assessed with human creativity ratings
(Beaty, Benedek, Kaufman, & Silvia, 2015)—the present
work contributes new insight, applying a data-driven WD
and link-level connectivity analysis to explore resting-state
network architecture related to DT assessed objectively via
computational semantic distance. Brain graphs provide a
powerful model to study the complex organization of the
cerebro humano; application of graph theory metrics may
deepen our understanding of the network interactions,
which facilitate creative thinking.
Present Research
New approaches in network neuroscience have begun to
elucidate the cognitive mechanisms that underlie creative
pensamiento; sin embargo, more work is needed to disentangle the
complex network interactions involved in the production
of novel ideas and how these interactions relate to objec-
tive assessments of creativity. This study applies WD and
link-level analysis to represent individual differences in
500
Revista de neurociencia cognitiva
Volumen 33, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
oh
C
norte
_
a
_
0
1
6
5
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
resting-state networks associated with DT ability. To objec-
tively quantify individual’s DT performance, we leverage
latent variable modeling and multiple computational
models of semantic distance, extracting a latent factor of
semantic distance from five semantic models previously
shown to have a strong (but non-unity) correlation with
human judgments of relatedness, novelty, and creativity
(Beaty & Johnson, 2020). Semantic distance has been
employed in a task-based fMRI study using a noun–verb
generation task, finding that the semantic distance
between nouns and verbs tracked increases in frontopolar
cortex activity and connectivity (Verde, cohen, Raab,
Yedibalian, & Gray, 2015). We do not seek to replicate
these findings in our study, given differences in the tasks,
semantic distance calculations, and fMRI data; sin embargo,
this work provides preliminary evidence that semantic
distance captures individual variation in task-relevant
neural response. Hasta la fecha, it is unknown how such auto-
mated assessments relate to individual differences in the
brain’s intrinsic functional architecture (es decir., durante el
resting state) and whether similar neural correlates corre-
spond to human versus automated creativity assessment.
The present research thus aimed to extend research on
the neural basis of creative thinking by combining WD
analysis of resting-state fMRI data with both human and
automated creativity assessments.
MÉTODOS
The data were collected as part of a larger study on individ-
ual differences in creativity and imagination (Beaty, Kenett,
et al., 2018). The larger project included a task-based fMRI
study of DT; the task fMRI data from this study are not
analyzed here but have been published elsewhere (Frith
et al., 2020; Adnan, Beaty, Silvia, Spreng, & Tornero, 2019;
Beaty, Chen, et al., 2018). Aquí, we analyze resting-state
fMRI data from the full sample of the larger project. Nosotros
have previously published subsets of the resting-state
fMRI sample in studies of personality (Beaty, Chen, et al.,
2018) and human ratings on the AUT (Kenett, Betzel, &
Beaty, 2020), but no study has examined resting-state data
in relation to computational semantic distance. The total
sample consisted of 186 participants from the University
of North Carolina, Greensboro and members of the com-
munity; arte, música, and science majors were oversampled
to broaden the representation of creative backgrounds.
Desde 177 participants who completed a resting-state
scan, two were excluded because of distortion of the struc-
tural image. The final sample consisted of 175 Participantes
(127 women, edad media = 22.67 años, DE = 6.37 años).
All participants were right-handed with normal or
corrected-to-normal vision and reported no history of any
neurological disorders, cognitive disabilities, or medica-
tions that affect the central nervous system (Beaty, Chen,
et al., 2018). All participants provided written informed
consentir. The study was approved by the University of
North Carolina, Greensboro, institutional review board.
Behavioral Assessment
DT performance was assessed by the AUT, conducted
during a separate task-based fMRI scan, as well as on a com-
puter outside scanner. Note that the task-based fMRI data
are not presented here (only the verbal responses; ver
Beaty, Chen, et al., 2018). During the task-based fMRI scan,
participants were presented with a series of everyday ob-
jects (p.ej., brick) and asked to imagine new and unusual
uses for each object. Participants had 12 sec to think of a
single alternate use for a list of 23 objects and then had
5 sec to verbally report their response via an MRI-compatible
microphone (Benedek, Christensen, Fink, & Beaty, 2019;
Beaty, Chen, et al., 2018). For the computer-based assess-
mento, consistent with conventional procedures, participar-
pants had 3 min to generate as many alternative uses for
two objects as possible (box and rope). Además, partícipe-
ipants completed a test of visuospatial intelligence (Gv) a
measure the ability to mentally manipulate visual stimuli,
assessed via three independent tasks: paper-folding,
block-rotations, and cube comparisons (Frith et al., 2020).
The MRI- and lab-based DT responses were pooled and
scored for creative quality by 1) trained human raters using
the subjective scoring method (Silvia, 2008) y 2) compu-
tational models using semantic distance (Beaty & Johnson,
2020). Regarding human ratings, four trained raters inde-
pendently scored the creative quality of each response
using a 1 (not at all creative) a 5 (very creative) escala
(Benedek, Mühlmann, Jauk, & Neubauer, 2013; Silvia, 2008).
Raters were instructed to provide a single rating for each
respuesta, focusing on uncommonness, remoteness, y
cleverness; the scoring rubric and guidance for raters can be
found on Open Science Framework (https://osf.io/vie7s/).
Regarding semantic distance, we followed the approach
described in Beaty and Johnson (2020). We thus computed
the semantic distance between each object cue word (p.ej.,
box) and each response using an on-line application called
SemDis, an open platform developed to automate creativ-
ity assessment via semantic distance (semdis.wlu.psu.edu).
SemDis leverages five compositional vector models to
compute the relatedness between inputted texts: tres
continuous bag of words (CBOW) predict models and
two count models. CBOW/predict models were built using
a neural network architecture (Mandera, Keuleers, &
Brysbaert, 2017) that employs a sliding window to move
through text corpora and aims to predict a central word
from surrounding context words (cf. word2vec); count
modelos, in contrast to predict models, compute the co-
occurrence of words within these large text corpora. El
three CBOW models included: 1) a concatenation of the
ukwac web crawling corpus (∼2 billion words) y
the subtitle corpus (∼385 million words; window size =
12 palabras, 300 dimensions, most frequent 150,000 palabras);
2) the subtitle corpus only (window size 12 palabras, 300
dimensions, most frequent 150,000 palabras); y 3) a con-
catenation of the British National Corpus (∼2 billion
palabras), ukwac corpus, y el 2009 Wikipedia dump
Orwig et al.
501
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
oh
C
norte
_
a
_
0
1
6
5
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(∼800 million tokens; window size = 11 palabras, 400 di-
mensions, most frequent 300,000 palabras). CBOW models
have previously demonstrated high correlations with
human relatedness judgments (Mandera et al., 2017) como
well as human creativity ratings across a range of common
creativity tasks, including the AUT (Beaty & Johnson, 2020).
The two count models include: 1) a latent semantic analysis
modelo, Touchstone Applied Science Associates, cual
computes word co-occurrences within a text corpus
(37,000 documentos, middle and high school textbooks
and literary words, 92,393 different words), followed by a
singular value decomposition on the resulting sparse
matrix (300 dimensions; cf. Prabhakaran et al., 2014); y
2) the global vectors (Glove; Pennington, Socher, &
Manning, 2014) modelo, which is trained on ∼6 billion
tokens (300 dimensions, arriba 400,000 palabras) and uses
weighted least squares to extract global information
across a concatenation of the 2014 Wikipedia dump and
the Gigaword corpus (news publications from 2009 a
2010).
All five spaces were used to compute the semantic dis-
tance between the AUT item (p.ej., box) and participants’
respuestas, where the cosine angle between the word
vectors represents semantic similarity; semantic distance
is then computed by subtracting this similarity from 1
(Kenett & Faust, 2019; Beaty, Christensen, Benedek,
Silvia, & Schacter, 2017; Verde, 2016; Prabhakaran et al.,
2014). Following Beaty and Johnson (2020), we used latent
variable modeling to extract the common variance from the
five semantic models. This approach has the benefit of
reducing the influence of any one model—which has been
shown to yield idiosyncratic values specific to the given
model and text corpus employed (Mandera et al., 2017)—
thus boosting the reliability and generalizability of results.
Using Mplus 8, we specified a confirmatory factor analysis
(CFA) that modeled semantic distance, human creativity
ratings, and Gv as three latent variables using the full sample
of participants with available data (norte = 186). For semantic
distancia, the mean values of the five semantic models
served as indicators for three lower order latent variables,
corresponding to the three AUT tasks (box, rope, y
resonancia magnética); a higher order variable was indicated by these three
lower order variables. For human ratings, the mean values
of the four raters served as indicators for three lower order
latent variables (box, rope, and MRI); a higher order vari-
able was indicated by these three lower order variables.
For Gv, the summed scores of the three tasks were mod-
eled as indicators of a Gv factor. The variance of the latent
variables was fixed to one; all indicators were standardized.
For the fMRI analyses below, we extracted factor scores
from the two higher order semantic distance and human
creativity variables.
MRI Acquisition and Preprocessing
Resting-state MRI data were acquired for all participants on
a 3 T Siemens Magnetom MRI system using a 16-channel
head coil (Cifra 1). High-resolution T1 scans were ac-
quired for anatomical normalization. BOLD T2*-weighted
functional images were acquired with gradient EPI se-
quence with the following parameters: repetition time =
2000 mseg, echo time = 30 mseg, flip angle = 78°, 192-
mm field of view, 32 axial slices, 3.5 × 3.5 × 4.0 mm, enterrar-
leaved slice ordering, sequence length = 5 mín..
Participants were instructed to relax awake in the scanner
with eyes closed for the duration of the scan.
MRI data for both anatomical and functional images were
preprocessed using FMRIB Software Library v5.0.7 and
MATLAB 2017a (The MathWorks Inc.). The anatomical
and functional preprocessing pipelines were adapted from
previous work (Diez et al., 2019). The anatomical T1 pre-
processing included: reorientation to right-posterior-
inferior; alignment to anterior and posterior commissures;
desmontaje del cráneo; gray matter, white matter, and cerebrospi-
nal fluid segmentation; and computation of nonlinear
transformation between individual skull-stripped T1 and
2-mm resolution Montreal Neurological Institute (MNI)
152 template images. The fMRI preprocessing pipeline
incluido: slice time correction, reorientation to right-
posterior-inferior, realigning functional volumes within
runs with a rigid body transformations (six parameters
linear transformation), computation of the transformation
between individual skull-stripped T1 and mean functional
images, normalización de intensidad, and removal of con-
founding factors from the data using linear regression—
incluido 12 motion-related covariates (rigid motion pa-
rameters and its derivatives), linear and quadratic terms,
and five components each from the lateral ventricles and
white matter. Global signal regression was not applied
because of the negative correlations this can introduce.
Transformation of resting-state data to MNI space was
performed, concatenating the transformation from func-
tional to structural and from structural to MNI, spatial
smoothing with an isotropic Gaussian kernel of 6-mm
FWHM, and band-pass filtering (0.01–0.08 Hz) to reduce
low-frequency drift and high-frequency noise were also ap-
plied. Head motion was quantified using realignment pa-
rameters obtained during image preprocessing, incluido
three translation and three rotation estimates. Scrubbing of
time points with excess head motion interpolated all time
points with a frame displacement > 0.2 mm was applied.
No participants demonstrated excessive head motion;
de este modo, none was removed from the study based on these
criteria. The distributions of the correlations across time
series were reviewed for possible contamination; no out-
liers were observed from the whole-brain connectivity
distributions.
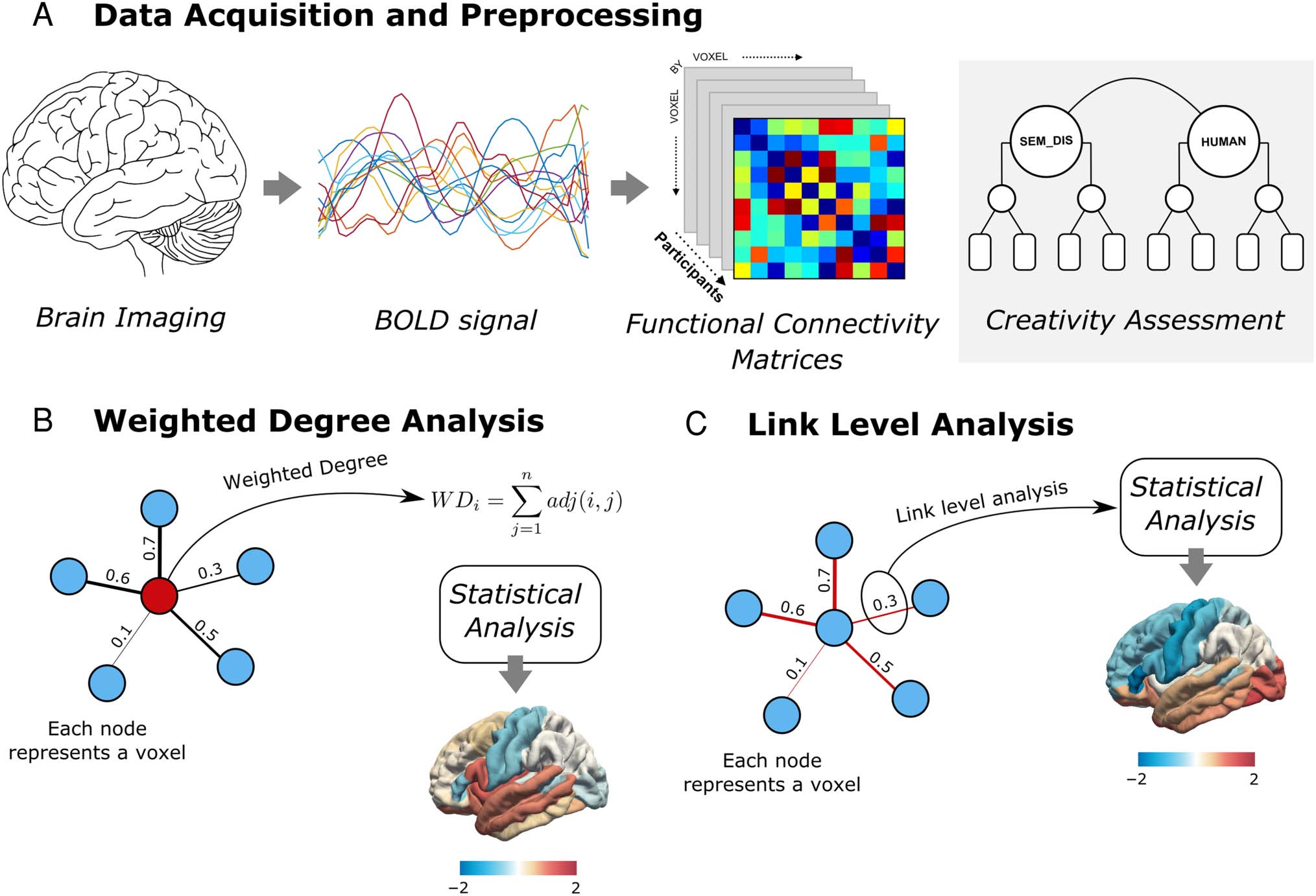
Weighted-Degree Functional Connectivity Analysis
Pearson correlation coefficients were used to calculate the
connectivity matrices of each participant using the time
series of all cortical gray matter voxels (Cifra 1). A r-to-z
Fisher transformation was applied to the resulting
502
Revista de neurociencia cognitiva
Volumen 33, Número 3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
oh
C
norte
_
a
_
0
1
6
5
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
oh
C
norte
_
a
_
0
1
6
5
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. Métodos. A sample of healthy adults underwent resting-state fMRI scans, from which we computed voxel-wise connectivity matrices
between all gray matter voxels. Behavioral assessment enabled the analysis of individual differences in functional connectivity associated with creative
actuación.
correlation matrix, and negative values were removed. A
minimize noise, we considered only the most significant
links using a false discovery rate at q-level < 0.005
(Benjamini & Hochberg, 1995). To evaluate the relative
prominence of each voxel in the whole-brain architecture
of each individual, voxel-level WD values were computed
(Figure 1). After obtaining a high-resolution 52,769 ×
52,769 connectivity matrix for each participant, we summed
all the weighted connections of each voxel to generate a
WD map showing the extent to which each voxel is func-
tionally connected to the rest of the brain (Ortiz-Terán
et al., 2017). A general linear model was used to compute
the association between WD and DT score. All statistical
analyses were corrected for participant age, sex, and Gv.
Whole-brain correction for multiple comparisons was com-
puted using Monte Carlo simulation with 10,000 iterations
to estimate the probability of false-positive clusters with a
two-tailed p < .05 (3dClustSim; afni.nimh.nih.gov).
Link-Level Functional Connectivity
Strength Analysis
To further investigate relationships between DT and corti-
cal regions identified in the WD analysis, we evaluated if DT
performance correlated with link-level connectivity
strength values across brain areas. To evaluate link-level
functional connectivity strength, MRI data were down-
sampled to 6-mm isotropic voxels to reduce dimension-
ality, resulting in a 6620 × 6620 connectivity matrix for
each participant. A general linear model was used at
every link of the network to evaluate the association of
the link weight and the DT scores (separately for seman-
tic distance and human ratings). Whole-brain correction
for multiple comparisons was computed adapting the
Monte Carlo simulation method to networks. Ten thou-
sand random networks were generated with the same
smoothing properties, to compute a false-positive cluster
size with a two-tailed p < .001. Compared to WD maps,
where clusters were defined as contiguous voxels, here,
clusters were defined as links that connect contiguous
voxel groups. We reduced the dimensionality of the sur-
viving links for visualization purposes. The statistically
significant links positively/negatively associated with DT
were then represented in a connectogram in Neuromarvl
(https://immersive.erc.monash.edu/neuromarvl/). Cortical
surfaces were visualized using the population-average
landmark and surface-based projections of CARET soft-
ware. We used Caret v5.65 software to represent the
Orwig et al.
503
results in a three-dimensional Population-Average Landmark
and Surface-based surface using the “enclosing voxel
algorithm” and “fiducial and flat mapping” settings.
Surface images were displayed using a color scale based
on T-scores.
RESULTS
Correlation between Semantic Distance and
Human Creativity Ratings
Before moving to the fMRI analysis, we examined the rela-
tionship between semantic distance and human creativity
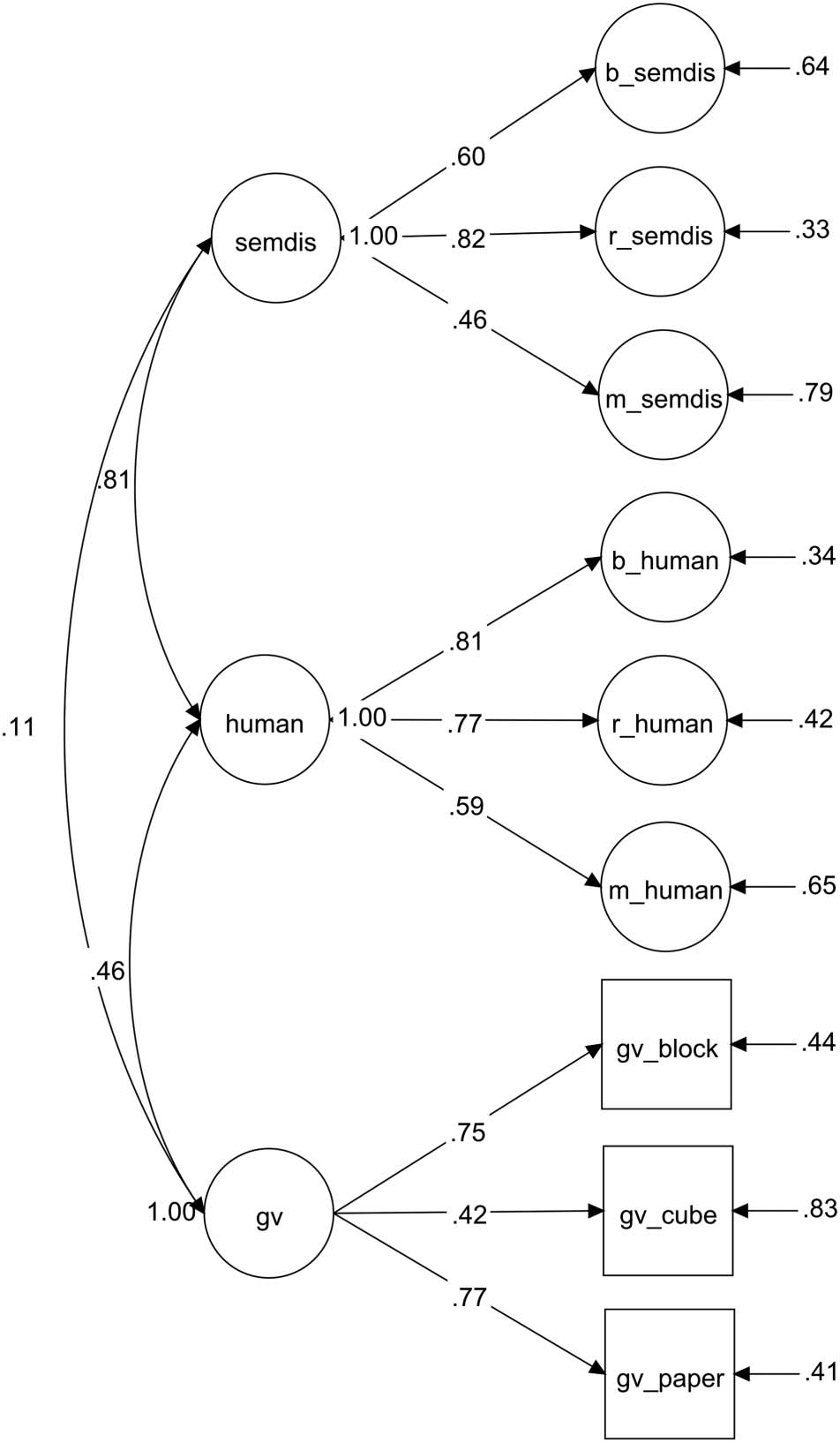
ratings. The CFA showed good fit: χ2 (396 df ) 643.242,
p < .001; comparative fit index = .945; root mean square
error of approximation = .058 (90% CI [0.05, .066]); stan-
dardized root means square residual = .074 (Figure 2). We
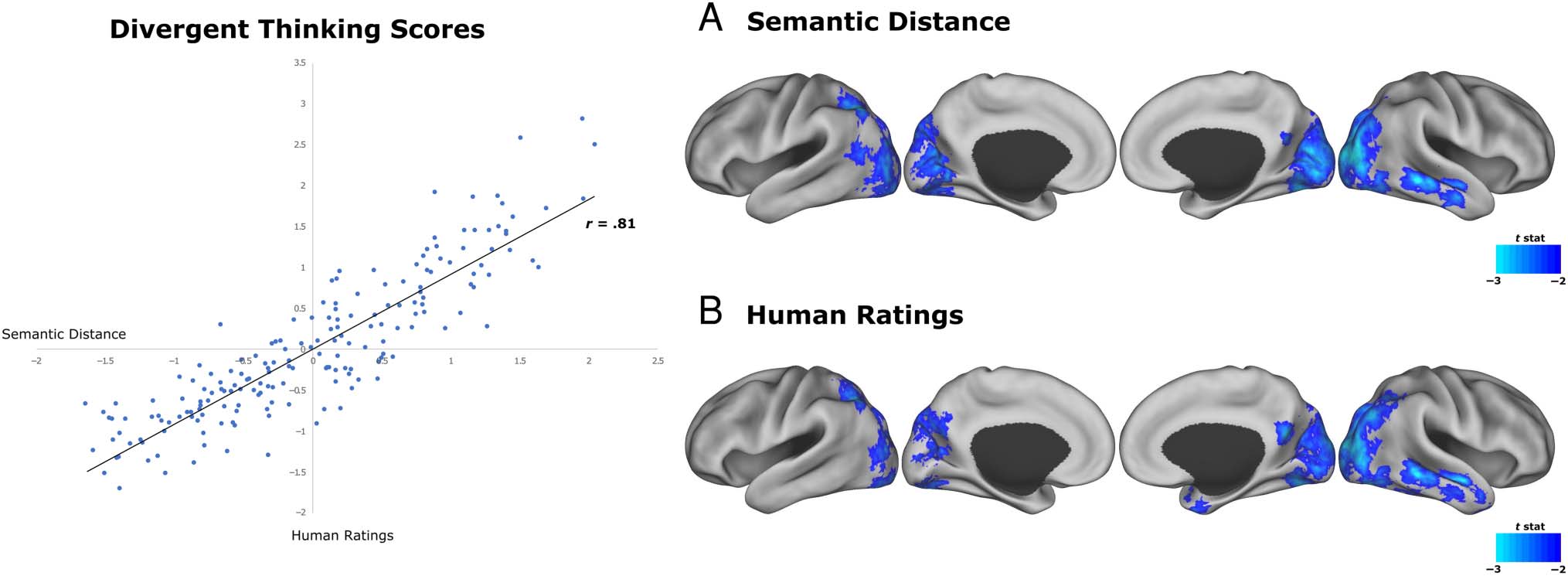
found a large latent correlation between semantic distance
and human creativity ratings, r = .81 ( p < .001), consistent
with the large effect reported in Beaty and Johnson (2020)
using only the laboratory-based AUT data. This result indi-
cates a high degree of overlap between semantic distance
and human ratings (see Figure 3). Gv correlated significantly
with human creativity ratings (r = .46, p < .001)—as previ-
ously reported (see Frith et al., 2020)—but not semantic dis-
tance (r = .11, p = .29), indicating that human ratings share
more variance with general cognitive ability than with auto-
mated assessments of creativity (cf. Beaty & Johnson, 2020).
Brain Hubs Associated with DT
Our first WD analysis aimed to identify cortical hubs related
to individual differences in DT assessed via semantic
distance. Results showed that DT semantic distance was
negatively correlated with WD of voxels in the occipital
cortex, along with parietal-occipital and temporal regions
(Figure 3A). WD across lateral occipital and superior
parietal regions was negatively correlated with DT. Similarly,
lower WD along the temporo-parietal junction and right
middle temporal gyrus was associated with higher DT
performance. WD of the left inferior parietal sulcus was neg-
atively associated with DT. There were no positive associa-
tions between WD and DT.
In addition, we performed WD analysis with human ratings
of DT (Figure 3B). WD maps generated with human ratings
of DT shared similar cortical distributions to the semantic
distance measure, with 51% of voxels shared between the
two maps. Results showed that human ratings of DT were
negatively correlated with WD of voxels in the occipital cortex
and right temporal regions. WD of the right temporal pole
was negatively associated with DT. There were no positive
associations between WD and human ratings of DT.
Individual Connectivity Patterns and DT
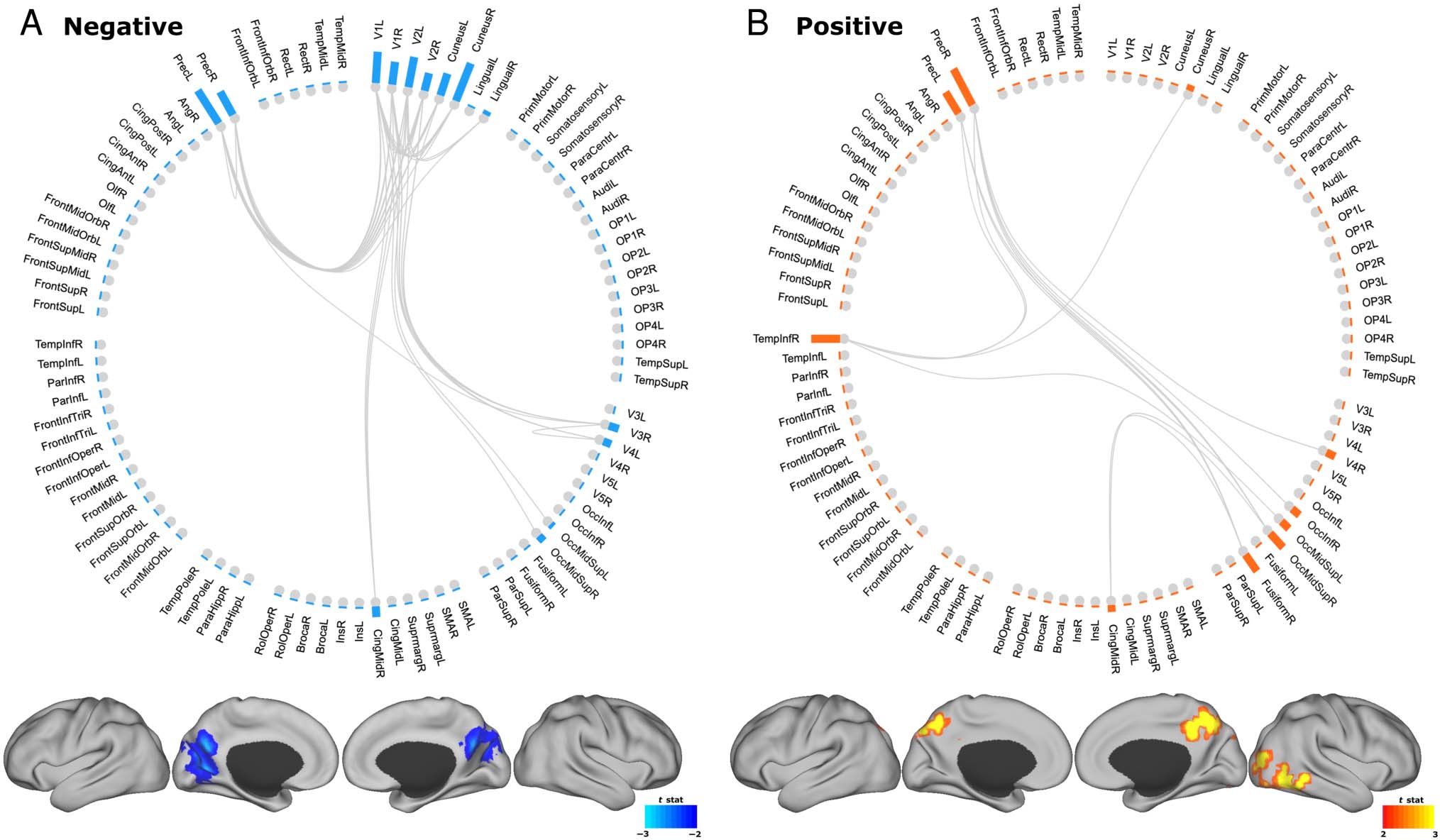
Link-level analysis revealed individual links between brain
regions negatively associated with semantic distance of
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
o
c
n
_
a
_
0
1
6
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. CFA of semantic distance, human creativity ratings, and
visuospatial intelligence. Observed variables are depicted as squares,
and latent variables are depicted as circles. For interpretability, the
observed variables for semantic distance and human creativity ratings
are not shown in the diagram. b_semdis = box task, semantic distance;
r_semdis = rope task, semantic distance; m_semdis = MRI tasks,
semantic distance; b_human = box task, human ratings; r_human =
rope task, human ratings; m_human = MRI tasks, human ratings;
gv_block = visuospatial intelligence, block rotation; gv_cube =
visuospatial intelligence, cube comparison; gv_paper = visuospatial
intelligence, paper folding. n = 186.
DT, corrected for multiple comparisons (Figure 4A).
Strength of connectivity within the primary visual cortex
was negatively correlated with DT. In other words, high
local connectivity within the visual area was associated
with lower DT performance. Projections from bilateral
V1 and V2 to the precuneus were negatively associated
with DT. Strength of connectivity between primary visual
areas and precuneus was negatively associated with DT.
In addition, we identified links positively associated with
DT semantic distance (Figure 4B). The strength of the con-
nection between the precuneus and right inferior temporal
504
Journal of Cognitive Neuroscience
Volume 33, Number 3
Figure 3. Weighted degree. WD across the lateral occipital and superior parietal cortex was negatively associated with DT. WD along the right middle
temporal gyrus negatively correlated with DT. Semantic distance and human ratings of DT were consistent in these findings.
gyrus was positively correlated with DT. Projections from
the precuneus to right inferior occipital cortex (fusiform),
right inferior temporal, and occipital cortices were posi-
tively correlated with DT semantic distance. Link-level
analysis with human ratings of DT revealed a negative
association similar to that of semantic distance measures.
Connectivity across the occipital cortex and projection to
the right primary motor and somatosensory areas were
negatively associated with human ratings of DT. We did
not find any positive association with human ratings of
DT. Despite the similar negative associations between
the two measures, semantic distance reveals a positive
association between individual connectivity strength and
DT, which human ratings do not capture.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
o
c
n
_
a
_
0
1
6
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Link level. Strength of connectivity within the primary visual cortex was negatively correlated with DT. Projections from the precuneus to
the right inferior occipital and temporal cortices were positively correlated with DT.
Orwig et al.
505
DISCUSSION
This study applies graph theory techniques in resting-state
functional connectivity MRI data to explore how individual
differences in assembly of resting-state networks are
related to creativity, using a new application to automat-
ically assess creativity via semantic distance. Critically,
WD results from the semantic distance measure were
largely consistent with human ratings—producing similar
functional connectivity maps—further validating this ob-
jective assessment as a viable measure of creative perfor-
mance. The findings thus contribute to the growing
neuroscience literature on individual differences in crea-
tive thinking and demonstrate the utility of automated
scoring approaches to capture variance in creativity at the
neural level.
Lateral Visual Cortex Centralizes the Emergence
of DT
Among the most salient findings, our analysis revealed a
negative association between DT and WD across lateral
occipital, inferior parietal, and right middle temporal
areas. Secondary link-level analyses revealed that func-
tional connectivity between the left precuneus and right
lateral visual pathway was positively associated with DT,
whereas strength of connections within primary visual re-
gions were negatively associated with DT. These results
indicate that there is less integration of visual information
during resting state in individuals with higher DT scores.
High local connectivity within the primary visual system
represents a high degree of segregation. One interpreta-
tion of these results could be that creative people are
more prone to engage in internally directed cognition
(such as mind wandering) in the absence of an external
task. Schooler et al. (2011) describe the phenomenon of
perceptual decoupling, the capacity to disengage atten-
tion from perception, during episodes of mind wander-
ing. Perceptual decoupling during resting-state may
indicate engagement in more abstract, internally directed
attention, which characterizes creative individuals.
Internally directed attention has been associated with ex-
tended deactivation in occipital and inferior parietal cor-
tices (Benedek et al., 2016). In addition, Sepulcre (2014)
identifies the lateral occipital cortex and the inferior pari-
etal lobule as integrators of sensory information, toward
more high-level cognition. One possibility is that partici-
pants with reduced functional connectivity to the primary
visual system are engaged in more creative, self-generated
cognitive processes during resting state.
Link-level analyses showed that strength of connec-
tions within the primary visual cortex was negatively as-
sociated with DT, whereas connectivity from the right
lateral and ventral occipital cortex to the precuneus was
positively associated with DT. At first glance, these results
may appear to be in conflict, as we see projections from
the precuneus both positively and negatively associated
with DT. This differential connectivity reflects the atten-
uation of primary visual areas, with higher connectivity
between lateral inferior occipital regions and multimodal
integration areas being positively associated with DT. The
precuneus is a core hub of the DMN and is thought to
play a role in multimodal integration. In a study examin-
ing gray matter density and verbal creativity, it was found
that increased gray matter density in precuneus was pos-
itively associated with DT (Benedek, 2018). Our results
suggest that regions of the precuneus may be differentially
involved in creative problem solving. Stronger connec-
tivity between visual regions and precuneus may reflect
tighter coupling between perception and generation sys-
tems, whereas decreased connectivity between the visual
cortex and semantic regions may reflect weaker commu-
nication between perception and object representation
along the ventral stream. Differential pathways for the in-
tegration of visual information may elucidate the neural
mechanisms involved in DT, although further research
is needed to support this claim.
Previous studies of task-based and resting-state data have
described consistent patterns of network interaction
between DMN and executive control networks in relation
to creative cognition (Christensen, Benedek, Silvia, &
Beaty, 2019; Beaty, Kenett, et al., 2018; Beaty et al., 2015).
DMN activity is thought to contribute to the generation of
candidate ideas, whereas executive control networks exert
top–down monitoring to meet external constraints (Beaty
et al., 2016). Notably, our results do not reflect this interac-
tion between default mode and executive control networks,
suggesting that the novel semantic distance measures and
graph theory analyses reported here capture variance in cre-
ative thinking that is distinct from these network dynamics.
The finding of a negative association between WD in
the right middle temporal gyrus and DT raises questions
regarding the involvement of this region in verbal creativ-
ity. Task-based studies of DT have applied dynamic causal
modeling to describe unidirectional control from the pFC
over the middle temporal gyrus ( Vartanian et al., 2018).
Other studies have implicated the middle temporal gyrus
in idea generation (Ellamil, Dobson, Beeman, & Christoff,
2012) and semantic integration ( Jung-Beeman, 2005). It
has been suggested the right hemisphere is preferentially
involved in the processing of distantly related “coarse” se-
mantic information (Beeman et al., 1994). The negative
association between DT performance and WD of right
middle temporal areas demonstrate that, at rest, these re-
gions are less functionally connected to the rest of the
brain in participants who scored higher on the AUT.
Link-level connectivity strength between inferior tempo-
ral areas and the precuneus was positively associated
with DT, which may reflect more coupling of semantic
processing and multimodal-integration areas. In summary,
our findings extend past research by identifying neural
correlates of individual creative thinking using objective
and automated assessments of creativity based on seman-
tic distance.
506
Journal of Cognitive Neuroscience
Volume 33, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
o
c
n
_
a
_
0
1
6
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Limitations and Future Directions
An important consideration in cognitive and neuroscience
studies of creativity concerns the operationalization of cre-
ativity. Many studies focus on the creative quality of ideas
(assessed via human creativity ratings or semantic distance)
as the primary dependent outcome, but this outcome alone
can only provide a modest window into the cognitive basis
of idea generation. Viewed as a high-level cognitive ability,
creative thought (or the output from creativity tasks and
their assessed quality) likely results from the complex
interplay of multiple “lower level” cognitive processes, such
as memory retrieval, cognitive control, and attention. We
encourage future research to further identify the cognitive
processes that give rise to creative ideas.
The observed overlap between WD maps generated
from human ratings and semantic distance measures
showed 51% of voxels common between the two maps.
Currently, there is no clear benchmark for determining
the magnitude or significance between these maps.
Although the cortical distributions appear visually similar
(see Figure 3), we cannot explain what is reflected by the
nonoverlapping regions. Speculatively, the distinction may
be because of features of ideas that humans consider when
rating creative responses that are not captured by compu-
tational semantic distance (e.g., utility or cleverness of an
idea). These unique neural features should be more deeply
explored in future work (cf. Vartanian et al., 2020). The
present research uses resting-state fMRI data to examine
functional networks of the brain at rest. Given the nature
of resting-state data, there is a considerable amount noise.
In collecting the data, we are not able to control for the par-
ticipant’s mood or state of mind during the resting-state
scan and there is known to be high variability between
scans of the same individual. Although this measure is sub-
ject to noise, the size of our sample (n = 175) allows us to
be confident that these findings are not simply the result of
noise and do provide new insight into individual differ-
ences in the creative brain.
Future studies may explore the causal relationship be-
tween identified regions and DT performance. For exam-
ple, transcranial magnetic stimulation to the right middle
temporal gyrus before completion of the AUT may pro-
vide further clarification into the role of this brain region
in the integration of semantically distant concepts (Luft,
Zioga, Thompson, Banissy, & Bhattacharya, 2018). In ad-
dition, future work in the field may explore cortical gene
expression related with patterns of functional connectiv-
ity to describe how specific genetic pathways may be re-
lated to individual variability in creativity.
Conclusions
This study applied graph theory techniques to resting-state
fMRI data to measure individual differences in functional
connectivity associated with DT. Computational semantic
models were applied to calculate the semantic distance
between objects and responses to obtain an automated
measure of DT performance. This automated measure of
DT provided results consistent with (but not identical to)
those of human ratings, highlighting the potential of
semantic distance as a reliable metric of creativity. The find-
ing of a negative association between resting-state connec-
tivity in the occipital cortex and DT may be explained by
perceptual decoupling, reflecting a higher degree of inter-
nally directed cognition in participants who scored higher
on the AUT. In addition, our findings suggest a positive
association between DT and connectivity from the pre-
cuneus to right inferior temporal regions. More work is
needed to validate these findings and further characterize
the complexities of the creative brain.
Acknowledgments
The authors declared no potential conflicts of interest with respect
to the research, authorship, and/or publication of this paper.
Reprint requests should be sent to William Orwig, 149 13th St,
Suite 5.209, Department of Radiology, Massachusetts General
Hospital and Harvard Medical School, Charlestown, MA 02129,
or via e-mail: worwig@mgh.harvard.edu.
Author Contributions
William Orwig: Conceptualization; Formal analysis;
Visualization; Writing – original draft; Writing – review &
editing. Ibai Diez: Formal analysis; Methodology. Patrizia
Vannini: Writing – review & editing. Roger Beaty: Data
curation; Supervision; Writing – review & editing. Jorge
Sepulcre: Supervision; Writing – review & editing.
Funding Information
This research was supported by grants from the National
Institutes of Health (R01AG061811 to J. S.; R01AG061083
to P. V. and J. S.). R. E. B. is supported by a grant from the
National Science Foundation (DRL-1920653). This research
was supported by grant RFP-15-12 to R. E. B., from the
Imagination Institute (www.imagination-institute.org),
funded by the John Templeton Foundation.
REFERENCES
Adnan, A., Beaty, R., Silvia, P., Spreng, R. N., & Turner, G. R.
(2019). Creative aging: Functional brain networks associated
with divergent thinking in older and younger adults.
Neurobiology of Aging, 75, 150–158. DOI: https://doi.org
/10.1016/j.neurobiolaging.2018.11.004, PMID: 30572185
Andrews-Hanna, J. R., Smallwood, J., & Spreng, R. N. (2014).
The default network and self-generated thought: Component
processes, dynamic control, and clinical relevance. Annals
of the New York Academy of Sciences, 1316, 29–52. DOI:
https://doi.org/10.1111/nyas.12360, PMID: 24502540, PMCID:
PMC4039623
Beaty, R. E., Benedek, M., Kaufman, S. B., & Silvia, P. J. (2015).
Default and executive network coupling supports creative
idea production. Scientific Reports, 5, 10964. DOI: https://
Orwig et al.
507
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
o
c
n
_
a
_
0
1
6
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
doi.org/10.1038/srep10964, PMID: 26084037, PMCID:
PMC4472024
Beaty, R. E., Benedek, M., Silvia, P. J., & Schacter, D. L. (2016).
Creative cognition and brain network dynamics. Trends in
Cognitive Sciences, 20, 87–95. DOI: https://doi.org/10.1016
/j.tics.2015.10.004, PMID: 26553223, PMCID: PMC4724474
Beaty, R. E., Chen, Q., Christensen, A. P., Qiu, J., Silvia, P. J.,
& Schacter, D. L. (2018). Brain networks of the imaginative
mind: Dynamic functional connectivity of default and cognitive
control networks relates to openness to experience. Human
Brain Mapping, 39, 811–821. DOI: https://doi.org/10.1002
/hbm.23884, PMID: 29136310, PMCID: PMC5764809
Beaty, R. E., Christensen, A. P., Benedek, M., Silvia, P. J., &
Schacter, D. L. (2017). Creative constraints: Brain activity and
network dynamics underlying semantic interference during
idea production. Neuroimage, 148, 189–196. DOI: https://
doi.org/10.1016/j.neuroimage.2017.01.012, PMID: 28082106,
PMCID: PMC6083214
Beaty, R. E., & Johnson, D. R. (2020). Automating creativity
assessment with SemDis: An open platform for computing
semantic distance. PsyArXiv Preprints. DOI: https://doi.org
/10.31234/osf.io/nwvps
Beaty, R. E., Kenett, Y. N., Christensen, A. P., Rosenberg, M. D.,
Benedek, M., Chen, Q., et al. (2018). Robust prediction of
individual creative ability from brain functional connectivity.
Proceedings of the National Academy of Sciences, U.S.A.,
115, 1087–1092. DOI: https://doi.org/10.1073/pnas
.1713532115, PMID: 29339474, PMCID: PMC5798342
Beaty, R. E., Silvia, P. J., Nusbaum, E. C., Jauk, E., & Benedek, M.
(2014). The roles of associative and executive processes in
creative cognition. Memory & Cognition, 42, 1186–1197.
DOI: https://doi.org/10.3758/s13421-014-0428-8, PMID:
24898118
Buckner, R. L., Sepulcre, J., Talukdar, T., Krienen, F. M., Liu, H.,
Hedden, T., et al. (2009). Cortical hubs revealed by intrinsic
functional connectivity: Mapping, assessment of stability, and
relation to Alzheimer’s disease. Journal of Neuroscience,
29, 1860–1873. DOI: https://doi.org/10.1523/JNEUROSCI
.5062-08.2009, PMID: 19211893, PMCID: PMC2750039
Bullmore, E. T., & Bassett, D. S. (2011). Brain graphs: Graphical
models of the human brain connectome. Annual Review
of Clinical Psychology, 7, 113–140. DOI: https://doi.org/10
.1146/annurev-clinpsy-040510-143934, PMID: 21128784
Bullmore, E. T., & Sporns, O. (2009). Complex brain networks:
Graph theoretical analysis of structural and functional
systems. Nature Reviews Neuroscience, 10, 186–198. DOI:
https://doi.org/10.1038/nrn2575, PMID: 19190637
Christensen, A. P., Benedek, M., Silvia, P. J., & Beaty, R. E. (2019).
Executive and default network connectivity reflects conceptual
interference during creative imagery generation. PsyArXiv
Preprints. DOI: https://doi.org/10.31234/osf.io/n438d
Diez, I., Ortiz-Terán, L., Williams, B., Jalilianhasanpour, R.,
Ospina, J. P., Dickerson, B. C., et al. (2019). Corticolimbic
fast-tracking: Enhanced multimodal integration in functional
neurological disorder. Journal of Neurology, Neurosurgery,
& Psychiatry, 90, 929–938. DOI: https://doi.org/10.1136
/jnnp-2018-319657, PMID: 30850473, PMCID: PMC6625895
Dumas, D., Organisciak, P., & Doherty, M. (2020). Measuring
divergent thinking originality with human raters and text-
mining models: A psychometric comparison of methods.
Psychology of Aesthetics, Creativity, and the Arts. DOI:
https://doi.org/10.1037/aca0000319
Ellamil, M., Dobson, C., Beeman, M., & Christoff, K. (2012).
Evaluative and generative modes of thought during the
creative process. Neuroimage, 59, 1783–1794. DOI: https://
doi.org/10.1016/j.neuroimage.2011.08.008, PMID: 21854855
Beeman, M., Friedman, R. B., Grafman, J., Perez, E., Diamond, S.,
Feng, Q., He, L., Yang, W., Zhang, Y., Wu, X., & Qiu, J. (2019).
& Lindsay, M. B. (1994). Summation priming and coarse
semantic coding in the right hemisphere. Journal of Cognitive
Neuroscience, 6, 26–45. DOI: https://doi.org/10.1162/jocn
.1994.6.1.26, PMID: 23962328
Benedek, M. (2018). Internally directed attention in creative
cognition. In R. E. Jung & O. Vartanian (Eds.), The Cambridge
handbook of the neuroscience of creativity (1st ed.,
pp. 180–194). Cambridge: Cambridge University Press.
DOI: https://doi.org/10.1017/9781316556238.011
Benedek, M., Christensen, A. P., Fink, A., & Beaty, R. E. (2019).
Creativity assessment in neuroscience research. Psychology
of Aesthetics, Creativity, and the Arts, 13, 218–226. DOI:
https://doi.org/10.1037/aca0000215
Benedek, M., Jauk, E., Beaty, R. E., Fink, A., Koschutnig, K., &
Neubauer, A. C. (2016). Brain mechanisms associated with
internally directed attention and self-generated thought.
Scientific Reports, 6, 22959. DOI: https://doi.org/10.1038
/srep22959, PMID: 26960259, PMCID: PMC4785374
Benedek, M., Mühlmann, C., Jauk, E., & Neubauer, A. C. (2013).
Assessment of divergent thinking by means of the subjective
top-scoring method: Effects of the number of top-ideas and
time-on-task on reliability and validity. Psychology of Aesthetics,
Creativity, and the Arts, 7, 341–349. DOI: https://doi.org
/10.1037/a0033644, PMID: 24790683, PMCID: PMC4001084
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false
discovery rate: A practical and powerful approach to multiple
testing. Journal of the Royal Statistical Society, Series B,
Methodological, 57, 289–300. DOI: https://doi.org/10.1111
/j.2517-6161.1995.tb02031.x
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L. (2008).
The brain’s default network: Anatomy, function, and
relevance to disease. Annals of the New York Academy of
Sciences, 1124, 1–38. DOI: https://doi.org/10.1196/annals
.1440.011, PMID: 18400922
Verbal creativity is correlated with the dynamic reconfiguration
of brain networks in the resting state. Frontiers in Psychology,
10, 894. DOI: https://doi.org/10.3389/fpsyg.2019.00894, PMID:
31068873, PMCID: PMC6491857
Fink, A., & Benedek, M. (2019). The neuroscience of creativity.
Neuroforum, 25, 231–240. DOI: https://doi.org/10.1515
/nf-2019-0006
Fink, A., Koschutnig, K., Hutterer, L., Steiner, E., Benedek, M.,
Weber, B., et al. (2014). Gray matter density in relation to
different facets of verbal creativity. Brain Structure and
Function, 219, 1263–1269. DOI: https://doi.org/10.1007
/s00429-013-0564-0, PMID: 23636224
Frith, E., Elbich, D. B., Christensen, A. P., Rosenberg, M. D.,
Chen, Q., Kane, M. J., et al. (2020). Intelligence and creativity
share a common cognitive and neural basis. Journal of
Experimental Psychology: General. DOI: https://doi.org
/10.1037/xge0000958, PMID: 33119355
Gao, Z., Zhang, D., Liang, A., Liang, B., Wang, Z., Cai, Y., et al.
(2017). Exploring the associations between intrinsic brain
connectivity and creative ability using functional connectivity
strength and connectome analysis. Brain Connectivity, 7,
590–601. DOI: https://doi.org/10.1089/brain.2017.0510,
PMID: 28950708
Green, A. E. (2016). Creativity, within reason: Semantic distance
and dynamic state creativity in relational thinking and
reasoning. Current Directions in Psychological Science,
25, 28–35. DOI: https://doi.org/10.1177/0963721415618485
Green, A. E., Cohen, M. S., Raab, H. A., Yedibalian, C. G., &
Gray, J. R. (2015). Frontopolar activity and connectivity
support dynamic conscious augmentation of creative state.
Human Brain Mapping, 36, 923–934. DOI: https://doi.org
/10.1002/hbm.22676, PMID: 25394198, PMCID: PMC6869232
Guilford, J. P. (1967). The nature of human intelligence.
New York: McGraw-Hill.
508
Journal of Cognitive Neuroscience
Volume 33, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
o
c
n
_
a
_
0
1
6
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Heinen, D. J. P., & Johnson, D. R. (2018). Semantic distance: An
automated measure of creativity that is novel and appropriate.
Psychology of Aesthetics, Creativity, and the Arts, 12, 144–156.
DOI: https://doi.org/10.1037/aca0000125
Jung-Beeman, M. (2005). Bilateral brain processes for
comprehending natural language. Trends in Cognitive
Sciences, 9, 512–518. DOI: https://doi.org/10.1016/j.tics
.2005.09.009, PMID: 16214387
Kenett, Y. N., Betzel, R. F., & Beaty, R. E. (2020). Community
structure of the creative brain at rest. Neuroimage, 210,
116578. DOI: https://doi.org/10.1016/j.neuroimage.2020
.116578, PMID: 31982579
Kenett, Y. N., & Faust, M. (2019). A semantic network cartography of
the creative mind. Trends in Cognitive Sciences, 23, 271–274.
DOI: https://doi.org/10.1016/j.tics.2019.01.007, PMID: 30803872
Li, J., Zhang, D., Liang, A., Liang, B., Wang, Z., Cai, Y., et al.
(2017). High transition frequencies of dynamic functional
connectivity states in the creative brain. Scientific Reports,
7, 46072. DOI: https://doi.org/10.1038/srep46072, PMID:
28383052, PMCID: PMC5382673
Liu, Z., Zhang, J., Xie, X., Rolls, E. T., Sun, J., Zhang, K., et al.
(2018). Neural and genetic determinants of creativity.
Neuroimage, 174, 164–176. DOI: https://doi.org/10.1016
/j.neuroimage.2018.02.067, PMID: 29518564
Luft, C. D. B., Zioga, I., Thompson, N. M., Banissy, M. J., &
Bhattacharya, J. (2018). Right temporal alpha oscillations as
a neural mechanism for inhibiting obvious associations.
Proceedings of the National Academy of Sciences, U.S.A.,
115, E12144–E12152. DOI: https://doi.org/10.1073/pnas
.1811465115, PMID: 30541890, PMCID: PMC6310824
Mandera, P., Keuleers, E., & Brysbaert, M. (2017). Explaining
human performance in psycholinguistic tasks with models of
semantic similarity based on prediction and counting: A review
and empirical validation. Journal of Memory and Language,
92, 57–78. DOI: https://doi.org/10.1016/j.jml.2016.04.001
Mednick, S. A. (1962). The associative basis of the creative
process. Psychological Review, 69, 220–232. DOI: https://
doi.org/10.1037/h0048850, PMID: 14472013
Ortiz-Terán, L., Diez, I., Ortiz, T., Perez, D. L., Aragón, J. I.,
Costumero, V., et al. (2017). Brain circuit–gene expression
relationships and neuroplasticity of multisensory cortices in
blind children. Proceedings of the National Academy of
Sciences, U.S.A., 114, 6830–6835. DOI: https://doi.org/10.1073
/pnas.1619121114, PMID: 28607055, PMCID: PMC5495230
Pennington, J., Socher, R., & Manning, C. (2014). Glove: Global
vectors for word representation. In Proceedings of the 2014
Conference on Empirical Methods in Natural Language
Processing (EMNLP) (pp. 1532–1543). Doha, Qatar:
Association for Computational Linguistics. DOI: https://
doi.org/10.3115/v1/D14-1162
Prabhakaran, R., Green, A. E., & Gray, J. R. (2014). Thin slices of
creativity: Using single-word utterances to assess creative
cognition. Behavior Research Methods, 46, 641–659. DOI:
https://doi.org/10.3758/s13428-013-0401-7, PMID: 24163211,
PMCID: PMC4105589
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. L. (2001). A default mode of
brain function. Proceedings of the National Academy of
Sciences, U.S.A., 98, 676–682. DOI: https://doi.org/10.1073
/pnas.98.2.676, PMID: 11209064, PMCID: PMC14647
Salvi, C., & Bowden, E. M. (2016). Looking for creativity: Where
do we look when we look for new ideas? Frontiers in
Psychology, 7, 161. DOI: https://doi.org/10.3389/fpsyg.2016
.00161, PMID: 26913018, PMCID: PMC4753696
Salvi, C., Bricolo, E., Franconeri, S. L., Kounios, J., & Beeman, M.
(2015). Sudden insight is associated with shutting out visual
inputs. Psychonomic Bulletin & Review, 22, 1814–1819.
DOI: https://doi.org/10.3758/s13423-015-0845-0, PMID:
26268431
Schooler, J. W., Smallwood, J., Christoff, K., Handy, T. C.,
Reichle, E. D., & Sayette, M. A. (2011). Meta-awareness,
perceptual decoupling and the wandering mind. Trends
in Cognitive Sciences, 15, 319–326. DOI: https://doi.org
/10.1016/j.tics.2011.05.006, PMID: 21684189
Sepulcre, J. (2014). Functional streams and cortical integration
in the human brain. Neuroscientist, 20, 499–508. DOI:
https://doi.org/10.1177/1073858414531657, PMID: 24737695
Sepulcre, J., Sabuncu, M. R., & Goñi, J. (2014). Hubs and
pathways. In M. Mesulam & S. Kastner (Eds.), Brain
mapping: An encyclopedic reference (Vol. 2, pp. 441–447).
San Diego, CA: Elsevier. DOI: https://doi.org/10.1016/B978
-0-12-397025-1.00023-3
Sepulcre, J., Sabuncu, M. R., Yeo, T. B., Liu, H., & Johnson, K. A.
(2012). Stepwise connectivity of the modal cortex reveals the
multimodal organization of the human brain. Journal of
Neuroscience, 32, 10649–10661. DOI: https://doi.org/10
.1523/JNEUROSCI.0759-12.2012, PMID: 22855814, PMCID:
PMC3483645
Shi, L., Sun, J., Xia, Y., Ren, Z., Chen, Q., Wei, D., et al. (2018).
Large-scale brain network connectivity underlying creativity
in resting-state and task fMRI: Cooperation between default
network and frontal-parietal network. Biological Psychology,
135, 102–111. DOI: https://doi.org/10.1016/j.biopsycho.2018
.03.005, PMID: 29548807
Silvia, P. J. (2008). Discernment and creativity: How well can
people identify their most creative ideas? Psychology of
Aesthetics, Creativity, and the Arts, 2, 139–146. DOI: https://
doi.org/10.1037/1931-3896.2.3.139
Sporns, O., Chialvo, D. R., Kaiser, M., & Hilgetag, C. C. (2004).
Organization, development and function of complex brain
networks. Trends in Cognitive Sciences, 8, 418–425. DOI:
https://doi.org/10.1016/j.tics.2004.07.008, PMID: 15350243
Sun, J., Liu, Z., Rolls, E. T., Chen, Q., Yao, Y., Yang, W., et al.
(2019). Verbal creativity correlates with the temporal
variability of brain networks during the resting state. Cerebral
Cortex, 29, 1047–1058. DOI: https://doi.org/10.1093/cercor
/bhy010, PMID: 29415253
Takeuchi, H., Taki, Y., Hashizume, H., Sassa, Y., Nagase, T.,
Nouchi, R., et al. (2012). The association between resting
functional connectivity and creativity. Cerebral Cortex, 22,
2921–2929. DOI: https://doi.org/10.1093/cercor/bhr371,
PMID: 22235031
van den Heuvel, M. P., & Hulshoff Pol, H. E. (2010). Exploring
the brain network: A review on resting-state fMRI functional
connectivity. European Neuropsychopharmacology, 20,
519–534. DOI: https://doi.org/10.1016/j.euroneuro.2010
.03.008, PMID: 20471808
Vartanian, O., Beatty, E. L., Smith, I., Blackler, K., Lam, Q., &
Forbes, S. (2018). One-way traffic: The inferior frontal
gyrus controls brain activation in the middle temporal gyrus
and inferior parietal lobule during divergent thinking.
Neuropsychologia, 118, 68–78. DOI: https://doi.org/10.1016
/j.neuropsychologia.2018.02.024, PMID: 29477840
Vartanian, O., Smith, I., Lam, T. K., King, K., Lam, Q., & Beatty,
E. L. (2020). The relationship between methods of scoring
the alternate uses task and the neural correlates of divergent
thinking: Evidence from voxel-based morphometry.
Neuroimage, 223, 117325. DOI: https://doi.org/10.1016
/j.neuroimage.2020.117325, PMID: 32882380
Wei, D., Yang, J., Li, W., Wang, K., Zhang, Q., & Qiu, J. (2014).
Increased resting functional connectivity of the medial
prefrontal cortex in creativity by means of cognitive
stimulation. Cortex, 51, 92–102. DOI: https://doi.org
/10.1016/j.cortex.2013.09.004, PMID: 24188648
Orwig et al.
509
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
9
9
1
8
6
2
6
1
5
/
j
o
c
n
_
a
_
0
1
6
5
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3