Asociados verbales emparejados y el hipocampo:
The Role of Scenes
Ian A. clark, Misun Kim, and Eleanor A. Maguire
Abstracto
■ It is widely agreed that patients with bilateral hippocampal
damage are impaired at binding pairs of words together. Consecuencia-
frecuentemente, the verbal paired associates ( VPA) task has become
emblematic of hippocampal function. This VPA deficit is not well
understood and is particularly difficult for hippocampal theories
with a visuospatial bias to explain (p.ej., cognitive map and scene
construction theories). Resolving the tension among hippo-
campal theories concerning the VPA could be important for
leveraging a fuller understanding of hippocampal function.
Notablemente, VPA tasks typically use high imagery concrete words
and so conflate imagery and binding. To determine why VPA
engages the hippocampus, we devised an fMRI encoding task
involving closely matched pairs of scene words, pairs of object
palabras, and pairs of very low imagery abstract words. Encontramos
that the anterior hippocampus was engaged during process-
ing of both scene and object word pairs in comparison to ab-
stract word pairs, despite binding occurring in all conditions.
This was also the case when just subsequently remembered
stimuli were considered. Además, for object word pairs,
fMRI activity patterns in anterior hippocampus were more
similar to those for scene imagery than object imagery. Este
was especially evident in participants who were high imagery
users and not in mid and low imagery users. En general, nuestros resultados
show that hippocampal engagement during VPA, even when
object word pairs are involved, seems to be evoked by scene
imagery rather than binding. This may help to resolve the issue
that visuospatial hippocampal theories have in accounting for
verbal memory. ■
INTRODUCCIÓN
The field of hippocampal neuroscience is characterized
by vigorous debates. But one point on which there is
general agreement is that people with bilateral hippo-
campal damage and concomitant amnesia (hippocampal
amnesia) are significantly impaired on verbal paired
associates ( VPA) tareas. The VPA task is a widely used
instrument for testing verbal memory and has been a
continuous subtest within the Wechsler Memory Scale
(WMS) from its initial inception ( Wechsler, 1945) to the
present day ( WMS-IV; Wechsler, 2009). Although the VPA
task has been revised many times (p.ej., increasing the
number of word pairs to be remembered, changing the
ratio of difficult to easy word pairs), the basic premise has
remained the same. The requirement is to encode pairs
of words (p.ej., bag–truck), memory for which is then
probado. Testing can be conducted in multiple ways, pero
one primary outcome measure is performance on a
delayed cued recall test (es decir., the experimenter asks for
the word that goes with bag) 30 min after the completion
of the learning trials. Compared with matched healthy
control participants, patients with hippocampal amnesia
show a consistent and reliable deficit on delayed cued
recall tests (Giovanello, Verfaellie, & Keane, 2003; Spiers,
University College London
Maguire, & Burgess, 2001; Zola-Morgan, Squire, & Amaral,
1986; Graf & Schacter, 1985), and consequently, the VPA
has become emblematic of hippocampal function.
The VPA task is typically regarded as a verbal memory
tarea. Sin embargo, many theories focus on elucidating the
role of the hippocampus in visuospatial rather than
verbal processing. This includes accounts that consider
spatial navigation (Maguire et al., 2000; O’Keefe & Nadel,
1978), memoria autobiográfica (Hassabis & Maguire,
2007; Squire, 1992; Scoville & Milner, 1957), scene per-
ception (McCormick, Rosenthal, Molinero, & Maguire, 2017;
graham, Barense, & Sotavento, 2010), the mental construction of
visual scene imagery (Zeidman & Maguire, 2016; Maguire
& Mullally, 2013), and more specific aspects of visuospatial
Procesando, including perceptual richness, a sense of re-
living, and imagery content (St-Laurent, Moscovitch, &
McAndrews, 2016; Andrews-Hanna, Reidler, Sepulcro,
Poulin, & Buckner, 2010; Calle. jacques, Conway, Lowder, &
Cabeza, 2010).
The cognitive map theory, por ejemplo, posits that the
hippocampus specifically supports flexible, allocentric
representations of spatial relationships (O’Keefe & Nadel,
1978). A diferencia de, the scene construction theory (ver
also the emergent memory account; Graham et al.,
2010) proposes that the anterior hippocampus con-
structs models of the world in the form of spatially coher-
ent scenes (Dalton & Maguire, 2017; Zeidman & Maguire,
© 2018 by Massachusetts Institute of Technology. Published under
a Creative Commons Attribution 4.0 no portado (CC POR 4.0) licencia.
Revista de neurociencia cognitiva 30:12, páginas. 1821–1845
doi:10.1162/jocn_a_01315
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
oh
C
norte
_
a
_
0
1
3
1
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
2016; Maguire & Mullally, 2013; Hassabis & Maguire,
2007). A scene in this context is a specific type of visual
image that represents a naturalistic 3-D space typically
populated by objects and that is viewed from an ego-
centric perspective. The construction of scene imagery
involves associative processing and binding, pero el
scene construction theory asserts that the hippocampus
is specifically required to perform these functions in the
service of creating scene representations (Maguire &
Mullally, 2013). The difficulty with theories such as cog-
nitive map and scene construction is that they do not
appear to be able to explain why VPA learning is invari-
ably compromised following hippocampal damage.
On the face of it, another hippocampal theory does
seem to account for the VPA findings. The relational the-
ory suggests that the hippocampus makes associations
between any elements, regardless of whether or not
space or scenes are involved (Konkel & cohen, 2009;
cohen & Eichenbaum, 1993). This generic associative
process could account for the creation of an association
between two unrelated words in the VPA task, while also
explaining the involvement of the hippocampus in visuo-
spatial tasks and the combining of individual elements
into a coherent memory or the recombination of different
elements from past experiences to simulate the future
(Roberts, Schacter, & Addis, 2018; Calle. jacques, Carpintero,
Szpunar & Schacter, 2018; Thakral, Benoit, & Schacter,
2017; Moscovitch, Cabeza, Winocur, & Nadel, 2016; Schacter
et al., 2012). Sin embargo, a purely associative account of
hippocampal function is not completely satisfactory,
given that patients with hippocampal damage retain an
ability to form associations in some circumstances. Para
ejemplo, intact performance has been reported for
Yes/No and forced choice recognition of both intraitem
associations and associations between items of the same
kind (Mayes et al., 2004), when creating basic associa-
tions in probabilistic learning (Kumaran et al., 2007;
Knowlton, Mangels, & Squire, 1996), in the rapid learn-
ing of arbitrary stimulus–response contingencies (Henson
et al., 2017), and in other contexts (see Clark & Maguire,
2016; Mullally & Maguire, 2014).
Resolving the tension among hippocampal theories
concerning the VPA could be important for leveraging a
fuller understanding of hippocampal function. In taking
this issue forward, it is worthwhile first to step back.
Examination of the words used in typical VPA tests shows
the vast majority are high imagery concrete words. Él
could be that people use visual imagery when processing
the word pairs (Maguire & Mullally, 2013). This specula-
tion has recently received indirect support from the finding
that patients with hippocampal amnesia used significantly
fewer high imagery words in their narrative descriptions of
real and imagined events (Hilverman, Cocinar, & Duff, 2017),
suggesting a potential link between verbal processing
and visual imagery.
Actualmente, por lo tanto, standardized VPA tests may be
conflating associative processes and imageability. Patients
with hippocampal damage are reportedly unable to imag-
ine fictitious and future scenes in addition to their well-
reported memory deficits (Schacter et al., 2012; Carrera,
Keane, & Verfaellie, 2011; Hassabis, Kumaran, Vann, &
Maguire, 2007). It would, por lo tanto, follow that their im-
poverished scene imagery ability may place them at a
disadvantage for processing high imagery concrete words.
One way to deal with the conflation of visual imagery
and binding is to examine very low imagery (abstract)
word pairs, which would assess binding outside the
realm of imagery. Sin embargo, abstract word pairs rarely fea-
ture in VPA tests used with patients or in neuroimaging
experimentos.
Además, different types of high imagery words are
not distinguished in VPA tests, with the majority of words
representing single objects. Sin embargo, the scene con-
struction theory links the anterior hippocampus specifi-
cally with constructing visual imagery of scenes (Dalton
& Maguire, 2017; Zeidman & Maguire, 2016). Por el contrario,
the processing of single objects is usually associated with
perirhinal and lateral occipital cortices (Murray, Bussey,
& Saksida, 2007; Malach et al., 1995). It could therefore
be that a scene word (p.ej., forest) in a pair engages the
hippocampus (via scene imagery) and not because of
binding or visual imagery in general. It has also been sug-
gested that even where each word in a pair denotes an
object (p.ej., cat–table), this might elicit imagery of both
objects together in a scene, and it is the generation of this
scene imagery that recruits the hippocampus (clark &
Maguire, 2016; Maguire & Mullally, 2013). Como consecuencia, si
visual imagery does play a role in the hippocampal depen-
dence of the VPA task, then it will be important to establish
not only whether visual imagery or binding is more relevant
but also the type of visual imagery being used.
To determine why VPA engages the hippocampus, nosotros
devised an fMRI task with three types of word pairs: dónde
both words in a pair denoted “Scenes,” where both words
represented single “Objects,” and where both words were
very low imagery “Abstract” words. This allowed us to sep-
arate imageability from binding and to examine different
types of imagery. Of particular interest were the Object
word pairs because we wanted to ascertain whether they
were processed using scene or object imagery. Para todos
word pairs, our main interest was during their initial pre-
sentation, when any imagery would likely be evoked.
Además, we conducted recognition memory tests
after scanning to investigate whether the patterns of
(hippocampal) activity were affected by whether pairs were
successfully encoded or not. Although the VPA memory
test used with patients typically involves cued recall, el
adaptation of the VPA task for fMRI necessitated the use
of recognition memory tests. This is because performing
a cued recall test for 135 word pairs that were each seen
only once is too difficult even for healthy participants.
We note that recognition memory for word pairs is not
often tested in patients, and in the few studies where it
has been examined, the results are mixed, with some
1822
Revista de neurociencia cognitiva
Volumen 30, Número 12
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
oh
C
norte
_
a
_
0
1
3
1
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
studies finding a deficit and others a preservation of per-
rendimiento (Mayes et al., 2001; Haist, Shimamura, & Squire,
1992). Sin embargo, we expected that the use of a recog-
nition memory test instead of cued recall would have
limited impact on the patterns of brain activity in this
study because we assessed brain activity during the initial
presentation of the word pairs and not during memory
retrieval. Finalmente, given that people vary in their use of
mental imagery (McAvinue & Robertson, 2007; Kosslyn,
Brunn, Cave, & Wallach, 1984; Marks, 1973), we also tested
groups of high, mid, and low imagery users to assess
whether this influenced hippocampal engagement during
VPA encoding.
In line with the scene construction theory, we hypoth-
esized that anterior hippocampal activity would be ap-
parent for Scene words pairs, given the likely evocation
of scene imagery. We also predicted that anterior hippo-
campal activity would be increased for Object word
pairs and that this would be best explained by the use
of scene imagery. Además, we expected that the effect
of scene imagery use on the hippocampus would be
most apparent in high imagery users. Por el contrario, nosotros
predicted that Abstract words pairs would engage areas
outside the hippocampus, even when only subsequently
remembered pairs were considered.
MÉTODOS
Participantes
Forty-five individuals took part in the fMRI study. All were
saludable, right-handed, and had normal or corrected-
to-normal vision. Given the verbal nature of the task,
all participants were highly proficient in English, had
English as their first language, and were educated in
English throughout their school years. Each partici-
pant gave written informed consent. The study was ap-
proved by the University College London Research Ethics
Mesa 1. Characteristics of the Participant Groups
Committee. Participants were recruited on the basis of
their scores on the Vividness of Visual Imagery Question-
naire ( VVIQ; Marks, 1973). The VVIQ is a widely used
self-report questionnaire, which asks participants to bring
images to mind and rate them on a 5-point scale as to their
vividness (anchored at 1 = Perfectly clear and as vivid as
normal vision and 5 = No image at all, you only “know”
that you are thinking of the object). Por lo tanto, a high
score on the VVIQ corresponds to low use of visual imag-
ery. The validity of the VVIQ has been demonstrated in
numerous ways. Por ejemplo, experimental studies have
found that high visualizers were able to match two pic-
tures more quickly than low visualizers when the first pic-
ture had to be retained as a mental image over a 20-sec
período (Gur & Hilgard, 1975). Además, significant cor-
relations between the VVIQ and the Betts’ Questionnaire
Upon Mental Imagery (another widely used imagery
cuestionario; Sheehan, 1967) have also been reported
(Campos & Pérez-Fabello, 2005; Burton & Fogarty, 2003).
Our fMRI participants comprised three subgroups (norte =
15 in each), low imagery users, mid imagery users, y
high imagery users. Initially, 184 people completed the
VVIQ. Fifteen of the highest and 15 of the lowest scorers
made up the low and high imagery groups. Otro 15
mid scorers served as the mid imagery group. We acknowl-
edge that these groups are relatively small for an fMRI
estudiar, but we were nevertheless interested to see whether
any differences would be observed. The groups did not
differ significantly on age, género, years of education,
and general intellect. Mesa 1 provides details of the three
grupos.
Estímulos
To ensure that any fMRI differences were due to our
imagery manipulation and not other word properties,
the word conditions were highly matched. Six hundred
Imagery Group
pag
Bajo
Mid
High
Low vs. Mid
Low vs. High Mid vs. High
Age, años
23.07 (2.31)
21.87 (2.20)
23.93 (5.26)
No. of male participants
6 (40.0%)
7 (46.67%)
8 (53.33%)
Years of education
16.0 (1.89)
15.8 (1.61)
16.0 (2.33)
Matrix Reasoning
12.47 (2.26)
11.47 (2.17)
12.07 (3.61)
TOPF
FSIQ
VCI
54.93 (5.13)
57.47 (5.49)
53.0 (9.47)
110.13 (5.48)
111.97 (6.13)
110.22 (5.99)
108.93 (5.32)
110.81 (6.18)
108.75 (6.0)
.16
.71
.76
.23
.20
.39
.38
.57
.46
1.0
.72
.49
.97
.93
.18
.72
.79
.59
.13
.44
.36
VVIQ mean score
3.08 (0.45)
2.15 (0.17)
1.51 (0.25)
<.001
<.001
<.001
Means (SDs). Two-tailed p values for t tests (χ2 test for the number of male participants). General intellect was measured using the Matrix Reasoning
subtest (scaled scores) of the Wechsler Adult Intelligence Scale-IV ( Wechsler, 2008) and the Test of Premorbid Function (TOPF; Wechsler, 2011),
provided an estimate of Full Scale IQ (FSIQ) and a Verbal Comprehension Index ( VCI).
Clark, Kim, and Maguire
1823
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
fifty-four words were required for the study—218 Scene
words, 218 Object words, and 218 Abstract words. Words
were initially sourced from databases created by Brysbaert
and colleagues, which provided ratings for concreteness,
word frequency, age of acquisition, valence, and arousal
(Brysbaert, Warriner, & Kuperman, 2014; van Heuven,
Mandera, Keuleers, & Brysbaert, 2014; Warriner, Kuperman,
& Brysbaert, 2013; Kuperman, Stadthagen-Gonzalez, &
Brysbaert, 2012). It was important to control for valence
and arousal given reports of higher emotional ratings
for abstract words, which could influence fMRI activity
(Vigliocco et al., 2014; Kousta, Vigliocco, Vinson, Andrews,
& Del Campo, 2011). We also used data from the English
Lexicon Project (Balota et al., 2007) to provide lexical in-
formation about each word—word length, number of
phonemes, number of syllables, number of orthographic
neighbors, and number of phonological and phonographic
neighbors with and without homophones.
To verify that each word induced the expected imagery
(i.e., scene imagery, object imagery, or very little/no
imagery for the abstract words), we collected two further
ratings for each word. First, a rating of imageability to
ensure that Scene and Object words were not only con-
crete but also highly imageable (although concreteness
and imageability are often interchanged, and although
they are highly related constructs, they are not the same;
Paivio, Yuille, & Madigan, 1968) and, additionally, that
Abstract words were low on imageability. Second, a
decision was elicited about the type of imagery the word
brought to mind. This was in response to the following
instruction:
“If you had an image we would like you to classify it
as either a ‘scene’ or an ‘object’. A scene is an image
in your mind that has a sense of space; that you
could step into or operate within. An object on the
other hand is more of an isolated image, without
additional background imagery. It is also likely that for
a number of words you will experience very little or
no imagery—please do select this option if this is
the case.”
These ratings were collected from 119 participants in
total using Amazon Mechanical Turk’s crowdsourcing
Web site, following the procedures used by Brysbaert
and colleagues for the databases described above. Words
were classified as a Scene or Object word when there was
a minimum of 70% agreement on the type of imagery
brought to mind, and the mean imageability rating was
greater than 3.5 (out of 5). For Abstract words, the mean
imageability had to be less than or equal to 2. An overview
of the word properties is shown in Table 2. This also in-
cludes summary comparison statistics. A list of the words
in each category can be found in Appendices A–C and at
www.fil.ion.ucl.ac.uk/Maguire/Clark_et_al_2018_Scene_
Words.pdf, www.fil.ion.ucl.ac.uk/Maguire/Clark_et_al_
2018_Object_Words.pdf, www.fil.ion.ucl.ac.uk/Maguire/
Clark_et_al_2018_Abstract_Words.pdf.
Scene, Object, and Abstract words were matched on
13 of the 16 measures. Scene and Object words were
matched on all 16 measures, whereas Abstract words,
as expected, were less concrete and less imageable than
Scene and Object words and had a higher age of acqui-
sition, as is normal for abstract words (Kuperman et al.,
2012; Stadthagen-Gonzalez & Davis, 2006). As well as
being matched at the overall word type level as shown
on Table 2, within each word type, words were assigned
to one of four lists (word pairs, single words, catch trials,
or postscan memory test lures), and all lists were matched
on all measures.
Experimental Design and Task
The fMRI task consisted of two elements, the main task
and catch trials. The latter were included to provide an
active response element and to encourage concentration
during the experiment. To match the WMS-IV Verbal
Paired Associate Test ( Wechsler, 2009), each stimulus
was presented for 4 sec. This was followed by a jittered
baseline (a central fixation cross) for between 2 and 5 sec,
which aided concentration by reducing the predictability
of stimulus presentation (Figure 1D). The scanning session
was split into four runs, three runs containing 80 trials
lasting 10 min each and a final run of 78 trials lasting 9 min
45 sec. Trials were presented randomly for each partici-
pant with no restrictions on what could precede or follow
each trial.
Unknown to participants, there were six categories of
stimuli—high imagery Scene words, high imagery Object
words, and very low imagery Abstract words, shown either
in pairs of the same word type (Figure 1A) or as single
words (Figure 1B). To equalize visual presentation be-
tween the word pairs and the single words, the latter
were presented with a random letter string that did not
follow the rules of the English language and did not re-
semble real words (Figure 1B). The average, minimum,
and maximum length of the letter strings was matched
to the real words. Letter strings could either be presented
at the top or the bottom of the screen. There were 45 tri-
als of each condition, with each word shown only once to
the participant. Our prime interest was in the word pair
conditions and, in particular, the Object word pairs, as
these related directly to our research question. The single-
word conditions were included for the purposes of specific
analyses, which are detailed in the Results section.
Participants were asked to try to commit the real words
to memory for later memory tests and were specifically
instructed that they would be asked to recall the pairs
of real words as pairs. They were explicitly told they
would not need to remember the random letter strings.
No further instructions about how to memorize the
stimuli were given (i.e., we did not tell participants to
use any particular strategy).
Participants were told that, occasionally, there would
be catch trials where they had to indicate, using a button
1824
Journal of Cognitive Neuroscience
Volume 30, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 2. Properties of Each Word Type
Type of Word
p
Scene
Object
Abstract
Scene vs.
Object
Scene vs.
Abstract
Object vs.
Abstract
Word Property
Lexical Criteria
No. of lettersa
No. of phonemesa
No. of syllablesa
6.73 (1.99)
6.64 (1.91)
6.75 (1.97)
5.61 (1.89)
5.41 (1.71)
5.66 (1.72)
2.09 (0.85)
2.06 (0.79)
2.16 (0.77)
No. of orthographic neighborsa
2.57 (4.97)
3.09 (4.96)
2.35 (4.33)
No. of phonological neighborsa
6.28 (11.71)
7.61 (11.80)
6.12 (11.31)
No. of phonological neighbors
(including homophones)a
6.88 (12.59)
8.13 (12.48)
6.66 (11.86)
No. of phonographic neighborsa
1.53 (3.51)
1.78 (3.48)
1.46 (3.21)
No. of phonographic neighbors
(including homophones)a
Word frequency: Zipf b
Age of acquisitionc
1.61 (3.67)
1.96 (3.63)
1.49 (3.36)
3.90 (0.71)
3.80 (0.61)
3.88 (0.82)
7.69 (2.14)
7.40 (2.12)
9.78 (2.46)
Emotional Constructs
Valenced
No. of positive wordsd,e
Hedonic valenced,f
Arousald
Imagery
Concretenessg
Imageabilityh
5.68 (1.08)
5.63 (1.02)
5.58 (1.12)
171 (78.44%)
173 (79.6%)
167 (76.61%)
1.07 (0.69)
0.98 (0.68)
1.04 (0.70)
4.07 (0.96)
3.99 (0.87)
4.04 (0.71)
4.65 (0.22)
4.68 (0.22)
1.83 (0.29)
4.38 (0.29)
4.41 (0.32)
1.53 (0.20)
.64
.25
.68
.28
.23
.30
.45
.32
.12
.15
.63
.81
.18
.34
.11
.31
.90
.77
.38
.62
.89
.85
.84
.72
.77
.55
.13
.18
.10
.18
.21
.32
.16
.27
<.001
<.001
.34
.65
.69
.73
.61
.49
.34
.46
<.001
<.001
<.001
<.001
Means (SDs). Two-tailed p values for t tests (χ2 test for the number of positive words). Note that each comparison was assessed separately to provide
a greater opportunity for any differences between conditions to be identified.
aFrom the English Lexion Project (Balota et al., 2007: exlexicon.wustl.edu).
bFrom van Heuven et al. (2014). The Zipf scale is a standardized measure of word frequency using a logarithmic scale. Values go from 1 (low-
frequency words) to 6 (high-frequency words).
cFrom Kuperman et al. (2012).
dFrom Warriner et al. (2013).
ePositive words were those that had a valence score greater than or equal to 5.
fHedonic valence is the distance from neutrality (i.e., from 5), regardless of being positive or negative, as per Vigliocco et al. (2014).
gFrom Brysbaert et al. (2014).
hCollected for the current study as detailed in the Methods.
press, if they saw a real word presented with a “pseudo-
word” (Figure 1C). The participants were informed that
they were not required to remember the real word or
the pseudoword presented in these catch trials. A pseudo-
word is a combination of letters that resembles a real
English word and follows the rules of the English language
but is not an actual real word. Pseudowords were gener-
ated using the English Lexicon Project (Balota et al.,
2007) and were paired with Scene, Object, or Abstract
words. They were presented at either the top or the
bottom of the screen to ensure that participants attended
to both. The number of letters and orthographic neighbors
of the pseudowords were matched to all of the real word
conditions and across the three pseudoword groups (all
Clark, Kim, and Maguire
1825
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
had seen it in the scanner (old) or not (new). Following
this, they rated their confidence in their answer on a
3-point scale—high confidence, low confidence, or guess-
ing. Any trials where a participant correctly responded
“old” and then indicated they were guessing were excluded
from subsequent analyses.
After the item memory test, memory for the pairs of
words was examined. This associative memory test pre-
sented all of the 135 word pairs shown to participants
in the scanner and an additional 66 lure pairs (22 of each
type), one pair at a time, for up to 5 sec. The word pairs
were presented in a different random order for each
participant. The lure pairs were constructed from the
single words that were presented to the participants in
the scanner. Therefore, the participants had seen all of
the words presented to them in the associative recog-
nition memory test, but not all were previously in pairs,
specifically testing whether the participants could remem-
ber the correct associations. Participants were asked to
indicate whether they saw that exact word pair presented
to them in the scanner (old) or not (new). They were
explicitly told that some pairs would be constructed from
the single words they had seen during scanning and not
to make judgments solely on individual words, but to
consider the pair itself. Confidence ratings were obtained
in the same way as for the item memory test, and trials
where a participant correctly responded “old” and then
indicated they were guessing were excluded from sub-
sequent analyses.
Debriefing
On completion of the memory tests, participants were
asked about their strategies for processing the words
while they were in the scanner. At this point, the partic-
ipants were told about the three different types of words
presented to them—Scenes, Objects, and Abstract. For
each word type and separately for single words and word
pairs, participants were presented with reminders of the
words. They were then asked to choose from a list of
options as to which strategy best reflected how they
processed that word type. Options included the follow-
ing: “I had a visual image of a scene related to this type
of single word” (scene imagery for single words), “I had
a visual image of the two entities that the words repre-
sented within a single visual scene” (scene imagery for
word pairs), “I had a visual image of a single entity (e.g.,
one specific object) for a word with no other background
imagery” (object imagery), “I read each word without
forming any visual imagery at all” (no imagery).
Statistical Analyses of the Behavioral Data
Stimuli Creation and Participant Group Comparisons
Comparisons between word conditions and between the
participant groups were performed using independent
samples t tests for continuous variables and χ2 tests
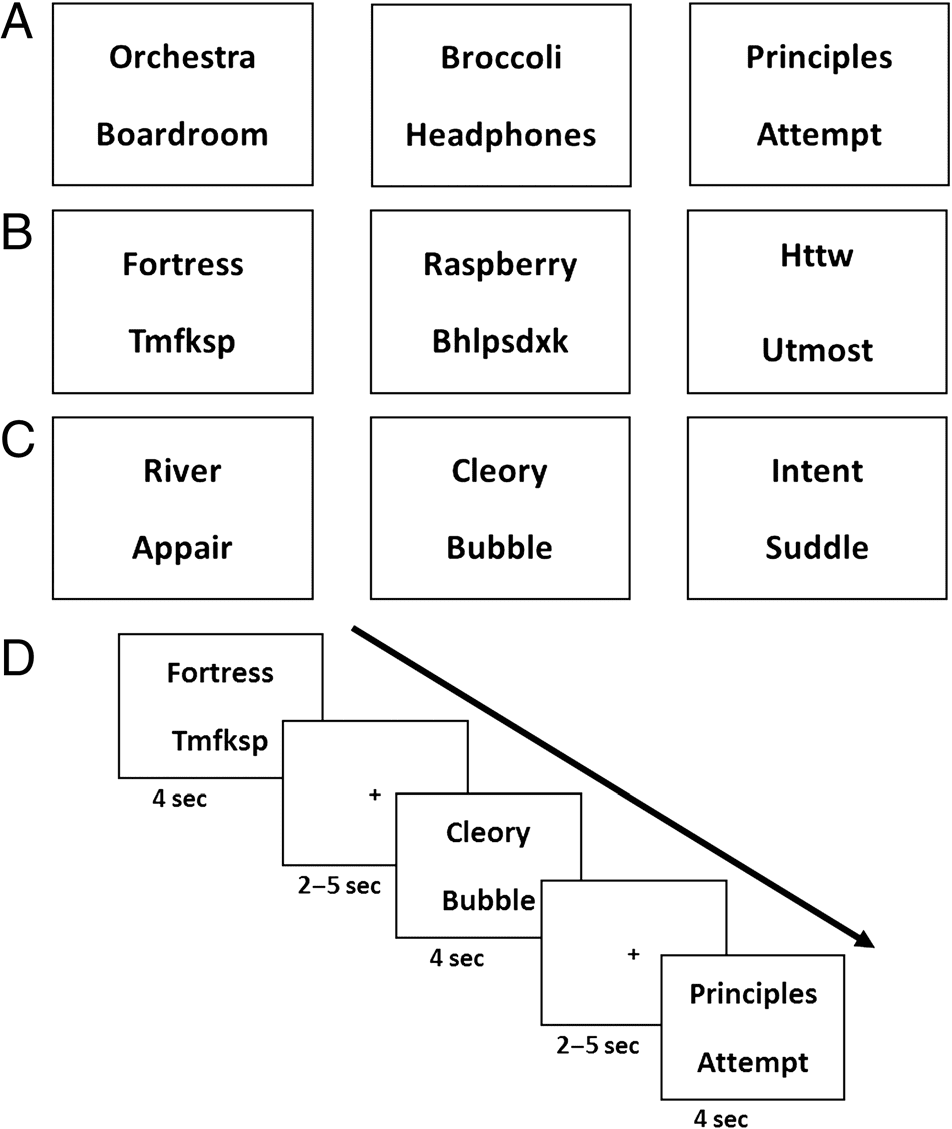
Figure 1. Example stimuli and trial timeline. (A) Examples of stimuli
from each of the word types in the order of (from left to right) Scene
word pair, Object word pair, and Abstract word pair. (B) Examples of
single word trials in the order of (from left to right) Scene single word,
Object single word, and Abstract single word. Single words were
shown with random letter strings (which could be presented at either
the top or the bottom) to be similar to the visual presentation of the word
pairs. (C) Examples of catch trials, where a real word was presented
with a pseudoword, which could be presented as either the top or
bottom word. (D) Example timeline of several trials.
ps > .3). Además, across the pseudoword groups, nosotros
matched the accuracy of pseudoword identification (todo
ps > .6) as reported in the English Lexicon Project (Votar
et al., 2007). Forty-eight catch trials were presented over
the course of the experiment, 16 trials with each of the
word types, ranging between 10 y 15 in each of the four
carreras. Catch trials were pseudorandomly presented to
ensure regular presentation, but not in a predictable man-
ner. Feedback was provided at the end of each scanning
run as to the number of correctly identified pseudowords
and incorrectly identified real words.
Postscan Recognition Memory Tests
Following scanning, participants had two recognition
memory tests. The first was an item recognition memory
test for all 405 words presented during scanning (45 palabras
for each of three single word types and 90 words for each
of three paired word types) and a further 201 foils (67 de
each word type). Each word was presented on its own in
the center of the screen for up to 5 segundo. Words were pre-
sented randomly in a different order for each participant.
Participants had to indicate for each word whether they
1826
Revista de neurociencia cognitiva
Volumen 30, Número 12
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
oh
C
norte
_
a
_
0
1
3
1
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
for categorical variables. An alpha level of p > .05 era
used to determine that the stimuli/groups were matched.
Note that each comparison was assessed separately
(using t tests or χ2 tests) to provide a greater oppor-
tunity for any differences between conditions to be
identificado.
Main Study
Both within- and between-participant designs were used.
The majority of analyses followed a within-participant
diseño, with all participants seeing all word conditions. En
addition, participants were split into three groups depen-
dent on their VVIQ score allowing for between-participant
analyses to be performed.
All data were assessed for outliers, defined as values
that were at least 2 standard deviations away from the
significar. If an outlier was identified, then the participant
was removed from the analysis in question (and this is
explicitly noted in the Results section). Memory per-
formance for each word condition was compared with
chance level (50%) using one-sample t tests. Para todos
within-participant analyses, when comparing across three
condiciones, repeated-measures ANOVAs with follow-up
paired t tests were used, and for comparison across two
condiciones, paired t tests were utilized. For between-
participant analyses, a one-way ANOVA was performed
with follow-up independent samples t tests.
All ANOVAs were subjected to Greenhouse–Geisser
adjustment to the degrees of freedom if Mauchly’s sphe-
ricity test identified that sphericity had been violated. Para
all statistical tests, alpha was set at .05. Effect sizes are
reported following significant results as Cohen’s d for
one sample and independent sample t tests, eta-squared
for repeated-measures ANOVA, and Cohen’s d for repeated
measures (drm) for paired samples t tests (Lakens, 2013).
All analyses were performed in IBM SPSS statistics v22.
Scanning Parameters and Data Preprocessing
T2*-weighted echo-planar images (EPI) were acquired
using a 3T Siemens Trio scanner (Siemens Healthcare)
with a 32-channel head coil. fMRI data were acquired over
four scanning runs using scanning parameters optimized
for reducing susceptibility-induced signal loss in the
medial-temporal lobe: 48 transverse slices angled at −30°,
repetition time = 3.36 segundo, tiempo de eco (EL) = 30 mseg, res-
olution = 3 × 3 × 3 mm, matrix size = 64 × 74, z-shim
gradient moment of −0.4 mT/m msec (Weiskopf, Hutton,
Josephs, & Deichmann, 2006). Fieldmaps were acquired
with a standard manufacturer’s double-echo gradient-
echo field map sequence (short TE = 10 mseg, largo
TE = 12.46 mseg, 64 axial slices with 2-mm thickness
and 1-mm gap yielding whole-brain coverage; in-plane
resolution 3 × 3 mm). After the functional scans, a 3D
MDEFT structural scan was obtained with 1-mm isotropic
resolution (Deichmann, Schwarzbauer, & Tornero, 2004).
Preprocessing of data was performed using SPM12
(www.fil.ion.ucl.ac.uk/spm). The output of the SPM image
realignment protocol showed that head motion was low
(significar [Dakota del Sur] in millimeters: x = 0.51 [0.32], y = 1.29
[0.33], z = 1.64 [0.80]; significar [Dakota del Sur] in degrees: pitch =
0.03 [0.03], roll = 0.01 [0.01], yaw = 0.01 [0.01]) y era
smaller than the voxel size. Functional images were co-
registered to the structural image and then realigned and
unwarped using field maps. The participant’s structural im-
age was segmented and spatially normalized to a standard
EPI template in MNI space with a voxel size of 2 × 2 × 2 mm
and the normalization parameters were then applied to
the functional data. For the univariate analyses, the func-
tional data were smoothed using an 8-mm FWHM Gaussian
kernel. In line with published representational similarity
análisis (RSA) literature (p.ej., Chadwick, Jolly, Amos,
Hassabis, & Spiers, 2015; Marchette, Vass, ryan, & Epstein,
2014; Kriegeskorte, Mur, Fallar, et al., 2008), the multivariate
analyses used unsmoothed data. We used unsmoothed
data to capture neural information in the form of spatially
distributed activity across multiple voxels. Smoothing
potentially washes out the fine activity differences between
vóxeles.
Where bilateral ROI analyses were performed, el
hippocampal ROIs were manually delineated on a pre-
viously collected (norte = 36) group-averaged structural MRI
scan (1 × 1 × 1 mm) using ITK-SNAP (www.itksnap.org)
and then resampled to our functional scans (2 × 2 ×
2 mm). The anterior hippocampus was delineated using
an anatomical mask that was defined in the coronal plane
and went from the first slice where the hippocampus can
be observed in its most anterior extent until the final slice
of the uncus. In terms of structural space, this amounted
a 3616 voxels and in functional space to 481 vóxeles. El
posterior hippocampus was defined as proceeding from
the first slice following the uncus until the final slice of
observation in its most posterior extent (see Dalton,
Zeidman, Barry, williams, & Maguire, 2017, for more
details). In terms of structural space, this amounted to
4779 voxels and in functional space to 575. The whole hip-
pocampus mask combined the anterior and posterior
masks and therefore contained 8395 voxels in structural
space and 1056 voxels in functional space.
fMRI Analysis: Univariate
The six experimental word conditions were Scene, Object,
and Abstract words, presented as either word pairs or
single words. As noted above, our prime interest was in
the word pair conditions and, En particular, the Object word
pares, as these related directly to our research question. Nosotros
therefore directly contrasted fMRI BOLD responses be-
tween the word pair conditions. The single-word condi-
tions were included for the purposes of specific analyses,
which are detailed in the Results section. We performed
two types of whole-brain analysis, one using all of the trials
(45 per condition) and the other using only trials where the
clark, kim, and Maguire
1827
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
oh
C
norte
_
a
_
0
1
3
1
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
items were subsequently remembered, not including
trials where the participant indicated they were guessing.
The average number of trials per condition were as
follows: Scene word pairs, 31.49 (DE = 6.25); Object
word pairs, 34.53 (DE = 5.84); Abstract word pairs,
27.42 (DE = 8.67). See Table 4 for comparisons of the
number of correct trials, not including guessing, across
the conditions.
For both analyses, the general linear model consisted of
the word condition regressors convolved with the hemo-
dynamic response function, in addition to participant-
specific movement regressors and physiological noise
regressors. The Artifact Detection Toolbox (www.nitrc.
org/projects/artifact_detect/ ) was used to identify spikes
in global brain activation, and these were entered as a
separate regressor. Participant-specific parameter esti-
mates for each regressor of interest were calculated for
each voxel. Second-level random effects analyses were then
performed using one-sample t tests on the parameter esti-
compañeros. For comparison across VVIQ imagery groups, nosotros
performed an ANOVA with follow-up independent sample
t tests. We report results at a peak-level threshold of p
less than .001 whole-brain uncorrected for our a priori
ROI—the hippocampus—and p less than .05 family-wise
error (FWE)-corrected at the voxel level elsewhere.
Además, several ROI analyses were performed on
a subset of the univariate analyses. Three ROIs were
considered—the whole hippocampus, the anterior hip-
pocampus, and the posterior hippocampus (all bilat-
eral). We used a peak-level threshold of p less than
.05 FWE-corrected at the voxel level for each mask
y, where indicated in the Results section, also a more
lenient threshold of p less than .001 uncorrected for
each mask.
fMRI Analysis: Multivariate
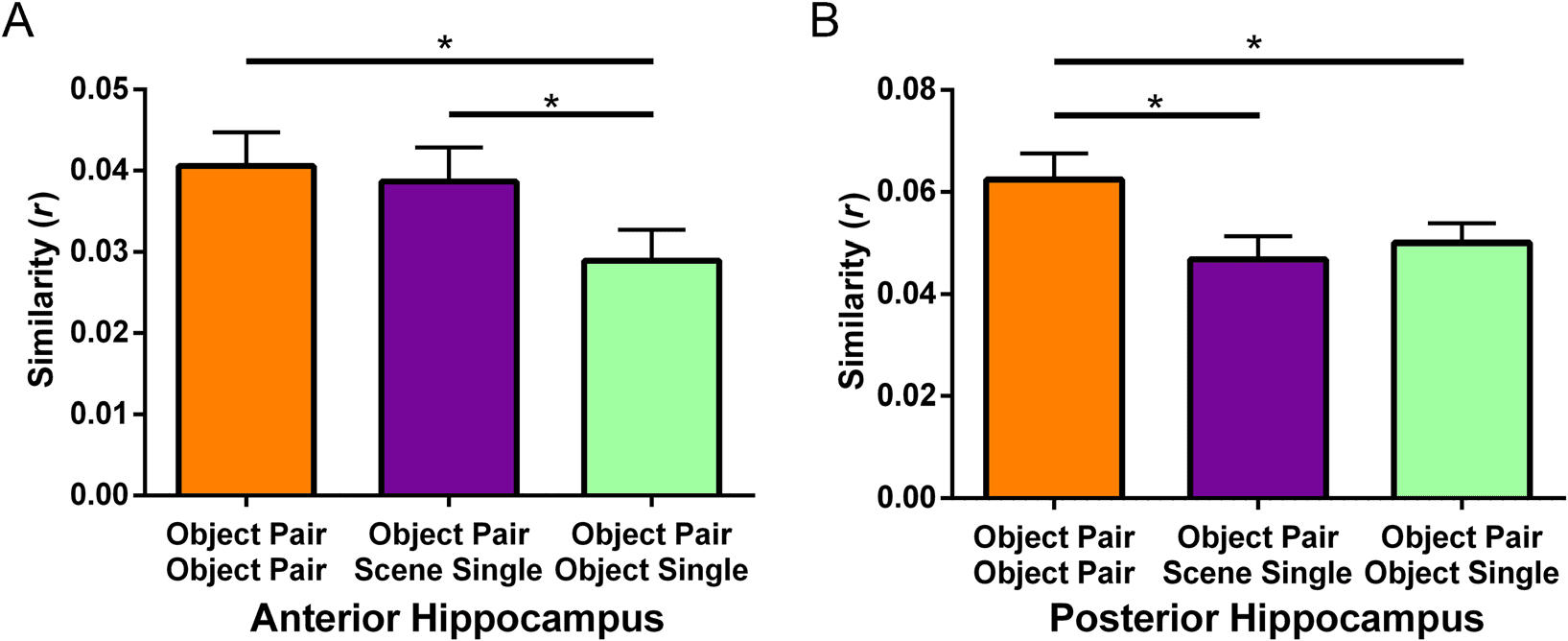
Multivoxel pattern analysis was used to test whether the
neural representations of the Object word pairs were
more similar to the Scene single words than the Object
single words when separately examining bilateral anterior
and posterior hippocampal ROIs. Para cada participante,
t statistics for each voxel in the ROI were computed for
each condition (Object word pair, Object single word,
Scene single word) and in each scanning run. The Pearson
correlation between each condition was then calculated
as a similarity measure (Object word pair/Object word pair,
Object word pair/Scene single word, Object word pair/
Object single word). The similarity measure was cross-
validated across different scanning runs to guarantee
the independence of each data set. Repeated-measures
ANOVA and paired t tests were used to compare the sim-
ilarity between conditions at the group level. This multi-
variate analysis was first applied to the data from all
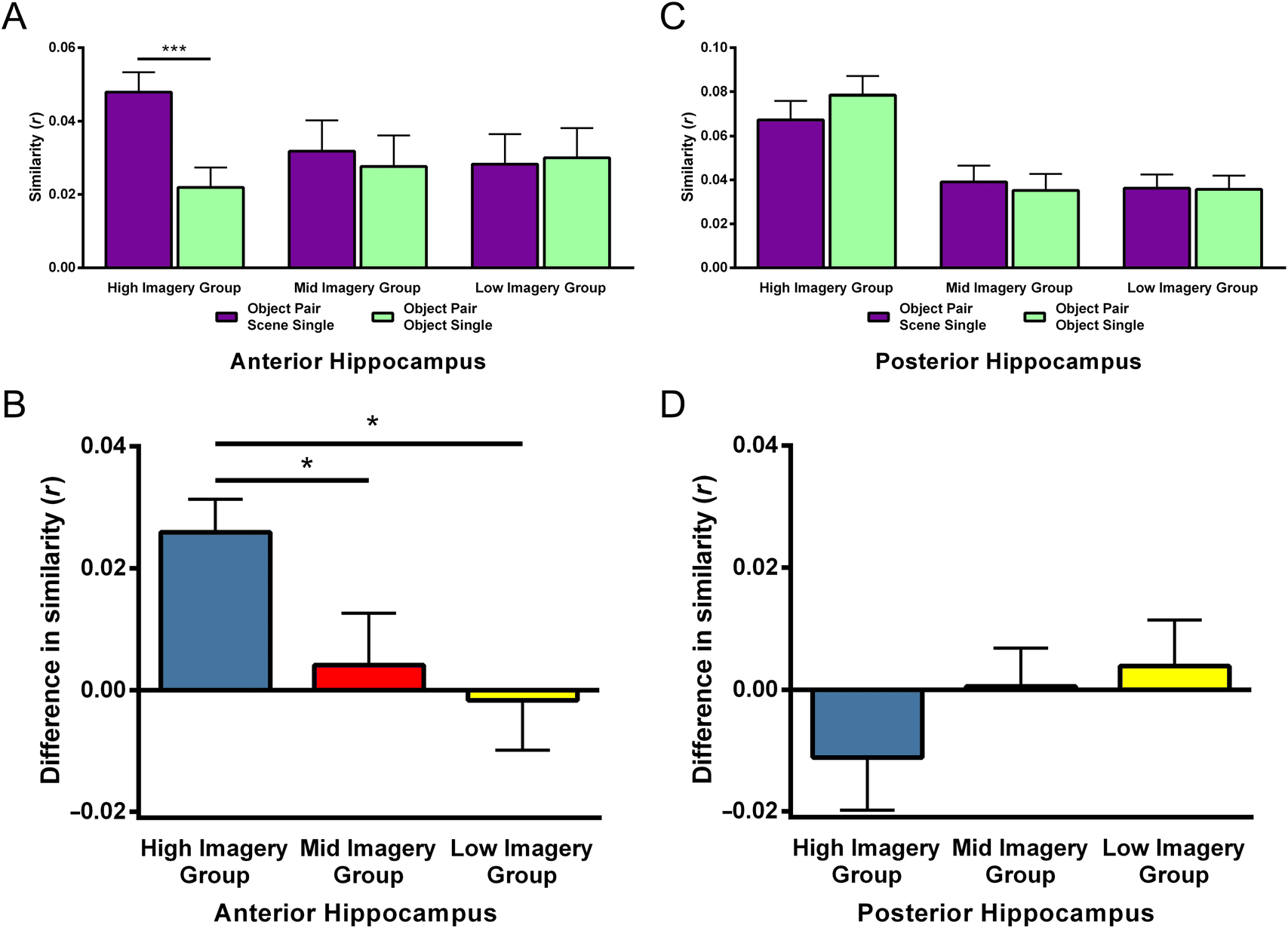
participants and then to the three subsets of participants
(bajo, mid, and high imagery users). All data were as-
sessed for outliers, defined as values that were at least
2 standard deviations away from the group mean. If an
outlier was identified, then the participant was removed
from the analysis in question (and this is explicitly noted
in the Results section).
Note that the absolute correlation of the similarity
value is expected to be low because of inherent neural
variability and the fact that a unique set of words was pre-
sented for each scanning run. Tal como, the important
measure is the comparison of the similarity value between
the conditions, not the absolute similarity value of a single
condición. The range of similarity values that we found was
entirely consistent with those reported in other studies uti-
lizing a similar representational similarity approach in a
variety of learning, memory, and navigation tasks in a wide
range of brain regions (kim, jeffery, & Maguire, 2017;
Bellmund, Deuker, Navarro Schröder, & Doeller, 2016;
Deuker, Bellmund, Navarro Schröder, & Doeller, 2016;
Schapiro, Turk-Browne, Norman, & Botvinick, 2016; Schuck,
Cai, wilson, & NVI, 2016; Chadwick et al., 2015; Hsieh &
Ranganath, 2015; Milivojevic, Vicente-Grabovetsky, &
Doeller, 2015; Hsieh, Gruber, Jenkins, & Ranganath,
2014; Staresina, Henson, Kriegeskorte, & Alink, 2012).
RESULTADOS
conductual
De término medio, participants identified 85.56% (DE = 11.52)
of the pseudowords during catch trials, showing that they
maintained concentration during the fMRI experiment.
On the postscan item memory test, Scene, Object, y
Abstract words were remembered above chance, y ahí
were no differences between the conditions (Mesa 3,
which includes the statistics). Performance on the asso-
ciative memory test also showed that Scene, Object, y
Abstract word pairs were remembered above chance
(Mesa 4, which includes the statistics). Considerando el
average performance across the four word conditions
used in the main univariate analyses (es decir., Scene word
pares, Object word pairs, Abstract word pairs, Abstracto
single words) then one participant performed below
chance. The fMRI analyses do not change whether this
participant is included or not. Comparison of memory
performance across the word types found differences in
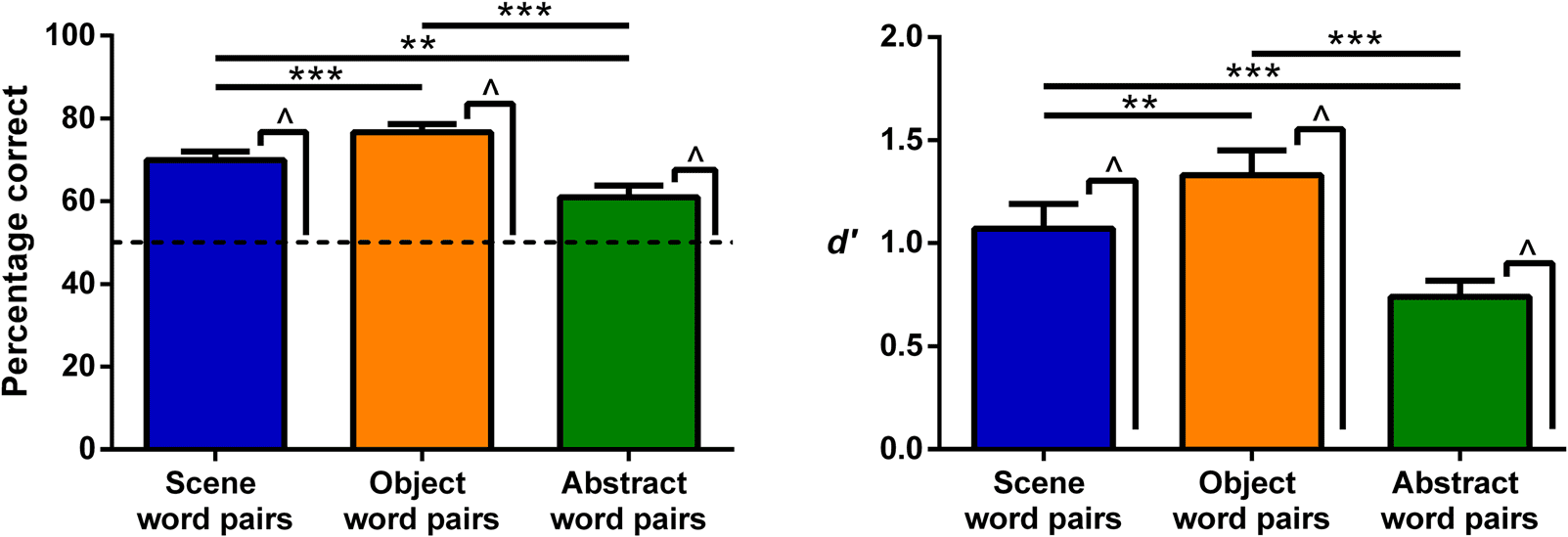
performance in line with the literature (Paivio, 1969). Ambos
types of high imagery word pairs (Scene and Object) eran
remembered better than Abstract word pairs (Cifra 2;
Mesa 4), whereas Object word pairs were remembered
better than Scene word pairs. Given that the word pair
memory lures were highly confusable with the actual word
pares (because the lure pairs were made up of the studied
values were also calculated for the word
single words), d
0
pares. Scene, Object, and Abstract word pairs all showed d
values greater than 0, representing the ability to discrimi-
nate between old and new pairs (Mesa 4). Both Scene
0 values than Abstract
and Object word pairs had greater d
0 values were greater
word pairs, and Object word pairs d
0
1828
Revista de neurociencia cognitiva
Volumen 30, Número 12
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
oh
C
norte
_
a
_
0
1
3
1
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Mesa 3. Actuación (% Correct) on the Postscan Item
Memory Test (Nonguessing Trials)
Scene
Single Words
Object
Single Words
Abstracto
Single Words
Significar
Dakota del Sur
67.41
14.93
66.37
17.71
67.61
16.06
than those for Scene word pairs (Mesa 4), showing the
same pattern as that calculated using the percentage
correcto. En general, these behavioral findings show that, de-
spite the challenging nature of the experiment with so
many stimuli, participants engaged with the task commit-
ted a good deal of information to memory and could suc-
cessfully distinguish between previously presented word
pairs and highly confusable lures.
Comparison to Chance (50%)
t(44)
7.82
6.20
7.36
pag
<.001
<.001
<.001
d
2.36
1.87
2.22
Scene single words
Object single words
Abstract single words
Comparison across the Word Types
F(1.76, 77.51)
Main effect
0.28
p
.73
fMRI
Univariate Analyses
We performed two whole-brain analyses, one using all of
the trials and another using only trials where the items
were subsequently remembered in the postscan memory
tests (the item memory test for the single word trials, the
associative memory test for the word pairs, excluding
trials where participants correctly responded “old” and
then indicated that they were guessing). The two analyses
yielded very similar results across the whole brain, even
though the analysis using only subsequently remembered
stimuli was less well powered because of the reduced
number of stimuli. Given that our interest was in the
point at which participants were initially processing the
word pairs and potentially using mental imagery to do
so, we focus on the results of the analysis using all of
Table 4. Performance (% Correct and d
0 ) on the Postscan Associative Memory Test (Nonguessing Trials)
Percent Correct
0
d
Scene
Word Pairs
Object
Word Pairs
Abstract
Word Pairs
Scene
Word Pairs
Object
Word Pairs
Abstract
Word Pairs
Mean
SD
Scene word pairs
Object word pairs
Abstract word pairs
69.98
13.88
t(44)
9.65
13.83
3.81
76.74
12.97
Comparison to Chance (50%)
p
<.001
<.001
<.001
60.94
19.27
d
2.91
4.17
1.15
1.07
0.81
t(44)
8.83
10.84
8.94
1.33
0.83
Comparison to Chance (0)
p
<.001
<.001
<.001
0.74
0.56
d
2.66
3.27
2.70
Comparison across the Word Types
Comparison across the Word Types
Main effect
Scene vs. Object
Scene vs. Abstract
Object vs. Abstract
F(1.35, 59.48)
24.21
t(44)
5.25
3.58
5.75
p
<.001
p
<.001
.001
<.001
η2
.36
drm
0.50
0.52
0.94
F(2, 88)
23.75
t(44)
3.06
4.35
6.22
p
<.001
p
.004
<.001
<.001
η2
.35
drm
0.32
0.42
0.78
Clark, Kim, and Maguire
1829
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Memory performance on the associative memory test shown by percentage correct (left) and d
significant difference from chance (for percentage correct the dashed line indicates chance at 50%, for d
differences across the word pair types: **p < .01, ***p < .001.
0 (right). Error bars are 1 SEM. ^ indicates a
0 it is 0) at p < .001. Stars show the significant
the trials. Results are also reported for the analyses using
only the remembered stimuli, which allowed us to con-
trol for any memory-related effects.
We first compared the high imagery (Scene, Object) and
very low imagery (Abstract) word pairs. All of the condi-
tions involved associative processing, and so we reasoned
that any differences we observed, particularly in hippocam-
pal engagement, would be due to the imageability of the
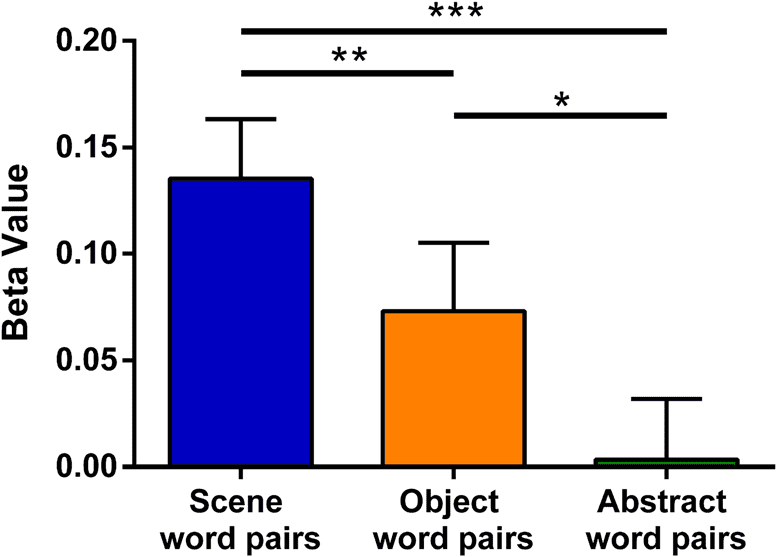
Scene and Object word pairs. As predicted, Scene word
pairs elicited greater bilateral anterior (but not posterior)
hippocampal activity compared with Abstract word pairs
(Figure 3A; see full details in Table 5A). Of note, increased
activity was also observed in bilateral parahippocampal,
fusiform, retrosplenial, and left ventromedial prefrontal
cortices. The analysis using only the remembered stimuli
showed very similar results, including for the anterior hip-
pocampus (Table 6A). The reverse contrast identified no
hippocampal engagement, but rather greater activity for
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
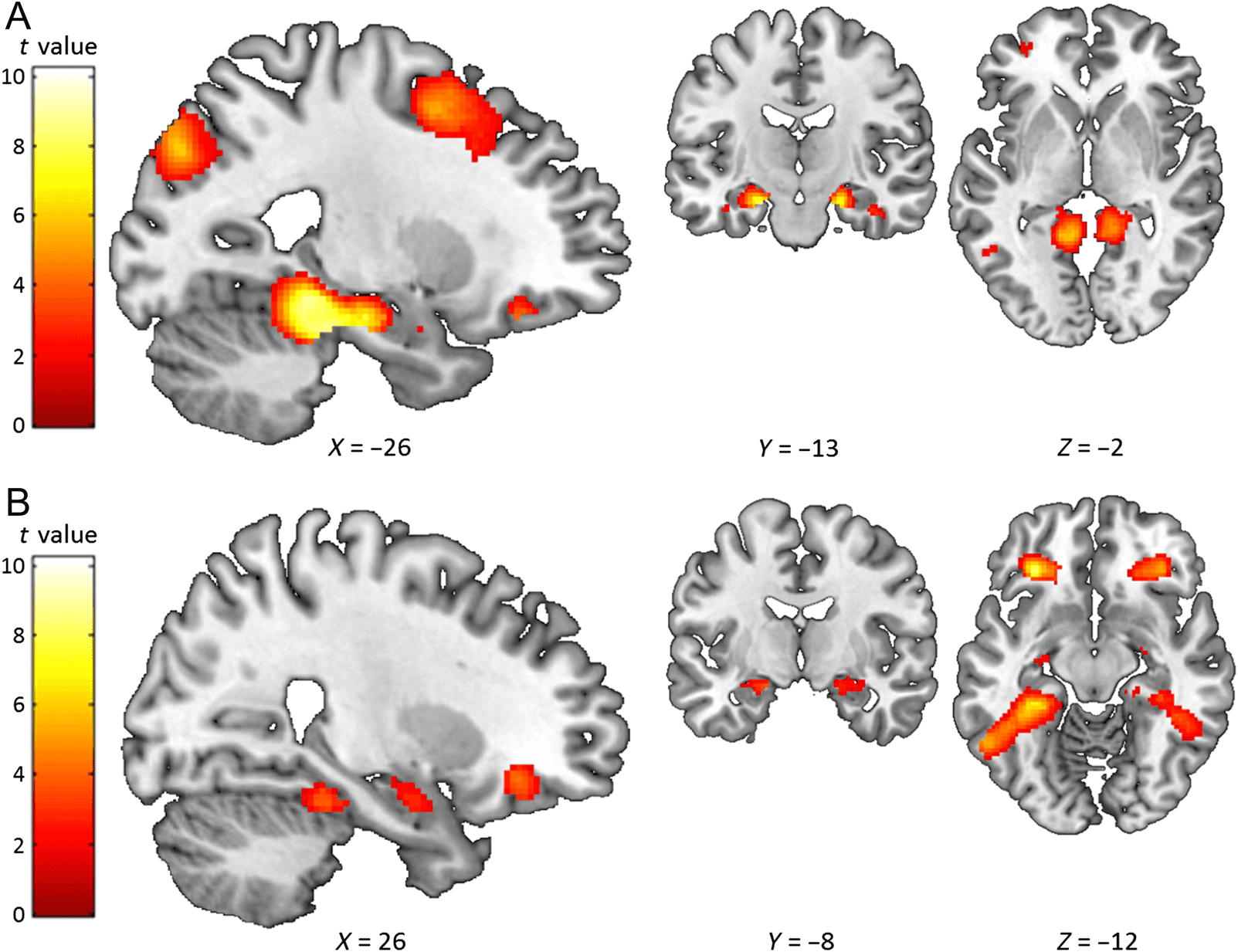
Figure 3. Comparison of high imagery Scene or Object word pairs with very low imagery Abstract word pairs. The sagittal slice is of the left
hemisphere, which is from the ch2better template brain in MRicron (Rorden & Brett, 2000; Holmes et al., 1998). The left of the image is the left side
of the brain. The colored bar indicates the t value associated with each voxel. (A) Scene word pairs > Abstract word pairs. (B) Object word pairs >
Abstract word pairs. Images are thresholded at p < .001 uncorrected for display purposes.
1830
Journal of Cognitive Neuroscience
Volume 30, Number 12
Table 5. High Imagery Word Pairs Compared with Abstract
Word Pairs
Region
Peak Voxel
Coordinates
A. Scene Word Pairs > Abstract Word Pairs
Left anterior hippocampus
Right anterior hippocampus
Left retrosplenial cortex
Left fusiform cortex
Right retrosplenial cortex
Left middle occipital cortex
Right parahippocampal cortex
Left inferior temporal cortex
Right fusiform cortex
Left ventrolateral prefrontal cortex
Right middle occipital cortex
Left middle frontal cortex
Left inferior frontal cortex
−20, −16, −20
20, −10, −20
−10, −52, 4
−22, −34, −20
10, −48, 6
−30, −74, 34
24, −34, −20
−56, −54, −10
32, −32, −14
−30, 32, −16
44, −70, 26
−26, 6, 50
−42, 32, 12
B. Object Word Pairs > Abstract Word Pairs
Left anterior hippocampus
Right anterior hippocampus
−20, −10, −18
20, −10, −18
Left ventral medial prefrontal cortex −32, 32, −14
Left fusiform cortex (extending to
−32, −34, −20
parahippocampal cortex)
Left middle occipital cortex
Right ventrolateral prefrontal cortex
Left inferior frontal cortex
−34, −80, 28
34, 32, −12
−40, 28, 14
Right fusiform gyrus (extending to
34, −32, −18
parahippocampal cortex)
t
8.79
7.58
9.15
9.03
8.73
8.49
8.40
8.03
7.72
6.51
6.48
6.19
5.74
4.45
3.98
9.45
8.88
6.17
6.05
6.05
5.72
hippocampal engagement, but rather greater activity for
Abstract word pairs in middle temporal cortex (−62, −32,
−2, T = 8) and temporal pole (−54, 10, −18, T = 7.12).
Increased anterior hippocampal activity was therefore
observed for both Scene and Object word pairs com-
pared with the very low imagery Abstract word pairs. Como
greater anterior hippocampal engagement was apparent
even when using just the remembered stimuli, it is un-
likely that this result can be explained by better associa-
tive memory or successful encoding for the high imagery
word pairs. Rather the results suggest that the anterior
hippocampal activity for word pair processing may be
related to the use of visual imagery.
All of the above contrasts involved word pairs, sugerir-
ing that associative binding per se cannot explain the
Mesa 6. Remembered High Imagery Word Pairs Compared
with Remembered Abstract Word Pairs
Region
A. Scene Word Pairs Remembered >
Abstract Word Pairs Remembered
Peak Voxel
Coordinates
t
Left hippocampus
Right hippocampus
−28, −22, −18
24, −20, −18
7.53
5.09
Left retrosplenial cortex
Left fusiform cortex (extending to
parahippocampal cortex)
−10, −50, 2
−30, −34, −14
10.20
8.21
Right retrosplenial cortex
Left middle occipital lobe
10, −48, 4
−30, −80, 40
Right fusiform cortex (extending to
26, −28, −20
parahippocampal cortex)
Left ventral medial prefrontal cortex −30, 34, −12
Right middle occipital lobe
Left inferior temporal cortex
44, −70, 28
−56, −54, −10
7.47
7.23
7.00
6.51
6.29
5.64
pag < .001 uncorrected for the hippocampus and p < .05 FWE-corrected
for the rest of the brain. Brain regions within the medial temporal lobe
were identified via visual inspection. For regions outside the medial
temporal lobe, the AAL atlas was used (Tzourio-Mazoyer et al., 2002).
B. Object Word Pairs Remembered >
Abstract Word Pairs Remembered
Left hippocampus
−32, −22, −12
5.05
Abstract word pairs in middle temporal cortex (−58, −36,
−2, T = 6.58) and temporal pole (−52, 10, −22, T = 6.16).
Object word pairs also showed greater bilateral anterior
(but not posterior) hippocampal activity compared with
the Abstract word pairs, along with engagement of bilat-
eral parahippocampal cortex, fusiform cortex, and ventro-
medial prefrontal cortex (Figura 3B; Table 5B), con
increased anterior hippocampal activity also apparent
when just the subsequently-remembered stimuli were
consideró (Table 6B). The reverse contrast identified no
Left ventral medial prefrontal cortex −30, 34, −12
Left fusiform cortex (extending to
−30, −32, −18
parahippocampal cortex)
Left middle occipital lobe
Left inferior temporal cortex
−34, −82, 30
−54, −58, −6
9.26
7.94
6.43
6.13
pag < .001 uncorrected for the hippocampus and p < .05 FWE-corrected for the rest of the brain. Brain regions within the medial temporal lobe were identified via visual inspection. For regions outside the medial temporal lobe, the AAL atlas was used (Tzourio-Mazoyer et al., 2002). Clark, Kim, and Maguire 1831 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / 3 0 1 2 1 8 2 1 1 7 8 7 9 0 1 / / j o c n _ a _ 0 1 3 1 5 p d . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 results. However, it could still be the case that binding Ab- stract word pairs does elicit increased hippocampal activity but at a lower level than Scene and Object word pairs. To address this point, we compared the Abstract word pairs with the Abstract single words, as this should reveal any hippocampal activity related to associative processing of the pairs. No hippocampal engagement was evident for the Abstract word pairs in comparison to the Abstract single words (Table 7). This was also the case when just the remembered stimuli were considered (Table 8), albeit with slightly lower power than the previous contrasts (num- ber of trials for the Abstract word pairs = 27.42 [SD = 8.67]; for the Abstract single words: 30.42 [SD = 7.23]). Given the difficulty of interpreting null results, in par- ticular when using whole-brain standard contrasts, we performed additional ROI analyses to further test whether any subthreshold hippocampal activity was evident for the Abstract word pairs compared with the Abstract single words. Using an anatomically defined bilateral whole hippocampal mask, no differences in hippocampal activity were apparent at a p < .05 FWE-corrected thresh- old at the voxel level for the mask or when a more lenient p < .001 uncorrected threshold was used. We then ex- tracted average beta values from across the whole hippocampus bilateral ROI and two additional smaller ROIs—anterior and posterior hippocampus—for the Abstract word pairs and Abstract single words. t Tests showed that there were no differences between condi- tions (whole hippocampus: t(44) = 0.16, p = .88; anterior hippocampus only: t(44) = 0.13, p = .89; posterior hippo- campus only: t(44) = 0.18, p = .86). Similar results were also observed when using just the remembered stimuli (whole hippocampus: t(44) = 1.16, p = .25; anterior hippo- campus only: t(44) = 1.36, p = .18; posterior hippocampus Table 7. Abstract Word Pairs Compared with Abstract Single Words Region Peak Voxel Coordinates Abstract Word Pairs > Abstract Single Words
Left middle temporal cortex
Left temporal pole
Left fusiform cortex
Left inferior frontal cortex
Left inferior occipital cortex
Right inferior occipital cortex
Right lingual cortex
Left precentral gyrus
−64, −36, 2
−52, 12, −16
−38, −46, −20
−54, 24, 12
−42, −68, −12
36, −74, −12
20, −82, −10
−50, 0, 48
t
8.39
6.72
6.64
6.54
6.52
6.11
5.87
5.84
pag < .001 uncorrected for the hippocampus (no activations found) and
p < .05 FWE-corrected for the rest of the brain. Brain regions within the
medial temporal lobe were identified via visual inspection. For regions
outside the medial temporal lobe, the AAL atlas was used (Tzourio-
Mazoyer et al., 2002).
Table 8. Remembered Abstract Word Pairs Compared with
Remembered Abstract Single Words
Region
Abstract Word Pairs Remembered >
Abstract Single Words Remembered
Peak Voxel
Coordinates
Left inferior frontal gyrus
Left precentral gyrus
Left middle temporal gyrus
Left inferior occipital lobe
Right inferior occipital lobe
Left supplementary motor area
Right inferior frontal gyrus
Right superior temporal pole
Right caudate nucleus
Left pallidum
−54, 14, 12
−48, −2, 48
−52, −46, 4
−38, −78, −8
34, −80, −6
−2, 4, 56
50, 10, 28
46, −30, 4
12, 10, 6
−18, 6, 0
t
9.50
8.02
8.21
7.23
7.11
6.72
6.44
6.11
6.07
6.04
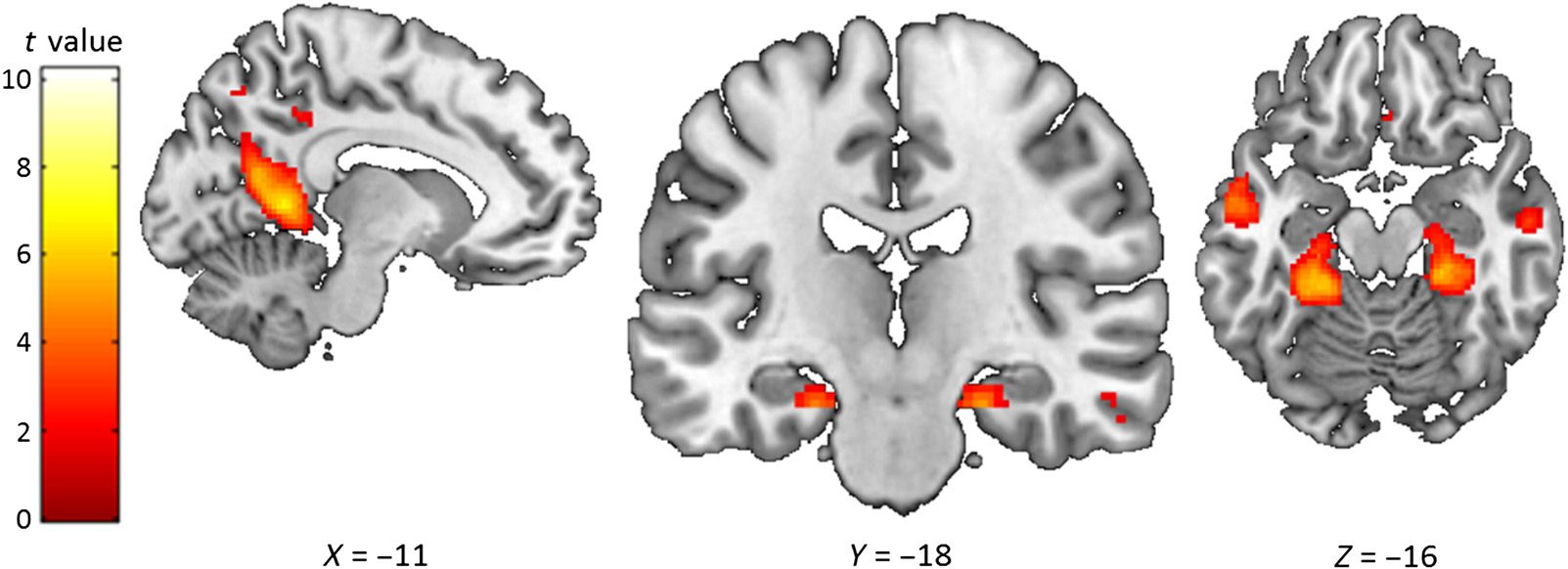
pag < .001 uncorrected for the hippocampus (no activations found) and p < .05 FWE-corrected for the rest of the brain. Brain regions within the medial temporal lobe were identified via visual inspection. For regions outside the medial temporal lobe, the AAL atlas was used (Tzourio- Mazoyer et al., 2002). only: t(44) = 0.63, p = .53). Overall, therefore, even at lenient thresholds and using an ROI approach, no hippo- campal engagement was identified for Abstract word pairs compared with the Abstract single words. Although the absence of evidence is not evidence of absence, this is in direct contrast to our findings of increased hippo- campal activity for the high imagery word pairs compared with the very low imagery Abstract word pairs. This, there- fore, lends support to the idea that the use of visual im- agery might be important for inducing hippocampal responses to word pairs. We also predicted that anterior hippocampal activity would be specifically influenced by the use of scene imagery, as opposed to visual imagery per se. The inclu- sion of both Scene and Object word pairs offered the op- portunity to test this. Scene word pairs would be expected to consistently evoke scene imagery (as both words in a pair represented scenes), whereas Object word pairs could evoke both or either object and scene imagery (e.g., object imagery by imagining the two objects without a back- ground context or scene imagery by creating a scene and placing the two objects into it), thus potentially diluting the hippocampal scene effect. Scene word pairs might there- fore activate the anterior hippocampus to a greater extent than Object word pairs. This comparison also provided an additional opportunity to contrast the effects of scene im- agery and memory performance on hippocampal activity, because Object word pairs were better remembered than the Scene word pairs. As such, if hippocampal activity could be better explained by word pair memory performance 1832 Journal of Cognitive Neuroscience Volume 30, Number 12 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / 3 0 1 2 1 8 2 1 1 7 8 7 9 0 1 / / j o c n _ a _ 0 1 3 1 5 p d . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / 3 0 1 2 1 8 2 1 1 7 8 7 9 0 1 / / j o c n _ a _ 0 1 3 1 5 p d . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 Figure 4. Brain areas more activated by Scene word pairs than Object word pairs. The sagittal slice is of the left hemisphere, which is from the ch2better template brain in MRicron (Rorden & Brett, 2000; Holmes et al., 1998). The left of the image is the left side of the brain. The colored bar indicates the t value associated with each voxel. Images are thresholded at p < .001 uncorrected for display purposes. rather than scene imagery, we would expect that Object word pairs would show greater hippocampal activity than Scene word pairs. Contrasting Scene and Object word pairs revealed that, in line with our prediction, Scene word pairs evoked greater bilateral anterior (but not posterior) hippocampal activity than the Object word pairs (Figure 4; Table 9A). Analysis using just the remembered stimuli gave sim- ilar results (Table 10A). Other areas that showed in- creased activity for the Scene word pairs included the retrosplenial and parahippocampal cortices. The reverse contrast examining what was more activated for Object word pairs compared with Scene word pairs found no evidence of hippocampal activity despite better sub- sequent memory performance for the Object word pairs (Table 9B), even when just the remembered stimuli were examined (Table 10B). It seems, therefore, that the ante- rior hippocampus may be particularly responsive to scene imagery and that increases in hippocampal activity in this task were not driven by greater memory performance. To summarize, our univariate analyses found that Scene word pairs engaged the anterior hippocampus the most, followed by the Object word pairs, with the Abstract word pairs not eliciting any significant increase in activation (Figure 5). This is what we predicted and may be sugges- tive of particular responsivity of the anterior hippocampus to scenes. Multivariate Analyses We next sought further, more direct evidence that our main condition of interest, Object word pairs, elicited hippo- campal activity via scene imagery. Given our univariate find- ings of increased anterior hippocampal activity for Scene word pairs and Object word pairs compared with Abstract word pairs and the extant literature showing the importance of the anterior hippocampus for processing scenes (e.g., Zeidman & Maguire, 2016, but see also Sheldon & Levine, 2016), we looked separately at anatomically defined bilat- eral anterior and posterior hippocampal ROIs. We then used multivariate RSA (Kriegeskorte, Mur, & Bandettini, 2008) to compare the neural patterns of activity associated with encoding Object word pairs with Scene or Object single words. We predicted that the neural representa- tions of Object word pairs in the anterior hippocampus would be more similar to Scene single words than Object single words, but that this would not be apparent in the posterior hippocampus. As our aim was to specifically investigate the contribution of different types of imagery to hippocampal activity, the scene and object single words Table 9. Scene Word Pairs Compared with Object Word Pairs Region Peak Voxel Coordinates A. Scene Word Pairs > Object Word Pairs
Left anterior hippocampus
Right anterior hippocampus
−22, −18, −20
22, −20, −20
Right retrosplenial cortex
Left retrosplenial cortex
16, −54, 20
−10, −50, 4
Left fusiform cortex (extending to
−28, −38, −12
parahippocampal cortex)
t
5.55
6.07
7.35
7.34
7.25
Right fusiform cortex (extending to
28, −26, −20
6.87
parahippocampal cortex)
Left middle temporal cortex
−58, −6, −14
5.77
B. Object Word Pairs > Scene Word Pairs
Left inferior temporal cortex
−42, −48, −16
7.16
pag < .001 uncorrected for the hippocampus and p < .05 FWE-corrected
for the rest of the brain. Brain regions within the medial temporal lobe
were identified via visual inspection. For regions outside the medial
temporal lobe, the AAL atlas was used (Tzourio-Mazoyer et al., 2002).
Clark, Kim, and Maguire
1833
Table 10. Remembered Scene Word Pairs Compared with
Remembered Object Word Pairs
Region
A. Scene Word Pairs Remembered >
Object Word Pairs Remembered
Peak Voxel
Coordinates
t
Right hippocampus
Left hippocampus
24, −20, −20
−22, −20, −18
Left retrosplenial cortex
−12, −50, 4
Right fusiform cortex (extending to
24, −28, −18
parahippocampal cortex)
Right retrosplenial cortex
10, −48, 6
Left fusiform cortex (extending to
−24, −38, −12
parahippocampal cortex)
5.18
4.26
6.74
6.49
6.46
6.37
B. Object Word Pairs Remembered >
Scene Word Pairs Remembered
Left inferior temporal gyrus
−42, −48, −16
6.12
pag < .001 uncorrected for the hippocampus and p < .05 FWE-corrected
for the rest of the brain. Brain regions within the medial temporal lobe
were identified via visual inspection. For regions outside the medial
temporal lobe, the AAL atlas was used (Tzourio-Mazoyer et al., 2002).
were chosen as comparators because they consistently
elicit either scene or object imagery respectively (see
Methods). Abstract words do not elicit much visual imag-
ery, so they were not included in the RSA analyses.
Three similarity correlations were calculated. First, the
similarity between Object word pairs and themselves,
which provided a baseline measure of similarity (i.e.,
the correlation of Object word pairs over the four runs
of the scanning experiment). The two similarities of inter-
est were the similarity between Object word pairs and
Scene single words and the similarity between Object
word pairs and Object single words. For the anterior hip-
pocampus ROI, two participants showed similarity scores
greater than 2 standard deviations away from the mean
and were removed from further analysis, leaving a sample
of 43 participants. For the posterior hippocampus ROI,
again two participants (one of whom was also excluded
from the anterior hippocampus analysis) showed similar-
ity scores greater than 2 standard deviations away from
the mean and were removed from further analysis, leav-
ing a sample of 43 participants.
For the anterior hippocampus, a repeated-measures
ANOVA found a significant difference between the three
similarities, F(2, 84) = 3.40, p = .038, η2 = .075. As pre-
dicted, the neural representations in the anterior hippo-
campus of Object word pairs were more similar to Scene
single words (Figure 6A, purple bar) than to Object single
words (Figure 6A, light green bar; t(42) = 2.09, p = .042,
drm = 0.21). In fact, representations of Object word pairs
were as similar to Scene single words as to themselves
(Figure 6A, orange bar; t(42) = 0.38, p = .71). Object
word pairs were significantly less similar to Object single
words than to themselves (t(42) = 2.54, p = .015, drm =
0.23). Of note, these results cannot be explained by sub-
sequent memory performance because Scene single
words and Object single words were remembered equally
well (t(42) = 0.68, p = .50).
For the posterior hippocampus, a repeated-measures
ANOVA also found a significant difference between the
three similarities, F(2, 84) = 4.83, p = .010, η2 = .10.
However, in contrast to the anterior hippocampus, the
neural representations in the posterior hippocampus of
Object word pairs were more similar to themselves
(Figure 6B, orange bar) than either Scene single words
(Figure 6B, purple bar; t(42) = 2.60, p = .013, drm =
0.32) or Object single words (Figure 6B, light green
bar; t(42) = 2.33, p = .025, drm = 0.26). Moreover, there
was no difference between the representations of Scene
and Object single words (t(42) = −0.71, p = .48). As
before, these results cannot be explained by subsequent
memory performance because Scene single words and
Object single words were remembered equally well (t(42) =
0.74, p = .46).
Overall, these multivariate results show that, within the
anterior hippocampus, Object word pairs were repre-
sented in a similar manner to Scene single words, but
not Object single words. On the other hand, within the
posterior hippocampus, Object word pairs were only
similar to themselves. This provides further support for
our hypothesis that Object word pairs evoke anterior
Figure 5. Comparison of each word pair condition with a fixation cross
baseline. Mean beta values extracted from a bilateral anatomical mask
of the anterior hippocampus for each of the word pair conditions
compared with the central fixation cross baseline. Error bars are 1 SEM.
A repeated-measures ANOVA showed significant differences between
the conditions, F(1.69, 74.51) = 16.06, p < .001, η2 = .27. Follow-up
paired t tests revealed significant differences between Scene word pairs
versus Abstract word pairs, t(44) = 6.46, p < .001, drm = 0.70; Scene
word pairs versus Object word pairs, t(44) = 2.97, p = .005, drm = 0.30;
Object word pairs versus Abstract word pairs, t(44) = 2.51, p = .016,
drm = 0.34. *p < .05, **p < .01, ***p < .001.
1834
Journal of Cognitive Neuroscience
Volume 30, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. The neural similarity of Object word pairs, Scene single words, and Object single words separately for the anterior and posterior
hippocampus. (A) Anterior hippocampus. (B) Posterior hippocampus. Object Pair Object Pair = the similarity between Object word pairs between
runs; Object Pair Scene Single = the similarity between Object word pairs and Scene single words; Object Pair Object Single = the similarity between
Object word pairs and Object single words. Error bars represent 1 SEM adjusted for repeated-measures (Morey, 2008). *p < .05.
(but not posterior) hippocampal activity when scene
imagery is involved.
VVIQ and the Use of Imagery
As well as examining participants in one large group, as
above, we also divided them into three groups based on
whether they reported high, mid, or low imagery ability
on the VVIQ. We found no differences in memory perfor-
mance among the groups on the word pair tasks (F < 0.4
for all contrasts). Similarly, fMRI univariate analyses involv-
ing the word pair conditions revealed no differences in
hippocampal activity. Voxel-based morphology (Ashburner,
2009; Mechelli, Price, Friston, & Ashburner, 2005; Ashburner
& Friston, 2000) showed no structural differences be-
tween the groups anywhere in the brain, including in the
hippocampus.
Interestingly, however, the imagery groups did differ
in one specific way—their strategy for processing the
Object word pairs. Although strategy use was similar
across the imagery groups for the other word conditions,
for the Object word pairs, twice as many participants
indicated using a scene imagery strategy in the high
imagery group (n = 12/15, 80%) than in the mid or low
imagery groups (n = 5/15, 33% and 6/15, 40%, respec-
tively). Comparison of scene strategy use compared with
other strategy use across the imagery groups revealed
this to be a significant difference, χ2(2) = 7.65, p = .022.
Given this clear difference in scene imagery use specif-
ically for the Object word pairs, we performed the ante-
rior and posterior hippocampus RSA analyses again for
the three imagery participant groups. We hypothesized
that, in the anterior hippocampus, the high imagery
group would represent Object word pairs in a similar
manner to Scene single words (as with our whole-group
analyses), whereas this would not be the case in the mid
or low imagery groups. For the posterior hippocampus,
on the other hand, we expected no differences between
the imagery groups. Participants with similarity values
greater than 2 standard deviations away from the mean
were again excluded. For the anterior hippocampus
ROI analyses, this resulted in one participant being re-
moved from each group. For the posterior hippocampus
two participants were excluded (both different partici-
pants to those excluded from the anterior hippocampus
analyses), one from the mid imagery group and one from
the low imagery group. Importantly, the pattern of scene
imagery strategy remained the same even after the re-
moval of these few participants (anterior hippocampus:
high imagery group, n = 11/14; mid imagery group, n =
5/14; low imagery group, n = 5/14; χ2(2) = 6.86, p = .032;
posterior hippocampus: high imagery group, n = 12/15;
mid imagery group, n = 4/14; low imagery group, n =
5/14; χ2(2) = 9.10, p = .011).
As predicted, in the anterior hippocampus for the high
imagery group, Object word pairs were more similar to
Scene single words than Object single words (Figure 7A;
t(13) = 4.63, p < .001, d = 0.78). This was not the case
for the mid or low imagery groups (t(13) = 0.472, p =
.65; t(13) = 0.20, p = .85, respectively). Of note, the inter-
action between the imagery groups was significant
F(2, 39) = 3.53, p = .039, η2 = 0.15). Inde-
(Figure 7B;
pendent samples t tests showed that the difference be-
tween the similarities was greater in the high imagery
group than in the mid and low imagery groups (t(26) =
2.09, p = .046, d = 0.79 and t(26) = 2.72, p = .011, d =
1.03, respectively). As before, these differences cannot
be explained by subsequent memory performance be-
cause all three groups showed no differences between
the Scene single and Object single words (high imagery
group: t(13) = 0.35, p = .74; mid imagery group: t(13) =
0.40, p = .69; low imagery group: t(13) = 1.18, p = .26).
Clark, Kim, and Maguire
1835
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. RSA comparisons of the three imagery groups separately for the anterior and posterior hippocampus. (A) The neural similarity of Object
word pairs, Scene single words, and Object single words in the anterior hippocampus when split by self-reported imagery use. Object Pair Scene
Single = the similarity between Object word pairs and Scene single words; Object Pair Object Single = the similarity between Object word pairs
and Object single words. (B) The difference in similarity between Object word pairs and Scene single words compared with Object words pairs and
Object single words in the imagery groups in the anterior hippocampus. (C) The neural similarity of Object word pairs, Scene single words, and Object
single words in the posterior hippocampus when split by self-reported imagery use. Object Pair Scene Single = the similarity between Object
word pairs and Scene single words; Object Pair Object Single = the similarity between Object word pairs and Object single words. (D) The difference
in similarity between Object word pairs and Scene single words compared with Object words pairs and Object single words in the imagery groups
in the posterior hippocampus. Error bars represent 1 SEM. *p < .05, ***p < .001.
For the posterior hippocampus, on the other hand,
there were no differences in similarities in any of the
imagery groups (Figure 7C; high imagery group: t(14) =
−1.29, p = .22; mid imagery group: t(13) = 0.50, p =
.63; low imagery group: t(13) = 0.084, p = .94). In line with
these findings, the interaction between the imagery groups
was also not significant (Figure 7D; F(2, 40) = 1.07, p = .35).
As before, there were no differences in subsequent mem-
ory performance between the Scene single and Object
single words, suggesting this was not influencing the activ-
ity patterns (high imagery group: t(14) = 0.40, p = .69; mid
imagery group: t(13) = −0.06, p = .95; low imagery group:
t(13) = 1.25, p = .24).
In summary, the neural patterns in anterior hippocam-
pus for Object word pairs showed greater similarity with
the Scene single words in the high imagery group, whereas
for the mid and low imagery groups, this was not the
case. On the other hand, we saw no differences in any of
the imagery groups in the posterior hippocampus. This
provides further evidence linking the anterior hippo-
campus with the processing of Object word pairs through
scene imagery.
DISCUSSION
The aim of this study was to understand the role of the
hippocampus in processing VPA. There were five findings.
First, we observed greater anterior (but not posterior)
hippocampal activity for high imagery (concrete) word
pairs than very low imagery (abstract) word pairs,
highlighting the influence of visual imagery. Second, very
low imagery abstract word pairs compared with very low
imagery abstract single words revealed no differences in
hippocampal engagement, despite the former involving
binding, adding further support for the significance of
visual imagery. Third, increased anterior (but not poste-
rior) hippocampal engagement was apparent for Scene
word pairs more than Object word pairs, implicating spe-
cifically scene imagery. Fourth, for Object word pairs, fMRI
activity patterns in the anterior (but not posterior) hippo-
campus were more similar to those for scene imagery than
object imagery, further underlining the propensity of the
anterior hippocampus to respond to scene imagery. Finally,
our examination of high, mid, and low imagery users
found that the only difference between them was the
1836
Journal of Cognitive Neuroscience
Volume 30, Number 12
use of scene imagery for encoding Object word pairs by
high imagers, which in turn was linked to scene-related
activity patterns in the anterior (but not posterior) hippo-
campus. Overall, our results provide evidence that ante-
rior hippocampal engagement during VPA seems to be
closely related to the use of scene imagery, even for
Object word pairs.
Previous findings have hinted that visual imagery might
be relevant in the hippocampal processing of verbal mate-
rial such as VPA. Work in patients with right temporal
lobectomies, which included removal of some hippo-
campal tissue, suggested that, although memory for high
imagery word pairs was impaired, memory for low imagery
word pairs was preserved (Jones-Gotman & Milner, 1978).
Furthermore, instructing these patients to use visual imag-
ery strategies impaired both high and low imagery word
pair performance ( Jones-Gotman, 1979). More recently,
detailed examination of the language use of patients with
bilateral hippocampal damage showed that the patients
used fewer high imagery words when producing verbal
narratives compared with both healthy controls and pa-
tients with damage elsewhere in the brain (Hilverman
et al., 2017), supporting a link between the hippocampus
and word imageability. In addition, higher than expected
word pair performance has been found in amnesic patients
for highly semantically related word pairs in comparison to
unrelated word pairs of the kind that are usually em-
ployed in VPA tasks (Shimamura & Squire, 1984; Winocur
& Weiskrantz, 1976). This suggests that when alternate
strategies can be used to remember word pairs (i.e.,
using their semantic relationship rather than constructing
scene imagery), amnesic patients do not show the typical
VPA impairment. We are, however, unaware of any study
that has examined VPA in patients with selective bilateral
hippocampal damage where high and low imagery word
pairs were directly compared (Clark & Maguire, 2016).
fMRI findings also support a possible distinction in hip-
pocampal engagement between high and low imagery
word pairs. Caplan and Madan (2016) investigated the
role of the hippocampus in boosting memory performance
for high imagery word pairs, concluding that imageability
increased hippocampal activity. However, greater hippo-
campal activity for high over low imagery word pairs was
only observed at a lenient whole-brain threshold ( p <
.01 uncorrected, cluster size ≥ 5), possibly because their
low imagery words (e.g., muck, fright) retained quite a
degree of imageability. Furthermore, they did not examine
the influence of different types of visual imagery on hippo-
campal engagement.
We did not find hippocampal engagement for the low
imagery Abstract word pairs compared with Abstract
single words, even when using ROI analyses and just
the remembered stimuli. We acknowledge that null re-
sults can be difficult to interpret and that an absence of
evidence is not evidence of absence. However, even our
lenient uncorrected ROI analyses found no evidence of
increased hippocampal activity. This is in clear contrast to
the finding of increased hippocampal activity for the high
imagery word pairs over the very low imagery Abstract
word pairs at the whole-brain level. The most parsimonious
interpretation is, therefore, that Abstract word pairs may
be processed differently to the high imagery word pairs,
in particular in terms of hippocampal engagement.
By contrast, activity associated with the Abstract word
pairs was evident outside the hippocampus, where re-
gions that included the left middle temporal cortex, the
left temporal pole, and the left inferior frontal gyrus were
engaged. These findings are in line with other fMRI stud-
ies that examined the representations of abstract words
and concepts in the human brain (Wang et al., 2017; Wang,
Conder, Blitzer, & Shinkareva, 2010; Binder, Westbury,
McKiernan, Possing, & Medler, 2005). Our results, there-
fore, align with the notion of different brain systems for
processing concrete (high imagery) and abstract (low im-
agery) concepts and stimuli.
Our different word types were extremely well matched
across a wide range of features, with the abstract words
being verified as eliciting very little imagery and the scene
and object words as reliably eliciting the relevant type of
imagery. Using these stimuli, we showed that hippocam-
pal involvement in VPA is not linked to visual imagery in
general but seems to be specifically related to scene im-
agery, even when each word in a pair denoted an object.
This supports a prediction made by Maguire and Mullally
(2013; see also Clark & Maguire, 2016), who noted that a
scene allows us to collate a lot of information in a quick,
coherent, and efficient manner. Consequently, they pro-
posed that people may automatically use scene imagery
during the processing of high imagery verbal material.
For instance, we might visualize the scene within which
a story is unfolding or place the objects described in
word pairs in a simple scene together.
If verbal tasks can provoke the use of imagery-based
strategies and if these strategies involve scenes, then
patients with hippocampal amnesia would be expected
to perform poorly on VPA tasks involving high imagery
concrete words because they are known to have difficulty
with constructing scenes in their imagination (e.g., Kurczek
et al., 2015; Mullally, Intraub, & Maguire, 2012; Race et al.,
2011; Andelman, Hoofien, Goldberg, Aizenstein, & Neufeld,
2010; Hassabis, Kumaran, Vann, et al., 2007). This impair-
ment, which was not apparent for single objects, prompted
the proposal of the scene construction theory, which
holds that scene imagery constructed by the hippocam-
pus is a vital component of memory and other functions
(Maguire & Mullally, 2013; Hassabis & Maguire, 2007).
Findings over the last decade have since linked scenes
to the hippocampus in relation to autobiographical mem-
ory (Hassabis, Kumaran, & Maguire, 2007; Hassabis &
Maguire, 2007) but also widely across cognition, including
perception (McCormick et al., 2017; Mullally et al., 2012;
Graham et al., 2010), future thinking (Irish, Hodges, &
Piguet, 2013; Schacter et al., 2012; Hassabis, Kumaran,
Vann, et al., 2007), spatial navigation (Clark & Maguire,
Clark, Kim, and Maguire
1837
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
2016; Maguire, Nannery, & Spiers, 2006), and decision-
making (McCormick, Rosenthal, Miller, & Maguire, 2016;
Mullally & Maguire, 2014). However, as the current study
was only designed to examine the role of the hippocampus
in the VPA task, we do not speculate further here as to
whether or not scene construction is the primary mecha-
nism at play within the hippocampus. For more on this is-
sue, we refer the reader to broader theoretical discussions
of the scene construction theory (McCormick, Ciaramelli,
De Luca, & Maguire, 2018; Dalton & Maguire, 2017; Clark
& Maguire, 2016; Maguire, Intraub, & Mullally, 2016) and
alternative accounts of hippocampal function (Moscovitch
et al., 2016; Sheldon & Levine, 2016; Eichenbaum & Cohen,
2014; Schacter et al., 2012).
Our hippocampal findings were located in the ante-
rior portion of the hippocampus. Anterior and posterior
functional differentiation is acknowledged as a feature of
the hippocampus, although the exact roles played by
each portion are not widely agreed (Ritchey, Montchal,
Yonelinas, & Ranganath, 2015; Strange, Witter, Lein, &
Moser, 2014; Poppenk, Evensmoen, Moscovitch, & Nadel,
2013; Fanselow & Dong, 2010; Moser & Moser, 1998). Of
note, the medial portion of the anterior hippocampus
contains the presubiculum and parasubiculum hippo-
campal subfields. These areas have been highlighted as
being consistently implicated in scene processing (re-
viewed in Zeidman & Maguire, 2016) and were recently
proposed to be neuroanatomically determined to process
scenes (Dalton & Maguire, 2017). The current results
seem to accord with these findings, although higher-
resolution studies are required to determine the specific
subfields involved.
An important point to consider is whether our results
can be explained by the effectiveness of encoding, as
measured in a subsequent memory test. It is certainly
true that people tend to recall fewer abstract than con-
crete words in behavioral studies of memory (Paivio,
Walsh, & Bons, 1994; Jones, 1974; Paivio, 1969). We tested
memory for both single words and paired words. Memory
performance for Scene, Object, and Abstract words was
comparable when tested singly. Memory for the word
pairs was significantly lower for the low imagery Abstract
word pairs compared with the Scene word pairs and Object
word pairs. Nevertheless, performance for all conditions
was above chance, which was impressive given the large
number of stimuli to be encoded with only one exposure.
Increased hippocampal activity was apparent for both
Scene word pairs and Object words pairs compared with
the Abstract word pairs when all stimuli or only the sub-
sequently remembered stimuli were analyzed. Further-
more, although Object word pairs were remembered
better than Scene word pairs, hippocampal activity was
nevertheless greater for the Scene word pairs. This shows
that our results cannot be explained by encoding success.
It is also worth considering why such a gradient in mem-
ory performance was observed within the word pairs, with
Object word pairs being remembered better than Scene
word pairs, which were remembered better than Abstract
word pairs. One possibility may be that memory perfor-
mance benefitted from the extent to which the two words
could be combined into some kind of relationship. This is
arguably easier for two objects than two scenes, both of
which are easier than for two abstract concepts.
Although differences between the performance of
amnesic patients and healthy participants on VPA tasks
are typically observed during cued recall, in the current
study we used recognition memory tests postscanning
to assess the success of encoding. This is because testing
cued recall for 135 word pairs that were each seen only
once is simply too difficult even for healthy participants.
For example, learning just 14 (high imagery concrete)
word pairs on the WMS-IV VPA task is performed over
four learning trials. We did, however, ensure that the
associative recognition memory test was challenging by
constructing the lure word pairs from the single words
that were presented to the participants during scanning.
Thus, all words were previously seen by participants, but
not all were previously seen in pairs.
Moreover, we believe that the use of a recognition
memory test instead of cued recall had little impact on
the patterns of brain activity we observed because brain
activity was assessed during the initial presentation of the
word pairs and not during memory retrieval. As partici-
pants were not told exactly how their memory would
be tested after the learning phase, it might be expected
that participants engaged in the most effortful encoding
that they could. That the involvement of the hippocam-
pus was identified when using all the trials in the fMRI
analysis or just the subsequently remembered stimuli
also points to the use of imagery at the time of stimuli
presentation as being of most relevance rather than en-
coding success.
There is a wealth of research linking the hippocampus
with associative binding (e.g., Palombo, Hayes, Peterson,
Keane, & Verfaellie, 2018; Roberts et al., 2018; Rangel
et al., 2016; Schwarb et al., 2015; Eichenbaum & Cohen,
2014; Addis, Cheng, Roberts, & Schacter, 2011; Konkel &
Cohen, 2009; Davachi, 2006). We do not deny this is the
case but suggest that our results provoke a reconsid-
eration of the underlying reason for apparent associative
effects. We found that the creation of associations be-
tween nonimageable Abstract word pairs did not elicit an
increase in hippocampal activity compared with Abstract
single words, even when only subsequently remembered
stimuli were considered. If binding per se was the reason
for hippocampal involvement in our study, then this con-
trast should have revealed it. We suggest instead that the
anterior hippocampus engages in associative binding spe-
cifically to create scene imagery and that this relationship
with scenes has been underestimated or ignored in VPA
and other associative tasks despite potentially having a
significant influence on hippocampal engagement.
Our participants were self-declared low, mid, or high
imagery users as measured by the VVIQ. They differed
1838
Journal of Cognitive Neuroscience
Volume 30, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
only in the degree of scene imagery usage, in particular
during the processing of Object word pairs, with high
imagers showing the greatest amount. Given that scene
imagery has been implicated in functions across cogni-
tion, it might be predicted that those who are able to
use scene imagery well might have more successful recall
of autobiographical memories and better spatial naviga-
tion. Individual differences studies are clearly required
to investigate this important issue in depth, as currently
there is a dearth of such work (a point also noted by
Palombo, Sheldon, & Levine, 2018). In this study, increased
use of scene imagery by the high imagery group did not
convey a memory advantage for the Object word pairs.
However, in the real world, with more complex memo-
randa like autobiographical memories, we predict that
scene imagery would promote better memory.
In conclusion, we showed a strong link between the
anterior hippocampus and processing words in a VPA
task mediated through scene imagery. This offers a way
to reconcile hippocampal theories that have a visuo-
spatial bias with the processing and subsequent memory
of verbal material. Moreover, we speculate that this could
hint at a verbal system in humans piggy-backing on top
of an evolutionarily older visual (scene) mechanism (see
also Corballis, 2017). We believe it is likely that other
common verbal tests, such as story recall and list learn-
ing, which are typically highly imageable, may similarly
engage scene imagery and the anterior hippocampus.
Greater use of low imagery abstract verbal material would
seem to be prudent in future verbal memory studies.
Indeed, an obvious prediction arising from our results
is that patients with selective bilateral hippocampal dam-
age would be better at recalling abstract compared with
imageable word pairs, provided care is taken to match
the stimuli precisely. Our data do not speak to the issue
of whether or not scene construction is the primary
mechanism at play within the hippocampus, as our main
interest was in examining VPA, a task closely aligned with
the hippocampus. What our results show, and we believe
in a compelling fashion, is that anterior hippocampal
engagement during VPA seems to be best explained by
the use of scene imagery.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Clark, Kim, and Maguire
1839
Appendix A. List of Scene Words Used in the Study
Airfield
Airport
Aisle
Alley
Apartment
Aquarium
Arcade
Arena
Attic
Auditorium
Avalanche
Avenue
Backyard
Bakery
Ballroom
Bank
Banquet
Barbecue
Barnyard
Basement
Bathroom
Battlefield
Bay
Beach
Bedroom
Blizzard
Boardroom
Bog
Bookshop
Boutique
Brewery
Brook
Buffet
Cabin
Cafeteria
Campsite
Canyon
Carnival
Casino
Castle
Cathedral
Catwalk
Cave
Cellar
Cemetery
Chapel
Chateau
Choir
Church
Cinema
Circus
City
Classroom
Cliff
Clinic
Coast
Cockpit
Coliseum
College
Constellation
Corridor
Cottage
Countryside
Courtroom
Courtyard
Cove
Crater
Creek
Crowd
Crypt
Cubicle
Dawn
Depot
Dock
Dormitory
Drawbridge
Driveway
Dungeon
Eclipse
Explosion
Factory
Farm
Festival
Fireworks
Fjord
Flood
Forest
Fortress
Foyer
Gallery
Garage
Garden
Glacier
Gorge
Graveyard
Gym
Gymnasium
Hairdresser
Hall
Harbor
Heliport
Herd
Highway
Hill
Horizon
Hospital
Hotel
Hurricane
Infirmary
Inn
Island
Jungle
Kitchen
Laboratory
Lagoon
Lake
Landscape
Lane
Lawn
Library
Lightning
Loft
Manor
Market
Mausoleum
Maze
Meadow
Monastery
Morgue
Motel
Mountain
Museum
Newsroom
Nightclub
Nursery
Oasis
Observatory
Ocean
Office
Orchard
Orchestra
Palace
Parade
Park
Passageway
Pasture
Patio
Peninsula
Pharmacy
Picnic
Planetarium
Plantation
Plateau
Playground
Playroom
Plaza
Pond
Port
Prairie
Pub
Quarry
Racetrack
Railroad
Railway
Ranch
Ravine
Reservoir
Restaurant
River
Road
Rodeo
Rooftop
Salon
Sandstorm
Sauna
School
Sea
Seaside
Sewer
Shipwreck
Shipyard
Shop
Sky
Snowstorm
Stadium
Stage
Station
Stockroom
Storm
Stream
Street
Studio
Sunrise
Sunset
Supermarket
Swamp
Tavern
Temple
Terrace
Terrain
Theatre
Tornado
Tournament
Tower
Town
Trail
Tunnel
Twilight
University
Valley
Villa
Village
Vineyard
Volcano
Warehouse
Waterfall
Waterfront
Zoo
1840
Journal of Cognitive Neuroscience
Volume 30, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Appendix B. List of Object Words Used in the Study
Ambulance
Amulet
Apron
Asterisk
Badger
Bandage
Banjo
Barrel
Bayonet
Beer
Bench
Biscuit
Blazer
Blindfold
Blossom
Book
Boomerang
Bouquet
Bowl
Brick
Broccoli
Brooch
Bubble
Buckle
Bullet
Buoy
Butter
Buttercup
Button
Cabinet
Calf
Camera
Canoe
Cap
Card
Cardinal
Carp
Carriage
Catalogue
Catapult
Cent
Certificate
Chain
Champagne
Chariot
Cheese
Cheetah
Cherry
Chestnut
Chisel
Cigarette
Cloak
Clock
Coaster
Cocktail
Cod
Coffee
Comet
Compass
Computer
Conditioner
Container
Cooker
Crepe
Crocodile
Crown
Crucifix
Cushion
Cylinder
Desk
Diamond
Dice
Dictionary
Dough
Doughnut
Dragon
Drainpipe
Driftwood
Drill
Dynamite
Embroidery
Emerald
Envelope
Espresso
Fairy
Fan
Flask
Frame
Generator
Guitar
Hammer
Hamper
Handle
Harness
Harpoon
Headphones
Heart
Helicopter
Honey
Honeycomb
Hook
Horseradish
Jigsaw
Kangaroo
Kayak
Ladle
Leech
Lemon
Leopard
Letter
Lifeboat
Limousine
Lobster
Locket
Lotus
Machine
Magazine
Marmalade
Medal
Medallion
Menu
Microphone
Microwave
Minibus
Monkey
Mosquito
Motorbike
Muffin
Nest
Newspaper
Noose
Nut
Olive
Ostrich
Package
Packet
Page
Painting
Pallet
Pamphlet
Panda
Parcel
Parchment
Pasta
Pendulum
Pepper
Peppermint
Photograph
Picture
Pint
Potato
Projector
Prune
Pterodactyl
Pudding
Pump
Pussycat
Rabbit
Racehorse
Radiator
Raspberry
Recorder
Reptile
Rifle
Rocket
Rose
Ruby
Sapphire
Satellite
Saucer
Scale
Scalpel
Scone
Screwdriver
Seat
Shampoo
Shell
Shrapnel
Shutter
Signature
Skirt
Skull
Sleigh
Smoothie
Snorkel
Socket
Soup
Spade
Staple
Stretcher
Submarine
Sunflower
Swing
Tabloid
Tangerine
Tapestry
Telescope
Thread
Ticket
Topaz
Tortoise
Treasure
Tripod
Trolley
Tulip
Turnip
Typewriter
Van
Vase
Vodka
Wand
Weasel
Wedge
Wheelbarrow
Whip
Whistle
Window
Witch
Clark, Kim, and Maguire
1841
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Appendix C. List of Abstract Words Used in the Study
Abstinence
Absurd
Accord
Affront
Ambition
Amends
Annual
Aptitude
Aspect
Assurance
Attempt
Attitude
Avail
Awareness
Basis
Behavior
Belief
Bias
Blame
Bother
Care
Cause
Certainty
Chance
Clarity
Closure
Commitment
Concept
Concern
Conduct
Confidence
Conjecture
Conscience
Contention
Context
Courtesy
Creed
Cunning
Debut
Decision
Decorum
Default
Desire
Despair
Destiny
Difference
Dignity
Dilemma
Discretion
Distinction
Distrust
Doubt
Dread
Duty
Effect
Ego
Empathy
Envy
Essence
Esteem
Eternity
Ethic
Euphemism
Existence
Expertise
Extent
Extreme
Fairness
Fate
Fault
Feel
Feeling
Finesse
Folly
Foresight
Forgiveness
Function
Gain
Gist
Godsend
Grandeur
Grudge
Guess
Guilt
Hint
Honesty
Honour
Idea
Ideal
Idealist
Importance
Infinity
Insight
Intellect
Intent
Intention
Interest
Intuition
Involvement
Irony
Issue
Judgment
Karma
Kind
Knack
Lack
Leeway
Legacy
Leniency
Likelihood
Logic
Luck
Manner
Meaning
Memory
Mercy
Merit
Metaphor
Method
Midst
Mistake
Mode
Moment
Mood
Moral
Morale
Motive
Mystique
Myth
Need
Neutral
Nonsense
Normal
Notion
Opinion
Origin
Outcome
Oversight
Paradigm
Paradox
Pardon
Patience
Pause
Penance
Piety
Pity
Plea
Precaution
Prestige
Principles
Prophecy
Prudence
Psyche
Pun
Purity
Purpose
Quality
Rarity
Readiness
Realism
Reason
Reasoning
Reckoning
Reform
Regard
Relevance
Remorse
Renown
Repression
Reproach
Resolve
Respect
Respite
Retrospect
Reverence
Rhetoric
Riddance
Risk
Role
Rumour
Sanctity
Sanity
Sarcasm
Seriousness
Skill
Snub
Splendour
Standard
Standpoint
Stoic
Strategy
Subconscious
Succession
Taboo
Tact
Tendency
Theory
Think
Thrift
Tradition
Transition
Trust
Truth
Try
Uncertainty
Unknown
Utmost
Validity
Value
Version
Virtue
Way
Whim
Willpower
Wisdom
Wish
Woe
Zeal
1842
Journal of Cognitive Neuroscience
Volume 30, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Acknowledgments
E. A. M. and I. A. C. were supported by a Wellcome Principal
Research Fellowship to E. A. M. (101759/Z/13/Z) and the Centre
by a Centre Award from Wellcome (203147/Z/16/Z). M. K. was
supported by a Wellcome PhD studentship (102263/ Z/13/ Z)
and a Samsung Scholarship.
Reprint requests should be sent to Eleanor A. Maguire, Wellcome
Centre for Human Neuroimaging, Institute of Neurology, Uni-
versity College London, 12 Queen Square, London, WC1N 3AR,
United Kingdom, or via e-mail: e.maguire@ucl.ac.uk.
REFERENCES
Addis, D. R., Cheng, T., Roberts, R. P., & Schacter, D. L. (2011).
Hippocampal contributions to the episodic simulation of
specific and general future events. Hippocampus, 21,
1045–1052.
Andelman, F., Hoofien, D., Goldberg, I., Aizenstein, O., &
Neufeld, M. Y. (2010). Bilateral hippocampal lesion and a
selective impairment of the ability for mental time travel.
Neurocase, 16, 426–435.
Andrews-Hanna, J. R., Reidler, J. S., Sepulcre, J., Poulin, R., &
Buckner, R. L. (2010). Functional-anatomic fractionation of
the brain’s default network. Neuron, 65, 550–562.
Ashburner, J. (2009). Computational anatomy with the SPM
software. Magnetic Resonance Imaging, 27, 1163–1174.
Ashburner, J., & Friston, K. J. (2000). Voxel based morphometry
—The methods. Neuroimage, 11, 805–821.
Balota, D. A., Yap, M. J., Hutchison, K. A., Cortese, M. J., Kessler,
B., Loftis, B., et al. (2007). The English Lexicon Project.
Behavior Research Methods, 39, 445–459.
Bellmund, J. L. S., Deuker, L., Navarro Schröder, T., & Doeller,
C. F. (2016). Grid-cell representations in mental simulation.
eLife, 5, e17089.
Binder, J. R., Westbury, C., McKiernan, K., Possing, E., & Medler,
D. (2005). Distinct brain systems for processing concrete and
abstract concepts. Journal of Cognitive Neuroscience, 17,
905–917.
Brysbaert, M., Warriner, A., & Kuperman, V. (2014). Concreteness
ratings for 40 thousand generally known English word
lemmas. Behavior Research Methods, 46, 904–911.
Burton, L. J., & Fogarty, G. J. (2003). The factor structure of
visual imagery and spatial abilities. Intelligence, 31, 289–318.
Campos, A., & Pérez-Fabello, M. J. (2005). The Spanish version
of Betts’ Questionnaire upon Mental Imagery. Psychological
Reports, 96, 51–56.
Caplan, J. B., & Madan, C. R. (2016). Word imageability enhances
association-memory by increasing hippocampal engagement.
Journal of Cognitive Neuroscience, 28, 1522–1538.
Chadwick, M. J., Jolly, A. E. J., Amos, D. P., Hassabis, D., &
Spiers, H. J. (2015). A goal direction signal in the human
entorhinal/subicular region. Current Biology, 25, 87–92.
Clark, I. A., & Maguire, E. A. (2016). Remembering preservation
in hippocampal amnesia. Annual Review of Psychology, 67,
51–82.
Cohen, N. J., & Eichenbaum, H. (1993). Memory, amnesia
and the hippocampal system. Cambridge, MA: MIT Press.
Corballis, M. C. (2017). Language evolution: A changing
perspective. Trends in Cognitive Sciences, 21, 229–236.
Dalton, M. A., & Maguire, E. A. (2017). The pre/parasubiculum:
A hippocampal hub for scene-based cognition? Current
Opinion in Behavioral Sciences, 17, 34–40.
Dalton, M. A., Zeidman, P., Barry, D. N., Williams, E., & Maguire, E. A.
(2017). Segmenting subregions of the human hippocampus on
structural magnetic resonance image scans: An illustrated
tutorial. Brain and Neuroscience Advances, 1, 1–36.
Davachi, L. (2006). Item, context and relational episodic encoding
in humans. Current Opinion in Neurobiology, 16, 693–700.
Deichmann, R., Schwarzbauer, C., & Turner, R. (2004).
Optimisation of the 3D MDEFT sequence for anatomical
brain imaging: Technical implications at 1.5 and 3 T.
Neuroimage, 21, 757–767.
Deuker, L., Bellmund, J. L. S., Navarro Schröder, T., & Doeller,
C. F. (2016). An event map of memory space in the
hippocampus. eLife, 5, e16534.
Eichenbaum, H., & Cohen, N. J. (2014). Can we reconcile
the declarative memory and spatial navigation views on
hippocampal function? Neuron, 83, 764–770.
Fanselow, M. S., & Dong, H.-W. (2010). Are the dorsal and
ventral hippocampus functionally distinct structures? Neuron,
65, 7–19.
Giovanello, K. S., Verfaellie, M., & Keane, M. M. (2003).
Disproportionate deficit in associative recognition relative
to item recognition in global amnesia. Cognitive, Affective, &
Behavioral Neuroscience, 3, 186–194.
Graf, P., & Schacter, D. L. (1985). Implicit and explicit memory
for new associations in normal and amnesic subjects.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 11, 501–518.
Graham, K. S., Barense, M. D., & Lee, A. C. H. (2010). Going
beyond LTM in the MTL: A synthesis of neuropsychological
and neuroimaging findings on the role of the medial temporal
lobe in memory and perception. Neuropsychologia, 48,
831–853.
Gur, R. C., & Hilgard, E. R. (1975). Visual imagery and the
discrimination of differences between altered pictures
simultaneously and successively presented. British Journal
of Psychology, 66, 341–345.
Haist, F., Shimamura, A. P., & Squire, L. R. (1992). On the
relationship between recall and recognition memory.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 18, 691–702.
Hassabis, D., Kumaran, D., & Maguire, E. A. (2007). Using
imagination to understand the neural basis of episodic
memory. Journal of Neuroscience, 27, 14365–14374.
Hassabis, D., Kumaran, D., Vann, S. D., & Maguire, E. A. (2007).
Patients with hippocampal amnesia cannot imagine new
experiences. Proceedings of the National Academy of
Sciences, U.S.A., 104, 1726–1731.
Hassabis, D., & Maguire, E. A. (2007). Deconstructing episodic
memory with construction. Trends in Cognitive Sciences, 11,
299–306.
Henson, R. N., Horner, A. J., Greve, A., Cooper, E., Gregori, M.,
Simons, J. S., et al. (2017). No effect of hippocampal lesions
on stimulus-response bindings. Neuropsychologia, 103,
106–114.
Hilverman, C., Cook, S. W., & Duff, M. C. (2017). The influence
of the hippocampus and declarative memory on word use:
Patients with amnesia use less imageable words.
Neuropsychologia, 106, 179–186.
Holmes, C. J., Hoge, R., Collins, L., Woods, R., Toga, A. W., &
Evans, A. C. (1998). Enhancement of MR images using
registration for signal averaging. Journal of Computer
Assisted Tomography, 22, 324–333.
Hsieh, L.-T., Gruber, M. J., Jenkins, L. J., & Ranganath, C. (2014).
Hippocampal activity patterns carry information about
objects in temporal context. Neuron, 81, 1165–1178.
Hsieh, L.-T., & Ranganath, C. (2015). Cortical and subcortical
contributions to sequence retrieval: Schematic coding of
temporal context in the neocortical recollection network.
Neuroimage, 121, 78–90.
Irish, M., Hodges, J. R., & Piguet, O. (2013). Episodic future
thinking is impaired in the behavioural variant of
frontotemporal dementia. Cortex, 49, 2377–2388.
Clark, Kim, and Maguire
1843
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Jones, M. K. (1974). Imagery as a mnemonic aid after left
temporal lobectomy: Contrast between material-specific
and generalized memory disorders. Neuropsychologia, 12,
21–30.
Jones-Gotman, M. (1979). Incidental learning of image-mediated
or pronounced words after right temporal lobectomy.
Cortex, 15, 187–197.
Jones-Gotman, M., & Milner, B. (1978). Right temporal-lobe
contribution to image-mediated verbal
Neuropsychologia, 16, 61–71.
learning.
Kim, M., Jeffery, K. J., & Maguire, E. A. (2017). Multivoxel
pattern analysis reveals 3D place information in the human
hippocampus. Journal of Neuroscience, 37, 4270–4279.
Knowlton, B. J., Mangels, J. A., & Squire, L. R. (1996). A
neostriatal habit learning system in humans. Science, 273,
1399–1402.
Konkel, A., & Cohen, N. J. (2009). Relational memory and the
hippocampus: Representations and methods. Frontiers in
Neuroscience, 3, 166–174.
Kosslyn, S. M., Brunn, J., Cave, K. R., & Wallach, R. W. (1984).
Individual differences in mental-imagery ability: A
computational analysis. Cognition, 18, 195–243.
Kousta, S.-T., Vigliocco, G., Vinson, D. P., Andrews, M., &
Del Campo, E. (2011). The representation of abstract words:
Why emotion matters. Journal of Experimental Psychology:
General, 140, 14–34.
Kriegeskorte, N., Mur, M., & Bandettini, P. A. (2008).
Representational similarity analysis—Connecting the branches
of systems neuroscience. Frontiers in Systems Neuroscience,
2, 4.
Kriegeskorte, N., Mur, M., Ruff, D. A., Kiani, R., Bodurka, J.,
Esteky, H., et al. (2008). Matching categorical object
representations in inferior temporal cortex of man and
monkey. Neuron, 60, 1126–1141.
Kumaran, D., Hassabis, D., Spiers, H. J., Vann, S. D., Vargha-
Khadem, F., & Maguire, E. A. (2007). Impaired spatial and
non-spatial configural learning in patients with hippocampal
pathology. Neuropsychologia, 45, 2699–2711.
Kuperman, V., Stadthagen-Gonzalez, H., & Brysbaert, M. (2012).
Age-of-acquisition ratings for 30,000 English words. Behavior
Research Methods, 44, 978–990.
Kurczek, J., Wechsler, E., Ahuja, S., Jensen, U., Cohen, N. J.,
Tranel, D., et al. (2015). Differential contributions of
hippocampus and medial prefrontal cortex to self-projection
and self-referential processing. Neuropsychologia, 73,
116–126.
Lakens, D. (2013). Calculating and reporting effect sizes to
facilitate cumulative science: A practical primer for t tests
and ANOVAs. Frontiers in Psychology, 4, 863.
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D.,
Ashburner, J., Frackowiak, R. S. J., et al. (2000). Navigation-
related structural change in the hippocampi of taxi drivers.
Proceedings of the National Academy of Sciences, U.S.A., 97,
4398–4403.
Maguire, E. A., Intraub, H., & Mullally, S. L. (2016). Scenes,
spaces, and memory traces: What does the hippocampus do?
The Neuroscientist, 22, 432–439.
Maguire, E. A., & Mullally, S. L. (2013). The hippocampus: A
manifesto for change. Journal of Experimental Psychology:
General, 142, 1180–1189.
Maguire, E. A., Nannery, R., & Spiers, H. J. (2006). Navigation
around London by a taxi driver with bilateral hippocampal
lesions. Brain, 129, 2894–2907.
Malach, R., Reppas, J. B., Benson, R. R., Kwong, K. K., Jiang, H.,
Kennedy, W. A., et al. (1995). Object-related activity revealed
by functional magnetic resonance imaging in human occipital
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 92, 8135–8139.
Marchette, S. A., Vass, L. K., Ryan, J., & Epstein, R. A. (2014).
Anchoring the neural compass: Coding of local spatial
reference frames in human medial parietal lobe. Nature
Neuroscience, 17, 1598–1606.
Marks, D. F. (1973). Visual imagery differences in the recall
of pictures. British Journal of Psychology, 64, 17–24.
Mayes, A. R., Holdstock, J. S., Isaac, C. L., Montaldi, D., Grigor,
J., Gummer, A., et al. (2004). Associative recognition in a
patient with selective hippocampal lesions and relatively
normal item recognition. Hippocampus, 14, 763–784.
Mayes, A. R., Isaac, C. L., Holdstock, J. S., Hunkin, N. M.,
Montaldi, D., Downes, J. J., et al. (2001). Memory for single
items, word pairs, and temporal order of different kinds in
a patient with selective hippocampal lesions. Cognitive
Neuropsychology, 18, 97–123.
McAvinue, L. P., & Robertson, I. H. (2007). Measuring visual
imagery ability: A review. Imagination, Cognition and
Personality, 26, 191–211.
McCormick, C., Ciaramelli, E., De Luca, F., & Maguire, E. A.
(2018). Comparing and contrasting the cognitive effects of
hippocampal and ventromedial prefrontal cortex damage:
A review of human lesion studies. Neuroscience, 374,
295–318.
McCormick, C., Rosenthal, C. R., Miller, T. D., & Maguire, E. A.
(2016). Hippocampal damage increases deontological
responses during moral decision making. Journal of
Neuroscience, 36, 12157–12167.
McCormick, C., Rosenthal, C. R., Miller, T. D., & Maguire, E. A.
(2017). Deciding what is possible and impossible following
hippocampal damage in humans. Hippocampus, 27, 303–314.
Mechelli, A., Price, C. J., Friston, K. J., & Ashburner, J. (2005).
Voxel based morphometry of the human brain: Methods
and applications. Current Medical Imaging Reviews, 1,
105–113.
Milivojevic, B., Vicente-Grabovetsky, A., & Doeller, C. F. (2015).
Insight reconfigures hippocampal-prefrontal memories.
Current Biology, 25, 821–830.
Morey, R. D. (2008). Confidence intervals from normalized data:
A correction to Cousineau (2005). Tutorial in Quantitative
Methods for Psychology, 4, 61–64.
Moscovitch, M., Cabeza, R., Winocur, G., & Nadel, L. (2016).
Episodic memory and beyond: The hippocampus and
neocortex in transformation. Annual Review of Psychology,
67, 105–134.
Moser, M.-B., & Moser, E. I. (1998). Functional differentiation
in the hippocampus. Hippocampus, 8, 608–619.
Mullally, S. L., Intraub, H., & Maguire, E. A. (2012). Attenuated
boundary extension produces a paradoxical memory
advantage in amnesic patients. Current Biology, 22, 261–268.
Mullally, S. L., & Maguire, E. A. (2014). Counterfactual thinking
in patients with amnesia. Hippocampus, 24, 1261–1266.
Murray, E. A., Bussey, T. J., & Saksida, L. M. (2007). Visual
perception and memory: A new view of medial temporal
lobe function in primates and rodents. Annual Review of
Neuroscience, 30, 99–122.
O’Keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive
map. Oxford: Clarendon Press.
Paivio, A. (1969). Mental imagery in associative learning and
memory. Psychological Review, 76, 241–263.
Paivio, A., Walsh, M., & Bons, T. (1994). Concreteness effects
on memory—When and why. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 20,
1196–1204.
Paivio, A., Yuille, J. C., & Madigan, S. A. (1968). Concreteness,
imagery and meaningfulness values for 925 nouns. Journal of
Experimental Psychology, 76, 1–25.
Palombo, D. J., Hayes, S. M., Peterson, K. M., Keane, M. M.,
& Verfaellie, M. (2018). Medial temporal lobe contributions to
1844
Journal of Cognitive Neuroscience
Volume 30, Number 12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
episodic future thinking: Scene construction or future
projection? Cerebral Cortex, 28, 447–458.
Palombo, D. J., Sheldon, S., & Levine, B. (2018). Individual
differences in autobiographical memory. Trends in Cognitive
Sciences, 22, 583–597.
Poppenk, J., Evensmoen, H. R., Moscovitch, M., & Nadel, L.
(2013). Long-axis specialization of the human hippocampus.
Trends in Cognitive Sciences, 17, 230–240.
Race, E., Keane, M. M., & Verfaellie, M. (2011). Medial temporal
lobe damage causes deficits in episodic memory and episodic
future thinking not attributable to deficits in narrative
construction. Journal of Neuroscience, 31, 10262–10269.
Rangel, L. M., Rueckemann, J. W., Riviere, P. D., Keefe, K. R.,
Porter, B. S., Heimbuch, I. S., et al. (2016). Rhythmic
coordination of hippocampal neurons during associative
memory processing. eLife, 5, e09849.
Ritchey, M., Montchal, M. E., Yonelinas, A. P., & Ranganath, C.
(2015). Delay-dependent contributions of medial temporal
lobe regions to episodic memory retrieval. eLife, 4, e05025.
Roberts, R. P., Schacter, D. L., & Addis, D. R. (2018). Scene
construction and relational processing: Separable constructs?
Cerebral Cortex, 28, 1729–1732.
Rorden, C., & Brett, M. (2000). Stereotaxic display of brain
lesions. Behavioural Neurology, 12, 191–200.
Schacter, D. L., Addis, D. R., Hassabis, D., Martin, V. C., Spreng,
R. N., & Szpunar, K. K. (2012). The future of memory:
Remembering, imagining, and the brain. Neuron, 76, 677–694.
Schapiro, A. C., Turk-Browne, N. B., Norman, K. A., & Botvinick,
M. M. (2016). Statistical learning of temporal community
structure in the hippocampus. Hippocampus, 26, 3–8.
Schuck, N. W., Cai, M. B., Wilson, R. C., & Niv, Y. (2016). Human
orbitofrontal cortex represents a cognitive map of state
space. Neuron, 91, 1402–1412.
Schwarb, H., Watson, P. D., Campbell, K., Shander, C. L.,
Monti, J. M., Cooke, G. E., et al. (2015). Competition and
cooperation among relational memory representations.
PLoS One, 10, e0143832.
Scoville, W. B., & Milner, B. (1957). Loss of recent memory
after bilateral hippocampal lesions. Journal of Neurology,
Neurosurgery and Psychiatry, 20, 11–21.
Sheehan, P. W. (1967). A shortened form of Betts Questionnaire
upon Mental Imagery. Journal of Clinical Psychology, 23,
386–389.
Sheldon, S., & Levine, B. (2016). The role of the hippocampus
in memory and mental construction. Annals of the New York
Academy of Sciences, 1369, 76–92.
Shimamura, A. P., & Squire, L. R. (1984). Paired-associate
learning and priming effects in amnesia: A neuropsychological
study. Journal of Experimental Psychology: General, 113,
556–570.
Spiers, H. J., Maguire, E. A., & Burgess, N. (2001). Hippocampal
amnesia. Neurocase, 7, 357–382.
Squire, L. R. (1992). Memory and the hippocampus: A synthesis
from findings with rats, monkeys, and humans. Psychological
Review, 99, 195–231.
St. Jacques, P. L., Carpenter, A. C., Szpunar, K. K., & Schacter,
D. L. (2018). Remembering and imagining alternative versions
of the personal past. Neuropsychologia, 110, 170–179.
St. Jacques, P. L., Conway, M. A., Lowder, M. W., & Cabeza, R.
(2010). Watching my mind unfold versus yours: An fMRI
study using a novel camera technology to examine neural
differences in self-projection of self versus other perspectives.
Journal of Cognitive Neuroscience, 23, 1275–1284.
Stadthagen-Gonzalez, H., & Davis, C. (2006). The Bristol norms
for age of acquisition, imageability, and familiarity. Behavior
Research Methods, 38, 598–605.
Staresina, B. P., Henson, R. N. A., Kriegeskorte, N., & Alink, A.
(2012). Episodic reinstatement in the medial temporal lobe.
Journal of Neuroscience, 32, 18150–18156.
St-Laurent, M., Moscovitch, M., & McAndrews, M. P. (2016).
The retrieval of perceptual memory details depends on
right hippocampal integrity and activation. Cortex, 84,
15–33.
Strange, B. A., Witter, M. P., Lein, E. S., & Moser, E. I. (2014).
Functional organization of the hippocampal longitudinal
axis. Nature Reviews Neuroscience, 15, 655–669.
Thakral, P. P., Benoit, R. G., & Schacter, D. L. (2017).
Characterizing the role of the hippocampus during episodic
simulation and encoding. Hippocampus, 27, 1275–1284.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject
brain. Neuroimage, 15, 273–289.
van Heuven, W. J. B., Mandera, P., Keuleers, E., & Brysbaert, M.
(2014). SUBTLEX-UK: A new and improved word frequency
database for British English. Quarterly Journal of Experimental
Psychology, 67, 1176–1190.
Vigliocco, G., Kousta, S.-T., Della Rosa, P. A., Vinson, D. P.,
Tettamanti, M., Devlin, J. T., et al. (2014). The neural
representation of abstract words: The role of emotion.
Cerebral Cortex, 24, 1767–1777.
Wang, J., Conder, J. A., Blitzer, D. N., & Shinkareva, S. V. (2010).
Neural representation of abstract and concrete concepts:
A meta-analysis of neuroimaging studies. Human Brain
Mapping, 31, 1459–1468.
Wang, X., Wu, W., Ling, Z., Xu, Y., Fang, Y., Wang, X., et al. (2017).
Organizational principles of abstract words in the human
brain. Cerebral Cortex, 1–14. doi:10.1093/cercor/bhx283.
Warriner, A. B., Kuperman, V., & Brysbaert, M. (2013). Norms of
valence, arousal, and dominance for 13,915 English lemmas.
Behavior Research Methods, 45, 1191–1207.
Wechsler, D. (1945). A standardized memory scale for clinical
use. Journal of Psychology, 19, 87–95.
Wechsler, D. (2008). Wechsler Adult Intelligence Scale–Fourth
Edition ( WAIS-IV). San Antonio, TX: NCS Pearson.
Wechsler, D. (2009). WMS-IV: Wechsler Memory Scale-
Administration and scoring manual. London: Psychological
Corporation.
Wechsler, D. (2011). Test of Premorbid Functioning–UK
version (TOPF UK). London: Pearson Assessment.
Weiskopf, N., Hutton, C., Josephs, O., & Deichmann, R. (2006).
Optimal EPI parameters for reduction of susceptibility-induced
BOLD sensitivity losses: A whole-brain analysis at 3 T and
1.5 T. Neuroimage, 33, 493–504.
Winocur, G., & Weiskrantz, L. (1976). An investigation of paired-
associate learning in amnesic patients. Neuropsychologia,
14, 97–110.
Zeidman, P., & Maguire, E. A. (2016). Anterior hippocampus:
The anatomy of perception, imagination and episodic
memory. Nature Reviews Neuroscience, 17, 173–182.
Zola-Morgan, S., Squire, L. R., & Amaral, D. G. (1986). Human
amnesia and the medial temporal region: Enduring memory
impairment following a bilateral lesion limited to field
CA1 of the hippocampus. Journal of Neuroscience, 6,
2950–2967.
Clark, Kim, and Maguire
1845
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
2
1
8
2
1
1
7
8
7
9
0
1
/
/
j
o
c
n
_
a
_
0
1
3
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3