Neural Integration in Body Perception
Richard Ramsey
Abstracto
■ The perception of other people is instrumental in guiding
social interactions. Por ejemplo, the appearance of the human
body cues a wide range of inferences regarding sex, edad, salud,
and personality, as well as emotional state and intentions, cual
influence social behavior. Hasta la fecha, most neuroscience research
on body perception has aimed to characterize the functional con-
tribution of segregated patches of cortex in the ventral visual
stream. In light of the growing prominence of network architec-
tures in neuroscience, the current article reviews neuroimaging
studies that measure functional integration between different
brain regions during body perception. The review demonstrates
that body perception is not restricted to processing in the ventral
visual stream but instead reflects a functional alliance between the
ventral visual stream and extended neural systems associated with
action perception, funciones ejecutivas, and theory of mind.
En general, these findings demonstrate how body percepts are con-
structed through interactions in distributed brain networks and
underscore that functional segregation and integration should
be considered together when formulating neurocognitive theo-
ries of body perception. Insight from such an updated model of
body perception generalizes to inform the organizational struc-
ture of social perception and cognition more generally and also
informs disorders of body image, such as anorexia nervosa, cual
may rely on atypical integration of body-related information. ■
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INTRODUCCIÓN
The appearance of the human body provides a rich
source of social information. Bodies signal cues to an
observed individual’s sex, edad, salud, and personality,
as well as his or her emotional states and intentions. Semejante
signals are important for social interactions, as they guide
human behavior in terms of approach/avoidance tenden-
cíes, mate selection, and cooperation. Given their instru-
mental influence on daily life, research has aimed to
identify the neurobiological mechanisms by which such
signals are detected, procesado, and utilized (Frith &
Frith, 2010).
Research investigating the perception of other people—
social perception—has been dominated by the study of
faces ( Jacobo & Schyns, 2017; duchaína & Yovel, 2015;
Kanwisher, 2010; Haxby, Hoffman, & Gobbini, 2000; bruce
& Joven, 1986). Faces play a central role in social interac-
ciones, and as a consequence, face perception research has
provided valuable insights. Sin embargo, bodies also cue a
range of information that is exploited during social inter-
comportamiento (de Gelder et al., 2010), cual, at times, faces
conceal (Aviezer, Trope, & todorov, 2012). Por lo tanto, si
a core aim of social perception research is to understand
how we read and navigate social signals in the real world,
bodies are also a vitally important cue to study. Además,
bodies, like faces, can be studied as a model system to
investigate the cognitive and neural processes that under-
pin social perception.
Bangor University
The majority of neuroscience research on body per-
ception has focused on understanding the role of seg-
regated patches of cortex in the ventral visual stream
(for reviews, see Downing & Peelen, 2011, 2016). Este
work has identified two regions of ventral temporal cor-
tex (fusiform body area [FBA], extrastriate body area
[EBA]) that respond more robustly to bodies than other
classes of stimuli, such as houses and chairs. FBA and
EBA, por lo tanto, are said to show category selectivity for
bodies. Although many functional claims have been made
for the role of these two regions, the majority of evidence
suggests that these regions primarily process body shape
and posture (Downing & Peelen, 2011).
Complicated mental processes, such as those under-
pinning aspects of social perception, are unlikely to rely
solely on segregated patches of cortex acting alone, cómo-
alguna vez (Ramsey, van Schie, & Cruz, 2011; Kanwisher,
2010). Bastante, mental processes are likely to involve the
integration of interacting signals that span across distrib-
uted neural networks (bullmore & despreciar, 2009; Fuster,
1997; Mesulam, 1990). En efecto, two cornerstones of brain
function are functional segregation and functional inte-
gration (Parque & Friston, 2013). Functional segregation is
characterized by information processing that is carried
out by functionally related brain regions that are arranged
in modules,1 whereas functional integration involves the
exchange of signals across a distributed set of such brain
networks or modules (Parque & Friston, 2013; despreciar,
2013). Given the range and complexity of social informa-
tion that bodies are associated with, responses in ventral
temporal cortex are likely to be a combined product of
© 2018 by the Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 Internacional (CC POR 4.0) licencia.
Revista de neurociencia cognitiva 30:10, páginas. 1442–1451
doi:10.1162/jocn_a_01299
local, as well as distributed, processing functions (despreciar,
2013). Hasta la fecha, sin embargo, little is known about the role of
functional integration in body perception.
The main aim of the current article is to review neuro-
imaging evidence for functional integration in body per-
ception and consider the implications of functional
integration research for understanding the neural bases
of social perception. The article is organized in four parts.
Primero, to provide a relevant context, a brief review of evi-
dence for functional segregation in body perception is
provided. Segundo, lines of evidence from fMRI studies
that have investigated functional integration in body
perception are reviewed. These studies show that brain
circuits in ventral temporal cortex and those in extended
networks associated with action perception, executive
funciones, and theory of mind integrate information
during body perception. Juntos, the first two sections
of the article suggest that, by considering functional
segregation and integration together, we will have a more
complete understanding of the neural systems that
support body perception. Tercero, the implications of such
an updated neurocognitive model of body perception for
understanding social perception and cognition more
generally, as well as disorders of body image, are dis-
cussed. Finalmente, future directions that embrace network
science approaches to understanding social perception
are outlined.
FUNCTIONAL SEGREGATION IN
BODY PERCEPTION
A primary neuroimaging method for identifying category
selectivity in the human brain has been to adopt a func-
tional ROI approach (Kanwisher, 2010, 2017). The func-
tional ROI approach typically uses univariate methods for
comparing responses across different categories of stim-
uli. Primero, ROIs are identified based on functional data
using a “localizer” scan, before the response in these re-
gions is interrogated using separate task data. This ap-
proach has identified two body-selective regions in
ventral temporal cortex (FBA and EBA), which respond
to bodies more than other object categories such as
houses and chairs (Figura 1A; Zhan, Goebel, & de
Gelder, 2018; Peelen & Downing, 2005; Downing, Jiang,
Shuman, & Kanwisher, 2001; para una revisión, see Downing &
Peelen, 2011). Functional divisions have also been iden-
tified within this body circuit with FBA showing greater
sensitivity to whole bodies and EBA showing greater sen-
sitivity to body parts (taylor, Wiggett, & Downing, 2007).
Although there is clear evidence for body shape and
posture processing in FBA and EBA, more elaborate
cognitive processes have also been ascribed to these
regions including identity, emotion, and action-related
procesos (Downing & Peelen, 2011). Sin embargo, hay
less convincing evidence for these more elaborate repre-
sentations in ventral temporal regions (Downing &
Peelen, 2011). Like the majority of brain networks, re-
sponses in ventral temporal cortex are likely to index a
local processing function as well as an exchange of signals
within a wider neural network (despreciar, 2013). Tal como,
claims based on univariate responses in EBA and FBA
may reflect the exchange of signals with wider brain
networks in addition to local processes (Parque & Friston,
2013). This is especially the case for more elaborate
representations associated with social cognition, cual
have been shown to recruit a widely distributed neural
architecture (Frith & Frith, 2010; Figura 1B). Evidencia
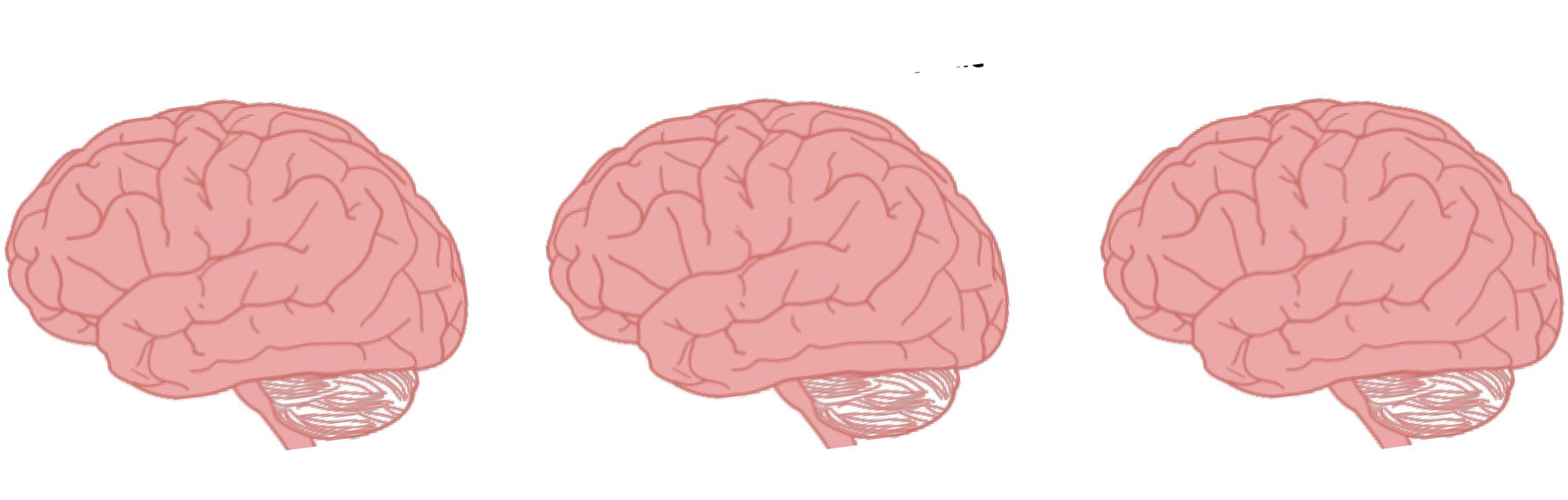
Cifra 1. Functional segregation in body perception and social cognition. The majority of research investigating body perception (A) and social
cognition (B) has focused on understanding the role of functional segregation. Functional segregation is characterized by information processing
that is carried out by functionally related brain regions that are arranged in modules. Less body perception research has investigated the role
of functional integration between brain networks (C). Functional integration is characterized by the exchange of signals across a distributed
set of brain networks or modules. Abbreviations: mPFC = medial pFC; TP = temporal pole; IFG = inferior frontal gyrus; IPL = inferior parietal
lobule; dlPFC = dorsolateral pFC. Color scheme: green = body-selective cortex; blue = theory-of-mind network; yellow = mirror neuron system;
red = executive control circuit.
Ramsey
1443
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
for interactions between body-selective areas in ventral
temporal cortex and wider networks associated with
social perception and cognition are reviewed in the next
sección.
FUNCTIONAL INTEGRATION IN
BODY PERCEPTION
Complex mental processes, such as those subserving
social perception and social inference, are unlikely to rely
on a narrow use of neural tissue that is restricted to ven-
tral temporal cortex (duchaína & Yovel, 2015; Ramsey
et al., 2011; Kanwisher, 2010; de Gelder, 2006; Haxby
et al., 2000). Models of emotional body perception, para
ejemplo, are based on a distributed and interacting
set of brain networks (de Gelder, de Borst, & watson,
2015; de Gelder, 2006). To measure network connec-
actividad, neuroimaging methods have been developed
that enable interactions between distinct anatomical or
functional regions to be estimated (Friston, 2011). Alabama-
though many connectivity studies measure resting state
actividad (Greicius, Krasnow, Reiss, & menón, 2003), otro
studies measure how connectivity changes as a function
of the experimental condition, such as the type of task or
stimulus (Friston, 2011; Friston et al., 1997).
Such task-based functional connectivity approaches
substantially extend univariate approaches by first identi-
fying functional ROIs using established localizers and
then estimating how these networks interact as a func-
tion of the task or stimulus set. At least two broad classes
of task-based connectivity have been developed: direc-
tional and correlational. Directional measures of func-
conectividad nacional, such as dynamic causal modeling
(DCM) and Granger causality, permit inferences to be
drawn regarding the direction of influence of one brain
region on another (Friston, 2009). A diferencia de, purely cor-
relational measures, such as psychophysiological inter-
comportamiento (PPI), are unable to provide an estimate of the
direction of influence (McLaren, Ries, Xu, & Johnson,
2012; Friston et al., 1997). En cambio, PPI relies on general
linear modeling to estimate how correlations between
brain regions vary as a function of task demands. Impor-
tantly, PPI modeling procedures typically include univari-
ate and PPI regressors within the same model, cual
means that, for PPI regressors to be of interest, they must
explain variance above and beyond that explained by the
univariate regressors (McLaren et al., 2012; O’Reilly,
lana rica, Behrens, Herrero, & Johansen-Berg, 2012).
Although it has been proposed that body perception
involves a distributed neural architecture that extends be-
yond ventral temporal cortex (p.ej., Ramsey et al., 2011;
de Gelder, 2006), fewer than 10 studies have investigated
functional integration during body perception using
resonancia magnética funcional. Univariate neuroimaging techniques as well as
neuropsychology lesion studies show that recognizing
emotional body postures relies on a distributed neural
architecture that extends beyond ventral temporal cortex
(for reviews, see de Gelder et al., 2015; de Gelder, 2006).
Sin embargo, the lack of functional connectivity studies
means that the boundary conditions that govern local
processing and distributed processing in body percep-
tion remain unclear (Figura 1C). En efecto, neural integra-
tion research in body perception has only just begun to
identify which neural circuits interact with ventral tempo-
ral cortex and in which social contexts. En esta sección, I
focus on studies that have used fMRI and measures of
task-based connectivity during body perception. Estos
studies have investigated the relationship between
body perception and a range of different topics including
identity recognition, action perception, executive con-
controlar, and theory of mind.
Integration within the Ventral Visual Stream
Ewbank and colleagues (2011) used a repetition suppres-
sion design to investigate functional interplay between
FBA and EBA during the processing of physical identity.
Repetition suppression is observed when a repeat stimu-
lus feature produces a reduced neural response and has
been used to test population coding models of percep-
tion and cognition (barrón, Garvert, & Behrens, 2016;
Grill-Spector, Henson, & Martín, 2006). In Ewbank and
colleagues’ (2011) estudiar, participants observed body
images that varied in size, orientación, and identity. Cuando
there was a repeat identity, both FBA and EBA showed a
reduced response, thus showing repetition suppression
for person identity. Además, Ewbank and colleagues
(2011) used DCM to show that FBA modulated responses
in EBA for a repeated compared with a novel identity.
This response was invariant to changing size and view
of the body. The authors suggest that FBA provides
top–down control over the response in EBA. Tal
interpretation is consistent with the view that FBA repre-
sents whole bodies (irrespective of size and viewpoint
cambios) and influences a more granular, body part-
specific representation in EBA that is tuned by body size
and view (Taylor et al., 2007). Por eso, este estudio muestra
that body identity processing is not only a product of
local responses in FBA and EBA but instead reflects
integration between these two nodes (Figura 2A).
Integration between the Ventral Visual Stream and
the Action Perception Network
In addition to integration between EBA and FBA, otro
studies have shown that body patches interact with wider
neural networks associated with action perception, exec-
utive functions, and theory of mind. In terms of action
percepción, Zimmermann, Toni, and de Lange (2013)
showed that body posture modulates the perception of
another’s action goals. The authors found that, cuando un
participant’s body posture matches an observed action,
1444
Revista de neurociencia cognitiva
Volumen 30, Número 10
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
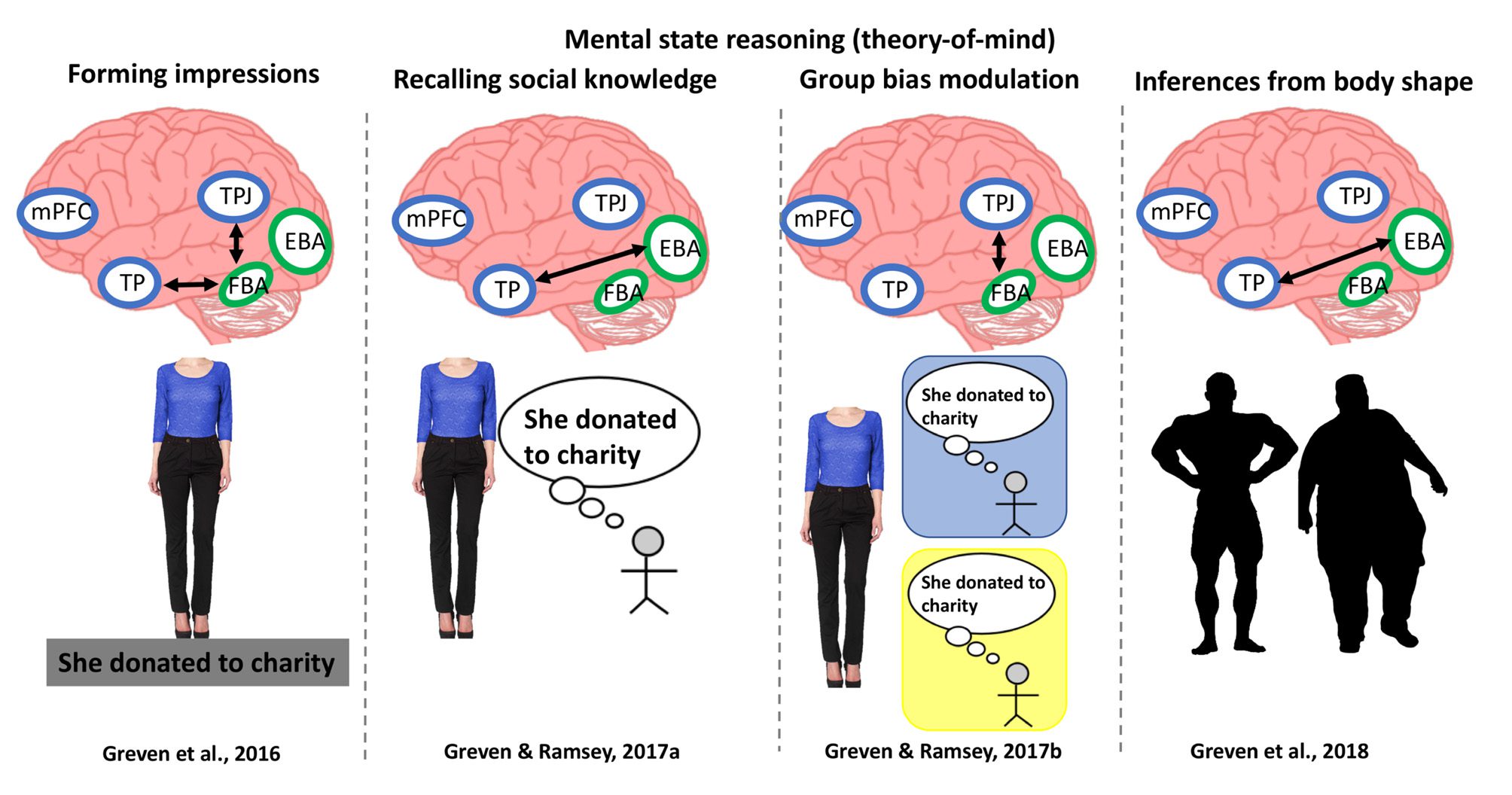
Cifra 2. Functional integration in body perception. A summary of fMRI studies that have investigated functional integration in body perception.
These studies have used measures of functional connectivity to estimate links within the ventral visual stream during identity processing (A) así como
between the ventral visual stream and networks associated with action perception (B), funciones ejecutivas (C), and theory of mind (D). Abbreviations
and color scheme as Figure 1.
the prediction of another’s action goal is facilitated. En
support of this goal ascription process, the intraparietal
sulcus was engaged more when there was a mismatch
between the participant’s body posture and the observed
action goal posture. Además, using PPI, the response
in intraparietal sulcus correlated with EBA as a function
of action frequency: observing low-frequency actions
increased coupling. The authors interpret the neuro-
imaging results within a predictive framework, bajo
the assumption that body perception signals in ventral
temporal cortex contribute to a prediction of a person’s
likely goal. The goals associated with more frequently
observed actions are less surprising and result in lower
prediction error. Por el contrario, less frequent actions pro-
duce a higher prediction error, y por lo tanto, a greater signal
exchange between intraparietal sulcus and EBA is re-
quired to update the goal estimate (Figura 2B). Estos
resultados, por lo tanto, document a link between ventral
temporal cortex and brain regions associated with the
perception of action goals.
Integration between the Ventral Visual Stream and
Executive Functions
Perception in general, whether of objects, escenas, o
gente, has been shown to involve interplay between
the visual stream and neural systems associated with ex-
ecutive functions (Baldauf & Desimone, 2014; Bar, 2004).
Executive functions are a set of mental processes that are
needed to accomplish difficult tasks, when relying on
Ramsey
1445
automated processes would be ineffective (Diamante,
2013). Using a paradigm that manipulated the presence
of sex-based stereotypes, processes associated with body
perception have been shown to have a similar interactive
relationship with executive functions (Quadflieg et al.,
2011). When we meet other people, we categorize them
into social groups based on many factors, such as sex,
edad, profesión, and race. We also hold stereotyped
expectations for such social groups, which influence
social interactions (Macrae & Quadflieg, 2010; Fiske &
Neuberg, 1990; Cervecero, 1988). Por ejemplo, we typically
expect nurses to be female and courtroom judges to
be male. In some instances, sin embargo, individuals violate
stereotypical expectations (p.ej., a male nurse). Cuando
performing sex judgments of others in situations that
violate sex-based stereotypes compared with those that
conform, Quadflieg et al. (2011) showed increased cou-
pling between dorsolateral pFC and body-selective
patches in the ventral visual stream. The authors suggest
that dorsolateral pFC modulates visual processing of
object categories, in this case bodies, to override the
initial expectation based on bodies and to modulate the
formation person percepts in the brain (Figura 2C).
Integration between the Ventral Visual Stream and
the Theory-of-Mind Network
Theory of mind is the attribution of mental states, como
creencias, desires, and attitudes, to others and has been
consistently associated with the engagement of medial
pFC, TPJ, temporal poles, and precuneus ( Van Overwalle,
2009; sajonia & Kanwisher, 2003; Frith & Frith, 1999). El
theory-of-mind network responds to a variety of tasks
involving mental state attribution and social inferences
( Van Overwalle, 2009) and can be reliably identified with
a short belief reasoning functional localizer during fMRI
(Dodell-Feder, Koster-Hale, Bedny, & sajonia, 2011).
Using body perception and theory-of-mind localizers, a
series of studies has investigated the relationship be-
tween body-selective patches in ventral temporal cortex
and the theory-of-mind network during body perception
(Figura 2D; Greven, Downing, & Ramsey, 2016, 2018;
Greven & Ramsey, 2017a, 2017b). Each study investi-
gated a distinct component of social information process-
ing during body perception, including the formation
(Greven et al., 2016) and recall (Greven & Ramsey,
2017a) of impressions, the impact of group bias on body
percepción (Greven & Ramsey, 2017b), as well as person
inferences that are based on body shape alone (Greven
et al., 2018). The broad hypothesis across these ex-
periments was the same: Social information processing
during body perception will not be restricted to uni-
variate responses in segregated networks but will also
be indexed by integration between body-selective and
theory-of-mind networks.
The first study investigated the formation of impres-
sions during body perception (Greven et al., 2016). en un
2 × 2 factorial design, bodies or names were shown to
participants alongside a short statement that described
behaviors that cued trait-based or neutral judgements
(Figura 2D). Por ejemplo, the statement “She gave
money to charity” cues a trait-based inference (p.ej.,
selfless, generous), much more than a trait-neutral state-
ment such as “She sharpened her pencil.” Therefore, el
type of inference (trait-based, neutral) and the social tar-
conseguir (body, name) were manipulated, and participants
were asked to form an impression of the person. Previo
work had demonstrated that, compared with neutral
statements, trait-based inferences engage the theory-of-
mind network (Mamá, Vandekerckhove, Van Overwalle,
Seurinck, & Fias, 2011; mitchell, Cloutier, Banaji, &
Macrae, 2006). Using PPI, Greven and colleagues (2016)
showed that FBA showed stronger functional coupling
with TPJ and temporal poles when participants formed
an impression of a body compared with when they
formed similar impressions based on a person’s name.
This suggests that, when forming impressions of others,
functional connectivity between FBA and nodes in the
theory-of-mind network are tuned to specific types of
social information (bodies more than names; trait infer-
ences more than neutral judgments).
Although first impressions are common, much of our
daily lives involve interactions with familiar people (p.ej.,
amigos, familia, and colleagues). Tal como, we have a rich
set of stored person associations, which we rely upon to
guide social exchanges. To assess recall of social knowl-
edge that is prompted by body perception, in a sub-
sequent study Greven and Ramsey (2017a) entrenado
participants before scanning to associate different bodies
with trait-based or neutral information. During scanning,
participants viewed the same bodies and were asked to
form an impression of the individual. PPI analyses
showed that perceiving bodies that prompted the recall
of social knowledge compared with bodies associated
with neutral knowledge engaged more functional cou-
pling between EBA and the temporal poles. Estos resultados
may suggest that the detection of body parts in EBA trig-
gers an exchange of signals with a node in the theory-of-
mind network that has consistently been associated with
the development of person knowledge (Olson, McCoy,
Klobusicky, & ross, 2013). One possible interpretation
of this result is that once identity is established based
on body shape and posture cues, there is a relatively
rapid exchange with a nonvisual person knowledge
representation in the temporal poles.
In addition to stored knowledge regarding trait-based
personaje, we readily recognize others as being part of an
ingroup or outgroup based on factors such as sex, profes-
sión, carrera, and age. Such group biases are prevalent in
social perception and cognition, and we typically per-
ceive ingroup members more favorably than outgroup
miembros (Cervecero, 1999; Allport, 1954). We are also
more likely to remember positive information about
ingroup members and more negative information about
1446
Revista de neurociencia cognitiva
Volumen 30, Número 10
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
outgroup members (Fyock & Estangor, 1994). In terms of
neural circuits, a distributed set of brain networks are
sensitive to group biases, which span visual, affective,
and cognitive systems (Amodio, 2014; Molenberghs,
2013). Sin embargo, little is known regarding functional con-
nectivity between these neural circuits during group bias
modulation of person perception. Greven and Ramsey
(2017b) used a minimal group manipulation (Tajfel,
Billig, Bundy, & Flament, 1971), whereby participants
were randomly assigned to a “blue” or “yellow” team
and given a long-sleeved t-shirt to wear, which matched
their team color. Participants were subsequently shown
images of ingroup and outgroup members (es decir., those
wearing blue or yellow t-shirts), who were previously
associated with positive or negative social information.
PPI results showed greater coupling between FBA and
TPJ for bias-consistent (ingroup-positive and outgroup-
negative) than inconsistent pairings. These results sug-
gest that coupling between the ventral visual stream
and the theory-of-mind network is tuned to social knowl-
edge and social group pairings. En efecto, interactions
between networks is not driven by main effects of group
or valence but instead reflects the combination of the
two types of information (ingroup, bien; outgroup, bad).
Impressions are not only formed based on explicit
knowledge of behavior; impressions are also formed
based on physical shape alone. Por ejemplo, body shape
and posture cue inferences regarding emotional state,
personality, and health (de Gelder et al., 2010; Puhl &
Heuer, 2009; Sell et al., 2009; Borkenau & Liebler,
1992). Using silhouette images of bodies, which empha-
size body shape and posture cues, Greven and colleagues
(2018) performed two fMRI experiments that investi-
gated the neural bases of inferences that are drawn from
body shape alone. Before scanning, three behavioral
experiments showed that different body types (obese,
muscular) were judged differently on dimensions of per-
sonality and health compared with slim bodies. Obese
bodies were rated as less extraverted, conscientious, y
saludable, whereas muscular bodies were rated as more
extraverted and healthy but less agreeable. Estos resultados
show that social inferences of slim bodies are more neu-
tral (es decir., closer to the middle of the rating scale) cuando
evaluating personality and health than muscular and
obese bodies. This does not imply an absence of social
inferences for slim individuals, just that inferences are
less extreme. En otras palabras, social inferences are made
for all body types, and only the content of these infer-
ences varies based on the physical attributes of the bod-
es. As part of the same study, two subsequent fMRI
experiments used the same stimuli but varied the task.
The first experiment used a 1-back recognition task and
showed no evidence for differential engagement of body
or theory-of-mind networks and no coupling between
body and theory-of-mind networks. In the second exper-
mento, which required participants to form an impression
of the person, evidence emerged for functional coupling
between EBA and the temporal poles, but it was a
relatively weak effect. Había, sin embargo, clearer evi-
dence for differential engagement of segregated neural
circuitos: the Muscular > Slim contrast engaged EBA
and FBA, whereas the Obese > Slim contrast engaged
medial pFC and temporal poles. These results suggest
that there is a division of labor between body and
theory-of-mind networks when forming an impression
based on body shape.
Juntos, this series of four fMRI studies shows that
different dimensions of body perception involve func-
tional interplay between body and theory-of-mind net-
obras. These dimensions include (1) stage of social
knowledge acquisition (formation vs. recordar), (2) the form
of social knowledge (written description vs. body shape),
(3) identity of the social target (ingroup vs. outgroup),
y (4) intentionality of social inference (unintentional
vs. intentional). Considering the results of these studies
together suggests that the ventral visual stream and the
theory-of-mind network do not act in isolation during
body perception but instead exchange signals across
multiple social information processing dimensions.
Además, the results permit speculation on a pos-
sible division of labor in functional network organization.
Forming impressions of another person’s character and
tagging such information to body shape is associated
with links between FBA and the theory-of-mind network,
including the temporal poles and TPJ (Greven et al.,
2016). It is possible that developing a richer representa-
tion of a person to include nonvisual information (es decir.,
impressions of trait-based character) involves exchange
between FBA and temporal poles, which is consistent
with the role of TP in stored person knowledge (Olson
et al., 2013) and FBA in a representation of whole bodies
(Taylor et al., 2007). It is also consistent with recent work
in the domain of face perception, whereby links between
the ventral visual stream and temporal poles have been
demonstrated to underpin the retrieval of social knowl-
edge that is associated with faces ( Wang y cols., 2017).
Por el contrario, recall of social knowledge that is prompted
by body shape involves links between EBA and temporal
poles (Greven et al., 2018; Greven & Ramsey, 2017a).
One interpretation is that, when bodies cue social infer-
ences, the detection of body parts in EBA (Taylor et al.,
2007) triggers an associated representation of stored
social knowledge in temporal poles (Olson et al., 2013).
This proposal is consistent with theories of impression
formation that posit links between representations of
facial features and trait knowledge (Encima & Cocinar,
2018). Sin embargo, the possibility that networks can be frac-
tionated into functionally distinct partitions remains spec-
ulative at the moment. En efecto, models of neural
integration between the ventral visual stream and the
theory-of-mind network are only just beginning to be
formulated, and it will be important for future work to
directly test these predictions using a range of methods
(see Limitations and Future Directions section).
Ramsey
1447
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Summary
En resumen, evidence is emerging that different dimen-
sions of body perception involve functional interplay
within the ventral visual stream, as well as between the
ventral visual stream and neural networks associated with
action perception, funciones ejecutivas, and theory of
mente (Cifra 2). These results demonstrate that the
ventral visual stream does not act alone in body percep-
tion but instead forms functional connections with
distributed neural networks that span anterior temporal,
frontal, and parietal cortices. Próximo, implications for
neurocognitive models of body perception are outlined.
IMPLICATIONS
The primary implication of the reviewed evidence is that
body percepts are constructed through relationships
between distributed and interacting neural networks. En-
deed, links between the visual stream and extended sys-
tems are suggestive that information processing in the
visual stream is not sufficient to perceive the outside en-
vironment (Sterzer, Kleinschmidt, & rees, 2009; Gilbert
& Sigman, 2007). A consequence of this suggestion for
neuroimaging research in general is that focusing on seg-
regation alone will produce skewed models of mental
processes that are biased toward a segregationist struc-
ture and underestimate complexity. This is not to suggest
that understanding functional segregation holds no value
in social perception. Bastante, these results underscore
eso, to understand complex mental processes, functional
segregation and integration need to be considered in
partnership (despreciar, 2013). En efecto, fMRI studies that
only use univariate approaches must keep in mind that
responses may not only reflect a local, segregated func-
ción, but also an integrative function.
Studying the perception of bodies, like faces, escenas,
palabras, and tools, is one way to understand organizing
principles of human brain function. Here we extend this
understanding to show how functionally segregated
modules connect to form functionally interacting net-
works during body perception. Por lo tanto, the reviewed
research uses body perception as a model system to in-
vestigate mechanisms of social perception, as well as a
means to study network models of human brain function
more generally. Como consecuencia, the results hold the po-
tential to inform other research domains that also rely
on distributed but interacting modules, such as face per-
ception (p.ej., duchaína & Yovel, 2015), object percep-
ción (Bar, 2004), and memory (Cabeza & Moscovitch,
2013). Por ejemplo, similarities are likely to exist be-
tween face and body perception (de Gelder et al.,
2010), which means core principles from the findings re-
ported here may readily apply to face perception. Relat-
edly, theories of impression formation, which specify
links between the acquisition of trait knowledge and
the representation of facial features (Encima & Cocinar,
2018), could be informed by the work reviewed here
on links between systems associated with body shape
perception and theory of mind. As a further example,
functional structures in the domains of memory (Cabeza
& Moscovitch, 2013) and object perception (Bar, 2004)
involve links between domain-specific and domain-
general systems, a picture that also emerges in the body
perception research reviewed here. Tal como, by compar-
ing different information processing domains, common
and distinct organizing principles of brain function can
emerge, which may lead to new hypotheses.
With regard to body perception research more specif-
icamente, it is becoming clearer that category selectivity in
ventral temporal cortex cannot be completely reduced
to task-invariant processing of visual features (Peelen &
Downing, 2017; Bi, Wang, & Caramazza, 2016; Harel,
Kravitz, & Panadero, 2014). En cambio, category-selective
responses reflect knowledge of what the object means
to the observer, as well as how they interact with it
(Peelen & Downing, 2017). Tal como, a wider neural archi-
tecture is likely to be important to consider. The re-
viewed studies begin to probe the boundary conditions
that control the relationship between functional seg-
regation and integration and identify which neural cir-
cuits interact with ventral temporal cortex and in which
social contexts. But integration research is only beginning
to scratch the surface of understanding this complex
topic and much more research is needed.
A deeper appreciation of network science approaches
to body perception may have clinical relevance for body-
related disorders. Por ejemplo, in anorexia nervosa,
reduced connectivity between FBA and EBA has been
associated with body image distortion (Suchan et al.,
2013). More generally, por lo tanto, when considering dis-
tortions in body image, it may prove useful to consider
the role of wider networks. Problems in body-related in-
formation processing may arise from altered integration
of body representations as much as altered responses in
the ventral visual stream alone.
LIMITATIONS AND FUTURE DIRECTIONS
The current review had a purposely narrow focus and did
not set out to provide a comprehensive review of body
perception research from a cognitive neuroscience
perspectiva. En cambio, the review targeted human fMRI re-
search that investigated body perception using measures
of functional connectivity. Tal como, a comprehensive re-
view of body perception research was beyond the scope
of this review. Además, detailed reviews have already
considered the proposed functions of EBA and FBA
(Downing & Peelen, 2011), as well as the contribution
from neurostimulation and patient studies to understand-
ing body perception (Downing & Peelen, 2016), y el
role of emotion in body perception (de Gelder et al.,
2010; de Gelder, 2006). Además, other work has used
direct intracranial recordings in humans (Pourtois,
1448
Revista de neurociencia cognitiva
Volumen 30, Número 10
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Peelen, Spinelli, Seeck, & Vuilleumier, 2007) and evi-
dence from nonhuman primates (Campana, Hadj-Bouziane,
Frihauf, Tootell, & Ungerleider, 2009; Pinsk et al., 2009;
Pinsk, DeSimone, moore, Bruto, & Kastner, 2005) to fur-
ther understand the neural bases of body perception.
A further consideration also relates to the intended
scope of the current review. The current article was
centered on understanding functional connectivity
within the ventral visual stream as well as between
the ventral visual stream and broader neural networks.
This focus was motivated by the dominance of the ventral
visual stream in person perception research to date
(Kanwisher, 2010). Sin embargo, recent body perception re-
search has also shown that coupling between extended
networks makes a contribution to emotional body per-
ception (Engelen, Zhan, Sack, & de Gelder, 2018; Poyo
Solanas et al., 2018). Por ejemplo, using fMRI, Poyo
Solanas and colleagues (2018) showed that, when faces
and bodies convey congruent compared with incongru-
ent emotional signals, there is greater functional coupling
between the amygdala and the ACC. This suggests that
the amygdala may provide a regulatory role in respond-
ing to unambiguous emotional signals, which are con-
veyed by face and body concurrently. The results also
suggest that coupling in body perception need not be
restricted functional interactions that involve the ventral
visual stream and future research should pursue this line
of research further.
Further future directions stem from three principle
limitations of the current evidence. Primero, a lack of empha-
sis on functional integration in body perception research
hampers understanding of social perception more broad-
ly. Except for models of emotional body perception,
which include distributed networks (de Gelder, 2006),
there is little research on body perception more generally
that considers functional integration and network ap-
se acerca. Building on the work reviewed here, further
research is required that investigates the boundary
conditions that demarcate the reliance on segregated
processing in local modules and information processing
that is distributed more widely across larger neural
redes.
Segundo, evidence for functional integration in body
perception is largely based on correlational data sets. Fur-
ther methodological development will circumvent a reli-
ance on correlational measures of functional connectivity
and increase the prevalence of measures that permit
inferences regarding directional (p.ej., DCM, Granjero
causality), structural (p.ej., imágenes de tensor de difusión),
and causal relationships (p.ej., using neurostimulation
techniques combined with fMRI). Finalmente, functional con-
nectivity studies should embrace best practice from open
ciencia (Munafò et al., 2017). Por ejemplo, an increase in
sample sizes will increase statistical power and may also
permit analyses based on individual differences across
the sample (Dubois & Adolphs, 2016). Además, usando
approaches from neuropsychology, as well as body dis-
orders, has shown promise in understanding mecha-
nisms of body perception and should be used wherever
posible.
Tercero, theories and models of body perception, cual
include functional integration, currently lack detail and
precisión. Updated theories of body perception should
consider integration as much as segregation, así como
the extent to which particular processes are positioned
along a segregation–integration continuum. By doing
entonces, this would build a model of social perception,
which stipulates a relative mix between segregation and
integración. To aid the articulation of such theories,
researchers may consider using theory mapping as a tool
to develop, illustrate, and compare theories (Gray, 2017;
www.theorymaps.org). Theory mapping provides a
common language to visualize theories and store them
en línea, thus promoting easier information exchange.
The development of theories and models will enable
more precise predictions to be made, thus providing a
stronger test of the underlying hypothesis (Meehl,
1990). Harnessing the extensive development of network
science approaches, which include graph theory, will
also be vital for more sophisticated techniques for
specifying and testing models of functional integration
with brain data (bullmore & despreciar, 2009).
Conclusión
Although bodies cue a range of inferences, which are in-
strumental for guiding social behavior, we currently know
little about the neural organization of body perception.
The current review of evidence from fMRI studies dem-
onstrates that body perception is not restricted to pro-
cessing in the ventral visual stream but instead reflects
a functional alliance between the ventral visual stream
and extended neural systems associated with action per-
ception, funciones ejecutivas, and theory of mind. En general,
these findings demonstrate how body percepts are con-
structed through interactions in distributed brain net-
works and underscore that functional segregation and
integration should be considered together when formu-
lating neurocognitive theories of body perception. Por
emphasizing the importance of network science ap-
se acerca, the findings have implications for understand-
ing network models of perception and cognition more
generally, as well as understanding the biological bases
of body image disturbances, such as anorexia nervosa,
which are likely to have a complex biological basis.
Expresiones de gratitud
This work was supported by a grant from the Economic and
Social Research Council (grant ES/K001884/1 to R. r.). I also
thank Emily Cross for feedback on an earlier version of this
manuscript.
Reprint requests should be sent to Richard Ramsey, School of
Psicología, Bangor University, Brigantia Building, Bangor, United
Kingdom, LL57 2AS, o por correo electrónico: r.ramsey@bangor.ac.uk.
Ramsey
1449
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Nota
1. The term “module” refers only to functionally related brain
regiones. It does not refer to additional features that were initially
proposed by Jerry Fodor to define information processing mod-
ules (Fodor, 1983).
REFERENCIAS
Allport, GRAMO. (1954). The nature of prejudice. Cambridge, MAMÁ:
Addison-Wesley Publication Company.
Amodio, D. METRO. (2014). The neuroscience of prejudice and
stereotyping. Naturaleza Reseñas Neurociencia, 15, 670–682.
Aviezer, h., Trope, y., & todorov, A. (2012). Body cues, no
facial expressions, discriminate between intense positive
and negative emotions. Ciencia, 338, 1225–1229.
Baldauf, D., & Desimone, R. (2014). Neural mechanisms of
object-based attention. Ciencia, 344, 424–427.
Bar, METRO. (2004). Visual objects in context. Reseñas de naturaleza
Neurociencia, 5, 617–629.
barrón, h. C., Garvert, METRO. METRO., & Behrens, t. mi. (2016).
Repetition suppression: A means to index neural
representations using BOLD? Philosophical Transactions
of the Royal Society of London, Serie B, Ciencias Biologicas,
371. doi:10.1098/rstb.2015.0355.
Campana, A. h., Hadj-Bouziane, F., Frihauf, j. B., Tootell, R. B., &
Ungerleider, l. GRAMO. (2009). Object representations in the
temporal cortex of monkeys and humans as revealed by
functional magnetic resonance imaging. Diario de
Neurofisiología, 101, 688–700.
Bi, y., Wang, X., & Caramazza, A. (2016). Object domain and
modality in the ventral visual pathway. Tendencias en Cognitivo
Ciencias, 20, 282–290.
Borkenau, PAG., & Liebler, A. (1992). Trait inferences: Fuentes
of validity at zero acquaintance. Journal of Personality and
Social Psychology, 62, 645–657.
Cervecero, METRO. (1988). A dual-process model of impression
formación. En R. S. Wyer Jr. & t. k. Skrull (Editores.), Avances
in social cognition (páginas. 1–36). Hillsdale, Nueva Jersey: Erlbaum.
Cervecero, METRO. B. (1999). The psychology of prejudice: Ingroup
love or outgroup hate? Revista de Temas Sociales, 55, 429–444.
bruce, v., & Joven, A. (1986). Understanding face recognition.
British Journal of Psychology, 77, 305–327.
bullmore, MI., & despreciar, oh. (2009). Complex brain networks:
Graph theoretical analysis of structural and functional
sistemas. Naturaleza Reseñas Neurociencia, 10, 186.
Cabeza, r., & Moscovitch, METRO. (2013). Memory systems, Procesando
modes, and components: Functional neuroimaging evidence.
Perspectives on Psychological Science, 8, 49–55.
de Gelder, B. (2006). Towards the neurobiology of emotional
body language. Naturaleza Reseñas Neurociencia, 7, 242–249.
de Gelder, B., de Borst, A., & watson, R. (2015). The perception
of emotion in body expressions. Wiley Interdisciplinary
Reseñas: Ciencia cognitiva, 6, 149–158.
de Gelder, B., Van den Stock, J., Meeren, h. K., Sinke, C. B.,
Kret, METRO. MI., & Tamietto, METRO. (2010). Standing up for the body.
Recent progress in uncovering the networks involved in the
perception of bodies and bodily expressions. Neurociencia &
Revisiones de biocomportamiento, 34, 513–527.
Diamante, A. (2013). Executive functions. Annual Review of
Psicología, 64, 135–168.
Dodell-Feder, D., Koster-Hale, J., Bedny, METRO., & sajonia, R. (2011).
fMRI item analysis in a theory of mind task. Neuroimagen,
55, 705–712.
Downing, PAG. MI., Jiang, y., Shuman, METRO., & Kanwisher, norte. (2001).
A cortical area selective for visual processing of the human
body. Ciencia, 293, 2470–2473.
Downing, PAG. MI., & Peelen, METRO. V. (2011). El rol de
occipitotemporal body-selective regions in person
percepción. Neurociencia Cognitiva, 2, 186–203.
Downing, PAG. MI., & Peelen, METRO. V. (2016). Body selectivity in
occipitotemporal cortex: Causal evidence. Neuropsicología,
83, 138–148.
Dubois, J., & Adolphs, R. (2016). Building a science of individual
differences from fMRI. Tendencias en Ciencias Cognitivas, 20, 425–443.
duchaína, B., & Yovel, GRAMO. (2015). A revised neural framework
for face processing. Annual Review of Vision Science,
1, 393–416.
Engelen, T., Zhan, METRO., Sack, A. T., & de Gelder, B. (2018).
Dynamic interactions between emotion perception, acción
percepción, and action preparation areas for reacting to social
amenaza: A combined cTBS-fMRI study. eNeuro, ENEURO-0408.
Ewbank, METRO. PAG., Lawson, R. PAG., Henson, R. NORTE., Rowe, j. B.,
Passamonti, l., & Calder, A. j. (2011). Changes in “top–down”
connectivity underlie repetition suppression in the ventral visual
pathway. Revista de neurociencia, 31, 5635–5642.
Fiske, S. T., & Neuberg, S. l. (1990). A continuum of impression
formación, from category-based to individuating processes:
Influences of information and motivation on attention and
interpretación. Advances in Experimental Social Psychology,
23, 1–74.
Fodor, j. A. (1983). The modularity of mind. Cambridge, MAMÁ:
CON prensa.
Friston, k. (2009). Causal modelling and brain connectivity
in functional magnetic resonance imaging. PLoS Biol, 7,
e1000033.
Friston, k. j. (2011). Functional and effective connectivity:
A review. Brain Connect, 1, 13–36.
Friston, k. J., Buechel, C., Fink, GRAMO. r., morris, J., Rolls, MI., &
Dolan, R. j. (1997). Psychophysiological and modulatory
interactions in neuroimaging. Neuroimagen, 6, 218–229.
Frith, C. D., & Frith, Ud.. (1999). Interacting minds—A biological
base. Ciencia, 286, 1692–1695.
Frith, Ud., & Frith, C. (2010). The social brain: Allowing humans
to boldly go where no other species has been. Philosophical
Transactions of the Royal Society, Serie B, Biológico
Ciencias, 365, 165–176.
Fuster, j. METRO. (1997). Network memory. Tendencias en
Neurosciences, 20, 451–459.
Fyock, J., & Estangor, C. (1994). The role of memory biases
in stereotype maintenance. British Journal of Social
Psicología, 33, 331–343.
Gilbert, C. D., & Sigman, METRO. (2007). Brain states: Top–down
influences in sensory processing. Neurona, 54, 677–696.
Gray, k. (2017). How to map theory: Reliable methods are
fruitless without rigorous theory. Perspectives on
ciencia psicológica, 12, 731–741.
Greicius, METRO. D., Krasnow, B., Reiss, A. l., & menón, V. (2003).
Functional connectivity in the resting brain: A network
analysis of the default mode hypothesis. Actas de la
Academia Nacional de Ciencias, EE.UU., 100, 253–258.
Greven, I. METRO., Downing, PAG. MI., & Ramsey, R. (2016). Enlace
person perception and person knowledge in the human
cerebro. Social Cognitive and Affective Neuroscience,
11, 641–651.
Greven, I. METRO., Downing, PAG. MI., & Ramsey, R. (2018). Neural
networks supporting social evaluation of bodies based on
body shape. Social Neuroscience, doi:10.1080/
17470919.2018.1448888.
Greven, I. METRO., & Ramsey, R. (2017a). Person perception involves
functional integration between the extrastriate body area and
temporal pole. Neuropsicología, 96, 52–60.
Greven, I. METRO., & Ramsey, R. (2017b). Neural network integration
during the perception of in-group and out-group members.
Neuropsicología, 106, 225–235.
1450
Revista de neurociencia cognitiva
Volumen 30, Número 10
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Grill-Spector, K., Henson, r., & Martín, A. (2006). Repetition
and the brain: Neural models of stimulus-specific effects.
Tendencias en Ciencias Cognitivas, 10, 14–23.
Harel, A., Kravitz, D. J., & Panadero, C. I. (2014). Task context
impacts visual object processing differentially across the
corteza. procedimientos de la Academia Nacional de Ciencias,
EE.UU., 111, E962–E971.
Haxby, j. v., Hoffman, mi. A., & Gobbini, METRO. I. (2000). El
distributed human neural system for face perception. Trends
in Cognitive Sciences, 4, 223–233.
Jacobo, R. MI., & Schyns, PAG. GRAMO. (2017). Toward a social
psychophysics of face communication. Annual Review of
Psicología, 68, 269–297.
Kanwisher, norte. (2010). Functional specificity in the human brain:
A window into the functional architecture of the mind.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
107, 11163–11170.
Kanwisher, norte. (2017). The quest for the FFA and where it led.
Revista de neurociencia, 37, 1056–1061.
Mamá, NORTE., Vandekerckhove, METRO., Van Overwalle, F., Seurinck, r., &
Fias, W.. (2011). Spontaneous and intentional trait inferences
recruit a common mentalizing network to a different degree:
Spontaneous inferences activate only its core areas. Social
Neurociencia, 6, 123–138.
Macrae, C. NORTE., & Quadflieg, S. (2010). Perceiving people.
In S. t. Fiske, D. t. Gilbert, & GRAMO. Lindzey (Editores.), Manual
of social psychology (páginas. 428–463). Hoboken, Nueva Jersey: wiley.
McLaren, D. GRAMO., Ries, METRO. l., Xu, GRAMO., & Johnson, S. C. (2012). A
generalized form of context-dependent psychophysiological
interactions (gPPI): A comparison to standard approaches.
Neuroimagen, 61, 1277–1286.
Meehl, PAG. mi. (1990). Appraising and amending theories: El
strategy of Lakatosian defense and two principles that
warrant it. Psychological Inquiry, 1, 108–141.
Mesulam, METRO. (1990). Large-scale neurocognitive networks and
distributed processing for attention, idioma, and memory.
Annals of Neurology, 28, 597–613.
mitchell, j. PAG., Cloutier, J., Banaji, METRO. r., & Macrae, C. norte. (2006).
Medial prefrontal dissociations during processing of trait
diagnostic and nondiagnostic person information. Social
Cognitive and Affective Neuroscience, 1, 49–55.
Molenberghs, PAG. (2013). The neuroscience of in-group bias.
Neurociencia & Revisiones de biocomportamiento, 37, 1530–1536.
Munafò, METRO. r., Nosek, B. A., obispo, D. V. METRO., Button, k. S.,
Chambers, C. D., Percie du Sert, NORTE., et al. (2017). A manifesto
for reproducible science. Nature Human Behaviour, 1, 0021.
Olson, I. r., McCoy, D., Klobusicky, MI., & ross, l. A. (2013).
Social cognition and the anterior temporal lobes: A review
and theoretical framework. Social Cognitive and Affective
Neurociencia, 8, 123–133.
O’Reilly, j. X., lana rica, METRO. w., Behrens, t. mi. J., Herrero, S. METRO.,
& Johansen-Berg, h. (2012). Tools of the trade:
Psychophysiological interactions and functional connectivity.
Social Cognitive and Affective Neuroscience, 7, 604–609.
Encima, h., & Cocinar, R. (2018). Where do spontaneous first
impressions of faces come from? Cognición, 170, 190–200.
Parque, H.-J., & Friston, k. (2013). Structural and functional
redes cerebrales: From connections to cognition. Ciencia,
342, 1238411.
Pinsk, METRO. A., Arcaro, METRO., Weiner, k. S., Kalkus, j. F., Inati, S. J.,
Bruto, C. GRAMO., et al. (2009). Neural representations of
faces and body parts in macaque and human cortex: A
comparative fMRI study. Revista de neurofisiología, 101,
2581–2600.
Pinsk, METRO. A., DeSimone, K., moore, T., Bruto, C. GRAMO., & Kastner,
S. (2005). Representations of faces and body parts in
macaque temporal cortex: A functional MRI study.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
102, 6996–7001.
Pourtois, GRAMO., Peelen, METRO. v., Spinelli, l., Seeck, METRO., & Vuilleumier,
PAG. (2007). Direct intracranial recording of body-selective
responses in human extrastriate visual cortex.
Neuropsicología, 45, 2621–2625.
Poyo Solanas, METRO., Zhan, METRO., Vaessen, METRO., Hortensius, r.,
Engelen, T., & de Gelder, B. (2018). Looking at the face and
seeing the whole body. Neural basis of combined face and
body expressions. Social Cognitive and Affective
Neurociencia, 13, 135–144.
Puhl, R. METRO., & Heuer, C. A. (2009). The stigma of obesity: A
review and update. Obesity, 17, 941–964.
Quadflieg, S., Flannigan, NORTE., Waiter, GRAMO. D., rossión, B., Peluca,
GRAMO. S., Turk, D. J., et al. (2011). Stereotype-based modulation
of person perception. Neuroimagen, 57, 549–557.
Ramsey, r., van Schie, h. T., & Cruz, mi. S. (2011). No two are
lo mismo: Body shape is part of identifying others. Cognitivo
Neurociencia, 2, 207–208.
sajonia, r., & Kanwisher, norte. (2003). People thinking about
thinking people. The role of the temporo-parietal junction in
“theory of mind”. Neuroimagen, 19, 1835–1842.
Sell, A., Cosmides, l., Tooby, J., Sznycer, D., von Rueden, C., &
Gurven, METRO. (2009). Human adaptations for the visual
assessment of strength and fighting ability from the body and
rostro. Actas de la Royal Society de Londres, Serie B,
Ciencias Biologicas, 276, 575–584.
despreciar, oh. (2013). Network attributes for segregation and
integration in the human brain. Opinión actual en
Neurobiología, 23, 162–171.
Sterzer, PAG., Kleinschmidt, A., & rees, GRAMO. (2009). el neural
bases of multistable perception. Tendencias en Cognitivo
Ciencias, 13, 310–318.
Suchan, B., Bauser, D. S., Busch, METRO., Schulte, D., Gronemeyer,
D., Herpertz, S., et al. (2013). Reduced connectivity between
the left fusiform body area and the extrastriate body area
in anorexia nervosa is associated with body image distortion.
Behavioural Brain Research, 241, 80–85.
Tajfel, h., Billig, METRO. GRAMO., Bundy, R. PAG., & Flament, C. (1971).
Social categorization and intergroup behavior. European
Journal of Social Psychology, 1, 149–177.
taylor, j. C., Wiggett, A. J., & Downing, PAG. mi. (2007). Funcional
MRI analysis of body and body part representations in the
extrastriate and fusiform body areas. Diario de
Neurofisiología, 98, 1626–1633.
Van Overwalle, F. (2009). Social cognition and the brain: A
meta-analysis. Mapeo del cerebro humano, 30, 829.
Wang, y., collins, j. A., Koski, J., Nugiel, T., Metoki, A., & Olson,
I. R. (2017). Dynamic neural architecture for social
knowledge retrieval. Proceedings of the National Academy
of Sciences, EE.UU., 114, E3305–E3314.
Peelen, METRO. v., & Downing, PAG. mi. (2005). Selectivity for the
Zhan, METRO., Goebel, r., & de Gelder, B. (2018). Ventral and dorsal
human body in the fusiform gyrus. Diario de
Neurofisiología, 93, 603–608.
Peelen, METRO. v., & Downing, PAG. mi. (2017). Category selectivity in
human visual cortex: Beyond visual object recognition.
Neuropsicología, 105, 177–183.
pathways relate differently to visual awareness of body
postures under continuous flash suppression. eNeuro, 5.
Zimmermann, METRO., Toni, I., & el largo, F. PAG. (2013). Body
posture modulates action perception. Diario de
Neurociencia, 33, 5930–5938.
Ramsey
1451
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
0
1
4
4
2
1
7
8
7
7
6
6
/
/
j
oh
C
norte
_
a
_
0
1
2
9
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3