It’s the Other Way Around! Early Modulation of
Sensory Distractor Processing Induced by
Late Response Conflict
Bernhard Pastötter and Christian Frings
Abstracto
■ Understanding the neural processes that maintain goal-
directed behavior is a major challenge for the study of attentional
control. Although much of the previous work on the issue has
focused on prefrontal brain areas, little is known about the
contribution of sensory brain processes to the regulation of atten-
tional control. The present EEG study examined brain oscillatory
activities invoked in the processing of response conflict in a later-
alized Eriksen single-flanker task, in which target letters were
presented at fixation and single distractor letters were presented
either left or right to the targets. Distractors were response
compatible, response incompatible, or neutral in relation to the
responses associated with the targets. The behavioral results
showed that responses to targets in incompatible trials were
slower and more error prone than responses in compatible trials.
The electrophysiological results revealed an early sensory lateral-
ization effect in (both evoked and induced) theta power (3–6 Hz)
that was more pronounced in incompatible than compatible
ensayos. The sensory lateralization effect preceded in time a mid-
frontal conflict effect that was indexed by an increase of (induced)
theta power (6–9 Hz) in incompatible compared with compatible
ensayos. The findings indicate an early modulation of sensory dis-
tractor processing induced by response conflict. Theoretical im-
plications of the findings, in particular with respect to the theory
of event coding and theories relating to stimulus–response bind-
En g [Henson, R. NORTE., Eckstein, D., Waszak, F., Frings, C., & Horner,
A. Stimulus-response bindings in priming. Tendencias en Cognitivo
Ciencias, 18, 376–384, 2014; Hommel, B., Müsseler, J., Aschersleben,
GRAMO., & Príncipe, W.. The theory of event coding (TEC): A framework
for perception and action planning. Behavioral and Brain
Ciencias, 24, 849–878, 2001], are discussed. ■
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
oh
C
norte
_
a
_
0
1
2
6
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INTRODUCCIÓN
Our brain is constantly bombarded with information from
the different senses while at the same time only a small
subset of the current information is actually relevant for
our goals. To make this situation even worse, humanos
typically have several goals activated at a time (p.ej., encima-
arching goals like “avoiding danger” that should be
always active and very specific goals like “searching for
your keys”). The different stimulus information must be
matched to our current goals to promote goal-directed
comportamiento. Obviamente, there might be conflict between
the processing of currently relevant and, for the task at
mano, irrelevant information, which can arise either at
the perceptual level or the level of response selection.
Stimulus conflict arises at the perceptual level when the
relevant information is perceptually different from the
irrelevant information, whereas response conflict arises
at the level of response selection when distracting stimuli
suggest interfering actions. Although it is clear that human
behavior outside the laboratory is quite complex—as we
can juggle several goals with different priorities at a time
with ease and evaluate and process stimuli accordingly—it
University of Trier
© 2018 Instituto de Tecnología de Massachusetts
is nevertheless important to pinpoint aspects of this
complex behavior and investigate the tools with which
humans achieve efficient behavior. Controlling stimulus
and response conflict is one process with which humans
act, although it is clear that there is much more to action
regulation than just controlling stimulus information (p.ej.,
behavioral routines due to binding, self-monitoring, tareas
conjuntos, attentional and intentional weighting, prioritization
due to self-relevance, etcétera).
Respectivamente, research in the cognitive sciences has
developed several experimental paradigms to pinpoint
the control processes with which humans can handle
these conflicts and successfully achieve goal-directed
comportamiento. One task in particular, the Eriksen flanker task,
is well suited to independently measure conflicts at the
level of perception and the level of response selection
(Eriksen & Eriksen, 1974; see Eriksen, 1995, para una revisión).
In the visual variant of this task, flanker or distractor stim-
uli are presented adjacent to a target stimulus. Stimulus
conflict is measured by comparing trials in which the
flankers are identical to the target (es decir., they are stimulus
congruent and response compatible) with trials in which
the flankers are perceptually different from the target
but still suggest the same response (es decir., they are stimulus
incongruent but response compatible). A diferencia de,
Revista de neurociencia cognitiva 30:7, páginas. 985–998
doi:10.1162/jocn_a_01267
response conflict is measured by comparing trials in
which the flankers are perceptually different from the
targets but suggest the same response (es decir., ellos son
stimulus incongruent and response compatible) y
trials in which the flankers are also perceptually different
from the target but, in addition, suggest a different re-
respuesta (es decir., they are stimulus incongruent and response
incompatible). Typically, both stimulus and response
conflict lead to cost effects in the Eriksen flanker, indexed
by slower RTs and more errors in conflict than non-
juicios de conflicto. The costs are typically explained by atten-
tional control processes. One idea is that the internal
representation of the flanker stimuli may be inhibited
or actively suppressed at the perceptual level and also
at the level of response selection, yet inhibition of inter-
fering information needs attentional resources, cual
leads to the observed cost effects (p.ej., Frings, Wentura, &
Wuhr, 2012; Lamers & Roelofs, 2011; Wuhr & Frings, 2008;
Heil, Osman, Wiegelmann, Rolke, & Hennighausen, 2000;
por supuesto, there are other interpretations of the flanker
effect in terms of response biases or response competition
or in terms of an attentional gradient, p.ej., Blanco, ratcliff,
& estrellas, 2011). The effects are not restricted to visual
estímulos. Both stimulus and response conflicts in the flanker
tarea (or variants thereof ) have been well documented in
the literature to arise across the senses, in vision (Eriksen
& Eriksen, 1974), in audition (chan, Merrifield, & Spence,
2005), in touch (evans & Craig, 1992), and even cross-
modally (Frings & Spence, 2010).
Understanding the neural processes that maintain
goal-directed behavior is a major challenge for the study
of attentional control. Much of the previous work on the
issue has focused on prefrontal brain areas, En particular
the ACC and associated regions of the medial frontal wall,
which are believed to play a vital role in the detection of
conflict and the regulation of attentional control. Es un
prominent idea that ACC functions as a conflict monitor
that detects conflicts between concurrently activated rel-
evant and irrelevant information (Botvinick, cohen, &
Carretero, 2004; Van Veen & Carretero, 2002a). Corresponding
evidence arose from fMRI studies showing that ACC is
more activated on conflict than nonconflict trials in
conflict tasks, including the Stroop task (macdonald,
cohen, Stenger, & Carretero, 2000), the Simon task (Peterson
et al., 2002), and the Eriksen flanker task (Botvinick,
nistrom, Fissell, Carretero, & cohen, 1999). Además,
EEG studies have demonstrated that conflicting stimuli or
responses can elicit larger negative deflections in early ERP
componentes (p.ej., N200 and N400) over midfrontal elec-
trodes, likely originating from ACC (p.ej., Apelbaum,
Herrero, Böhler, Chen, & Wolforff, 2011; Hanslmayr et al.,
2008; Van Veen & Carretero, 2002b; Liotti, Wolforff, Pérez, &
Mayberg, 2000). Consistently, both EEG and MEG studies
on brain oscillatory activity have shown that midfrontal
theta oscillations (4–8 Hz) typically show a power increase
in conflict compared with nonconflict trials, an effect that
has been source-localized to ACC (p.ej., Pastötter, Berchtold,
& Bäuml, 2012; Nigbur, Ivanova, & Stürmer, 2011; Pastötter,
Hanslmayr, & Bäuml, 2010; Hanslmayr et al., 2008; ver
Cavanagh & Franco, 2014, para una revisión). Thereby, Tiene
been shown that mainly induced (es decir., not phase-locked)
but less evoked (es decir., phase-locked) theta oscillations are
related to the processing of (respuesta) conflicto (cohen &
Donner, 2013; Cavanagh, Zambrano-Vazquez, & allen,
2012). Although there is evidence that ACC responds to
both stimulus and response conflict, ACC conflict effects
seem to be strongest for conflict arising at the level of
response selection (Nigbur, cohen, Ridderinkhof, &
Stürmer, 2012; Van Veen, cohen, Botvinick, Stenger, &
Carretero, 2001).
In addition to ACC, other brain regions are involved in
conflict processing and attentional control, including the
dorsolateral prefrontal cortex (DLPFC), inferior frontal
gyrus, posterior parietal cortex, anterior insula, and visual
corteza, with the different structures being differentially
related to stimulus and response conflict (see Nee, Apostar,
& En Jonia, 2007, para una revisión). En tono rimbombante, both fMRI and
EEG studies employing the flanker task (or variants thereof )
have shown that early sensory distractor processing in visual
brain areas can be modulated by conflicting stimuli (Marini,
Demeter, Roberts, Chelazzi, & Wolforff, 2016; Donohue,
Todisco, & Wolforff, 2013; Appelbaum et al., 2011; kelly
& La vida, 2011). In the study by Appelbaum et al. (2011),
stimulus and response conflict were varied nonorthog-
onally. Occipital brain activity was contrasted between
conditions with conflicting and (parcialmente) nonconflicting
flankers in a lateralized version of the visual Eriksen
flanker task, in which the distractors were presented left
and right to the targets. In no-conflict trials, all distractors
were nonconflicting (stimulus congruent and response
compatible); in conflict trials, all distractors were conflict-
En g (stimulus incongruent and response incompatible). En
a third condition, the distractors were conflicting in one
hemifield but nonconflicting in the other hemifield. En
these partial-conflict trials, the results showed a sensory lat-
eralization effect over occipital electrode sites, with more
positive ERPs contralateral than ipsilateral to the conflicting
distractor side, that emerged simultaneously in time with a
midfrontal N200 conflict effect. The sensory laterali-
zation effect was suggested to indicate attentional dis-
traction caused by the conflicting flankers. The findings
by Appelbaum et al. (2011), together with the findings
from other fMRI and EEG work (p.ej., Marini et al., 2016;
Donohue et al., 2013; kelly & La vida, 2011), support the
view that occipital brain activity in general and sensory
lateralization in particular are modulated by stimulus
conflict arising at the perceptual level. Sin embargo, porque
of the nonorthogonal manipulation of stimulus and re-
sponse conflict in the earlier work, in principle, both stim-
ulus and response conflict could have contributed to the
modulation of the sensory lateralization effect.
This study examined whether (lateralized) occipital
brain activity is modulated by response conflict arising at
the level of response selection; eso es, we propose the
986
Revista de neurociencia cognitiva
Volumen 30, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
oh
C
norte
_
a
_
0
1
2
6
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
counterintuitive idea that response conflict arising at later
processing stages can influence early sensory processing.
Todavía, with respect to the literature on action control in the
last decade, one can make a clear argument for this idea. En
particular, it has been established that the links between
perception and action are much closer and more inter-
twined than previously assumed. A theory that has been
widely used to investigate action control is the theory of
event coding (TEC; Hommel, Müsseler, Aschersleben, &
Príncipe, 2001), which provides a framework for understand-
ing the linkages between perception and action. Based on
the ideomotor principle (see Shin, Proctor, & Capaldi,
2010; Existencias & Existencias, 2004, for reviews), TEC assumes
that actions are represented by codes of their perceptual
consequences and further that perceived events and
produced actions may share the same representations
(Hommel, 2004, 2009). TEC has been used to explain a
range of phenomena, including stimulus–response (S-R)
effects in general and priming effects in particular (for re-
puntos de vista, see Henson, Eckstein, Waszak, Frings, & Horner,
2014; Henson, 2009). For our purpose here, it is relevant
that TEC assumes that, even after a single pairing of a
stimulus and a response, the S-R episode is bound into a
memory (or event file) and upon repetition of any part
of this episode is again reinstated. En efecto, correspondiente
evidence arose from fMRI and EEG studies, mostrando que
the repetition of a stimulus directly retrieves the response
that was previously associated to it, thereby suggesting
that—after an initial pairing—reencountering a stimulus
makes the previous response instantly accessible (Horner
& Henson, 2012; Dobbins, Schnyer, Verfaellie, & Schacter,
2004).
Concerning our study, we therefore assume that, en un
typical response conflict task with fixed S-R mappings,
reencountering a stimulus after a few trials directly re-
trieves the particular response. Given that research on
action control yields ample evidence that also flanker
stimuli can directly activate responses due to S-R bindings
(p.ej., Frings & Rothermund, 2011; Frings, Rothermund, &
Wentura, 2007; Mayr & Buchner, 2006), we hypothesized
that reencountering a flanker might instantly activate the
associated response, and because of S-R bindings, enterrar-
ference due to response conflict might influence the
processing of flankers at very early levels of sensory infor-
procesamiento de datos. Por supuesto, such view is in contrast to
established sequential information processing models
from research on the psychological refractory period (ver
Pashler, 1994).
This study examined the behavioral and electrophysio-
logical effects of response conflict in a lateralized single-
flanker version of the Eriksen task. This version was
chosen because previous imaging work has shown that
it can produce reliable distractor competition effects in
early visual cortex (kelly & La vida, 2011). Targets were
presented at central fixation, and single flanking dis-
tractors were presented either left or right to the targets.
Distractors were always stimulus incongruent, eso es,
perceptually different from the targets. Response conflict
effects were examined by comparing behavioral and EEG
data between response-incompatible (INC), respuesta-
compatible (COM), and neutral (NEU) condiciones. Regard-
ing the electrophysiological effects of response conflict,
oscillatory brain activities and lateralized readiness poten-
tials (LRPs) were examined. With regard to LRPs, onset
times of stimulus-locked and response-locked potentials
(S-LRP and LRP-R, respectivamente) were analyzed as chrono-
metric markers for the duration of premotor (S-LRP) y
motor processing stages (LRP-R), respectivamente (see Eder,
Leuthold, Rothermund, & Schweinberger, 2012). Con
regard to oscillatory activities, both stimulus-related and
response-related power changes across EEG frequencies
de 1 a 30 Hz were computed and compared between
experimental conditions. Both evoked and induced parts
of the oscillatory signal were calculated. Evoked oscilla-
tions are phase-locked to stimulus or response onset (como
is the ERP), whereas induced oscillations are not (ver
Tallon-Baudry & Bertrand, 1999). Both nonlateralized
and lateralized (to distractor presentation side) effects of
response conflict were examined.
Several expectations arose. Behaviorally, response con-
flict effects were expected for both RT and response accu-
racy, reflected in slower and more error-prone responses
on incompatible than compatible (and neutral) ensayos. En
the electrophysiological level, with respect to LRP onsets,
we expected a response conflict effect on S-LRP onset,
due to later S-LRP onset on incompatible than compatible
and neutral trials, but no conflict effect on LRP-R onset
(p.ej., Kopp, Rist, & Mattler, 1996; graton, Coles, Sirevaag,
Eriksen, & Donchin, 1988). With regard to nonlateralized
EEG activity, we expected an increase of induced (pero
not evoked) theta power over midfrontal electrodes on
incompatible compared with compatible (and neutral)
ensayos (p.ej., cohen & Donner, 2013; Cavanagh et al.,
2012). Most important, with regard to lateralized EEG activ-
idad, we expected a modulation of sensory distractor pro-
cessing induced by response conflict. The hypothesis was
based on the finding that sensory distractor processing can
be modulated by the processing of conflicting information
(p.ej., Appelbaum et al., 2011) and current theories on ac-
tion control and S-R binding, which suggest a close link
between perception and action, and the processing of
stimulus and response conflict, respectivamente (Henson et al.,
2014; Hommel, 2004, 2009; Hommel et al., 2001). Porque
mainly low-frequency oscillations in the theta (alrededor 5 Hz)
and alpha band (alrededor 10 Hz) have been linked to latera-
lized control of visuospatial attention (theta: p.ej., Van der
Lubbe, Bundt, & abrahámico, 2014; Verde & McDonald,
2008; alfa: p.ej., Thut, Nietzel, Brandt, & Pascual Leone,
2006; Worden, Foxe, Wang, & Simpson, 2000), we expected
to find a sensory lateralization effect in theta or alpha
power over occipital electrode sites. As argued above,
theories from action control research would predict that
the flanker stimuli directly activate the associated responses
in parallel with the sensory processing. De este modo, interference
Pastötter and Frings
987
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
oh
C
norte
_
a
_
0
1
2
6
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
due to response conflict might already impact visual
processing of the irrelevant distractors, leading to enhanced
sensory lateralization during the processing of response
incompatible distractors.
MÉTODOS
Participantes
Thirty-four students from the University of Trier, Alemania,
were included in the study (29 women, 6 left-handed,
edad media = 20.7 años, DE = 2.2 años). Six additional
participants were tested but eliminated from analysis
because of excess physiological noise in the EEG data (four
Participantes) or not following task instructions (two par-
ticipants). All participants reported normal or corrected-
to-normal vision; no participant reported any history of
neurological disease. All participants gave written informed
consent before examination and received course credit for
participación. The study was conducted in accordance
with the Declaration of Helsinki and approved by the local
ethics review committee at the University of Trier.
Stimuli and Task
Participants performed a lateralized Eriksen single-flanker
tarea, in which target letters were presented at central fix-
ation and flanking distractor letters were presented either
left or right to the targets. En cada prueba, only a single dis-
tractor letter was shown. Three experimental conditions
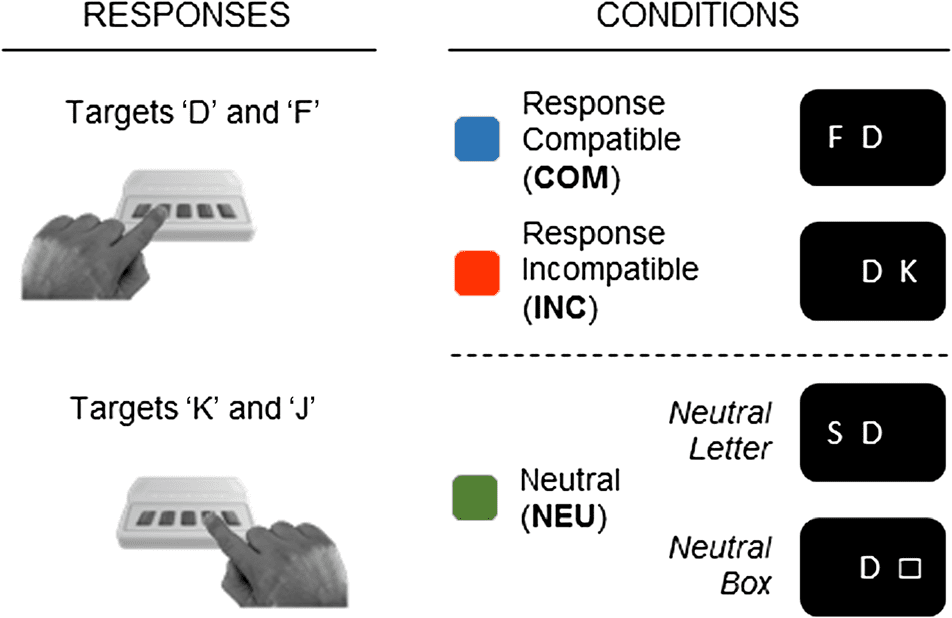
were included in this study (Cifra 1). In all conditions,
target stimuli were the letters D, F, j, and K. In the COM
Cifra 1. Experimental task. Participants performed a lateralized
Eriksen single-flanker task, in which target letters were presented at
central fixation and flanking distractor letters were presented at either
left or right to the targets. Participants responded to target letters
with their left and right index fingers. In the COM condition, targets and
distractors were mapped to the same response, whereas in the INC
condición, targets and distractors were mapped to different responses.
Además, an NEU condition with two neutral-trial types was included,
in which the distractors were not mapped to any responses. Letras
were used as distractors in neutral-letter trials; a box was presented
as distractor in neutral-box trials.
and INC conditions, distractor stimuli were also the letters
D, F, j, and K. The letters “D” and “F” were mapped to a
left-hand response, the letters “J” and “K” were mapped
to a right-hand response. In the COM condition, el dis-
tractors were response compatible to the targets, eso es,
targets and distractors were mapped to the same response
(p.ej., target “D” and distractor “F”). En tono rimbombante, distractors
never matched the targets, eso es, distractors were always
stimulus incongruent to the targets at the perceptual level.
In the INC condition, distractors were response incompat-
ible to the targets, eso es, targets and distractors were
mapped to different responses (p.ej., target “D” and distrac-
tor “K”). The assignment of the single distractor letters to
the target letters was chosen to be one-to-one and kept
constant for all participants (“D”–“K” and “J”–“F”), as it
naturally was in the COM condition (“D”–“F” and
“J”–“K”). Response (en)compatibility effects were exam-
ined by comparing data between compatible and incom-
patible trials in the COM and INC conditions, respectivamente.
Además, a NEU condition with two neutral trial types
was included in the study. In neutral-letter trials, el dis-
tractors were the letters “S,” “G,” “L,” and “H,” which were
not mapped to any response; the assignment of the single
distractor letters to the target letters was chosen to be
one-to-one and kept constant for all participants
(“D”–“‘S,” “F”–“G,” “J”–“L,” and “K”–“H”). In neutral-box
ensayos, a box was presented as distractor (□); the box
was not mapped to any response.
All stimuli were shown in white on black background,
at viewing distance of 65 cm. Stimuli were approximately
1.2° in size. Letters were written in Arial font. Targets
were presented in the center of the screen. Distractors
were centered 3.5° to the left or right of the targets. Ser-
tween displays, a fixation cross was shown in the center
of the screen, which was 0.5° in size. Two response but-
tons were marked on a Chronos response box (Psychol-
ogy Software Tools). Participants were instructed to place
their left index finger on the left button and their right
index finger on the right button of the Chronos response
box (ver figura 1). They were asked to press the left but-
ton in response to the target letters “D” and “F,” and the
right button in response to the target letters “J” and “K.”
Participants were asked to respond as quickly and accu-
rately as possible to the targets and to ignore the flankers.
They were asked to maintain fixation on the center of the
screen at all times and to not move the eyes.
An experimental session consisted of 320 single trials,
presented in 10 blocks of 32 trials each. Within each
block, all predefined combinations of target and distractor
estímulos, with the same number of left- and right-hemifield
distractor presentations, were realized in a counter-
balanced manner. Order of compatible, incompatible,
neutral-letter, and neutral-box trials was randomized with
the constraints that no trial type, target letter, or distractor
hemifield was repeated more than three trials in a row.
Each single trial began with a fixation cross, which was
shown in the center of the screen for an interval of variable
988
Revista de neurociencia cognitiva
Volumen 30, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
oh
C
norte
_
a
_
0
1
2
6
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
duration of 2–2.5 sec. If a participant did not fixate the
cross at the end of this interval, the duration of the inter-
val was prolonged until fixation. Eye gaze was controlled
by simultaneous eye tracking (Eyegaze System, LC Tech-
nológico; calibration was done before the experiment).
Maximum permissible gaze deviation was set to 1.75°
radius from the fixation cross. After presentation of the
fixation cross, the target letter was shown together with
one single distractor letter. The target was shown in the
middle of the screen, and the distractor was shown either
left or right to the target. Target and distractor stimuli re-
mained on the screen until a response was made by the
partícipe. No feedback was provided. After the re-
respuesta, the next single trial started with presentation of
the fixation cross. To familiarize participants with the
procedimiento, they were given one block of 32 practice trials
before the experiment. Presentation and recording of
behavioral responses was done with E-Prime software
(v2.0, Psychology Software Tools).
Analysis of Behavioral Data
Both mean RT and error rate were analyzed. For each
partícipe, trials with RT greater or less than 2.5 standard
deviations from the individual mean RT were excluded
from further analysis. For RT analysis, only trials were in-
cluded for which responses on both the current trial and
the previous trial were correct. Mean numbers of trials
included in the final analysis were 74.9 compatible trials
(DE = 4.1, Min = 65, Max = 80), 74.3 incompatible trials (DE =
4.4, Min = 61, Max = 80), 75.0 neutral-letter trials (DE = 3.5,
Min = 63, Max = 80), y 74.7 neutral-box trials (DE = 4.0,
Min = 66, Max = 80).
Behavioral data were analyzed with repeated-measures
ANOVAs with the factor of Experimental condition (COM,
INC, NEU). Greenhouse–Geisser correction was applied
where appropriate. Post hoc comparisons were conducted
using Tukey honestly significant difference (HSD) pruebas,
which accounts for the problem of multiple testing and
maintains the significance threshold at 5% (HSD.05). Nota
eso, a priori, the behavioral (and EEG) data in the NEU
condition were contrasted between neutral-letter and
neutral-box trials. Como se vio despues, there were no significant
differences between the two neutral trial types, neither in
the behavioral data nor in the EEG data. Por lo tanto, el
present results were based on a single NEU condition,
including both neutral-letter and neutral-box trials.
Recording of EEG Data
Electrophysiological data were recorded from 65 Ag/AgCl
electrodes, which were positioned according to the 10–
10 electrode system with reference to FCz (EC80, Montage
No. 1, Easycap). The ground was placed at location AFz.
The EOG was recorded from four bipolar channels, posi-
tioned on the inferior and superior regions of the left eye
and the outer canthi of both eyes, to monitor the vertical
and horizontal EOG. Electrode-skin impedance was kept
abajo 5 kΩ for all electrodes. Signals were digitalized with
a sampling rate of 500 Hz and amplified between 0.016
y 250 Hz (BrainAmp, BrainVision Recorder, v1.20, Cerebro
Products).
Preprocessing of EEG Data
EEG recordings were re-referenced offline against aver-
age reference and EOG corrected by using calibration
data and generating individual EOG artifact coefficients,
as implemented in BESA Research (v6.1, BESA Software;
see Ille, Iceberg, & Scherg, 2002). Remaining artifacts were
marked by careful visual inspection. EEG data were seg-
mented into epochs ranging from −2.5 to 2.5 sec around
the onsets of stimuli and responses. To avoid filter arti-
facts at the edges of the segments, further analyses were
restricted to intervals ranging from −1.5 to 1.5 sec around
stimulus and response onsets. Segments containing arti-
facts and segments with response errors either on the cur-
rent or the previous trial were discarded from further
análisis.
LRP Analysis
The EEG data were averaged time-locked to stimulus on-
set for S-LRP analysis and response onset for LRP-R anal-
ysis. For each participant and experimental condition, el
ERP at the recording site over the motor cortex ipsilateral
to the response hand was subtracted from the ERP at the
homologous contralateral recording site (C4 minus C3
for left-hand responses, C3 minus C4 for right-hand re-
sponses). The LRP was calculated by averaging the result-
ing difference waveforms across hands, separately for
condiciones. Negative deviations of the resulting LRP indi-
cate stronger activation of the correct response hand
than of the incorrect response hand at the level of the
motor activation.
LRP onsets were measured after low-pass filtering (5 Hz,
12 dB/octave) and analyzed by applying the jackknife-
based procedure (Molinero, Patterson, & Ulrich, 1998). El
S-LRP was aligned to a 0.25-sec baseline before stimulus
onset; the LRP-R was aligned to a 0.25-sec baseline interval
starting at 1.25 sec before response onset (note that the
results and conclusions were the same with an LRP-R base-
line that started 0.55 sec before response onset; see Eder
et al., 2012). S-LRP onset was determined at the point in
time when the LRP amplitude reached 50% of the LRP peak
amplitude, whereas LRP-R onset was calculated with a
fraction of 30% as suggested by Miller et al. (1998). LRP
onset latency measures were submitted to ANOVAs with
F values corrected to Fcorr = F/(n − 1)2, where Fcorr
denotes the corrected F value and n denotes the number
of participants (see Ulrich & Molinero, 2001). Post hoc compar-
isons between conditions were calculated using jackknife-
based tcorr statistics (see Miller et al., 1998).
Pastötter and Frings
989
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
oh
C
norte
_
a
_
0
1
2
6
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Spectral EEG Analysis
The EEG data were transformed into the time–frequency
domain using a complex demodulation algorithm, cual
is implemented in BESA Research (v6.1; see Hoechstetter
et al., 2004). The algorithm consists of a multiplication of
the time domain signal with a complex periodic exponen-
tial function, having a frequency equal to the frequency
under analysis, and subsequent low-pass filtering. El
low-pass filter is a finite impulse response filter of Gaussian
shape in the time domain, which is related to the envelope
of the moving window in wavelet analysis. The data were
filtered in a frequency range from 1 a 30 Hz. Time resolu-
tion was set to 78.8 mseg (FWHM), and frequency resolu-
tion was set to 1.42 Hz (FWHM). Time–frequency data
were exported in bins of 50 msec and 1 Hz.
Both stimulus- and response-locked power changes
were calculated, time-locked to stimulus or response on-
colocar, respectivamente. Stimulus- and response-locked changes
in power were determined by calculating the temporal-
spectral evolution, eso es, power changes for all time–
frequency points with power increases or decreases at
time point t and frequency f related to mean power at fre-
quency f over a preceding baseline interval (Pfurtscheller
& Aranibar, 1977). Stimulus-locked power changes were
determined in relation to a prestimulus baseline interval
that was set from −0.25 to 0 sec time-locked to stimulus
onset, whereas response-locked power changes were de-
termined in relation to a baseline interval that was set
from −1.25 to −1 sec time-locked to response onset. En
addition to total power, both induced and evoked power
changes were estimated. Evoked power changes were
estimated by calculating the temporal-spectral evolution
of the ERP, separately for each condition, electrodo, y
partícipe. Induced power changes were estimated by
calculating the temporal-spectral evolution of the EEG
signal after subtracting the ERP on each trial, separately
for each condition, electrodo, and participant (Kalcher &
Pfurtscheller, 1995).
The following steps were taken to examine the effects
of response (en)compatibility between the COM and INC
condiciones. Primero, the topography of response conflict
effects in total power around response onset (−0.5 to
0.5 segundo, response-locked) between compatible and in-
compatible trials was examined with permutation-based
cluster analysis, which controls for the problem of multiple
pruebas (Maris & Oostenveld, 2007) using BESA Statistics
(v2.0, BESA Software). Específicamente, paired t tests for all
data points (21 [time bins] × 30 [contenedores de frecuencia] × 65 [elec-
trodes]) were calculated and 3-D clusters of contiguous
data points that fell below a p value of .01 in the single
t tests were derived. For each empirical cluster, la suma
of t values of the single significant data points was kept
as a test statistic. Próximo, 5000 random permutations were
run in which the sum test statistic was repeatedly calcu-
lated for randomly shuffled data sets, with the data ran-
domly reordered across COM and INC conditions and
the permutation-based cluster with the highest sum of
t values was kept. By these means, a null distribution was
created from the random permutation runs, and the crit-
ical pcrit values for the empirically derived clusters were
calculated. Empirical clusters with a critical pcrit value
abajo .05 went into further analysis. For those clusters,
mean power changes were calculated for total, evoked,
and induced power, averaged across data points of the
cluster’s maximum time range, maximum frequency
range, and corresponding electrodes, separately for each
experimental condition. Time courses of total power
changes in a cluster were compared between the three
experimental conditions (COM, INC, NEU); time courses
of evoked and induced power change in a cluster were
compared between the COM and INC conditions. Fuerza
changes were time-locked to stimulus and response on-
sets in different analyses. Repeated-measures ANOVAs
that examined the interactions of Experimental condi-
ciones, Time, and Activity (evoked vs. induced) were cal-
culated. In all ANOVAs, Greenhouse–Geisser correction
was applied where appropriate, and Tukey HSD.05 era
used for post hoc testing.
Permutation-based cluster analysis was also used to ex-
amine the topography of (sensorial) lateralization effects in
total power changes related to stimulus onset (0–0.5 sec)
between trials with left-hemifield distractor presentation
and right-hemifield distractor presentation, independiente
of experimental condition (13 [time bins] × 30 [frequency
bins] × 65 [electrodes]). In the permutation analysis, el
data were randomly reordered across left-hemifield and
right-hemifield distractor presentations. Only clusters with
a critical pcrit value below .05 went into further analysis. Como
it turned out, two clusters over the left and right occipital
cortex did show very similar time–frequency characteris-
tics, indicating a sensory lateralization effect over occipital
sites. Por lo tanto, a sensory lateralization index (SLI) era
calculated by subtracting mean power changes of the
electrodes from the cluster ipsilateral to distractor presen-
tation from mean power changes of the electrodes from
the cluster contralateral to distractor presentation, aver-
aged across left- and right-hemifield distractor presenta-
ciones. SLIs were calculated for total, evoked, and induced
power changes, separately for conditions, and the time
courses of sensory lateralization effects were contrasted
time-locked to the onset of stimuli or responses. En todo
ANOVAs, Greenhouse–Geisser correction was applied
where appropriate. Tukey HSD.05 was used for post hoc
pruebas.
RESULTADOS
Behavioral Results
RTs
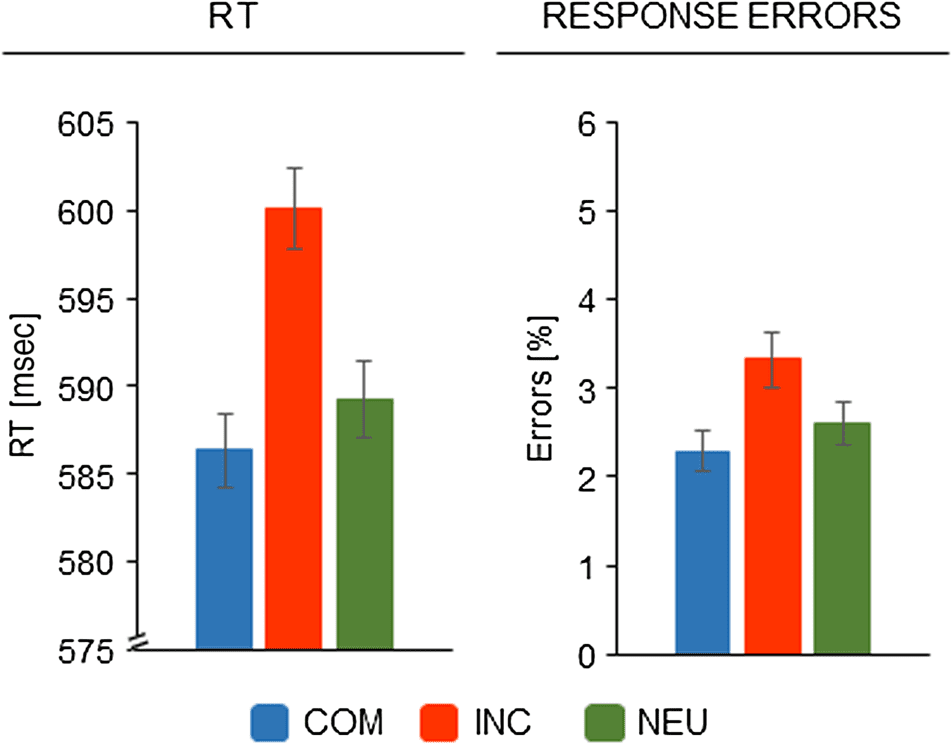
Mean RTs are depicted in Figure 2. A one-way repeated-
measures ANOVA with the factor of Experimental con-
condición (COM vs. INC vs. NEU) revealed a significant effect,
990
Revista de neurociencia cognitiva
Volumen 30, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
oh
C
norte
_
a
_
0
1
2
6
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
suggest a longer duration of premotor processes in
incompatible trials than in compatible and neutral trials.
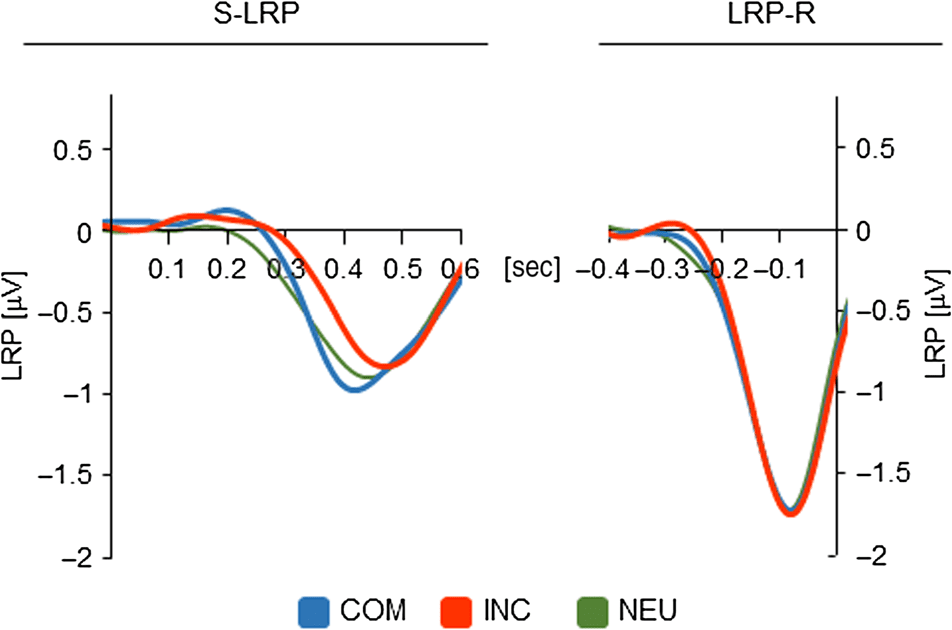
LRP-R
Cifra 3 also shows the LRP-R waveforms. A one-way
repeated-measures ANOVA on LRP-R onset with the factor
of Experimental condition (COM vs. INC vs. NEU) presentado
no significant effect, Fcorr(2, 66) = 0.44, pag = .645. LRP-R onset
was −192.8 msec in the COM condition, −186.8 msec in
the INC condition, and −197.8 msec in the NEU condition
(time-locked to response onset, 30% peak amplitude).
De este modo, the LRP-R results suggest no difference in the dura-
tion of motor processes between experimental conditions.
Physiological Results: Fuerza
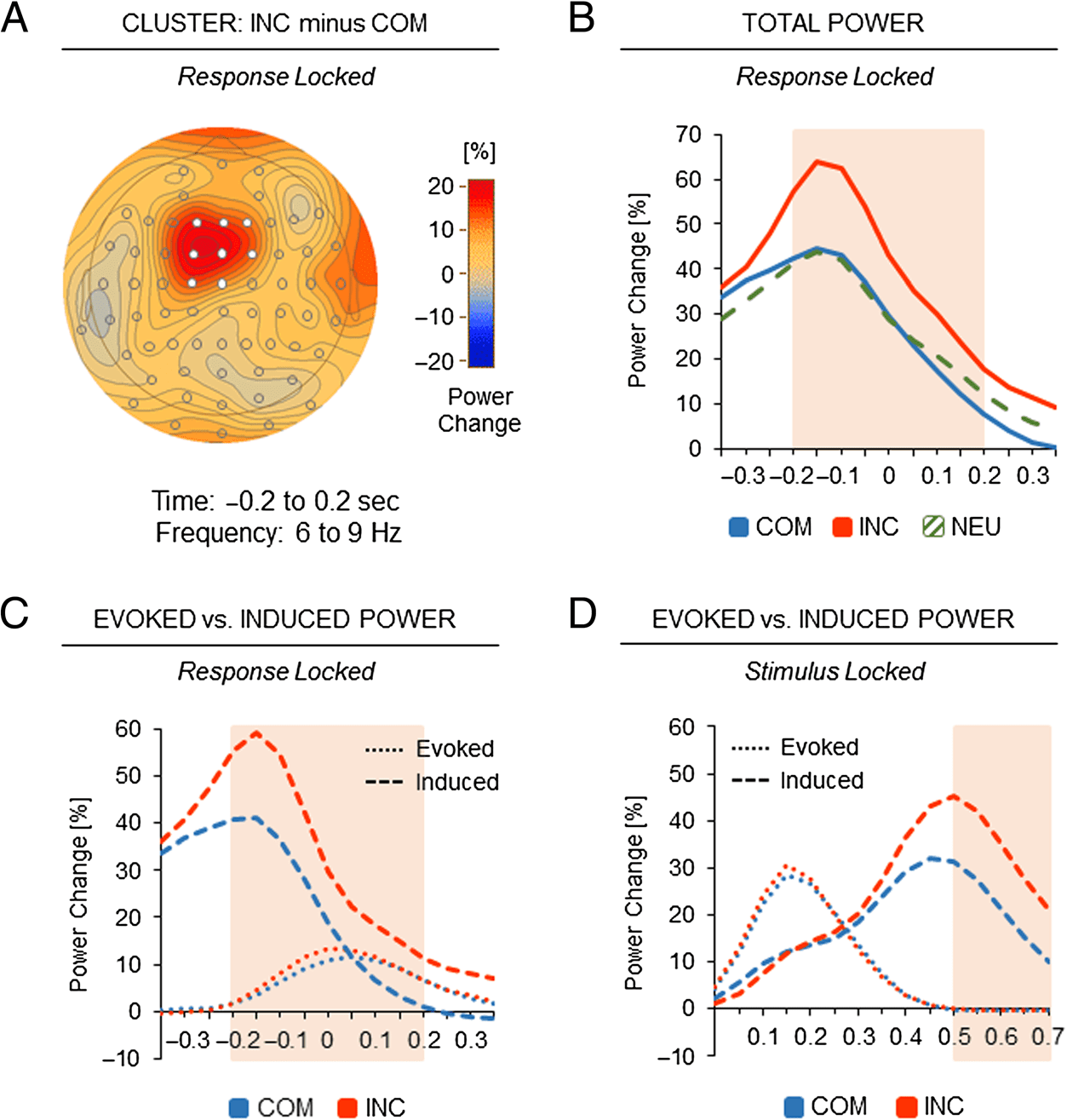
Response Conflict
In the time–frequency analysis of response-locked con-
flict effects, the first step was to examine significant clus-
ters of differences in total power between the COM and
INC conditions. Permutation-based cluster analysis re-
vealed a single significant cluster over midfrontal elec-
trode sites (F1, Fz, F2, FC1, FCz, FC2, C1, and Cz),
which showed a larger response-locked theta power in-
crease (6–9 Hz) in the INC condition than in the COM
condition from −0.2 to 0.2 sec around response onset,
pcrit < .001 (Figure 4A). In the said time window, the
cluster’s total theta power increase in the NEU condition
(31.1%) was significantly smaller than in the INC condi-
tion (45.1%), t(33) = 3.53, p = .001, but comparable with
the COM condition (30.5%), t(33) < 1. Therefore, to in-
crease statistical power in the analysis of conflict effects
in the midfrontal cluster, subsequent analyses contrasted
theta activities between the COM and INC conditions only.
Time courses of total theta power changes in the three
experimental conditions are depicted in Figure 4B.
Figure 3. S-LRP and LRP-R for the three experimental conditions.
S-LRP onset was generally later in the INC condition than in the
other conditions; LRP-R onset did not differ between conditions (COM,
INC, NEU).
Pastötter and Frings
991
Figure 2. Behavioral results. Both mean RTs and error rates showed
reliable response conflict effects, that is, slower and more error-prone
responses on incompatible than compatible trials. In neutral trials,
responding was reliably faster than in incompatible trials; other
differences were not significant. Error bars represent within-subject
standard errors (Cousineau, 2005).
F(2, 66) = 9.38, p < .001. Post hoc testing indicated
that the differences in RT between incompatible trials
(600.1 msec) and compatible trials (586.3 msec) and be-
tween incompatible and neutral trials (589.3 msec) were
significant, whereas the difference between compatible
and neutral trials was not significant (Tukey HSD.05 =
8.0 msec).
Response Errors
Error rates are also shown in Figure 2. A one-way repeated-
measures ANOVA with the factor of Experimental condi-
tion (COM vs. INC vs. NEU) revealed a significant effect,
F(2, 66) = 3.44, p = .038. Post hoc testing showed that
there was a significant difference in error rates between
incompatible (3.32%) and compatible trials (2.29%); the
other differences were not significant (neutral trials: 2.60%;
Tukey HSD.05 = 0.97%).
Physiological Results: LRPs
S-LRP
Figure 3 depicts the S-LRP waveforms for the three exper-
imental conditions. A one-way repeated-measures ANOVA
on S-LRP onset with the factor of Experimental condi-
tion (COM vs. INC vs. NEU) revealed a significant effect,
Fcorr(2, 66) = 3.68, p = .031. Indeed, S-LRP onset in the
INC condition (364.5 msec; time-locked to stimulus onset,
50% peak amplitude) was significantly later than in the
COM condition (330.1 msec), tcorr(33) = 2.05, p = .048,
and the NEU condition (319.0 msec), tcorr(33) = 2.25, p =
.031. The difference between the COM and NEU conditions
was not significant, tcorr(33) < 1. Thus, the S-LRP results
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
o
c
n
_
a
_
0
1
2
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Response conflict.
(A) Topography of the response
conflict effect in midfrontal
theta power (difference
between the INC and COM
conditions). Electrodes in white
depict the significant cluster in
theta power (6–9 Hz; −0.2 to
0.2 sec around response
onset). (B) Response-locked
time course of midfrontal
theta power changes in the
significant cluster as a function
of experimental condition
(NEU). The red-shaded area
indicates significant differences
between incompatible and
compatible trials, p < .05.
(C, D) Response-locked and
stimulus-locked time courses
of midfrontal theta power
changes in the significant cluster
as a function of response conflict
(COM vs. INC). Dotted lines
refer to evoked theta power
modulations; dashed lines
refer to induced theta power
modulations. The red-shaded
areas indicate significant
differences in induced theta
power between compatible and
incompatible trials, p < .05.
Contrasting the time courses of theta activities be-
tween the COM and INC conditions, a repeated-measures
ANOVA with the factors of Condition (COM vs. INC) and
Time (four 100-msec time intervals from −0.2 to 0.2 sec
around response onset) revealed a significant main effect
of Condition, F(1, 33) = 13.94, p < .001, and a significant
main effect of Time, F(3, 99) = 12.67, p < .001, but no sig-
nificant interaction between the two factors, F(3, 99) =
1.21, p = .309. These results suggest that the midfrontal
conflict effect in total theta power was equally present
before and after response execution.
The contribution of evoked and induced theta activi-
ties to the midfrontal conflict effect around response
onset (−0.2 to 0.2 sec) was analyzed by calculating a
repeated-measures ANOVA with the factors of Condition
(COM vs. INC) and Activity (evoked vs. induced). The
analysis showed a significant main effect of Condition,
F(1, 33) = 13.76, p < .001, and a significant main effect
of Activity, F(1, 33) = 11.70, p = .002, due to generally
higher induced than evoked theta power. More im-
portant, the analysis revealed a significant interaction
between the two factors, F(1, 33) = 13.07, p < .001.
Indeed, a significant conflict effect emerged for induced
(COM: 23.0% vs. INC: 36.5%), but not for evoked theta
power (7.5% vs. 8.6%; Tukey HSD.05 = 6.6%). Time
courses of induced and evoked theta activities are depicted
in Figure 4C.
Figure 4D shows the results of the stimulus-locked
analyses. Repeated-measures ANOVAs with the factors
of Condition (COM vs. INC) and Time (seven 100-msec
time intervals from 0 to 0.7 sec following stimulus onset)
were calculated separately for induced and evoked theta
power changes. Concerning induced theta power, the
analysis showed a significant main effect of Condition,
F(1, 33) = 4.60, p = .039, a significant main effect of Time,
F(6, 198) = 10.64, p < .001, and also a significant interac-
tion between the two factors, F(6, 198) = 6.71, p = .001.
Indeed, reliable differences between the COM and INC
conditions arose for the later time intervals from 500 to
700 msec only (means: 23.7% vs. 37.3%; Tukey HSD.05 =
9.55%). Regarding evoked theta power, there was a signif-
icant main effect of time, F(6, 198) = 46.30, p < .001, but
neither a significant main effect of Condition nor a sig-
nificant interaction, both Fs < 1, suggesting that evoked
theta power time-locked to stimulus onset was unaffected
by response conflict.1
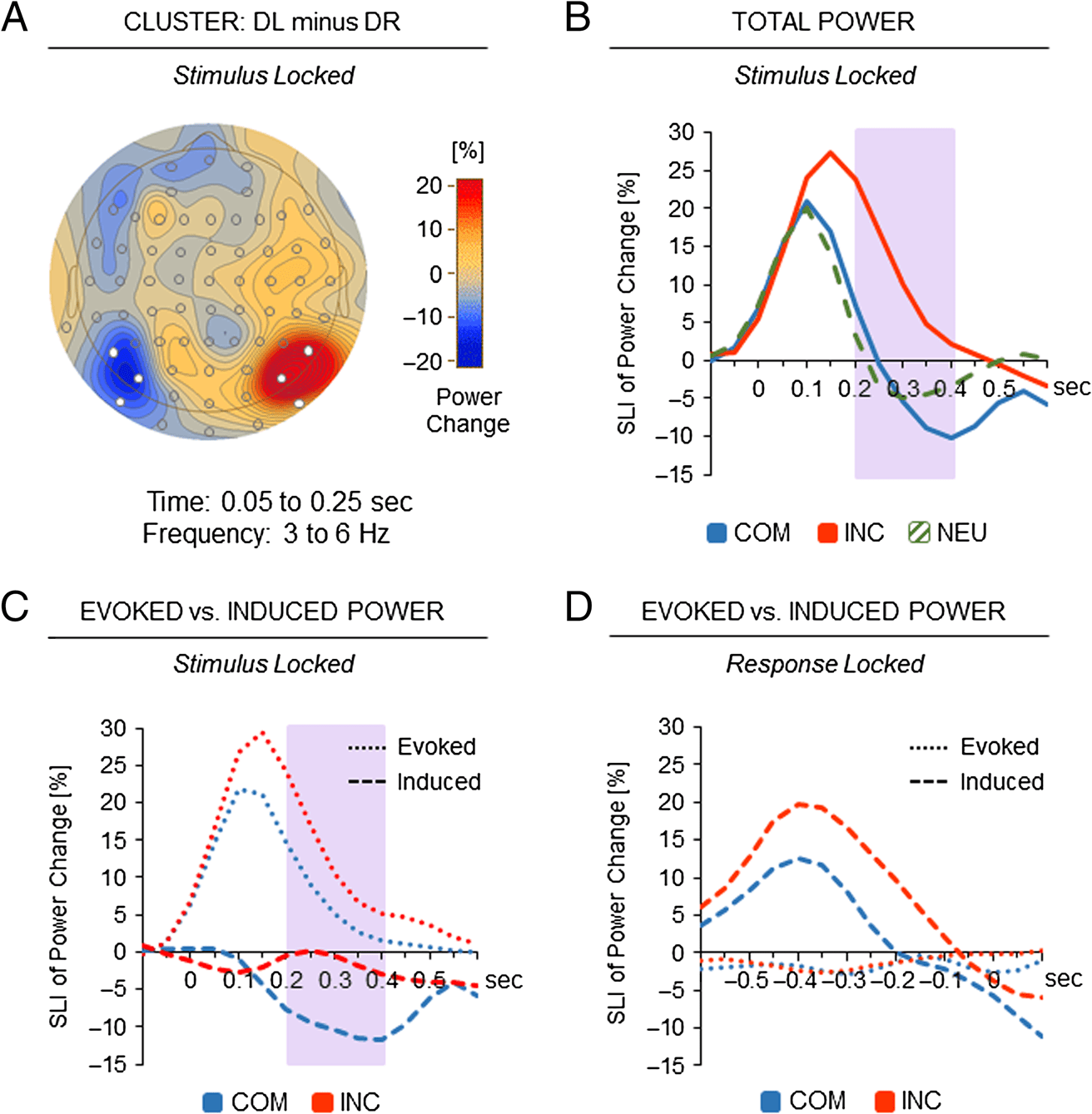
Sensory Lateralization
Examination of lateralization effects in stimulus-locked
power changes between trials with left- and right-hemifield
distractor presentations revealed two significant clusters,
one in the left hemisphere over occipital electrodes P5,
P7, and PO7, pcrit = .015, and one in the right hemisphere
992
Journal of Cognitive Neuroscience
Volume 30, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
o
c
n
_
a
_
0
1
2
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
over occipital electrodes P6, P8, and PO8, pcrit = .019
(Figure 5A). The two clusters showed concurrent theta
power effects with similar time–frequency characteristics
(3–6 Hz; minimum of 0.05 to maximum of 0.25 sec after
stimulus onset), thus indicating a sensory lateralization
effect. Therefore, for each time point, an SLI was calculated
by subtracting mean power changes of the cluster ipsi-
lateral to distractor presentation side from mean power
changes of the cluster contralateral to distractor presen-
tation side, averaged across left and right hemifield presen-
tations. Time courses of the SLI for total power changes
are depicted in Figure 5B, separately for the three experi-
mental conditions.
Following the same steps as in the analysis of the mid-
frontal conflict effect reported above, next, we compared
the time courses of the SLI between the COM and INC
conditions. First, a repeated-measures ANOVA with the
factors of Condition (COM vs. INC) and Time (six 100-msec
time intervals from 0 to 0.6 sec following stimulus onset)
was calculated. The analysis showed a significant main
effect of Condition, F(1, 33) = 4.39, p = .044, a significant
main effect of Time, F(5, 165) = 10.95, p < .001, and also
a significant interaction between the two factors, F(5,
165) = 3.00, p = .013. Indeed, reliable differences be-
tween conditions arose from 0.2 to 0.4 sec after stimulus
onset with stronger sensory lateralization of total theta
power in incompatible than compatible trials (means:
14.1% vs. −1.4%; Tukey HSD.05 = 14.2%). Additional anal-
ysis showed that, in the said time interval, reliable differ-
ences between conditions were similarly present in the
first and second half of the experiment, F(1, 33) = 1.26,
p = .270.
In the analysis of the evoked and induced parts of the
stimulus-locked sensory lateralization effect, theta activi-
ties were averaged from 0.2 to 0.4 sec following stimulus
onset and contrasted between the COM and INC condi-
tions. A repeated-measures ANOVA with the factors of Con-
dition (COM vs. INC) and Activity (evoked vs. induced)
revealed significant main effects of Condition, F(1, 33) =
5.30, p = .028, and activity, F(1, 33) = 23.20, p < .001,
but no significant interaction between the two factors,
F(1, 33) < 1. The results indicate that evoked theta activity
showed a stronger stimulus-locked sensory lateralization
effect than induced theta activity. More important, compa-
rable conflict effects arose for evoked (8.4% vs. 15.0%) and
induced theta activities (−9.7% vs. −0.9%) contributing
Figure 5. Sensory lateralization.
(A) Topography of the sensory
lateralization effect in occipital
theta power (difference
between left distractor
presentation and right
distractor presentation trials).
Electrodes in white depict
the two significant clusters
in theta power (3–6 Hz;
0.05–0.25 sec after stimulus
onset). (B) Stimulus-locked
time course of the SLI (theta
power changes in the cluster
contralateral minus theta power
changes in the cluster ipsilateral
to distractor presentation side)
as a function of experimental
condition (COM, INC, NEU).
The purple-shaded area
indicates significant differences
between incompatible and
compatible trials, p < .05.
(C, D) Response-locked and
stimulus-locked time courses
of the SLI as a function of
response conflict (COM vs.
INC). Dotted lines refer to
the SLI of evoked theta
power changes; dashed lines
refer to the SLI of induced
theta power changes. The
purple-shaded areas indicate
significant differences in
the SLIs of both evoked and
induced theta power changes
between compatible and
incompatible trials, p < .05.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
o
c
n
_
a
_
0
1
2
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Pastötter and Frings
993
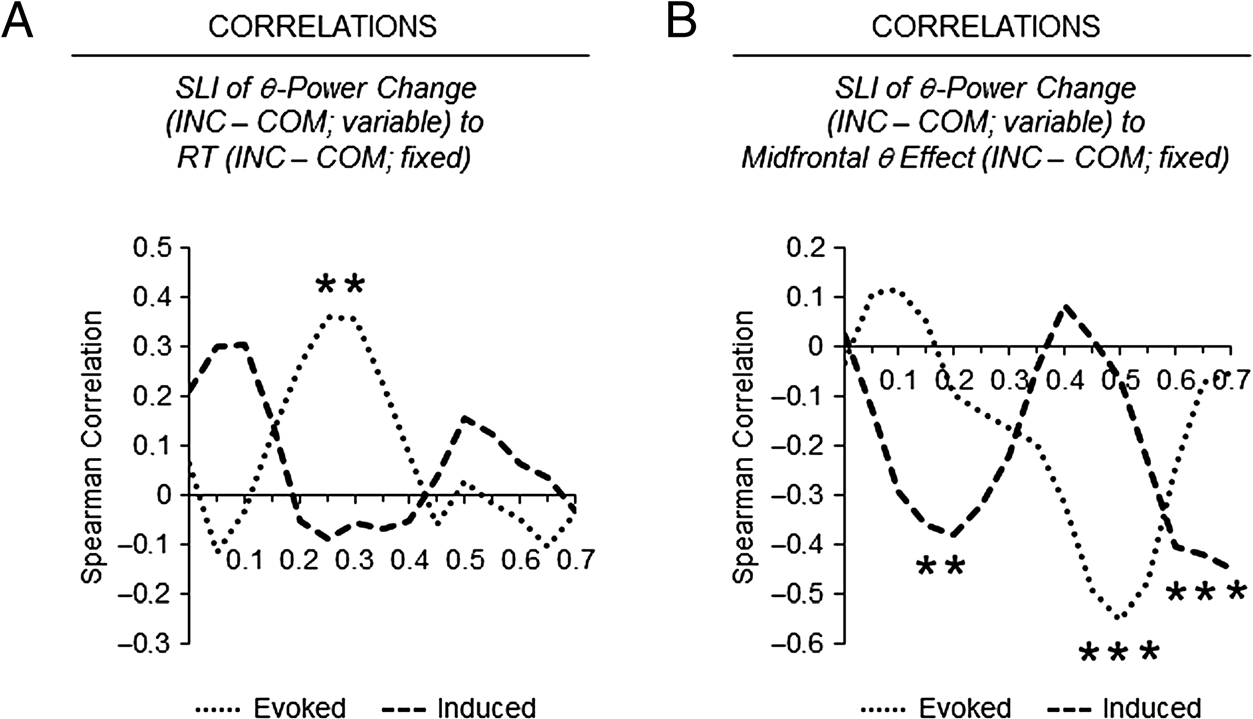
Figure 6. Spearman correlations.
Relationship of the sensory
lateralization effect in evoked
and induced theta power (INC
minus COM; variable across
time) with (A) the behavioral
conflict effect in RT (INC minus
COM; fixed) and (B) the
midfrontal conflict effect in
induced theta power (INC minus
COM; fixed; based on the
significant cluster in the
response-locked analysis shown
in Figure 4A). Asterisks mark
significant correlations, p < .05,
uncorrected.
to the stimulus-locked sensory lateralization effect. The
time courses are depicted in Figure 5C.
In addition, the evoked and induced parts of the
response-locked sensory lateralization effect were calcu-
lated in the 500-msec interval before response onset (see
Figure 5D). Repeated-measures ANOVAs with the factors
of Condition (COM vs. INC) and Time (five 100-msec
time intervals from −0.5 to 0 sec before response onset)
were calculated separately for evoked and induced theta
activities. The analyses showed a main effect of Time for
induced theta, F(4, 132) = 5.93, p = .008, but not for
evoked theta, F(4, 132) < 1. No significant main effect
of Condition was found, neither for induced theta,
F(1, 33) = 2.42, p = .130, nor for evoked theta, F(1,
33) < 1, and also no significant interaction arose, both
Fs < 1. Together with the stimulus-locked analyses
above, these results suggest that the modulation of sen-
sory distractor processing induced by response conflict
was primarily driven by stimulus-related theta activities
and less, if at all, by response-related activities.2
Finally, correlational brain–behavior and brain–brain
analyses were calculated. Figure 6 shows the results. Re-
garding brain–behavior correlations, the results showed
that the behavioral conflict effect in RT (individual
differences between the INC and COM conditions) was
positively related to the conflict effect in the SLI of
evoked theta power change (individual differences
between the INC and COM conditions) from 0.25 to
0.3 sec following stimulus onset (see Figure 6A). Regard-
ing brain–brain correlations, the midfrontal conflict effect
in induced theta power (response-locked; averaged from
−0.2 to 0.2 sec around response onset) was negatively
related to the conflict effect in the SLIs of both evoked
and induced theta power change in different time inter-
vals following the onset of stimuli (see Figure 6B). No
significant correlation between the midfrontal conflict
effect in induced theta power and the behavioral conflict
effect in RT arose, r = .09, p = .599.
DISCUSSION
This study examined the behavioral and electrophysio-
logical effects of response conflict in a lateralized version
of the Eriksen flanker task. Behaviorally, the results
showed effects of response (in)compatibility in both RT
and accuracy, characterized by slower and more error-
prone responses on incompatible than compatible trials,
which is consistent with the findings from previous
behavioral work (e.g., Frings & Spence, 2010; Evans &
Craig, 1992; Eriksen & Eriksen, 1974). At the electro-
physiological level, the results showed a conflict effect
on S-LRP onset, but no conflict effect on LRP-R onset, in-
dicating that conflict affected the duration of premotor
processes but did not influence the duration of motor
processes (see Kopp et al., 1996; Gratton et al., 1988,
for similar findings). With regard to EEG oscillatory activ-
ities, we found a prominent effect of response conflict in
(induced) midfrontal theta power (6–9 Hz), which is con-
sistent with the findings from previous EEG work (e.g.,
Nigbur et al., 2011, 2012). Going beyond the previous
work, the results showed that sensory lateralization of
(both evoked and induced) theta activities (3–6 Hz) over
occipital electrode sites was stronger on incompatible
than compatible trials, thus indicating a modulation of
early sensory distractor processing induced by late re-
sponse conflict.
Earlier research has shown that occipital brain activity in
general and sensory lateralization in particular can be mod-
ulated by the processing of conflicting information in the
Eriksen flanker task (e.g., Marini et al., 2016; Appelbaum
et al., 2011). In this earlier research, occipital brain activity
was contrasted between trials with conflicting and (par-
tially) nonconflicting distractor information. In partial-
conflict trials, the distractors were conflicting (stimulus
incongruent and response incompatible) in one hemifield
but nonconflicting (stimulus congruent and response
compatible) in the other hemifield. Examining ERPs,
994
Journal of Cognitive Neuroscience
Volume 30, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
o
c
n
_
a
_
0
1
2
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Appelbaum et al. (2011) demonstrated a sensory lateraliza-
tion effect in partial-conflict trials, with more positive ERPs
over occipital sites contralateral than ipsilateral to the
conflicting distractor presentation side. The effect arose
from 0.2 to 0.55 sec after stimulus onset, simultaneously
in time with the midfrontal N200 conflict effect. The
researchers proposed that the sensory lateralization effect
reflects attentional distraction toward the conflicting infor-
mation. However, because of the nonorthogonal manipu-
lation of stimulus and response conflict, it is not clear
whether the distraction is caused by stimulus conflict,
response conflict, or both. In this study, we examined
whether occipital brain activity is modulated by response
conflict arising at the response selection level. The results
showed a clear modulation of early sensory distractor
processing induced by response conflict. Indeed, both
evoked and induced theta power over occipital electrode
sites showed a stronger lateralization effect in incompati-
ble than compatible trials from 0.2 to 0.4 sec following
stimulus onset. On the individual subject level, the sensory
lateralization effect in evoked theta power was correlated
with the behavioral conflict effect in RT. Because theta
oscillations in the visual system have been linked to later-
alized control of visuospatial attention (e.g., Van der Lubbe
et al., 2014; Green & McDonald, 2008), the sensory lateral-
ization effect in theta power is suggested to reflect atten-
tional distraction to conflicting distractor information.
This view is consistent with the view by Appelbaum et al.
(2011), with the additional proposal that it is response
conflict at the response selection level that influences the
processing of sensory distractor information. Future EEG
work is needed that manipulates stimulus and response
conflict orthogonally to evaluate the separate contribution
of stimulus and response conflict to the sensory lateraliza-
tion effect in a single experiment.
TEC provides a theoretical framework for explaining
the present results. TEC suggests a close link between
the processes of stimulus perception and response activa-
tion and the processing of stimulus and response conflict,
respectively. TEC assumes that the perception of a stimulus
and the action associated with this stimulus share a com-
mon code and therefore are representationally and func-
tionally related (Hommel, 2004, 2009). The idea is that,
even after a single pairing of a stimulus and a response,
an S-R episode is bound into a common memory, and
upon repetition of any part of the episode, this memory
is instantly retrieved. Imaging studies have suggested that
posterior prefrontal, ventral temporal, and higher-order
visual brain areas contribute to the processing of S-R
bindings (Horner & Henson, 2008; Dobbins et al., 2004).
These brain areas show an increase in activation when
the response associated to a stimulus changes between
stimulus presentations. With respect to the present flanker
task, TEC predicts that—after an initial S-R pairing—
reencountering a flanker stimulus should instantly activate
the S-R episode and directly retrieve the associated re-
sponse. This should lead to fast processing of response
conflict in incompatible trials. In addition, TEC predicts
that—due to S-R bindings—interference due to response
conflict in incompatible trials could influence the process-
ing at early levels of (sensory) information processing. This
is exactly what the present results suggest, showing an
early modulation of lateralized theta power over occipital
electrodes induced by response conflict. Moreover, the
modulation was found to be robust, that is, equally present
in the first and second half of the experiment, which
further corroborates the S-R binding view on the effect.
Indeed, S-R bindings do not need to be gradually learned
but are promptly constructed and stably retained (see
Henson et al., 2014).
In addition to the sensory lateralization effect, a promi-
nent nonlateralized conflict effect in induced theta power
(6–9 Hz) over midfrontal electrode sites was observed.
This finding is in line with previous EEG studies that have
demonstrated reliable conflict effects for induced theta
power around response onset in various conflict tasks
(e.g., Cohen & Donner, 2013; Cavanagh et al., 2012). The
finding is also in line with previous studies that have re-
ported conflict effects in induced theta power regardless
of whether the evoked part of the signal was removed in
the analysis or not (e.g., Cohen & Donner, 2013; Pastötter,
Dreisbach, & Bäuml, 2013; Pastötter et al., 2012). The mid-
frontal conflict effect in theta power has been source local-
ized to ACC (e.g., Töllner et al., 2017; Nigbur et al., 2011;
Pastötter et al., 2010; Hanslmayr et al., 2008), which is be-
lieved to monitor and detect conflict between concurrent
stimuli and responses (Botvinick et al., 2004; Van Veen &
Carter, 2002a). According to the conflict-monitoring view,
ACC is the key structure to initiate the temporal cascade of
processes by which the brain responds to conflict and
regulates attentional control. In this study, the midfrontal
conflict effect in induced theta power began 0.5 sec after
stimulus onset and thus followed in time the occipital con-
flict effect in lateralized theta power. This finding suggests
that the occipital cortex, but not ACC, was the first struc-
ture to “detect” the conflict in the present task. Arguably,
ACC thus may not have “detected” but may have “regis-
tered” conflicting information and “communicated” the
information to other brain areas in the present task. In
fact, the present correlational analyses suggest that the
conflict effect in lateralized theta power over occipital sites
was negatively related to the midfrontal conflict effect in
theta power, which indicates that conflict processing in
the occipital cortex may have even reduced the processing
of conflict in ACC. The finding of a negative correlation is
consistent with recent work showing that early stimulus-
related interregional phase coupling between parietal and
frontal electrode sites in the theta-to-alpha frequency range
is negatively related to midfrontal theta power increase in-
duced by response conflict that was elicited by task-irrelevant
motion direction of stimuli in a feature-based Simon task
(Vissers, Ridderinkhof, Cohen, & Slagter, 2018).
The proposal that ACC registers and communicates
the conflict is in line with the cognitive control literature,
Pastötter and Frings
995
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
o
c
n
_
a
_
0
1
2
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
which assumes that ACC detects (or registers) conflict on a
current trial n and engages control functions in the DLPFC
that bias future processing and reduce (potential) conflict
on the following trial n + 1 (see Carter & van Veen, 2007,
for a review). Consistent with this view, both behavioral
and physiological conflict effects on trial n + 1 have been
found to be modulated by conflict in trial n, indicating
conflict adaptation effects (e.g., Gratton, Coles, & Donchin,
1992). Indeed, imaging studies have shown that ACC activ-
ity is typically reduced on conflict trials following conflict
trials as compared with conflict trials following nonconflict
trials (e.g., Kerns, 2006; Kerns et al., 2004; Botvinick et al.,
1999). Corresponding evidence also arose from EEG stud-
ies that examined the oscillatory signature of the conflict
adaptation effect and showed a sequential modulation in
midfrontal theta power (Töllner et al., 2017; van Driel,
Swart, Egner, Ridderinkhof, & Cohen, 2015; Pastötter et al.,
2013; Cohen & Cavanagh, 2011). Addressing the interplay
between ACC and DLPFC, an intracranial EEG study by
Oehrn et al. (2014) further showed that, in the Stroop task,
the detection (or registration) of conflict in trial n is re-
lated to theta power increase in ACC, whereas the com-
munication of conflict to the DLPFC in trial n + 1 is
mediated by theta phase synchronization between ACC
and DLPFC and an increase in DLPFC gamma power
(30–100 Hz). Addressing the contribution of visual brain
regions to the sequential conflict adaptation effect in a
Stroop task, an fMRI study by Egner and Hirsch (2005)
showed that the processing of relevant targets (i.e., prom-
inent faces) in high-level visual brain regions (i.e., the
fusiform face area) can be enhanced in trials that followed
the processing of conflicting distractors (i.e., incongruent
names), whereas activity in the early visual cortex showed
no conflict adaptation effect. In this study, we found a
modulation of visual processes induced by conflict that
was induced in the same trial n. Future EEG work is needed
that examines possible sequential modulations of early
visual processing in the present flanker and other conflict
tasks (e.g., the accessory Simon task; see Footnote 2) with
higher temporal resolution than fMRI.
To conclude, previous fMRI and EEG work employing
lateralized versions of the Eriksen flanker task have re-
vealed modulations of early sensory distractor processing
induced by conflict but did not further evaluate the sepa-
rate contribution of stimulus and response conflict to
sensory distraction. In the present EEG study, we took
a first step in this direction and examined the contribu-
tion of response conflict to the lateralized sensory dis-
traction effect. The results showed a clear conflict
modulation in lateralized theta power over occipital sites,
indicating that early sensory distractor processing is
modulated by late response conflict. The finding is con-
sistent with theories of action control emphasizing S-R
bindings (Henson et al., 2014; Hommel, 2004, 2009), which
suggest a close link between stimulus and response repre-
sentations, and the processing of stimulus and response
conflict, respectively.
Acknowledgments
The authors thank B. Hommel and one anonymous reviewer
for their comments on a previous version of the manuscript
and T. Brinkmann, J. Paulus, J. Becker, and J. Lohner for their
help with experimental setup and data collection.
Reprint requests should be sent to Bernhard Pastötter, Depart-
ment of Psychology, University of Trier, Universitätsring 15,
54286 Trier, Germany, or via e-mail: pastoetter@uni-trier.de.
Notes
1. Regarding evoked theta power time-locked to stimulus
onset, Figure 4D shows that there was a small but not significant
difference between compatible and incompatible trials at peak
power latency of 150 msec (28.4% vs. 30.6%), t(33) = 1.12, p =
.273. Bayes hypothesis testing showed that the Bayes factor was
3.074, in favor of the null hypothesis over the two-sided alter-
native hypothesis (calculated with default settings in the JASP soft-
ware package; Wagenmakers et al., 2018). This indicates that the
observed data were 3.074 times more likely under the null hypoth-
esis than under the alternative hypothesis, thus providing moder-
ate evidence for the null hypothesis (see Lee & Wagenmakers,
2013), which assumes that evoked theta power (150 msec after
stimulus onset) was unaffected by response conflict.
2. The present flanker task additionally featured a Simon-like
conflict effect, which has been called accessory Simon effect in
the literature (e.g., Nishimura & Yokosawa, 2010). The effect
refers to the finding that responses to centrally located targets
are faster and less error-prone when the presentation side of
the lateralized distractors is spatially congruent with the re-
sponses to the targets. Therefore, we examined whether Simon
congruency interacted with the major outcomes of the present
task. ANOVAs on (median) RT, error rate, SLI of total theta power
change, and induced midfrontal theta power change were calcu-
lated with the additional factor of Simon congruency. All analyses
showed a main effect of Simon congruency, due to slower RT,
more response errors, stronger theta SLI, and higher midfrontal
theta power in Simon-incongruent than Simon-congruent trials,
all ps < .030. Importantly, none of the analyses showed a signif-
icant interaction between Simon congruency and Experimental
condition, all ps > .156, thus suggesting that the major outcomes
in the present task were unaffected by Simon congruency. El
nonfinding of an interaction between Simon congruency and
experimental condition is consistent with the findings from an
earlier behavioral study by Diedrichsen, Ivry, cohen, and Danziger
(2000), which also showed no modulation of response conflict
effects induced by Simon congruency (but did find such modula-
tion of stimulus-conflict effects). Notablemente, the present EEG finding
of a main effect of Simon congruency on theta SLI indicates that
the modulation of early sensory distractor processing induced by
(respuesta) conflict may not be restricted to the present flanker
task but may generalize to other (respuesta) conflict tasks.
REFERENCIAS
Apelbaum, l. GRAMO., Herrero, D. v., Böhler, C. NORTE., Chen, W.. D.,
& Wolforff, METRO. GRAMO. (2011). Rapid modulation of sensory
processing induced by stimulus conflict. Revista de Cognitivo
Neurociencia, 23, 2620–2628.
Botvinick, METRO., nistrom, l. MI., Fissell, K., Carretero, C. S., & cohen,
j. D. (1999). Conflict monitoring versus selection-for-action
in anterior cingulate cortex. Naturaleza, 402, 179–181.
Botvinick, METRO. METRO., cohen, j. D., & Carretero, C. S. (2004). Conflicto
monitoring and anterior cingulate cortex: An update. Trends
in Cognitive Sciences, 8, 539–546.
996
Revista de neurociencia cognitiva
Volumen 30, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
oh
C
norte
_
a
_
0
1
2
6
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Carretero, C. S., & van Veen, V. (2007). Anterior cingulate cortex
and conflict detection: An update of theory and data.
Cognitivo, Afectivo, & Neurociencia conductual, 7, 367–379.
Cavanagh, j. F., & Franco, METRO. j. (2014). Theta frontal como
mecanismo de control cognitivo. Tendencias en Cognitivo
Ciencias, 18, 414–421.
Cavanagh, j. F., Zambrano-Vazquez, l., & allen, j. j. (2012).
Theta lingua franca: A common mid-frontal substrate for
action monitoring processes. Psychophysiology, 49, 220–238.
chan, j. S., Merrifield, K., & Spence, C. (2005). Auditory spatial
attention assessed in a flanker interference task. Acta
Acustica United With Acustica, 91, 554–563.
cohen, METRO. X., & Cavanagh, j. F. (2011). Single-trial regression
elucidates the role of prefrontal theta oscillations in response
conflicto. Fronteras en psicología, 2, 30.
cohen, METRO. X., & Donner, t. h. (2013). Midfrontal conflict-related
theta-band power reflects neural oscillations that predict
comportamiento. Revista de neurofisiología, 110, 2752–2763.
Cousineau, D. (2005). Confidence intervals in within-subject
designs: A simpler solution to Loftus and Masson’s method.
Tutorial in Quantitative Methods for Psychology, 1, 42–45.
Diedrichsen, J., Ivry, R. B., cohen, A., & Danziger, S. (2000).
Asymmetries in a unilateral flanker task depend on the
direction of the response: The role of attentional shift and
perceptual grouping. Revista de Psicología Experimental:
Percepción y desempeño humanos, 26, 113–126.
Dobbins, I. GRAMO., Schnyer, D. METRO., Verfaellie, METRO., & Schacter, D. l.
(2004). Cortical activity reductions during repetition priming
can result from rapid response learning. Naturaleza, 428, 316–319.
Donohue, S. MI., Todisco, A. MI., & Wolforff, METRO. GRAMO. (2013). El
rapid distraction of attentional resources toward the source
of incongruent stimulus input during multisensory conflict.
Revista de neurociencia cognitiva, 25, 623–635.
Eder, A. B., Leuthold, h., Rothermund, K., & Schweinberger,
S. R. (2012). Automatic response activation in sequential
affective priming: An ERP study. Social Cognitive and
Affective Neuroscience, 7, 436–445.
Adecuado, T., & Hirsch, j. (2005). Mecanismos de control cognitivo
resolver conflictos a través de la amplificación cortical de tareas relevantes
información. Neurociencia de la naturaleza, 8, 1784–1790.
Eriksen, B. A., & Eriksen, C. W.. (1974). Effects of noise letters
on the identification of target letters in a non-search task.
Percepción & Psicofísica, 16, 143–149.
Eriksen, C. W.. (1995). The flankers task and response
competition: A useful tool for investigating a variety of
cognitive problems. Visual Cognition, 2, 101–118.
evans, PAG. METRO., & Craig, j. C. (1992). Response competition: A
major source of interference in a tactile identification task.
Atención, Percepción, & Psicofísica, 51, 199–206.
Frings, C., & Rothermund, k. (2011). To be or not to be…
included in an event file: Integration and retrieval of
distractors in stimulus-response episodes is influenced by
perceptual grouping. Revista de Psicología Experimental:
Aprendiendo, Memoria, & Cognición, 37, 1209–1227.
Frings, C., Rothermund, K., & Wentura, D. (2007). Distractor
repetitions retrieve previous responses to targets. Quarterly
Revista de Psicología Experimental, 60, 1367–1377.
Frings, C., & Spence, C. (2010). Crossmodal congruency effects
based on stimulus identity. Brain Research, 1354, 113–122.
Frings, C., Wentura, D., & Wuhr, PAG. (2012). On the fate of
distractor representations. Revista de experimentación
Psicología: Percepcion humana & Actuación, 38, 570–575.
graton, GRAMO., Coles, METRO. GRAMO., & Donchin, mi. (1992). Optimizing
the use of information: Control estratégico de activación de
respuestas. Revista de Psicología Experimental: General,
121, 480–506.
graton, GRAMO., Coles, METRO. GRAMO., Sirevaag, mi. J., Eriksen, C. w., &
Donchin, mi. (1988). Pre- and poststimulus activation of
response channels: A psychophysiological analysis. Diario
of Experimental Psychology: Human Perception and
Actuación, 14, 331–344.
Verde, j. J., & McDonald, j. j. (2008). Electrical neuroimaging
reveals timing of attentional control activity in human brain.
Biología PLoS, 6, e81.
Hanslmayr, S., Pastötter, B., Bäuml, K.-H., Gruber, S., Wimber,
METRO., & Klimesch, W.. (2008). The electrophysiological
dynamics of interference during the Stroop task. Diario
of Cognitive Neuroscience, 20, 215–225.
Heil, METRO., Osman, A., Wiegelmann, J., Rolke, B., & Hennighausen,
mi. (2000). N200 in the Eriksen task: Inhibitory executive
procesos? Journal of Psychophysiology, 14, 218–225.
Henson, R. norte. (2009). Priming. en l. Squire, t. Albright, F.
Bloom, F. Gage, & norte. Spitzer (Editores.), Encyclopedia of
neurociencia (páginas. 1055–1063). Ámsterdam: Elsevier.
Henson, R. NORTE., Eckstein, D., Waszak, F., Frings, C., & Horner, A.
(2014). Stimulus-response bindings in priming. Tendencias en
Cognitive Sciences, 18, 376–384.
Hoechstetter, K., Bornfleth, h., Weckesser, D., Ille, NORTE., Iceberg, PAG.,
& Scherg, METRO. (2004). BESA source coherence: A new method
to study cortical oscillatory coupling. Brain Topography, 16,
233–238.
Hommel, B. (2004). Event files: Feature binding in and across
perception and action. Tendencias en Ciencias Cognitivas, 8,
494–500.
Hommel, B. (2009). Action control according to TEC (theory
of event coding). Investigación psicológica, 73, 512–526.
Hommel, B., Müsseler, J., Aschersleben, GRAMO., & Príncipe, W.. (2001).
The theory of event coding (TEC): A framework for
perception and action planning. Behavioral and Brain
Ciencias, 24, 849–878.
Horner, A. J., & Henson, R. norte. (2008). Priming, respuesta
learning and repetition suppression. Neuropsicología, 46,
1979–1991.
Horner, A. J., & Henson, R. norte. (2012). Incongruent abstract
stimulus–response bindings result in response interference:
fMRI and EEG evidence from visual object classification
priming. Revista de neurociencia cognitiva, 24,
760–773.
Ille, NORTE., Iceberg, PAG., & Scherg, METRO. (2002). Artifact correction of the
ongoing EEG using spatial filters based on artifact and brain
signal topographies. Journal of Clinical Neurophysiology,
19, 113–124.
Kalcher, J., & Pfurtscheller, GRAMO. (1995). Discrimination between
phase-locked and non-phase-locked event-related EEG activity.
Electroencephalography and Clinical Neurophysiology, 94,
381–384.
kelly, t. A., & La vida, norte. (2011). Working memory load
modulates distractor competition in primary visual cortex.
Corteza cerebral, 21, 659–665.
Kerns, j. GRAMO. (2006). Corteza cingulada anterior y prefrontal
activity in an fMRI study of trial-to-trial adjustments on the
tarea simon. Neuroimagen, 33, 399–405.
Kerns, j. GRAMO., cohen, j. D., macdonald, A. w., III, Dar, R. y.,
Stenger, V. A., & Carretero, C. S. (2004). cingulado anterior
Monitoreo de conflictos y ajustes en el control.. Ciencia, 303,
1023–1026.
Kopp, B., Rist, F., & Mattler, Ud.. (1996). N200 in the flanker task
as a neurobehavioral tool for investigating executive control.
Psychophysiology, 33, 282–294.
Lamers, METRO. j. METRO., & Roelofs, A. (2011). Attentional control
adjustments in Eriksen and Stroop task performance can be
independent of response conflict. The Quarterly Journal of
Psicología experimental, 64, 1056–1081.
Sotavento, METRO. D., & carpinteros, E.-J. (2013). Bayesian cognitive
modelado: A practical course. Cambridge: Cambridge
Prensa universitaria.
Pastötter and Frings
997
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
oh
C
norte
_
a
_
0
1
2
6
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Liotti, METRO., Wolforff, METRO. GRAMO., Pérez, r., & Mayberg, h. S. (2000).
An ERP study of the temporal course of the Stroop color-word
interference effect. Neuropsicología, 38, 701–711.
macdonald, A. w., cohen, j. D., Stenger, V. A., & Carretero, C. S.
(2000). Dissociating the role of the dorsolateral prefrontal
and anterior cingulate cortex in cognitive control. Ciencia,
288, 1835–1838.
Marini, F., Demeter, MI., Roberts, k. C., Chelazzi, l., & Wolforff,
METRO. GRAMO. (2016). Orchestrating proactive and reactive
mechanisms for filtering distracting information:
Brain–behavior relationships revealed by a mixed-design
fMRI study. Revista de neurociencia, 36, 988–1000.
Maris, MI., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Revista de neurociencia
Métodos, 164, 177–190.
Mayr, S., & Buchner, A. (2006). Evidence for episodic retrieval
of inadequate prime responses in auditory negative priming.
Revista de Psicología Experimental: Percepcion humana
& Actuación, 32, 932–943.
Molinero, J., Patterson, T., & Ulrich, R. (1998). Jackknife-based
method for measuring LRP onset latency differences.
Psychophysiology, 35, 99–115.
Nee, D. MI., Apostar, t. D., & En Jonia, j. (2007). Interference
resolution: Insights from a meta-analysis of neuroimaging tasks.
Cognitivo, Afectivo, & Neurociencia conductual, 7, 1–17.
Nigbur, r., cohen, METRO. X., Ridderinkhof, k. r., & Stürmer, B.
(2012). Theta dynamics reveal domain-specific control over
stimulus and response conflict. Revista de Cognitivo
Neurociencia, 24, 1264–1274.
Nigbur, r., Ivanova, GRAMO., & Stürmer, B. (2011). Theta power as a
marker for cognitive interference. Clinical Neurophysiology,
122, 2185–2194.
Nishimura, A., & Yokosawa, k. (2010). Visual and auditory
accessory stimulus offset and the Simon effect. Atención,
Percepción, & Psicofísica, 72, 1965–1974.
Oehrn, C. r., Hanslmayr, S., Fell, J., Deuker, l., Kremers, norte. A.,
Do Lam, A. T., et al. (2014). Neural communication patterns
underlying conflict detection, resolution, and adaptation.
Revista de neurociencia, 34, 10438–10452.
Pashler, h. (1994). Dual-task interference in simple tasks: Datos
and theory. Boletín Psicológico, 116, 220–244.
Pastötter, B., Berchtold, F., & Bäuml, K.-H. t. (2012). Oscillatory
correlates of controlled speed-accuracy tradeoff in a
response-conflict task. Mapeo del cerebro humano, 33,
1834–1849.
Pastötter, B., Dreisbach, GRAMO., & Bäuml, K.-H. t. (2013). Dynamic
adjustments of cognitive control: Oscillatory correlates of the
efecto de adaptación al conflicto. Revista de Cognitivo
Neurociencia, 25, 2167–2178.
Pastötter, B., Hanslmayr, S., & Bäuml, K.-H. t. (2010). Conflicto
processing in the anterior cingulate cortex constrains
response priming. Neuroimagen, 50, 1599–1605.
Peterson, B. S., kane, METRO. J., Alexander, GRAMO. METRO., Lacadie, C.,
Skudlarski, PAG., Leung, H.-C., et al. (2002). An event-related
functional MRI study comparing interference effects in the
Simon and Stroop tasks. Cognitive Brain Research, 13,
427–440.
Pfurtscheller, GRAMO., & Aranibar, A. (1977). Event-related cortical
desynchronization detected by power measurements of scalp
EEG. Electroencephalography and Clinical Neurophysiology,
42, 817–826.
espinilla, Y. K., Proctor, R. w., & Capaldi, mi. j. (2010). Una revisión de
contemporary ideomotor theory. Boletín Psicológico,
136, 943–974.
Existencias, A., & Existencias, C. (2004). A short history of ideo-motor
acción. Investigación psicológica, 68, 176–188.
Tallon-Baudry, C., & Bertrand, oh. (1999). Oscillatory gamma
activity in humans and its role in object representation.
Tendencias en Ciencias Cognitivas, 3, 151–162.
Thut, GRAMO., Nietzel, A., Brandt, S. A., & Pascual Leone, A. (2006).
Alpha-band electroencephalographic activity over occipital
cortex indexes visuo-spatial attention bias and predicts visual
target detection. Revista de neurociencia, 26, 9494–9502.
Töllner, T., Wang, y., Makeig, S., Müller, h. J., Jung, t. PAG., &
Gramann, k. (2017). Two independent frontal midline theta
oscillations during conflict detection and adaptation in a
Simon-type manual reaching task. Revista de neurociencia,
37, 2504–2515.
Ulrich, r., & Molinero, j. (2001). Using the jackknife-based scoring
method for measuring LRP onset effects in factorial designs.
Psychophysiology, 38, 816–827.
Van der Lubbe, R. h., Bundt, C., & abrahámico, mi. l. (2014).
Internal and external spatial attention examined with lateralized
EEG power spectra. Brain Research, 1583, 179–192.
van Driel, J., Swart, j. C., Adecuado, T., Ridderinkhof, k. r., &
cohen, METRO. X. (2015). (No) time for control: Frontal theta
dynamics reveal the cost of temporally guided conflict
anticipation. Cognitivo, Afectivo, & conductual
Neurociencia, 15, 787–807.
Van Veen, v., & Carretero, C. S. (2002a). The anterior cingulate as a
conflict monitor: fMRI and ERP studies. Physiology &
Comportamiento, 77, 477–482.
Van Veen, v., & Carretero, C. S. (2002b). The timing of action-
monitoring processes in the anterior cingulate cortex.
Revista de neurociencia cognitiva, 14, 593–602.
Van Veen, v., cohen, j. D., Botvinick, METRO. METRO., Stenger, V. A., &
Carretero, C. S. (2001). Anterior cingulate cortex, conflicto
supervisión, and levels of processing. Neuroimagen, 14,
1302–1308.
Vissers, METRO. MI., Ridderinkhof, k. r., cohen, METRO. X., & Slagter, h. A.
(2018). Oscillatory mechanisms of response conflict elicited
by color and motion direction: An individual differences
acercarse. Revista de neurociencia cognitiva, 30, 468–481.
carpinteros, E.-J., Amar, J., Marsman, METRO., Jamil, T., Ly, A.,
Verhagen, A. J., et al. (2018). Bayesian inference for
psicología. Part II: Example applications with JASP.
Boletín psiconómico & Revisar, 25, 58–76.
Blanco, C. NORTE., ratcliff, r., & estrellas, j. j. (2011). Modelos de difusión
de la tarea de flanqueador: Atención discreta versus gradual
selección. Psicología cognitiva, 4, 210–238.
Worden, METRO. S., Foxe, j. J., Wang, NORTE., & Simpson, GRAMO. V. (2000).
Anticipatory biasing of visuospatial attention indexed by
retinotopically specific alpha-band electroencephalography
increases over occipital cortex. Revista de neurociencia, 20,
RC63.
Wuhr, PAG., & Frings, C. (2008). A case for inhibition: Visual
attention suppresses the processing of irrelevant objects.
Revista de Psicología Experimental: General, 137, 116–130.
998
Revista de neurociencia cognitiva
Volumen 30, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
0
7
9
8
5
1
7
8
7
5
8
3
/
j
oh
C
norte
_
a
_
0
1
2
6
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3