La memoria a corto plazo resistente a los distractores está respaldada por
Transient Changes in Neural Stimulus Representations
Jan Derrfuss1,2, Matthias Ekman1, Michael Hanke3,4,
Marc Tittgemeyer5, and Christian J. Fiebach1,6,7
Abstracto
■ Goal-directed behavior in a complex world requires the mainte-
nance of goal-relevant information despite multiple sources of dis-
traction. Sin embargo, the brain mechanisms underlying distractor-
resistant working or short-term memory (STM) are not fully under-
permaneció. Although early single-unit recordings in monkeys and fMRI
studies in humans pointed to an involvement of lateral prefrontal
cortices, more recent studies highlighted the importance of poste-
rior cortices for the active maintenance of visual information also in
the presence of distraction. Aquí, we used a delayed match-to-
sample task and multivariate searchlight analyses of fMRI data to
investigate STM maintenance across three extended delay phases.
Participants maintained two samples (either faces or houses) across
an unfilled pre-distractor delay, a distractor-filled delay, and an un-
filled post-distractor delay. STM contents (faces vs. houses) podría
be decoded above-chance in all three delay phases from occipital,
temporal, and posterior parietal areas. Classifiers trained to distin-
guish face versus house maintenance successfully generalized from
pre- to post-distractor delays and vice versa, but not to the distrac-
tor delay period. Además, classifier performance in all delay
phases was correlated with behavioral performance in house, pero
not face, ensayos. Our results demonstrate the involvement of distrib-
uted posterior, but not lateral prefrontal, cortices in active mainte-
nance during and after distraction. They also show that the neural
code underlying STM maintenance is transiently changed in the
presence of distractors and reinstated after distraction. El
correlation with behavior suggests that active STM maintenance
is particularly relevant in house trials, whereas face trials might rely
more strongly on contributions from long-term memory. ■
INTRODUCCIÓN
Short-term memory is the ability to actively maintain task-
relevant information over brief periods of time. Monkeys
and humans can maintain such information even when
distractors intervene between encoding and recall. en un
now classic study, Molinero, Erickson, and Desimone
(1996) showed that individual lateral prefrontal neurons
maintained sample-selective delay activity in a delayed
match-to-sample (DMS) task despite multiple distractors
intervening between sample and probe, with sample-
selective delay activity defined as increased delay-related
activity to the “preferred” stimulus of the respective neu-
ron. Recording from prefrontal neurons that showed a
modulation of activity in at least one phase of their DMS
tarea, Miller et al. (1996) showed sample- selective delay
activity in 28% of these neurons. This finding led them
to conclude that the lateral pFC plays an important role
in distractor-resistant STM.
1Radboud University Nijmegen, 2University of Nottingham, 3Otón
von Guericke University, Magdeburg, Alemania, 4Center for
Behavioral Brain Sciences, Magdeburg, Alemania, 5Max Planck
Institute for Metabolism Research, Cologne, Alemania, 6Goethe
University Frankfurt, 7Center for Individual Development and
Adaptive Education, Frankfurt am Main, Alemania
© 2017 Instituto de Tecnología de Massachusetts
Curiosamente, sin embargo, it seems that neurons with
sample-selective delay activity did not show sample selec-
tivity while the distractors were presented (ver figura 5
in Miller et al., 1996). De este modo, the question arises—how is
information about the sample maintained during distrac-
tor presentation—a question that was not addressed by
Miller and colleagues. We hypothesize that, in principle,
there are two possible mechanisms by which sample-
related information may survive distractor delays: active
and passive maintenance. Active maintenance would in-
volve persistent neuronal activity, although the level of
neuronal activity might be substantially reduced relative
to stimulus presentation. As this form of maintenance re-
quires ongoing neuronal activity, its metabolic effects
should in principle be detectable with fMRI. Passive main-
tenance, por otro lado, could be described as a state
of heightened accessibility of information (como, p.ej.,
assumed in the working memory model of Cowan,
2001), in the absence of active maintenance and thus also
without persistent neuronal firing or increased fMRI
activación.
Only a limited number of studies have so far investi-
gated short-term or working memory maintenance in
the presence of distractors. In one of these studies, Jacob
and Nieder (2014) used a delayed match-to-numerosity
Revista de neurociencia cognitiva 29:9, páginas. 1547–1565
doi:10.1162/jocn_a_01141
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
oh
6
C
5
norte
9
_
6
a
/
_
j
0
oh
1
C
1
norte
4
1
_
a
pag
_
d
0
1
b
1
y
4
gramo
1
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
.
t
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
paradigm to study distractor-resistant STM in macaque
monkeys while recording simultaneously from lateral
pFC and the ventral intraparietal area ( VIP). Monkeys
had to maintain information about the number of dots
shown as sample across a pre-distractor delay, a distrac-
tor delay, and a post-distractor delay; distractors were
also dots, but their numerosity was task-irrelevant. In line
with Miller et al. (1996), Jacob and Nieder found that pre-
frontal neurons displayed sample-selective delay activity
even after distraction. Going beyond Miller et al., Jacob
and Nieder also studied neuronal activity during the
distractor delay itself and found that distractors interfered
with numerosity representations in pFC, suggesting that
STM representations may not be actively maintained in
the pFC while distractors are presented. Unexpectedly,
sin embargo, Jacob and Nieder found that VIP neurons
displayed distractor-resistant response properties: Many
VIP neurons maintained sample selectivity even in
the distractor delay. These findings suggest that poste-
rior, not lateral prefrontal, cortices play a major role in
representing distractor-resistant information in STM.
Initial support for this hypothesis for humans comes
from an fMRI DMS study with faces and shoes as samples
y distractores (2 × 2 factorial design), reported by Jha,
Fabian, and Aguirre (2004). These authors observed in-
creased activity in the right fusiform face area (FFA) cuando
participants maintained faces in memory and faces were
at the same time presented as distractors. This could sug-
gest that FFA was involved both in maintaining the mem-
oranda and processing the distractors. Sin embargo, más
detailed analyses did not fully support this assumption,
leading the authors to conclude that heightened FFA ac-
tivity might in fact be due to increased FFA inhibition to
filter out distractors when samples and distractors come
from the same category, or due to an interaction of main-
tenance and inhibition. More recently, Bettencourt and
Xu (2016) investigated the roles of visual areas V1 to V4
and of a load-sensitive segment of the posterior intra-
parietal sulcus in distractor-resistant STM. They demon-
strated that the orientation of grating stimuli maintained
in memory could be decoded from the posterior intra-
parietal sulcus during the presence of distraction (DMS
tarea; delay length = 11 segundo). Además, cuando el
presence of distractors was not predictable, distractor-
resistant maintenance was also found in lower visual
areas V1–V4. Estos hallazgos, de este modo, provide converging
evidence that, also in humans, posterior cortices are
involved in the distractor-resistant maintenance of infor-
mation in STM.
Sin embargo, although the studies of Bettencourt and Xu
(2016) and Jha et al. (2004) suggested that—at least un-
der some conditions—there is active maintenance of
information during distractor delays, this has not been
a consistent finding. Using a variant of the DMS task,
Lewis-Peacock, Drysdale, Oberauer, and Postle (2012;
see also Lewis-Peacock & Postle, 2012) found that the
sample category could not be decoded once the distrac-
tor presentation commenced, despite successful perfor-
mance on the subsequent probe presentation. En cambio,
these authors were able to decode the category of the
distractor pictures, even though these were irrelevant
to the task. In Lewis-Peacock et al.’s (2012) Experimento
2, two samples from two different stimulus categories
were encoded. Participants initially maintained both
muestras (Delay 1: 8 segundo), then were cued to attend to
one of the samples (Delay 2: 7.5 segundo), before being cued
to continue attending to the same sample or to switch
their attention to the other sample (Delay 3: 8 segundo). Dur-
ing Delay 2, only the category of the attended sample
could be decoded, even though the unattended sample
might again become relevant again in Delay 3 and in that
case also could be decoded during Delay 3. The results of
Lewis-Peacock et al. suggest that successful performance
in STM tasks does not always have to rely on active main-
tenance of sample information; in tasks involving diver-
sion of attention away from STM contents, it seems to
rely partly on what we have described above as passive
maintenance and a subsequent reactivation or recovery
stage once the distraction ends (cf. sprague, Ester, &
serencias, 2016). As a result of this, the sample information
would then again be held in STM (and thus become acces-
sible to fMRI-based decoding). There is some evidence that
such recovery after distraction is supported by medial-
temporal lobe structures (cf. Sakai & Passingham, 2004;
Sakai, Rowe, & Passingham, 2002a).
Previously, a number of further influential fMRI studies
have investigated the effects of distractors presented in
the encoding phase or the effects of briefly presented dis-
tractors in the delay phase (p.ej., Bloemendaal et al., 2015;
Clapp, Rubens, & Gazzaley, 2010; Zanto & Gazzaley,
2009; Yoon, Curtis, & D'Esposito, 2006; Gazzaley,
Cooney, Rissman, & D'Esposito, 2005). Sin embargo, as a
result of their design, these studies provide no further
evidence regarding whether or not STM contents are
actively maintained in the distractor delay and, if so,
whether or not the maintenance-related neural processes
in the distractor delay differ from those in unfilled delays.
To investigate these questions, the present fMRI study
utilized a variant of the DMS task in which participants
were asked to maintain two faces or two houses over
three extended delay periods, with distractors being pre-
sented in the middle delay period (Cifra 1). This design
allowed us to apply multivariate pattern analysis (MVPA)
a (i) decode which type of stimulus (faces vs. houses) es
maintained in memory within each individual delay
período (within-delay analyses) as well as to (ii) investigate
the generalization of activity patterns across delay pe-
riods (across-delay analyses). By presenting pictures of
faces and houses as well as scrambled pictures as distrac-
tores, we furthermore investigated (iii) how the percep-
tual similarity of distractors and samples affected the
decodability of sample information. In addition to the
DMS task, we acquired an independent data set to func-
tionally localize face-preferential and house-preferential
1548
Revista de neurociencia cognitiva
Volumen 29, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
oh
6
C
5
norte
9
_
6
a
/
_
j
0
oh
1
C
1
norte
4
1
_
a
pag
_
d
0
1
b
1
y
4
gramo
1
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
t
/
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Cifra 1. The DMS task
employed in this study.
Participants had to encode and
maintain two sample pictures
(faces or houses) across three
delay periods. The first delay
period was unfilled (pre-
distractor delay). For both face
and house trials, the middle
delay period was filled with
distractor pictures, which were
either intact or scrambled
(distractor delay). Overlaid on
the distractors was a slight blue
or red color gradient (exaggerated in the figure to increase visibility) to which participants responded with index or middle finger button presses,
respectivamente. The final delay period was again unfilled (post-distractor delay). At the end of the trial, a probe picture was presented that
matched one of the sample pictures with a probability of 50%. Acerca de 25% of the probe stimulus was unpredictably covered to discourage the
reliance on specific, salient features during stimulus encoding. All pictures were presented individually in the center of the screen.
brain regions to investigate if these stimulus-preferential
areas (in particular FFA and the parahippocampal place
área [PPA]) overlap with areas exhibiting above-chance
decoding in the DMS task.
MÉTODOS
Participantes
Twenty-two participants (10 women) took part in the ex-
perimento. One male participant was excluded because of
multiple movements clearly exceeding the size of one
voxel in the functional runs. The mean age of the remain-
En g 21 participants was 27.1 años (DE = 3.3, range =
22–34 years). All participants were right-handed (laterality
quotient of >50 in the Edinburgh Inventory; Oldfield,
1971), had normal or corrected-to-normal vision, and re-
ported no deficits in color vision. No participant reported
a history of neurological, major medical, or psychiatric dis-
orden. The study was approved by the local ethics commit-
tee, and written consent was obtained from all participants.
Design and Stimuli of the DMS Task
This study employed a variant of the DMS task (Cifra 1).
On each trial, participants encoded either two faces or
two houses (2.1 sec each, intertrial interval = 0.1 segundo)
and maintained these stimuli across three delay periods.
The first delay was unfilled (fixation cross, 8.8 segundo). El
second delay was filled with six distractor pictures
(6.6 segundo, 1.1 sec per distractor): In the object condition,
pictures of three faces and three houses were sequential-
ly presented as distractors (distractors came from both
categories to keep the distractor-related visual input for
faces and houses constant in all trials). In the scrambled
condición, six phase-scrambled pictures were presented.
To ensure that distractors were attended, all distractor
pictures were overlaid with a slight blue or red color gra-
dient, and participants had to indicate the color by a but-
ton press. Responses were given with the right index
finger (azul) or the right middle finger (rojo); maximum
RT was 1.1 segundo. The order of colors and distractor catego-
ries was randomized with the constraint that the same
color or distractor category could not appear more than
twice in a row. The distractor delay was followed by a
final delay phase that was again unfilled (fixation cross,
8.8 segundo). Próximo, a probe picture was presented. The prob-
ability of a match to one of the samples was 50%, equated
over the two samples. Responses were again given with
the right index finger (match) or the right middle finger
(no-match; maximum RT = 3 segundo). Acerca de 25% del
probe stimulus were unpredictably covered, either in ver-
tical or horizontal direction (cf. Cifra 1), to ensure that
participants could not successfully perform the task by
simply encoding a specific feature of the stimulus (p.ej.,
an unusually shaped mouth) and compare this feature
with the probe stimulus. En general, there were four condi-
ciones: face samples/object distraction, face samples/
scrambled distraction, house samples/object distraction,
and house samples/scrambled distraction. Each trial was
followed by intertrial intervals of varying lengths (4.4, 6.6,
o 8.8 segundo), resulting in overall trial lengths of 35.2, 37.4,
o 39.6 segundo (uniform distribution, controlled on a per-
condition basis).
The task was presented in two runs. Each run lasted
24.2 min and began with four dummy scans (to achieve
steady-state magnetization), followed by a rest phase of
44 segundo (used for echo weighting, see below). cada carrera
involved three task blocks of 7.5 min each. After each
task block, participants received feedback about their ac-
curacy (separately for the memory probes and the color
decision task; 2 segundo) and rested for 17.6 segundo. Each block
consistió en 12 ensayos (three per condition). Respectivamente,
72 trials were presented overall (18 per condition). En
each block, direct repetitions of conditions were exclud-
ed, and the remaining transitions were counterbalanced.
Each block had six match and six no-match trials. Más-
encima, after two blocks the number of match and no-
match trials for each condition was equalized (as there
were three trials per condition per block, this was not
possible within a single block).
Derrfuss et al.
1549
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
oh
6
C
5
norte
9
_
6
a
/
_
j
0
oh
1
C
1
norte
4
1
_
a
pag
_
d
0
1
b
1
y
4
gramo
1
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
/
t
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
All pictures were presented individually in grayscale in
the center of the screen and subtended a visual angle of
2.6 × 3.5°. Pictures were shown only once as samples
(de este modo, every encoding period involved two pictures that
were not seen before). Sin embargo, the no-match probes
were previously presented as sample stimuli to make
sure that participants could not base their decision solely
on stimulus familiarity (es decir., on whether or not they had
seen a particular face or house before). En general, 72 rostro
y 72 house pictures were presented as samples and
probes (if the very first trial of the experiment was a
no-match trial, one additional probe picture was present-
ed). The face pictures came from a set assembled by Endl
et al. (1998), depicting men photographed in front of a
uniform gray background (ver figura 1). House pictures
came from a set of face and house pictures assembled by
Piekema, Kessels, Rijpkema, and Fernandez (2009). El
left and right edges of house pictures were cropped to
achieve the same aspect ratio as that of the face pictures
(ver figura 1). We only chose pictures in which the
house was still clearly visible after cropping.
Distractors also came from the set assembled by Piekema
et al. (2009) but were not used as samples or probes. Dis-
tractors were shown twice, with a minimum of 36 enterrar-
vening distractors before a repetition of the same picture
could occur. To generate scrambled distractors, MATLAB
version 2014b (The MathWorks, Natick, MAMÁ) fue usado.
For each picture, a fast Fourier transform was performed.
Entonces, random phase information was added and an in-
verse fast Fourier transform was performed to generate
the scrambled picture. The color gradient overlaid onto
the distractors was Gaussian-shaped with the highest
color intensity located in the middle of the image. Encima-
todo, 54 face and 54 house distractors were used.
Design and Stimuli of the Functional Localizer Task
In the functional localizer task, participants had to decide
for each presented stimulus whether or not it matched
the immediately preceding stimulus (1-back task; bien
index finger: match; right middle finger: nonmatch).
There were separate blocks for face and house stimuli,
y 10 trials were presented per block. Half of the blocks
of each stimulus category had one 1-back target and the
other half had two 1-back targets, to ensure that partici-
pants would maintain attention up to the end of the
block. Participants were instructed not to respond to
the first stimulus in each block (as a 1-back decision is
not possible). The 1-back run lasted about 7 min and be-
gan with four dummy scans (to achieve steady-state mag-
netization), followed by a rest phase of 44 segundo (used for
echo weighting, see below). Participants performed six
face and six house blocks in alternating order; 50% comenzar-
ing with a face block and the other half with a house
block. A block lasted for 15 sec and was followed by a
15-sec rest period. Pictures were presented for 1 segundo,
followed by a blank screen of 500 mseg. The pictures
used were a randomly selected subset of the pictures
used as samples in the DMS task. No pictures apart from
the 1-back targets were repeated.
Procedures
After being welcomed, participants were given general in-
formation about the study, received an MRI participant
information form, and gave informed consent. Próximo, ellos
were given detailed instructions for the DMS task. El
instructions stressed the requirement to encode and
maintain the stimuli in a holistic manner (as opposed
to individual features) and to actively rehearse the stimuli
across all three delay phases. Después, Participantes
were given 12 practice trials (three of each condition) a
familiarize themselves with the task. Stimuli used in the
practice trials were not used in the experiment proper.
Próximo, participants received instructions for and per-
formed one face block and one house block from the
1-back task (again using stimuli not presented in the
scanner). Entonces, participants were taken to the MRI room
and scanning commenced. All participants performed the
DMS task first, followed by the 1-back task. After leaving
the scanner, participants filled in a postexperimental
questionnaire and were debriefed.
Data Acquisition
Imaging was performed using a 3-T Siemens Magnetom
Trio scanner (Siemens Medical Solutions, Erlangen,
Alemania). In both runs of the DMS task, 660 images with
28 axial slices (3.2 mm in-plane resolution, 3.3 mm slice
thickness, 20% spacing) parallel to the AC–PC plane were
acquired using a multiecho EPI sequence (Poser,
Versluis, Hoogduin, & Norris, 2006) and a 32-channel
head coil. The repetition time was 2,200 mseg, the flip an-
gle was 90°, and the echo times were 9.4, 21.2, 33, 45, y
57 mseg. The field of view was 205 × 205 mm2. The first
four images were discarded from the analysis. The next
20 images were rest scans that were later used to calculate
weighting images (see below). Identical scanning param-
eters were used for the 1-back task, but only 190 images
were acquired per run. In a separate scanning session, a
high-resolution 3-D T1-weighted data set with 1 × 1 ×
1.25 mm3 resolution was acquired.
Análisis de los datos
Behavioral Data Analysis
Behavioral and ROI data were analyzed with IBM SPSS 22
(IBM Corp., Armonk, Nueva York) and IPython notebook (Pérez
& Granjero, 2007). Unless otherwise noted, error bars in
line and bar plots represent 95% within-subject confi-
dence intervals (CIs; Morey, 2008; Cousineau, 2005).
The calculations of Cohen’s d disregarded correlations,
1550
Revista de neurociencia cognitiva
Volumen 29, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
oh
6
C
5
norte
9
_
6
a
/
_
j
0
oh
1
C
1
norte
4
1
_
a
pag
_
d
0
1
b
1
y
4
gramo
1
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
/
.
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
eso es, the data were treated as between-subject data
(Dunlap, Cortina, Vaslow, & Burke, 1996).
Preprocessing of fMRI Data
fMRI data processing was carried out using FSL (FMRIB’s
Software Library; Smith et al., 2004). As we used a multi-
echo sequence with five echoes, the reconstructed data
consisted of five complete time series for each run, uno
for every echo time. In the first preprocessing step, estos
time series were combined into a single time series per
run. Para tal fin, we used the time series from the first
echo to compute motion correction parameters using
MCFLIRT ( Jenkinson, Bannister, Brady, & Herrero, 2002).
These parameters were then applied to all time series.
Próximo, the five time series were split into two parts, corre-
sponding to the rest periods acquired at the beginning of
each run (the “weighting time series”) and the task prop-
es (the “task time series”). The data corresponding to the
weighting time series were then used to compute weight-
ing images as described previously (Poser et al., 2006).
Brevemente, the weight of a voxel in a particular weighting im-
age depends on its mean signal strength and variability
relative to this voxel’s values in the other weighting time
series. The weight will be high if a voxel’s signal is strong
and stable in a particular time series relative to the other
time series. One weighting image was computed for each
weighting time series. Próximo, the five task time series were
multiplied by their corresponding weighting images and
added up to create a single time series that was used for
the remainder of the analysis.
The following preprocessing steps were then applied
within FEAT (versión 5.98): nonbrain removal using
BET (Herrero, 2002), grand mean intensity normalization
of the entire 4-D data set by a single multiplicative factor,
and high-pass temporal filtering (Gaussian-weighted least
squares straight line fitting, 128 segundo). Finalmente, spatial
smoothing was applied with a Gaussian kernel of 5 mm
FWHM.
Modeling and Multivariate Analysis
Following the approach proposed by Mumford, Tornero,
Ashby, and Poldrack (2012), the sample phase and all
three delay phases of every single trial were modeled in
separate general linear models (GLM) using a double-
gamma function; for each such GLM, all other events con-
tributed to a single regressor of no interest. The sample
(es decir., memory encoding) phase was modeled by an epoch
de 4.4 sec length. Unfilled delays (es decir., the pre-distractor
delay and the post-distractor delay) were modeled by a
4.4-sec epoch placed in the middle of the 8.8-sec delay
período (Zarahn, Aguirre, & D'Esposito, 1997). Distractor
delays were modeled by 6.6-sec epochs. Motion correc-
tion parameters obtained during echo combination were
used as confound regressors. Para cada participante, the pa-
rameter estimates (es decir., one beta image per trial period)
from Run 2 were then spatially registered to Run 1, based
on transformation matrices obtained from registering the
middle volume of Run 2 to the middle volume of Run 1
using six degrees of freedom and normalized correlation
as a cost function. To prepare the data for the multivariate
análisis, the parameter estimates from all trial phases of
all correctly answered trials were concatenated in the
order in which they were acquired. If a participant made
no errors, this resulted in a 4-D file with 288 volumes (es decir.,
72 ensayos, parameter estimates for sample phase and three
delay phases). For each image in the concatenated file, un
attribute file labeled its sample category (es decir., face or
house) and its phase in the trial (es decir., sample, pre-distractor,
distractor, post-distractor). A separate set of attribute files
was created that also labeled the type of distraction (object
or scrambled).
PyMVPA (Hanke et al., 2009; www.pymvpa.org/) era
then used for linear detrending and z-transforming the
data for each cross-validation fold. A searchlight analysis
(Kriegeskorte, Goebel, & Bandettini, 2006) with a radius
of two voxels (33 voxels per searchlight) and a support
vector machine (SVM) classifier with PyMVPA’s default
C parameter (which automatically scales C according to
the norm of the data) with cross-validation was per-
formed to identify voxels that distinguished between
maintenance of face versus house information in the dif-
ferent delay phases. The chunks used for cross-validation
were the blocks into which the runs were subdivided.
De este modo, for each participant, there were six chunks overall.
For each cross-validation fold, five chunks were used for
classifier training and the remaining chunk was used for
classifier testing. For each fold of the cross-validation pro-
cedure, we made sure that an equal number of training
samples was available for face and house trials (a different
approach was used for the analysis that correlated behav-
ioral and decoding performance—see below). An exam-
ple might help to illustrate this: There was a maximum of
30 correct trials per sample category in five chunks (es decir.,
5 blocks × 6 trials per condition and block). If a partici-
pant had made no house trial errors but two errors on
face trials, solo 28 face trials remained for training. El
training procedure ensured that an equal number of
house trials (es decir., 28) was then randomly chosen from
el 30 available ones. To mitigate the effects of randomly
selecting a subset for one category, we repeated this
selection process five times for each fold and averaged
the classifier performance across the five repetitions
(given that differences in trial numbers were typically
very small, further increasing the number of repetitions
had very little effect on the results).
Classifier performance was evaluated by calculating the
balanced accuracy averaged across the six folds. The bal-
anced accuracy results from calculating accuracies per
target category initially, before averaging over target cat-
egories. The advantage of this measure is that it is inde-
pendent of the relative frequency of the target categories
in a chunk used for testing. Por ejemplo, if the classifier
Derrfuss et al.
1551
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
oh
6
C
5
norte
9
_
6
a
/
_
j
0
oh
1
C
1
norte
4
1
_
a
pag
_
d
0
1
b
1
y
4
gramo
1
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
t
/
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
categorized zero out of three faces correctly (0% acumular-
racy) and six out of six houses (100% exactitud), the bal-
anced accuracy will be 50%, reflecting the fact that the
classifier failed at correctly identifying faces and simply pre-
dicted “house” each time. For an unbalanced measure of
exactitud, sin embargo, the classification accuracy will be
66.6% (6 out of 9 correcto), suggesting successful decoding.
To speed up searchlight computations, the analysis
was run using a Monte Carlo approach similar to the
one described by Björnsdotter, Rylander, and Wessberg
(2011): PyMVPA’s “scatter-rois” parameter was set to 1,
indicating that at least one voxel had to be located be-
tween two neighboring searchlight centers (thus reduc-
ing computation time). Each voxel was then assigned
the mean balanced accuracy of all searchlights to which
it contributed (cf. Björnsdotter et al., 2011). Single-
subject balanced accuracy maps were then nonlinearly
registered to MNI152 space using ANTs (Avants, Tustison,
Wu, Cocinar, & Gee, 2011), and one-sample t tests were
performed to identify voxels with above-chance decoding
actuación. Correction for multiple comparisons
was done using FSL’s cluster tool (fsl.fmrib.ox.ac.uk/fsl/
fslwiki/Cluster) with a voxel-wise z-threshold of 3.1 en
combination with a cluster level threshold of p < .05
(Worsley, 2001). For visualization, thresholded z-maps
were then overlaid on the PALS-B12 atlas ( Van Essen,
2005) using Caret ( Van Essen et al., 2001).
We report two main types of decoding analyses. In
within-delay analyses, the classifier was trained and tested
on independent trials of the same type of delay phase
(i.e., pre-distractor, distractor, or post-distractor) to as-
sess whether brain activity in the respective phase carries
information about STM contents (faces vs. houses).
Across-delay analyses, on the other hand, tested if a clas-
sifier trained on one delay phase (e.g., the pre-distractor
delay) generalizes to another (e.g., the post-distractor
delay). To ensure that generalization works in both direc-
tions, we always ran both possible analyses (e.g., training
on the pre-distractor delay and testing on post-distractor
delay, and vice versa) and averaged the results.
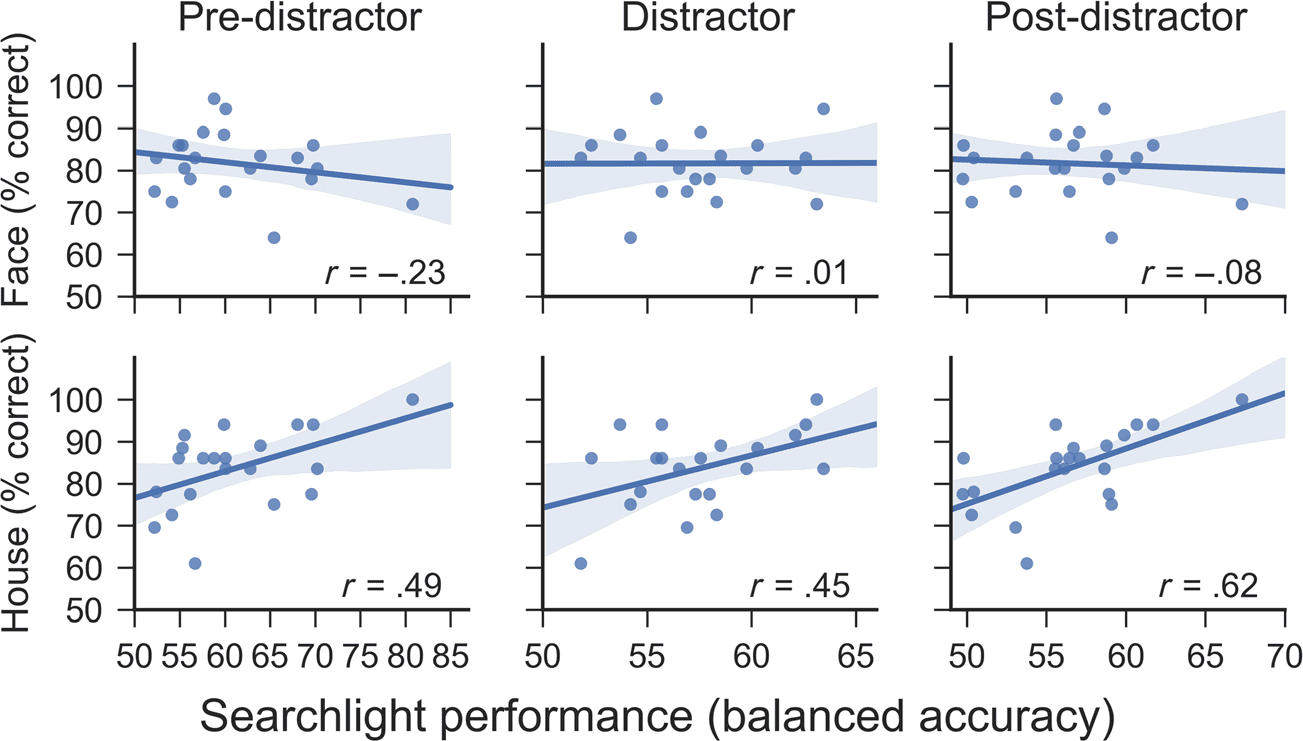
For the analysis that correlated behavioral and decod-
ing performance, a slightly different approach for the se-
lection of data samples was chosen. Although the number
of training data samples was still equal for face and house
trials, we randomly selected a predefined number of data
samples from the available sets. This predefined number
was the number of data samples available in five chunks
(as five chunks were used for training) for the lowest-
performing participant (16 correct house trials). This pro-
cedure ensured that the number of data samples available
for training was independent of the performance of the
participant. Without this procedure, a positive correlation
between behavioral and decoding performance might re-
flect nothing but a confound between performance and
the number of trials available in the training set. In addi-
tion, to mitigate the effects of randomly selecting a rela-
tively small set of samples, we increased the number of
repetitions per fold from 5 to 15 (again, further increasing
the number of repetitions had very little effect on the
results). Apart from these changes, this analysis was iden-
tical to the previously described approach. Correction for
multiple comparisons of correlation coefficients used the
permutation approach described by Yoder, Blackford,
Waller, and Kim (2004). Behavioral performance scores
were randomly permuted, whereas classifier accuracies
for the different delays were not (leaving the interrela-
tions between these variables intact). The number of
permutations was set to 100,000.
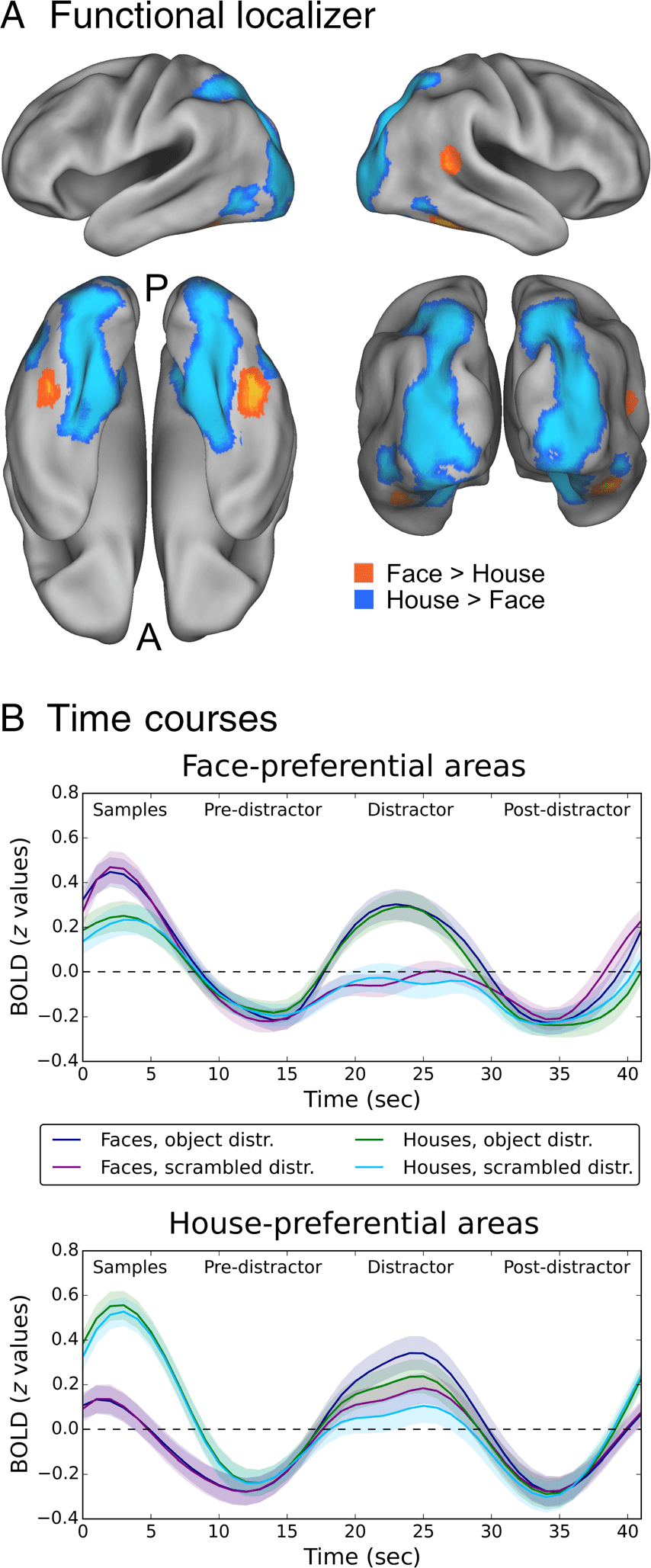
Analysis of the Functional Localizer Task and
Time Course Analysis
Preprocessing for the functional localizer task was identi-
cal to that of the main task, with the exceptions that the
temporal filter was set to 100 sec and that the blocked
design was modeled with epoch durations of 15 sec. A
univariate GLM (with motion correction parameters as
confound regressors) was estimated using FILM with lo-
cal autocorrelation correction ( Woolrich, Ripley, Brady,
& Smith, 2001); z-statistic images were thresholded as
described above. Face blocks were contrasted with house
blocks to identify face- and house-preferential processing
areas. To exclude significant clusters based on relative de-
activations in the control condition, the resulting z-maps
were additionally masked with the simple contrast of face
or house, respectively, versus the implicit baseline. It
turned out that the cluster size correction was too con-
servative for the left FFA. On the basis of the strong
a priori hypothesis about FFA location, we decided to re-
run the face versus house whole-brain analysis with z >
3.1, but without cluster size correction to identify the left
FFA. Using this approach, the left FFA was clearly identi-
fiable (there were no other above-threshold clusters
within several centimeters) at the mid-fusiform sulcus
( Weiner et al., 2014). We isolated the left FFA cluster
using fslmaths and merged the resulting map with the
face versus house map generated with cluster size cor-
rection. This combined map was used for visualization
purposes and for computing the multivariate analysis
focusing on FFA described below. To calculate time
courses, the time series from each run were shifted by
two repetition times, detrended, and z-scored. Próximo, todo
events belonging to the same condition were averaged
within and then across runs. Finalmente, across-subject means
and standard errors were calculated. For visualization,
time courses were up-sampled using cubic interpolation.
RESULTADOS
Behavioral Results
A 2 × 2 repeated-measures ANOVA with the factors Sam-
ple category (face or house) and Type of distraction (transmisión exterior-
ject or scrambled) indicated that mean RTs to probes
1552
Revista de neurociencia cognitiva
Volumen 29, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
oh
6
C
5
norte
9
_
6
a
/
_
j
0
oh
1
C
1
norte
4
1
_
a
pag
_
d
0
1
b
1
y
4
gramo
1
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
t
.
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Mesa 1. Probe RT (mseg) and Accuracy (%) as a Function of Distractor Type (Object or Scrambled) and Sample Category
(Face or House)
Probe RT (M and SD)
Probe Accuracy (M and SD)
Distractor Type
Face Sample
House Sample
Face Sample
House Sample
Object
Scrambled
1170 (216)
1117 (193)
1196 (217)
1181 (213)
80.7 (7.1)
82.7 (11)
85.1 (9.8)
82.2 (12.2)
(Mesa 1) displayed a significant main effect of Distraction,
F(1, 20) = 5.3, pag = .03, ηp
2 = .21, with RTs being slower
after object relative to scrambled distraction. Había
also a trend toward a main effect of Stimulus category,
F(1, 20) = 3.8, pag = .07, ηp
2 = .16, with RTs tending to
be slower in house than face trials. The interaction was
not significant, F(1, 20) = 1.6, pag = .22. For mean probe
accuracies (Mesa 1), there were no significant main ef-
fects or interactions (all ps > .17). Finalmente, a Pearson cor-
relation showed that accuracies for face and house trials
were not significantly related, r(19) = .28, pag = .22.
Performance on the color task during distractor pre-
sentation (Mesa 2) was analyzed using 2 × 3 repeated-
measures ANOVAs with the factors Sample category (rostro
or house) and Type of distraction (rostro, house, or scram-
bled). RTs showed a significant main effect of Distraction
tipo, F(1, 20) = 32.9, pag < .001, ηp
2 = .78, because of
faster color judgments for scrambled relative to face
(t(20) = 7.1, p < .001) or house distractors (t(20) = 6,
p < .001). No other effects were significant (all ps > .4).
Analogously, higher mean accuracies were found for scram-
bled distractors (main effect distraction type, F(1, 20) =
13, pag < .001, ηp
2 = .58; scrambled vs. face: t(20) = 4.6,
p < .001; scrambled vs. house: t(20) = 4.1, p = .001). Again,
no other effects were significant (all ps > .8).
fMRI Results
Within-delay Analyses
pendiente). For the distractor delay, this initial analysis
collapsed across distractor type (object or scrambled)
to maximize power. Results showed that the SVM classi-
fier could indeed decode whether participants main-
tained faces or houses with above-chance accuracy in
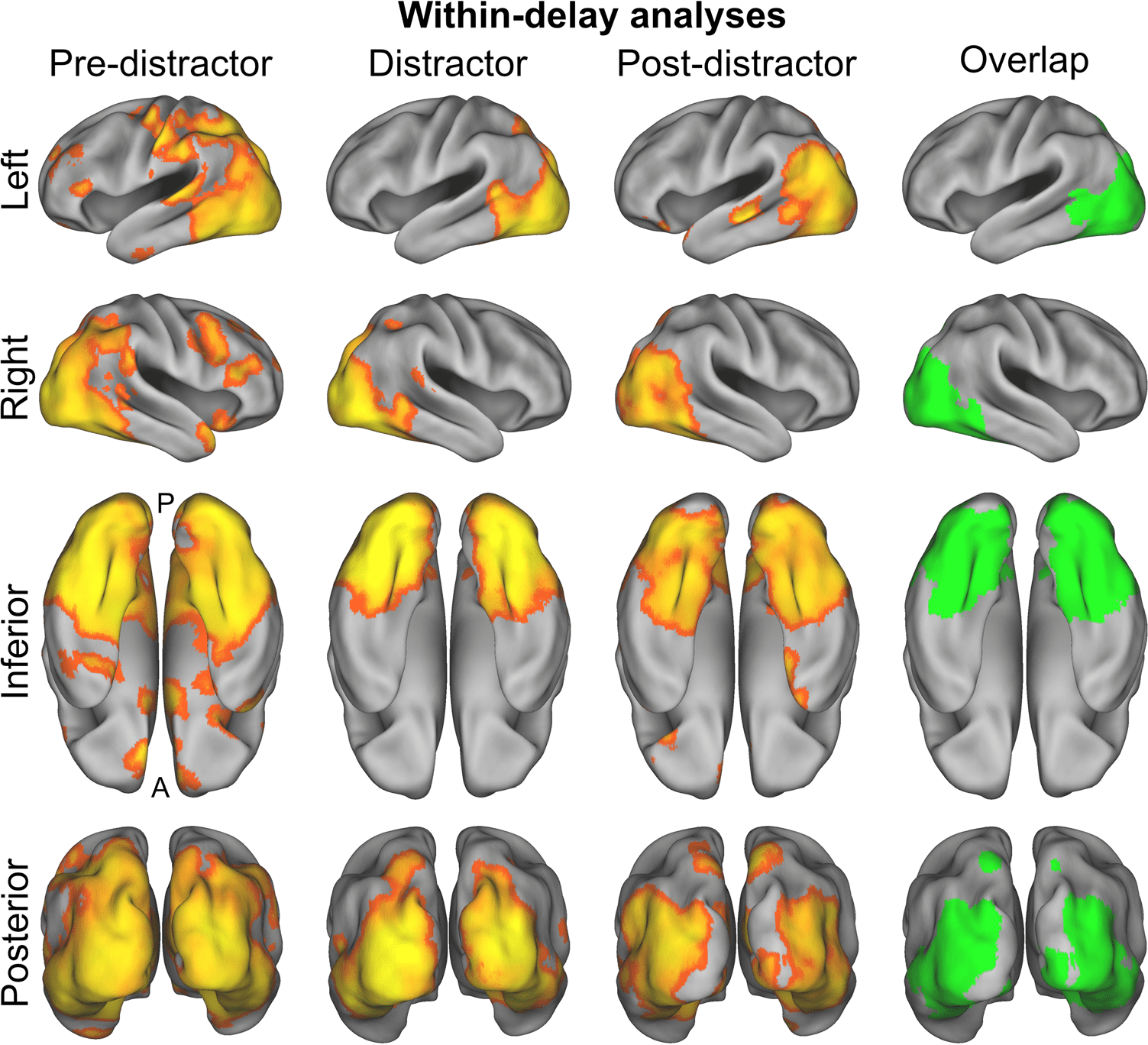
all three delays. For the pre-distractor delay, the search-
light analysis revealed that information about STM con-
tent was present in a broad range of brain regions,
including extensive parts of the occipital cortex, también
as parts of the parietal, temporal, and pFC (ver tabla 3
y figura 2). As this initial delay phase is not the main
focus of the present article, we will not describe these
activations in further detail. For the distractor delay, re-
gions with above-chance decoding strongly overlapped
with the results for pre-distractor delay but were restrict-
ed to posterior cortex (Mesa 3 y figura 2), incluido
parts of the occipital cortex; parts of the parahippocam-
pal, lingual, and fusiform gyri; posterior parts of the tem-
poral cortex; and the posterior parietal cortex around the
paroccipital segment of the intraparietal sulcus (Zlatkina
& Petrides, 2014). For the post-distractor delay, regiones
with above-chance decoding were very similar to those
of the distractor delay, with additional clusters found in
the mid-portion of the left STS, the left inferior frontal
gyrus (pars orbitalis), and the anterior frontomedian cor-
tex (Mesa 3 y figura 2). Regions common to the three
individual analyses are shown as an overlap map in the
rightmost column of Figure 2.
As a first step, we examined whether or not it would be
possible to decode, within the different delay phases,
which stimulus type (face vs. house) was maintained in
STM (please note that the cross-validation procedure en-
sured that training and testing data samples were inde-
Across-delay Analyses of Pre- and Post-distractor Delays
As a second step, we explored whether or not the MVPA
classifier generalizes from pre- to post-distractor delay
periods and vice versa. Successful decoding in these
Mesa 2. Distractor RT (mseg) and Accuracy (%) as a Function of Distractor Type (Face, House, or Scrambled) and Sample Category
(Face or House)
Distractor RT (M and SD)
Distractor Accuracy (M and SD)
Distractor Type
Face Sample
House Sample
Face Sample
House Sample
Face
House
Scrambled
503 (56)
498 (47)
471 (51)
508 (47)
497 (46)
474 (45)
93.5 (4.8)
93.8 (6.2)
97.5 (2.9)
93.7 (5.4)
94.3 (4.6)
97.4 (2.9)
Derrfuss et al.
1553
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
oh
6
C
5
norte
9
_
6
a
/
_
j
0
oh
1
C
1
norte
4
1
_
a
pag
_
d
0
1
b
1
y
4
gramo
1
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
.
/
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Mesa 3. Within-delay Analyses
Location
Pre-distractor Delay
Volumen (mm3)
z-Value
X,y,z
Inferior temporal gyrus, cerebellum
555,840
Parahippocampal gyrus, collateral sulcus, cerebellum
Collateral sulcus
Intraparietal sulcus
Postcentral gyrus
Transverse occipital sulcus
Cerebellum, collateral sulcus
Lateral occipital gyrus
Cerebellum
Fusiform gyrus
Superior frontal gyrus
Superior frontal sulcus
Inferior precentral sulcus
Inferior frontal gyrus
Superior frontal gyrus
Middle frontal gyrus
Paracingulate sulcus
Middle frontal gyrus
Distractor Delay
Fusiform gyrus
Lateral occipital gyrus
Fusiform gyrus
Superior temporal sulcus
Lateral occipital gyrus
Fusiform gyrus
Superior occipital gyrus
Inferior temporal gyrus
Middle temporal gyrus
Lingual gyrus, parahippocampal gyrus
Post-distractor Delay
Lateral occipital gyrus
Superior temporal sulcus
Fusiform gyrus
Lateral occipital gyrus
Fusiform gyrus
Lingual gyrus
21,376
8,144
230,008
231,592
6.44
5.79
5.47
5.33
5.27
5.25
5.94
5.54
5.35
5.26
4.6
4.46
4.39
4.21
3.7
4.44
3.82
3.77
6.8
6.55
6.44
4.26
6.86
6.72
4.9
4.78
4.68
4.66
5.74
5.04
4.94
4.89
4.77
5.03
−44,−46,−30
−20,−36,−22
−26,−50,0
−22,−56,44
−56,−14,46
−20,−82,16
16,−44,−18
28,−88,20
52,−58,−30
38,−28,−22
30,58,30
24,22,34
54,10,44
52,32,14
0,58,42
−36,40,38
−8,40,32
−38,56,18
−42,−58,−14
−32,−94,14
−26,−80,−10
−50,−46,10
34,−86,10
30,−66,−14
12,−94,24
56,−66,2
68,−44,4
16,−38,−14
−22,−86,14
−56,−58,18
−22,−74,−12
−44,−78,30
−28,−46,−14
12,−58,4
1554
Revista de neurociencia cognitiva
Volumen 29, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
oh
6
C
5
norte
9
_
6
a
/
_
j
0
oh
1
C
1
norte
4
1
_
a
pag
_
d
0
1
b
1
y
4
gramo
1
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
t
F
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Mesa 3. (continued )
Location
Lingual gyrus
Inferior temporal gyrus
Occipitotemporal cortex
Fusiform gyrus
Superior parietal lobule
Superior parietal lobule
Cingulate sulcus
Parahippocampal gyrus
Temporal pole
Rostral gyrus
Volumen (mm3)
z-Value
4.97
4.93
4.81
4.73
3.76
4.89
3.81
4.13
5.15
3.74
8,312
3,856
3,840
2,424
X,y,z
12,−82,−12
56,−60,−18
58,−64,18
30,−42,−20
−8,−70,58
18,−60,60
14,−36,42
26,−6,−32
−50,22,−18
−4,46,−14
The z maps were thresholded using clusters determined by z > 3.1 and a corrected cluster significance threshold of p < .05. Peaks are at least
25 mm apart. Up to 10 peaks per cluster are reported.
analyses would provide evidence that the neuronal code
representing sample category information was either ro-
bustly maintained across distraction or reinstated after
the distractor delay. We found that sample categories
could be successfully decoded when training on the
pre-distractor delay and testing on the post-distractor de-
lay, and vice versa. Regions with above-chance decoding
in both analyses were found in the parahippocampal and
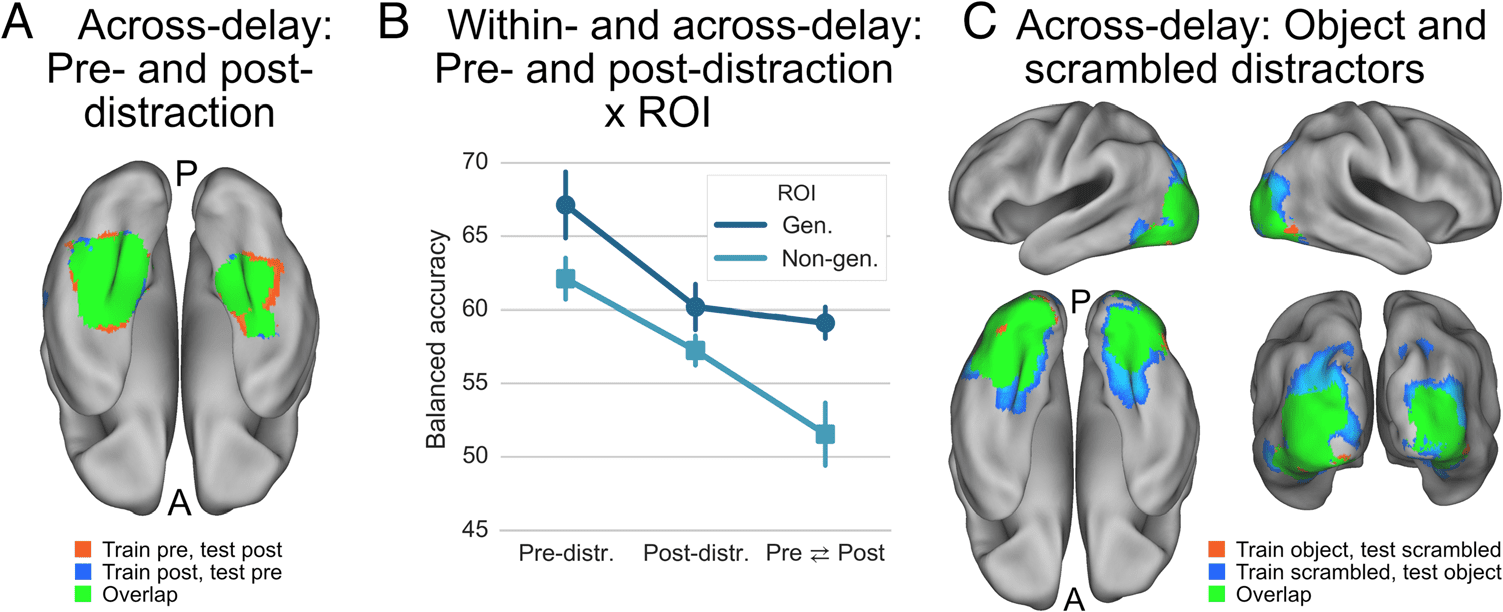
fusiform gyri (Table 4; Figure 3A) and overlapped with
functionally defined FFA and PPA (see below).
The areas identified in the present across-delay analy-
ses overlapped with the anterior inferotemporal regions
Figure 2. Results of the within-
delay searchlight analyses
decoding STM representations
of faces versus houses. For
these analyses, a multivariate
classifier was trained and tested
on independent chunks of the
same delay phase. Depicted in
color are areas where decoding
performance was above chance
(z > 3.1, cluster threshold p <
.05). The rightmost column
depicts the overlap in all three
delay phases.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Derrfuss et al.
1555
Table 4. Across-delay Analyses for the Pre- and Post-distractor Delays
Location
Volume (mm3)
z-Value
x,y,z
Train on Pre-, Test on Post-distractor Delay
Collateral sulcus
Fusiform gyrus
Collateral sulcus
Parahippocampal gyrus
Lateral occipitoparietal cortex
Train on Post-, Test on Pre-distractor Delay
Occipitotemporal sulcus
Fusiform gyrus
Inferior temporal sulcus
Collateral sulcus
Fusiform gyrus
18,400
9,568
2,096
19,808
6,560
4.91
4.15
4.31
3.9
3.68
4.46
4.29
3.55
3.72
3.57
−30,−38,−16
−30,−58,−14
26,−38,−16
36,−22,−24
−40,−84,26
−44,−44,−10
−28,−44,−16
−54,−30,−10
22,−42,−14
36,−24,−26
The z maps were thresholded using clusters determined by z > 3.1 and a corrected cluster significance threshold of p < .05. Peaks are at least
15 mm apart.
identified in all three within-delay analyses. This suggests
that only in ventral temporal areas the category-specific
pattern of activation could survive or be reinstated,
whereas the pattern of activation in more posterior areas
was modified after the presentation of the distractors. To
formally test this observation, we created two ROIs based
on the overlap map shown in Figure 2, that is, an ROI
consisting of those voxels showing significant across-
delay generalization and another ROI consisting of all re-
maining voxels in the overlap map (i.e., that did not
generalize from pre- to post-distractor delay). We per-
formed a 2 × 3 repeated-measures ANOVA on the mean
balanced accuracies (cf. Methods) retrieved from these
ROIs with factors ROI (generalization or nongeneraliza-
tion) and Analysis (pre-distractor, post-distractor, or
across-delay analysis).
The results showed a main effect of ROI, F(1, 20) =
2 = .7, and a main effect of Analysis,
46.8, p < .001, ηp
Figure 3. (A) Results of the
across-delay analysis of face
versus house representations
involving the pre- and post-
distractor delays. Depicted in
color are areas where decoding
performance was above chance
(z > 3.1, cluster threshold p <
.05), indicating that in these
areas the pattern of pre-
distractor maintenance-related
activity was reinstantiated after
distraction. A ventral view of the
PALS-B12 atlas brain is shown.
(B) Results of an ROI analysis
further investigating the
reinstatement of maintenance-
related activity patterns. The analyses labeled “Pre-distr.” and “Post-distr.” are the respective within-delay analyses. The analysis labeled “Pre ⇆ Post”
is the across-delay analysis involving training and testing on the pre-distractor delay and the post-distractor delay. The ROIs were defined based
on the overlap map shown in Figure 2. The generalization ROI (“Gen.”) corresponds to areas within the overlap map where evidence was
reinstated after the distractor delay, the nongeneralization ROI (“Non-gen.”) corresponds to areas within the overlap map where evidence was
not reinstated. The main result of this analysis is that the balanced classifier accuracy in the nongeneralization ROI drops significantly in the
Pre ⇆ Post analysis relative to the post-distractor analysis, whereas this is not the case for the generalization ROI. Error bars represent 95%
within-subject CIs. (C) Results of the across-delay analysis involving object and scrambled distractor delays. Depicted in color are areas where
decoding performance was above chance (z > 3.1, cluster threshold p < .05), indicating that these areas represented sample information in the
distractor delay in a similar way irrespective of the type of distractor.
1556
Journal of Cognitive Neuroscience
Volume 29, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
F(2, 40) = 30.2, p < .001, ηp
2 = .6 (Figure 3B). Crucially,
there was also a significant interaction effect, F(2, 40) =
9.4, p < .001, ηp
2 = .59. For the across-delay analyses, the
less accurate of the two within-delay analyses likely pro-
vides an upper limit for the generalization classifier per-
formance. Therefore, in our post hoc analyses of the
interaction effect, the across-delay results will be evalu-
ated relative to the less accurate within-delay analysis, that
is, the post-distractor delay. Note that these analyses are
independent of how the ROIs were selected. In the non-
generalization ROI, decoding performance was indeed
worse for the across-delay analysis, compared with the
post-distractor delay analysis, t(20) = 4.9, p < .001,
Cohen’s d = 1.17, whereas there was no significant differ-
ence in the generalization ROI, t(20) = 1.2, p = .23,
Cohen’s d = 0.15. As a t test cannot provide evidence
for the null hypothesis, we also calculated the scaled JZS
Bayes factor (Cauchy prior width = 0.707). The Bayes
factor was 2.3, indicating that, given the data, the null hy-
pothesis of no difference between the conditions resulted
in being twice as likely, which can be considered weak ev-
idence in favor of the null hypothesis. For the nongener-
alization ROI, a control analysis showed that the drop in
accuracy in the pre–post analysis (5.7%) was almost iden-
tical (5.6%) when voxels with lower accuracies in the post-
distractor analysis were omitted from the analysis (and, as
a result, the mean accuracy of the generalization and non-
generalization ROIs in the post-distractor analysis was
matched). This suggests that the observed pre–post anal-
ysis drop in accuracy in the nongeneralization ROI is not
driven by a subset of voxels with lower overall accuracies
or higher levels of noise. Taken together, these analyses
indicate that in bilateral ventral temporal cortex (fusiform
and parahippocampal gyri) the patterns of activity for the
two categories were stable across the pre-distractor delay
and the post-distractor delay (and thus a classifier trained
on distinguishing these categories in one type of delay
could also distinguish them in another), whereas in more
posterior areas (involving occipital, posterior temporal,
and parietal cortex) the patterns of activity changed over
time, suggesting that in these latter areas the neural rep-
resentations of memory contents were unique to the
respective delay.
Across-delay Analyses Involving the Distractor Delay
As a third step, we tested whether or not generalization
was still possible when the analyses involved the distrac-
tor delay. First, we trained on the pre-distractor delay and
attempted to classify in the distractor delay, and vice ver-
sa. The searchlight analysis did not show any areas with
above-chance decoding. Next, we trained on the distrac-
tor delay and attempted to classify in the post-distractor
delay, and vice versa. Again, sample category could not
be successfully decoded. An inspection of the mean bal-
anced accuracies for the above analyses in the overlap
areas shown in Figure 2 revealed that these accuracies
were slightly below chance (pre-distractor and distractor:
M = 47.3%, SD = 2.2; post-distractor and distractor: M =
49.2%, SD = 2.4). To further explore the null effects in
the decoding analyses, directional Bayes factors (alterna-
tive hypothesis: M > 50%; Cauchy prior width = 0.707)
for the mean balanced accuracies in the overlap ROI were
computed. The Bayes factors were 19.4 y 10, respetar-
activamente, constituting strong evidence in favor of the null
hypothesis. De este modo, although it was possible to decode
STM contents when training and testing were based on
the distractor delay, decoding did not generalize between
unfilled delay periods and the distractor delay period,
suggesting that some information about the sample cat-
egory is maintained during the distractor delay, but that
the pattern of activity representing this information is
fundamentally different from the other delays. This indi-
cates that activation patterns representing STM contents
in the ventral temporal cortex are not maintained across
all delays, but are reinstated after distraction.
A potential problem of the previous across-delay anal-
yses involving the distractor delay is that the distractor
delay is the only delay in which stimuli were presented.
De este modo, Por ejemplo, training on the pre-distractor delay
and testing on the distractor delay involves training in
the absence of perceptual input and testing in the pres-
ence of perceptual input. To address this issue, we re-
peated the analyses after training on the encoding
phase. Using this approach, the classifier generalized to
the pre- and post-distractor delay phases (in both direc-
ciones). The regions found in these analyses overlapped
with those identified in the above-reported within-delay
analiza (results not shown). Sin embargo, decoding for the
distractor phase was still not successful. Estos resultados
strengthen our conclusion that the pattern of activation
representing information about samples is qualitatively
different in the distractor delay.
Decoding Sample Category during Object and
Scrambled Distraction
To maximize power, our initial analysis collapsed across
distractor type (es decir., object and scrambled distractors). Como
a result, successful decoding during the distractor phase
might be dominated by one of the distractor types. A
more rigorous demonstration of maintenance during
the distractor delay would be to show that it is possible
to train the classifier to distinguish the sample categories
during one type of distraction and then decode them
during the other. As Figure 3C shows (see also Table 5),
this analysis was indeed successful. The regions common
to both analyses were highly similar to the distractor-type
independent analysis (Cifra 2, second column) y
involved the occipital cortex and the posterior fusiform
gyrus. This suggests that, in these regions, a common
pattern of activity is present during both types of distrac-
tor delay.
Derrfuss et al.
1557
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
oh
6
C
5
norte
9
_
6
a
/
_
j
0
oh
1
C
1
norte
4
1
_
a
pag
_
d
0
1
b
1
y
4
gramo
1
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
F
.
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Mesa 5. Across-delay Analyses for the Object and Scrambled Distractor Delays
Location
Train Object, Test Scrambled
Lateral occipital gyrus
Fusiform gyrus
Fusiform gyrus
Lateral occipital gyrus
Fusiform gyrus
Train Scrambled, Test Object
Fusiform gyrus
Fusiform gyrus
Lateral occipital gyrus
Paroccipital segment of the intraparietal sulcus
Fusiform gyrus
Lateral occipital gyrus
Parieto-occipital fissure
Paroccipital segment of the intraparietal sulcus
Volumen (mm3)
z-Value
X,y,z
35,360
21,840
59,554
45,592
4.95
4.82
4.6
4.64
4.27
5.95
5.71
5.64
4.11
5.99
5.54
4.42
3.82
−34,−92,12
−22,−86,−12
−34,−70,−12
34,−90,12
28,−68,−18
−26,−80,−8
−36,−60,−8
−26,−94,24
−22,−72,24
30,−62,−12
30,−84,10
22,−62,18
22,−60,42
The z maps were thresholded using clusters determined by z > 3.1 and a corrected cluster significance threshold of p < .05. Peaks are at least 20 mm
apart.
Relationships between Behavior and Decoding
Our decoding analyses showed that it was possible to dis-
tinguish face from house maintenance in all three delay
periods. However, these analyses cannot tell us if face
and house information was actively maintained through-
out all delay periods or if perhaps different strategies
were employed for the different stimulus categories.
For example, previous studies have shown that there is
a long-term memory contribution to short-term mainte-
nance of faces ( Warrington & Taylor, 1973; but see Race
et al., 2013). This could suggest that active maintenance
was more relevant for house trials than for face trials. To
investigate this possibility, we correlated behavioral and
decoding performance. To evaluate the decoding perfor-
mance, the mean balanced accuracy across those voxels
involved in all three delay periods (see Figure 2, right)
was retrieved. Note that all decoding analyses were based
exclusively on correct trials and thus cannot be influenced
by error trials. In addition, we ensured that the number of
trials contributing to a training set was orthogonal to the
behavioral performance of a participant (see Methods).
Permutation analyses corrected for multiple comparisons
showed that for face trials there was no relationship be-
tween behavioral and classifier performance (Figure 4,
top panel). For house trials (Figure 4, bottom panel), be-
havioral and classifier performance in pre-distractor delay
and post-distractor delay was significantly correlated
( p = .045 and p = .007, respectively; one-tailed p values).
For the distractor delay, the relationship was marginally sig-
nificant ( p = .062, one-tailed; but see semipartial correla-
tion reported below). This result suggests that participants
who were good at correctly identifying old versus new
house pictures also had superior representations of the
sample pictures during the delay periods.
As the across-delay analysis for the pre-distractor delay
and the post-distractor delay indicated that for a subset of
brain areas in the temporal lobe the pattern of activity
was recovered following distraction, we investigated if
the success in recovering the pattern of activity (as in-
dexed by the accuracy achieved by training on the pre-
distractor delay and testing on the post-distractor delay,
and vice versa) would also be related to the behavioral
performance in house trials. To this end, we correlated
the average balanced accuracy for the across-delay analy-
ses of the pre- and post-distractor delays in the general-
ization ROI with the performance in house trials. Results
showed that this correlation was significant, r(21) = .45,
p = .018 (one-tailed permutation test), suggesting that
participants whose patterns of activity were more similar
in the pre-distractor delay and the post-distractor delay
performed better on house trials.
Localizer and Time Course Analyses
Table 6 and Figure 5A present the results of the localizer
contrasts. Face-preferential regions were relatively small
1558
Journal of Cognitive Neuroscience
Volume 29, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 4. Analyses correlating
classifier performance in the
three different delay intervals
with behavioral performance
in face and house memory
trials. Notably, behavioral
accuracy in face trials is never
correlated with classification
accuracy, whereas behavioral
accuracy in house trials is
correlated with classification
accuracy in all delay phases.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
(as noted in the Methods section, the left FFA could only
be identified when no cluster size correction was used)
and restricted to the mid-fusiform sulcus ( Weiner et al.,
2014) and the right STS. House-preferential regions on
the other hand were extensive, covering parts of the
parahippocampal gyrus (corresponding to the PPA), the
medial fusiform gyrus, the inferior temporal gyrus, the
occipital gyri, and the superior parietal lobule.
As apparent when comparing Figures 5A and 2, both
face- and house-preferential areas overlapped with regions
Table 6. Results of the Functional Localizer Analysis
Location
Face vs. House
Mid-fusiform sulcus
Mid-fusiform sulcus
Superior temporal sulcus
House vs. Face
Collateral sulcus
Lateral occipital gyrus
Fusiform gyrus
Paroccipital segment of the intraparietal sulcus
Parieto-occipital fissure
Inferior temporal gyrus
Occipital pole
Collateral sulcus
Lateral occipital gyrus
Superior parietal lobule
Parieto-occipital fissure
Fusiform gyrus
Inferior temporal gyrus
Volume (mm3)
z-Value
x,y,z
240
1,400
544
53,232
53,280
632
4.3
4.61
3.95
6.69
6.42
6.39
5.74
5.02
4.99
4.71
7.42
6.61
6.49
6.31
5.45
4.3
−44,−46,−22
44,−48,−18
46,−46,18
−26,−46,−8
−38,−88,14
−26,−80,−14
−26,−76,40
−16,−62,12
−50,−62,−6
−18,−100,0
24,−46,−10
34,−84,20
20,−72,52
18,−58,16
26,−88,−10
52,−60,−12
The z maps were thresholded using clusters determined by z > 3.1. A corrected cluster significance threshold of p < .05 was applied to the house
versus face map. No cluster correction was applied to the face versus house map (see Methods). In addition, both maps were exclusively masked by
contrasts against the implicit baseline. Peaks are at least 25 mm apart.
Derrfuss et al.
1559
tential issue, we ran an additional analysis restricted to the
left and right FFA, considering all voxels in the ROI simul-
taneously. Results showed that sample category could be
decoded significantly above chance from all delay periods,
that is, pre-distractor delay: mean balanced accuracy =
66.7%, t(21) = 7.1, p < .001, 95% between-subject CI
[62.1, 71.4], Cohen’s d = 1.54; distractor delay: 57.1%,
t(21) = 4, p = .001, CI [53.6, 60.5], Cohen’s d = 0.87;
post-distractor delay: 59.3%, t(21) = 4.9, p < .001, CI
[55.6, 63.1], Cohen’s d = 1.06.
Finally, to evaluate the potential contribution of univar-
iate effects in stimulus-preferential ROIs, we also analyzed
univariate BOLD effects. The results (Figure 5B) showed
that, although there were clear univariate category effects
during the sample phase and early in the pre-distractor
delay, these effects were absent in the remainder of the
delay periods. In the distractor phase, face-preferential
areas showed a response only in object distractor trials
(where faces and houses were presented), whereas
house-preferential areas were also activated by scrambled
distractors. Interestingly, responses to distractors in
house-preferential areas were slightly reduced when
houses were maintained.
As the time-course analysis suggested that activity in
house-preferential areas tended to be generally sup-
pressed in the distractor delay of house maintenance tri-
als, we aimed to investigate if this suppression might (a)
help the classifier to distinguish the two sample catego-
ries and (b) drive the correlation observed between be-
havioral house accuracy and classifier performance in the
distractor delay (see Figure 4). First, we calculated the
degree of suppression in the distractor delay by calculat-
ing a mean beta image separately for face and house trials
for every participant across the whole brain. The mean
house beta image was then subtracted from the mean
face beta image to calculate the suppression effect for
every voxel (thus, higher scores correspond to more sup-
pression). The resulting whole-brain maps were masked
by the overlap map shown in Figure 2 (i.e., the resulting
map included the same voxels that were used to calculate
the correlations reported in Figure 4), and the suppres-
sion scores in the remaining voxels were averaged. The
result of these processing steps is, for every participant,
a single number reflecting the degree of suppression ob-
served in house trials relative to face trials during the dis-
tractor delay. Next, we calculated a Pearson correlation
between the suppression scores and the decoding accu-
racy in the distractor interval. The result showed that
these were indeed correlated, r(21) = .60, p = .004, sug-
gesting that the suppression effect may have helped the
classifier to distinguish the sample categories in the dis-
tractor delay. We then asked if this effect underlies the
correlation observed for behavioral and decoding accu-
racy shown in Figure 4. To investigate this, we computed
a semipartial correlation between behavioral and decod-
ing accuracy, controlling for the effect of suppression on
decoding accuracy. The result showed that behavioral
Figure 5. (A) Results of the 1-back task used as a functional localiser
(z > 3.1, cluster threshold p < .05). Face-preferential activations were
restricted to the FFA and the STS. House-preferential activations were
much more extensive, involving parts of the temporal, occipital, and
parietal cortex. (B) Mean univariate BOLD time courses (±SEM ) for
face- and house-preferential areas.
involved in all three delay periods. Given the small size of
the FFA, it is possible that information for distinguishing
between sample categories that is actually represented out-
side the FFA may have been assigned to voxels within the
FFA (as voxels were assigned the mean accuracy of all
searchlights to which they contributed; cf. Etzel, Zacks, &
Braver, 2013, for detailed discussion). To address this po-
1560
Journal of Cognitive Neuroscience
Volume 29, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
and decoding accuracy in the distractor delay were signif-
icantly correlated, rY(1.2)(21) = .62, p = .004. This sug-
gests that, whereas suppression in house trials has an
influence on the decoding accuracy, the suppression
effect does not drive the correlation of behavioral and
decoding accuracy.
DISCUSSION
This study investigated the neural correlates of distractor-
resistant STM employing a DMS task with face or house
stimuli. The task had three delay phases, a pre-distractor,
a distractor, and a post-distractor delay. Using fMRI and
multivariate searchlight analyses, we found that a support
vector machine classifier could successfully distinguish
face from house maintenance in all three delay phases
when trained and tested on the same delay phase (within-
delay analyses). Thus, our results provide support for
active maintenance in all three delay intervals. Using
across-delay analyses (where the classifier was trained
on one delay and tested on another), we found that the
classifier generalized from the pre-distractor delay to the
post-distractor delay (and vice versa) in ventral temporal
lobe areas overlapping with functionally defined face- and
house-preferential areas. However, generalization failed
when the training or the testing data set included the dis-
tractor delay, suggesting that patterns of maintenance-
related activity were different when distractors were
present. Our task employed two different types of distrac-
tors, scrambled and object pictures. Results showed that
it was possible to train on one type of distractor and de-
code the memory content while the other distractor type
was presented. This result suggests that the type of dis-
tractor had no major effect on how memoranda were
maintained in the distractor delay.
In a correlational analysis, we were furthermore able to
demonstrate that behavioral performance in house mem-
ory trials was positively correlated with decoding accu-
racy in posterior cortical areas. This suggests that, for
house trials, the patterns of activity in the different delay
intervals reflected how well sample-related information
was maintained. Presumably, these patterns allowed the
classifier to distinguish between the sample categories,
leading to improved classifier performance, and allowed
the participant to more precisely match sample and
probe, leading to improved behavioral performance. An-
other correlational analysis showed that the success of
classifier generalization in ventral temporal areas where
activity patterns were recovered after distraction was also
correlated with performance in house trials. This sug-
gests that for these temporal areas the fidelity of recovery
was relevant for task performance. It is noteworthy that
no analogous brain–behavior correlation was observed
for performance on face maintenance trials. We discuss
possible reasons for this differential finding below (see
Limitations and Open Questions section).
The remainder of our discussion will initially focus on
how our study relates to previous STM research (focusing
on distractor resistance) and what it contributes to this
research. We will then move on to a discussion of possi-
ble reasons as to why some previous studies failed to find
evidence for active maintenance in distractor delays.
Finally, we will discuss limitations of our approach and
open questions.
Relation to Previous Studies
Miller et al.’s (1996) classic study showed that lateral pre-
frontal neurons in macaques are involved in the recovery
of STM contents after distraction. However, this study did
not address the question if STM contents are actively
maintained during distraction. Recently, Jacob and
Nieder (2014) showed that parietal area VIP neurons do
maintain such distractor-resistant representations in a nu-
merosity task, thus highlighting the role of posterior cor-
tices in distractor-resistant memory. Our fMRI results
obtained in humans concur with these results in that they
also implicate posterior cortices. However, using a task
that required maintenance of information about visual
appearance (as opposed to numerosity), we found that
more posterior and ventral areas were involved in STM
maintenance during distraction.
This study differs from previous fMRI studies investi-
gating distractor-resistant STM (Bettencourt & Xu, 2016;
Jha et al., 2004; Sakai, Rowe, & Passingham, 2002b) by
having three extended delay phases, which allowed us
to use MVPA not only to study delay-specific STM main-
tenance but also to study the generalization of activity
patterns across delays. The results showed that the pres-
ence of distractors altered the neural code for sample
maintenance in a way that classifiers that could decode
STM contents during maintenance-only (and, in a control
analysis, during stimulus encoding) could not decode the
contents of STM in the presence of distractors. This sug-
gests that the pattern of maintenance-related activity is
modified when STM contents have to be shielded against
distraction. Generally, STM is thought to operate by
maintaining activity that is similar to the activity originally
elicited by the sample (D’Esposito & Postle, 2015). Thus,
the presentation of distractors might elicit activity similar
to the maintained samples, and this might lead to inter-
ference between the neural representations of the infor-
mation in memory and the distractor. One strategy to
deal with this may be to alter the maintenance-related
neuronal code in a way that makes it less similar to activ-
ity elicited by distractors.
Another important difference between our study and
previous studies investigating distractor-resistant mainte-
nance is that we required participants to actively process
the distractor pictures, whereas the studies by Bettencourt
and Xu (2016) and by Jha et al. (2004) used passive dis-
tractor conditions where participants were instructed to
simply watch the distractors. We chose an active distractor
Derrfuss et al.
1561
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
condition (i) to ensure that participants attended the dis-
tractors and (ii) to investigate if we could replicate the
results of Lewis-Peacock et al. (2012, Experiment 1),
who, using an active distractor task, had found that task-
irrelevant distractors made it impossible to decode the cur-
rently maintained category. In contrast, our results show
that even when the distractor task is attention-demanding,
there are conditions under which it is possible to decode
STM contents. Further below, we will address possible
reasons as to why our results differ from those of Lewis-
Peacock and colleagues.
We will now turn to a discussion of the areas from
which above-chance decoding during the distractor delay
was possible. Bettencourt and Xu (2016) report data from
individually defined intraparietal sulcus ROIs and ROIs re-
presenting joint visual areas V1 to V4. The intraparietal
sulcus ROIs were defined based on multiple regression
analyses for working memory capacity and based on their
vicinity to coordinates previously reported by Todd and
Marois (2004) and Xu and Chun (2006). Bettencourt and
Xu do not report individual peak coordinates for these
ROIs, but an inspection of the coordinates in Todd and
Marois (2004) and Xu and Chun (2006) suggests that the
ROIs were presumably located in the paroccipital seg-
ment of the intraparietal sulcus (Zlatkina & Petrides,
2014). In their Experiment 1, Bettencourt and Xu report
a dissociation between the two ROIs, such that sample
orientation could only be decoded from the parietal
but not from the visual ROI. In their Experiment 3, how-
ever, sample orientation could be decoded from both re-
gions. Bettencourt and Xu attribute this difference to a
change in participant strategy (as distractors were pre-
dictable in Experiment 1 but not in Experiment 3) and
suggest that the parietal ROI might be more relevant
for distractor-resistant maintenance as it was implicated
in both experiments.
In our study, above-chance decoding during distrac-
tion was possible from the paroccipital segment of the
intraparietal sulcus and from the occipital cortex. How-
ever, in our study, STM contents could also be decoded
from the temporal cortex, including the FFA and the PPA.
Likely, this extension along the ventral and medial-
temporal cortex is a consequence of the stimulus mate-
rial used, as ventral and medial-temporal cortices are
known to be involved in processing object information
(Reddy & Kanwisher, 2006). Our results show that these
areas are also involved in distractor-resistant STM
maintenance.
Contrary to Bettencourt and Xu’s (2016) results, we
found that it was possible to decode during the distractor
delay from visual cortex (similar to their Experiment 3)
even though the distraction was predictable (similar to
their Experiment 1). This shows that distractor predict-
ability does not necessarily lead to a null decoding result
for the visual cortex. As Ester, Rademaker, and Sprague
(2016) point out, Bettencourt and Xu’s null result is dif-
ficult to interpret and might indicate that these areas did
not contribute to the task, but it might also indicate that
the classifier failed to learn relevant patterns that may
nevertheless be present in the data. Furthermore,
Bettencourt and Xu’s (2016) Experiment 4 showed that
there is a positive correlation between the ability of a
multivariate fMRI classifier and of a participant to distin-
guish two gratings. This relationship was observed both
in V1 to V4 and in the intraparietal sulcus ROI. Bettencourt
and Xu explain the effect in V1 to V4 as “a result of its
role in the initial processing of the orientation informa-
tion” (p. 7). Our results show that the role of visual cor-
tices in distractor-resistant STM maintenance likely goes
beyond initial stimulus processing. Our distractor interval
onset was 8.8 sec after stimulus encoding. Still, we ob-
served a significant correlation between behavioral perfor-
mance in house trials and decoding accuracy in the
distractor delay. Thus, our results strengthen the hypoth-
esis that not only the intraparietal sulcus but also “lower-
level” visual cortices contribute to distractor-resistant STM
representations.
Turning to the post-distractor delay, our human fMRI
results differ from those reported for the macaque. Both
Miller et al. (1996) and Jacob and Nieder (2014) reported
recovery of STM contents in lateral prefrontal neurons af-
ter distraction. In contrast, we were not able to decode
from lateral prefrontal cortices in the post-distractor de-
lay. Again, such a null result is difficult to interpret, and a
failure to successfully decode could simply be related to
the methods employed (fMRI and/or MVPA). However,
there is also evidence to suggest that the human pFC
plays a less important role for simple STM maintenance
(D’Esposito, Cooney, Gazzaley, Gibbs, & Postle, 2006). In
addition, delays in our paradigm were much longer than
the delays used in the monkey studies. If the pFC would
be particularly relevant at the onset of the post-distractor
delay, our modeling approach would have been unable
to identify this involvement. Future studies specifically
designed to elucidate the role of the lateral pFC in
post-distractor recovery of STM contents might be able
to shed more light on this issue.
When Is Distractor-resistant Active
Maintenance Found?
We will now turn to a discussion of possible reasons as to
why some previous studies failed to find evidence for ac-
tive maintenance in distractor delays. As explained in the
introduction, we propose that there are two different,
but not necessarily mutually exclusive, strategies to main-
tain stimulus information across a distractor delay. We
referred to these strategies as active (associated with
persistent neuronal activity) and passive (not requiring
persistent firing) STM, and the degree to which one or
the other strategy is used may depend on the require-
ments of the task. The fact that we could decode the
stimulus category of STM contents during distraction sug-
gests that our task has encouraged active maintenance,
1562
Journal of Cognitive Neuroscience
Volume 29, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
which we attribute to the following reasons: (1) The sam-
ples on each trial were unique and had never been seen
by the participants before—thus precluding familiarity-
based task performance. (2) Distractors needed to be at-
tended and were presented in the same location as the
samples. Therefore, lower-level stimulus representations
were presumably “overwritten,” which might have im-
peded passive STM based on lower-level perceptual rep-
resentations. (3) About 25% of the probe picture were
covered by a black bar at an unpredictable location and
nonmatch probe pictures were selected from the set of
pictures previously presented as samples. Again, these
factors presumably made the use of a passive strategy
less likely to be successful. (4) Memory load was relatively
low (2 items), being well within the typical working
memory capacity (4 ± 1 items; Cowan, 2001).
Some of these factors might explain why, as men-
tioned above, a recent study by Lewis-Peacock et al.
(2012) was not successful at decoding memory content.
In their Experiment 1, distractors always came from a cat-
egory that was not currently maintained. Moreover, non-
match probes were not different exemplars of the same
category but came from an irrelevant category. Further-
more, there were only 18 stimuli overall which had been
learned before the fMRI scan, potentially allowing the for-
mation of long-term memory representations. All of these
factors may have reduced the likelihood that participants
relied on active maintenance. In a second experiment by
the same authors (Experiment 2), two samples were spa-
tially separated, and apart from cues, no further stimuli
were presented in the location of the samples. This might
have left lower-level perceptual representations mostly
intact and thus have facilitated reliance on passive mem-
ory mechanisms.
None of the factors discussed as possible reasons for
the inability to decode STM contents in Lewis-Peacock
et al. (2012) apply to the study by Sakai et al. (2002b),
raising the question why these authors observed no acti-
vation differences between correct and incorrect trials in
the distractor delay. One possible explanation is that the
authors applied a purely univariate approach to data anal-
ysis, which might be less sensitive than current multi-
variate approaches. However, there is also a potentially
more interesting explanation based on the nature of their
tasks: In Sakai et al.’s study, the combined memory load
from main task and distractor task was 10 items, which is
considerably above the typical working memory capacity
(Cowan, 2001) and makes it very unlikely that an active
memory strategy could have succeeded.
naire, 59% of participants reported that one of the strat-
egies they employed was to try to associate sample faces
with persons known to them (e.g., friends or celebrities);
an analogous strategy was only reported by 14% of partic-
ipants in the house condition. As a result, although all of
the samples were new to participants, long-term memory
representations might have played a more important role
for face trials. This might explain why decoding accuracy
was correlated with behavioral accuracy for house trials,
but not for face trials.
Our results suggest that posterior cortices play a role in
the active maintenance of information in long distractor
delays. Currently, few functional imaging studies have in-
vestigated this issue and a number of open questions re-
main. For example, it is unclear for how long active
maintenance during distraction is possible. In our task,
the distractor delay was 6.6 sec (using an active distractor
task), and in the study by Bettencourt and Xu (2016), it
was 10.2 sec (using passive distraction). Furthermore, it
is uncertain which factors determine if active mainte-
nance is used or not. The timing of the task, the memory
load, the material to be remembered, the preexistence of
long-term memory representations, and the participant
strategy might influence the results. Future research
should also continue to investigate which areas contrib-
ute to distractor-resistant STM. Although Bettencourt and
Xu (2016) stressed the role of the posterior intraparietal
sulcus, we could successfully decode in occipital, temporal,
and parietal cortices.
Conclusion
This study investigated the maintenance of visual infor-
mation in STM in unfilled and distractor-filled delays. A
multivariate searchlight analysis successfully decoded
STM contents (faces or houses) in all delay phases. Re-
gions with above-chance decoding in all delay phases
were located in the occipital, temporal, and posterior pa-
rietal lobes. In ventral temporal cortex, including func-
tionally defined areas FFA and PPA, activity patterns
were reactivated after distraction. In more posterior re-
gions, activity patterns were more flexible and depended
on the delay phase. Classifier performance in all delay
phases was correlated with the behavioral performance
in house trials, but not face trials. The present results
highlight the role posterior cortices play in the online
maintenance of STM contents both in the presence and
absence of distractors.
Acknowledgments
Limitations and Open Questions
Some of our results suggest that faces and houses were
treated differently by our participants. Performance in
face and house trials was only weakly correlated, and de-
coding accuracy was only related to behavioral house, but
not face trial performance. In a postexperiment question-
This research was supported by a VIDI grant from the Netherlands
Organization for Scientific Research (Grant 452-09-006 to C. J. F.).
J. D. was further supported by the German Research Foundation
(Grant DE 1141/2–1). M. H. was supported by funds from the
German Federal State of Saxony-Anhalt and the European
Regional Development Fund (ERDF; project: Center for Behavioral
Brain Sciences). M. T. was supported by the German Research
Derrfuss et al.
1563
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Foundation in the Transregional Collaborative Research Center
134. C. J. F. was further supported by an ERC consolidator grant
(No. 617891).
Reprint requests should be sent to Jan Derrfuss, School of
Psychology, University of Nottingham, Nottingham, NG7 2RD,
UK, or via e-mail: jan.derrfuss@nottingham.ac.uk.
REFERENCES
Avants, B. B., Tustison, N. J., Wu, J., Cook, P. A., & Gee, J. C.
(2011). An open source multivariate framework for
n-tissue segmentation with evaluation on public data.
Neuroinformatics, 9, 381–400.
Bettencourt, K. C., & Xu, Y. (2016). Decoding the content of
visual short-term memory under distraction in occipital and
parietal areas. Nature Neuroscience, 19, 150–157.
Björnsdotter, M., Rylander, K., & Wessberg, J. (2011). A Monte
Carlo method for locally multivariate brain mapping.
Neuroimage, 56, 508–516.
Bloemendaal, M., van Schouwenburg, M. R., Miyakawa, A.,
Aarts, E., D’Esposito, M., & Cools, R. (2015). Dopaminergic
modulation of distracter-resistance and prefrontal delay
period signal. Psychopharmacology, 232, 1061–1070.
Clapp, W. C., Rubens, M. T., & Gazzaley, A. (2010). Mechanisms
of working memory disruption by external interference.
Cerebral Cortex, 20, 859–872.
Cousineau, D. (2005). Confidence intervals in within-subject
designs: A simpler solution to Loftus and Masson’s method.
Tutorials in Quantitative Methods for Psychology, 1, 42–45.
Cowan, N. (2001). The magical number 4 in short-term
memory: A reconsideration of mental storage capacity.
Behavioral and Brain Sciences, 24, 87–114.
D’Esposito, M., Cooney, J. W., Gazzaley, A., Gibbs, S. E. B., &
Postle, B. R. (2006). Is the prefrontal cortex necessary for
delay task performance? Evidence from lesion and fMRI data.
Journal of the International Neuropsychological Society,
12, 248–260.
D’Esposito, M., & Postle, B. R. (2015). The cognitive
neuroscience of working memory. Annual Review of
Psychology, 66, 115–142.
Dunlap, W. P., Cortina, J. M., Vaslow, J. B., & Burke, M. J. (1996).
Meta-analysis of experiments with matched groups or repeated
measures designs. Psychological Methods, 1, 170–177.
Endl, W., Walla, P., Lindinger, G., Lalouschek, W., Barth, F. G.,
Deecke, L., et al. (1998). Early cortical activation indicates
preparation for retrieval of memory for faces: An event-
related potential study. Neuroscience Letters, 240, 58–60.
Ester, E. F., Rademaker, R. L., & Sprague, T. C. (2016). How do
visual and parietal cortex contribute to visual short-term
memory? eNeuro, 3, e1.
Etzel, J. A., Zacks, J. M., & Braver, T. S. (2013). Searchlight analysis:
Promise, pitfalls, and potential. Neuroimage, 78, 261–269.
Gazzaley, A., Cooney, J. W., Rissman, J., & D’Esposito, M.
(2005). Top–down suppression deficit underlies working
memory impairment in normal aging. Nature Neuroscience,
8, 1298–1300.
Hanke, M., Halchenko, Y. O., Sederberg, P. B., Hanson, S. J.,
Haxby, J. V., & Pollmann, S. (2009). PyMVPA: A python
toolbox for multivariate pattern analysis of fMRI data.
Neuroinformatics, 7, 37–53.
Jacob, S. N., & Nieder, A. (2014). Complementary roles for
primate frontal and parietal cortex in guarding working
memory from distractor stimuli. Neuron, 83, 226–237.
Jenkinson, M., Bannister, P., Brady, M., & Smith, S. (2002).
Improved optimization for the robust and accurate linear
registration and motion correction of brain images.
Neuroimage, 17, 825–841.
Jha, A. P., Fabian, S. A., & Aguirre, G. K. (2004). The role of
prefrontal cortex in resolving distractor interference. Cognitive,
Affective & Behavioral Neuroscience, 4, 517–527.
Kriegeskorte, N., Goebel, R., & Bandettini, P. (2006).
Information-based functional brain mapping. Proceedings of
the National Academy of Sciences, U.S.A., 103, 3863–3868.
Lewis-Peacock, J. A., Drysdale, A. T., Oberauer, K., & Postle,
B. R. (2012). Neural evidence for a distinction between
short-term memory and the focus of attention. Journal of
Cognitive Neuroscience, 24, 61–79.
Lewis-Peacock, J. A., & Postle, B. R. (2012). Decoding the
internal focus of attention. Neuropsychologia, 50, 470–478.
Miller, E. K., Erickson, C. A., & Desimone, R. (1996). Neural
mechanisms of visual working memory in prefrontal cortex of
the macaque. Journal of Neuroscience, 16, 5154–5167.
Morey, R. D. (2008). Confidence intervals from normalized data:
A correction to Cousineau (2005). Tutorials in Quantitative
Methods for Psychology, 4, 61–64.
Mumford, J. A., Turner, B. O., Ashby, F. G., & Poldrack, R. A.
(2012). Deconvolving BOLD activation in event-related
designs for multivoxel pattern classification analyses.
Neuroimage, 59, 2636–2643.
Oldfield, R. C. (1971). Assessment and analysis of handedness—
Edinburgh inventory. Neuropsychologia, 9, 97–113.
Pérez, F., & Granger, B. E. (2007). IPython: A system for
interactive scientific computing. Computing in Science &
Engineering, 9, 21–29.
Piekema, C., Kessels, R. P., Rijpkema, M., & Fernandez, G.
(2009). The hippocampus supports encoding of between-
domain associations within working memory. Learning and
Memory, 16, 231–234.
Poser, B. A., Versluis, M. J., Hoogduin, J. M., & Norris, D. G.
(2006). BOLD contrast sensitivity enhancement and
artifact reduction with multiecho EPI: Parallel-acquired
inhomogeneity-desensitized fMRI. Magnetic Resonance in
Medicine, 55, 1227–1235.
Race, E., LaRocque, K. F., Keane, M. M., & Verfaellie, M. (2013).
Medial temporal lobe contributions to short-term memory
for faces. Journal of Experimetal Psychology-General, 142,
1309–1322.
Reddy, L., & Kanwisher, N. (2006). Coding of visual objects in the
ventral stream. Current Opinion in Neurobiology, 16, 408–414.
Sakai, K., & Passingham, R. E. (2004). Prefrontal selection and
medial temporal lobe reactivation in retrieval of short-term
verbal information. Cerebral Cortex, 14, 914–921.
Sakai, K., Rowe, J. B., & Passingham, R. E. (2002a).
Parahippocampal reactivation signal at retrieval after
interruption of rehearsal. Journal of Neuroscience, 22,
6315–6320.
Sakai, K., Rowe, J. B., & Passingham, R. E. (2002b). Active
maintenance in prefrontal area 46 creates distractor-resistant
memory. Nature Neuroscience, 5, 479–484.
Smith, S. M. (2002). Fast robust automated brain extraction.
Human Brain Mapping, 17, 143–155.
Smith, S. M., Jenkinson, M., Woolrich, M. W., Beckmann, C. F.,
Behrens, T. E., Johansen-Berg, H., et al. (2004). Advances
in functional and structural MR image analysis and
implementation as FSL. Neuroimage, 23(Suppl. 1),
S208–S219.
Sprague, T. C., Ester, E. F., & Serences, J. T. (2016). Restoring
latent visual working memory representations in human
cortex. Neuron, 91, 694–707.
Todd, J. J., & Marois, R. (2004). Capacity limit of visual short-
term memory in human posterior parietal cortex. Nature,
428, 751–754.
Van Essen, D. C. (2005). A Population-Average, Landmark- and
Surface-based (PALS) atlas of human cerebral cortex.
Neuroimage, 28, 635–662.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1564
Journal of Cognitive Neuroscience
Volume 29, Number 9
Van Essen, D. C., Drury, H. A., Dickson, J., Harwell, J., Hanlon, D.,
& Anderson, C. H. (2001). An integrated software suite for
surface-based analyses of cerebral cortex. Journal of the
American Medical Informatics Association, 8, 443–459.
Warrington, E. K., & Taylor, A. M. (1973). Immediate memory
for faces: Long- or short-term memory? Quarterly Journal of
Experimental Psychology, 25, 316–322.
Weiner, K. S., Golarai, G., Caspers, J., Chuapoco, M. R.,
Mohlberg, H., Zilles, K., et al. (2014). The mid-fusiform
sulcus: A landmark identifying both cytoarchitectonic and
functional divisions of human ventral temporal cortex.
Neuroimage, 84, 453–465.
Woolrich, M. W., Ripley, B. D., Brady, M., & Smith, S. M. (2001).
Temporal autocorrelation in univariate linear modeling of
fMRI data. Neuroimage, 14, 1370–1386.
Xu, Y., & Chun, M. M. (2006). Dissociable neural mechanisms
supporting visual short-term memory for objects. Nature,
440, 91–95.
Yoder, P. J., Blackford, J. U., Waller, N. G., & Kim, G. (2004).
Enhancing power while controlling family-wise error: An
illustration of the issues using electrocortical studies.
Journal of Clinical and Experimental Neuropsychology,
26, 320–331.
Yoon, J. H., Curtis, C. E., & D’Esposito, M. (2006). Differential
effects of distraction during working memory on delay-period
activity in the prefrontal cortex and the visual association
cortex. Neuroimage, 29, 1117–1126.
Zanto, T. P., & Gazzaley, A. (2009). Neural suppression of
irrelevant information underlies optimal working memory
performance. Journal of Neuroscience, 29, 3059–3066.
Zarahn, E., Aguirre, G., & D’Esposito, M. (1997). A trial-based
experimental design for fMRI. Neuroimage, 6, 122–138.
Zlatkina, V., & Petrides, M. (2014). Morphological patterns of
the intraparietal sulcus and the anterior intermediate parietal
sulcus of Jensen in the human brain. Proceedings of the
Royal Society, Series B, Biological Sciences, 281, e1.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
9
2
9
1
/
5
9
4
/
7
1
1
5
9
4
5
7
2
/
9
1
4
7
6
8
o
6
c
5
n
9
_
6
a
/
_
j
0
o
1
c
1
n
4
1
_
a
p
_
d
0
1
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Derrfuss et al.
1565