Se comparte el circuito cerebral para el control cognitivo
by Task and Language Switching
Wouter De Baene1,2,3, Wouter Duyck1, Marcel Brass1, and Manuel Carreiras2,4
Abstracto
■ Controlling multiple languages during speech production is
believed to rely on functional mechanisms that are (al menos
partly) shared with domain-general cognitive control in early,
highly proficient bilinguals. Recent neuroimaging results have
indeed suggested a certain degree of neural overlap between
language control and nonverbal cognitive control in bilinguals.
Sin embargo, this evidence is only indirect. Direct evidence for
neural overlap between language control and nonverbal cogni-
tive control can only be provided if two prerequisites are met:
Language control and nonverbal cognitive control should be
compared within the same participants, and the task require-
ments of both conditions should be closely matched. To pro-
vide such direct evidence for the first time, we used fMRI to
examine the overlap in brain activation between switch-specific
activity in a linguistic switching task and a closely matched non-
linguistic switching task, within participants, in early, highly
proficient Spanish–Basque bilinguals. The current findings pro-
vide direct evidence that, in these bilinguals, highly similar
brain circuits are involved in language control and domain-
general cognitive control. ■
INTRODUCCIÓN
A key question in bilingual language production is how
bilingual speakers are able to control their two languages
during speech processing and why they are so efficient
in avoiding language conflicts or unintended nontarget
language intrusions. The nature of the cognitive pro-
cesses underlying this bilingual language control is still
a matter of debate and has generated a substantial body
of research during the last decade. Several language
control mechanisms have been proposed, such as inhibi-
tion of the unintended language (Verde, 1998).
One of the most frequently used paradigms to study
the cognitive mechanisms underlying bilingual and multi-
lingual language control in language production has been
the language-switching paradigm (Abutalebi et al., 2008,
2013; Gollan & Ferreira, 2009; Verhoef, Roelofs, & Chwilla,
2009; Costa, Santesteban, & Ivanova, 2006; Costa &
Santesteban, 2004; Jackson, Swainson, Cunnington, &
Jackson, 2001; Hernández, Martinez, & Kohnert, 2000;
Meuter & Allport, 1999). Recent neuroimaging research
has suggested that brain areas involved in language switch-
ing are similar to those implicated in nonverbal cognitive
control, as measured for instance with (nonverbal) tarea-
switching paradigms (p.ej., Garbin et al., 2010, 2011; guo,
Liu, Misra, & Kroll, 2011; Abutalebi & Verde, 2008; Wang,
Xue, Chen, Xue, & Dong, 2007; Crinion et al., 2006;
1Ghent University, 2Basque Center on Cognition, Brain and
Idioma, Donostia-San Sebastián, España, 3Tilburg University,
4Basque Foundation for Science, Bilbao, España
© 2015 Instituto de Tecnología de Massachusetts
Hernández, Dapretto, Mazziotta, & Bookheimer, 2001;
Hernandez et al., 2000). The language control network
involves lateral and medial prefrontal areas, parietal areas,
and the caudate nucleus (see Abutalebi & Verde, 2008, para
a review). Por otro lado, in task switching, a fronto-
parietal network is generally observed, including lateral
and medial prefrontal, premotor, and anterior and pos-
terior parietal regions as well as the BG (De Baene, Albers,
& Brass, 2012; De Baene & Brass, 2011; Shi, zhou, Müller, &
Schubert, 2010; Crone, Wendelken, Donohue, & Bunge,
2006; Yeung, nistrom, Aronson, & cohen, 2006; Barber
& Carretero, 2005; Ruge et al., 2005; Más valiente, Reynolds, &
Donaldson, 2003; Brass & por cramon, 2002; Dreher &
Berman, 2002; Dreher, Koechlin, Alí, & Grafman, 2002;
Rushworth, Hadland, Paus, & Sipila, 2002; Rushworth,
Paus, & Sipila, 2001; Dove, Pollmann, Schubert, Wiggins, &
Yves von Cramon, 2000; Kimberg, Aguirre, & D'Esposito,
2000; Sohn, Ursu, anderson, Stenger, & Carretero, 2000).
Curiosamente, up to now there is only indirect evidence
that the neural regions supporting language control are
the same as those supporting cognitive control in non-
verbal domains. Only a few studies have directly exam-
ined the neural regions involved in a nonverbal cognitive
control task in bilinguals (Garbin et al., 2010; Luk, anderson,
Craik, grady, & Bialystok, 2010; Bialystok et al., 2005).
Garbin et al. (2010), por ejemplo, compared the brain
regions involved in a non-linguistic switching task between
bilinguals and monolinguals and reported fundamental
differences in the brain network engaged in task switching
between both groups. Whereas monolinguals activated the
right inferior frontal gyrus, CAC, and left inferior parietal
Revista de neurociencia cognitiva 27:9, páginas. 1752–1765
doi:10.1162/jocn_a_00817
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
oh
3
C
5
norte
5
_
3
a
/
_
j
0
oh
0
C
8
norte
1
7
_
a
pag
_
d
0
0
b
8
y
1
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
/
t
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
lobule, bilinguals only displayed switch-specific activity in
the left inferior frontal gyrus and the left striatum. Given
that the left inferior frontal gyrus and the left striatum have
been consistently related to bilingual language control
(p.ej., Abutalebi & Verde, 2007), according to Garbin
et al. (2010), these results suggest a certain degree of
neural overlap between language control and nonverbal
cognitive control in bilinguals.
Sin embargo, to directly examine the link between the
regions involved in control of language conflict and
those involved in general cognitive control, we need to
examine the regions involved in both domains, dentro
the same participants. Until now, only Abutalebi et al.
(2012) followed this rationale. They examined within
the same participants whether language control and the
cognitive control processes involved in the flanker task
have a common neural substrate. The dorsal ACC was
found to be common to language switching and conflict
monitoring in the flanker task. Sin embargo, comparing
a language-switching task with a conflict task confounds
a number of cognitive control processes that are not
related to the specific requirements of language switch-
En g. En particular, language switching relies mainly on
the executive function of mental shifting (Miyake et al.,
2000), whereas the flanker task is more strongly asso-
ciated with inhibition of distractors or responses. A pesar de
these executive functions (together with updating of
working memory) are moderately correlated (see also
Friedman et al., 2006), they are clearly distinct, separable
funciones. Respectivamente, these functions seem to rely on a
partly shared–partly selective neural circuit. Hay
several brain areas involved commonly in different execu-
tive processes, whereas other brain areas are involved
only in specific executive processes (p.ej., only for shift-
En g; Hedden & Gabrieli, 2010). To capture those specific
processes that relate language control with nonverbal
control, one needs to compare language switching with
a closely matched nonlinguistic switching paradigm.
Por lo tanto, we examined the neural overlap between
two closely matched linguistic and nonlinguistic switch-
ing paradigms within the same participants.
En tono rimbombante, the occurrence and manifestation of lan-
guage conflict might depend on the proficiency of the
bilinguals (Van Heuven, Schriefers, Dijkstra, & Hagoort,
2008). De hecho, the precise nature of the language conflict
(Abutalebi & Verde, 2007) and the associated control
mechanism (Costa et al., 2006; Costa & Santesteban,
2004) might even alter qualitatively with proficiency. En
the neural level, the activation of the regions involved in
language control or the specific network involved might
also be modulated by language proficiency (Abutalebi
et al., 2013; Garbin et al., 2011). Como consecuencia, uno
might assume that also the overlap between the regions
involved in language control and the regions involved in
cognitive control might vary as a function of language
proficiency. en este estudio, we only considered early profi-
cient bilinguals who switch frequently between languages.
This choice was motivated by the fact that several stud-
ies have claimed that bilinguals outperform mono-
linguals on a range of cognitive control tasks (p.ej., Previo
& Gollan, 2011; Previo & MacWhinney, 2010; Bialystok &
Viswanathan, 2009; Bialystok, Craik, & Luk, 2008; Costa,
Hernández, & Sebastián-Gallés, 2008; but see Antón
et al., 2014; Duñabeitia et al., 2014; Hernández, Martín,
Barceló, & Costa, 2013; Paap & Greenberg, 2013). Cómo-
alguna vez, this bilingual advantage might be more salient and
might spread across a wider range of attention-demanding
tareas (Bialystok, Craik, & ryan, 2006) for those bilinguals
who constantly exercise language control functions on a
daily basis (Verreyt, Woumans, Vandelanotte, Szmalec, &
Duyck, 2015).
En resumen, in the current study, we wanted to examine
the overlap in brain activation between a language-
switching paradigm and a nonverbal task-switching
paradigm with a closely matched procedure, using a
within-subject paradigm with early proficient bilinguals.
This approach allows us to directly relate brain acti-
vation in a linguistic switching task to brain activation
in a nonlinguistic switching task, providing the strongest
test possible of the generalizability of the language con-
trol system developed by early proficient bilinguals to
the cognitive control domain.
en este estudio, we opted for a paradigm with three tasks
or three languages. This was motivated by the fact that
two-task or two-language experiments might be a special
caso, because switching away from one task or language
automatically involves switching back to the only other
task or language (Ruthruff, Remington, & Johnston,
2001). With three tasks or languages, a switch requires
that participants choose which of the remaining tasks
or languages to perform, which might be more repre-
sentative of natural language processing. Because early,
proficient bilinguals seem to apply the same language-
switching mechanism not only to the most proficient lan-
guages but also to weaker (L3) idiomas (Costa et al.,
2006; Costa & Santesteban, 2004), including a weaker
third language here should not imply qualitatively dif-
ferent language switches.
MÉTODOS
Participantes
Thirty-six healthy right-handed college students partici-
pated in this study for monetary reimbursement. Four
participants were excluded from the analyses because
of excessive movement during scanning. All remaining
Participantes (13 hombres; edad media = 22.4 años, range =
18–33 years) had Spanish as their L1, Basque as their L2
and had a good knowledge of English (L3). All participants
were early, highly proficient bilinguals: They acquired L2
at an early age (on average before the age of 3 años; uno
participant at the age of 8 años) and were regularly (en
promedio 4.3 days/week, range = 1–7 days/week)
De Baene et al.
1753
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
oh
3
C
5
norte
5
_
3
a
/
_
j
0
oh
0
C
8
norte
1
7
_
a
pag
_
d
0
0
b
8
y
1
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
F
.
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
confronted with contexts in which (enterrar- and intrasen-
tential) code switching between their L1 and L2 occurred.
The participants acquired L3 on average after the age of
6 años (range = 4–12 years) and were rarely confronted
with contexts in which code switching between their L1
and L3 or between their L2 and L3 occurred.
All participants had normal or corrected-to-normal
visión. None of them used medication or had a history
of drug abuse, head trauma, or neurological or psychiat-
ric illness. All participants gave informed consent before
pruebas. The study was approved by the institutional
ethical committee.
Materials
Language proficiency has many different dimensions
(word processing, syntactic processing, etc.), making it
a complex concept to measure. A pesar de, optimally,
proficiency should be defined using different tasks that
measure proficiency at different representational levels
of the language, only single word processing tasks were
included here given that the focus of this study is on
switching at the word level. Next to the self-reported
proficiency measures, language proficiencies in Spanish,
Basque, and English were therefore measured with the
Rapid Automatized Naming (RAN) test and the Boston
Naming Test (BNT; ver tabla 1 for results on these tests).
RAN Test
Both a digits RAN test and a color RAN test were admin-
istered in Spanish, Basque, and English in all participants.
The order of the language to be used was counter-
balanced across participants. The RAN test (Denckla &
Rudel, 1974) is assumed to measure the ability to access
and retrieve phonological representations from long-term
memory (p.ej., Torgesen, Wagner, Rashotte, Burgess, &
Hecht, 1997; Wagner & Torgesen, 1987) así como el
ability to form orthographic representations (Bowers,
Sunseth, & Golden, 1999; Bowers, Golden, Kennedy, &
Joven, 1994).
In each rapid naming test, participants were asked to
name, as quickly as possible, six visual stimuli displayed
en la pantalla, in a random order in four rows of nine
stimuli each. Before each test, all stimuli were shown
once to the participant to verify that he or she was able
to name them in the languages to be used.
The stimuli for the digits RAN test were 1, 2, 3, 5, 7,
y 8, each presented six times. The stimuli of the color
RAN test were red, negro, verde, brown, azul, and yellow
squares, also presented six times each. Naming times
were measured. The digits RAN test was always adminis-
tered before the color RAN test.
As a proficiency measure, the ratio between the
average naming times across the digits and color RAN
tests in L2 and L1 was calculated. Perfectly balanced par-
ticipants have a RAN ratio of 1, whereas larger RAN
ratios indicate a larger proficiency difference between
L1 and L2.
BNT
The BNT was administered in Spanish, Basque, e inglés
to all participants. The order of the language to be used
was counterbalanced across participants. The BNT is
assumed to measure word retrieval abilities (Kaplan,
Goodglass, & Weintraub, 1983).
The BNT contains 60 pictures presented one by one in
order of word frequency and grade of difficulty (de
common, high frequent, [p.ej., “bed”] to less familiar,
low frequent, [p.ej., “abacus”]). Participants were asked
to name them in the appropriate language. The scoring
was done according to standard instructions.
Language-switching Task
For the language-switching task, eight pictures (size =
3.27 × 3.27 visual degrees) of common objects with
noncognate names in Spanish, Basque, and English were
selected from the Snodgrass and Vanderwart pictures set
(rossión & Pourtois, 2004). The stimuli were selected
based on the following matching criteria across the three
idiomas: frequency, number of letters, number of
phonemes, number of orthographic neighbors, age of
adquisición, and concreteness. For Spanish and Basque,
information was extracted from the BaSp database
(Duñabeitia et al., in preparation). Para ingles, infor-
mation was provided by the N-Watch program (davis,
2005).
Mesa 1. Overview of Language Proficiency Scores
Self-ratings
Naming times (colores), segundo
Naming times (numbers), segundo
BNT
Español
9.56 (0.62)
21.59 (2.75)
14.04 (2.79)
50.72 (3.79)
Basque
8.13 (1.29)
25.65 (4.22)
17.30 (4.80)
39.97 (8.71)
Inglés
6.59 (1.16)
26.70 (6.61)
19.06 (2.95)
24.72 (7.41)
1754
Revista de neurociencia cognitiva
Volumen 27, Número 9
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
oh
3
C
5
norte
5
_
3
a
/
_
j
0
oh
0
C
8
norte
1
7
_
a
pag
_
d
0
0
b
8
y
1
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
t
.
/
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Cifra 1. Design of the
experimento. The language-
switching condition is
presented on the left. El
task-switching condition is
presented on the right. A trial
started with the presentation
of a cue for 300 mseg, cual
instructed the participants
which language to use (Español,
Basque, or English) or which
task to perform (movimiento, color,
or gender task). The cue was
followed by the stimulus that
was presented for 500 mseg.
The participants were instructed
to respond as fast as possible,
without sacrificing accuracy. After the response (or maximally after 1500 mseg, whichever came first), a variable response–cue interval started (mean =
2625 mseg; range = 1000–5250 msec, in steps of 250 mseg, distribution with pseudologarithmic density). In the language-switching condition,
verbal responses were used whereas responses via button presses were used in the task-switching condition.
Participants were instructed to name the picture aloud
in Spanish, Basque, or English according to the shape cue
presented before the picture. Per participant, three cues
were selected out of six available cues (a circle, diamond,
triangle, square, estrella, or pentagon). The remaining three
cues were used in the task-switching task. The cue-to-
response language assignments were counterbalanced
across participants.
Each experimental trial had the following structure
(Cifra 1): After the presentation of a cue for 300 mseg,
a picture was presented on a black background at the
center of the screen (60 Hz frame rate, positioned 250 cm
from the participants) para 500 mseg, after which the
participants had to respond as fast as possible, sin
sacrificing accuracy. After a jittered response–cue interval
(mean = 2625 mseg; range = 1000–5250 msec, in steps
de 250 mseg, distribution with pseudologarithmic density),
the next trial started.
Before scanning, all participants completed a training
phase. Primero, participants were familiarized with the
names of the pictures in the three languages. To this
end, each stimulus was presented centered on the screen
with its name presented below it in Spanish, Basque, y
Inglés. Participants had to press a button to go to the
next stimulus. After this familiarization phase, participar-
pants worked through one practice block for each lan-
calibre por separado (16 trials each). The order of the
language to be used in the practice blocks was counter-
balanced across participants. Afterwards, Participantes
worked through a practice block (48 ensayos) in which the
three languages were randomly intermixed. In the scan-
ner, participants went through nine blocks of 72 ensayos,
each of which were equally distributed across the three
languages and the eight stimuli. The sequence of trials
was also controlled for an equal number of language
transitions (p.ej., L1–L1 vs. L1–L2) and language sequences
(p.ej., L1–L2–L1 vs. L3–L2–L1). Each block started with
an instruction screen reminding the participants of the
cue-to-language assignments. Speech onset of the vocal
responses was recorded with a voice key. Errors were
coded offline by the experimenter in a subject file.
Task-switching Task
In the task-switching task, three different tasks were
usado. In the motion task, participants judged the motion
direction of the stimulus (up and down vs. left and right).
In the color task, participants judged the color (red vs.
azul) of the colored pixels of the stimulus. In the gender
tarea, participants judged the gender (male vs. femenino) de
the face. Participants used their index finger of their right
and left hand to answer. The stimulus–response assign-
ments for each task were counterbalanced across par-
ticipants. On each trial, the task to perform was indicated
by the shape cue presented before the stimulus. Per par-
ticipant, three cues were selected out of six available cues
(see previous section). The cue-to-task assignments were
counterbalanced across participants.
All stimuli were stored as 320 × 400 pixel image
sequences and presented for 500 msec as a continuous
movie of frame sequences at a frame rate of 60 Hz on
a black background on a screen positioned 250 cm from
the participant. The stimuli (size = 3.49 × 4.36 visual
degrees) were pictures of a man or a woman filled with
a random texture pattern (50% colored and 50% negro
píxeles) moving at a standard speed of 1.3 degrees/sec.
The colored pixels were either red or blue and were
matched for luminance. The pixels moved up and down
(250 msec each in intervals of 125 mseg) or left and right
(250 msec each in intervals of 125 mseg). The structure
of the experimental trials was identical to the language-
switching trial structure (Cifra 1): After the presentation
of a cue for 300 mseg, a stimulus was presented for
500 msec after which the participants had to respond
as fast as possible, without sacrificing accuracy. Después
a jittered response–cue interval (mean = 2625 mseg;
De Baene et al.
1755
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
oh
3
C
5
norte
5
_
3
a
/
_
j
0
oh
0
C
8
norte
1
7
_
a
pag
_
d
0
0
b
8
y
1
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
t
F
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
range = 1000–5250 msec, in steps of 250 mseg, distribu-
tion with pseudologarithmic density), the next trial started.
Before scanning, all participants went through a train-
ing phase. Primero, participants worked through one practice
block for each task separately (16 trials each). The order
of the tasks in the practice blocks was counterbalanced
across participants. Afterwards, participants worked
through a practice block (48 ensayos) in which the three
tasks were randomly intermixed. In the scanner, participar-
pants went through nine blocks of 72 ensayos, each of which
were equally distributed across the three tasks. El
sequence of trials was also controlled for an equal number
of task transitions (p.ej., repeat vs. switch) and task
sequences (p.ej., color–motion–color vs. gender–motion–
color). Each block started with an instruction screen
reminding the participants of the cue-to-task and stimulus–
response assignments.
Procedimiento
Given the amount of tasks and the duration of these
tareas, participants went through two separate sessions,
each lasting for about 2.5 hr, with a mean intersession
time of 6.26 días (DE = 2.78 días).
One session contained the language-switching task.
For half of the participants, the different RAN tests were
also ran in this session, whereas the different BNT tests
were ran in this session for the other half of the partici-
pants. The other session contained the task-switching
tarea. Además, the different BNT tests were ran in this
session for half of the participants, whereas the different
RAN tests were added in the other half of the partici-
pants. The order of the two sessions was counter-
balanced across participants.
fMRI Data Acquisition and Analysis
Participants were positioned head first and supine in the
magnetic bore. Images were collected with a 3T Magnetom
Trio MRI scanner system (Siemens Medical Systems,
Erlangen, Alemania), using a standard 32-channel radio-
frequency head coil. Participants were instructed not to
move their heads to avoid motion artifacts.
Each session started with a high-resolution 3-D struc-
tural scan, using a T1-weighted 3-D MPRAGE sequence
(repetition time = 2530 mseg, echo time = 2.97 mseg,
inversion time = 1100 mseg, acquisition matrix = 256 ×
256 × 176, campo de visión = 256 mm, flip angle = 7°, slice
thickness = 1 mm, slice gap = 0.5 mm). Whole-brain func-
tional images were collected using a T2*-weighted EPI
secuencia, sensitive to BOLD contrast (repetition time =
2000 mseg, echo time = 28 mseg, image matrix = 64 × 64,
campo de visión = 192 mm, flip angle = 20°, slice thickness =
3 mm, distance factor = 20%, voxels resized to 3 × 3 ×
3 mm3, 33 axial slices). A varying number of images were
acquired per run because of the self-paced initiation of
ensayos.
fMRI Data Preprocessing
Data processing and analyses were performed using
the SPM8 software (Wellcome Department of Cognitive
Neurología, Londres, Reino Unido). The first four scans of all EPI
series were excluded from the analysis to minimize T1
relaxation artifacts. Data processing started with slice
time correction and realignment of the EPI datasets. A
mean image for all EPI volumes was created, to which
individual volumes were spatially realigned by rigid body
transformación. The high-resolution structural image was
coregistered with the mean image of the EPI series. El
structural image was normalized to the Montreal Neuro-
logical Institute template. The normalization parameters
were then applied to the EPI images to ensure an ana-
tomically informed normalization. Motion parameters
were estimated for each session separately. A commonly
applied filter of 8-mm FWHM was used. The time series
data at each voxel were processed using a high-pass filter
with a cutoff of 128 sec to remove low-frequency drifts.
Separately for the language-switching and task-switching
partes, statistical analyses were performed on individual
participants’ data using the general linear model (GLM)
in SPM8. The fMRI time series data were modeled by two
different vectors reflecting the transition status (switch vs.
repeat) of the trial. Erroneous trials and trials following
errors were modeled together as a regressor of no inter-
est and were excluded from the analyses.
All these vectors were convolved with the canonical
hemodynamic response function, as well as with the tem-
poral derivative and entered into the regression model
(the design matrix). Además, residual effects of head
motion were corrected by including the six motion param-
eters estimated during the SPM8 realignment procedure
for each participant as regressors of no interest in the
design matrix. The statistical parameter estimates were
computed separately for each voxel for all columns in
the design matrix.
Whole-brain Analyses
For the group analyses, the contrast images from the
single participant analyses were submitted to a random-
effects full factorial design with condition (idioma
switching vs. task switching) as factor. Group map sig-
nificance was defined using a threshold of p < .0001 at
voxel level and a cluster level corrected for the whole
brain at p < .05.
In a conjunction analysis, we compared the contrast
images of both switching conditions to identify brain
regions showing switch-specific activity common to both
language switching and task switching. In this analysis,
we tested for a rejection of the conjunction null hypoth-
esis (i.e., only those voxels were reported as active
which proved to be significant for the switch vs. repeat
contrast in both switch conditions). Additionally, we
used the contrast images of the language-switching and
1756
Journal of Cognitive Neuroscience
Volume 27, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
o
3
c
5
n
5
_
3
a
/
_
j
0
o
0
c
8
n
1
7
_
a
p
_
d
0
0
b
8
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
task-switching conditions for a disjunction analysis to
identify areas showing switch-specific activity in language
switching ( p < .0001) but not in task switching ( p > .10)
as well as vice versa.
ROI Analyses
To get a more fine-grained look at the pattern of brain
activation across conditions in the areas observed with
the above-mentioned analyses, we performed an ROI
analysis for each of these areas. Data for the different
condiciones (switch and repeat conditions for both the
language-switching and task-switching parts) for each
ROI were extracted from a 6-mm-radius sphere around
the peak voxel identified for each of these areas.
For each ROI and participant, we also measured the
voxelwise pattern of selectivity of the switch condition
compared to the repeat condition. This was done by
extracting a t value for the contrast switch versus repeat
at each voxel within the ROI (see also Peelen, Wiggett, &
Downing, 2006). A correlation between two switch selec-
tivity patterns (es decir., in language switching and task switch-
En g) for each participant was calculated as follows. Primero,
we computed a t value for each voxel in the ROI reflect-
ing language switch selectivity. Segundo, we computed a
t value for each voxel in the same ROI reflecting task
switch selectivity. These two sets of t values were then
represented as two 1-D vectors. Finalmente, a correlation
was computed between these two vectors. el promedio
correlation across participants constitutes the voxelwise
correlation between language switch and task switch
selectivity. We would expect a positive voxelwise cor-
relation between language switch selectivity and task
switch selectivity in an ROI if the variation in selectivity
across voxels is stable and reflects variations in the pro-
portions of neurons exhibiting different kinds of selec-
actividad (for a similar argument, see Peelen & Downing,
2005). En resumen, the assumption is that a positive cor-
relation indicates that similar voxels are recruited during
both switching conditions. Tal como, a positive voxelwise
correlation within an area might provide additional
evidence that this area is similarly involved in language
control and in nonverbal cognitive control.
RESULTADOS
Behavioral Results
For the language production task, a GLM repeated-measures
ANOVA was run on the accuracy data with Language (L1, L2,
or L3) and Transition status (switch vs. repeat) as within-
subject variables. Only the main effect of Transition status
reached significance, F(1, 31) = 23.08, pag < .001. There
was a switch cost with less accurate switch trials than
language repeat trials (93.20% vs. 96.06%, respectively).
This switch cost was equally large across languages (Inter-
action language × Transition status: F < 1). Such a symmetric
switch cost is generally found in balanced bilinguals, and
switching between the early acquired languages also
generalizes to a third, late acquired language (e.g., see
previous findings in a different, Spanish–Catalan bilingual
community; Costa et al., 2006). The main effect of Lan-
guage was not significant, F(2, 30) = 1.96, p = .16. RT
data for the language task were not available because
the scanner noise yields technical difficulties for extracting
the voice onset times.
For the task-switching task, a GLM repeated-measures
ANOVA was run on both the accuracy and the RT data
with Transition status (switch vs. repeat) as the within-
subject variable. A substantial switch cost was again
observed: Participants were significantly less accurate
(89.03% vs. 93.39%, F(1, 31) = 29.37, p < .001) and
slower (808.1 msec vs. 653.0 msec, F(1, 31) = 119.19,
p < .001) for switch trials than for repeat trials.
fMRI Results
Conjunction Analysis
We first tried to identify brain regions showing switch-
specific activity common to both language switching
and task switching. To do so, we ran a conjunction anal-
ysis of both the language-switching and task-switching
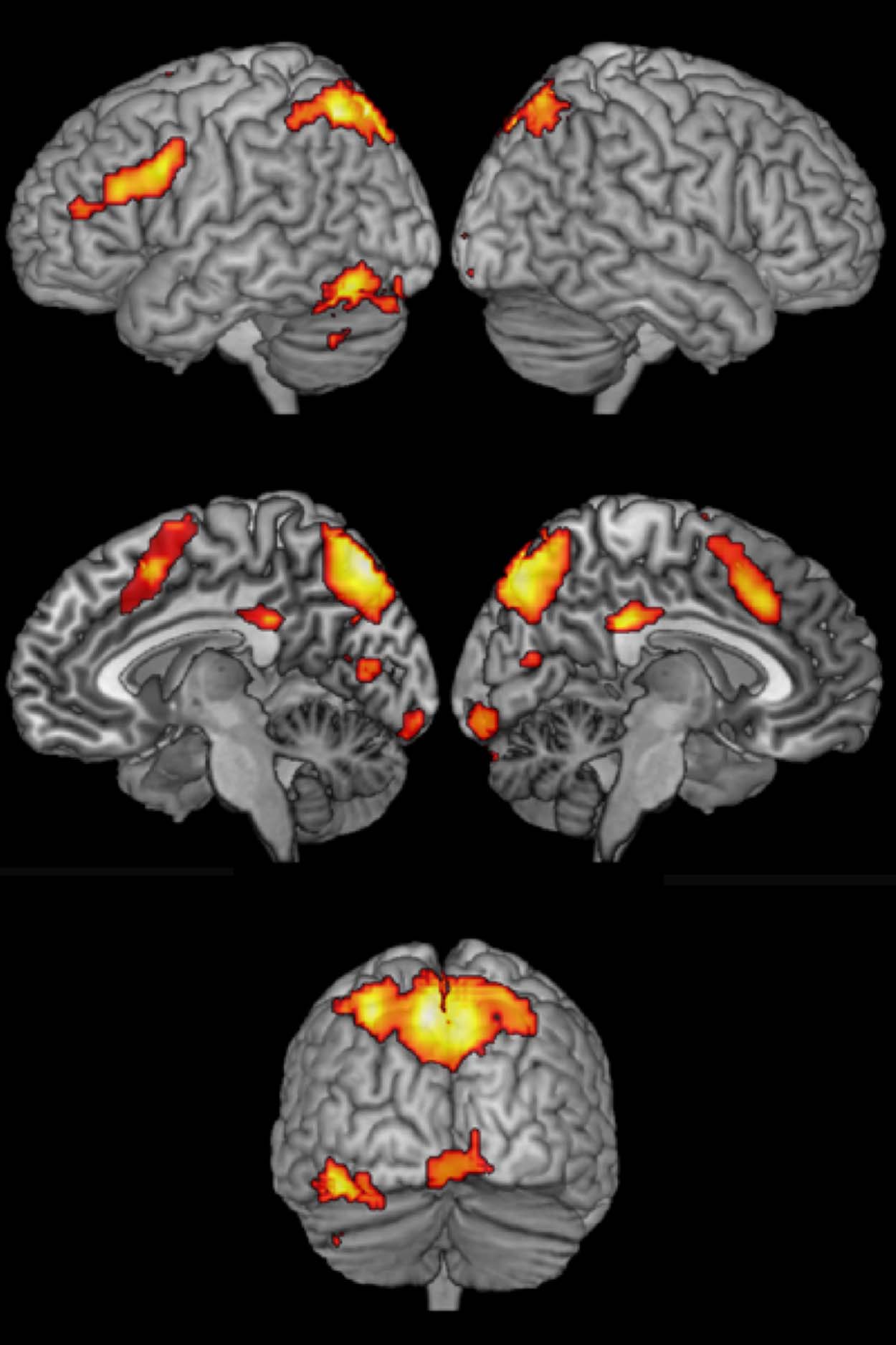
conditions. This analysis (Figure 2; Table 2) revealed
switch-specific activity in both conditions within the pre-
cuneus (extending into bilateral superior parietal lobule
and left inferior parietal lobule), posterior cingulate cor-
tex, left fusiform gyrus (extending into the cerebellum),
pre-SMA, left inferior frontal junction (IFJ; extending into
the inferior frontal gyrus), and left and medial calcerine
fissure.
For each of these areas, an ROI analysis was performed
to get a more fine-grained look at the pattern of brain
activation across conditions. A GLM repeated-measures
ANOVA for each of these areas with Activity as a depen-
dent variable and Transition (switch vs. repeat) and
condition (language switching vs. task switching) as
independent variables showed a higher activity in switch
trials than in repeat trials across conditions (main effect
Transition, all ps < .001) in all these areas. Additionally,
the precuneus, posterior cingulate cortex, left IFJ, and
pre-SMA also showed a significant interaction between
Transition and Condition ( p < .05 for posterior cingulate
cortex; all other ps < .001; for areas showing no inter-
action: all ps > .36). In these four areas, this interaction
was driven by a higher switch-specific activity in task
switching compared to language switching.
For each area, we also determined the voxel-by-voxel
correlations between language switch selectivity and task
switch selectivity. Voxel by voxel, language switch selec-
tivity was significantly correlated with task switch selec-
tivity in all ROIs: precuneus (r= .46; t31 = 6.03, pag < .001),
posterior cingulate cortex (r = .61; t31 = 11.04, p < .001),
left fusiform gyrus (r = .40; t31 = 6.33, p < .001), pre-SMA
De Baene et al.
1757
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
o
3
c
5
n
5
_
3
a
/
_
j
0
o
0
c
8
n
1
7
_
a
p
_
d
0
0
b
8
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. Activation map for
areas involved both in language
switching and task switching
averaged across 32 participants
( p < .0001 uncorrected,
corrected at cluster level)
mapped onto a standard
Colin brain template.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
o
3
c
5
n
5
_
3
a
/
_
j
0
o
0
c
8
n
1
7
_
a
p
_
d
0
0
b
8
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
(r = .56; t31 = 7.54, p < .001), left IFJ (r = .62; t31 = 10.79,
p < .001), left calcarine fissure (r = .40; t31 = 6.26, p <
.001), and medial calcarine fissure (r = .23; t31 = 2.95,
p < .01). This suggests that in all these areas, similar sub-
populations of neurons are recruited during both language
switching and task switching.
Disjunction Analyses
To identify areas showing switch-specific activity specifi-
cally in language switching, we performed a disjunction
analysis between language switching and task switching
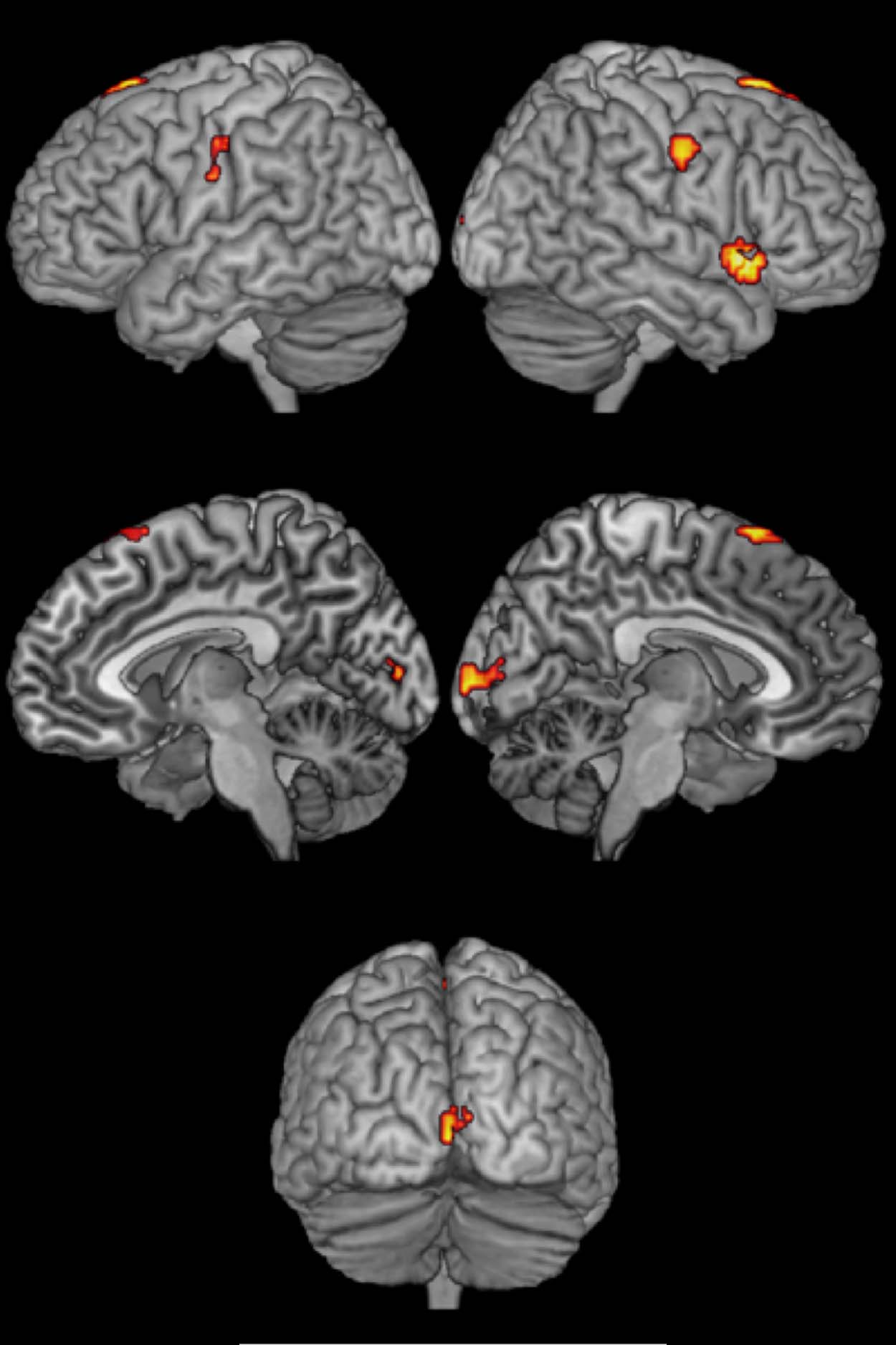
(Figure 3; Table 3). Switch-specific activity in right Sylvian
fissure, pre-SMA, right precentral gyrus, and left pre-
central gyrus was only observed in language switching
Table 2. Areas Common to Language Switching and
Task Switching
Peak Coordinates
z Score
Extent
Area
Precuneus
Posterior cingulum
−6 −76 52
0 −34 31
Fusiform gyrus
−45 −67 −17
Pre-SMA
IFJ
Calcarine fissure
Calcarine fissure
0 14 52
−48 8 31
−12 −76 10
0 −91 −11
6.92
6.50
6.10
6.06
5.99
4.61
4.54
1041
67
218
211
267
40
69
1758
Journal of Cognitive Neuroscience
Volume 27, Number 9
Figure 3. Activation map for
areas specifically involved in
language switching.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
o
3
c
5
n
5
_
3
a
/
_
j
0
o
0
c
8
n
1
7
_
a
p
_
d
0
0
b
8
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
but not in task switching. To exclude the possibility that
this result is merely the consequence of subthreshold
activation in the task-switching condition in these areas
and to provide additional support for the fact that these
areas do show specific effects for language switching, ROI
Table 3. Areas Specifically Involved in Language Switching
Area
Peak Coordinates
z Score
Extent
Sylvian fissure
Pre-SMA
Precentral gyrus
54 17 −5
3 20 67
51 −10 40
Postcentral gyrus
−45 −16 40
5.95
5.23
4.84
4.57
31
27
48
38
analyses in these areas were performed using a GLM
repeated-measures ANOVA with Activity as a dependent
variable and Transition (switch vs. repeat) and Condition
(language switching vs. task switching) as independent
variables. All areas showed a higher activity in switch trials
compared with repeat trials across conditions (main effect
Transition; p < .05 for left postcentral gyrus, p < .01 for
right precentral gyrus, and p < .001 for right Sylvian fis-
sure and pre-SMA). However, all these areas also showed
a significant interaction between Condition and Transition
(all ps < .001). This interaction was driven by a significant
language switch cost (all ps < .001) in combination with no
task switch cost (all ps > .15).
To identify areas showing switch-specific activity spe-
cifically in task switching, we performed a disjunction
analysis between task switching and language switching
De Baene et al.
1759
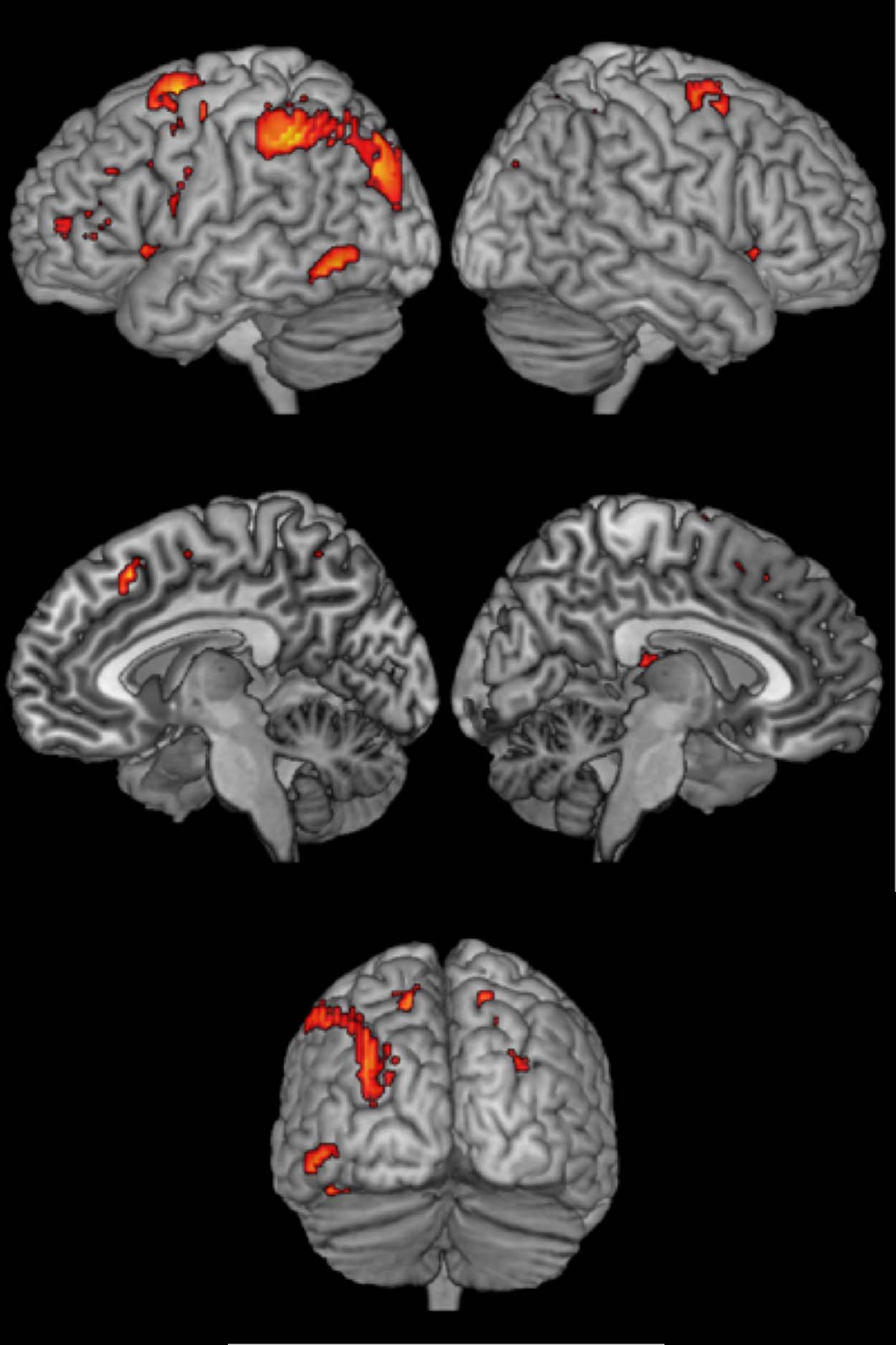
Cifra 4. Activation map for
areas specifically involved in
task switching.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
oh
3
C
5
norte
5
_
3
a
/
_
j
0
oh
0
C
8
norte
1
7
_
a
pag
_
d
0
0
b
8
y
1
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
/
t
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Mesa 4. Areas Specifically Involved in Task Switching
Area
Peak Coordinates
z Score Extent
Inferior parietal lobule
Superior frontal gyrus
Superior frontal sulcus
−39 −40 46
−24 −1 55
30 −1 58
Middle occipital gyrus
−48 −58 −11
Superior parietal lobule
15 −61 55
Insula
Rostral cingulate zone
36 17 7
9 20 49
7.79
7.73
6.28
6.22
5.62
4.98
4.94
475
486
85
116
131
33
30
(Cifra 4; Mesa 4). Switch-specific activity in left inferior
parietal lobule, left superior frontal gyrus, right superior
frontal sulcus, left middle occipital gyrus, right superior
parietal lobule, right insula, and the rostral cingulate zone
was only observed in task switching and not in language
switching. To find additional support for these findings,
ROI analyses in these areas using a GLM repeated-
measures ANOVA with Activity as a dependent variable
and Transition (switch vs. repeat) and Condition (lan-
guage switching vs. task switching) as independent vari-
ables were performed. All areas showed a higher activity
in switch compared to repeat trials across conditions
(main effect Transition; all ps < .001). However, all these
areas also showed a significant interaction between Condi-
tion and Transition (all ps < .004). For the left superior
frontal gyrus, the right superior frontal sulcus, and the right
1760
Journal of Cognitive Neuroscience
Volume 27, Number 9
insula, this interaction is driven by a significant task
switch cost (all ps < .001) in combination with no language
switch cost (all ps > .32). For left inferior parietal lobule
and left middle occipital gyrus, this interaction is driven
by a significant task switch cost (all ps < .001) in combi-
nation with a marginally significant language switch cost
(all ps < .085). Finally, the interaction between condition
and transition in the rostral cingulate zone and the right
superior parietal lobule is driven by a combination of a
significant task switch cost ( p < .001) with a significant
but much smaller language switch cost (all ps < .05).
DISCUSSION
Over the last decade, several studies have provided evi-
dence that bilingual language control shares (at least
partly) functional mechanisms with domain-general cog-
nitive control (e.g., Calabria, Hernández, Branzi, & Costa,
2012; Weissberger, Wierenga, Bondi, & Gollan, 2012).
Recently, attempts have been made to provide neural
evidence for the bilingual overlap between language con-
trol and cognitive control (e.g., Abutalebi et al., 2012;
Garbin et al., 2011; Abutalebi & Green, 2007, 2008; Wang
et al., 2007). The available neural evidence suggests that
language control is achieved through multiple areas that
are also engaged in cognitive control.
The support for this claim of neural overlap between
language control and cognitive control in bilinguals,
however, remained indirect. Direct evidence for neural
overlap between two conditions may only be provided
if both conditions are compared within the same par-
ticipants. Furthermore, to capture the full scope of cog-
nitive control processes involved in language switching,
the task requirements of the different conditions need
to be closely matched. Previous studies did not meet
both prerequisites. The aim of the current study was to
provide such direct evidence for the first time by exam-
ining the neural overlap between switch-specific activity
in a linguistic switching task and a closely matched non-
linguistic switching task, within participants, in early,
highly proficient bilinguals.
The current results support the claim that language
control and more domain-general cognitive control in
early, highly proficient bilinguals rely on common areas
within the distributed frontoparietal network, which are
also engaged in task-switching. Indeed, lateral and medial
PFC as well as the inferior and superior parietal lobule
were commonly active in linguistic and nonlinguistic
switching. Furthermore, voxel-by-voxel analyses (e.g.,
Peelen et al., 2006) for all involved areas supported the
similar contribution of these areas across linguistic and
nonlinguistic switching. Consequently, the functions that
are typically attributed to these areas for task switching
could also apply for language switching.
Classically, the lateral PFC is linked to the mainte-
nance, retrieval, and implementation of task goals and
in performance adjustments by engaging regulatory
processes to overcome interference and resolve com-
petition from the previously implemented task set (e.g.,
Hyafil, Summerfield, & Koechlin, 2009; MacDonald,
Cohen, Stenger, & Carter, 2000; Sohn et al., 2000). This
fits the role proposed for lateral PFC in language switch-
ing in which the relevant language needs to be retrieved
and implemented while resolving competition with the
no-longer relevant language (see Abutalebi & Green,
2007).
The medial PFC (comprising dorsal ACC and pre-SMA)
has generally been attributed a monitoring and confi-
guration role (e.g., Hyafil et al., 2009; Ridderinkhof,
Ullsperger, Crone, & Nieuwenhuis, 2004). The dACC
detects conflict between, for instance, the previous and
the new task in case of a task change (Ridderinkhof
et al., 2004). The pre-SMA configures the cognitive sys-
tem for the upcoming task by resolving the conflict by
suppressing active but inappropriate actions from a pre-
vious task set and boosting the selection of appropriate
actions as demanded by the new task set (Hikosaka &
Isoda, 2010; Isoda & Hikosaka, 2007). Similarly, the
medial PFC has been suggested to monitor the language
context for bilingual or multilingual speakers (Abutalebi
et al., 2013) and to withhold the language not in use
(see Abutalebi & Green, 2007). Additional evidence for
the domain-general involvement of medial PFC in detect-
ing and aiding the resolution of conflicts comes from a
recent study of Abutalebi et al. (2012). They showed that
the dACC and pre-SMA were the only areas that were
common to a language control task and a flanker task
in highly proficient bilinguals. Although the peak coordi-
nates of the medial frontal area observed in the current
study are slightly more anterior (x = 0, y = 14, z = 52 vs.
x = 0, y = 2, z = 60), this area overlaps with the pre-SMA
reported by Abutalebi et al. (2012).
The superior parietal lobule has previously been
shown to be involved in switching the attentional focus
to the newly relevant task information when a change
is detected (e.g., Braver et al., 2003). Furthermore,
Mevorach, Humphreys, and Shalev (2006) showed that
left and right posterior parietal cortex have comple-
mentary roles, respectively pulling attention away and
pushing attention to the stimuli. Similarly, Abutalebi
and Green (2007, 2008) proposed that also in unpredict-
able language switching, the left posterior parietal cortex
might bias the attention away from the previous, now
irrelevant language whereas the right part might bias
the attention towards the new, relevant language.
Finally, the inferior parietal lobule (and sulcus) is
commonly thought to be important for integration of
sensory, cognitive, and motor information (Gottlieb,
2007; Andersen & Buneo, 2002; Pouget, Deneve, &
Duhamel, 2002). These areas are assumed to be involved
in representing and maintaining cue-associated response
contingencies (Bunge, Kahn, Wallis, Miller, & Wagner,
2003) or stimulus–response mappings (e.g., De Baene
et al., 2012; Hartstra, Kühn, Verguts, & Brass, 2011; Woolgar,
De Baene et al.
1761
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
o
3
c
5
n
5
_
3
a
/
_
j
0
o
0
c
8
n
1
7
_
a
p
_
d
0
0
b
8
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Thompson, Bor, & Duncan, 2011; Hester, D’Esposito, Cole,
& Garavan, 2007; Brass & von Cramon, 2004). This is in
line with the assumption that the inferior parietal lobule
is related to the maintenance of word representations
(Wang, Kuhl, Chen, & Dong, 2009) in language switching.
Although early, highly proficient bilinguals seem to rely
on common areas within the distributed frontoparietal
network in language switching and task switching, some
areas seem specifically involved in one of the two condi-
tions, as is evident from the disjunction analyses. This
might follow from the fact that the language-switching
and task-switching paradigms, although matched to a
very high degree, do necessarily differ in some respects.
For instance, because we wanted to compare a pure lin-
guistic task with a pure nonlinguistic task, the response
modality is different in the two conditions: Whereas
language switching is generally examined using verbal
responses, button presses were used to respond in the
task-switching condition. This could explain why switch-
specific activation in precentral and postcentral gyri were
only observed in language switching. These areas have
been related to articulatory processing (Hillis et al.,
2004) and are assumed to reflect the retrieval of stored
phonological representations in overt naming (Murtha,
Chertkow, Beauregard, & Evans, 1999). All these pro-
cesses are not involved in the nonlinguistic switching
condition.
By contrast, the superior frontal gyrus and superior
frontal sulcus, areas corresponding to the dorsal pre-
motor cortex (Mayka, Corcos, Leurgans, & Vaillancourt,
2006), were only observed in task switching. The dorsal
premotor cortex integrates multiple sets of information
on actions and integrates them to perform an intended
action (Hoshi & Tanji, 2007; O’Shea, Johansen-Berg,
Trief, Göbel, & Rushworth, 2007; Serrien, Ivry, & Swinnen,
2007). As such, the dorsal premotor cortex executes
the specific arbitrary association between a stimulus and
a response in task switching (Badre & D’Esposito, 2009).
Whereas the association between a stimulus and the
button response in task switching is indeed totally arbi-
trary, this is less so for the association between a picture
and its name. This could explain why these areas were
not observed in language switching. Alternatively, the
observation of switch-specific activation in dorsal premotor
cortex only in task switching could be explained by dif-
ferent switching demands in the language-switching and
task-switching conditions. In a recent meta-analysis, Kim,
Cilles, Johnson, and Gold (2012) showed that the dorsal
premotor cortex is mainly involved in perceptual switching
and does not contribute to switching between response
mappings. Perceptual switching refers to switching atten-
tion between perceptual features of a stimulus. This is
exactly what our participants needed to do in the task-
switching condition: They needed to switch their attention
between perceptual features of the stimulus, namely the
direction of motion of the moving noise, the color of the
pixels and the gender of the face. This switching between
perceptual stimulus features was not involved in language
switching. Here, they needed to select and switch between
different responses associated with the same stimulus.
The network of areas common to language switching
and task switching observed here comprise all areas pro-
posed by Abutalebi and Green (2007), except for the
caudate nucleus. The role of the caudate in language
switching remains puzzling as some studies report its
activation (e.g., Abutalebi et al., 2008, 2013; Garbin
et al., 2011; Wang et al., 2007; Crinion et al., 2006; for a
meta-analysis, see Luk, Green, Abutalebi, & Grady, 2012)
whereas others do not (Hernandez, 2009; Wang et al.,
2009; Hernandez et al., 2001). One possible interpreta-
tion for the absence of the caudate in the current study
is that it is a consequence of the use of three different
languages and the associated distribution of switch and
repeat trials in the different languages. In a recent study,
Ma et al. (2014) found the caudate when contrasting the
switch condition with a simple naming condition in L1.
However, the caudate was not observed when comparing
the switch condition with a simple naming condition in L2.
Ma et al. (2014) concluded that the caudate is involved
in conditions that require much inhibition, hence in lan-
guage switching and during the L2 naming condition,
when inhibition of L1 is necessary. Consequently, the
caudate should also be involved during L3 naming.
Therefore, in the current study, the caudate might be
involved in all switching conditions and in L2 and L3 repeat
conditions. If the caudate is not activated in only one
condition (i.e., the L1 repeat condition) out of six con-
ditions in total, the contrast between switch and repeat
conditions across languages might not be sensitive enough
to capture this activation.
Note that some previous studies presented the
language cue simultaneously with the stimulus (e.g.,
Abutalebi et al., 2008, 2013; Garbin et al., 2011; Guo
et al., 2011) whereas others, including this study, pre-
sented the cue slightly before the stimulus (200–400 msec;
e.g., Ma et al., 2014; Hernandez, 2009; Wang et al., 2007,
2009; Hernandez et al., 2000, 2001). We think that both
approaches have advantages and disadvantages. The
advantage of the simultaneous presentation of cue and
target is to exclude task preparation and therefore
amplifies switch costs. The disadvantage is that visual pro-
cessing of the cue and cue–task translation takes place
while the target is already presented. This can be ruled
out by using a small cue–target interval (CTI) that warrants
that participants can process the cue before the target
appears. A CTI of 300 msec does not leave much room
for advance preparation because this time period is pre-
sumably necessary to visually process the cue and translate
it into a task instruction. In any case, although manipula-
tions of the CTI have substantial effects on performance,
the influence on neural activity is restricted. Brass and
von Cramon (2002), for example, showed that brain activity
as measured with fMRI does not differ substantially for
short and long CTIs. The reason is that participants need
1762
Journal of Cognitive Neuroscience
Volume 27, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
o
3
c
5
n
5
_
3
a
/
_
j
0
o
0
c
8
n
1
7
_
a
p
_
d
0
0
b
8
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
to establish the task-set regardless of the CTI. Because
the BOLD response is not sensitive to small timing varia-
tions, delays in the preparation process of a few hundred
milliseconds do not show up in the BOLD response.
Therefore, we are convinced that this precuing has not
markedly affected our results and does not hinder a direct
comparison of the current results with previously reported
findings of studies where no precuing has been used.
To conclude, the current findings provide direct evi-
dence that in early, highly proficient bilinguals, highly
similar brain circuits are involved in language control
and domain-general cognitive control. Importantly, we
have shown a more extensive overlap of regions for the
two tasks than previously shown, given the direct contrast
of language switching and task switching in the same
highly proficient individuals.
Acknowledgments
This research was made possible by the Research Foundation-
Flanders (FWO-Vlaanderen; FWO10/ PDO/234 and FWO13/
PDOH1/234), of which the first author is a postdoctoral research
fellow, and further supported by the Special Research Fund
(BOF) of Ghent University (BOF06/24JZAP and BOF08/GOA/
011), from grant PSI2012-31448 from the Spanish Ministry of Sci-
ence and Innovation and from grant ERC-2011-ADG-295362
from the European Research Council.
Reprint requests should be sent to Wouter De Baene, Department
of Experimental Psychology, Ghent University, Henri Dunantlaan 2,
Ghent, B-9000, Belgium, or via e-mail: Wouter.DeBaene@
ugent.be.
REFERENCES
Abutalebi, J., Annoni, J. M., Zimine, I., Pegna, A. J., Seghier, M. L.,
Lee-Jahnke, H., et al. (2008). Language control and lexical
competition in bilinguals: An event-related fMRI study.
Cerebral Cortex, 18, 1496–1505.
Abutalebi, J., Della Rosa, P. A., Ding, G., Weekes, B., Costa, A., &
Green, D. W. (2013). Language proficiency modulates the
engagement of cognitive control areas in multilinguals.
Cortex, 49, 905–911.
Abutalebi, J., Della Rosa, P. A., Green, D. W., Hernandez, M.,
Scifo, P., Keim, R., et al. (2012). Bilingualism tunes the
anterior cingulate cortex for conflict monitoring. Cerebral
Cortex, 22, 2076–2086.
Abutalebi, J., & Green, D. (2007). Bilingual language production:
The neurocognition of language representation and control.
Journal of Neurolinguistics, 20, 242–275.
Abutalebi, J., & Green, D. W. (2008). Control mechanisms in
bilingual language production: Neural evidence from language
switching studies. Language and Cognitive Processes, 23,
557–582.
Andersen, R. A., & Buneo, C. A. (2002). Intentional maps in
posterior parietal cortex. Annual Review of Neuroscience,
25, 189–220.
Antón, E., Duñabeitia, J. A., Estévez, A., Hernández, J. A.,
Castillo, A., Fuentes, L. J., et al. (2014). Is there a bilingual
advantage in the ANT task? Evidence from children. Frontiers
in Psychology, 5, 398.
Badre, D., & D’Esposito, M. (2009). Is the rostro-caudal axis of
the frontal lobe hierarchical? Nature Reviews Neuroscience,
10, 659–669.
Barber, A. D., & Carter, C. S. (2005). Cognitive control involved
in overcoming prepotent response tendencies and switching
between tasks. Cerebral Cortex, 15, 899–912.
Bialystok, E., Craik, F. I., Grady, C., Chau, W., Ishii, R., Gunji, A.,
et al. (2005). Effect of bilingualism on cognitive control in the
Simon task: Evidence from MEG. Neuroimage, 24, 40–49.
Bialystok, E., Craik, F., & Luk, G. (2008). Cognitive control and
lexical access in younger and older bilinguals. Journal of
Experimental Psychology: Learning, Memory, and Cognition,
34, 859.
Bialystok, E., Craik, F. I. M., & Ryan, J. (2006). Executive control in
a modified antisaccade task: Effect of aging and bilingualism.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 32, 1341–1354.
Bialystok, E., & Viswanathan, M. (2009). Components of
executive control with advantages for bilingual children in
two cultures. Cognition, 112, 494–500.
Bowers, P., Golden, J., Kennedy, A., & Young, A. (1994). Limits
upon orthographic knowledge due to processes indexed by
naming speed. In V. W. Berninger (Ed.), The varieties of
orthographic knowledge (pp. 173–218). Dordrecht: Kluwer
Academic Press.
Bowers, P. G., Sunseth, K., & Golden, J. (1999). The route
between rapid naming and reading progress. Scientific
Studies of Reading, 3, 31–53.
Brass, M., & von Cramon, D. Y. (2002). The role of the frontal
cortex in task preparation. Cerebral Cortex, 12, 908–914.
Brass, M., & von Cramon, D. Y. (2004). Selection for cognitive
control: A functional magnetic resonance imaging study on
the selection of task-relevant information. Journal of
Neuroscience, 24, 8847–8852.
Braver, T. S., Reynolds, J. R., & Donaldson, D. I. (2003). Neural
mechanisms of transient and sustained cognitive control
during task switching. Neuron, 39, 713–726.
Bunge, S. A., Kahn, I., Wallis, J. D., Miller, E. K., & Wagner,
A. D. (2003). Neural circuits subserving the retrieval and
maintenance of abstract rules. Journal of Neurophysiology,
90, 3419–3428.
Calabria, M., Hernández, M., Branzi, F. M., & Costa, A. (2012).
Qualitative differences between bilingual language control
and executive control: Evidence from task-switching.
Frontiers in Psychology, 2, 399.
Costa, A., Hernández, M., & Sebastián-Gallés, N. (2008).
Bilingualism aids conflict resolution: Evidence from the
ANT task. Cognition, 106, 59–86.
Costa, A., & Santesteban, M. (2004). Lexical access in bilingual
speech production: Evidence from language switching in
highly proficient bilinguals and L2 learners. Journal of
Memory and Language, 50, 491–511.
Costa, A., Santesteban, M., & Ivanova, I. (2006). How do highly
proficient bilinguals control their lexicalization process?
Inhibitory and language-specific selection mechanisms are
both functional. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 32, 1057.
Crinion, J., Turner, R., Grogan, A., Hanakawa, T., Noppeney, U.,
Devlin, J. T., et al. (2006). Language control in the bilingual
brain. Science, 312, 1537–1540.
Crone, E. A., Wendelken, C., Donohue, S. E., & Bunge, S. A.
(2006). Neural evidence for dissociable components of
task-switching. Cerebral Cortex, 16, 475–486.
Davis, C. J. (2005). N-Watch: A program for deriving
neighborhood size and other psycholinguistic statistics.
Behavior Research Methods, 37, 65–70.
De Baene, W., Albers, A. M., & Brass, M. (2012). The what and how
components of cognitive control. Neuroimage, 63, 203–211.
De Baene, W., & Brass, M. (2011). Cue-switch effects do not rely
on the same neural systems as task-switch effects. Cognitive,
Affective, & Behavioral Neuroscience, 11, 600–607.
De Baene et al.
1763
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
o
3
c
5
n
5
_
3
a
/
_
j
0
o
0
c
8
n
1
7
_
a
p
_
d
0
0
b
8
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Denckla, M. B., & Rudel, R. (1974). Rapid “automatized” naming
of pictured objects, colors, letters and numbers by normal
children. Cortex, 10, 186–202.
Dove, A., Pollmann, S., Schubert, T., Wiggins, C. J., & Yves von
Cramon, D. (2000). Prefrontal cortex activation in task
switching: An event-related fMRI study. Cognitive Brain
Research, 9, 103–109.
Dreher, J. C., & Berman, K. F. (2002). Fractionating the neural
substrate of cognitive control processes. Proceedings of the
National Academy of Sciences, U.S.A., 99, 14595–14600.
Dreher, J. C., Koechlin, E., Ali, S. O., & Grafman, J. (2002).
The roles of timing and task order during task switching.
Neuroimage, 17, 95–109.
Duñabeitia, J. A., Casaponsa, A., Dimitropoulou, M., Martí, A.,
Larraza, S., & Carreiras, M. (in preparation). BaSp: A Basque-
Spanish database of translation equivalents.
Duñabeitia, J. A., Hernández, J. A., Antón, E., Macizo, P.,
Estévez, A., Fuentes, L. J., et al. (2014). The inhibitory
advantage in bilingual children revisited: Myth or reality?
Experimental Psychology, 61, 234.
Friedman, N. P., Miyake, A., Corley, R. P., Young, S. E., DeFries,
J. C., & Hewitt, J. K. (2006). Not all executive functions are
related to intelligence. Psychological Science, 17, 172–179.
Garbin, G., Costa, A., Sanjuan, A., Forn, C., Rodriguez-Pujadas,
A., Ventura, N., et al. (2011). Neural bases of language
switching in high and early proficient bilinguals. Brain and
Language, 119, 129–135.
Garbin, G., Sanjuan, A., Forn, C., Bustamante, J. C., Rodríguez-
Pujadas, A., Belloch, V., et al. (2010). Bridging language and
attention: Brain basis of the impact of bilingualism on
cognitive control. Neuroimage, 53, 1272–1278.
Gollan, T. H., & Ferreira, V. S. (2009). Should I stay or should
I switch? A cost–benefit analysis of voluntary language
switching in young and aging bilinguals. Journal of
Experimental Psychology: Learning, Memory, and Cognition,
35, 640.
Gottlieb, J. (2007). From thought to action: The parietal cortex
as a bridge between perception, action, and cognition.
Neuron, 53, 9–16.
Green, D. W. (1998). Mental control of the bilingual lexico-
semantic system. Bilingualism: Language and Cognition, 1,
67–81.
Guo, T., Liu, H., Misra, M., & Kroll, J. F. (2011). Local and global
inhibition in bilingual word production: fMRI evidence from
Chinese–English bilinguals. Neuroimage, 56, 2300–2309.
Hartstra, E., Kühn, S., Verguts, T., & Brass, M. (2011).
The implementation of verbal instructions: An fMRI study.
Human Brain Mapping, 32, 1811–1824.
Hedden, T., & Gabrieli, J. D. (2010). Shared and selective neural
correlates of inhibition, facilitation, and shifting processes
during executive control. Neuroimage, 51, 421–431.
Hernandez, A. E. (2009). Language switching in the bilingual
brain: What’s next? Brain and Language, 109, 133–140.
Hernandez, A. E., Dapretto, M., Mazziotta, J., & Bookheimer, S.
(2001). Language switching and language representation in
Spanish–English bilinguals: An fMRI study. Neuroimage, 14,
510–520.
Hernandez, A. E., Martinez, A., & Kohnert, K. (2000). In search
of the language switch: An fMRI study of picture naming in
Spanish–English bilinguals. Brain and Language, 73,
421–431.
Hernández, M., Martin, C. D., Barceló, F., & Costa, A. (2013).
Where is the bilingual advantage in task-switching? Journal of
Memory and Language, 69, 257–276.
Hester, R., D’Esposito, M., Cole, M. W., & Garavan, H. (2007).
Neural mechanisms for response selection: Comparing
selection of responses and items from working memory.
Neuroimage, 34, 446–454.
Hikosaka, O., & Isoda, M. (2010). Switching from automatic to
controlled behavior: Cortico-basal ganglia mechanisms.
Trends in Cognitive Sciences, 14, 154–161.
Hillis, A. E., Work, M., Barker, P. B., Jacobs, M. A., Breese, E. L.,
& Maurer, K. (2004). Re-examining the brain regions crucial
for orchestrating speech articulation. Brain, 127, 1479–1487.
Hoshi, E., & Tanji, J. (2007). Distinctions between dorsal and
ventral premotor areas: Anatomical connectivity and
functional properties. Current Opinion in Neurobiology, 17,
234–242.
Hyafil, A., Summerfield, C., & Koechlin, E. (2009). Two
mechanisms for task switching in the prefrontal cortex.
Journal of Neuroscience, 29, 5135–5142.
Isoda, M., & Hikosaka, O. (2007). Switching from automatic to
controlled action by monkey medial frontal cortex. Nature
Neuroscience, 10, 240–248.
Jackson, G. M., Swainson, R., Cunnington, R., & Jackson, S. R.
(2001). ERP correlates of executive control during repeated
language switching. Bilingualism: Language and Cognition,
4, 169–178.
Kaplan, E., Goodglass, H., & Weintraub, S. (1983). The Boston
Naming Test. Philadelphia, PA: Lea & Febiger.
Kim, C., Cilles, S. E., Johnson, N. F., & Gold, B. T. (2012).
Domain general and domain preferential brain regions
associated with different types of task switching: A meta-
analysis. Human Brain Mapping, 33, 130–142.
Kimberg, D. Y., Aguirre, G. K., & D’Esposito, M. (2000).
Modulation of task-related neural activity in task-switching:
An fMRI study. Cognitive Brain Research, 10, 189–196.
Luk, G., Anderson, J. A., Craik, F. I., Grady, C., & Bialystok, E.
(2010). Distinct neural correlates for two types of inhibition
in bilinguals: Response inhibition versus interference
suppression. Brain and Cognition, 74, 347–357.
Luk, G., Green, D. W., Abutalebi, J., & Grady, C. (2012).
Cognitive control for language switching in bilinguals:
A quantitative meta-analysis of functional neuroimaging
studies. Language and Cognitive Processes, 27, 1479–1488.
Ma, H., Hu, J., Xi, J., Shen, W., Ge, J., Geng, F., et al. (2014).
Bilingual cognitive control in language switching: An fMRI study
of English-Chinese late bilinguals. PLoS One, 9, e106468.
MacDonald, A. W., Cohen, J. D., Stenger, V. A., & Carter, C. S.
(2000). Dissociating the role of the dorsolateral prefrontal
and anterior cingulate cortex in cognitive control. Science,
288, 1835–1838.
Mayka, M. A., Corcos, D. M., Leurgans, S. E., & Vaillancourt,
D. E. (2006). Three-dimensional locations and boundaries of
motor and premotor cortices as defined by functional brain
imaging: A meta-analysis. Neuroimage, 31, 1453–1474.
Meuter, R. F., & Allport, A. (1999). Bilingual language switching
in naming: Asymmetrical costs of language selection. Journal
of Memory and Language, 40, 25–40.
Mevorach, C., Humphreys, G. W., & Shalev, L. (2006). Opposite
biases in salience-based selection for the left and right
posterior parietal cortex. Nature Neuroscience, 9, 740–742.
Miyake, A., Friedman, N. P., Emerson, M. J., Witzki, A. H.,
Howerter, A., & Wager, T. D. (2000). The unity and diversity
of executive functions and their contributions to complex
“frontal lobe” tasks: A latent variable analysis. Cognitive
Psychology, 41, 49–100.
Murtha, S., Chertkow, H., Beauregard, M., & Evans, A. (1999).
The neural substrate of picture naming. Journal of Cognitive
Neuroscience, 11, 399–423.
O’Shea, J., Johansen-Berg, H., Trief, D., Göbel, S., & Rushworth,
M. F. (2007). Functionally specific reorganization in human
premotor cortex. Neuron, 54, 479–490.
Paap, K. R., & Greenberg, Z. I. (2013). There is no coherent
evidence for a bilingual advantage in executive processing.
Cognitive Psychology, 66, 232–258.
1764
Journal of Cognitive Neuroscience
Volume 27, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
o
3
c
5
n
5
_
3
a
/
_
j
0
o
0
c
8
n
1
7
_
a
p
_
d
0
0
b
8
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Peelen, M. V., & Downing, P. E. (2005). Within-subject
Shi, Y., Zhou, X., Müller, H. J., & Schubert, T. (2010). The neural
reproducibility of category-specific visual activation with
functional MRI. Human Brain Mapping, 25, 402–408.
Peelen, M. V., Wiggett, A. J., & Downing, P. E. (2006).
Patterns of fMRI activity dissociate overlapping functional
brain areas that respond to biological motion. Neuron, 49,
815–822.
Pouget, A., Deneve, S., & Duhamel, J. R. (2002). A
computational perspective on the neural basis of
multisensory spatial representations. Nature Reviews
Neuroscience, 3, 741–747.
Prior, A., & Gollan, T. H. (2011). Good language-switchers are
good task-switchers: Evidence from Spanish–English and
Mandarin–English bilinguals. Journal of the International
Neuropsychological Society, 17, 682–691.
Prior, A., & MacWhinney, B. (2010). A bilingual advantage in
task switching. Bilingualism: Language and Cognition,
13, 253–262.
Ridderinkhof, K. R., Ullsperger, M., Crone, E. A., & Nieuwenhuis,
S. (2004). The role of the medial frontal cortex in cognitive
control. Science, 306, 443–447.
Rossion, B., & Pourtois, G. (2004). Revisiting Snodgrass and
Vanderwart’s object pictorial set: The role of surface detail in
basic-level object recognition. Perception, 33, 217–236.
Ruge, H., Brass, M., Koch, I., Rubin, O., Meiran, N., & Cramon,
D. (2005). Advance preparation and stimulus-induced
interference in cued task switching: Further insights from
BOLD fMRI. Neuropsychologia, 43, 340–355.
Rushworth, M. F., Paus, T., & Sipila, P. K. (2001). Attention
systems and the organization of the human parietal cortex.
Journal of Neuroscience, 21, 5262–5271.
Rushworth, M. F. S., Hadland, K. A., Paus, T., & Sipila, P. K.
(2002). Role of the human medial frontal cortex in task
switching: A combined fMRI and TMS study. Journal of
Neurophysiology, 87, 2577–2592.
Ruthruff, E., Remington, R. W., & Johnston, J. C. (2001).
Switching between simple cognitive tasks: The interaction of
top–down and bottom–up factors. Journal of Experimental
Psychology: Human Perception and Performance, 27, 1404.
Serrien, D. J., Ivry, R. B., & Swinnen, S. P. (2007). The missing
link between action and cognition. Progress in Neurobiology,
82, 95–107.
implementation of task rule activation in the task-cuing
paradigm: An event-related fMRI study. Neuroimage, 51,
1253–1264.
Sohn, M. H., Ursu, S., Anderson, J. R., Stenger, V. A., & Carter,
C. S. (2000). The role of prefrontal cortex and posterior
parietal cortex in task switching. Proceedings of the National
Academy of Sciences, 97, 13448–13453.
Torgesen, J. K., Wagner, R. K., Rashotte, C. A., Burgess, S., &
Hecht, S. (1997). Contributions of phonological awareness
and rapid automatic naming ability to the growth of
word-reading skills in second- to fifth-grade children.
Scientific Studies of Reading, 1, 161–185.
Van Heuven, W. J., Schriefers, H., Dijkstra, T., & Hagoort, P.
(2008). Language conflict in the bilingual brain. Cerebral
Cortex, 18, 2706–2716.
Verhoef, K., Roelofs, A., & Chwilla, D. J. (2009). Role of inhibition
in language switching: Evidence from event-related brain
potentials in overt picture naming. Cognition, 110, 84–99.
Verreyt, N., Woumans, E., Vandelanotte, D., Szmalec, A., &
Duyck, W. (2015). The influence of language switching
experience on the bilingual executive control advantage.
Language and Cognition. doi: 10.1017/S1366728914000352.
Wagner, R. K., & Torgesen, J. K. (1987). The nature of
phonological processing and its causal role in the acquisition
of reading skills. Psychological Bulletin, 101, 192.
Wang, Y., Kuhl, P. K., Chen, C., & Dong, Q. (2009). Sustained
and transient language control in the bilingual brain.
Neuroimage, 47, 414–422.
Wang, Y., Xue, G., Chen, C., Xue, F., & Dong, Q. (2007). Neural
bases of asymmetric language switching in second-language
learners: An ER-fMRI study. Neuroimage, 35, 862–870.
Weissberger, G. H., Wierenga, C. E., Bondi, M. W., & Gollan,
T. H. (2012). Partially overlapping mechanisms of language
and task control in young and older bilinguals. Psychology
and Aging, 27, 959.
Woolgar, A., Thompson, R., Bor, D., & Duncan, J. (2011).
Multi-voxel coding of stimuli, rules, and responses in
human frontoparietal cortex. Neuroimage, 56, 744–752.
Yeung, N., Nystrom, L. E., Aronson, J. A., & Cohen, J. D. (2006).
Between-task competition and cognitive control in task
switching. Journal of Neuroscience, 26, 1429–1438.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
9
2
7
1
/
7
9
5
/
2
1
1
7
9
5
4
2
9
/
8
1
1
7
0
8
o
3
c
5
n
5
_
3
a
/
_
j
0
o
0
c
8
n
1
7
_
a
p
_
d
0
0
b
8
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
De Baene et al.
1765