The Timing and Neuroanatomy of Conscious Vision as
Revealed by TMS-induced Blindsight
Christopher P. GRAMO. allen, Petroc Sumner, and Christopher D. Chambers
Abstracto
■ Following damage to the primary visual cortex, some patients
exhibit “blindsight,” where they report a loss of awareness while
retaining the ability to discriminate visual stimuli above chance.
Transient disruption of occipital regions with TMS can produce
a similar dissociation, known as TMS-induced blindsight. El
neural basis of this residual vision is controversial, with some
studies attributing it to the retinotectal pathway via the superior
colliculus whereas others implicate spared projections that origi-
nate predominantly from the LGN. Here we contrasted these
accounts by combining TMS with visual stimuli that either acti-
vate or bypass the retinotectal and magnocellular (R/M) camino-
maneras. We found that the residual capacity of TMS-induced
blindsight occurs for stimuli that bypass the R/M pathways, en-
dicating that such pathways, which include those to the superior
colliculus, are not critical. We also found that the modulation of
conscious vision was time and pathway dependent. TMS applied
either early (0–40 msec) or late (280–320 msec) after stimulus
onset modulated detection of stimuli that did not bypass R/M
pathways, whereas during an intermediate period (90–130 msec)
the effect was pathway independent. Our findings thus suggest
a prominent role for the R/M pathways in supporting both the
preparatory and later stages of conscious vision. This may help
resolve apparent conflict in previous literature by demonstrating
that the roles of the retinotectal and geniculate pathways are
likely to be more nuanced than simply corresponding to the
unconscious/conscious dichotomy. ■
INTRODUCCIÓN
Blindsight (Weiskrantz, 1986) and TMS-induced blindsight
(p.ej., Boyer, harrison, & Ro, 2005; Jolij & Lamme, 2005) son
intriguing phenomena in which observers can successfully
identify visual stimuli for which they deny awareness. Este
dissociation between ability and awareness typically fol-
lows damage or disruption to the early visual cortex and
offers insights into the neural bases of consciousness and
subliminal perception (Dretske, 2000). Despite decades
of research, a consensus explanation of blindsight re-
mains elusive, and two central problems remain unsolved.
The first, long-standing question concerns which neu-
ral pathways are crucial for vision lacking in aware-
ness (Cowey, 2010). El segundo, more recent, pregunta
concerns the temporal dynamics of vision—whether un-
conscious and conscious processing are supported, respetar-
activamente, by feedforward and recurrent phases of processing
(Lamme, 2001).
The classic anatomical debate in blindsight research is
whether residual reportedly “unseen” vision depends on
the ancient retinotectal pathway via the superior colliculus
(CAROLINA DEL SUR) or whether it instead arises from other spared projec-
tions that are thought to predominantly involve the LGN.
Empirical support has emerged for both the retinotectal
(Leh, Ptito, Schonwiesner, Chakravarty, & Mullen, 2010;
Universidad de Cardiff
Tamietto et al., 2010; Leh, Johansen-Berg, & Ptito, 2006;
Leh, Mullen, & Ptito, 2006; Ro, Shelton, Sotavento, & Chang,
2004; Rafal, Herrero, Krantz, cohen, & Brennan, 1990)
and geniculate hypotheses (Railo, Salminen-Vaparanta,
Henriksson, Revonsuo, & Koivisto, 2012; Schmid et al.,
2010; Radoeva, Prasad, Brainard, & Aguirre, 2008; Wessinger,
Fendrich, & Gazzaniga, 1997). Although some authors have
offered a more nuanced view (p.ej., Cowey, 2010; Danckert
& Rossetti, 2005), many have argued for one standpoint
over the other, with the majority favoring the retinotectal
cuenta.
Two previous TMS studies probed the retinotectal ver-
sus geniculate basis of blindsight but arrived at opposite
conclusions (Boyer et al., 2005; Ro et al., 2004). The first
of these studies (Ro et al., 2004) showed that reportedly
“unseen” distractors slowed saccadic responses, pero no
manual button presses. Because the retinotectal pathway
is thought to drive saccades, the authors concluded that
the preserved capacity of TMS-induced blindsight neces-
sarily originated from retinotectal information. Sin embargo,
the absence of a manual distractor effect was not definitive,
and subsequent evidence demonstrates that saccadic
distractor effects can also be generated via geniculate
pathways (Bompas & Sumner, 2009; Sumner, Nachev,
Castor-Perry, Isenman, & Kennard, 2006). In contrast to
Ro et al. (2004), Boyer et al. (2005) used chromatic stim-
uli that did not activate the retinotectal pathway and
demonstrated preserved capacity under TMS, sugerencia
© 2014 Massachusetts Institute of Technology Published under a
Creative Commons Attribution 3.0 no portado (CC POR 3.0) licencia
Revista de neurociencia cognitiva 26:7, páginas. 1507–1518
doi:10.1162/jocn_a_00557
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
oh
1
C
3
norte
3
_
9
a
/
_
j
0
oh
0
C
5
norte
5
7
_
a
pag
_
d
0
0
b
5
y
5
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
7
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
t
/
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
a geniculate role. Sin embargo, they also showed that the
confidence participants placed in discrimination judgments
correlated with the capacity they claimed to represent
TMS-induced blindsight, and during the collection of these
data some of their participants acknowledged partial
awareness of the stimuli. Partial conscious perception, como
opposed to blindsight, may therefore be a more plausible
interpretation of their results. Clear interpretations of both
studies are further hampered by a lack of control for non-
specific effects of TMS (p.ej., sham TMS condition or a con-
trol site) and the absence of robust evidence that TMS was
effective in suppressing awareness during the conditions
designed to demonstrate blindsight. Together with the
divergence in conclusions, these limitations leave open
the question as to the anatomical basis of blindsight.
Here we directly contrasted the retinotectal and genic-
ulate accounts in TMS-induced blindsight by comparing
responses to visual stimuli that activate or bypass the
retinotectal pathway (Sumner, Adamjee, & Mollon, 2002).
The retinotectal pathway has previously been demon-
strated not to receive color opponent input from short-
wave cones (s-cones) of the retina (Sumner et al., 2002;
de Monasterio, 1978; Schiller & Malpeli, 1977). This means
that s-cone-specific stimuli, when accompanied by lumi-
nance noise, will be invisible to the retinotectal route
(Sumner et al., 2002). Por lo tanto, if the residual capacity
of blindsight requires retinotectal processing, it should
not be demonstrable with “s-cone” stimuli. En tono rimbombante,
our question addresses whether critical information is
carried in the fast and direct pathway to the SC, bastante
than concerning all processing within the SC. Información
can reach the SC via the visual cortex as well as via the
direct retinotectal pathway. Respectivamente, chromatic infor-
mation can activate the SC when it is the target for an
eye movement, and does so with a delay consistent with
the cortical route (Blanco, Boehnke, Marino, Itti, & Munoz,
2009). The geniculate account of blindsight does not,
por lo tanto, require that there is no SC processing, pero
simply that the retinotectal pathway is not the critical
conduit for blindsight.
Además, it is noteworthy that magnocellular and
possibly parvocellular projections of the LGN are also blind
to s-cone stimuli (Dacey, 2000; Mollon, 1989). De este modo, si
TMS-induced blindsight were eliminated for s-cone stim-
uli, then this finding would be consistent with retinotectal
mediation but also consistent with spared magnocellular
fibers. Por otro lado, if TMS-induced blindsight were
the same for s-cone stimuli compared with luminance
estímulos, then this would rule out a critical role for the
retinotectal pathway. It would not rule out a retinotectal
contribution for other types of stimulus that we do not test
aquí, but it would be sufficient to show that retinotectal
mediation is not always required.
The anatomical basis of blindsight may also depend
critically on temporal dynamics. According to a popular
cuenta (Lamme, 2001; Lamme & Roelfsema, 2000), visual
stimuli trigger an initial feedforward volley, likely includ-
ing retinotectal signals, that is sufficient for some degree
of unconscious processing; this is then followed (de
∼100 msec poststimulus) by recurrent feedback that facili-
tates awareness. By manipulating both the stimulus color
(s-cone vs. luminance) and timing of TMS, we sought to
test how the functional anatomy of blindsight interacts
with these putative phases of visual processing.
Both the retinotectal and geniculate hypotheses predict
that later occipital TMS (≥100 msec) should disrupt recur-
rent processing, suppressing conscious awareness while
leaving reportedly “unseen” abilities for luminance stim-
uli above chance (and thus demonstrating TMS-induced
blindsight; ver Figura 1B). According to the retinotectal
hypothesis, “unseen” abilities under these conditions
should be relatively suppressed for s-cone stimuli because
the retinotectal route is blind to these stimuli. A diferencia de,
if the geniculate hypothesis is correct, then blocking the
retinotectal pathway should be inconsequential and blind-
sight should be observed for s-cone stimuli. Because an
equivalence of TMS-induced blindsight for s-cone and
luminance stimuli would be theoretically informative, nosotros
adopted Bayesian statistical methods that assess the like-
lihood of the null hypothesis (Dienes, 2008b, 2011) en
addition to more common Neyman–Pearson analyses.
MÉTODOS
Participantes
Sixteen neurologically healthy participants with normal
color vision were recruited (seven men, edad media =
24.0 años, DE = 3.04). All procedures were approved
by the research ethics committee at the School of Psy-
chology, Universidad de Cardiff. Initially, a further 10 participar-
pants were enlisted but did not participate in the main
experiment because occipital TMS did not elicit phos-
phenes within the safety limits set by ethics approval
(seven participants) or because TMS applied at the required
intensity caused large contractions of peripheral facial
nerves that could cause an experimental confound (two
Participantes). One participant withdrew from the study
following a mild adverse reaction to the TMS (Maizey
et al., 2013).
Tarea
In our paradigm, active or sham (control) TMS was applied
in 25-Hz pulse pairs over the occipital cortex. To modu-
late the earliest phases of visual processing, TMS was
applied at 0–40 msec and 40–80 msec after stimulus onset
(Corthout, Hallett, & Cowey, 2002). Recurrent process-
ing was targeted by applying TMS at 90–130 msec (p.ej.,
Amassian et al., 1989) and during a later period at 280–
320 mseg (p.ej., Chambers, allen, Maizey, & williams,
2012). These four periods were tested to capture possible
independent early and late phases (Koivisto, Mantyla, &
Silvanto, 2010). TMS applied at ∼100 msec poststimulus
1508
Revista de neurociencia cognitiva
Volumen 26, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
oh
1
C
3
norte
3
_
9
a
/
_
j
0
oh
0
C
5
norte
5
7
_
a
pag
_
d
0
0
b
5
y
5
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
7
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
t
/
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
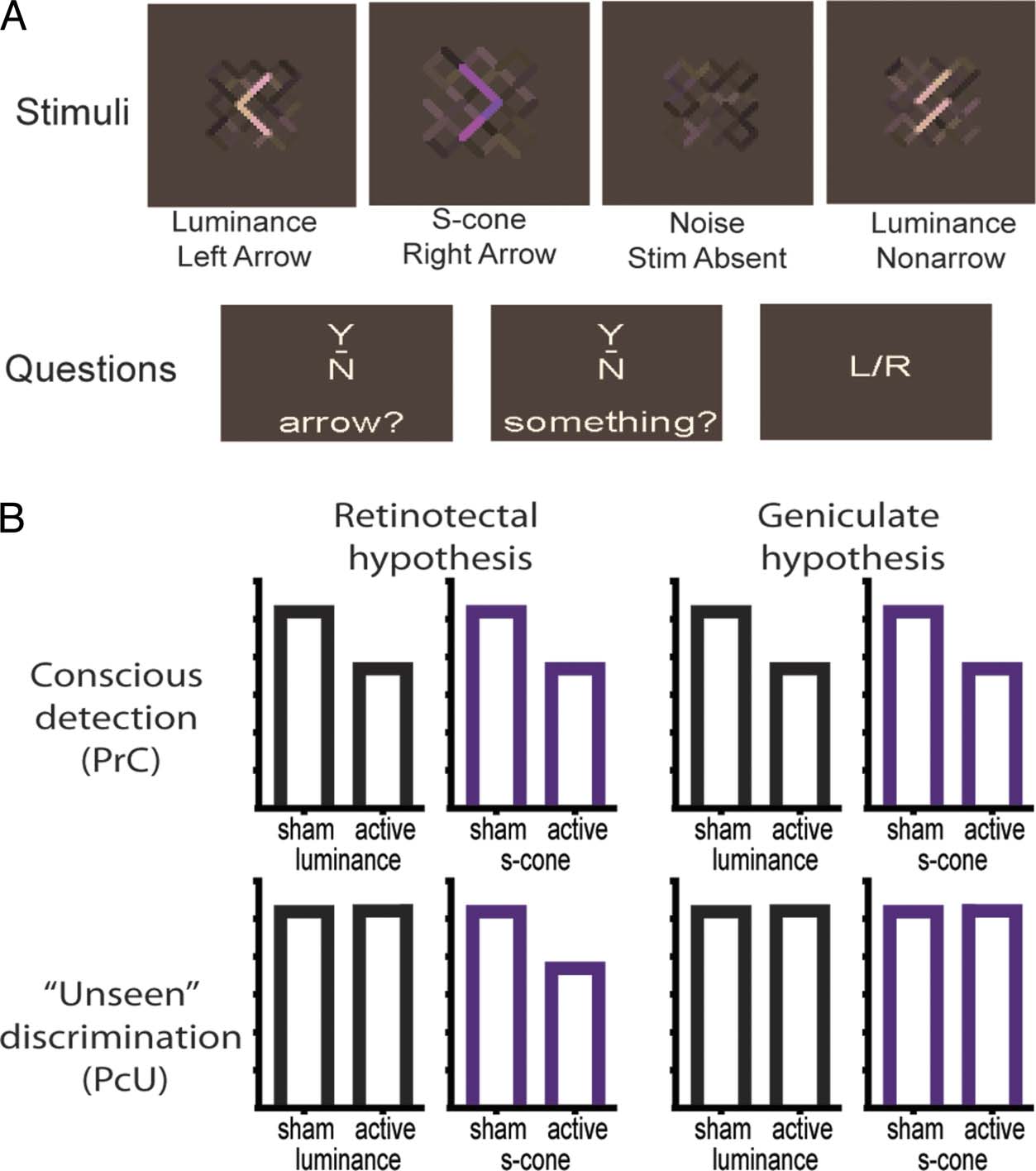
Cifra 1. (A) Examples of
stimuli and questions. El
measure of conscious detection
was derived from application
of signal detection theory
(Corwin, 1994) in response
to the questions: “Did you
consciously see the arrow?"
(Arrow Y/N) and “Did you see
something that might have
been a stimulus?" (Something
Y/N). The measure of “unseen”
discrimination was the
proportion of correct
discriminations of arrow
direction (L/R) on trials in
which participants reported
“No” to both the “arrow?"
and “something?” questions.
The nonarrow condition
ensured that participants
perceived an arrow when
reporting one, Opuesto a
simply responding to a change
in luminance or s-cone
saturation. (B) Illustration
of predictions. TMS-induced
blindsight as defined by
a statistically significant
suppression in the measure
of conscious detection (PrC)
for both stimulus types, en
conjunction with above-chance
“unseen” discrimination (PcU)
for luminance stimuli. Under
the retinotectal hypothesis,
“unseen” discrimination for
s-cone stimuli may not be demonstrated as being greater than chance. Además, according to the retinotectal account, concurrent “unseen”
discrimination in the presence of s-cone stimuli should be suppressed relative to sham, and any change in capacity may differ from that
observed in the presence of luminance stimuli. According to the geniculate hypothesis, concurrent “unseen” discrimination ability for s-cone
stimuli should be relatively preserved and comparable to that observed in the presence of luminance stimuli.
has formed the basis for many of the previous demonstra-
tions of TMS-induced blindsight (Christensen, Kristiansen,
Rowe, & Nielsen, 2008; Boyer et al., 2005; Jolij & Lamme,
2005; Ro et al., 2004).
The target stimulus was an arrow embedded in lumi-
nance noise, presented on 50% of all trials (Figura 1A).
Trials could include an arrow target, a nonarrow target,
or no target. On each trial, participants were asked if they
were aware of the arrow stimulus and in which direction
it was pointing (left or right). Además, we asked par-
ticipants whether they were aware of “something” having
been presented (including the nonarrow target). Este
question probed a lower level of awareness than the
“arrow?” question (Overgaard, Rote, Mouridsen, & Ramsoy,
2006). Using negative responses to both questions to in-
dicate lack of awareness is more stringent than merely
using the “arrow?” question alone. Forced-choice discrimi-
nation judgments (left/right) on these “double negative”
awareness trials constituted the “unseen” measure (PcU).
This discrimination capacity therefore represents a rela-
tively conservative form of unconscious processing (ver
Dienes, 2008a). The conscious awareness measure was
participantsʼ sensitivity in their reported awareness of stim-
uli (PrC). In this design, blindsight would thus manifest
as a TMS-induced impairment of conscious detection (PrC)
while leaving the ability to discriminate reportedly “unseen”
arrows (PcU) above chance and relatively unaffected by
TMS.
Two classes of target stimuli were employed (Figura 1A):
luminance stimuli, which were defined by an increase in
luminance, and s-cone stimuli, which were defined by an
increase in color saturation that stimulated s-cones, pero
no (or minimally) long and medium wave cones (Sumner,
2006; Sumner, Nachev, Vora, Husain, & Kennard, 2004;
Sumner et al., 2002). In addition to retinotectal path-
maneras, magnocellular layers of the LGN—and possibly
parvocellular layers (Dacey, 2000; Mollon, 1989)—are also
blind to s-cone stimuli embedded in luminance noise
(Derrington, Krauskopf, & Lennie, 1984); de este modo, estos
retinotectal and magnocellular pathways bypassed by
s-cone stimuli are collectively denoted hereafter as “R/M.”
Because of the isolation of magnocellular pathways, cualquier
allen, Sumner, and Chambers
1509
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
oh
1
C
3
norte
3
_
9
a
/
_
j
0
oh
0
C
5
norte
5
7
_
a
pag
_
d
0
0
b
5
y
5
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
7
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
.
F
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
s-cone-dependent effect cannot be attributed exclusively
to the retinotectal pathway. Sin embargo, if a capacity were
to depend upon retinotectal input, then a deficit would
be expected when s-cone stimuli are used. To obscure
the s-cone stimuli from R/M channels (Bompas & Sumner,
2009; Sumner et al., 2006), all stimuli were presented
against a background of luminance noise (mean luminance
of noise and background: 25 cd/m2; range of noise: 10 cd/
m2). To our knowledge, this range is larger and therefore
has a greater capacity to obscure a luminance transition
than any previous comparable study. The noise occupied
a 1.91° × 1.91° area at fixation, whereas the arrows and
nonarrows subtended 0.81° × 1.43°. To aid comparability
of stimulus classes, the noise also contained low levels of
color noise along the tritan axis (range: ±8% of mean
s-cone saturation of stimuli). Arrow stimuli were present
en 50% of all trials, whereas nonarrows and stimulus-
absent trials comprised 25% of trials each.
Note that here we use “s-cone stimuli” to denote the
transition between gray and purple in the context of lumi-
nance noise, rather than the presence of the purple arrow
per se. This is because the retinotectal pathway is thought
to be blind to such a transition: Even if a small s-cone sig-
nal were to enter the pathway, there are no chromatically
opponent cells in this pathway to distinguish a chromatic
change from a luminance change (Smithson, Sumner, &
Mollon, 2003; Sumner et al., 2002; Marrocco & li, 1977;
Schiller & Malpeli, 1977). Any chromatic information that
does reach the SC is likely to be indirect, initially entering,
and therefore dependent upon cortical areas (Blanco y col.,
2009). Consistente con esto, Sumner et al. (2006) presentado
that s-cone stimuli (calibrated and embedded in luminance
noise using the same methods we employ here) affect eye
movements in a pattern distinct from any strength of
luminance stimulus (es decir., the effect of s-cone stimuli could
not be accounted for by weak retinotectal signals). Ad-
ditionally, White et al. (2009) showed that, although chro-
matic information can activate the SC when it is the target
for an eye movement, it does so with a delay consistent
with an indirect cortical route. Por lo tanto, as noted in the
Introducción, our question addresses whether critical
information is carried in the retinotectal pathway to the
CAROLINA DEL SUR, rather than assessing SC processing per se. The genicu-
late account of blindsight does not require an absence of
SC processing, but rather that the retinotectal pathway is
not critical for the residual capacity.
Trials commenced with a fixation cross (1.5 segundo) fol-
lowed by luminance noise, which was produced by alter-
nating the luminance of the bars at 50 Hz. The noise
continued for 800 msec before onset of the target stim-
ulus (20 mseg; or noise of equal duration on stimulus-
absent trials). Following target offset, the noise continued
for a further 380 mseg. The three questions were then
presentado, and responses were collected on a standard
computer keyboard.
The pattern of behavior that characterizes blindsight
could potentially be attributed to changes in response
inclinación (Campion, Latto, & Herrero, 1983; Nisbett & wilson,
1977). To address these concerns, we applied signal de-
tection theory (SDT) to responses to the “something?"
and “arrow?” questions. This provided a measure of
conscious detection (PrC) that is largely independent of
response criteria and corresponds to the rate at which
participants report awareness of the arrow when it is pre-
sent (hit) minus the rate at which they report awareness
when no arrow was presented (false alarm; Corwin, 1994;
MacMillan & Creelman, 1990). This measure was pre-
dominantly driven by responses to the arrow question,
in line with task instructions and discursive reports made
by the participants about their experience of the task
(Gallagher, 2003; Varela, 1996). Por ejemplo, responding
“No” to “arrow?” and “Yes” to “something?” in the pre-
sence of a nonarrow was classified as a “correct rejection”
of a potential arrow target as opposed to being a “hit” for
a nonarrow target. This categorization of responses into
SDT classes is summarized in Table 1 and was established
before data analysis. Nonparametric versions of SDT
were used because the imbalance in stimulus-absent con-
ditions violated the assumptions of classic SDT (Pr; ver
Corwin, 1994). “Unseen” discrimination performance
was defined as the proportion correct when participants
responded negatively to both “arrow?” and “something?"
preguntas (PcU).
Procedimiento
Following calibration, participants completed six experi-
mental sessions of ∼1 hr. Each session consisted of four
bloques, one for each of the experimental conditions: s-cone
stimuli or luminance stimuli combined with sham or active
TMS, with the order randomized between sessions. Blocks
consistió en 80 ensayos, incluido, in a randomized order,
the four stimulus types (left arrow, right arrow, nonarrow,
Mesa 1. SDT Classes for Measures of Conscious
Awareness (PrC)
Response
Stimulus
Something?
Flecha?
SDT Class
Flecha
Nonarrow
Nothing
Sí
No
Sí
Sí
Sí
Sí
No
Sí
No
Sí
No
Sí
No
No
HIT
MISS
FA
CR
FA
FA
CR
FA = false alarms; CR = correct rejections. Hit Rate = Hits/(Hits +
Miss); False Alarm Rate = FA/(FA + CR); Pr = Hit Rate − False Alarm
Rate (Corwin, 1994).
1510
Revista de neurociencia cognitiva
Volumen 26, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
oh
1
C
3
norte
3
_
9
a
/
_
j
0
oh
0
C
5
norte
5
7
_
a
pag
_
d
0
0
b
5
y
5
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
7
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
F
t
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
and stimulus-absent) and four TMS onset times (0–40,
40–80, 90–130, and 280–320 msec).
The order of the response questions (Flecha? Alguno-
thing? Left/Right?) was different for each session, de modo que
across the experiment each participant completed a fully
counterbalanced set of question orders. Los participantes fueron
given 10 practice trials at the start of each session to
become familiar with the order. The sequence in which
these question orders were presented was randomized
between participants.
Task Calibration
Calibration sessions lasted 2–4 hr and took place at least
24 hr before experimental sessions. During this session,
stimulus levels were calibrated to ensure that all partici-
pants performed at comparable levels of detection for
both luminance and s-cone stimuli and participants were
screened for suitability for TMS. Para cada participante
we also established the TMS phosphene threshold (PT;
Franca, Koch, Mochizuki, Huang, & Rothwell, 2006) y
tritan line in color space for calibration of s-cone stimuli
(Smithson et al., 2003).
Detection thresholds for stimuli were calculated using a
psychophysical method of constants. Following a period
of familiarization with the task, participants completed
aproximadamente 10 blocks over a range of stimulus inten-
ciudades, separately for s-cone and luminance stimuli. Para
luminance stimuli, the threshold was obtained by adjust-
ing stimulus intensity, whereas for s-cone stimuli, el
color saturation was varied along the s-cone axis/tritan
line. Data were then regressed using a sigmoidal or
linear function (depending on fit quality), solving for the
intensidad, which produced a PrC of 0.5; this corresponded
to the stimulus level at which participants reported con-
sciously seeing the stimulus on 50% of target-present trials
if no false alarms were reported. Following the initial cali-
bration, participants then completed several blocks at the
derived intensity, and minor adjustments were made to
the stimulus levels to compensate for any learning effects,
thus maintaining PrC ≅ 0.5. At the start of each experi-
mental session, participants completed at least one pre-
block with sham TMS (40 ensayos), and similar adjustments
were made as required (criteria > ±0.1 PrC from 0.5). Si
participants completed two sessions in 1 día, then the
mean value for the previous sham block was taken as
the starting point for subsequent recalibration. The mean
luminance for luminance stimuli was 36.5 cdm2 (DE =
3.5). Participant-specific s-cone stimuli were produced fol-
lowing the methods described in Smithson et al. (2003)
and used in several previous studies (Bompas & Sumner,
2008, 2009, 2011; anderson, Husain, & Sumner, 2008;
Bompas, Sterling, Rafal, & Sumner, 2008; Sumner, 2006;
Sumner et al., 2002, 2004). This procedure yielded indi-
vidual s-cone colors with the following coordinates in CIE
1931 espacio: significar (±SD): 0.249 (0.009), 0.186 (0.022),
25.067 (0.809).
The method used to determine PT closely resembled
that of Franca et al. (2006). Primero, we assessed each partici-
pantʼs susceptibility to phosphenes within safety limits
set by our ethics committee and international guidelines
for 25-Hz stimulation (160% of motor threshold). El
coil was oriented with the handle pointing upward and
side “B” facing the participant, so that the induced cur-
rent passed initially in a left-to-right direction. Coil posi-
tioning was initially based on anatomical proximity to
the midhemispheric convergence of the calcarine sulci,
localized in individual structural MRI scans. Immediately
before each active TMS block, the intensity was set to
120% of PT (or ∼130% of motor threshold if a PT had
not yet been established) and the coil was moved so that
it produced a phosphene that the participant reported
as being “reasonably clear” and “at least in part, covering
the center of their visual field,” with their eyes closed.
This procedure usually required ∼10 pulses. The coil
position was then recorded using a Brainsight system
(Rogue Research, Cª, Montréal, Canada) and used for
the subsequent block of trials. If the participant moved
beyond a 5-mm tolerance of the original position, entonces
the block was paused and the coil repositioned to the re-
corded site. Single pulse TMS was applied approximately
cada 5 segundo. An approximate PT was obtained using an
up–down staircase method, starting at 50% of maximum
output and adjusting TMS intensity in reducing steps of
5%, 2%, y 1%, so that participants verbally reported see-
ing five phosphenes from 10 pulses. This level was then
used as the basis of a more accurate threshold where the
number of reported phosphenes arising from 10 pulses
was recorded at −10%, −5%, 0%, +5%, +10%, y +15%
of the approximate PT. The orders of these sets of 10 pulses
were randomized, and the full range of intensities was
repeated three times in separate blocks with breaks in
entre. The coil was repositioned at the start of each
block. Averaging across blocks yielded a function rep-
resenting the number of phosphenes out of 10 over a
range of intensities, to which a curve was fitted (sigmoid
or linear depending on goodness of fit). Solving this curve
para 5/10 phosphenes thus provided the PT used in the sub-
sequent experiments.
Equipment
Cortical stimulation was delivered with a Magstim Super
Rapid biphasic stimulator in conjunction with a Magstim
high-power 90 mm round coil. Round coils have pre-
viously been shown to effectively produce TMS-induced
blindsight (p.ej., Christensen et al., 2008; Boyer et al.,
2005; Jolij & Lamme, 2005) and are more likely to affect
both hemispheres than are figure-eight coils. TMS delivery
was controlled with a Cambridge Research Systems Visage
running Real-time Sequencer software on a Matlab plat-
forma, which also governed stimulus presentation on a
gamma-corrected 21-in. Mitsubishi CRT monitor (100 Hz
vertical refresh rate). Pairs of TMS pulses were applied at
allen, Sumner, and Chambers
1511
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
oh
1
C
3
norte
3
_
9
a
/
_
j
0
oh
0
C
5
norte
5
7
_
a
pag
_
d
0
0
b
5
y
5
gramo
7
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
7
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
F
/
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
95% of PT (group mean = 54.4% of maximum stimulator
producción, DE = 11.5%). This protocol was adopted because
higher TMS intensities in pilot experiments caused blinks,
twitches, and phosphenes that the participants reported
as visible during the task. In pilot experiments, más bajo
intensities and single pulses of TMS were found not to
produce reliable effects on performance. The coil was
positioned using the same procedure as described for
the calibration procedure. An approximation of this posi-
tion was used in the sham condition but with the coil
perpendicular to the scalp so that the rim pointed toward
the head, with a 9-mm plastic spacer inserted between the
scalp and coil to replicate the contact artifact.
To exclude effects of TMS-induced blinks on perfor-
mance, eye tracking was undertaken throughout the ex-
periment using a Cambridge Research Systems chin-rest
mounted infrared eye tracker (250 Hz). Trials were ex-
cluded on the basis of blinks identified by a shift in the
trace of pupil position followed by a transitory loss of pupil
señal, coincident with the stimulus presentation. En general,
241 of a total 30,720 trials were excluded from the analysis
(<0.8%).
Statistical Analyses
The temporal and anatomical dynamics of effects were
assessed using a combination of Neyman–Pearson signif-
icance tests and complementary Bayesian analyses, which
can directly estimate the likelihood of the null hypothesis
(Gallistel, 2009; Rouder, Speckman, Sun, Morey, & Iverson,
2009; Dienes, 2008b). We adopted this combined ap-
proach because our hypotheses (Figure 1B) required test-
ing for a specific pattern of results that included both
variance and invariance between experimental conditions.
TMS-induced blindsight is defined here as a significant
suppression (i.e., for active TMS vs. sham) in conscious
detection, whereas concurrent reportedly “unseen” discrim-
ination remains above chance and ideally unperturbed by
the TMS. We expected to find this pattern for luminance
stimuli at least for the mid timed intervention at 90–
130 msec (Christensen et al., 2008; Boyer et al., 2005; Jolij
& Lamme, 2005; Ro et al., 2004). Only under such con-
ditions, where conscious detection is suppressed for both
stimuli types, can the retinotectal and geniculate hypoth-
esis be tested by assessing whether blindsight occurs in
the presence of s-cone stimuli—that is, when direct input
via the SC is withdrawn. If “unseen” perception is not pre-
served, then the retinotectal hypothesis is upheld, but if
“unseen” capacity is demonstrated, then the geniculate
hypothesis is supported. Note that the geniculate hypoth-
esis does not require the extent to which “unseen” perfor-
mance is above chance for s-cone stimuli to be the same
as for luminance stimuli, because there may be differences
in the way the different geniculate pathways contribute
to perception (Merigan & Maunsell, 1993). However, the
conclusions would be strengthened if the two stimulus
types produce equivalent effects. Additionally, pilot ex-
periments demonstrated that participants were able to
discriminate “unseen” s-cone stimuli above chance levels
when no TMS was applied. This is a prerequisite of the
current paradigm because the lack of such capacity in the
baseline condition would make it impossible to ask which
pathways support residual capacity when early visual areas
are disrupted with TMS.
Therefore, following the observation of TMS-induced
blindsight with luminance stimuli and the observation
that conscious detection of s-cone stimuli is suppressed
by TMS, the critical question is whether or not “unseen”
discrimination performance is maintained above chance
in the presence of s-cone stimuli. Second, any potential
disruption of “unseen” discrimination of s-cone arrows
was assessed by comparing active to sham “unseen” per-
formance and by comparing any change in “unseen” per-
formance between the stimuli types.
Effects were assessed using repeated-measure ANOVAs
with TMS site (two levels: active and sham), stimulus type
(two levels: luminance and s-cone), and time of TMS rela-
tive to stimulus onset (four levels: 0–40, 40–80, 90–130,
and 280–320 msec) as factors, conducted separately on
measures of conscious detection (PrC; hit rate − false
alarm rate) and “unseen” discrimination (PcU; proportion
correct when negative responses to both “arrow?” and
“something?” questions were given). Analyses of simple
main effects exploited the relevant Greenhouse–Geisser
corrected error terms from the ANOVAs (Winer, Brown,
& Michels, 1991) and the sham condition as a baseline.
“Unseen” discrimination performance was assessed rela-
tive to chance using single-sample t tests, which are re-
ported in the text. All t tests and analyses of simple main
effects applied the Holm–Bonferroni method of correction
for multiple comparisons (Holm, 1979).
The positive comparisons described above only assess
the likelihood of differences between conditions. By using
a Bayesian approach, we can additionally assess the oppos-
ing hypothesis that two conditions are equivalent (Dienes,
2008b). This approach involves deriving a Bayes factor (B)
which represents the strength of support for the alterna-
tive hypothesis (H1) relative to the null (Dienes, 2008b),
with B > 1 indicating evidence in favor of H1 and B <
1 indicating evidence in favor of the null. In practice,
values of B greater than 3 or smaller than 1/3 are regarded
as providing substantial evidence in favor of H1 or the null
hypothesis, respectively (Dienes, 2011; Jeffreys, 1961).
Bayesian analyses, complementary to the Neyman–
Pearson statistics, were based upon prior models rep-
resenting the critical hypotheses. Bayesian tests were
conducted separately on both positive and negative
changes from the sham baseline with the hypotheses
represented by a uniform distribution (Dienes, 2008b).
Because the critical measures were calculated relative
to a baseline, zero was selected as the starting point for
the distributions. The maximum reasonable shift on both
measures was 0.5 and was therefore used as the upper
limit for the theoretical distributions. For PcU, these
1512
Journal of Cognitive Neuroscience
Volume 26, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
o
1
c
3
n
3
_
9
a
/
_
j
0
o
0
c
5
n
5
7
_
a
p
_
d
0
0
b
5
y
5
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
values ranged from 1 to 0.5 (chance performance) and
for PrC from the calibrated level of 0.5 to chance at 0.
Because it is conceivably possible that performance could
fluctuate beyond this range (e.g., if false alarm rates were
found to be higher than hit rates or if discrimination
performance fell well below chance), the adoption of
the 0.5 limit does not unfairly favor evidence for the null
(see Dienes, 2008b). Sham baselined data (active − sham)
is denoted as Δ sham.
To assess TMS-dependent effects, we analyzed the
change in measures from sham at each level of TMS
onset time and stimulus condition. The vector for com-
parison between stimuli types, as affected by the TMS,
was calculated as (Luminanceactive- sham) − (s-coneactive-
sham). The hypothesis for this analysis was represented
by a uniform distribution between 0 and 0.5, correspond-
ing to a benefit bestowed by the additional input of lumi-
nance stimuli. In addition, for these comparisons, the
complementary t tests were reported, as were the B statis-
tics representing effects in the opposite direction. Because
each comparison involved 16 independent data points
(participants), appropriate standard error adjustments
were applied, as recommended by Dienes (2008b).
Tests for outlier rejection were applied at a participant
level using Chauvenetʼs criterion (Taylor, 1997). No partici-
pants were excluded.
RESULTS
The two principal measures were conscious detection and
“unseen” discrimination ability. As an overview, conscious
detection was suppressed by TMS during the middle and
later periods, whereas “unseen” performance remained
above chance and statistically unaffected. This pattern in-
dicates characteristic TMS-induced blindsight. Our key
result was that s-cone stimuli produced blindsight during
the principal epoch (∼100 msec, Figure 2C and D), indi-
cating that it does not rely on the retinotectal pathway.
Moreover, the pattern for s-cone stimuli in this epoch
was indistinguishable from that for luminance stimuli.
Beyond this, we also found that TMS facilitated rather
than impaired conscious detection at the earliest time
point (0–40 msec) for luminance stimuli only (Figure 2A
and B). Finally, TMS impaired conscious detection at the
latest time point (280–320 msec), again for luminance
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
o
1
c
3
n
3
_
9
a
/
_
j
0
o
0
c
5
n
5
7
_
a
p
_
d
0
0
b
5
y
5
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
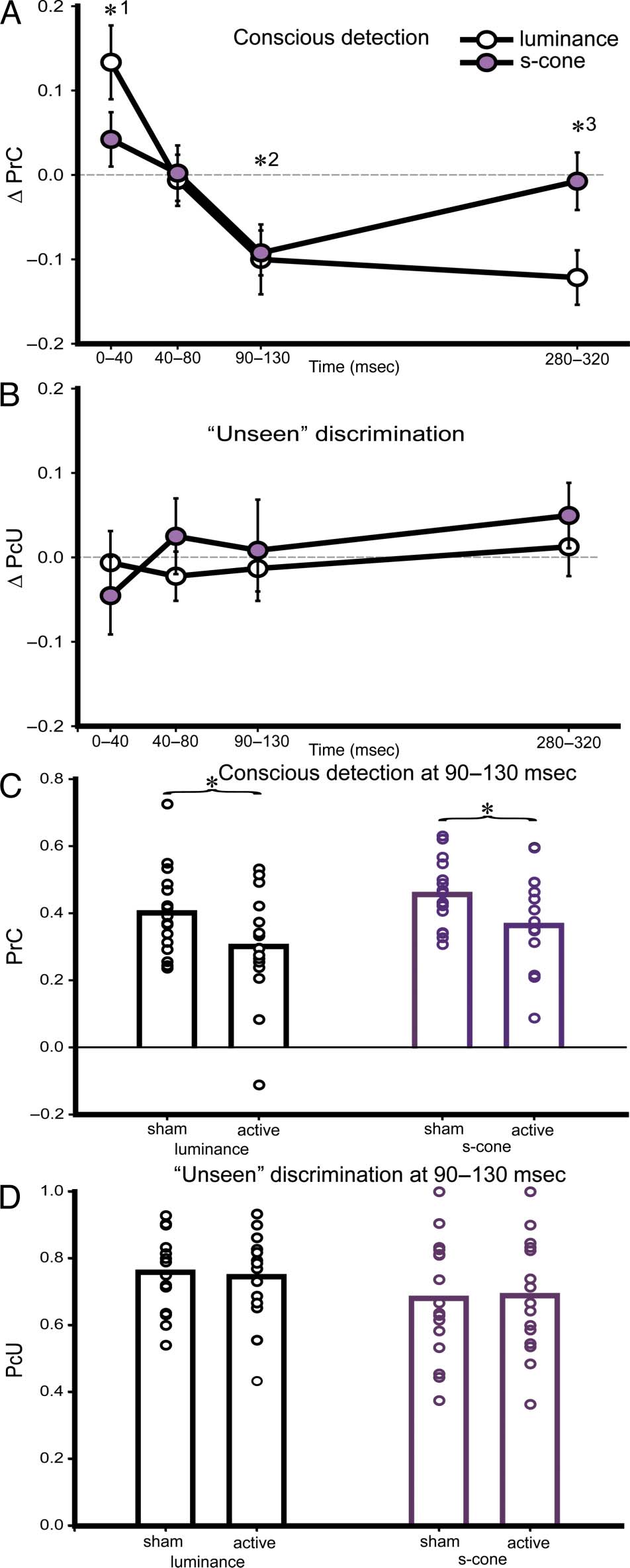
Figure 2. Sham-normalized effects of occipital TMS on (A) conscious
detection (PrC) and (B) “unseen” discrimination (Proportion correct
“Unseen,” PcU), plotted according to stimulus type (s-cone or luminance)
and TMS onset time. PrC is calculated as a nonparametric equivalent
of d0 (Corwin, 1994). Error bars are the within-subject SEM (Loftus
& Masson, 1994). An impairment in conscious detection for both
stimulus types, combined with preserved “unseen” abilities (TMS-induced
blindsight) was observed only at the 90–130 msec epoch (*2).
Occipital TMS selectively enhanced (0–40 msec, *1) or impaired PrC
(280–320 msec, *3) for luminance stimuli relative to s-cone stimuli.
No significant effect of TMS was observed for PrC at 40–80 msec or
for PcU at any TMS onset times. C and D illustrate raw data during
TMS-induced blindsight at the 90–130 msec period, where the measure
of conscious detection is suppressed (C) in the active TMS condition
relative to sham for both stimulus types. D depicts the preservation
of “unseen” discrimination ability in this condition, again for both
stimulus types. Symbols denote individual data points. *p < .05.
Allen, Sumner, and Chambers
1513
stimuli only. Data associated with this article can be
downloaded from dx.doi.org/10.6084/m9.figshare.802837.
The modulation of awareness by TMS was thus both
time- and stimulus-dependent: A significant three-way
interaction, F(3, 45) = 3.3, p = .031 (see Figure 2A), was
observed between TMS Condition (occipital, sham) ×
TMS Onset Time (0–40, 40–80, 90–130, 280–320) and Stim-
ulus Type (luminance, s-cone). Meanwhile, discrimination
performance on “unseen” trials was consistently above
chance at all 16 combinations of TMS condition, TMS onset
time, and stimulus type (mean proportion correct = 0.71,
SD = 0.15, all t(15) > 3.14, pag < .007 with Holm–Bonferroni
correction). A three-way ANOVA of “unseen” discrimination
revealed no significant interaction of TMS Condition × TMS
Onset Time × Stimulus Type, F(3, 45) = 0.36, p = .75
(Figure 2B), and no significant main effects or lower-order
interactions (all F < 2.37, all p > .142). Given the signifi-
cant three-way interaction for awareness, we now consider
these data in relation to the anatomical and temporal
hypotheses by detailing the effects of occipital stimulation
during each TMS epoch.
0–40 msec
The application of early TMS did not disrupt conscious
detection or “unseen” discrimination. Bastante, occipital
stimulation significantly facilitated conscious detection
relative to sham in the presence of luminance stimuli only
(Figura 2A *1, PrC luminance, active vs. sham t(15) = 5.59,
pag < .001, B(active > sham) = 8.97, B(active < sham) = 0.03; PrC
s-cone, active vs. sham t(15) = 1.76, p = .10, B(active > sham) =
0.31, B(active < sham) = 0.04; PrC Δ sham luminance vs.
s-cone t(15) = 2.70, p = .02, B(Δ sham, luminance > s-cone) =
1.20, B(Δ sham, luminance < s-cone) = 0.05). A TMS-induced
early suppression of “unseen” abilities would have sup-
ported a link between the feedforward sweep of activity
and unconscious processing. Additionally any stimulus
specificity of such an effect would implicate the role of the
retinotectal pathway in such processing (Lamme, 2001). In
contrast, “unseen” performance during this intervention
was above chance (for luminance stimuli t(15) = 6.25, p <
.001, for s-cone stimuli t(15) = 3.55, p = .003) and was
unaffected by the TMS (see Figure 2B, PcU luminance, active
vs. sham t(15) = 0.16, p = .88, B(luminance, active > sham) =
0.10, B(luminance, active < sham) = 0.12. PcU s-cone, active vs.
sham t(15) = 0.92, p = .37, B(s-cone, active > sham) = 0.08,
B(s-cone, active < sham) = 0.31). Moreover, no discernible
difference in “unseen” discrimination was observed
between stimuli types (PcU Δ sham luminance vs. s-cone
t(15) = 0.55, p = .59, B(Δ sham, luminance > s-cone) = 0.30,
B(Δ sham, luminance < s-cone) = 0.13). Therefore, these results
do not directly support a correspondence between early
occipital processing and unconscious vision. The disso-
ciation between stimulus types for conscious detection,
however, does provide confirmation that the s-cone stim-
uli were successfully excluded from luminance pathways.
That this dissociation occurred at the earliest time point is
consistent with s-cone stimuli being transmitted by a
slower pathway, as we would expect (e.g., Bompas &
Sumner, 2008, 2009; Anderson et al., 2008; Sumner et al.,
2002, 2004, 2006).
40–80 msec
No significant effects were observed as a result of apply-
ing TMS at 40–80 msec. Neither conscious detection
nor “unseen” discrimination showed any difference from
sham, and both stimulus types were equally unaffected
by the TMS (see Figure 2A and B, all t < 0.99, all p >
.34, all B < 0.32). “Unseen” discrimination was above
chance under active TMS (for luminance stimuli t(15) =
5.62, p < .001, for s-cone stimuli t(15) = 4.84, p < .001).
90–130 msec
TMS applied at 90–130 msec produced a blindsight effect
for both stimulus types and hence adjudicates between
the retinotectal versus geniculate accounts. A reliable sup-
pression of conscious detection was observed relative to
sham for both s-cone and luminance stimuli (see Figure 2A
*2 and C, PrC luminance, active vs. sham t(15) = 4.19,
p < .001, PrC s-cone; active vs. sham t(15) = 3.87, p =
.002, B(luminance, active < sham) = 1.58, B(s-cone, active < sham) =
59.98). This effect did not significantly dissociate be-
tween stimulus types (PrC Δ sham luminance vs. s-cone
t(15) = 0.23, p = .82, B(Δ sham, luminance > s-cone) = 0.15,
B(Δ sham, luminance < s-cone) = 0.18).
The key question in this study is whether above-chance
“unseen” discrimination is maintained in the context of
such a TMS-induced impairment of conscious detection
—that is, whether TMS-induced blindsight occurred—
and whether these “unseen” abilities are stimulus inde-
pendent (as predicted by the geniculate hypothesis) or
differentially impaired in the presence of s-cone versus
luminance stimuli (as predicted by the retinotectal hy-
pothesis). “Unseen” discrimination at 90–130 msec was
maintained above chance for both stimulus types (lumi-
nance: t(15) = 7.51, p < .001, s-cone: t(15) = 4.47, p <
.001; Figure 2D). This indicates the characteristic signa-
ture of TMS-induced blindsight and is the primary indica-
tion that residual capacity is preserved even when direct
input via the SC is withdrawn. “Unseen” ability was un-
affected by occipital TMS in the presence of both s-cone
stimuli and luminance stimuli (PcU luminance, active vs.
sham t(15) = 0.39, p = .70, B(luminance, active < sham) =
0.13, B(luminance, active > sham) = 0.07. PcU s-cone, active
vs. sham t(15) = 0.14, pag = .89, B(s-cone, active < sham) =
0.15, B(s-cone, active > sham) = 0.18). Además, compari-
sons between sham-normalized discrimination perfor-
mance confirmed no reliable difference between stimulus
types (see Figure 2B and D, PcU Δ sham luminance vs.
s-cone t(15) = 0.30, pag = .77, B(Δ sham luminance > s-cone) =
0.16, B(Δ sham luminance < s-cone) = 0.24). The strongest
trend in this comparison was a tendency toward reduced
1514
Journal of Cognitive Neuroscience
Volume 26, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
o
1
c
3
n
3
_
9
a
/
_
j
0
o
0
c
5
n
5
7
_
a
p
_
d
0
0
b
5
y
5
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
performance for luminance relative to s-cone stimuli
(B = 0.24), contrary to the pattern predicted by the retino-
tectal hypothesis. Taken together, the findings demon-
strate the persistence of “unseen” discrimination ability
when informative retinotectal input is prevented.
280–320 msec
The latest TMS intervention suppressed conscious de-
tection but only in the presence of luminance stimuli
(see Figure 2A *3, PrC luminance, active vs. sham
t(15) = 5.09, p < .001, B(luminance, active < sham) = 168.5,
B( luminance, active > sham) = 0.02. PrC s-cone, active vs.
sham t(15) = 0.31, pag = .76, B(s-cone, active < sham) = 0.11
B(s-cone, active > sham) = 0.08, PrC Δ sham luminance vs. s-cone
t(15) = 3.38, pag = .004, B(Δ sham, luminance > s-cone) = 0.03,
B(Δ sham, luminance < s-cone) = 10.62). This later disruption
of conscious processing, together with the disruption
caused by TMS at 90–130 msec, is consistent with the
notion that conscious processing is supported by recur-
rent activity (Lamme, 2001). “Unseen” discrimination was
above chance under active TMS (for luminance stimuli
t(15) = 6.98, p < .001; for s-cone stimuli t(15) = 6.90,
p < .001). Again “unseen” discrimination appeared to be
unaffected by the TMS and did not differ between stimulus
types (see Figure 2B, PcU luminance, active vs. sham
t(15) = 0.35, p = .73, B( luminance active < sham) = 0.08,
B( luminance active > sham) = 0.13, PcU s-cone, active vs.
sham t(15) = 1.13, pag = .28, B(s-cone active < sham) = 0.06,
B(s-cone active > sham) = 0.35, PcU Δ sham luminance vs.
s-cone t(15) = 0.60, pag = .55, B(Δ sham luminance > s-cone) =
0.11, B(Δ sham luminance < s-cone) = 0.28).
Discrimination performance when participants acknowl-
edged awareness of both the “arrow” and “something”
was close to ceiling (proportion correct; 0.97 ± 0.05 SD).
Performance when partial awareness of stimuli was
acknowledged, through a positive responses to the “some-
thing” question but denial of having seen the “arrow,”
was also high (proportion correct: 0.88 ± 0.15 SD). This
apparent correlation between level of awareness and dis-
crimination capacity is to be expected but does not violate
the zero correlation criteria for “unconscious” processing
(see Dienes, 2008a). This is because the claim is that
unconscious processing and blindsight are only observed
here when participants report “no” to both the “arrow”
and the “something” questions and therefore is consistent
with the “guessing criterion” definition of unconscious
capacity (Dienes, 2008a).
It is notable that PcU is calculated from “unseen” trials
only, whereas more trials contribute to the measure of
conscious detection (PrC and PcU were calculated from
a total of 22,275 and 5525 trials, respectively). Thus, it
might be argued that the analysis of PcU is hindered by
a relative lack of statistical power at an individual level. To
test this hypothesis, estimates of standard error based on
these specific trial numbers (the number of data points
convolved for the number of contributing trials) can be
compared with test whether heterogeneity of variance
could be responsible for the observation of significant
effects on one measure and not another. Keppel (1982)
recommends a 3:1 ratio of such variances (Fmax ratio)
as the cut-off point, beyond which the variances should
be considered heterogeneous and the conditions non-
comparable. For our data, the Fmax ratio was calculated
as 1.84:1 (0.032/0.017), thus satisfying this requirement.
DISCUSSION
Our results provide evidence that the residual abilities of
TMS-induced blindsight shown here are predominantly
geniculate (e.g., Schmid et al., 2010; Boyer et al., 2005)
rather than retinotectal in origin (e.g., Leh et al., 2010;
Ro et al., 2004). Blindsight was demonstrated most clearly
by the suppression of reported awareness at 90–130 msec
for both stimulus types, in the presence of above chance
“unseen” discrimination. This corresponds to the expected
epoch of visual suppression by occipital TMS (Amassian
et al., 1989). The central question posed was whether
the concurrent “unseen” abilities were dependent upon
stimulus type, that is, whether the residual abilities were
maintained for stimuli that are invisible to the retinotectal
route. The preservation of such abilities for s-cone stimuli
indicates that chromatic geniculate pathways rather than
the retinotectal route supported the residual abilities of
TMS-induced blindsight shown here. Moreover, it seems
unlikely that the absence of stimulus-specific effects on
the measure of “unseen” discrimination was due to the
stimuli not isolating the R/M pathways or the ineffec-
tiveness of TMS, because both TMS effects and stimulus
specificity were reliably demonstrated upon the contrast-
ing measure of conscious detection.
How can we reconcile previous evidence in favor of the
retinotectal hypothesis with the current evidence in sup-
port of the geniculate hypothesis? Much of the evidence
consistent with retinotectal mediation does not in fact
logically exclude a geniculate role, but in patients where
the geniculate pathways are surgically cut the evidence
for retinotectal mediation is particularly strong (Leh et al.,
2010; Tamietto et al., 2010; Leh, Mullen, et al., 2006). Our
results are not inconsistent with a role for the retinotectal
pathway in certain kinds of residual ability (i.e., for certain
types of stimuli), particularly following permanent brain
injury where plasticity may alter the functional contribu-
tion of different pathways (Silvanto & Rees, 2011; Huxlin,
2008; Mittmann & Eysel, 2001).
Rather than exclusively supporting the geniculate hy-
pothesis and disconfirming the retinotectal hypothesis,
we view our data as disconfirming the core idea implicit
in the long debate between these theories—that because
blindsight has been categorized as a single phenomenon
we should expect a single pathway to support it. Different
pathways are likely to support unconscious abilities that
depend on different types of information (Cowey, 2010;
Danckert & Rossetti, 2005). Some residual abilities for
Allen, Sumner, and Chambers
1515
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
o
1
c
3
n
3
_
9
a
/
_
j
0
o
0
c
5
n
5
7
_
a
p
_
d
0
0
b
5
y
5
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
some types of stimuli, within the wider scope of blindsight,
may be supported by input through the SC. However, the
positive demonstration of residual abilities when such
input is withdrawn indicates that the SC cannot be neces-
sary for the preserved capacity of TMS-induced blindsight,
as has been previously claimed (Ro et al., 2004).
The proposed correspondence between early feed-
forward activity and the residual “unseen” abilities was
neither supported nor refuted by the current data (Lamme
& Roelfsema, 2000). Our results do, however, indicate that
this period and these abilities are not wholly dependent
upon initial input provided by the SC. In contrast to un-
conscious abilities, R/M pathways did make a time-specific
contribution to conscious vision, thus validating the effi-
cacy of the chromatic intervention. No specific predictions
were cast in terms of s-cone dependency; hence, the con-
clusions that can be drawn in relation to these effects
are qualified as speculative. However, such effects upon
conscious awareness are clearly a possibility given the
significant contribution of R/M pathways to cortical input
(Nealey & Maunsell, 1994).
Interestingly, the earliest effect we observed (0–40 msec)
was a facilitation of conscious detection. Early TMS
has been reported to cause either perceptual suppres-
sion (Corthout, Uttl, Walsh, Hallett, & Cowey, 1999;
Corthout, Uttl, Ziemann, Cowey, & Hallett, 1999) or
enhancement (Abrahamyan, Clifford, Arabzadeh, & Harris,
2011; Schwarzkopf, Silvanto, & Rees, 2011). Such en-
hancement effects have been attributed to the addition
of neuronal noise, producing stochastic resonance and
driving cortical activity across a threshold for detection
(see Schwarzkopf et al., 2011). Earlier potentiation and
later suppression of functionality can be further under-
stood if the role of the early period is seen as preparatory
(Marr, 1982), potentially in the service of later conscious
processing. Because the state of activation leading up to
the arrival of information is crucial in determining whether
or not that information is perceived (Romei, Gross, & Thut,
2010; Mathewson, Gratton, Fabiani, Beck, & Ro, 2009), the
application of early TMS may possibly provide a pedestal
of activity, priming the region for subsequent processing.
This might not be the case for later applications because
increasing activity without interfering with information is
only possible when TMS is applied before the arrival of that
information. Here this rapid facilitation was only realized
in the presence of luminance stimuli, indicating that this
early period was dependent upon input provided by R/M
pathways. This is consistent with the relatively high speeds
at which these pathways transmit information (Bompas &
Sumner, 2008; McKeefry, Parry, & Murray, 2003; Maunsell
et al., 1999; Cottaris & De Valois, 1998), suggesting that
the R/M pathways may play a particularly prominent role
in the early preparatory period, potentially in the service
of conscious processing.
The observed disruption of conscious detection at later
TMS onset times is consistent with existing proposals
(Lamme, 2001) that delayed re-entrant processing sup-
ports conscious vision. The latest of these effects (280–
320 msec) occurred only in the presence of luminance
stimuli, indicating that information conveyed via R/M path-
ways supports this later period. What then provides this
late R/M input? Our findings provide no definitive answer
to this question. However, it seems unlikely to be depen-
dent upon feedforward or recurrent activity that has first
passed through the affected early visual cortex because
s-cone inputs are believed to be integrated with other
signals within these regions (Sincich & Horton, 2005;
Moutoussis & Zeki, 2000, 2002; Cottaris & De Valois,
1998). It is more plausible to speculate that R/M infor-
mation is first passed through unaffected areas—possibly
via the aforementioned extrastriate pathways—and is only
then fed back to interact with feedforward processing.
In this way, the later stage of conscious processing may
have been susceptible to later TMS because it is supported
by R/M inputs, present for luminance stimuli, that initially
enter unaffected frontoparietal networks (Corbetta, Patel,
& Shulman, 2008) before later feeding back into the dis-
rupted occipital cortex.
In conclusion, the residual visual abilities of TMS-
induced blindsight shown here originate primarily from
spared geniculate projections. Our findings thus stand in
contrast to the explanations of blindsight that rely on the
retinotectal pathway (Ro et al., 2004) and instead add
weight to evidence that spared regions in the geniculate
pathway can support unconscious vision following occipital
disruption (Schmid et al., 2010; Fendrich, Wessinger, &
Gazzaniga, 1992). Our results also indicate that conscious
vision depends on activity within the early occipital cortex
from ∼100 msec after stimulus onset and that R/M path-
ways make a contribution to awareness during early pre-
paratory (0–40 msec) and later feedback (280–320 msec)
stages of visual processing.
Acknowledgments
We thank Jacky Boivin, Aline Bompas, Zoltan Dienes, and Siân
Robson for their helpful comments on this research. This
research was supported by a BBSRC David Phillips Fellowship
(C. D. C.), BBSRC Grant BB/E020291/1 (C. D. C.), and the Wales
Institute of Cognitive Neuroscience (C. P. G. A./C. D. C.).
Reprint requests should be sent to Christopher P. G. Allen,
Cardiff University Brain Research Imaging Center, School of
Psychology, Cardiff University, Park Place, Cardiff CF10 3AT,
UK, or via e-mail: allencp@cardiff.ac.uk.
REFERENCES
Abrahamyan, A., Clifford, C. W., Arabzadeh, E., & Harris,
J. A. (2011). Improving visual sensitivity with subthreshold
transcranial magnetic stimulation. Journal of Neuroscience,
31, 3290–3294.
Amassian, V. E., Cracco, R. Q., Maccabee, P. J., Cracco,
J. B., Rudell, A., & Eberle, L. (1989). Suppression of
visual perception by magnetic coil stimulation of human
occipital cortex. Electroencephalography and Clinical
Neurophysiology, 74, 458–462.
1516
Journal of Cognitive Neuroscience
Volume 26, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
o
1
c
3
n
3
_
9
a
/
_
j
0
o
0
c
5
n
5
7
_
a
p
_
d
0
0
b
5
y
5
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Anderson, E. J., Husain, M., & Sumner, P. (2008). Human
intraparietal sulcus (IPS) and competition between
exogenous and endogenous saccade plans. Neuroimage,
40, 838–851.
Bompas, A., Sterling, T., Rafal, R. D., & Sumner, P. (2008).
Naso-temporal asymmetry for signals invisible to the
retinotectal pathway. Journal of Neurophysiology, 100,
412–421.
Bompas, A., & Sumner, P. (2008). Sensory sluggishness
dissociates saccadic, manual, and perceptual responses:
An S-cone study. Journal of Vision, 8, 10.1–10.13.
Bompas, A., & Sumner, P. (2009). Oculomotor distraction
by signals invisible to the retinotectal and magnocellular
pathways. Journal of Neurophysiology, 102, 2387–2395.
Bompas, A., & Sumner, P. (2011). Saccadic inhibition reveals
the timing of automatic and voluntary signals in the human
brain. Journal of Neuroscience, 31, 12501–12512.
Boyer, J. L., Harrison, S., & Ro, T. (2005). Unconscious
processing of orientation and color without primary visual
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 102, 16875–16879.
Campion, J., Latto, R., & Smith, Y. M. (1983). Is blindsight an
effect of scattered-light, spared cortex, and near-threshold
vision. Behavioral and Brain Sciences, 6, 423–447.
Chambers, C. D., Allen, C. P., Maizey, L., & Williams, M. A.
(2012). Is delayed foveal feedback critical for extra-foveal
perception? Cortex, 49, 327–335.
Christensen, M. S., Kristiansen, L., Rowe, J. B., & Nielsen,
J. B. (2008). Action-blindsight in healthy subjects after
transcranial magnetic stimulation. Proceedings of the
National Academy of Sciences, U.S.A., 105, 1353–1357.
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The
reorienting system of the human brain: From environment
to theory of mind. Neuron, 58, 306–324.
Corthout, E., Hallett, M., & Cowey, A. (2002). Early visual
cortical processing suggested by transcranial magnetic
stimulation. NeuroReport, 13, 1163–1166.
Corthout, E., Uttl, B., Walsh, V., Hallett, M., & Cowey, A.
(1999). Timing of activity in early visual cortex as revealed
by transcranial magnetic stimulation. NeuroReport, 10,
2631–2634.
Corthout, E., Uttl, B., Ziemann, U., Cowey, A., & Hallett, M.
(1999). Two periods of processing in the (circum)striate
visual cortex as revealed by transcranial magnetic stimulation.
Neuropsychologia, 37, 137–145.
Corwin, J. (1994). On measuring discrimination and response
bias: Unequal numbers of targets and distractors and two
classes of distractors. Neuropsychology, 8, 7.
Cottaris, N. P., & De Valois, R. L. (1998). Temporal dynamics
of chromatic tuning in macaque primary visual cortex.
Nature, 395, 896–900.
Cowey, A. (2010). The blindsight saga. Experimental Brain
Research, 200, 3–24.
Dacey, D. M. (2000). Parallel pathways for spectral coding
in primate retina. Annual Review of Neuroscience, 23,
743–775.
Danckert, J., & Rossetti, Y. (2005). Blindsight in action:
What can the different sub-types of blindsight tell us about
the control of visually guided actions? Neuroscience &
Biobehavioral Reviews, 29, 1035–1046.
de Monasterio, F. M. (1978). Properties of ganglion cells with
atypical receptive-field organization in retina of macaques.
Journal of Neurophysiology, 41, 1435–1449.
Derrington, A. M., Krauskopf, J., & Lennie, P. (1984). Chromatic
mechanisms in lateral geniculate nucleus of macaque.
The Journal of Physiology, 357, 241–265.
Dienes, Z. (2008a). Subjective measures of unconscious
knowledge. Progress in Brain Research, 168, 49–64.
Dienes, Z. (2008b). Understanding psychology as a science:
An introduction to scientific and statistical inference
(pp. 147–180). Basingstoke, UK: Palgrave Macmillan.
Dienes, Z. (2011). Bayesian versus orthodox statistics: Which
side are you on? Perspectives on Psychological Science,
6, 274–290.
Dretske, F. (2000). Perception without awareness. In T. S. G. J.
Hawthorne (Ed.), Perceptual experience (pp. 147–180).
Oxford: Oxford University Press.
Fendrich, R., Wessinger, C. M., & Gazzaniga, M. S. (1992).
Residual vision in a scotoma: Implications for blindsight.
Science, 258, 1489–1491.
Franca, M., Koch, G., Mochizuki, H., Huang, Y. Z., & Rothwell,
J. C. (2006). Effects of theta burst stimulation protocols
on phosphene threshold. Clinical Neurophysiology, 117,
1808–1813.
Gallagher, S. (2003). Phenomenology and experimental
design toward a phenomenologically enlightened
experimental science. Journal of Consciousness Studies,
10, 85.
Gallistel, C. R. (2009). The importance of proving the null.
Psychological Review, 116, 439–453.
Holm, S. (1979). A simple sequentially rejective multiple
test procedure. Scandinavian Journal of Statistics, 6,
65–70.
Huxlin, K. R. (2008). Perceptual plasticity in damaged adult
visual systems. Vision Research, 48, 2154–2166.
Jeffreys, E. T. (1961). The theory of probability (3rd ed.).
Oxford: Oxford University Press.
Jolij, J., & Lamme, V. A. (2005). Repression of unconscious
information by conscious processing: Evidence from affective
blindsight induced by transcranial magnetic stimulation.
Proceedings of the National Academy of Sciences, U.S.A.,
102, 10747–10751.
Keppel, G. (1982). Design and analysis: A researcherʼs
handbook (2nd ed.). Englewood Cliffs, NJ: Prentice-Hall.
Koivisto, M., Mantyla, T., & Silvanto, J. (2010). The role of
early visual cortex ( V1/ V2) in conscious and unconscious
visual perception. Neuroimage, 51, 828–834.
Lamme, V. A. (2001). Blindsight: The role of feedforward and
feedback corticocortical connections. Acta Psychologica,
107, 209–228.
Lamme, V. A., & Roelfsema, P. R. (2000). The distinct modes
of vision offered by feedforward and recurrent processing.
Trends in Neurosciences, 23, 571–579.
Leh, S. E., Johansen-Berg, H., & Ptito, A. (2006). Unconscious
vision: New insights into the neuronal correlate of blindsight
using diffusion tractography. Brain, 129, 1822–1832.
Leh, S. E., Mullen, K. T., & Ptito, A. (2006). Absence of S-cone
input in human blindsight following hemispherectomy.
European Journal of Neuroscience, 24, 2954–2960.
Leh, S. E., Ptito, A., Schonwiesner, M., Chakravarty, M. M.,
& Mullen, K. T. (2010). Blindsight mediated by an
S-cone-independent collicular pathway: An fMRI study
in hemispherectomized subjects. Journal of Cognitive
Neuroscience, 22, 670–682.
Loftus, G. R., & Masson, M. E. J. (1994). Using confidence-intervals
in within-subject designs. Psychonomic Bulletin & Review,
1, 476–490.
MacMillan, N. A., & Creelman, C. D. (1990). Detection theory:
A userʼs guide. Cambridge: Cambridge University Press.
Maizey, L., Allen, C. P. G., Dervinis, M., Verbruggen, F., Varnava,
A., Kozlov, M., et al. (2013). Comparative incidence rates
of mild adverse effects to transcranial magnetic stimulation.
Clinical Neurophysiology, 124, 8.
Marr, D. (1982). Vision: A computational investigation into
the human representation and processing of visual
information. San Francisco: W.H. Freeman.
Allen, Sumner, and Chambers
1517
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
o
1
c
3
n
3
_
9
a
/
_
j
0
o
0
c
5
n
5
7
_
a
p
_
d
0
0
b
5
y
5
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Marrocco, R. T., & Li, R. H. (1977). Monkey superior colliculus:
Properties of single cells and their afferent inputs. Journal
of Neurophysiology, 40, 844–860.
Mathewson, K. E., Gratton, G., Fabiani, M., Beck, D. M., & Ro, T.
(2009). To see or not to see: Prestimulus alpha phase predicts
visual awareness. Journal of Neuroscience, 29, 2725–2732.
Maunsell, J. H., Ghose, G. M., Assad, J. A., McAdams, C. J.,
Boudreau, C. E., & Noerager, B. D. (1999). Visual response
latencies of magnocellular and parvocellular LGN neurons
in macaque monkeys. Visual Neuroscience, 16, 1–14.
McKeefry, D. J., Parry, N. R., & Murray, I. J. (2003). Simple
reaction times in color space: The influence of chromaticity,
contrast, and cone opponency. Investigative Ophthalmology
& Visual Science, 44, 2267–2276.
Merigan, W. H., & Maunsell, J. H. (1993). How parallel are the
primate visual pathways? Annual Review of Neuroscience,
16, 369–402.
Mittmann, T., & Eysel, U. T. (2001). Increased synaptic plasticity
in the surround of visual cortex lesions in rats. NeuroReport,
12, 3341–3347.
Mollon, J. D. (1989). “Thoʼ she kneelʼd in that place where
they grew…” The uses and origins of primate colour vision.
The Journal of Experimental Biology, 146, 21–38.
Moutoussis, K., & Zeki, S. (2000). A psychophysical dissection
of the brain sites involved in color-generating comparisons.
Proceedings of the National Academy of Sciences, U.S.A.,
97, 8069–8074.
Moutoussis, K., & Zeki, S. (2002). Responses of spectrally
selective cells in macaque area V2 to wavelengths and colors.
Journal of Neurophysiology, 87, 2104–2112.
Nealey, T. A., & Maunsell, J. H. (1994). Magnocellular and
parvocellular contributions to the responses of neurons
in macaque striate cortex. Journal of Neuroscience, 14,
2069–2079.
Nisbett, R. E., & Wilson, T. D. (1977). Telling more than we can
know: Verbal reports on mental processes. Psychological
Review, 84, 28.
Overgaard, M., Rote, J., Mouridsen, K., & Ramsoy, T. Z.
(2006). Is conscious perception gradual or dichotomous?
A comparison of report methodologies during a visual
task. Consciousness and Cognition, 15, 700–708.
Radoeva, P. D., Prasad, S., Brainard, D. H., & Aguirre, G. K.
(2008). Neural activity within area V1 reflects unconscious
visual performance in a case of blindsight. Journal of
Cognitive Neuroscience, 20, 1927–1939.
Rafal, R., Smith, J., Krantz, J., Cohen, A., & Brennan, C.
(1990). Extrageniculate vision in hemianopic humans:
Saccade inhibition by signals in the blind field. Science,
250, 118–121.
Railo, H., Salminen-Vaparanta, N., Henriksson, L., Revonsuo, A.,
& Koivisto, M. (2012). Unconscious and conscious processing
of color rely on activity in early visual cortex: A TMS study.
Journal of Cognitive Neuroscience, 24, 819–829.
Ro, T., Shelton, D., Lee, O. L., & Chang, E. (2004).
Extrageniculate mediation of unconscious vision in
transcranial magnetic stimulation-induced blindsight.
Proceedings of the National Academy of Sciences,
U.S.A., 101, 9933–9935.
Romei, V., Gross, J., & Thut, G. (2010). On the role of
prestimulus alpha rhythms over occipito-parietal areas in
visual input regulation: Correlation or causation? Journal
of Neuroscience, 30, 8692–8697.
Rouder, J. N., Speckman, P. L., Sun, D., Morey, R. D., &
Iverson, G. (2009). Bayesian t tests for accepting and
rejecting the null hypothesis. Psychonomic Bulletin &
Review, 16, 225–237.
Schiller, P. H., & Malpeli, J. G. (1977). Properties and tectal
projections of monkey retinal ganglion cells. Journal of
Neurophysiology, 40, 428–445.
Schmid, M. C., Mrowka, S. W., Turchi, J., Saunders, R. C.,
Wilke, M., Peters, A. J., et al. (2010). Blindsight depends
on the lateral geniculate nucleus. Nature, 466, 373–377.
Schwarzkopf, D. S., Silvanto, J., & Rees, G. (2011). Stochastic
resonance effects reveal the neural mechanisms of
transcranial magnetic stimulation. Journal of Neuroscience,
31, 3143–3147.
Silvanto, J., & Rees, G. (2011). What does neural plasticity
tell us about role of primary visual cortex ( V1) in visual
awareness? Frontiers in Psychology, 2, 6.
Sincich, L. C., & Horton, J. C. (2005). The circuitry of V1 and
V2: Integration of color, form, and motion. Annual Review
of Neuroscience, 28, 303–326.
Smithson, H., Sumner, P., & Mollon, J. D. (2003). How to find
a tritan line. In J. D. Mollon (Ed.), Normal and defective
colour vision (pp. 279–287). Oxford: Oxford University Press.
Sumner, P. (2006). Inhibition versus attentional momentum
in cortical and collicular mechanisms of IOR. Cognitive
Neuropsychology, 23, 1035–1048.
Sumner, P., Adamjee, T., & Mollon, J. D. (2002). Signals invisible
to the collicular and magnocellular pathways can capture
visual attention. Current Biology, 12, 1312–1316.
Sumner, P., Nachev, P., Castor-Perry, S., Isenman, H.,
& Kennard, C. (2006). Which visual pathways cause
fixation-related inhibition? Journal of Neurophysiology,
95, 1527–1536.
Sumner, P., Nachev, P., Vora, N., Husain, M., & Kennard, C.
(2004). Distinct cortical and collicular mechanisms
of inhibition of return revealed with S cone stimuli.
Current Biology, 14, 2259–2263.
Tamietto, M., Cauda, F., Corazzini, L. L., Savazzi, S., Marzi,
C. A., Goebel, R., et al. (2010). Collicular vision guides
nonconscious behavior. Journal of Cognitive Neuroscience,
22, 888–902.
Taylor, J. R. (1997). An introduction to error analysis:
The study of uncertainties in physical measurements
(2nd ed.). Sausalito, CA: University Science.
Varela, F. J. (1996). Neurophenomenology: A methodological
remedy for the hard problem. Journal of Consciousness
Studies, 3, 20.
Weiskrantz, L. (1986). Blindsight: A case study and
implications ( Vol. 12). Oxford: Oxford University Press.
Wessinger, C. M., Fendrich, R., & Gazzaniga, M. S. (1997).
Islands of residual vision in hemianopic patients. Journal
of Cognitive Neuroscience, 9, 203–221.
White, B. J., Boehnke, S. E., Marino, R. A., Itti, L., & Munoz,
D. P. (2009). Color-related signals in the primate superior
colliculus. Journal of Neuroscience, 29, 12159–12166.
Winer, B. J., Brown, D. R., & Michels, K. M. (1991). Statistical
principles in experimental design (3rd ed.). New York:
McGraw-Hill.
1518
Journal of Cognitive Neuroscience
Volume 26, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
7
2
6
1
/
5
7
0
/
7
1
1
5
9
0
4
7
7
/
2
1
9
7
4
8
o
1
c
3
n
3
_
9
a
/
_
j
0
o
0
c
5
n
5
7
_
a
p
_
d
0
0
b
5
y
5
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1