What Role Does the Anterior Temporal Lobe Play
in Sentence-level Processing? Neural Correlates
of Syntactic Processing in Semantic Variant
Primary Progressive Aphasia
Stephen M. Wilson1, Andrew T. DeMarco1, Maya L. Henry2,
Benno Gesierich2, Miranda Babiak2, Maria Luisa Mandelli2,
Bruce L. Miller2, and Maria Luisa Gorno-Tempini2
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
oh
1
C
2
norte
8
_
4
a
/
_
j
0
oh
0
C
5
norte
5
0
_
a
pag
_
d
0
0
b
5
y
5
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
.
t
/
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Abstracto
■ Neuroimaging and neuropsychological studies have im-
plicated the anterior temporal lobe (ATL) in sentence-level
Procesando, with syntactic structure-building and/or combinatorial
semantic processing suggested as possible roles. A potential
challenge to the view that the ATL is involved in syntactic aspects
of sentence processing comes from the clinical syndrome of
semantic variant primary progressive aphasia (semantic PPA; también
known as semantic dementia). In semantic PPA, bilateral neuro-
degeneration of the ATLs is associated with profound lexical
semantic deficits, yet syntax is strikingly spared. The goal of this
study was to investigate the neural correlates of syntactic pro-
cessing in semantic PPA to determine which regions normally
involved in syntactic processing are damaged in semantic PPA
and whether spared syntactic processing depends on preserved
functionality of intact regions, preserved functionality of atrophic
regiones, or compensatory functional reorganization. We scanned
20 individuals with semantic PPA and 24 age-matched controls
using structural MRI and fMRI. Participants performed a sentence
comprehension task that emphasized syntactic processing and
minimized lexical semantic demands. We found that, in controls,
left inferior frontal and left posterior temporal regions were
modulated by syntactic processing, whereas anterior temporal
regions were not significantly modulated. In the semantic PPA
grupo, atrophy was most severe in the ATLs but extended to
the posterior temporal regions involved in syntactic processing.
Functional activity for syntactic processing was broadly similar
in patients and controls; En particular, whole-brain analyses re-
vealed no significant differences between patients and controls
in the regions modulated by syntactic processing. The atrophic
left ATL did show abnormal functionality in semantic PPA
patients; sin embargo, this took the unexpected form of a failure to
deactivate. Tomados juntos, our findings indicate that spared
syntactic processing in semantic PPA depends on preserved func-
tionality of structurally intact left frontal regions and moderately
atrophic left posterior temporal regions, but no functional re-
organization was apparent as a consequence of anterior temporal
atrophy and dysfunction. These results suggest that the role of
the ATL in sentence processing is less likely to relate to syntactic
structure-building and more likely to relate to higher-level pro-
cesses such as combinatorial semantic processing. ■
INTRODUCCIÓN
In the classical literature on aphasia and the neural sub-
strates of language, the anterior temporal lobe (ATL) era
not considered to be an important language region. Cor-
tical stimulation of the ATL during presurgical language
mapping did not typically induce speech errors or apha-
sic errors, and the ATL was frequently resected to treat
intractable epilepsy without resulting in obvious aphasias
(Penfield & Roberts, 1959). Además, focal strokes to
the ATL are very uncommon because of vascular anatomy,
so there were no relevant data from cerebrovascular
patients (Holanda & Lambon Ralph, 2010). Sin embargo, el
past two decades have seen an increasing body of evi-
dence for critical involvement of the ATL in two aspects
1University of Arizona, 2Universidad de California, San Francisco
© 2014 Instituto de Tecnología de Massachusetts
of language: primero, in the representation and processing of
lexical semantic information (para una revisión, see Patterson,
Nestor, & Rogers, 2007), y segundo, in sentence-level
Procesando (Humphries, Binder, Medler, & Liebenthal,
2006; Dronkers, Wilkins, Van Valin, Redfern, & Jaeger,
2004; Vandenberghe, Nobre, & Precio, 2002; Mazoyer
et al., 1993).
The role of the ATL in sentence-level processing has
been primarily motivated by numerous PET and fMRI
studies that have shown increased ATL activation for
sentences compared with matched word lists (Pallier,
Devauchelle, & Dehaene, 2011; Rogalsky & Hickok, 2009;
Humphries et al., 2006; Humphries, Amar, Swinney, &
Hickok, 2005; Xu, Kemeny, Parque, Frattali, & Braun, 2005;
Vandenberghe et al., 2002; Humphries, Willard, Buchsbaum,
& Hickok, 2001; Friederici, Meyer, & por cramon, 2000;
Stowe et al., 1999; Mazoyer et al., 1993). As many of these
Revista de neurociencia cognitiva 26:5, páginas. 970–985
doi:10.1162/jocn_a_00550
authors have pointed out, it is difficult to determine what
aspect of sentence processing is responsible for the greater
activation of the ATL for sentences than word lists, porque
sentence comprehension entails not only syntactic pro-
cesses to parse the sentence and build the syntactic struc-
tura, but also combinatorial semantic processes that
integrate the meanings of the words in the sentence to
arrive at a global meaning.
The strongest evidence that the ATL might play a spe-
cifically syntactic role in sentence comprehension has
come from several studies reporting that even sentences
in which content words were replaced with pseudo-
palabras (“Jabberwocky sentences”) activated the ATL more
than matched pseudoword lists (Humphries et al., 2006;
Friederici et al., 2000; Mazoyer et al., 1993). Estos hallazgos
suggest that ATL activation for sentences reflects syntactic
procesos, because the pseudowords in the jabberwocky
sentences lack any lexical semantic representations, cual
should limit the recruitment of combinatorial semantic
procesos. Additional findings supporting a syntactic role
for the ATL include two voxel-based lesion-symptom
mapping studies showing associations between anterior
temporal damage and sentence comprehension deficits
(Magnusdottir et al., 2013; Dronkers et al., 2004), adap-
tation effects for repeated syntactic structures in the ATL
(Noppeney & Precio, 2004), and ATL activation associated
with time-varying syntactic structure-building in natural-
istic contexts (Brennan et al., 2012).
A potential challenge to the view that the ATL is in-
volved in syntactic aspects of sentence processing comes
from the clinical syndrome of semantic variant primary
progressive aphasia (semantic PPA; also known as seman-
tic dementia; Hodges, Patterson, Oxbury, & Funnell,
1992; Snowden, Goulding, & Neary, 1989; Warrington,
1975). Semantic PPA is characterized by dramatic bilateral
atrophy of the ATL and profound lexical semantic deficits,
yet syntactic function is almost completely spared (for a re-
vista, see Wilson, Galantucci, Tartaglia, & Gorno-Tempini,
2012). Patients with semantic PPA almost invariably score
close to ceiling on sentence comprehension measures so
long as lexical demands are minimized ( Wilson et al.,
2011; Gorno-Tempini et al., 2004; Hodges et al., 1992;
Warrington, 1975) and they show normal sensitivity to
syntactic violations in on-line and off-line tasks (Cotelli
et al., 2007; Grossman, Rhee, & moore, 2005). Otro
patient groups with ATL damage have also been reported
to have spared sentence comprehension, including pa-
tients who have had anterior temporal lobectomies (Kho
et al., 2008) and patients with herpes simplex encephalitis
(Kapur et al., 1994).
Spared syntax in the face of ATL damage constitutes a
challenge to the view that the ATL is involved in syntactic
Procesando. Sin embargo, it is important to determine whether
the specific regions thought to be involved in syntactic
processing are damaged and whether any compensatory
functional reorganization might have taken place that
could compensate for the damaged ATL. The goal of this
study was to investigate the neural correlates of syntactic
processing in semantic PPA to address these two ques-
ciones. Específicamente, we first determined which regions that
are normally involved in syntactic processing are damaged
in semantic PPA by comparing activations for syntactic
processing in age-matched controls with patterns of
atrophy in semantic PPA. Segundo, we asked whether
spared syntactic processing in semantic PPA depends on
preserved functionality of intact regions, preserved func-
tionality of atrophic regions, or functional reorganiza-
ción. If the ATL is critical for syntactic processing, cual
is largely spared in semantic PPA, then we would expect
that syntactic processing in semantic PPA would depend
on preserved functionality in the ATL despite atrophy
and/or functional reorganization such that relatively intact
regions compensate for the damaged ATL.
MÉTODOS
Participantes
We successfully acquired structural and functional imag-
ing data in 20 patients with semantic PPA and 24 edad-
matched control participants over a 4-year period. Patients
and age-matched controls were recruited through the
Memory and Aging Center at the University of California,
San Francisco (UCSF). All participants gave written
informed consent, and the study was approved by the
institutional review boards at UCSF and the University of
Arizona. Patients and controls received a comprehensive
multidisciplinary evaluation including neurological his-
tory and examination, neuropsychological testing, y
neuroimaging.
Patients were diagnosed with semantic variant PPA based
on recent guidelines (Gorno-Tempini et al., 2011). A diag-
nosis of PPA required progressive deterioration of speech
and/or language functions and that deficits be largely
restricted to speech and/or language for at least 2 años. A
diagnosis of semantic variant PPA required two core fea-
tures to be present: impaired confrontation naming and
impaired single word comprehension. At least three of the
four following features were also required: impaired object
conocimiento, surface dyslexia or dysgraphia, spared repeti-
ción, and spared speech production. Neuroimaging results
were not used for diagnostic purposes, but only to rule
out other causes of focal brain damage. Sin embargo, anterior
temporal atrophy was evident in all 20 patients.
Additional inclusion criteria were fluency in English and
a Mini-Mental State Examination score of at least 15. A
total of 23 patients met these criteria and were scanned,
but two patients requested that the scanning session be
stopped before the functional run was complete, y
one patient had excessive head motion that could not
be corrected, so all analyses were based on the remaining
20 patients.
Demographic, clinical, and neuropsychological char-
acteristics for all participants are provided in Table 1.
Wilson et al.
971
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
oh
1
C
2
norte
8
_
4
a
/
_
j
0
oh
0
C
5
norte
5
0
_
a
pag
_
d
0
0
b
5
y
5
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
.
/
F
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Mesa 1. Demographic, Clinical, and Neuropsychological Characteristics of Patients and Controls
Semantic PPA
Demographic
Age
Sex (M/F)
Handedness (R/L)
Educación (años)
Clinical
Mini Mental Status Examination (30)
Clinical Dementia Rating
Age at disease onset
Years from first symptom
Language Production
Confrontation naming (BNT, 15)b
Phonemic fluency (D words in 1 mín.)
Semantic fluency (Animals in 1 mín.)b
Speech fluency ( WAB, 10)
Apraxia of speech rating (MSE, 7)
Dysarthria rating (MSE, 7)
Repetition ( WAB, 100)
Language Comprehension
Auditory word recognition (PPVT, 16)b
Sequential commands ( WAB, 80)
Semantic knowledge (PPT-P, 52)b
Visuospatial Function
Modified Rey-Osterrieth copy (17)
Visual Memory
Modified Rey-Osterrieth delayed copy (17)
Verbal Memory
CVLT-MS Trials 1–4 (40)
CVLT-MS 30 sec free recall (10)
CVLT-MS 10 min free recall (10)
Executive Function
Digit span backwards
Modified Trail Making Test (lines per min)
Calculation ( WAB, 5)
63.4 ± 6.5
11/9
17/3
16.5 ± 2.5
26.3 ± 3.0a
0.6 ± 0.2
58.8 ± 7.5
4.7 ± 2.7
5.4 ± 3.8a
7.5 ± 5.0a
7.8 ± 4.7a
8.7 ± 0.9a
0.0 ± 0.0
0.0 ± 0.0
92.2 ± 8.3a
8.9 ± 4.4a
75.7 ± 8.0a
42.7 ± 6.4a
15.7 ± 1.0
8.2 ± 4.3a
16.4 ± 7.7a
2.9 ± 2.5a
1.9 ± 2.0a
5.3 ± 1.3
24.5 ± 9.8a
4.7 ± 0.6
Control S
66.8 ± 4.2
7/17
22/2
17.2 ± 2
29.3 ± 0.8
N/A
N/A
N/A
14.5 ± 0.7
17.6 ± 4.1
23.8 ± 4.3
10.0 ± 0.0c
N/A
N/A
99.5 ± 0.9c
15.7 ± 0.7
80.0 ± 0.0c
51.8 ± 0.4c
15.0 ± 1.1
11.7 ± 2.3
28.7 ± 3.1
7.9 ± 1.6
7.3 ± 1.6
5.6 ± 1.2
37.8 ± 12.6
4.8 ± 0.4
Values are means ± standard deviation. Variables were compared between patients and controls using Welchʼs t tests where appropriate, the Wilcoxon
signed-rank test for measures with floor or ceiling effects, and Fisherʼs exact test for discrete variables. See Kramer et al. (2003) for detailed description
of neuropsychological testing procedures and Gorno-Tempini et al. (2004) for detailed description of language testing procedures.
BNT = Boston Naming Test; WAB = Western Aphasia Battery; MSE = Motor Speech Evaluation; PPVT = Peabody Picture Vocabulary Test; PPT-P =
Pyramids and Palm Trees-Pictures; CVLT-MS = California Verbal Learning Test-Mental Status.
aSignificantly impaired relative to controls, pag < .05.
bVariables included in composite semantic score.
cData from Gorno-Tempini et al. (2004) because present control group was not tested on these variables.
972
Journal of Cognitive Neuroscience
Volume 26, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
There were no significant differences between patients
and controls in age, sex, handedness, or education.
As expected, semantic PPA patients had significant
deficits on all measures related to lexical semantic func-
tion, including confrontation naming and auditory word
recognition.
In addition to the 24 controls who took part in functional
imaging, structural images from an additional 50 age-
matched controls were used to create a template for
intersubject normalization and voxel-based morphometry.
Experimental Design
The design of the fMRI study has been described in detail
previously (Wilson, Dronkers, et al., 2010), so only a brief
summary is presented here. We scanned participants with
fMRI as they performed an auditory sentence-to-picture
matching task. Each trial began with the presentation of
two pictures: a target and a foil, one on the left and one
on the right of the screen. One second later, a sentence
was presented auditorily that matched one of the two
pictures. Participants selected the matching picture
at any point by pressing one of two buttons with their
right hand.
There were seven conditions, which differed in the syn-
tactic processing required as well as in length (Table 2). In
two conditions (Short Lexical, Long Lexical), the correct
response could be determined based on lexical infor-
mation alone, because the foil pictures involved different
lexical items than the target pictures. In two more con-
ditions (Short Active, Long Easy), the foil pictures con-
tained the same lexical items as the target pictures, so it
was necessary to attend to syntactic structures to deter-
mine the correct response, but the syntactic structures
involved were canonical: Elements were arranged in con-
figurations that are prototypical in English. The final three
conditions (Short Passive, Long Medium, Long Hard) also
required processing of syntactic structures to determine
the correct response, but now the structures involved were
noncanonical: Elements were displaced from their proto-
typical positions. Within each length category (short or
long), all conditions were matched for length, for lexical
content, and for the point at which the sentence disambig-
uated between the target and foil pictures. To keep lexical
demands to a minimum, all sentences were constructed
using just two high-frequency nouns (boy, girl), one of
seven high-frequency verbs (push, pull, kiss, kick, chase,
wash, hug), and for the long sentences, one of three
high-frequency color adjectives (red, green, blue). It was
important to minimize lexical demands because semantic
PPA patients typically have severe lexical deficits. Note
that no “scrambled” condition was included, because
pilot studies had revealed that neurodegenerative patients
were often confused by such conditions and responded
to them in wildly variable ways.
Before scanning, participants were trained on the task.
They were then scanned on a Siemens 3-T Trio scanner.
T1-weighted anatomical images and T2*-weighted fMRI
images (repetition time = 2000 msec; whole-brain cover-
age) were acquired using standard sequences as described
previously (Wilson, Dronkers, et al., 2010). Auditory stim-
uli were presented via headphones, visual stimuli were
presented via a screen that was viewed through a mirror,
and button presses were recorded. There were three
blocks per condition, for a total of 21 blocks, presented
in random order. Each block was 28 sec in length and
contained four equally spaced trials, and there were 16-sec
rest periods between blocks and at the beginning and end
of the experiment. The total duration of the functional
sequence was 15 m 40 sec.
Table 2. Sentence Structures and Foils in the Seven Conditions
Condition
Short Lexical
Short Active
Short Passivea
Long Lexical
Sentence Example
Foil Picture
The boy is hugging the girl.
The girl is kissing the boy.
The boy is kicked by the girl.
Different action (e.g., pulling)
Agent and patient reversed
Agent and patient reversed
The girl who is green is pushing the boy.
Different action (e.g., chasing)
The boy is kissing the girl who is red.
Different colored patient (e.g., green girl)
Long Easy
The girl is pulling the boy who is red.
Color assigned to wrong participant
Long Mediuma
The girl who is washed by the boy is green.
Color assigned to wrong participant
The boy who is red is chasing the girl.
Agent and patient reversed
Long Harda
The boy who the girl is pushed by is blue.
Agent and patient reversed
The girl who the boy is hugging is red.
Agent and patient reversed
The girl who the boy is kissed by is green.
Color assigned to wrong participant
aNoncanonical structures. See Wilson, Dronkers, et al. (2010) for a more detailed description and linguistic analysis of the stimuli.
Wilson et al.
973
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Analysis of Behavioral Data
Overall accuracy and RT and differences in accuracy and
RT between noncanonical and canonical conditions were
compared between semantic PPA patients and controls
using two-tailed Welchʼs t tests. Correlations were cal-
culated between overall accuracy and disease duration
and between overall accuracy and a composite semantic
measure (see Table 1).
Analysis of Structural Imaging Data
The T1-weighted structural images were bias-corrected; seg-
mented into gray matter, white matter, and cerebrospinal
fluid; and initially normalized to Montreal Neurological
Institute (MNI) space using the unified segmentation
algorithm in SPM5 (Ashburner & Friston, 2005). More
anatomically precise intersubject registration was then
performed with the DARTEL (Diffeomorphic Anatomical
Registration Through Exponentiated Lie algebra) toolbox
(Ashburner, 2007) by warping each participantʼs image to
a template created from the 50 additional control partici-
pants. Gray matter and white matter probability maps were
scaled by Jacobians, smoothed with a Gaussian kernel of
12 mm FWHM, then summed together to obtain a map
of brain parenchyma (Wilson, Henry, et al., 2010).
To map atrophy at the group level, we compared the
semantic PPA patients with 74 age-matched controls (the
24 controls who took part in the functional study plus
the 50 additional controls), with age, sex, and total intra-
cranial volume included as covariates. Percent volume
loss was plotted for regions with at least 10% volume loss.
To identify regions where volume loss was predictive
of syntactic deficits, we calculated voxelwise correlations
of tissue volume with overall accuracy on the sentence
comprehension task in the group of 20 semantic PPA
patients only, with the same covariates. The resulting
t map was thresholded at voxelwise p < .005, then cor-
rected for multiple comparisons based on cluster size with
respect to 1000 permutations in which behavioral scores
were randomly reassigned (Wilson, Henry, et al., 2010).
Analysis of Functional Imaging Data
The functional data were visually inspected, and volumes
where there was excessive head motion (visible inter-
leaving artifact) or other artifacts were replaced with inter-
polated data before preprocessing and later excluded from
analysis. The number of volumes excluded was 19.5 ±
22.7 in semantic PPA patients and 11.5 ± 15.9 in controls
(ns). The functional imaging data were preprocessed with
standard methods in AFNI (Cox, 1996): The data were
corrected for slice timing differences, realigned to account
for head movement, smoothed with a Gaussian kernel of
8 mm FWHM, high pass filtered (cut-off = 100 sec) and
detrended (Legendre polynomials of order up to and
including 2). Independent component analysis was then
carried out using the FSL tool melodic (Beckmann &
Smith, 2004), generating 30 components. Noise compo-
nents were manually identified with reference to published
criteria (Kelly et al., 2010) and removed using fsl_regfilt.
The mean number of components removed was 9.1 ±
2.4 in semantic PPA patients and 9.0 ± 2.4 in controls
(ns). Because the ATL is sensitive to susceptibility arti-
facts, we calculated a map of temporal signal-to-noise ratio.
The mean temporal signal-to-noise ratio in the ATL ROI
described below was 165, sufficient to detect functional
changes.
We analyzed the data using two different whole-brain
general linear models (GLMs), each of which was fit using
the fmrilm procedure from FMRISTAT ( Worsley et al.,
2002). The first was a conventional parametric block de-
sign model, and the second was based on individual par-
ticipantsʼ RTs on individual trials. We also carried out ROI
analyses in nine left hemisphere ROIs to quantify the
distribution of structural and functional imaging measures
in these key regions and to identify any correlations be-
tween different imaging measures and behavior.
In the first whole-brain GLM analysis, the design matrix
contained one explanatory variable for each of the seven
conditions, each consisting of a boxcar function convolved
with a standard hemodynamic response function. Ad-
ditional covariates of no interest were included to reduce
error variance: three translation and three rotation param-
eters (saved during realignment), signal time courses from
a white matter ROI, a cerebrospinal fluid ROI, and the
whole-brain global signal and three cubic spline temporal
trends. The primary contrast of interest was between the
three conditions involving noncanonical structures and
the four conditions that involved only canonical struc-
tures. The purpose of this contrast was to identify regions
modulated by syntactic complexity. Note that the Short
Passive condition was multipled by 2 in the contrast, so
that the contrast was balanced across short and long con-
ditions. A second contrast of interest was between all
seven conditions and the resting baseline. This contrast
identified regions involved in sentence comprehension
in general, along with nonlinguistic functions related to
the task, such as visual, auditory, motor, and executive
processes.
In the second whole-brain GLM analysis, we used RT as
a proxy for allocation of syntactic processing resources.
RT was measured from the onset of the first word in the
sentence that disambiguated the target from the foil, and
we assumed that more syntactic processing took place on
trials with longer RTs. This approach enabled a direct
comparison of the regions that patients and controls re-
cruited to perform the task, despite individual and group
differences in performance. The four trials within each
block were treated as separate events with a duration of
4 sec each, convolved with a hemodynamic response
function. Short and long trials were modeled with separate
explanatory variables, each of which was parametrically
modulated by another variable coding RT for each trial.
974
Journal of Cognitive Neuroscience
Volume 26, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
RT was demeaned based on the mean of all short or long
trials and clipped at 1.5 standard deviations from the
mean. Correct and incorrect trials were included, because
we were interested in identifying regions recruited for
syntactic processing regardless of whether that processing
was successful. (Note that including only correct trials
yielded very similar results, because performance was gen-
erally accurate for both groups.) Trials on which there
was no response were not included; they were modeled
with a separate explanatory variable of no interest. The
same covariates of no interest were included as in the
first analysis. The contrast of interest was signal change
per second of RT, which was averaged across the short
and long parametric variables.
Random effects analyses were carried out on contrast
images from individual participants, which were normal-
ized to MNI space by applying the transformations de-
scribed above. Patients and controls were compared
with Welchʼs t tests. All statistical maps were thresholded
at voxelwise p < .005 and then corrected for multiple
comparisons at p < .05 based on cluster extent accord-
ing to Gaussian random field theory implemented in
SPM5. To ameliorate loss of power due to multiple com-
parisons, clusters in liberally defined potential language
areas were corrected for the total combined volume of
these language areas, whereas clusters outside these areas
were corrected based on whole-brain volume. Potential
language areas were defined as almost the entire lateral
left hemisphere, specifically the left superior temporal
gyrus (STG), middle temporal gyrus (MTG), inferior tem-
poral gyrus (ITG), Heschlʼs gyrus, temporal pole, supra-
marginal gyrus, angular gyrus, inferior frontal gyrus (IFG),
middle frontal gyrus, premotor cortex, and insula (Tzourio-
Mazoyer et al., 2002).
Analyses identifying regions with increased activation in
semantic PPA relative to controls were inclusively masked
with regions showing positive activations in semantic PPA
(to identify abnormally activated regions) or deactivations
in age-matched controls (to identify failure to deactivate
normally). Conversely, analyses identifying regions with
decreased activation in semantic PPA relative to controls
were inclusively masked with regions showing deactiva-
tions in semantic PPA (to identify abnormally deactivated
regions) or positive activations in age-matched controls
(to identify lack of normal activations). All masks were
defined at voxelwise p < .005, uncorrected.
ROI analyses were carried out using MATLAB in nine
left hemisphere ROIs, each defined as a sphere with a
radius of 8 mm. Six of the ROIs (anterior STS, posterior
STS, TPJ, IFG pars opercularis, premotor cortex, and ante-
rior insula) were centered on activation peaks in the con-
trast of noncanonical versus canonical conditions, averaged
across semantic PPA patients and controls, so as not to
bias them with respect to comparisons between patients
and controls. A temporal pole ROI was centered on the
mean coordinates (−50, 3, −20) of ATL activations related
to syntactic or combinatorial semantic processing in nine
previous studies (Brennan et al., 2012; Pallier et al.,
2011; Rogalsky & Hickok, 2009; Humphries et al., 2006;
Xu et al., 2005; Dronkers et al., 2004; Noppeney & Price,
2004; Vandenberghe et al., 2002; Stowe et al., 1999).
Finally, anterior MTG and IFG pars triangularis ROIs were
centered on the peaks of regions that differed between
semantic PPA patients and controls in whole-brain analyses
of regions activated for sentence comprehension relative
to rest (see below); our intention was to examine measures
other than the contrast on the basis of which the regions
had been defined.
For each of the nine ROIs, patients and controls were
compared with two-tailed Welchʼs t tests on one structural
imaging measure (tissue volume) and three functional
imaging measures (modulation by syntactic complexity,
modulation by RT, and activation for the sentence com-
prehension task relative to rest). These 36 comparisons
were corrected for multiple comparisons using the false
discovery rate (FDR) procedure implemented in MATLAB.
For each ROI, correlations were computed between (1)
tissue volume and accuracy, (2) tissue volume and each
of the three functional measures, and (3) each of the three
functional measures and accuracy. These 63 correlations
were corrected for multiple comparisons using the FDR
procedure.
RESULTS
Behavioral Data
Semantic PPA patients performed the sentence com-
prehension task quite accurately (86.1% ± 5.8% correct);
however, they were less accurate than controls (92.8% ±
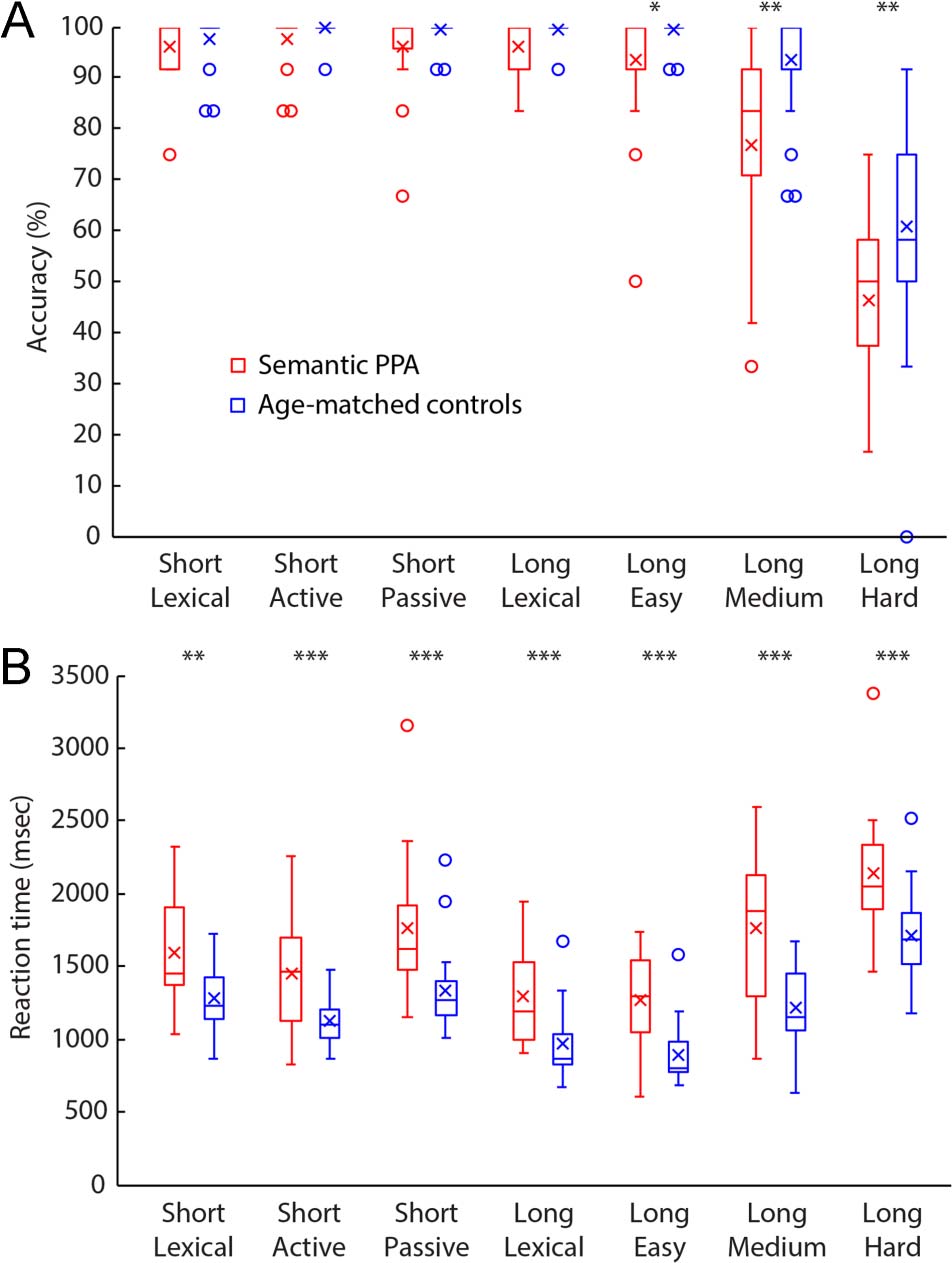
3.1%), t(27.72) = −4.61, p < .0001 (Figure 1A). Accuracy
in the patient group was not related to disease duration
(r = −0.21, p = .39) or the composite semantic mea-
sure (r = −0.07, p = .78). Semantic PPA patients also
performed disproportionately poorly relative to controls
on more syntactically complex (noncanonical) conditions
compared with less complex (canonical) conditions,
t(30.21) = −2.75, p = .0099.
Semantic PPA patients also responded more slowly over-
all (1612 ± 321 msec) than controls (1218 ± 175 msec),
t(28.18) = 4.90; p < .0001, and they responded dispropor-
tionately more slowly on syntactically complex conditions,
t(33.01) = 2.12; p = .042 (Figure 1B).
Although semantic PPA patients did not perform as well
as controls, it is noteworthy that they performed much
better than the nonfluent/agrammatic PPA patients we
investigated using this task in a prior study, whose mean
accuracy was just 69.8% ± 14.0% (Wilson, Dronkers, et al.,
2010).
Atrophy in Semantic PPA Patients
The most markedly atrophic regions in the semantic PPA
patients were the temporal lobes, with atrophy much
Wilson et al.
975
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
brain region. Note that these control data have been pub-
lished previously (Wilson, Dronkers, et al., 2010); how-
ever, we used improved image analysis procedures in
this study, so the results are not identical.
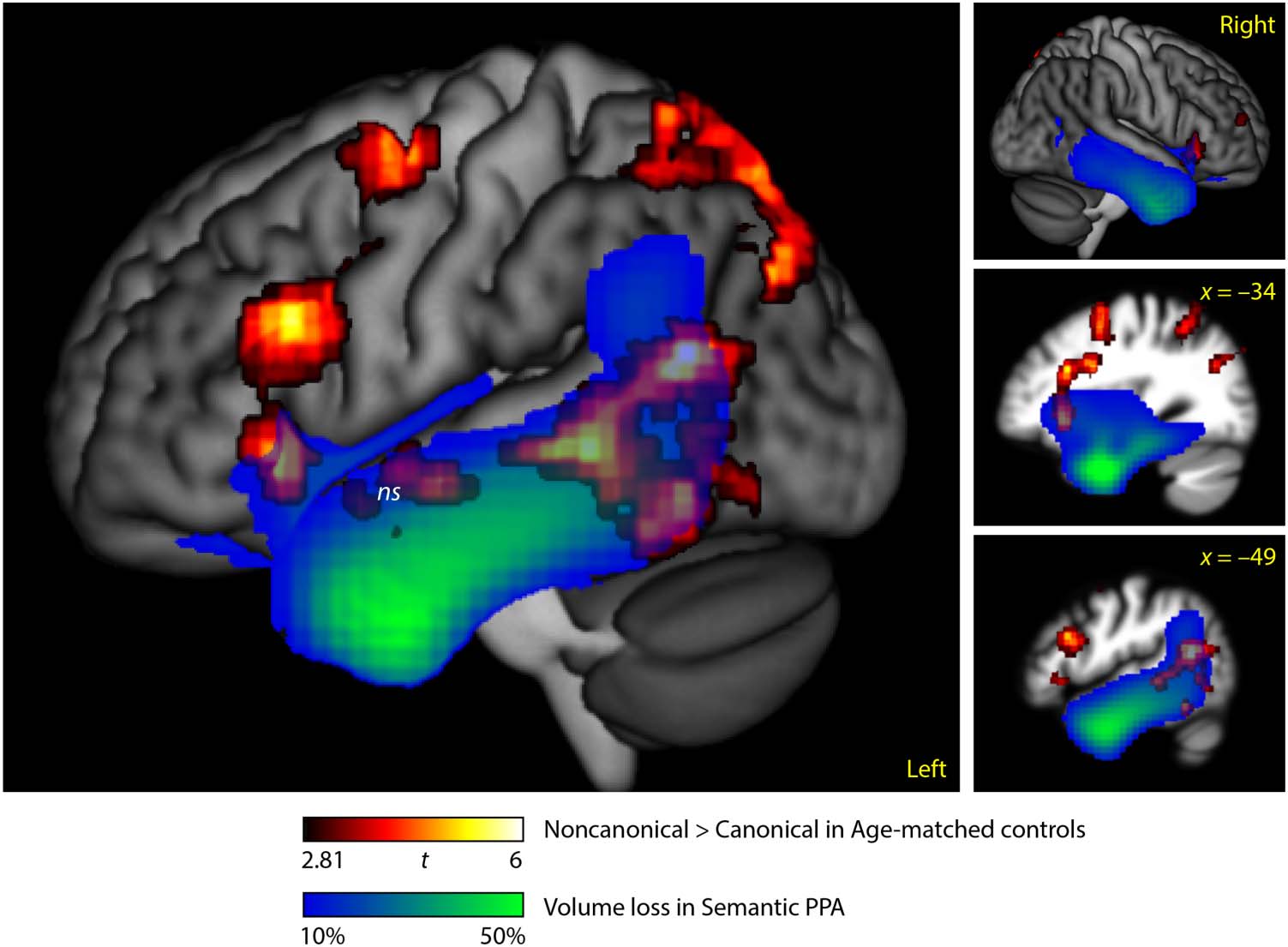
Overlap between Brain Regions Modulated by
Syntactic Complexity in Controls and Atrophy
in Semantic PPA Patients
There was extensive overlap between regions modulated
by syntactic complexity in controls and atrophic regions
in semantic PPA. In particular, the left posterior temporal
regions modulated in controls were largely atrophic, as
was the nonsignificant cluster in the left anterior STS.
There was also overlap in the left and right anterior insula
(Figure 2).
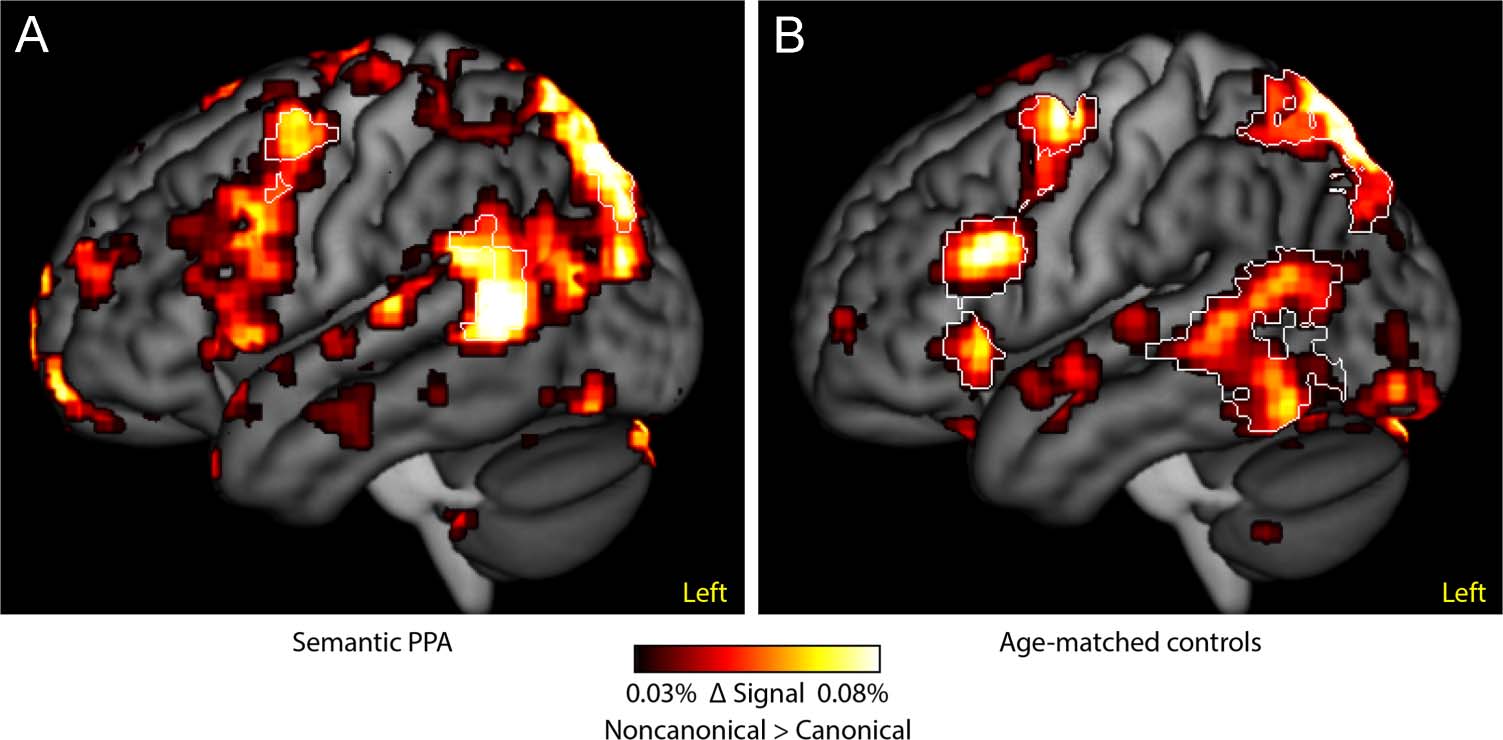
Comparison of Brain Regions Modulated by
Syntactic Complexity in Semantic PPA Patients
and Controls
The regions modulated by syntactic complexity were
strikingly similar in semantic PPA patients and controls
(Figure 3). Unthresholded effect size maps are presented
to depict the broad similarities in the regions activated
(Figure 3, hot). Similar to the control group, the regions
significantly modulated by syntactic complexity in seman-
tic PPA patients included left inferior frontal and poste-
rior temporal regions (Figure 3, white outlines; Table 3).
When the two groups were compared directly, there
were no significant differences between semantic PPA
patients and controls in the regions modulated by syn-
tactic complexity. This suggests that despite atrophy
extending to several regions involved in syntactic process-
ing in controls, these regions continued to show normal
functional activity in semantic PPA.
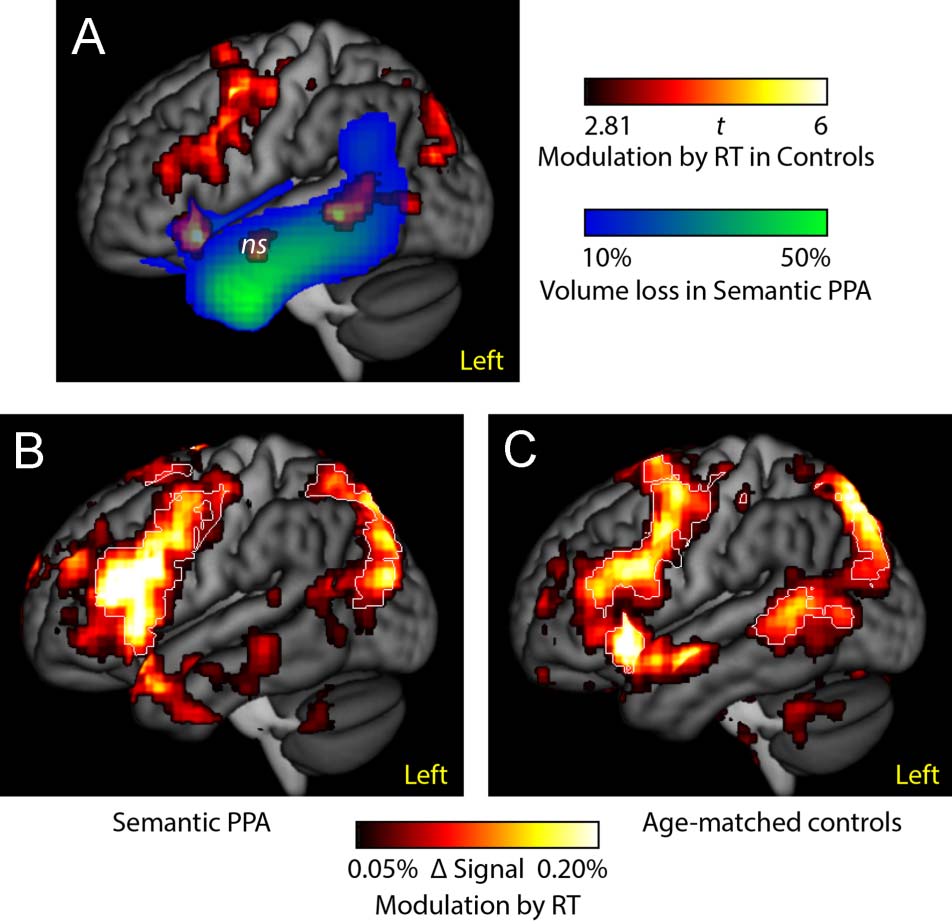
RT as a Proxy for Syntactic Processing
We carried out a second analysis in which RT was used as
a proxy for allocation of syntactic processing resources
(Figure 4). Brain regions modulated by RT in controls
were similar to those activated for noncanonical versus
canonical conditions and included left inferior frontal
and posterior temporal regions (Figure 4A, hot). Similar
to the syntactic complexity contrast, there were no ante-
rior temporal regions significantly modulated by RT,
but there was a nonsignificant cluster in the anterior STS
(MNI coordinates: −49, −5, −15; extent = 1640 mm3;
max t = 4.76; p = .13) that we included in the figure. Also
similar to the syntactic complexity contrast, posterior
temporal and insular regions overlapped with atrophy in
semantic PPA (Figure 4A, blue-green).
The regions modulated by RT were similar in semantic
PPA patients and controls. Unthresholded effect size
maps are presented to depict the broad similarities in
Figure 1. Behavioral data obtained during scanning. Accuracy (A) and
RT (B) on the syntactic comprehension task for semantic PPA patients
(red) and age-matched controls (blue). Boxes = interquartile range;
whiskers = range not including outliers; circles = outliers; horizontal
lines = medians; crosses = means. *p < .05; **p < .01; ***p < .001.
more extensive anteriorly than posteriorly (Figure 2, blue-
green). The entire left temporal lobe was atrophic, in-
cluding lateral and medial aspects; atrophy extended
posteriorly as far as the angular gyrus. In the right temporal
lobe, atrophy was somewhat less extensive: most of the
STG was spared, and the atrophy did not extend as far
posteriorly. Ventromedial prefrontal cortex and the insula
were also atrophic bilaterally.
Brain Regions Modulated by Syntactic Complexity
in Controls
Brain regions modulated by syntactic complexity in con-
trols (i.e., more active for noncanonical than canonical
conditions) included the left dorsal posterior IFG, inferior
frontal sulcus, precentral gyrus and anterior insula, and the
left posterior STS and adjacent MTG and ITG (Figure 2,
hot; Table 3). No anterior temporal regions were signifi-
cantly modulated by syntactic complexity; however, there
was a nonsignificant cluster in the anterior STS/STG (ex-
tent = 1392 mm3; p = .24) that we included in the figure
and table because of our specific questions related to this
976
Journal of Cognitive Neuroscience
Volume 26, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. Overlap between
regions modulated by syntactic
complexity in controls and
atrophic regions in semantic
PPA. Regions activated for
noncanonical versus canonical
sentences in controls (n = 24,
voxelwise p < .005, corrected
p < .05; hot) and regions with
tissue loss of 10% or greater in
semantic PPA patients (n = 20;
blue-green).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the regions activated (Figure 4B and C, hot). As in con-
trols, left inferior frontal and posterior temporal regions
were significantly modulated by RT in the patients (Fig-
ure 4B, white outlines).
When the two groups were compared directly,
there were no significant differences between semantic
PPA patients and controls in the regions modulated
by RT.
Table 3. Brain Regions Modulated by Syntactic Complexity in Controls and Semantic PPA Patients
MNI Coordinates
Brain Region
x
y
z
Extent (mm3) Max t
p
Modulated by Syntactic Complexity in Controls
Left posterior IFG (pars opercularis) and anterior insula −39
Left precentral gyrus
Left posterior STS, MTG, and ITG
Left anterior STS and STG
Left superior parietal lobule
Bilateral pre-SMA
Right anterior insula
−36
−53
−62
−19
0
39
Modulated by Syntactic Complexity in Semantic PPA Patients
Left precentral gyrus
Left posterior STS
Left superior parietal lobule
Left precuneus
MNI coordinates for clusters are centers of mass.
−37
−60
−28
−10
19
−1
−51
−3
−66
14
27
1
−50
−82
−54
16
56
3
−10
49
53
2
50
12
44
44
11984
4792
13256
1392
16664
3112
3568
1944
2376
3784
3408
5.58
4.95
6.40
4.81
5.43
4.37
4.03
4.52
4.67
6.49
4.74
< .001
.001
< .001
.24 (ns)
< .001
.033
.017
.048
.035
.019
.032
Wilson et al.
977
Figure 3. Effect size maps of
regions modulated by syntactic
complexity in semantic PPA (A)
and controls (B). Uncorrected
signal change is plotted to
illustrate the similarities
between activation patterns
in the two groups. The white
outlines indicate regions that
were statistically significant in
each group (voxelwise p < .005,
corrected p < .05). When the
groups were compared directly,
there were no significant
differences between them.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
Comparison of Brain Regions Activated or
Deactivated for Sentence Comprehension
versus Rest in Semantic PPA Patients
and Controls
We next examined the regions that were activated for the
sentence comprehension task relative to rest in the two
groups (Figure 5). Unthresholded effect size maps are
presented showing activations (Figure 5A and B, hot)
and deactivations (Figure 5A and B, cool), along with
statistical significance (Figure 5A and B, white and cyan
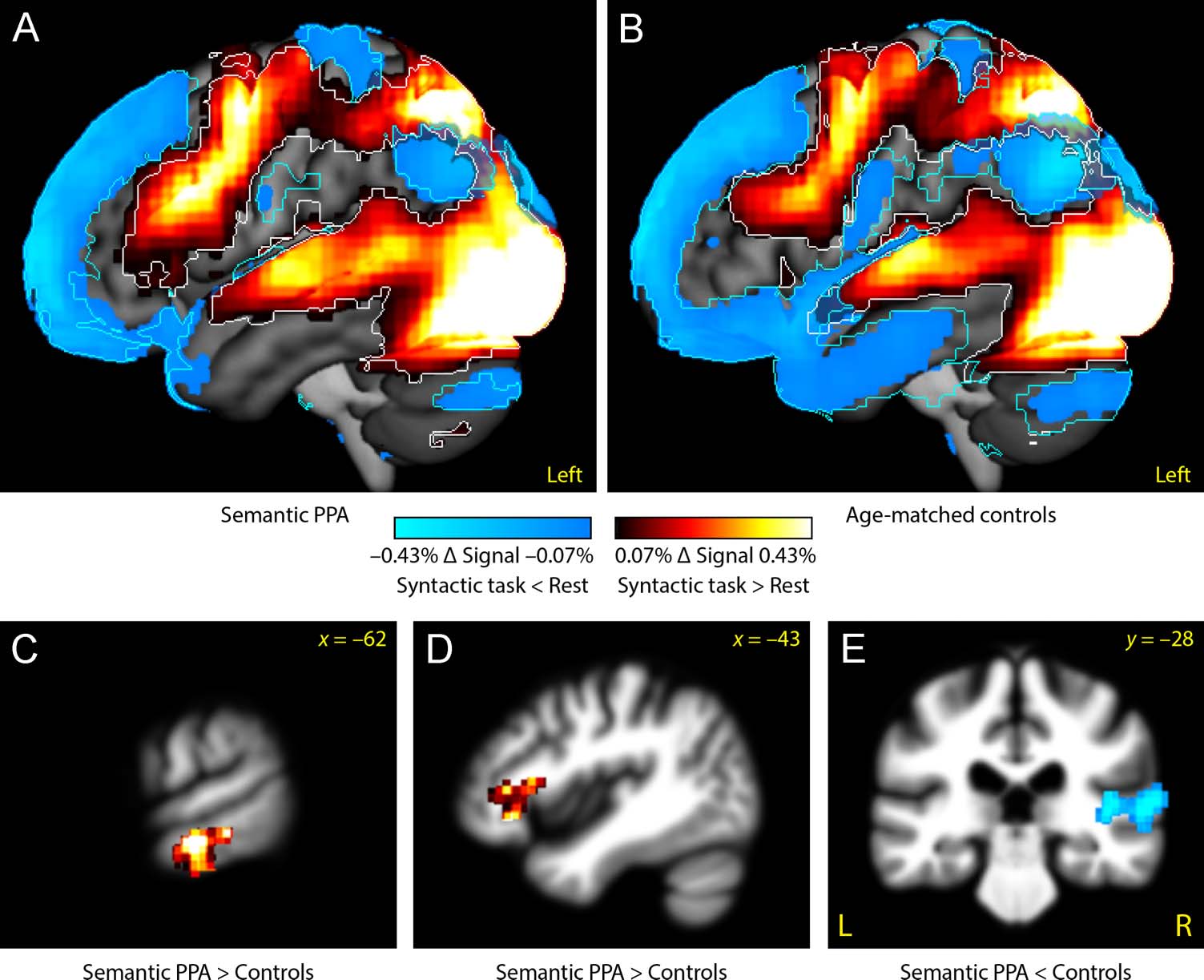
outlines). There were no activations in the ATL (anterior
to auditory regions), and surprisingly, the ventral ATL
was robustly deactivated in controls.
Although broad similarities between the two groups
can be readily appreciated, there were also three regions
that differed significantly when the two groups were
directly compared (Table 4). First, semantic PPA patients
did not show the ventral ATL deactivation that was
observed in controls (Figure 5C); the region where this
difference was observed was atrophic in the semantic
PPA patients (compare Figure 2A). Second, left inferior
frontal activation extended more anteriorly in semantic
PPA patients than in controls (Figure 5D); this region
was not atrophic (compare Figure 2A). Third, there was
less activation in the right STG in patients than in con-
trols (Figure 5E); this region was also not atrophic.
ROI Analyses
In nine left hemisphere ROIs, we investigated the distribu-
tion of tissue volume, modulation of signal by syntactic
processing, modulation of signal by RT, and signal change
for sentence comprehension relative to rest (Figure 6).
Semantic PPA patients showed significant volume loss in
all temporal lobe regions and the insula, but not in any of
the lateral frontal regions (Figure 6A). No regions differed
between patients and controls in terms of modulation by
syntactic complexity (Figure 6B). However, for modula-
tion by RT, the posterior STS differed between groups:
This region was modulated more in controls than in se-
mantic PPA patients ( p = .038; Figure 6C). Signal change
for sentence comprehension relative to rest differed
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
Table 4. Differences between Semantic PPA Patients and Controls in Functional Activations for the Sentence Comprehension Task
versus Rest
Brain Region
Semantic PPA > Control S
MNI Coordinates
X
y
z
Extent (mm3)
Max t
pag
Left IFG (pars triangularis and orbitalis)
Left anterior MTG
−48
−60
26
−19
9
−22
3128
6024
4.47
5.18
.017
.002
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Semantic PPA < Controls
Right STG
MNI coordinates for clusters are centers of mass.
56
−30
8
10872
5.94
< .001
978
Journal of Cognitive Neuroscience
Volume 26, Number 5
between groups in the two ROIs that had been defined
based on that contrast—the anterior MTG and the IFG
pars triangularis—and also in the anterior STS, which
was activated more in semantic PPA patients than controls
( p = .021 (Figure 6D). This region is adjacent to the ante-
rior MTG region that failed to deactivate in patients, so this
difference likely reflects extension of deactivation into the
anterior STS in some controls, but not in patients.
We next investigated, in semantic PPA patients only,
whether atrophy predicted accuracy, whether atrophy
predicted any of the three functional measures, and
whether any of the three functional measures predicted
accuracy (Figure 7).
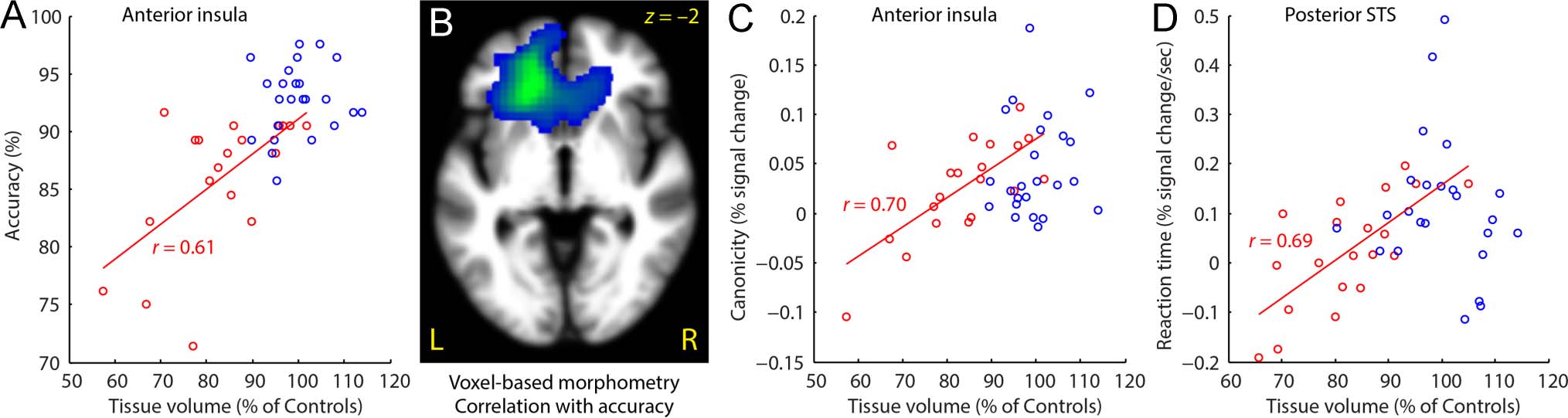
The only ROI where atrophy predicted accuracy was the
left anterior insula, where patients with greater volume
loss were less accurate (r = 0.61, p = .043; Figure 7A).
On the basis of that finding, we also carried out a whole-
brain voxel-based morphometry analysis to determine
where atrophy was associated with decreased accuracy,
and we found that tissue loss not only in the left anterior
insula, but throughout left frontal cortex, was associated
with decreased accuracy ( p = .002; Figure 7B).
There were two instances in which atrophy was pre-
dictive of functional imaging measures. In the left anterior
insula, patients with more atrophy showed less modula-
tion by syntactic complexity (r = 0.70, p = .021; Fig-
ure 7C), and in the left posterior STS, patients with more
atrophy showed less modulation by RT (r = 0.69, p = .012;
Figure 7D).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Wilson et al.
979
Figure 4. Analysis with RT as a proxy for syntactic processing.
(A) Overlap between regions modulated by RT in age-matched
controls (hot) and atrophic regions in semantic PPA (blue-green).
(B) Effect size map of regions modulated by RT in semantic PPA.
(C) Effect size map of regions modulated by RT in controls. The
white outlines indicate regions that were statistically significant in
each group (voxelwise p < .005, corrected p < .05). When the
groups were compared directly, there were no significant differences
between them.
Figure 5. Activations for the
syntactic task relative to rest.
(A) Effect size map of regions
activated for task versus rest
in semantic PPA. (B) Effect
size map of regions activated
for task versus rest in semantic
PPA. Uncorrected signal change
is plotted to illustrate the
similarities between activation
patterns in the two groups. The
white and cyan outlines indicate
regions that were statistically
activated (white) or deactivated
(cyan) in each group (voxelwise
p < .005, corrected p < .05).
(C) When the groups were
compared directly, there was
greater activation for semantic
PPA patients than controls in
the left anterior MTG, reflecting
deactivation in controls but not
patients. (D) There was greater
activation for semantic PPA
patients than controls in the
left IFG. (E) There was less
activation for semantic PPA
patients than controls in the
right STG.
Figure 6. ROI analyses for nine regions shown projected onto the cortical surface in the inset. (A) Tissue volume as a percentage of the control mean
for semantic PPA patients (red) and age-matched controls (blue). (B) Signal change for processing noncanonical sentences relative to canonical sentences.
(C) Signal change per second of RT. (D) Signal change for sentence comprehension task versus rest. Boxes = interquartile range; whiskers = range not including
outliers; circles = outliers; horizontal lines = medians; crosses = means. *p < .05; **p < .01; ***p < .001 (corrected for multiple comparisons, FDR).
There were no regions in which functional activity was
predictive of accuracy, although prior to correction for
multiple comparisons, there was some indication that,
in the left anterior insula, modulation by syntactic com-
plexity ( p = .056, uncorrected) and modulation by RT
( p = .032, uncorrected) were predictive of accuracy.
DISCUSSION
The overall goal of this study was to investigate the role of
the ATL in sentence processing by identifying the neural
correlates of syntactic processing in semantic PPA, a clini-
cal syndrome in which the ATL is damaged yet syntactic
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 7. Correlations between atrophy, behavior, and functional activation. (A) Tissue volume in the left anterior insula was predictive of accuracy
on the sentence comprehension task in semantic PPA patients. (B) Voxel-based morphometry revealed that left frontal atrophy was predictive of
accuracy on the sentence comprehension task. (C) Tissue volume in the left anterior insula was predictive of the difference in functional activity
between noncanonical and canonical sentences in semantic PPA patients. (D) Tissue volume in the left posterior STS was predictive of the extent
to which this region was modulated by RT in semantic PPA patients. Note that all correlations reflect only the semantic PPA group (red); data for
controls (blue) are shown for reference.
980
Journal of Cognitive Neuroscience
Volume 26, Number 5
processing is largely spared. We aimed to address two
specific questions. The first question was, which regions
normally involved in syntactic processing are damaged
in semantic PPA? We found that, in controls, left inferior
frontal and left posterior temporal regions were func-
tionally modulated by syntactic processing (quantified in
terms of syntactic complexity or trial-by-trial RT). Anterior
temporal regions were not consistently modulated. In the
semantic PPA group, atrophy was most severe in the ATLs
but extended to the posterior temporal regions involved
in syntactic processing. Atrophy also overlapped with
regions involved in syntactic processing in controls in the
anterior insula bilaterally.
The second question was whether spared syntactic pro-
cessing in semantic PPA depends on preserved functional-
ity of intact regions, preserved functionality of atrophic
regions, or compensatory functional reorganization. We
found that the brain regions modulated by syntactic pro-
cessing in semantic PPA were similar in most respects to
those observed in controls; in particular, there were no
significant differences in whole-brain analyses comparing
patients and controls in the regions modulated by syn-
tactic complexity or RT. This suggests that spared syntactic
processing in semantic PPA depends on preserved func-
tionality of intact regions (lateral frontal regions) as well
as preserved functionality of moderately atrophic regions
(posterior temporal cortex and the anterior insula).
Although there were some significant differences in func-
tional activity between semantic PPA patients and controls
(discussed below), there were no patterns suggestive of
functional reorganization, that is, no novel regions were
modulated by syntactic processing in semantic PPA.
The Role of the ATL in Sentence Processing
In controls and in semantic PPA patients, the ATL was only
inconsistently modulated by syntactic complexity and RT
(Figure 6), so no ATL regions were significantly activated
in either controls or semantic PPA patients. This is con-
sistent with prior studies of syntactic processing, which
have shown that frontal and posterior temporal regions
are activated for syntactically complex conditions relative
to simpler ones, but not the ATL (e.g., Bornkessel, Zysset,
Friederici, von Cramon, & Schlesewsky, 2005; Stromswold,
Caplan, Alpert, & Rauch, 1996).
Because the ATL is not modulated by syntactic com-
plexity, yet is activated for sentences relative to word lists,
some authors have raised the possibility that the ATL
may be involved in early stages of parsing and syntactic
structure-building that may not differ as a function of syn-
tactic complexity (Humphries et al., 2006; Friederici &
Kotz, 2003). In our view, it would be surprising if the
processes of reactivation and/or reanalysis that take place
in complex sentence processing did not make additional
demands on basic parsing mechanisms. Besides this theo-
retical consideration, there are two respects in which our
data are inconsistent with a role for the ATL in basic syn-
tactic structure-building. First, the semantic PPA patients
showed severe atrophy of the ATL, including almost 40%
volume loss in a temporal pole ROI that was defined based
on coordinates from the relevant literature. Although this
atrophy was sufficient to cause severe lexical semantic
deficits, syntactic processing was largely spared, with a
mean accuracy of 86.1% correct. The difference in accuracy
between patients and controls was just 7%, and although
this difference was statistically significant, its small mag-
nitude confirmed that syntactic processing is largely spared
in semantic PPA, consistent with previous literature (Wilson
et al., 2011; Cotelli et al., 2007; Grossman et al., 2005;
Gorno-Tempini et al., 2004; Hodges et al., 1992; Warrington,
1975; see Wilson et al., 2012, for a review). Second, the
ATL was not activated for sentence comprehension rela-
tive to rest in the controls or semantic PPA patients. If the
ATL were involved in basic syntactic structure-building,
we would have expected activation in all seven condi-
tions, because even canonical sentences require syntactic
structure-building.
Not only was the ATL not activated by our sentence
comprehension task relative to rest, but in the control
group, the ventral ATL was robustly deactivated. This
was a surprising finding, because most of the previous
studies comparing sentences and word lists have re-
ported positive ATL activation for sentences compared
with rest (Rogalsky & Hickok, 2009; Humphries et al.,
2001, 2005, 2006; Xu et al., 2005; Friederici et al., 2000;
Stowe et al., 1999; Mazoyer et al., 1993). The deactivation
of the ATL in our study is almost certainly a consequence
of our sentence-to-picture matching task. The ventral
ATL is one of the set of regions consistently deactivated
for attention-demanding cognitive tasks relative to rest
(Shulman et al., 1997). Why then has the ATL not shown
deactivation in previous sentence processing studies? A
likely explanation is that the ATL is an important node
of the semantic system (Binder, Desai, Graves, & Conant,
2009), and its deactivation in attention-demanding cog-
nitive tasks reflects reduced conceptual processing (Binder
et al., 1999). Many previous sentence processing studies
used passive listening (e.g., Humphries et al., 2005), very
simple tasks (e.g., Vandenberghe et al., 2002), or explicitly
semantic tasks (Humphries et al., 2006), all of which likely
promote semantic processing of the stimuli.
Intriguingly, the ventral ATL was not deactivated for sen-
tence comprehension relative to rest in the semantic PPA
patients. This indicates that the ATL is not just atrophic
in semantic PPA, but it is also functionally abnormal. If
our speculation is correct that the ATL deactivation in
controls reflects a reduction of resting state conceptual
processing, this lack of deactivation could be taken to
imply reduced resting state conceptual processing in
semantic PPA, which would be consistent with the funda-
mentally semantic nature of this syndrome.
On the basis of the arguments above, the role of the
ATL in sentence processing is unlikely to relate to syn-
tactic structure-building. Why then is the ATL so reliably
Wilson et al.
981
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
activated for sentences relative to word lists? As many
authors have suggested, we believe the role of the activa-
tion of the ATL in these contrasts reflects combinatorial
semantic processing (Vandenberghe et al., 2002). How-
ever, it is important to define what is meant by “combina-
torial semantic processing,” because syntactic processing
is also combinatorial: building a syntactic structure for a
sentence tells us “who did what to whom.” That is, build-
ing a syntactic structure relates the words in the sentence
to one another, connecting determiners to nouns, noun
phrases to slots in verb argument structure, and so on.
An insightful illustration of combinatorial semantic pro-
cessing that clearly distinguishes it from combinatorial
syntactic processing is provided by Humphries, Binder,
Medler, and Liebenthal (2007), drawing on earlier work
from Barsalou (1982). They note that, in the sentence
The shipwreck victim survived by clinging to a basketball,
the meaning of basketball takes on a crucial aspect of
meaning that is absent in isolation: The property of basket-
balls that they float is made salient. In another example,
The man on vacation lost a bag and a wallet, the word
bag in isolation has many possible meanings (e.g., garbage
bag, tea bag, etc.), but the sentential context refines the
meaning to suitcases, duffel bags, and the like.
In support of the view that combinatorial seman-
tic processing explains ATL activation for sentences,
Vandenberghe et al. (2002) reported that a subregion of
the ATL showed more activity for sentences than word
lists only when the sentences were semantically coherent.
Pallier et al. (2011) reported that the ATL was modulated
by syntactic constituent size only when stimuli contained
real words and not when they contained pseudowords.
This appears to contradict previous studies showing
activation for syntactic structure even with pseudowords
(Humphries et al., 2006; Friederici et al., 2000; Mazoyer
et al., 1993). The discrepancy between these studies
almost certainly reflects task differences. For instance,
Humphries et al. (2006) asked participants to rate the
meaningfulness of stimuli on a 4-point scale. We speculate
that being asked to rate the meaningfulness of pseudo-
word sentences leads participants to attempt to infer
meanings for pseudowords, possibly based on formal
similarities to real words, and to attempt to relate the
pseudowords to one another. Consider, for instance,
Twas brillig, and the slithy toves, the first line of The
Jabberwocky. The syntactic structure implies that slithy
is an adjective, and lexical neighbors such as sly, slimy,
slippery, and sleazy come to mind. These tend to give
rise to the impression that whatever toves are; they are
probably not very nice. Indeed later in Through the Look-
ing Glass, Humpty Dumpty tells Alice that slithy is a port-
manteau of slimy and lithe and that toves are something
like lizards, something like badgers, and something like
corkscrews. We can speculate that when participants
make inferences about the meaning of slithy that restrict
the meaning of toves, they are engaging in combinatorial
semantic processing that relies on the ATL.
Further support for a semantic function of the ATL
comes from studies of discourse-level comprehension that
have shown greater activation of the ATL for sentences
that were linked together to form a coherent narrative than
sentences that bore no relation to one another (Xu et al.,
2005; Fletcher et al., 1995) and strong correlations in the
time course of ATL activity across participants listening to
the same narratives (Wilson, Molnar-Szakacs, & Iacoboni,
2008).
We noted above that the ATL has been implicated in two
aspects of language: lexical semantics (Patterson et al.,
2007) and sentence-level processing. The literatures re-
lated to these two seemingly distinct functions have rarely
intersected. However, if the sentence-level role of the
ATL involves combinatorial semantic processing as de-
fined above, then we can suggest that the role of the
ATL in sentence processing is not distinct from its role in
lexical semantics, but rather follows directly from it. When
lexical items are brought together in relation to one
another in sentences and in discourses, their meanings
are narrowed, expanded, shifted, and refined by the con-
texts in which they occur. If the core function of the ATL
is the representation and processing of lexical semantic
information, then it follows that it will be more activated
for sentences than word lists, and discourses than sen-
tences, because the more context there is, the greater
the extent to which lexical semantic representations can
be refined.
Our sentence comprehension task was deliberately
limited in lexical semantic content: There were only two
nouns, seven verbs, and three adjectives employed in
the entire set of stimuli. Furthermore, because it was a
sentence-to-picture matching task, the entities and actions
referred to by the lexical items were apparent in advance
of hearing each sentence. Under these conditions, little
to no combinatorial semantic processing is necessary or
perhaps even possible, which may explain why the ATL
was not modulated by syntactic complexity, or even acti-
vated for the sentence comprehension task relative to
rest, and why severe damage to the ATL in semantic PPA
has little effect on performance on this task.
Lateral Frontal Cortex: Preserved Functionality
of Structurally Intact Regions
Left inferior frontal regions were modulated by syntactic
complexity, modulated by RT, and activated for the sen-
tence comprehension task relative to rest in controls and
in patients with semantic PPA. Numerous functional im-
aging studies have shown inferior frontal regions to be
differentially recruited for the processing of syntactically
complex sentences relative to simpler ones (Bornkessel
et al., 2005; Stromswold et al., 1996).
Although there were no differences between patients
and controls in frontal regions modulated by syntactic
complexity or RT, activation for the sentence comprehen-
sion task relative to rest extended more anteriorly into
982
Journal of Cognitive Neuroscience
Volume 26, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the pars triangularis in patients with semantic PPA (Fig-
ure 5D). An almost identical region showed increased
activation for this contrast in nonfluent/agrammatic PPA
patients relative to controls (Wilson, Dronkers, et al.,
2010). Increased prefrontal activation has been observed
in many studies of normal aging as well as dementia and
other neurological conditions and may reflect domain-
general compensatory processes (Park & Reuter-Lorenz,
2009).
The preserved functionality of lateral frontal regions in
semantic PPA, in particular the posterior IFG, contrasts
with our previous findings in patients with the nonfluent/
agrammatic variant of PPA, in whom these regions showed
abnormal functional activity (Wilson, Dronkers, et al.,
2010). Taken together with those findings, our present
results suggest that spared syntax in semantic PPA depends
in part on preserved functionality in these structurally
intact lateral frontal regions.
extension of atrophy into posterior temporal cortex in
semantic PPA has minimal consequences for the func-
tional status of this region, but an alternative explanation
could be that posterior temporal regions are less critical
for syntactic processing than left frontal cortex.
Volume loss in the left anterior insula, as well as other
frontal regions, was associated with less accurate perfor-
mance on the sentence comprehension task (Figure 7A
and B). Volume loss in the anterior insula was also as-
sociated with less functional modulation by syntactic
complexity, and there was some indication that these
functional changes mediated the performance deficits.
The role of the anterior insula in this context likely re-
lates to domain-general cognitive control and attention
(Dosenbach et al., 2006). Some of the minimal syntactic
deficits observed in semantic PPA may reflect the exten-
sion of atrophy into the anterior insula and other frontal
regions important for cognitive and executive processes
that support syntactic processing.
Posterior Temporal Cortex and Anterior Insula:
Preserved Functionality of Atrophic Regions
Conclusion
The left posterior STS, MTG, and adjacent regions were
modulated by syntactic complexity, modulated by RT,
and activated for the sentence comprehension task rela-
tive to rest in controls and in patients with semantic
PPA. Posterior temporal regions have been implicated
in syntactic processing in functional neuroimaging (e.g.,
Friederici, Makuuchi, & Bahlmann, 2009; Bornkessel
et al., 2005) and neuropsychological (Dronkers et al.,
2004) studies and may play a central role in syntactic
structure-building (Bornkessel-Schlesewsky & Schlesewsky,
2013) or may constitute an interface between syntax and
the lexicon (Friederici et al., 2009; Snijders et al., 2009).
The preserved functionality of posterior temporal cortex
in patients is noteworthy because these regions showed
significant atrophy. Although volume loss was most severe
in the ATLs (∼40% volume loss), posterior temporal re-
gions were not spared; almost every patient had less tissue
in this region than the control mean, and mean volume
loss was approximately 20% (Figure 6A). This suggests
that spared syntactic processing in semantic PPA depends
in part on preserved functionality of moderately atrophic
left posterior temporal regions.
Although whole-brain analyses did not show any dif-
ferences between semantic PPA patients and controls in
posterior temporal cortex, the more sensitive ROI analy-
sis showed that there was less modulation by RT in the
posterior STS (Figure 6C) and that patients with more
extensive atrophy in this region showed less modulation
by RT (Figure 7D). These findings should be treated with
caution, however, because there was no such effect for
modulation by syntactic complexity (Figure 6B). It is
noteworthy that neither atrophy nor functional changes
in the posterior STS were correlated with accuracy (but
cf. Bright, Moss, Stamatakis, & Tyler, 2008; Peelle et al.,
2008). This is consistent with our interpretation that the
In summary, our results show that spared syntactic pro-
cessing in semantic PPA depends on preserved functional-
ity of structurally intact left frontal regions and moderately
atrophic left posterior temporal regions, but there is no
functional reorganization apparent as a consequence of
anterior temporal atrophy and dysfunction. We have
argued that the role of the ATL in sentence processing is
less likely to relate to syntactic structure-building and
more likely to relate to higher level processes such as
combinatorial semantic processing.
Acknowledgments
This research was supported in part by the National Institutes
of Health (NIDCD R03 DC010878, NINDS R01 NS050915, NIA
P50 AG03006, NIA P01 AG019724). We thank Nina Dronkers
for helpful discussions; Jenny Ogar for patient testing; Laura
Calverley for drawing the stimulus pictures; Max Besbris, Paul
Keselman, and Lara Stables for assistance with functional im-
aging; Matthew Growdon, Jung Jang, Michael Neylan, and Henry
Rodriguez for administrative assistance; all of the members of
the UCSF Memory and Aging Center who contributed to patient
evaluation and care; and the patients, caregivers, and volunteers
for their participation in our research.
Reprint requests should be sent to Stephen M. Wilson, De-
partment of Speech, Language and Hearing Sciences, University
of Arizona, P.O. Box 210071, Tucson, AZ 85721, or via e-mail:
smwilson@u.arizona.edu.
REFERENCES
Ashburner, J. (2007). A fast diffeomorphic image registration
algorithm. Neuroimage, 38, 95–113.
Ashburner, J., & Friston, K. J. (2005). Unified segmentation.
Neuroimage, 26, 839–851.
Barsalou, L. W. (1982). Context-independent and context-
dependent information in concepts. Memory & Cognition,
10, 82–93.
Wilson et al.
983
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Beckmann, C. F., & Smith, S. M. (2004). Probabilistic
Hodges, J. R., Patterson, K., Oxbury, S., & Funnell, E. (1992).
independent component analysis for functional magnetic
resonance imaging. IEEE Transactions on Medical
Imaging, 23, 137–152.
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L.
(2009). Where is the semantic system? A critical review
and meta-analysis of 120 functional neuroimaging studies.
Cerebral Cortex, 19, 2767–2796.
Binder, J. R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S. F.,
Rao, S. M., & Cox, R. W. (1999). Conceptual processing
during the conscious resting state: A functional MRI study.
Journal of Cognitive Neuroscience, 11, 80–93.
Bornkessel, I., Zysset, S., Friederici, A. D., von Cramon, D. Y.,
& Schlesewsky, M. (2005). Who did what to whom? The
neural basis of argument hierarchies during language
comprehension. Neuroimage, 26, 221–233.
Bornkessel-Schlesewsky, I., & Schlesewsky, M. (2013).
Reconciling time, space and function: A new dorsal-ventral
stream model of sentence comprehension. Brain and
Language, 125, 60–76.
Brennan, J., Nir, Y., Hasson, U., Malach, R., Heeger, D. J., &
Pylkkänen, L. (2012). Syntactic structure building in the
anterior temporal lobe during natural story listening.
Brain and Language, 120, 163–173.
Bright, P., Moss, H. E., Stamatakis, E. A., & Tyler, L. K.
(2008). Longitudinal studies of semantic dementia:
The relationship between structural and functional
changes over time. Neuropsychologia, 46, 2177–2188.
Cotelli, M., Borroni, B., Manenti, R., Ginex, V., Calabria, M.,
Moro, A., et al. (2007). Universal grammar in the
frontotemporal dementia spectrum: Evidence of a selective
disorder in the corticobasal degeneration syndrome.
Neuropsychologia, 45, 3015–3023.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers
and Biomedical Research, 29, 162–173.
Dosenbach, N. U., Visscher, K. M., Palmer, E. D., Miezin,
F. M., Wenger, K. K., Kang, H. C., et al. (2006). A core
system for the implementation of task sets. Neuron, 50,
799–812.
Dronkers, N. F., Wilkins, D. P., Van Valin, R. D., Jr., Redfern, B. B.,
& Jaeger, J. J. (2004). Lesion analysis of the brain areas involved
in language comprehension. Cognition, 92, 145–177.
Fletcher, P. C., Happe, F., Frith, U., Baker, S. C., Dolan, R. J.,
Frackowiak, R. S. J., et al. (1995). Other minds in the brain:
A functional imaging study of “theory of mind” in story
comprehension. Cognition, 57, 109–128.
Friederici, A. D., & Kotz, S. A. (2003). The brain basis of
syntactic processes: Functional imaging and lesion studies.
Neuroimage, 20(Suppl. 1), S8–S17.
Friederici, A. D., Makuuchi, M., & Bahlmann, J. (2009). The
role of the posterior superior temporal cortex in sentence
comprehension. NeuroReport, 20, 563–568.
Friederici, A. D., Meyer, M., & von Cramon, D. Y. (2000).
Auditory language comprehension: An event-related fMRI
study on the processing of syntactic and lexical information.
Brain and Language, 75, 289–300.
Gorno-Tempini, M. L., Dronkers, N. F., Rankin, K. P., Ogar, J. M.,
Phengrasamy, L., Rosen, H. J., et al. (2004). Cognition
and anatomy in three variants of primary progressive
aphasia. Annals of Neurology, 55, 335–346.
Gorno-Tempini, M. L., Hillis, A. E., Weintraub, S., Kertesz, A.,
Mendez, M., Cappa, S. F., et al. (2011). Classification of
primary progressive aphasia and its variants. Neurology,
76, 1006–1014.
Grossman, M., Rhee, J., & Moore, P. (2005). Sentence
processing in frontotemporal dementia. Cortex, 41,
764–777.
Semantic dementia. Progressive fluent aphasia with
temporal lobe atrophy. Brain, 115, 1783–1806.
Holland, R., & Lambon Ralph, M. A. (2010). The anterior
temporal lobe semantic hub is a part of the language
neural network: Selective disruption of irregular
past tense verbs by rTMS. Cerebral Cortex, 20,
2771–2775.
Humphries, C., Binder, J. R., Medler, D. A., & Liebenthal, E.
(2006). Syntactic and semantic modulation of neural activity
during auditory sentence comprehension. Journal of
Cognitive Neuroscience, 18, 665–679.
Humphries, C., Binder, J. R., Medler, D. A., & Liebenthal, E.
(2007). Time course of semantic processes during
sentence comprehension: An fMRI study. Neuroimage,
36, 924–932.
Humphries, C., Love, T., Swinney, D., & Hickok, G. (2005).
Response of anterior temporal cortex to syntactic and
prosodic manipulations during sentence processing.
Human Brain Mapping, 26, 128–138.
Humphries, C., Willard, K., Buchsbaum, B., & Hickok, G.
(2001). Role of anterior temporal cortex in auditory
sentence comprehension: An fMRI study. NeuroReport,
12, 1749–1752.
Kapur, N., Barker, S., Burrows, E. H., Ellison, D., Brice, J.,
Illis, L. S., et al. (1994). Herpes simplex encephalitis: Long
term magnetic resonance imaging and neuropsychological
profile. Journal of Neurology, Neurosurgery, and
Psychiatry, 57, 1334–1342.
Kelly, R. E., Jr., Alexopoulos, G. S., Wang, Z., Gunning, F. M.,
Murphy, C. F., Morimoto, S. S., et al. (2010). Visual inspection
of independent components: Defining a procedure for
artifact removal from fMRI data. Journal of Neuroscience
Methods, 189, 233–245.
Kho, K. H., Indefrey, P., Hagoort, P., van Veelen, C. W. M.,
van Rijen, P. C., & Ramsey, N. F. (2008). Unimpaired
sentence comprehension after anterior temporal cortex
resection. Neuropsychologia, 46, 1170–1178.
Kramer, J. H., Jurik, J., Sha, S. J., Rankin, K. P., Rosen, H. J.,
Johnson, J. K., et al. (2003). Distinctive neuropsychological
patterns in frontotemporal dementia, semantic dementia,
and Alzheimer disease. Cognitive and Behavioral
Neurology, 16, 211–218.
Magnusdottir, S., Fillmore, P., den Ouden, D. B., Hjaltason, H.,
Rorden, C., Kjartansson, O., et al. (2013). Damage to left
anterior temporal cortex predicts impairment of complex
syntactic processing: A lesion-symptom mapping study.
Human Brain Mapping, 34, 2715–2723.
Mazoyer, B. M., Tzourio, N., Frak, V., Syrota, A., Murayama, N.,
Levrier, O., et al. (1993). The cortical representation of
speech. Journal of Cognitive Neuroscience, 5, 467–479.
Noppeney, U., & Price, C. J. (2004). An fMRI study of
syntactic adaptation. Journal of Cognitive Neuroscience,
16, 702–713.
Pallier, C., Devauchelle, A. D., & Dehaene, S. (2011). Cortical
representation of the constituent structure of sentences.
Proceedings of the National Academy of Sciences, U.S.A.,
108, 2522–2527.
Park, D. C., & Reuter-Lorenz, P. (2009). The adaptive brain:
Aging and neurocognitive scaffolding. Annual Review of
Psychology, 60, 173–196.
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do
you know what you know? The representation of semantic
knowledge in the human brain. Nature Reviews
Neuroscience, 8, 976–987.
Peelle, J. E., Troiani, V., Gee, J., Moore, P., McMillan, C.,
Vesely, L., et al. (2008). Sentence comprehension and
voxel-based morphometry in progressive nonfluent aphasia,
984
Journal of Cognitive Neuroscience
Volume 26, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
semantic dementia, and nonaphasic frontotemporal
dementia. Journal of Neurolinguistics, 21, 418–432.
anatomical parcellation of the MNI MRI single-subject
brain. Neuroimage, 15, 273–289.
Penfield, W., & Roberts, L. (1959). Speech and brain-mechanisms.
Vandenberghe, R., Nobre, A. C., & Price, C. J. (2002). The
Princeton, NJ: Princeton University Press.
Rogalsky, C., & Hickok, G. (2009). Selective attention to
semantic and syntactic features modulates sentence
processing networks in anterior temporal cortex.
Cerebral Cortex, 19, 786–796.
Shulman, G. L., Fiez, J. A., Corbetta, M., Buckner, R. L.,
Miezin, F. M., Raichle, M. E., et al. (1997). Common blood
flow changes across visual tasks: II. Decreases in cerebral
cortex. Journal of Cognitive Neuroscience, 9, 648–663.
Snijders, T. M., Vosse, T., Kempen, G., Van Berkum, J. J.,
Petersson, K. M., & Hagoort, P. (2009). Retrieval and
unification of syntactic structure in sentence comprehension:
An fMRI study using word-category ambiguity. Cerebral
Cortex, 19, 1493–1503.
Snowden, J. S., Goulding, P. J., & Neary, D. (1989). Semantic
dementia: A form of circumscribed cerebral atrophy.
Behavioural Neurology, 2, 167–182.
Stowe, L. A., Paans, A. M., Wijers, A. A., Zwarts, F., Mulder, G.,
& Vaalburg, W. (1999). Sentence comprehension and
word repetition: A positron emission tomography
investigation. Psychophysiology, 36, 786–801.
Stromswold, K., Caplan, D., Alpert, N., & Rauch, S. (1996).
Localization of syntactic comprehension by positron
emission tomography. Brain and Language, 52,
452–473.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D.,
Crivello, F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
response of left temporal cortex to sentences. Journal of
Cognitive Neuroscience, 14, 550–560.
Warrington, E. K. (1975). The selective impairment of semantic
memory. Quarterly Journal of Experimental Psychology,
27, 635–657.
Wilson, S. M., Dronkers, N. F., Ogar, J. M., Jang, J., Growdon,
M. E., Agosta, F., et al. (2010). Neural correlates of syntactic
processing in the nonfluent variant of primary progressive
aphasia. Journal of Neuroscience, 30, 16845–16854.
Wilson, S. M., Galantucci, S., Tartaglia, M. C., & Gorno-Tempini,
M. L. (2012). The neural basis of syntactic deficits in primary
progressive aphasia. Brain and Language, 122, 190–198.
Wilson, S. M., Galantucci, S., Tartaglia, M. C., Rising, K., Patterson,
D. K., Henry, M. L., et al. (2011). Syntactic processing depends
on dorsal language tracts. Neuron, 72, 397–403.
Wilson, S. M., Henry, M. L., Besbris, M., Ogar, J. M., Dronkers,
N. F., Jarrold, W., et al. (2010). Connected speech production
in three variants of primary progressive aphasia. Brain,
133, 2069–2088.
Wilson, S. M., Molnar-Szakacs, I., & Iacoboni, M. (2008). Beyond
superior temporal cortex: Intersubject correlations in narrative
speech comprehension. Cerebral Cortex, 18, 230–242.
Worsley, K. J., Liao, C. H., Aston, J., Petre, V., Duncan, G. H.,
Morales, F., et al. (2002). A general statistical analysis for
fMRI data. Neuroimage, 15, 1–15.
Xu, J., Kemeny, S., Park, G., Frattali, C., & Braun, A. (2005).
Language in context: Emergent features of word, sentence,
and narrative comprehension. Neuroimage, 25, 1002–1015.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
5
2
6
9
/
7
5
0
/
1
9
9
7
4
0
7
/
3
1
1
7
1
8
o
1
c
2
n
8
_
4
a
/
_
j
0
o
0
c
5
n
5
0
_
a
p
_
d
0
0
b
5
y
5
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Wilson et al.
985