Nonspatial Cueing of Tactile STM Causes Shift of
Spatial Attention
Tobias Katus1, Søren K. Andersen1,2, and Matthias M. Müller1
Abstracto
■ The focus of attention can be flexibly altered in mnemonic re-
presentations of past sensory events. We investigated the neural
mechanisms of selection in tactile STM by applying vibrotactile
sample stimuli of different intensities to both hands, seguido
by a symmetrically shaped visual retro-cue. The retro-cue indi-
cated whether the weak or strong sample was relevant for sub-
sequent comparison with a single tactile test stimulus. Locations
of tactile stimuli were randomized, and the required response did
not depend upon the spatial relation between cued sample and
test stimulus. Selection between spatially segregated items in
tactile STM was mirrored in lateralized activity following visual
retro-cues (N2pc) and influenced encoding of task-irrelevant tac-
tile probe stimuli (N140). Our findings support four major con-
clusions. Primero, retrospective selection results in transient shifts
of spatial attention. Segundo, retrospective selection is functionally
dissociable from attention-based rehearsal of locations. Tercero,
selection mechanisms are linked across processing stages, como
attention shifts in STM influence encoding of sensory signals.
Cuatro, selection in tactile STM recruits attentional control
mechanisms that are, at least partially, supramodal. ■
INTRODUCCIÓN
STM defines the capability to maintain and manipulate
previously perceived information in mind. The content of
STM is subject to attentional influences and similar to the
perceptual domain; the focus of attention can be flexibly
altered in STM to allow for adaptive behavior (kuo, alimenta,
& Nobre, 2012; kuo, Rao, Lepsien, & Nobre, 2009;
Theeuwes, Belopolsky, & Olivers, 2009; Awh, Vogel, &
Oh, 2006; Lepsien & Nobre, 2006; Curtis & DʼEsposito,
2003; Griffin & Nobre, 2003; Awh & En Jonia, 2001). Espacial
selection in mnemonic representations is known to influ-
ence perceptual encoding of external signals ( Jha, 2002;
Awh, Anllo-Vento, & Hillyard, 2000), which is consistent
with the proposed functional and anatomical overlap
of neural mechanisms mediating perceptual attention
and STM (Gazzaley & Nobre, 2012; Theeuwes et al.,
2009; Sörös et al., 2007; Awh et al., 2006; Müller & Caballero,
2006; Naghavi & Nyberg, 2005; Pasternak & verdelee,
2005; Curtis & DʼEsposito, 2003; Awh & En Jonia, 2001).
There are two lines of evidence suggesting an involve-
ment of spatial attention in STM tasks: The attention-based
rehearsal account states that covert spatial orienting to
memorized locations supports the voluntary maintenance
of spatial information (Theeuwes et al., 2009; Awh et al.,
2006; Awh & En Jonia, 2001; sin embargo, see also Belopolsky
& Theeuwes, 2009a). The investigation of attention-based
rehearsal therefore requires the implementation of spatial
STM tasks, in which locations have to be memorized and,
1Universität Leipzig, 2Universidad de California, San Diego
de este modo, are directly task-relevant. A series of experiments using
probe designs examined the influence of attention-based
rehearsal on sensory encoding ( Jha, 2002; Awh et al.,
2000). In this context, probe refers to sensory events that
probe the deployment of spatial attention and do not re-
quire a behavioral response. The studies mentioned above
reported that, during retention, probes presented to mem-
orized locations were preferentially encoded, comparado
with probes at nonmemorized locations. Such links be-
tween STM and perception suggest that spatial selection in-
volves neural processes that are common to both domains.
Además, spatial attention effects have also been
reported in STM paradigms in which spatial attributes were
not directly task-relevant, as memorization of locations for
subsequent comparison with test stimuli was not required.
These studies used retro-cues, presented after a set of
spatially segregated sample stimuli (Astle, Scerif, kuo, &
Nobre, 2009; Kuo et al., 2009; Griffin & Nobre, 2003). Como
a consequence, physiological activity following retro-cues
reflects control processes operating on mnemonic rather
than external stimulus representations (see Lepsien &
Nobre, 2006, para una revisión). Retro-cues that informed about
the target sample via cueing of nonspatial (Astle et al.,
2009; Kuo et al., 2009) or spatial attributes (Griffin & Nobre,
2003) caused lateralized electrophysiological activity in-
dicative for the directionality of attention shifts in visual
STM. This finding implies that STM preserves the spatial
configuration of eliciting peripheral events (Astle et al.,
2009; Kuo et al., 2009), even when memory for locations
is not explicitly instructed. Además, this observation
opens the intriguing possibility that retro-cues guiding
© 2012 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 24:7, páginas. 1596–1609
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
t
F
.
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
selection in STM (retrospective selection) trigger shifts of
spatial attention that might be unrelated to the voluntary
maintenance of locations (attention-based rehearsal). En
summary, retrospective selection and attention-based
rehearsal are both associated with spatial attention but
might represent functionally separable control processes,
en otras palabras, dissociable roles of spatial attention in
STM. Sin embargo, although all of the studies listed so far
investigated the visual modality, retrospective selection
has not been examined in touch up to today.
Evidence for a functional dissociation of retrospective
selection and attention-based rehearsal has been observed
in our previous somatosensory memory experiment
(Katus, andersen, & Muller, 2012). In a spatial STM task,
participants received bilateral sample pulses followed by
a retro-cue to control for exogenous orienting effects.
The retro-cue consisted of a color change of the symmetri-
cally shaped fixation cross, indicating whether the left- o
right-hand sample was relevant for subsequent compari-
son with a single delayed test stimulus. Los participantes fueron
asked to decide whether the test stimulus was applied
to the same location (finger) as the retrospectively cued
sample. The observed time course of spatial attention
effects was rather unusual for attention-based rehearsal
because the greatest spatial bias of encoding (indexed by
the N140) was evident for early probes, presented shortly
after the retro-cue. In vision, spatial STM experiments
consistently reported sustained attention-based rehearsal
efectos (Awh et al., 2000, 2006; Jha, 2002). One might argue
that such time-variant results are attributable to the tactile
nature of the task in our study. En cambio, we concluded that
retrospective selection triggers transient attention shifts
time-locked to the retro-cue, contributing to the sustained
impact of attention-based rehearsal. It is important to point
out that retrospective selection and attention-based re-
hearsal were confounded in the design of our previous
experiment because participants were asked to memorize
the retrospectively selected location. This confound is com-
mon to spatial STM paradigms, which therefore are problem-
atic approaches to examine retrospective selection apart
from possible attention shifts related to voluntary mainte-
nance of locations (attention-based rehearsal) y espacial
anticipation of test stimuli (more details in Katus et al., 2012).
The design of the task in this study differs from our
earlier experiment in three key aspects. First and most
importantly, participants were asked to memorize a non-
spatial attribute, namely stimulus intensity, y ubicaciones
of all tactile stimuli were unpredictable. This modification
circumvents the drawbacks of spatial STM paradigms and
rules out effects that might be attributable to attention-
based rehearsal or spatial anticipation. Si, nevertheless,
retro-cues elicited transient attention shifts, this finding
would substantiate the speculation that retrospective se-
lection engages a time-variant control process linked to
spatial attention yet dissociable from potentially con-
founding factors described above. Segundo, cue stimuli
were uninformative as to the location of the target (es decir.,
sample) relevant for delayed comparison with the test
stimulus. Tactile sample stimuli of different intensity were
simultaneously applied to both hands, and the color
change of the retro-cue indicated the category of the rele-
vant sample stimulus in terms of intensity (weak vs. strong
sample) rather than space (left vs. right hand). Tercero, nosotros
introduced a variable delay between samples and retro-
cue to counter a systematic contamination of the N2pc to
visual cues by neural responses to tactile sample stimuli.
Our hypotheses focused on physiological measures sen-
sitive to spatial selection between the left and right hands
(somatosensory N140, p.ej., Forster & Eimer, 2005) o
hemifields (N2pc, p.ej., Luck & Hillyard, 1994a).
Recent evidence suggests that the N2pc of the visual ERP
( VEP) reflects selection in visual STM, eso es, at a post-
perceptual stage (DellʼAcqua, Sessa, Toffanin, Luria, &
Jolicoeur, 2010; Kuo et al., 2009). In these studies, el
N2pc was measured in response to centrally presented
retro-cues that provided nonspatial information (p.ej., forma
and color) necessary to distinguish between targets and
distracters within a set of spatially segregated sample stim-
uli. The N2pc indexed spatially specific access of information
in visual STM, as this component reflected the location of
the target sample (p.ej., Kuo et al., 2009). Most intriguingly,
selection in tactile STM modulated the N2pc to visual retro-
cues in our earlier experiment. Such cross-modal links at a
relatively early stage might shed light on the neural origins
of the N2pc, suggesting contributions from neural pro-
cesses that are, at least to some degree, detached from fun-
damental characteristics of the accessed information.
Desafortunadamente, a decisive interpretation of the N2pc was
compromised in our previous study because of a contami-
nation by the P300 component to tactile sample stimuli.
The present somatosensory experiment sought to eluci-
date the neural mechanisms mediating retrospective selec-
tion between spatially segregated sample stimuli. Spatially
specific biasing of neural activity in the period after retro-
cues would point to an involvement of spatial attention in
a scenario designed to avoid shortcomings of spatial STM
paradigms, En particular, confounding factors such as
attention-based rehearsal and spatial anticipation of test
estímulos. We examined lateralized neural activity evoked by
the visual retro-cue. Recent STM experiments reported
modulations of the N2pc (DellʼAcqua et al., 2010; kuo
et al., 2009) and the anterior directing attention negativity
(ADAN; see Griffin & Nobre, 2003). Como consecuencia, semejante
lateralized components were assumed to mirror selection
in tactile STM. Segundo, attention shifts in STM influence
selection in perception (Theeuwes et al., 2009; Awh et al.,
2000, 2006; Jha, 2002; Awh & En Jonia, 2001). The somato-
sensory N140 is reliably affected by spatial selection be-
tween the left and right hands in the perceptual domain
(Forster & Eimer, 2004, 2005; Eimer, Forster, Fieger, &
Harbich, 2004; Zopf, Giabbiconi, Gruber, & Muller, 2004;
Eimer & Forster, 2003a; Michie, Bearpark, Crawford, &
Glue, 1987; Josiassen, Shagass, Roemer, Ercegovac, &
Straumanis, 1982; Desmedt & Robertson, 1977). Porque
Katus, andersen, and Müller
1597
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
/
t
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
retrospective selection was associated with transient at-
tention (Katus et al., 2012), we expected spatially spe-
cific encoding effects in a brief period after the retro-cue,
influencing the N140 to probes but not to test stimuli.
Además, we examined the decision-related P300
component in response to tactile test stimuli. The P300 is
greater for targets compared with distracters (Polich, 2007;
Desmedt & Debecker, 1979), and because participants were
asked to compare intensities, we expected enhanced P300
amplitudes when test stimuli were presented with the
memorized relative to nonmemorized intensity. The spatial
relation between cued sample and test stimulus did not
convey useful information for the task (intensity com-
parison) y, de este modo, should not influence the P300. If retro-
spective selection is dissociable from attention-based
rehearsal, electrophysiological measures (p.ej., N2pc, ADAN,
and N140) should mirror the location of the cued item,
even when memorized attributes belong to a nonspatial
feature dimension.
MÉTODOS
Participantes
Twenty participants took part in the experiment. Two
participants reported that they were unable to distinguish
between the samplesʼ intensities in more than approxi-
mately one third of trials. Because of the resulting low
actuación, we excluded these participants from statis-
tical analysis. A further participant ignored instructions
and placed index rather than middle fingers on the stim-
ulators during the last two blocks of the experiment. Seven-
teen participants remained in the sample (13 women, edad
range = 18–37 years, average = 25 años). All were right-
handed according to the Edinburgh Handedness Inventory
(Oldfield, 1971), reported no history of neurological illness,
and had normal or corrected-to-normal vision. Participantes
gave informed written consent and received payment or
course credit. The experiment was conducted in accor-
dance with the Declaration of Helsinki.
Experimental Setup
The experimental setup is illustrated in Figure 1A. Tactile
stimulation was provided by two miniature loudspeakers
(Ekulit Elektrotechnik, Ostfildern, Alemania) with cut-
out membranes. Participants were asked to rest the distal
phalanx of each middle finger on the inner plateau of a
stimulator. This even part in the center of the speakerʼs
basket forms the upper structural element of the magnetic
core. Participantsʼ index and ring fingers were located
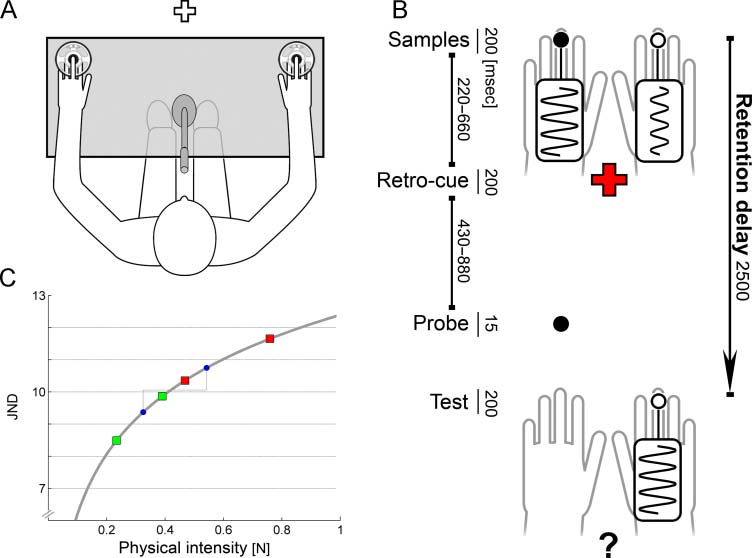
Cifra 1. Experimental
paradigma. (A) Experimental
setup: Tactile stimuli were
delivered to the middle fingersʼ
distal phalanges; both hands
were covered from sight.
Visual stimuli were shown
on a monitor that could be
seen through a window in
the recording chamber.
Retro-cues consisted of color
changes of the white fixation
cross to green or red.
(B) Stimulation procedure:
Tactile sample stimuli with
different intensities were
simultaneously applied to
both hands. The color of the
visual retro-cue informed
about the intensity of the
target sample, eso es, weak
versus strong. Participantes
were asked to compare
intensity of cued sample
and test, irrespective of
their spatial relation. En el
illustrated example, the red
retro-cue determined the
strong sample as target.
Tactile events were either spatially congruent (black dots) or incongruent (white dots) relative to the location of the target sample. El
exemplary trial required a “yes” response, because test and target had equal intensity. (C) Intensity scaling: Sample and test stimuli were
scaled according to the Weber–Fechner Law. For each trial, a reference was randomly drawn from a predefined range of physical intensities
(bounded by blue dots). The reference determined the intensity of both sample stimuli: Weak and strong samples were equidistantly below
and above the reference, in terms of perceived intensity ( y axis). In a given trial, sample stimuli differed by 1.7–2 JNDs. Throughout the
experimento, intensities of weak and strong samples ranged between the green and red squares, respectivamente. Test stimuli were presented
with the memorized intensity (50%) or differed by two JNDs (50%).
1598
Revista de neurociencia cognitiva
Volumen 24, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
t
.
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
beneath the basket, providing a gentle upward pressure.
Participants were encouraged to rest their middle fingers
comfortably on the center plateaus. They were asked to
neither lift the fingers upward nor press them against the
stimulator. Participantʼs hands and stimulators were cov-
ered from sight by towels. Stimulators were located on
additional towels on a table in front of the participantʼs
silla. Distance between both hands was kept constant at
80 cm. White noise was presented via headphones to
mask any sounds produced by the stimulators. Stimulators
were driven by a dual channel amplifier connected to a
digital-to-analog conversion card. Stimulation was con-
trolled by MATLAB (The MathWorks, Natick, MAMÁ).
Task Design and Stimulation Material
Stimulation Procedure and Task
Participants received bilateral sample stimuli with differing
intensities. Participants were instructed to selectively mem-
orize the intensity of a single retrospectively cued sample
(objetivo) for subsequent comparison with a test stimulus.
The target sample was determined by the categorical cue-
ing of a nonspatial stimulus attribute, namely intensity.
Spatial properties of all tactile stimuli were unpredictable
and without influence on the required response. Figura 1B
illustrates the stimulation sequence. A trial started with
simultaneous presentation of bilateral tactile sample stim-
uli. Between 220 y 660 msec after sample offset, el
fixation cross changed its color for 200 mseg, thereby serv-
ing as retro-cue. The retro-cue informed about the in-
tensity category (weak, strong) of the to-be-memorized
sample via an arbitrary color-mapping rule. A probe stim-
ulus was randomized to either hand between 430 y
880 msec after retro-cue offset. Participants had been
informed in advance that probes were task-irrelevant. A
unilateral test stimulus was presented to the left (50%)
or right (50%) hand at a fixed latency of 2500 mseg después
sample onset. Test stimuli were either presented with the
same physical intensity as the cued sample (50%) or scaled
to a nonmemorized intensity (50%). A question mark
prompted participants to respond verbally during the sub-
sequent 1500-msec period. Participants were instructed
to compare the intensity of the test stimulus with the retro-
spectively cued sample, regardless of their spatial relation.
Participants were asked to say the vowel “a” (correspond-
ing to a “yes” response) if intensities matched and “i” if
they differed (“no” response). The response period ended
when the question mark was replaced by the fixation
cross. Between 400 y 500 msec thereafter, the next trial
started.
The main experiment consisted of 560 trials divided
into eight blocks, aproximadamente 6 min and 30 sec each.
Feedback on average hit and false alarm rates was provided
after each block. Before recordings, three to five training
blocks of 50 trials were performed until stable performance
was reached. During the main experiment, Participantes
were instructed to avoid head and arm movements and
maintain fixation while the cross was present.
Tactile and Visual Stimuli
Mechanical tactile stimulation involved samples, probes,
and test stimuli. Sample and test stimuli consisted of
200-msec-long vibrotactile sinusoids, driven at 200 Hz.
The intensity of sample and test stimuli varied across ex-
perimental trials; the intensity scaling procedure will be
described below (Psychometric Scaling of Intensities sec-
ción). A sample pair consisted of a weak and strong stim-
ulus, simultaneously applied to the left and right hands.
The spatial configuration of sample pairs was unpredict-
capaz: Sample stimuli with a specific intensity (p.ej., strong)
were equiprobably presented to the left or right hand.
Unilateral test stimuli were randomized to the left (50%)
or right (50%) mano. Tactile probe stimuli consisted of
concise 15-msec pulses at 200 Hz, eso es, two successive
5-msec pulses separated by a 5-msec delay. Probes were
presented with fixed intensity of 0.84 N and randomized
to the left (50%) or right (50%) mano. Probes were not
delivered in 17% of the trials to reduce predictability of
the stimulation procedure.
Color change of the white fixation cross to either green
or red served as retro-cue. The symmetric retro-cue was
centrally presented on the monitor for 200 mseg. At a
given viewing distance of 140 cm, the retro-cue had a size
of 1.23° of visual angle. One of two mapping rules was
randomly assigned to each participant before the experi-
mento. Ten participants received the instruction to mem-
orize the weak versus strong sample stimulus when the
retro-cue turned green versus red, respectivamente. Para el
remaining seven participants, this mapping rule was re-
versed (es decir., red and green cued weak and strong samples,
respectivamente). The number of trials with green or red cues
was balanced for a particular spatial configuration of sam-
ple intensities. Within a given subject, the same color cued
tactile samples at the left (50%) y correcto (50%) manos.
Psychometric Scaling of Intensities
The scaling of tactile sample and test stimuli was based on
a psychometric function (see Figure 1C), which had been
approximated according to the Weber–Fechner Law with
help of four nonnaive participants (staff members) antes
the experiment. Parameters of the transfer function, semejante
as the Weber fraction and absolute threshold, were ob-
tained using QUEST ( watson & Pelli, 1983), a Bayesian
adaptive psychometric method. The scaling of sample
stimuli had to ensure that participants (a) were able to
discriminate between bilaterally presented samples in
terms of intensity and (b) memorized sensory signals
rather than abstract category labels. For a given trial, el
scaling of tactile stimuli was based on a reference inten-
sity randomly drawn from a range of physical intensities,
specifically 0.32–0.54 N (blue circles in Figure 1C). El
Katus, andersen, and Müller
1599
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
t
.
F
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
reference determined the intensities of both sample stim-
uli. Sample pairs differed between 1.7 y 2 just noticeable
diferencias ( JNDs) and were equidistant with respect to
the reference intensity, Por ejemplo, one JND below and
above the reference in terms of perceived intensity. Verde
and red squares in Figure 1C represent the total range of
weak and strong sample intensities across experimental
ensayos, respectivamente. With equal probability, test stimuli were
scaled to the intensity of the target or differed by approxi-
mately two JNDs. For the majority (75%) of nontarget test
estímulos, intensities were biased toward the uncued inten-
sity category: Weak nontargets exceeded the memorized
intensidad, whereas strong nontargets were set to a weaker
intensidad. This constraint sought to discourage partici-
pants to memorize the intensity of the distracter along with
the target.
Data Recording and Analysis
Recording of Electrophysiological Data and
Treatment of Artifacts
Participants were seated in a comfortable chair in a dimly
lit electrically shielded chamber. Brain activity was re-
corded at a sampling rate of 256 Hz from 64 Ag/AgCl elec-
trodes mounted in an elastic cap using an ActiveTwo
amplifier system (BioSemi, Ámsterdam, Los países bajos).
During recordings, all scalp channels were referenced to
an electrode next to POz. A bipolar outer canthus montage
(HEOG) monitored lateral eye movements. A bipolar
montage below and above the right eye (VEOG) grabado
vertical eye movements and blinks. Data processing was
performed off-line with the help of the EEGLab toolbox
(Delorme & Makeig, 2004) and custom-written MATLAB
routines. Continuous EEG data were digitally band-pass
filtered (0.5–40 Hz, Kaiser windowed finite impulse re-
respuesta). Somatosensory evoked potentials (SEPs) in re-
sponse to tactile probe and test stimuli were segmented
into 500-msec epochs, starting 100 msec before stimulus
onset and ending 400 msec after stimulus onset. VEPs elic-
ited by visual retro-cues were epoched into 800-msec
periods, starting 100 msec before cue onset and extending
700-msec postcue. A 100-msec prestimulus baseline was
subtracted from SEPs and VEPs.
Trials with horizontal eye movements exceeding 25 μV
(roughly corresponding to 2° of visual angle) were rejected
before further artifact correction procedures. The in-
fluence of eye blinks on the EEG signal was corrected
by means of independent component analysis (Campana &
Sejnowski, 1995). Independent components associated
with eye blinks are known to have stereotypical scalp
topographies and frequency characteristics (Delorme,
Sejnowski, & Makeig, 2007). Such components were iden-
tified by visual inspection and subtracted from the data
colocar. Después, epochs exceeding a voltage threshold
of ±150 μV were rejected. The remaining data were sub-
jected to SCADS (statistical control of artifacts in dense
array EEG/magnetoencephalography studies; see Junghöfer,
Elbert, Tucker, & Rockstroh, 2000). SCADS identified arti-
facts on the basis of statistical parameter distributions ob-
tained from a given data set. Spherical spline interpolation
of artifact-free electrodes was used to correct contaminated
signals. Epochs with more than 12 artifactual electrodes
were rejected. De término medio, 6%, 6.2%, y 4.2% de todo
epochs to cue, probe, and test stimuli were rejected, re-
spectively. These rejection rates did not differ between
experimental conditions. After artifact correction and rejec-
ción, the EEG signal was rereferenced to the arithmetic
mean of both mastoids. To ensure that data were not in-
fluenced by small but systematic eye movements, cual
might have escaped the above 25-μV threshold, HEOG
waveforms were separately averaged for trials in which
the retro-cue indicated the left- versus right-hand sample.
A paired t test running across sampling points postcue
revealed no difference between these trials during the
analyzed time windows.
Analysis of Electrophysiological Data
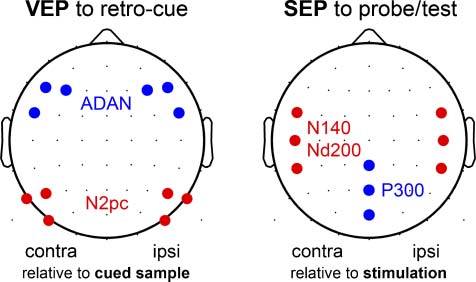
Mesa 1 summarizes latency range and electrode selection
used for the acquisition of specific VEP and SEP compo-
nents. The stated parameters were chosen in accordance
with previous findings with regards to N2pc (DellʼAcqua
et al., 2010; Kuo et al., 2009) and N140 (p.ej., Forster &
Eimer, 2005). Additional analyses assessed modulations
of the ADAN (see Forster, Sambo, & Pavone, 2009; Griffin
& Nobre, 2003; van Velzen, Forster, & Eimer, 2002); el
Mesa 1. Time Window and Electrode Selection for the Measurement of VEP and SEP Components
Eliciting Stimulus
Component
Electrodes
Latency Range (mseg)
Retro-cue ( VEP)
Retro-cue ( VEP)
Probe, prueba (SEP)
Probe, prueba (SEP)
Prueba (SEP)
N2pc
ADAN
N140
Nd200
P300
P7/8, P5/6, PO7/8
F3/4, F5/6, FC5/6
FC5/6, C5/6, CP5/6
FC5/6, C5/6, CP5/6
CPz, Pz, POz
250–310
350–700
124–156
184–216
260–340
Mean amplitudes in the stated latency ranges entered statistical testing. All components were measured at contralateral and ipsilateral ROIs, excepto
for the P300.
1600
Revista de neurociencia cognitiva
Volumen 24, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
t
.
F
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Nd200, a negative difference wave emerging at around
200-msec poststimulus (cf. Gillmeister & Forster, 2012;
Forster & Eimer, 2004; Eimer & Forster, 2003b); y el
P300 (Polich, 2007; Desmedt & Debecker, 1979).
Statistical analyses were conducted on the average of
three neighbored electrodes, as illustrated in Figure 2.
The design factor electrode cluster was included in the
analyses of VEP and SEP components, except for the
P300 that was measured at a single ROI. VEPs and SEPs
acquired at a particular electrode cluster were averaged
across both hands, because preliminary analyses of electro-
physiological and behavioral data revealed no significant
differences dependent on the location of the cued sample
stimulus. Mean amplitudes were calculated for the time
windows specified in Table 1 and submitted to repeated
measures ANOVAs. Statistical analyses included trials with
correct and incorrect behavioral responses because these
did not differ with regards to the modulation of physio-
logical measures under investigation. Within-subject con-
fidence intervals (CIs) in figures reflect the error terms
associated with particular effects of interest, independiente
of between-subject variance (see Jarmasz & Hollands,
2009; Loftus & Masson, 1994). Aquí, conditional means
with nonoverlapping CIs represent a significant effect ( pag <
.05) of the experimental manipulation specified in the
respective figure caption.
Lateralized components of the VEP.
Separate two-way
ANOVAs assessed lateralized VEP components to retro-
cues, such as N2pc and ADAN. The statistical design com-
prised the factors of Electrode Cluster (relative to cued
sample: contralateral, ipsilateral) and Cued Intensity
(weak, strong). Importantly, Electrode Cluster main ef-
fects reflect differential activity related to the location
of the target sample.
Figure 2. Electrode selection. Electrode clusters defined for statistical
assessment of VEPs (left) and SEPs (right). Three neighboring electrodes
were averaged for a particular recording cluster. Each VEP/SEP component
was measured at symmetrical contralateral and ipsilateral electrode
clusters, except for the P300 of the SEP. Contra = contralateral;
ipsi = ipsilateral.
SEP components elicited by probe stimuli. Modulations
of the SEP to task-irrelevant probe stimuli occurred dur-
ing the latency range of the N140 and Nd200. Separate
three-way ANOVAs tested the influence of the design fac-
tors Spatial Congruency (spatial relation with respect to
the cued sample: congruent, incongruent), Stimulation
Sequence (intensity of sample stimulus preceding the
probe: weak, strong), and Electrode Cluster. Stimulation
sequence served as a control factor for prior exposure to
weak versus strong stimulation at a given finger. Most im-
portantly, we hypothesized that selection in tactile STM
leads to spatially specific sensory encoding, reflected by
spatial congruency main effects.
SEP components elicited by test stimuli.
Separate three-
way ANOVAs assessed the SEP in response to task-relevant
test stimuli at latencies corresponding to the N140 and
Nd200. The statistical design comprised the factors of
Spatial Congruency (congruent, incongruent), Intensity
Congruency (target, distracter), and Electrode Cluster. In-
tensity congruency compared test stimuli with memorized
versus nonmemorized intensity. The P300 component was
obtained from a single recording cluster and assessed by a
two-way ANOVA with factors of Intensity Congruency and
Spatial Congruency.
Recording and Analysis of Behavioral Data
Vocal responses during the 1500-msec response pe-
riod were recorded by a microphone located in front of
the participant. Voice key routines written in MATLAB
code analyzed speech signals with respect to their for-
mant characteristics. Estimated resonance frequency
profiles of the first two formants were categorized by
means of multinomial logistic regression. Regression
parameters had been individually determined for each
participant in a calibration session at the beginning of
the experiment. Categorizations were checked again
off-line and corrected if necessary. “Yes” and “no” re-
sponses were associated with the vowels “a” and “i,” re-
spectively. A hit was defined as “yes” response, given
that the test stimulus was presented with the target inten-
sity. A correct rejection was considered as “no” response
following a distracter, that is, test with nonmemorized
intensity. Hit and correct rejection rates were submitted
to separate repeated-measures ANOVAs comprising
the factors of Cued Intensity (weak, strong), Probe Con-
gruency (congruent, incongruent, no probe), and Test
Congruency (congruent, incongruent). Note that con-
gruency referred to the spatial relation between a par-
ticular stimulus and a target sample. The location of
the cued sample (left vs. right hand) was not included
as a factor because preliminary analyses did not find
significant differences in hit as well as correct rejection
rates.
Katus, Andersen, and Müller
1601
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
o
7
c
9
n
4
_
0
a
/
_
j
0
o
0
c
2
n
3
4
_
a
p
_
d
0
0
b
2
y
3
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
RESULTS
Behavioral Data
Hit rates were higher when both the cued sample and test
stimulus had been presented to the same hand compared
with different hands (test congruency: F(1, 16) = 6.01,
p = .026). This effect was driven by responses to targets
of strong intensity and was absent for weak targets (Cued
Intensity × Test Congruency: F(1, 16) = 8.57, p = .010).
No further effects or interactions were observed in hit
rates. The analysis of correct rejection rates revealed a
different pattern. No significant effects or interactions
were found except for performance differences that de-
pended on the cued intensity category. Participants more
often responded correctly when the target intensity
was weak as compared with strong (cued intensity: F(1,
16) = 16.55, p = .001).
Electrophysiological Data
Lateralized Activity after Retro-cues ( VEP)
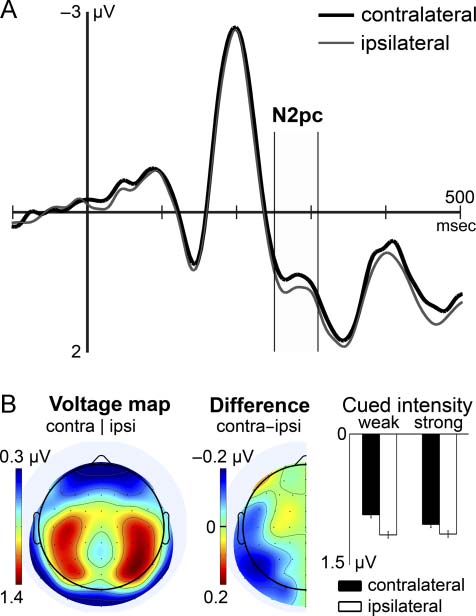
Figure 3A shows the VEP to visual retro-cues, recorded
at posterior scalp regions contralateral and ipsilateral to
the target sample. A highly significant N2pc modulation
mirrored the location of the retrospectively cued sample,
indicated by more negative amplitudes in the N2pc time
range contralateral to the target as compared with ipsi-
lateral (electrode cluster: F(1, 16) = 24.78, p < .001).
This effect generalized across retro-cues that directed
attention to weak versus strong samples, as an interaction
between electrode cluster and cued intensity was not
significant ( p > .1). The topographical extent of the
N2pc modulation is shown in the difference map, Higo-
ure 3B. Posterior lateralized activity was accompanied
by the ADAN, a differential negativity contralateral to
the cued sample (electrode cluster: F(1, 16) = 5.92,
pag = .027). There was no significant interaction of elec-
trode cluster and cued intensity in the ADAN time range
( p > .1). Tomados juntos, retro-cues elicited lateralized
activity over posterior and anterior scalp regions, cual
consistently reflected selection in tactile STM.
Cifra 3. (A) Grand mean VEP to retro-cues measured at posterior
scalp regions contralateral (thick black line) and ipsilateral (thin gray
line) to the retrospectively cued sample. Negative values above midline.
(B) Grand mean isocontour voltage map (left column), diferencia
map (middle), and mean amplitudes (bien) during the N2pc latency
range. The arrangement of data in the voltage map is equivalent to the
cueing of the right-hand sample. Contralateral and ipsilateral electrodes
correspond to left and right scalp regions, respectivamente. la diferencia
map illustrates amplitudes of contralateral minus mirrored ipsilateral
recordings; redundant parts are omitted. The right column shows
mean amplitudes in the N2pc latency range measured contralateral
(black bars) and ipsilateral (white bars) to the target sample, grouped
by the cued intensity category. Error bars depict within-subject
CIs associated with the main effect of electrode cluster. Note the
differing scales of voltage maps and bar plots. Contra = contralateral;
ipsi = ipsilateral.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
t
F
.
Components of the SEP to Probe Stimuli
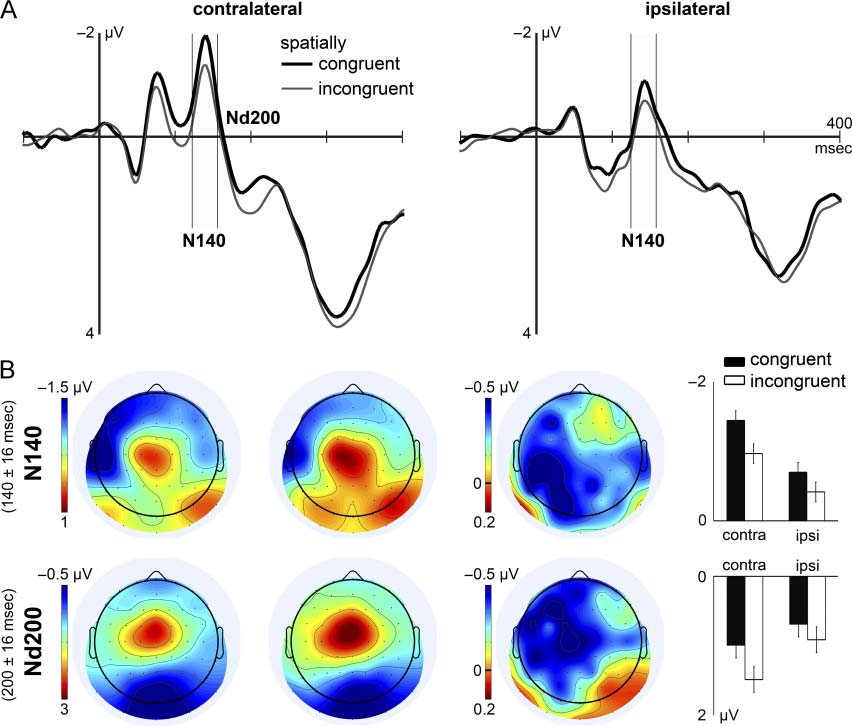
Figure 4A shows SEPs to task-irrelevant probe stimuli, re-
corded contralateral and ipsilateral to stimulation. El
N140 latency range is marked in these graphs because
major predictions were based on this component. Higo-
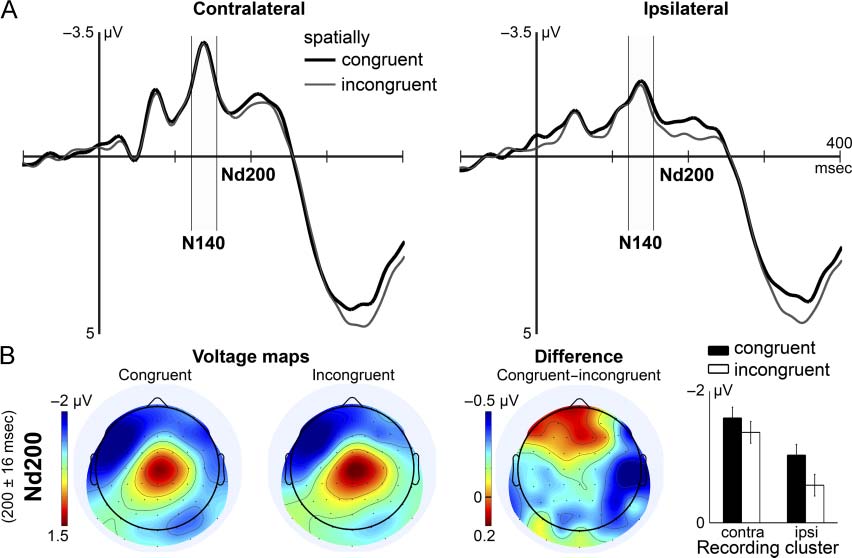
ure 4B depicts scalp distributions of N140 and Nd200
along with the topographic extent of spatial congruency
effects in these time ranges.
Somatosensory N140. The N140 was subject to the spa-
tial relation between probe stimulus and target sample.
Probes presented to the same hand as the retrospectively
cued sample elicited an enhanced N140 as compared
with probes delivered to the other hand (Spatial Con-
gruency: F(1, 16) = 8.30, pag = .011). The influence of
spatial congruency on the N140 was comparable across
recording sites, although in tendency greater contra-
lateral to stimulation (Spatial Congruency × Electrode
Cluster: F(1, 16) = 3.19, pag = .093). Probes elicited greater
contralateral and smaller ipsilateral N140 amplitudes
when the preceding weak sample was presented to the
same hand and the strong sample to the opposite hand
(Stimulation Sequence × Electrode Cluster: F(1, 16) =
7.08, pag = .017). No further effects or interactions were
significativo, except for the well-known asymmetry of
N140 amplitudes toward scalp regions contralateral to
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
1602
Revista de neurociencia cognitiva
Volumen 24, Número 7
stimulation (Electrode Cluster: F(1, 16) = 6.27, pag = .024).
De este modo, selection in STM influenced the N140. Probes
with congruent spatial attributes as the selected item elic-
ited an enhanced N140, as compared with incongruent
probes. This modulation was independent of effects re-
lated to the stimulation sequence used in the present
experimento.
Negative difference wave (Nd200). Spatial congruency
led to a near-significant modulation of the Nd200. Probe
stimuli with congruent spatial properties tended to result
in more negative amplitudes compared with incongruent
probes (spatial congruency: F(1, 16) = 4.31, pag = .055).
This modulation was predominantly found contralateral
to stimulation, as indicated by a significant interaction
between spatial congruency and recording site (Espacial
Congruency × Electrode Cluster: F(1, 16) = 5.46, pag =
.039). No further effects were found in the Nd200 latency
range, except for a tendency of amplitude differences
across contralateral and ipsilateral regions (Electrode
Cluster: F(1, 16) = 4.43, pag = .052).
Components of the SEP to Test Stimuli
Figure 5A shows SEP waveforms in response to tactile test
estímulos, recorded at scalp regions over somatosensory
areas contralateral and ipsilateral to stimulation. N140
amplitudes did not differ for test stimuli presented to
the same or different hand as the cued sample (Espacial
Congruency: F(1, 16) = 0.51, p > .1); compare with Fig-
ure 4B. Además, the N140 was not significantly
different for test stimuli with memorized as compared
with nonmemorized intensity (Intensity Congruency: F(1,
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
t
/
.
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Cifra 4. (A) Grand mean SEPs to task-irrelevant tactile probe stimuli recorded at parietal electrodes contralateral (izquierda) and ipsilateral (bien)
to stimulation. Spatially congruent probe stimuli were presented to the same hand as the cued sample (bold black line), whereas incongruent
probes were delivered to the other hand (thin gray line). (B) Separate rows represent averages during the latency ranges of N140 and Nd200.
Columns from left to right illustrate grand mean isocontour voltage maps for congruent and incongruent probes, difference map, and mean
amplitudes in the particular latency range. The arrangement of data in voltage maps corresponds to stimulation of the right hand. la diferencia
map represents congruent minus incongruent probe stimulus topographies. The right graph shows mean amplitudes in response to spatially
congruente (black bars) and incongruent (white bars) probe stimuli. Within-subject CIs are based on spatial congruency main effects, which were
separately estimated for N140 and Nd200. Note the differing scales of voltage maps and bar plots. Contra = contralateral; ipsi = ipsilateral.
Katus, andersen, and Müller
1603
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
.
t
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Cifra 5. (A) Grand mean SEPs to task-relevant tactile test stimuli acquired at parietal electrodes contralateral (izquierda) and ipsilateral (bien)
to stimulation. Spatially congruent test stimuli were presented to the same hand as the cued sample (bold black line), whereas incongruent
tests were delivered to the other hand (thin gray line). (B) From left to right: grand mean voltage maps, difference map, and averaged
amplitudes in the latency range of the Nd200. The arrangement of data corresponds to stimulation of the right hand. The difference map
represents subtracted topographies to congruent minus incongruent test stimuli. The right graph depicts mean Nd200 amplitudes elicited by
spatially congruent (black bars) and incongruent (white bars) test stimuli, measured contralateral and ipsilateral to stimulation. Within-subject
CIs are based on the spatial congruency main effect for test stimuli. Note the differing scales of voltage maps and bar plots.
16) = 0.00, p > .1). Except for amplitude differences across
recording sites (Electrode Cluster: F(1, 16) = 15.52, pag =
.001), no further effects were present during the N140
latency range. The spatial relation between cued sample
and test stimulus resulted in a significant modulation of
the Nd200: Amplitudes were more negative when test
and cued sample were presented to the same hand com-
pared with different hands (spatial congruency: F(1, 16) =
4.75, pag = .045). No further effects or interactions were
present for the Nd200, except for amplitude differences
between contralateral and ipsilateral recordings (electrodo
grupo: F(1, 16) = 10.59, pag = .005).
Participants were instructed to compare intensity of
cued sample and test stimulus. The P300 to tests was
the only component sensitive to congruency in terms
of the task-relevant feature dimension, intensidad. El
P300 was enhanced when test stimuli were scaled to
memorized intensity compared with distracter intensity
(Intensity Congruency: F(1, 16) = 7.55, pag = .014). En
contrast, the P300 was insensitive to the spatial relation
between cued sample and test stimulus (Spatial Con-
gruency: F(1, 16) = 0.01, p > .1). Intensity congruency
affected the P300 to a greater degree when test and
cued sample were presented to the same hand, as indi-
cated by a significant interaction between both factors
(Intensity Congruency × Spatial Congruency: F(1, 16) =
7.92, pag = .012).
DISCUSIÓN
This study examined effects of spatial attention in tactile
retrospective selection. The paradigm was designed to
rule out attention-based rehearsal: Because the delayed
judgment involved a comparison of intensities rather
than locations, the attention-based rehearsal account
would not predict spatially specific biasing of neural ac-
actividad. Sin embargo, visual retro-cues elicited lateralized com-
ponents (N2pc, ADAN) that mirrored the location of the
target sample. Somatosensory processing during the re-
tention period was also subject to a spatial encoding bias
(indexed by the N140), favoring probes that were deliv-
ered to the same location as the cued sample. Contrary
to that, spatial congruency did not affect the N140 to test
estímulos, indicating that spatial attention was not directed
to one or the other side at the end of the trial. Estos
findings substantiate the interpretation that retrospective
selection leads to transient attention shifts, Opuesto a
the sustained impact of attention-based rehearsal (Awh
et al., 2000, 2006; Jha, 2002). Because the memorization
of locations was not explicitly required, we conclude that
1604
Revista de neurociencia cognitiva
Volumen 24, Número 7
retrospective selection is mediated by a time-variant pro-
cess that is functionally dissociable from attention-based
rehearsal.
Neural Mechanisms of Retrospective Selection
Convergent modulation of physiological measures evoked
by visual and tactile stimuli suggests that retro-cues trigger
focal attention shifts toward the location of the target sam-
por ejemplo. Notablemente, we focused on electrophysiological measures
sensitive to locations rather than nonspatial features or
objects. For the interpretation of spatial attention effects,
it is important to remind that this study adopted a design
in which target and distracter sample were associated
with distinct locations (DellʼAcqua et al., 2010; Kuo et al.,
2009; Griffin & Nobre, 2003). Our results demonstrate
that the neural circuitry mediating the retrospective selec-
tion of nonspatial information is linked to spatial attention
mechanisms.
One interpretation of the transient time course of spa-
tial attention effects in retrospective selection observed
here and in our previous study (Katus et al., 2012) is that
a decay of spatial attributes in STM leads to attenuation
of spatially specific measures. Sin embargo, our results show
that spatial attributes in STM are preserved throughout
the entire retention delay, as the Nd200 component to
tactile probe as well as test stimuli reflected the location
of the cued sample. De este modo, we conclude that transient
spatial attention effects reflect the engagement of a time-
variant executive process.
The ADAN mirrors selection in tactile STM, y esto
finding is in line with evidence from vision (Griffin &
Nobre, 2003). Además, the ADAN modulation points
to the involvement of supramodal networks of selective
atención (Eimer et al., 2004; Eimer & van Velzen, 2002;
van Velzen et al., 2002). Mechanisms of supramodal
attention assert top–down biasing signals that regulate
activity in modality-specific regions (Sambo & Forster,
2011; Lakatos et al., 2009). Because probe stimuli were
presented after the retro-cue, it seems plausible that supra-
modal influences contributed to the modulation of the
somatosensory N140. Todavía, high-level biasing signals gener-
ated in the ADAN time range cannot explain cross-modal
effects at a preceding latency: The N2pc to visual retro-
cues mirrors spatially specific access of information in
tactile STM. Similar results have been reported in purely
visual STM experiments (DellʼAcqua et al., 2010; kuo
et al., 2009). Por eso, the N2pc appears to index a selective
process that is, at least partially, detached from the nature
of mnemonic content.
Cue-related Activity
Visual retro-cues elicited the lateralized N2pc and ADAN
componentes, which consistently indicated the location of
the cued sample. Unlike our previous experiment, retro-
cues did not inform about the location of the relevant
sample here, and for each participant, the same physical
stimulus cued samples at the left and right hands with
equal probability. Our findings support the notion that
retrospective selection engages spatially specific biasing
mechanisms with similarity to those involved in the pro-
cessing of peripheral events, Por ejemplo, in visual search
(N2pc, cf. Kuo et al., 2009) or anticipatory attention tasks
(ADAN; cf. Griffin & Nobre, 2003). Most interestingly,
symmetric retro-cues elicited the N2pc, an observation
extending mere replication of a phenomenon described
previamente (DellʼAcqua et al., 2010; Kuo et al., 2009), ser-
cause retrospective selection was based on mnemonic
representations of tactile rather than visual signals. Our re-
sults show that the N2pc is sensitive to selection in STM,
even when the accessed information is related to a non-
visual modality.
The N2pc is thought to originate in occipito-temporal re-
gions, presumably in extrastriate visual cortex (Oostenveld,
Praamstra, Stegeman, & van Oosterom, 2001; Luck &
Hillyard, 1994a, 1994b). Sources downstream to visual
sensory areas are known to contribute to the N2pc: Hopf
et al. (2000) reported engagement of parietal regions pre-
ceding activity in occipito-temporal cortex. Los autores
related the early parietal source to neural networks con-
trolling the directionality of attention shifts. cohen, Heitz,
Schall, and Woodman (2009) demonstrated that feedback
from FEF neurons influences the generation of the ma-
caque N2pc homologue in posterior cortical regions. En
aggregate, these findings point toward a hierarchy of neu-
ral processes shaping the N2pc. Top–down influence from
attentional control structures might regulate sensory-
specific mechanisms in visual cortex, such as spatial filter-
ing of visual signals (p.ej., Hopf et al., 2000). In accordance
with findings in vision (DellʼAcqua et al., 2010; Kuo et al.,
2009), we show that N2pc effects can be obtained in ab-
sence of lateralized visual events, supporting the view
that the N2pc is sensitive to selection at a postperceptual
stage. The early engagement of posterior regions points
to an involvement of neural processes related to the visual
sistema.
Spatial attributes of tactile events are describable in
terms of anatomical and extrapersonal coordinates because
touch operates on separable—yet linked—spatial reference
marcos (Sambo & Forster, 2011; Heed & Röder, 2010; Eimer,
Forster, & van Velzen, 2003). The N2pc is primarily as-
sociated with vision where spatial selection is based on an
extrapersonal frame (Sambo & Forster, 2011). Visual space
is furthermore of relevance for neural structures guiding
eye movements, such as the FEFs (Bisley, 2011; van der
Lubbe, Neggers, Verleger, & Kenemans, 2006; Corbetta,
1998) and the superior colliculus (p.ej., Belopolsky &
Theeuwes, 2011). Saccade studies provided evidence for
an active role of the oculumotor system in STM tasks that
involve retrospective selection (Belopolsky & Theeuwes,
2011) as well as attention-based rehearsal (Belopolsky &
Theeuwes, 2009b). The overlap between eye movements,
spatial attention, and spatial STM in vision is consistent with
Katus, andersen, and Müller
1605
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
.
t
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
the notion that these domains rely on common spatial
codes (Theeuwes et al., 2009). This study demonstrates
that the N2pc to visual retro-cues mirrors selection in
mnemonic representations of tactile signals. We specu-
late that such early cross-modal links are attributable to
the common coding of extrapersonal coordinates in touch
and vision.
Processing of Somatosensory Events
The analyses of tactile stimuli (probes, pruebas) revealed
two results of interest. Primero, a spatial bias in tactile encod-
En g (indexed by the N140) was evident for probes but not
for test stimuli. This finding is consistent with the pro-
posal that, unlike attention-based rehearsal, retrospective
selection leads to transient fluctuations of spatial atten-
tion time-locked to the retro-cue (Katus et al., 2012). Sec-
ond, spatial congruency influenced the SEP to test stimuli
at latencies before intensity congruency, Indicando que
spatial and nonspatial attributes were evaluated in a se-
quential fashion. Because we examined the role of spatial
attention in retrospective selection, we first discuss spa-
tial congruency effects (p.ej., N140) before we address
modulations of intensity congruency (P300).
The somatosensory N140 is a reliable marker of spatial
selection between the left and right hands in perception
(Forster & Eimer, 2004, 2005; Eimer et al., 2004; Zopf
et al., 2004; Eimer & Forster, 2003a; Michie et al., 1987;
Josiassen et al., 1982; Desmedt & Robertson, 1977). En el
present experiment, the N140 to probe stimuli mirrored
the location of the cued sample: Amplitudes were en-
hanced when sample and probe were presented to the
same hand, relative to different hands. Spatial congruency
did not affect the N140 to test stimuli. En resumen, estos
findings suggest that retrospective selection guides sen-
sory encoding for a brief period after the retro-cue. En
our previous spatial STM study, greatest attentional N140
modulations were observed for probes that were pre-
sented early after the retro-cue, as compared with late
in the retention delay. Aquí, the timing of probe stimuli
roughly corresponds to the latencies of such early probes.
We replicate a transient spatial encoding bias because of
selection in STM in a paradigm that did not explicitly re-
quire spatial STM, specifically the maintenance of a loca-
tion for subsequent comparison with a delayed stimulus.
Test stimuli of memorized intensity led to higher hit
rates when test and cued sample were presented to the
same hand, compared with different hands. Does this be-
havioral finding suggest that retrospective selection leads
to sustained—rather than transient—spatial attention ef-
efectos? Notablemente, hit rates are associated with responses to
objetivos (memorized intensity), whereas correct rejections
relate to distracters (nonmemorized intensity). Sin embargo,
the enhancement of hit rates was not accompanied by a
comparable modulation of the proportion of correct re-
jections. It is implausible to assume that spatial attention
was deployed in a sustained fashion in half of the ex-
perimento (objetivos) and transiently in the remaining trials
(distracters), because test stimuli of memorized and non-
memorized intensities were presented in a random se-
quence. Además, the N140 component is a reliable
marker of spatially specific encoding (p.ej., Forster & Eimer,
2005). If test stimuli with target but not distracter intensity
were subject to a spatial encoding bias, the N140 should
reflect an interaction between spatial congruency and
intensity congruency, which was not the case. En cambio,
we found no physiological evidence for spatially specific
encoding at the end of the retention delay.
The spatial congruency effect in hit rates might be attrib-
utable to sensitivity differences between the dominant and
nondominant hands: The recognition of a memorized in-
tensity (objetivo) should be easier when the sensory system
(mano) is kept constant. De lo contrario, the altered sensory
system might introduce variability in the sensation elicited
by a particular physical intensity. In light of these consid-
erations, we presume that the spatial congruency effect
on hit rates is not caused by perceptual spatial attention.
The N140 to probe stimuli exhibited a spatial encoding
inclinación, but the N140 to test stimuli did not. The absence of
a spatial encoding bias for test stimuli suggests that, en el
end of retention, spatial attention was equally divided
between the left and right hands. This is not surprising
because spatial randomization of test stimuli eliminates a
confounder in spatial STM paradigms: the conceivable
strategy to anticipate test stimuli at memorized locations.
As to test stimuli, the experimental design manipulated
congruency in separate feature dimensions, space and
intensidad. The task required a delayed comparison of inten-
sities rather than locations. Por eso, only nonspatial attri-
butes of test stimuli conveyed crucial information for
the judgment. It is established that the decision-related
P300 is enhanced for target stimuli as compared with dis-
tracters (Sambo & Forster, 2011; Polich, 2007; Desmedt &
Debecker, 1979). Intensity congruency modulated the
P300, leading to enhanced amplitudes when tests were
scaled to memorized relative to nonmemorized intensity.
A diferencia de, spatial congruency did not affect the P300.
Respectivamente, the P300 reflects attributes of task-relevant
features but not attributes of feature dimensions that are
arbitrary for a cognitive decision.
Are the Nd200 and P300 indicative for successive phases
in the evaluation of tactile information? Curiosamente,
numerous somatosensory studies that found a relative
negativity around 200-msec poststimulus did not report
an enhanced P300 for targets as compared with distracters
(Gillmeister & Forster, 2012; Forster & Gillmeister, 2011;
Gillmeister, Adler, & Forster, 2010; Forster & Eimer,
2004; Eimer & Forster, 2003b). This observation led to
the speculation that the negative difference wave repre-
sents a reversed attention effect at longer latencies (Sambo
& Forster, 2011). Aquí, the P300 was clearly separable
from the rather short-lived Nd200 modulation, y más
importantly, these long latency components mirrored
congruency in separate feature dimensions of space
1606
Revista de neurociencia cognitiva
Volumen 24, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
.
t
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
(Nd200) and intensity (P300). The Nd200 was only sen-
sitive to the task-irrelevant feature dimension, with a con-
sistent pattern of modulation across behaviorally irrelevant
probes and test stimuli that required decision and re-
respuesta. One speculation is that the Nd200 reflects a phase
of rapid evaluation of stimulus properties against templates
in STM before an exhaustive processing of task-relevant
information and decision (indexed by the P300). This in-
terpretation would be consistent with the finding that
the Nd200 attenuates when participants attend to two lo-
cations, relative to a single location (Eimer & Forster,
2003b). En resumen, spatial congruency and intensity con-
gruency modulated successive components of the SEP to
test stimuli. This result points to an asynchronous process-
ing of spatial and nonspatial attributes (pero, see Forster &
Eimer, 2004).
General Conclusions
The focus of attention can be flexibly altered in STM
(Lepsien & Nobre, 2006). Retro-cues that inform about
a nonspatial attribute of the target sample initiate a cas-
cade of spatially specific biasing signals indicative for the
targetʼs location. Retrospective selection affects markers of
spatial attention with neural origins in posterior (N2pc),
anterior (ADAN), and fronto-parietal (N140) redes.
Intensity information is accessed in a spatially specific
moda, which demonstrates that spatial properties are
preserved in mnemonic representations, even if memory
for locations is not explicitly instructed (Astle et al., 2009;
Kuo et al., 2009).
The earliest influence of retrospective selection was
found at posterior scalp regions: The N2pc to visual retro-
cues is sensitive to selection in tactile STM, and this effect
is not attributable to the physical asymmetries of cue stim-
uli (cf. van Velzen & Eimer, 2003). We speculate that the
N2pc reflects involvement of early selection mechanisms
that operate on coordinates in external space. Además,
we found evidence for a functional overlap of selection
mechanisms in memory and perception (Gazzaley &
Nobre, 2012; Gazzaley, 2011; Awh et al., 2006), as encod-
ing of peripheral tactile probe stimuli (indexed by the
N140) was biased in favor of events that shared spatial
properties with the retrospectively cued sample. El
N140 to test stimuli was insensitive to spatial congruency.
Por eso, these results suggest that selection in tactile STM
triggers transient shifts of spatial attention.
Our findings support a close link between retrospective
selection and covert spatial orienting. Although attention-
based rehearsal is similarly known to lead to spatial atten-
tion effects (Theeuwes et al., 2009; Awh et al., 2000, 2006;
Jha, 2002), maintenance of locations was not required
aquí. Our results point in the direction that spatially spe-
cific biasing of neural activity is caused by a time-variant
process triggered by the presentation of the retro-cue.
We conclude that the role of spatial attention in retro-
spective selection is functionally dissociable from attention-
based rehearsal of locations.
Expresiones de gratitud
We thank Renate Zahn, Henriette Greischel, and Elizabeth
Lafrentz for assistance with data acquisition and Salvador Soto-
Faraco, Björn Herrmann, Artem Belopolsky, and the anonymous
reviewer for helpful comments and advices. This work was sup-
ported by a grant of the German Research Foundation (DFG).
Reprint requests should be sent to Matthias M. Müller, Institut für
Psychologie, Universität Leipzig, Seeburgstrasse. 14–20, 04103
Leipzig, Alemania, o por correo electrónico: m.mueller@rz.uni-leipzig.de.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
F
t
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
REFERENCIAS
Astle, D. MI., Scerif, GRAMO., kuo, B. C., & Nobre, A. C. (2009). Espacial
selection of features within perceived and remembered
objects. Frontiers in Human Neuroscience, 3, 6.
Awh, MI., Anllo-Vento, l., & Hillyard, S. A. (2000). The role
of spatial selective attention in working memory for
locations: Evidence from event-related potentials. Diario
of Cognitive Neuroscience, 12, 840–847.
Awh, MI., & En Jonia, j. (2001). Overlapping mechanisms
of attention and spatial working memory. Tendencias en
Cognitive Sciences, 5, 119–126.
Awh, MI., Vogel, mi. K., & Oh, S. h. (2006). Interactions
between attention and working memory. Neurociencia,
139, 201–208.
Campana, A. J., & Sejnowski, t. j. (1995). An information-
maximization approach to blind separation and blind
deconvolution. Computación neuronal, 7, 1129–1159.
Belopolsky, A. v., & Theeuwes, j. (2009a). No functional role of
attention-based rehearsal in maintenance of spatial working
memory representations. Acta Psicológica, 132, 124–135.
Belopolsky, A. v., & Theeuwes, j. (2009b). Inhibition of
saccadic eye movements to locations in spatial working
memory. Atención, Perception and Psychophysics, 71,
620–631.
Belopolsky, A. v., & Theeuwes, j. (2011). Selection within
visual memory representations activates the oculomotor
sistema. Neuropsicología, 49, 1605–1610.
Bisley, j. W.. (2011). The neural basis of visual attention.
Revista de fisiología, 589, 49–57.
cohen, j. y., Heitz, R. PAG., Schall, j. D., & Woodman, GRAMO. F. (2009).
On the origin of event-related potentials indexing covert
attentional selection during visual search. Diario de
Neurofisiología, 102, 2375–2386.
Corbetta, METRO. (1998). Frontoparietal cortical networks for
directing attention and the eye to visual locations: Identical,
independiente, or overlapping neural systems? Actas
of the National Academy of Sciences, EE.UU., 95, 831–838.
Curtis, C. MI., & DʼEsposito, METRO. (2003). Persistent activity in
the prefrontal cortex during working memory. Tendencias en
Cognitive Sciences, 7, 415–423.
DellʼAcqua, r., Sessa, PAG., Toffanin, PAG., Luria, r., & Jolicoeur, PAG.
(2010). Orienting attention to objects in visual short-term
memory. Neuropsicología, 48, 419–428.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Revista de neurociencia
Métodos, 134, 9–21.
Delorme, A., Sejnowski, T., & Makeig, S. (2007). Enhanced
detection of artifacts in EEG data using higher-order statistics
and independent component analysis. Neuroimagen, 34,
1443–1449.
Katus, andersen, and Müller
1607
Desmedt, j. MI., & Debecker, j. (1979). Wave form and
neural mechanism of the decision P350 elicited without
pre-stimulus CNV or readiness potential in random
sequences of near-threshold auditory clicks and
finger stimuli. Electroencephalography and Clinical
Neurofisiología, 47, 648–670.
Desmedt, j. MI., & Robertson, D. (1977). Differential
enhancement of early and late components of the
cerebral somatosensory evoked potentials during
forced-paced cognitive tasks in man. Diario de
Physiology, 271, 761–782.
Eimer, METRO., & Forster, B. (2003a). Modulations of early
somatosensory ERP components by transient and
sustained spatial attention. Cerebro experimental
Investigación, 151, 24–31.
Eimer, METRO., & Forster, B. (2003b). The spatial distribution of
attentional selectivity in touch: Evidence from somatosensory
ERP components. Clinical Neurophysiology, 114, 1298–1306.
Eimer, METRO., Forster, B., Fieger, A., & Harbich, S. (2004). Effects
of hand posture on preparatory control processes and
sensory modulations in tactile-spatial attention. Clinical
Neurofisiología, 115, 596–608.
Eimer, METRO., Forster, B., & van Velzen, j. (2003). Anterior and
posterior attentional control systems use different spatial
reference frames: ERP evidence from covert tactile-spatial
orienting. Psychophysiology, 40, 924–933.
Eimer, METRO., & van Velzen, j. (2002). Crossmodal links in spatial
attention are mediated by supramodal control processes:
Evidence from event-related potentials. Psychophysiology,
39, 437–449.
Forster, B., & Eimer, METRO. (2004). The attentional selection of
spatial and non-spatial attributes in touch: ERP evidence for
parallel and independent processes. Biological Psychology,
66, 1–20.
Forster, B., & Eimer, METRO. (2005). Covert attention in touch:
Behavioral and ERP evidence for costs and benefits.
Psychophysiology, 42, 171–179.
Forster, B., & Gillmeister, h. (2011). ERP investigation
of transient attentional selection of single and multiple
locations within touch. Psychophysiology, 48, 788–796.
Forster, B., Sambo, C. F., & Pavone, mi. F. (2009). ERP correlates
of tactile spatial attention differ under intra- and intermodal
condiciones. Biological Psychology, 82, 227–233.
Gazzaley, A. (2011). Influence of early attentional modulation
on working memory. Neuropsicología, 49, 1410–1424.
Gazzaley, A., & Nobre, A. C. (2012). Top–down modulation:
Bridging selective attention and working memory. Trends
in Cognitive Sciences, 16, 129–135.
Gillmeister, h., Adler, J., & Forster, B. (2010). Object-guided
spatial attention in touch: Holding the same object with
both hands delays attentional selection. Diario de
Neurociencia Cognitiva, 22, 931–942.
Gillmeister, h., & Forster, B. (2012). Hands behind your
atrás: Effects of arm posture on tactile attention in the
space behind the body. Experimental Brain Research,
216, 489–497.
Griffin, I. C., & Nobre, A. C. (2003). Orienting attention to
locations in internal representations. Revista de Cognitivo
Neurociencia, 15, 1176–1194.
Heed, T., & Röder, B. (2010). Common anatomical and
external coding for hands and feet in tactile attention:
Evidence from event-related potentials. Diario de
Neurociencia Cognitiva, 22, 184–202.
Hopf, j. METRO., Luck, S. J., Girelli, METRO., Hagner, T., Mangun,
GRAMO. r., Scheich, h., et al. (2000). Neural sources of focused
attention in visual search. Corteza cerebral, 10, 1233–1241.
Jarmasz, J., & Hollands, j. GRAMO. (2009). Confidence intervals in
repeated-measures designs: The number of observations
principle. Canadian Journal of Experimental Psychology,
63, 124–138.
Jha, A. PAG. (2002). Tracking the time-course of attentional
involvement in spatial working memory: An event-related
potential investigation. Brain Research, Cognitive Brain
Investigación, 15, 61–69.
Josiassen, R. C., Shagass, C., Roemer, R. A., Ercegovac, D. v.,
& Straumanis, j. j. (1982). Somatosensory evoked potential
changes with a selective attention task. Psychophysiology,
19, 146–159.
Junghöfer, METRO., Elbert, T., Tucker, D. METRO., & Rockstroh, B.
(2000). Statistical control of artifacts in dense array
EEG/MEG studies. Psychophysiology, 37, 523–532.
Katus, T., andersen, S. K., & Muller, METRO. METRO. (2012). Maintenance
of tactile short-term memory for locations is mediated by
spatial attention. Biological Psychology, 89, 39–46.
kuo, B. C., Rao, A., Lepsien, J., & Nobre, A. C. (2009). Searching
for targets within the spatial layout of visual short-term
memory. Revista de neurociencia, 29, 8032–8038.
kuo, B. C., alimenta, METRO. GRAMO., & Nobre, A. C. (2012). Atención
modulates maintenance of representations in visual
short-term memory. Revista de neurociencia cognitiva,
24, 51–60.
Lakatos, PAG., OʼConnell, METRO. NORTE., Barczak, A., Mills, A., Javitt,
D. C., & Schroeder, C. mi. (2009). The leading sense:
Supramodal control of neurophysiological context by
atención. Neurona, 64, 419–430.
Lepsien, J., & Nobre, A. C. (2006). Cognitive control of attention
en el cerebro humano: Insights from orienting attention to
mental representations. Brain Research, 1105, 20–31.
Loftus, GRAMO. r., & Masson, METRO. mi. j. (1994). Using confidence
intervals in within-subject designs. Boletín psiconómico
and Review, 1, 476–490.
Luck, S. J., & Hillyard, S. A. (1994a). Spatial filtering during
visual search: Evidence from human electrophysiology.
Revista de Psicología Experimental: Percepcion humana
y rendimiento, 20, 1000–1014.
Luck, S. J., & Hillyard, S. A. (1994b). Electrophysiological
correlates of feature analysis during visual search.
Psychophysiology, 31, 291–308.
Michie, PAG. T., Bearpark, h. METRO., Crawford, j. METRO., & Glue, l. C.
(1987). The effects of spatial selective attention on the
somatosensory event-related potential. Psychophysiology,
24, 449–463.
Müller, norte. GRAMO., & Caballero, R. t. (2006). The functional
neuroanatomy of working memory: Contributions of
human brain lesion studies. Neurociencia, 139, 51–58.
Naghavi, h. r., & Nyberg, l. (2005). Common fronto-parietal
activity in attention, memory, and consciousness: Shared
demands on integration? Consciousness and Cognition,
14, 390–425.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsicología,
9, 97–113.
Oostenveld, r., Praamstra, PAG., Stegeman, D. F., & van Oosterom,
A. (2001). Overlap of attention and movement-related
activity in lateralized event-related brain potentials.
Clinical Neurophysiology, 112, 477–484.
Pasternak, T., & verdelee, METRO. W.. (2005). Working memory in
primate sensory systems. Naturaleza Reseñas Neurociencia,
6, 97–107.
Polich, j. (2007). Updating P300: An integrative theory of
P3a and P3b. Clinical Neurophysiology, 118, 2128–2148.
Sambo, C. F., & Forster, B. (2011). Sustained spatial attention
in touch: Modality-specific and multimodal mechanisms.
The Scientific World Journal, 11, 199–213.
Sörös, PAG., Marmurek, J., Tam, F., Panadero, NORTE., Staines, W.. r., &
graham, S. j. (2007). Functional MRI of working memory
1608
Revista de neurociencia cognitiva
Volumen 24, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
t
.
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
and selective attention in vibrotactile frequency discrimination.
BMC Neuroscience, 8, 48.
Theeuwes, J., Belopolsky, A., & Olivers, C. norte. (2009).
Interactions between working memory, attention and
eye movements. Acta Psicológica, 132, 106–114.
van der Lubbe, R. h., Neggers, S. F., Verleger, r., & Kenemans,
j. l. (2006). Spatiotemporal overlap between brain activation
related to saccade preparation and attentional orienting.
Brain Research, 1072, 133–152.
van Velzen, J., & Eimer, METRO. (2003). Early posterior ERP
components do not reflect the control of attentional shifts
toward expected peripheral events. Psychophysiology, 40,
827–831.
van Velzen, J., Forster, B., & Eimer, METRO. (2002). Temporal
dynamics of lateralized ERP components elicited during
endogenous attentional shifts to relevant tactile events.
Psychophysiology, 39, 874–878.
watson, A. B., & Pelli, D. GRAMO. (1983). QUEST: Un bayesiano
adaptive psychometric method. Perception and
Psicofísica, 33, 113–120.
Zopf, r., Giabbiconi, C. METRO., Gruber, T., & Muller, METRO. METRO.
(2004). Attentional modulation of the human somatosensory
evoked potential in a trial-by-trial spatial cueing and sustained
spatial attention task measured with high density 128
channels EEG. Brain Research, Cognitive Brain Research,
20, 491–509.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
4
/
7
2
4
1
/
5
7
9
/
6
1
1
5
9
9
4
6
4
/
3
1
7
7
7
7
oh
7
C
9
norte
4
_
0
a
/
_
j
0
oh
0
C
2
norte
3
4
_
a
pag
_
d
0
0
b
2
y
3
gramo
4
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
/
F
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Katus, andersen, and Müller
1609