Actúe rápidamente, decidir más tarde: Visual de larga latencia
Processing Underlies Perceptual Decisions
but Not Reflexive Behavior
Jacob Jolij1,2,3,4, h. Steven Scholte2, Simon van Gaal2,
Timothy L. Hodgson3, and Victor A. F. Lamme2
D
oh
w
norte
yo
oh
a
d
mi
d
Abstracto
■ Humans largely guide their behavior by their visual represen-
tation of the world. Recent studies have shown that visual in-
formation can trigger behavior within 150 mseg, suggesting that
visually guided responses to external events, En realidad, precede con-
scious awareness of those events. Sin embargo, is such a view correct?
By using a texture discrimination task, we show that the brain relies
on long-latency visual processing in order to guide perceptual
decisiones. Decreasing stimulus saliency leads to selective changes
in long-latency visually evoked potential components reflecting
scene segmentation. These latency changes are accompanied
by almost equal changes in simple RTs and points of subjective
simultaneity. Además, we find a strong correlation between
individual RTs and the latencies of scene segmentation related
components in the visually evoked potentials, showing that the
processes underlying these late brain potentials are critical in trig-
gering a response. Sin embargo, using the same texture stimuli in
an antisaccade task, we found that reflexive, but erroneous, pro-
saccades, but not antisaccades, can be triggered by earlier visual
procesos. En otras palabras: The brain can act quickly, but decides
late. Differences between our study and earlier findings suggest-
ing that action precedes conscious awareness can be explained
by assuming that task demands determine whether a fast and
unconscious, or a slower and conscious, representation is used
to initiate a visually guided response. ■
INTRODUCCIÓN
Visual information plays an important role in guiding our
comportamiento. Sin embargo, processing and extracting relevant in-
formation from visual input is a highly complex process
that can take up a considerable amount of time. Reciente
studies have shown that it can take up to almost 400 mseg
before visual information is available for conscious report
(p.ej., Scharnowski et al., 2009; Heinen, Jolij, & Lamme, 2005).
It is therefore not surprising that reacting to external
events does not require awareness of those events. En
the literature, there are numerous examples of dissocia-
tions between action and awareness. Goodale and Milner
(1992), Por ejemplo, describe a patient with visual agnosia
(es decir., this patient has no conscious access to object iden-
tity) who is, nevertheless, perfectly able to manipulate
objects. Another example is blindsight, the remarkable
capability of some patients with a lesion to primary visual
cortex to guess correctly about attributes of visual stimuli,
such as color, orientación, or even facial expression (p.ej.,
Weiskrantz, 1996).
Dissociations between action and awareness have been
demonstrated in healthy individuals, también. Por ejemplo,
1Universidad de Groninga, Los países bajos, 2University of Am-
sterdam, Los países bajos, 3University of Exeter, Reino Unido, 4Swiss Federal
Institute of Technology Lausanne (EPFL), Lausanne, Suiza
when observers are instructed to pick up an object, el
aperture between the thumb and the index finger is not
influenced by distortions in the perceived size of the object,
mimicking the situation in which conscious access to an
objectʼs features is accurate, but manipulation is still in-
tact (Goodale & Milner, 1992). Complete dissociations be-
tween action and awareness have been demonstrated, también:
Using TMS of primary visual cortex, blindsight has been
reported in normal observers for color, orientación, y
emotional expression (Boyer, harrison, & Ro, 2005; Jolij
& Lamme, 2005).
It seems that conscious awareness is not a prerequisite
for action: Estrictamente hablando, there is no need to see some-
thing in order to respond to it. Presently, there is general
consensus in the literature that there are different modes
of processing in the visual system: visual processes that
drive action and visual processes that result in conscious
percepción. To some extent, these processes are indepen-
dent and rely on different neural structures. Visual pro-
cessing for action is done in the dorsal cortical areas, y
driven by fast magnocellular visual inputs, allowing for quick
respuestas. Visual processing for perception, en el otro
mano, is done in more ventral cortical areas, and driven by
parvocellular inputs, that are capable of representing more
spatial details, but slower in processing visual information
(Goodale & Milner, 1992).
© 2011 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 23:12, páginas. 3734–3745
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
r
t
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
2
C
3
yo
1
mi
2
–
3
pag
7
d
3
F
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
oh
3
C
4
norte
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
0
oh
0
norte
3
0
4
8
.
S
pag
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
/
F
.
t
oh
norte
1
8
METRO
a
y
2
0
2
1
Sin embargo, within the “vision-for-perception” system, rapid
stimulus processing preceding awareness is possible as
Bueno. Thorpe, Fize, and Marlot (1996) demonstrated that
EEG responses to different categories of stimuli, como
animals versus inanimate objects, already deviate as early
como 150 msec after stimulus presentation, showing that ob-
ject classification can be accomplished extremely fast. Usando
an analysis of RT distributions to masked and unmasked
estímulos, VanRullen and Koch (2003) demonstrated that a
single feedforward sweep through the visual hierarchy is
sufficient to trigger a behavioral response to the presence
of a target. They presented masked and unmasked images
of natural scenes for 26 mseg, while subjects were engaged
in go/no-go task: They had to respond only when the
scene contained an animal. The analysis of the RT pattern
revealed that the moment at which scenes containing an
animal were discriminated from scenes not containing an
animal (es decir., the first moment in time in which number of
hits is significantly higher than the number of false alarms)
was the same for both the masked and the unmasked stim-
uli, showing that the information extracted during the first
26 msec of visual processing can trigger a behavioral re-
sponse related to a perceptual decision, independent of
visual awareness.
Tomados juntos, there is ample evidence that we can
respond to visual events in our environment before we
become aware of them—it almost seems that in order to
explain human perceptual decision-making, awareness
can be left out of the equation. En efecto, recent models of
perceptual decision-making do not assign any special value
to perceptual awareness. En cambio, when confronted with
a stimulus, the system simply responds when sufficient
information is available in order to make a decision, based
on internally set thresholds (p.ej., ratcliff & McKoon, 2008).
Whether or not that information is represented in con-
scious awareness is irrelevant.
Sin embargo, is this idea correct? Here we used texture
checkerboards in order to investigate what information
triggers behavior. Visual processing of texture checker-
boards requires texture segregation, which is a two-stage
proceso: Primero, borders of figures are detected, and sub-
sequently, the figures are filled in. Border detection occurs
around 80–90 msec, and is likely to be the result of lateral
inhibition within cortical areas, whereas figure filling-in can
take up to 200 msec and depends on re-entrant processing
(Scholte, Jolij, Fahrenfort, & Lamme, 2008; Jehee, Roelfsema,
decoración, Murre, & Lamme, 2007; Heinen et al., 2005; Roelfsema,
Lamme, Spekreijse, & Bosch, 2002; Caputo & Casco, 1999;
Lamme, 1995). This latter stage has been linked to per-
ceptual awareness of texture stimuli, whereas the former
stage could be sufficient in order to detect presence of
a texture stimulus ( Jehee, Lamme, & Roelfsema, 2007;
Heinen et al., 2005; Lamme, 1995, 2003; Supèr, Lamme, &
Spekreijse, 2001).
Curiosamente, evoked potential components related to
filling-in have been shown to increase in latency when de-
creasing perceived stimulus segregation strength, mientras
earlier components reflecting border detection remain un-
affected ( Jolij et al., 2007; Caputo & Casco, 1999). We in-
vestigated the relation between texture processing and
behavior in three experiments, in which we varied per-
ceived segmentation strength of texture and the task at
mano. In the first experiment, participants had to press a
button whenever they detected a texture, irrespective of
perceived segregation strength. EEG was measured during
este experimento. In Experiment 2, we measured points of
subjective simultaneity for two types of textures differing in
perceived segmentation strength and the onset of a sound.
In the last experiment, we used the two texture types as
target stimuli in an antisaccade task.
We found that simple detection RTs to textures corre-
spond strongly with latency of filling-in related component
(Experimento 1). Points of subjective simultaneity for tex-
tures and a sound also showed a similar correspondence
(Experimento 2). Sin embargo, an analysis of single-trial EEG
data revealed that after approximately 130 mseg, sufficient
information is available to reliably detect the presence of
a texture, irrespective of perceived segregation strength.
Data from the antisaccade experiment (Experimento 3)
show that this early activity can be used to trigger reflexive
comportamiento, en este caso, erroneous express-saccades toward
texture stimuli. Juntos, our findings suggest that re-
sponses to external events may, en algunos casos, not be trig-
gered as soon as information is available. En cambio, visually
guided responses to external events seem to be initiated
only when these external events are registered in visual
awareness, unless it concerns more reflexive behavior.
MÉTODOS
Participantes
All participants were healthy freshman psychology students
from the University of Amsterdam (experimentos 1 y 2)
and the University of Exeter (Experimento 3), with no
reported history of neurological or psychiatric health
problemas, and normal or corrected-to-normal vision. Ellos
received either study-credit or A7 per hour for participa-
ción. Thirty-three participants (18–24 years, 21 women)
participated in Experiment 1, 9 Participantes (18–19 years,
6 women) participated in Experiment 2, y 11 participar-
pants (18–34 years, all women) participated in Experi-
mento 3. None of them participated in any of the other
experiments of this study. All participants were naive ob-
servers and had never participated in texture discrimina-
tion experiments before. Written informed consent was
obtained from all participants prior to the experiments,
and all experiments were approved by the respective local
Ethics Committees.
Visual Stimulation
In all experiments, we used stimuli on the basis of the
stimuli used by Lamme, Van Dijk, and Spekreijse (1992),
Jolij et al.
3735
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
r
t
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
2
C
3
yo
1
mi
2
–
3
pag
7
d
3
F
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
oh
3
C
4
norte
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
0
oh
0
norte
3
0
4
8
.
S
pag
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
/
F
t
.
oh
norte
1
8
METRO
a
y
2
0
2
1
who used a stimulus in which orientation of line seg-
ments was modulated in such a way that a texture checker-
board appeared and disappeared from a homogeneous
texture background. Sin embargo, we increased the spacing
between the squares of the checkerboard, thus induc-
ing a strong percept of texture-defined squares against
a background. We manipulated the saliency of the tex-
ture squares by using two orientation differences be-
tween the squares and the background, being 90° or 20°
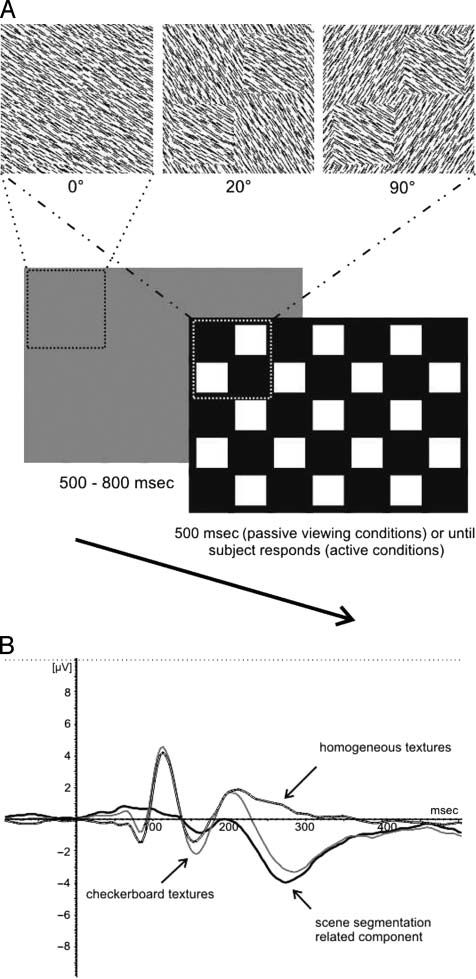
(ver Figura 1A).
Activity related to scene segmentation can be isolated
by subtracting the homogeneous visually evoked poten-
tials (VEPs) from the checkerboard VEPs (ver Figura 1B).
Sin embargo, to isolate these components, all local elements
of the textures (homogeneous and checkerboards) need
to be balanced, eso es, all local orientations should be pre-
sented an equal number of times as part of the background
and as part of the texture squares in the foreground. En
order to achieve this, we used line orientations of 35°,
55°, 125°, and 145°, yielding two pairs of 20° checkerboards
(35° foreground and 55° background, y viceversa; 125°
foreground and 145° background and vice versa), two
pairs of 90° checkerboards (35° foreground and 125° back-
ground and vice versa; 55° foreground and 145° back-
ground and vice versa), and four homogeneous textures.
All types of stimuli were presented an equal number of
veces.
Stimuli were generated on a PC and displayed on a 19-in.
Iiyama monitor, with a refresh rate of 100 Hz and a screen
resolution of 1024 por 768 píxeles (21.8° by 16° visual arc).
Subjects were seated 100 cm from the monitor. Cada prueba
started with a homogeneous texture which was displayed
for 300–800 msec. Entonces, the homogeneous texture was
replaced by either another homogeneous texture, a 90°
orientation contrast checkerboard or a 20° orientation con-
trast checkerboard, each with a 33% random probabil-
idad. The stimuli were always chosen in such a way that all
local line elements were replaced by new ones in order to
prevent static parts in the display. VEPs were computed
for the second textures only. The second texture remained
on screen until the subject responded by pressing a but-
ton with the dominant hand, after which a new trial started.
The size of the individual squares of the checkerboard was
2.5° by 2.5° visual arc. Although this results in relatively
long presentation times for the texture stimuli compared
to earlier work, we cannot expect that the longer pre-
sentation times will have a large effect on the processing
of textures (compare e.g., Fahrenfort, Scholte, & Lamme,
2007, who have used very brief presentation times, y
Scholte et al., 2008, using long presentation times, yielding
very similar results).
During the temporal order judgment task (Experimento 2),
white noise was played through speakers with different
SOAs with respect to the onset of the second texture,
que van desde 200 msec before the onset of the second
texture to 200 msec after the onset of the second texture
in 20-msec intervals. Noise lasted to the end of the trial
Cifra 1. Typical trial run and VEP analysis. (A) Example of a typical
trial run. After a homogeneous texture, presented for 300–800 msec,
a 20° orientation contrast, a 90° orientation contrast, or a homogeneous
texture could appear. Background and foreground elements were always
replaced with new elements. In the passive condition, the texture was
presented for 500 mseg, whereas in the active condition, it was presented
until the subject pressed a button. (B) Rationale of the VEP analysis.
By averaging all homogeneous orientations (white) and by subtracting
this signal from the average of all checkerboard presentations of a given
orientation contrast (gray), we could isolate the segregation-specific signal
(solid black). Here the activity in channel POz for a 90° orientation
contrast texture is shown.
to avoid effects of sound offset ( Jaskowski, 1996). The par-
ticipantʼs task was to indicate whether the noise started
before or after the onset of the second texture, but only
when this texture contained a checkerboard. Cuando el
3736
Revista de neurociencia cognitiva
Volumen 23, Número 12
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
r
t
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
2
C
3
yo
1
mi
2
–
3
pag
7
d
3
F
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
oh
3
C
4
norte
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
0
oh
0
norte
3
0
4
8
.
S
pag
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
/
t
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
second texture was a homogeneous texture, Participantes
had to withhold their response. The next trial was started
after the participant made a response. Before the experi-
mental session, participants were trained to perform at
∼100% at the −200 msec and 200 msec SOAs. At least
40 trials per texture–sound SOA per condition (20° or
90° orientation contrast) were collected per participant.
In Experiment 3, texture stimuli could only appear on
the left or on the right side of the screen. No fixation dot
was present in the target stimuli. The mask used in the
experiment was made out of line segments of all orienta-
tions used in the experiments, eso es, 35°, 55°, 125°, y
145° with a red fixation dot in the middle of the screen.
A typical trial run started with presentation of the mask
para 250 mseg. Después, the target stimulus was shown
para 200 mseg, followed by the mask again for 1000 mseg.
Participants were required to make a saccade away from
the target as quickly as possible after target presentation,
and return fixation to the fixation dot afterward.
Stimuli were presented using the SR Research Experi-
ment Builder (SR Research, Osgoode, Canada) on a 17-in.
Iiyama CRT monitor, running at a 100-Hz refresh rate with
a resolution of 1024 × 768 píxeles. Participants were seated
approx. 1 meter away from the screen. The experiment
was run in 13 blocks of 160 trials each. Stimulus location
and orientation difference were randomized on a trial-to-
trial basis. Participants did 2080 trials in total.
EEG Recording and Analysis
EEG was recorded using a 48-channel ActiveOne EEG
(Biosemi, Los países bajos) system with active electrodes.
Horizontal and vertical EOG was measured to control for
eye blinks. The EEG signal was digitized and sampled
en 256 Hz on a separate acquisition computer. Markers were
sent with the EEG signal by the stimulation computer to al-
low for off-line segmentation. Per condition, three 10-min
blocks were recorded; each block contained 450 ensayos.
EEG data were analyzed using Analyzer (BrainVision Prod-
ucts GmbH, Alemania). The raw signal was filtered be-
entre 1 y 15 Hz and then segmented in epochs of
1200 mseg (200 msec prestimulus–1000 msec poststimu-
lus) on the basis of markers sent with visual stimulation.
Bad segments were rejected automatically when the max-
imum amplitude in a segment exceeded 100 μV. A base-
line correction (base 20 mseg) was performed for each
segmento. VEPs were computed for homogeneous and
checkerboard trials.
To compute the scene segmentation specific signal,
VEPs from homogeneous trials were subtracted from
checkerboard trial VEPs. The resulting traces were aver-
aged, thus resulting in a VEP representing neural activity
related to scene segmentation (ver Figura 1B). For both
the 90° and the 20° orientation differences, and per sub-
ject, peak latencies in this subtraction signal were com-
puted by finding the minimum amplitude in the interval
200–400 msec. Peak latencies for the P100 component
(for both 90 y 20 orientation contrasts) fueron calculados
by finding the maximum amplitude in the interval 90–
130 msec in the checkerboard VEPs. To test for differ-
ences between VEPs in different conditions, we used
paired t tests per sample, with an alpha of .05, Bonferroni-
corrected for the number of channels and samples com-
pared. We used SPSS version 16.0 for Windows (SPSS,
chicago) to compute Pearson correlations between peak
and onset latencies and RTs. All statistics are based on
the activity in channel Oz according to the International
10–20 System. This electrode was selected on the basis of
estudios previos (p.ej., Scholte et al., 2008; Caputo & Casco,
1999; Lamme, Van Dijk, & Spekreijse, 1992).
RTs were measured using a push-button and were
sampled via the parallel port of the stimulus computer
for millisecond-precision analysis. These data were used
to compute RTs. With every button press, a marker was
sent to the EEG acquisition computer as well. Porque
the EEG acquisition computer was sampled at 256 Hz,
these markers have a 4-msec temporal resolution. Estos
markers were used to segment data based on RT. Par-
ticipants were instructed to press the button with their
dominant hand.
Because the brain does not use VEP averaging and sub-
sequent statistical testing in order to detect the presence
of texture stimuli, we ran a single-trial analysis using a
nearest mean classifier approach in order to estimate the
amount of information needed to detect the presence of
a texture stimulus (Bandt, Weymar, Samaga, & Hamm,
2009). For this analysis, we used segmented EEG data from
a single channel (Oz), filtered between 0.5 y 100 Hz of
el 14 last participants in Experiment 1. This dataset was
selected for technical reasons.
Individual trials were normalized so that the mean ac-
tivity in each trial was zero. Templates for both individual
checkerboard textures and the homogeneous textures
were computed per participant, and individual trials were
classified based on a template function that reflects the
difference between checkerboard (C) and homogeneous
(h) estímulos (es decir., C − H). For each trial x, a classifier score
S was obtained by multiplying activity in each sample t
with this template function, and taking the mean over
the total number of samples T:
X
SðxÞ ¼ ð1=TÞ
ðCðtÞ − HðtÞÞ xðtÞ
To avoid autocorrelation, we adopted a “leave-one-out”
approach for computing the templates: For each trial N,
this trial was excluded from the templates used to classify
él. To determine the moment at which sufficient infor-
mation is available to make above-chance perceptual de-
decisiones, we systematically increased the number of data
points per trial fed into the algorithm. The moment of
above-chance level target detection was determined per
0, averaged over
target type (20° or 90°) by testing when d
all participants, exceeded 0.
Jolij et al.
3737
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
oh
h
w
t
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
r
t
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
r
mi
mi
r
C
t
C
.
metro
h
a
i
mi
r
d
.
tu
C
oh
oh
metro
C
norte
/
j
a
oh
r
t
C
i
C
norte
mi
/
–
a
pag
r
d
t
i
2
C
3
yo
1
mi
2
–
3
pag
7
d
3
F
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
oh
3
C
4
norte
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
pag
/
d
j
oh
b
C
y
norte
gramo
_
tu
a
mi
_
s
0
t
0
oh
0
norte
3
0
4
8
.
S
pag
d
mi
F
pag
mi
b
metro
y
b
mi
gramo
r
tu
2
mi
0
s
2
t
3
/
j
/
.
F
t
oh
norte
1
8
METRO
a
y
2
0
2
1
Eye Tracking
Eye movements in Experiment 3 were recorded with
a head-mounted eye-tracking system (Eyelink II, SR Re-
buscar, Osgoode, Canada) at a sample rate of 500 Hz.
Saccadic RTs were computed off-line by taking the mo-
ment of the first eye movement exceeding a velocity
de 30 deg /sec. Any saccades with a saccadic RT smaller
than 75 msec were discarded, as were saccades with a
saccadic RT greater than mean ± 3 × SD. Trials in which
participants blinked were excluded from the analysis. Sac-
cade direction was classified on the basis of the saccade
termination point; subsequently, saccades were classified
as antisaccades or prosaccades on the basis of information
of the target position.
RESULTADOS
Experimento 1: Differences in RT Correspond
to Latency Differences in Long-latency
VEP Components
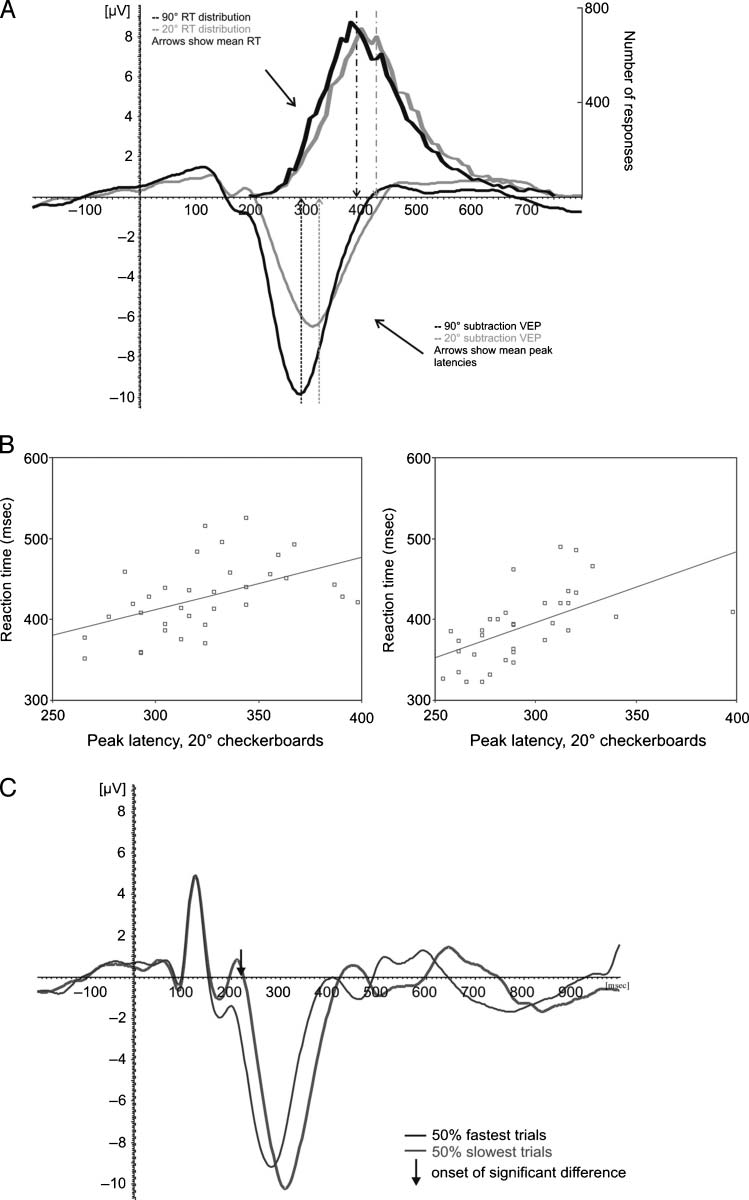
In the RT experiment (Experimento 1), reducing orienta-
tion contrast from 90° to 20° increased RTs from 391.1
a 427.6 mseg [t(32) = 21.4, pag = .000; see Figure 3A].
Detection performance was not affected by reducing orien-
tation contrast and was 97% for both conditions.
The subtraction potentials to both the 20° and 90° ori-
entation contrasts showed the characteristic pattern ob-
served in texture segmentation tasks: a small positivity
around 80–100 msec, followed by a large negative deflec-
tion between 200 y 300 mseg, localized in the posterior
electrodes. These two timeframes are linked respectively
to boundary detection and surface segregation (Scholte
et al., 2008; Vandenbroucke, Scholte, van Engeland, Lamme,
& Kemner, 2008).
Peak latencies of the scene segmentation-related VEP
components increased from 293.4 a 322.8 mseg [t(32) =
7.7, pag = .000; see Figure 2A]. Correlations between RTs
and latencies of scene segmentation signals of individual
subjects are highly significant: .51 ( pag = .002) y .68 ( pag =
.000) for the 20° and 90° orientation contrast checkerboard
textures, respectivamente (Figura 2B).
The relation between manual RT and peak latency of
scene segmentation-related VEP components showed up
even more explicitly in a reanalysis of the RT/EEG data.
We split trials in two groups of the 50% fastest versus
50% slowest trials and recomputed the VEPs for the 20°
and 90° checkerboards. Please note that for this analy-
sis we did not compute the segregation specific sig-
nal, but only took VEPs for checkerboard stimuli into
cuenta, because only in these trials was a manual re-
sponse given. The checkerboard VEPs were significantly
different only from 230 a 300 mseg ( pag < .05, corrected
for 256 samples), showing that differences in RTs are
explained by differences in relatively long-latency pro-
cesses, and not by differences in earlier cortical processing
(Figure 2C).
Finally, a single-trial analysis using a nearest mean clas-
sification algorithm (Bandt et al., 2009) revealed that suf-
ficient information is available in unfiltered single trials to
reliably detect presence of a texture after 113 msec for the
90° targets [t(13) = 2.38, p < .05], and after 117 msec for
the 20° targets [t(13) = 2.16, p < .05]. Only after 238 msec
was the classification performance better for 90° targets
than for 20° targets [t(13) = −2.33, p < .05], suggesting
that before that moment, the amount of information pres-
ent in a single trial does not differ for both types of targets
(Figure 3).
These results show a clear relation between scene seg-
mentation related components in the VEP and visually
guided behavior: Reducing orientation contrast had similar
effects on the latency of segregation-specific VEP compo-
nents and on RTs. The analysis of slow versus fast trials
shows that fast trials are accompanied by a latency decrease
of scene segmentation related VEP components, whereas
slow trials are characterized by an increased latency of
scene segmentation related VEP components. In all experi-
ments and analyses, earlier components of the VEP re-
mained unaffected: Latencies of the P100 did not show
any relation with RT, thus suggesting that in texture dis-
crimination tasks, visually guided behavior depends on
processes taking place approximately 175–275 msec after
stimulus presentation. As argued earlier, these processes
reflect scene segmentation, a process critically dependent
on recurrent interactions between higher and lower visual
areas, and possibly related to visual awareness (Scholte
et al., 2008; Heinen et al., 2005). As previous work has
shown that scene segmentation does not depend on at-
tention, it is unlikely that changes in attentional capture
can explain the pattern of results we observed (Scholte,
Witteveen, Spekreijse, & Lamme, 2006; Schubö, Meinecke,
& Schröger, 2001).
Experiment 2: Differences in Subjective
Simultaneity Correspond to Differences
in VEP Latency
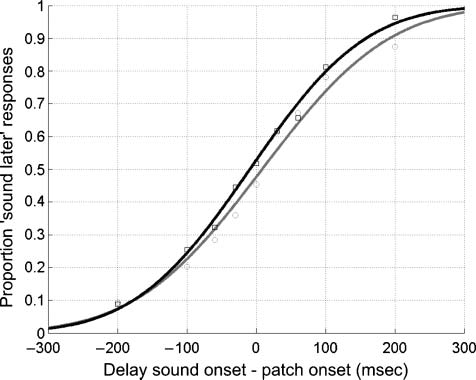
Points of subjective simultaneity (PSSs) for the onset of
a sound and the onset of checkerboards with 20° and
90° orientation contrast were determined in Experiment 2.
Differences in PSSs are believed to reflect differences in
perceptual latency (see Jaskowski, 1996). If the long-latency
VEP components we report in Experiments 1 do reflect the
perceived segmentation of a texture stimulus, one would
expect that comparing results from the 20° and 90° textures
would yield comparable effects on the VEP components
and PSSs.
We found a difference of 18 msec between PSSs, sug-
gesting that a 20° orientation contrast checkerboard is
perceived 18 msec later than a 90° orientation contrast
checkerboard (Figure 4). This difference was only margin-
ally significant, however; Monte Carlo analyses yielded a
p value of .09. Post hoc analyses showed that four of nine
individual subjects did show a significant difference of
3738
Journal of Cognitive Neuroscience
Volume 23, Number 12
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
2
-
3
p
7
d
3
f
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
o
3
c
4
n
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
3
0
4
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
.
f
t
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Differences in
RT correspond to differences
in visual processing speed.
(A) RT distributions and
subtraction VEPs (channel Oz)
for the 20° (gray lines) and 90°
(black lines) checkerboards.
Population averages are
indicated by dotted arrows.
Differences in mean RTs and
mean peak latencies are almost
identical. (B) Correlations
between RTs and VEP latencies.
Latency of scene segmentation
related VEP components of
each subject is plotted against
manual RT. Circles show
individual data points, lines
show best linear fit. (C) VEPs
for the 50% fastest (black
trace) and 50% slowest (gray
trace) trials, 90° orientation
contrast textures. Channel
Oz is shown. Black arrow
indicates onset of the significant
difference ( p < .05, corrected
for 256 samples). Earlier
components do not differ,
whereas later VEP components,
related to scene segmentation,
are slower in “slow” trials
than in “fast” trials.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
2
-
3
p
7
d
3
f
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
o
3
c
4
n
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
3
0
4
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
.
f
o
n
1
8
M
a
y
2
0
2
1
around 30 msec in PSSs; data of two subjects did not show
any difference; the data of the remaining two subjects
were too noisy to reliably estimate the PSS. The differ-
ences in PSSs (ca. 18 msec) we report here correspond well
to the differences in latencies in scene segmentation re-
lated components we report in Experiment 1 (24 msec),
suggesting that differences in PSSs between 20° and 90°
orientation contrast checkerboards are to be attributed
to differences in latencies of scene segmentation related
processing.
Jolij et al.
3739
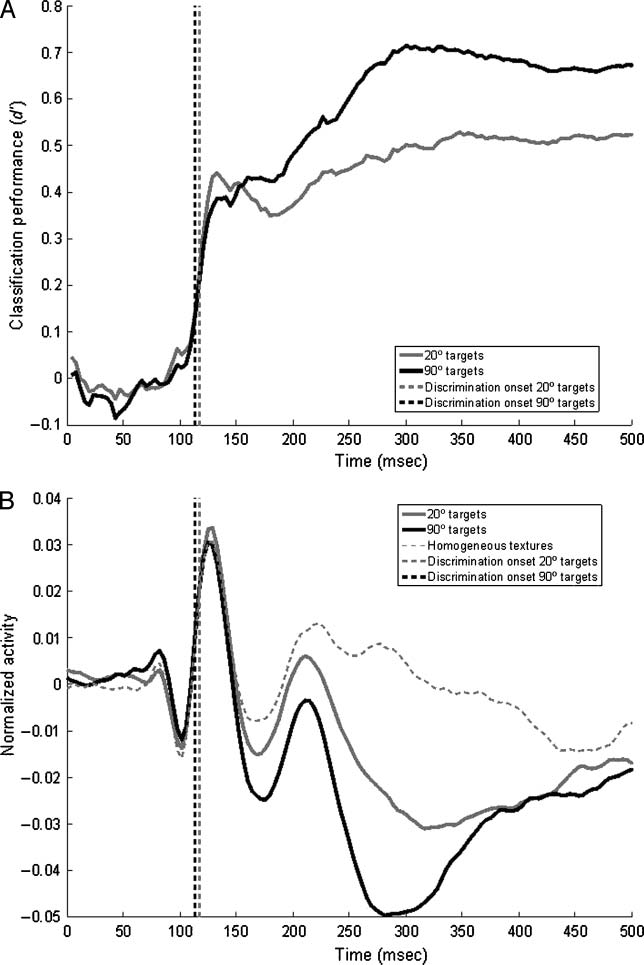
Experiment 3: Reflexive Behavior May Be Triggered
by Early Visual Processing
In the antisaccade experiment, participants failed to in-
hibit the tendency to make an eye movement toward the
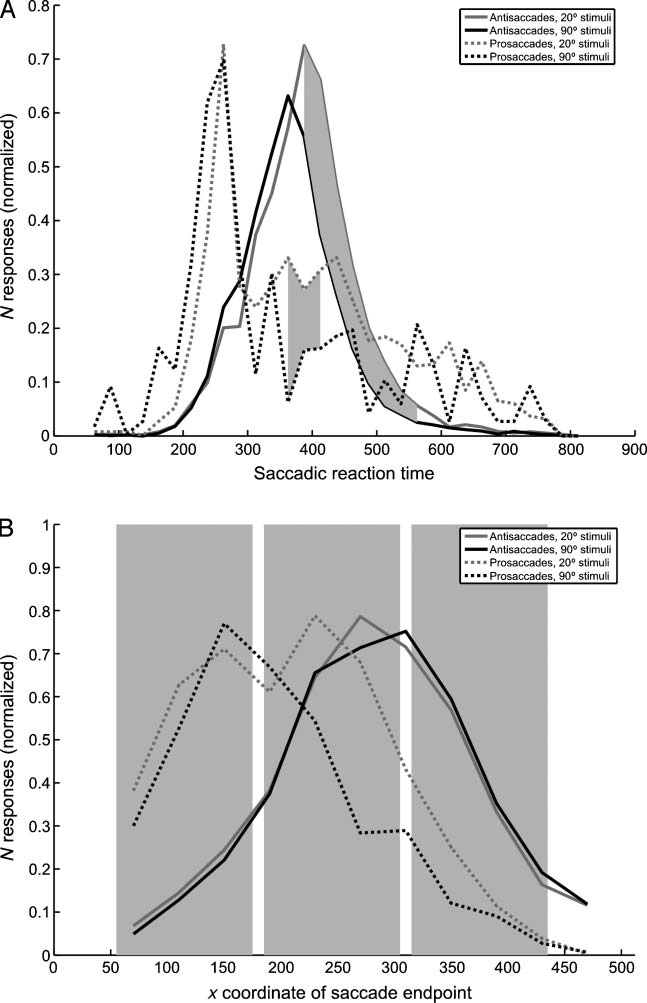
stimulus in 12.7% of the trials, and made a saccade to
the target stimulus. We computed RTs for antisaccades
and erroneous prosaccades for the 20° and 90° orientation
difference stimuli. Antisaccades based on 90° orientation
contrast textures (mean RT 364.39 msec) were 27.19 msec
faster than on 20° orientation contrast textures (mean RT =
391.58 msec) [t(10) = 8.37, p = .000], a difference strik-
ingly similar to the latency difference in long-latency tex-
ture processing related evoked potential components for
these stimuli (approx. 30 msec) reported in Experiment 1.
For the prosaccades, subjects were 47.91 msec faster
in making a saccade toward the 90° orientation con-
trast stimuli (mean RT = 327.24 msec) than to the 20°
orientation contrast stimuli (mean RT = 375.14 msec)
[t(10) = 4.12, p = .000]. However, the distribution of
RTs (Figure 5A) reveals that the RT distribution of erro-
neous saccades toward 20° orientation contrast stimuli
is, in fact, bimodal, and that the peaks of the RT distribu-
tions of erroneous saccades toward 20° and 90° orienta-
tion contrast stimuli overlap. Indeed, paired t tests reveal
no significant differences between the RT distributions
(25 msec bins) up to 325 msec, but between 350 and
400 msec significantly more prosaccades are made toward
20° targets than to 90° targets. Apparently, there are two
types of erroneous saccades—one generated on the basis
0 units. Vertical
Figure 3. Single-trial
classification analysis for 20°
and 90° orientation difference
textures. (A) Classification
performance of the nearest
mean classifier algorithm
for 20° (gray line) and
90° textures (black line)
based on a single-trial analysis
of segmented EEG data
expressed in d
lines indicate onset of
above-chance discrimination
for 20° and 90° textures (gray
and black lines, respectively).
(B) Template functions (i.e.,
normalized VEPs) used for
the classification algorithm
for 20° textures (gray line),
90° textures (black line),
and homogeneous textures
(dotted line). Vertical lines
again indicate onset of
above-chance discrimination
for targets.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
2
-
3
p
7
d
3
f
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
o
3
c
4
n
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
3
0
4
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
f
.
t
o
n
1
8
M
a
y
2
0
2
1
3740
Journal of Cognitive Neuroscience
Volume 23, Number 12
most the same amount of time as the latency of long-latency
VEP components when the perceived strength of segmen-
tation is decreased, strongly suggesting that both respond-
ing to and conscious perception of a texture require the
processes underlying these VEP components. However, re-
flexive behavior seems to be based on earlier stages of in-
formation processing, as indicated by the absence of a
difference in RT for erroneous prosaccades to 20° and 90°
targets. A single-trial analysis of EEG data shows that suffi-
cient information is indeed processed approx. 130 msec
after stimulus onset to reliably detect the presence of a tar-
get. Summarizing, we can state that long-latency visual pro-
cesses, linked to visual awareness, do play an important role
in detecting and responding to visual events in our study.
Interestingly, this suggests that instead of accumulating
evidence from the earliest moment of sensory processing,
neural decision-making systems seem to rely on visual
representations that are formed at longer latencies. In
the literature, there is growing support for the accumulator
model of perceptual decision-making (see, e.g., Ratcliff &
McKoon, 2008, for a recent review). In the accumulator
model, sensory evidence is accumulated over time until
an internal threshold is reached and a motor response is
initiated. A recent neuroimaging study in humans demon-
strated that a network of occipital, parietal, and prefrontal
areas is involved in this process. Critically, neurons in the
intraparietal sulcus seem to function as accumulators of
perceptual evidence, but exactly what feeds into this ac-
cumulating system, in terms of visual information, remains
unspecified (Kayser, Buchsbaum, Erickson, & DʼEsposito,
2010). Our experiments seem to suggest that only visual in-
formation that has been processed quite extensively feeds
into the decision-making process. However, this seems to
contrast with earlier findings, suggesting that perceptual
decisions can be made extremely fast. How can we explain
this discrepancy?
As argued, a large amount of information appears to
be extracted from visual input during the earliest stages
of visual processing (see e.g., Thorpe et al., 1996). Recent
studies suggest that visual information processing under-
lying rapid image classification is based on the statistical
properties of visual input: The majority of variability in
EEG responses up to approx. 100 msec after stimulus pre-
sentation can be explained by the distribution of contrast
values of an image. Interestingly, this underlying statistical
structure contains a large amount of ecologically meaningful
information about the general lay-out of a visual scene—for
example, whether a scene is cluttered, or contains just one
single object. It has been proposed that the statistical struc-
ture of visual input provides the system with sufficient in-
formation to do coarse categorization tasks. However, if
such statistical structure is absent, deeper processing is re-
quired for perceptual decision-making (Scholte, Ghebreab,
Waldorp, Smeulders, & Lamme, 2009; VanRullen & Koch,
2003; Lamme & Roelfsema, 2000; Thorpe et al., 1996).
A similar conceptual distinction between coarse cate-
gorization versus more precise stimulus identification has
Jolij et al.
3741
Figure 4. Differences in PSSs for 20° and 90° orientation differences.
Black line (probit fit) and symbols show the proportion of “sound
later” responses for 90° checkerboards; gray line (probit fit) and
symbols show the proportion of “sound later” responses for 20°
checkerboards.
of an early signal, and a second, smaller subset, based on
a later signal.
The distribution of antisaccade RTs, on the other hand,
shows two distinct peaks around 350–400 msec for anti-
saccades based on 90° and 20° orientation contrast stimuli,
with significantly more antisaccades away from 20° stimuli
(gray area in Figure 2A; p < .01) for bins later than 400 msec,
suggesting that these saccades are based on a relatively
long-latency visual signal. The different RT patterns for anti-
saccades and prosaccades show that they are based on dif-
ferent visual representations. The absence of a difference
in peak RTs for 20° and 90° textures suggests that most
prosaccades are based on detection of borders, around
100 msec. This is corroborated by the fact that most pro-
saccades to the texture stimuli are made toward the texture
squares closest to fixation, which would have captured
attention and evoked eye movements (Figure 5B).
On the other hand, the striking similarity between the
latency difference in the peak latencies for antisaccades
toward 20° and 90° stimuli and the VEP components re-
flecting filling-in of 20° and 90° stimuli strongly suggests
that antisaccades, and a small portion of erroneous pro-
saccades is based on the representation of a filled-in fig-
ure. Apparently, making an antisaccade requires more
than simply inhibiting a prosaccade and reprogramming
the eye movement: Antisaccades actually require deeper
visual processing of the stimulus on which they are based
than prosaccades.
DISCUSSION
In three experiments, we have investigated the relation
between behavior, perception, and neural processing using
texture stimuli. Both simple RTs and PSSs increase by al-
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
2
-
3
p
7
d
3
f
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
o
3
c
4
n
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
3
0
4
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
t
f
o
n
1
8
M
a
y
2
0
2
1
been suggested in the context of feedforward versus re-
current processing in object-sensitive visual areas. Neurons
in temporal cortex, for example, respond to faces, in gen-
eral, 150 msec after stimulus presentation; however, ap-
proximately 50 msec later, the same neurons respond to
individual faces (Sugase, Yamane, Ueno, & Kawano,
1999). Neural network models of texture segregation tasks
show a similar behavior: Global discriminations between
shapes can be done on the basis of a single feedforward
sweep from lower to higher layers in the network, analo-
gous to the feedforward transfer of visual information
from lower-tier to higher visual areas. However, in order
to allow for finer discriminations, for example, between
two exemplars of the same category, feedback process-
ing from higher to lower visual areas seems to be required
in order to obtain sufficient spatial detail ( Jehee et al.,
2007; Hochstein & Ahissar, 2002). In addition, feedback
processing may also provide a mechanism to highlight in-
formation that is relevant for a certain task (Roelfsema,
Lamme, & Spekreijse, 1998).
In general, we could state that visual information pro-
cessing may be characterized as a two-stage process: Dur-
ing the first stage, coarse information based on low-level
features and image statistics is extracted. This information
may be sufficient to trigger perceptual decisions, depend-
ing on the task at hand. However, in order to do finer
Figure 5. (A) Averaged
distribution of pro- and
antisaccadic RTs. y-axis gives
the number of responses
given in a given 20-msec bin,
normalized per participant.
Dotted lines show prosaccades,
solid lines show antisaccades;
gray lines show responses to
20° orientation contrast stimuli,
black lines show responses
to 90° orientation contrast
stimuli. Shaded area denotes a
significant difference between
the number of responses in
a given bin for 20° vs. 90°
stimuli, p < .05 (corrected
for multiple comparisons).
(B) Averaged distribution of
saccade termination points.
y-axis gives the number
of responses in a given
40 pixel bin, collapsed over
both leftward and rightward
saccades, x-axis gives horizontal
position. Further legend as
in (A). Shaded areas indicate
horizontal position of texture
squares.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
2
-
3
p
7
d
3
f
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
o
3
c
4
n
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
3
0
4
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
.
t
f
o
n
1
8
M
a
y
2
0
2
1
3742
Journal of Cognitive Neuroscience
Volume 23, Number 12
discrimination, such as within-category discriminations, ad-
ditional processing is required, and perceptual decision-
making will require more time (VanRullen, Reddy, & Koch,
2004; Hochstein & Ahissar, 2002). Although this model
seems plausible, and may be able to explain why in some
tasks perceptual decisions are based on coarse repre-
sentations, it does not completely explain the behavior
of participants in our experiments. In our study, we may
assume that information extracted up to approximately
130 msec after stimulus presentation should be sufficient
to trigger a behavioral response (Experiment 1), and
indeed, we find that reflexive saccades may, in fact, be
triggered by this early cortical processing (Experiment 3).
However, initiation of a simple button press, or an anti-
saccade, appears to be based on longer-latency visual pro-
cessing, despite the fact that coarse information is present
in the system.
We theorize that the selection of what information feeds
into the perceptual decision-making systems is not simply
a matter of using any information that is available. Instead,
there seems to be a preference for using information that
represents the visual environment with high spatial detail
when such information, in general, is available (see also
Ratcliff & Smith, 2010; Jolij, 2008; Jolij & Lamme, 2005). In
our experiments, all stimuli were presented long enough
to be clearly seen by the participants. Although partici-
pants were instructed to respond as fast as possible in the
RT and antisaccade tasks, we did not press for faster
responses by penalizing slow responses. Therefore, it is
very well possible that participants employed a strategy
in which they waited until they consciously perceived the
target, and then pressed the button or made an eye move-
ment. Given the very good performance on both tasks,
this is quite likely. However, in the studies by VanRullen
et al. (2004), briefly presented targets were used, which in
some trials were even masked. Therefore, overall visibility
was much lower. In fact, they even report that some partici-
pants in their study could reliably detect targets in the
absence of visual awareness, resembling a blindsight-like
condition. We propose that the poor overall quality of
visual information may have triggered the perceptual deci-
sion network to start accumulating evidence as soon as
sensory information became available, instead of waiting
for a more precise representation that does not become
available as a result of masking (Fahrenfort et al., 2007;
Lamme, Zipser, & Spekreijse, 2002).
In a study on blindsight in normal participants using
TMS, Jolij and Lamme (2005) report a very similar contrast
between availability of unconscious (or preconscious) in-
formation for perceptual decision-making. In this study,
TMS of primary visual cortex was used to induce artifi-
cial blindsight in normal observers. Interestingly, subjects
showed the blindsight capability only in a context in which
the stimuli were generally hard to see, and the subjects
were mostly guessing even without magnetic stimulation.
When stimulus visibility was increased, and subjects could
base their response on their conscious percept, the blind-
sight capability disappeared, probably because they ignored
unconsciously processed information. This suggests that, in
a context where conscious representations are available,
these are preferred over unconscious representations to
initiate a response, most likely due to the fact that conscious
representations have a higher accuracy ( Jolij, 2008).
Interestingly, not all visually guided behavior seems to
be governed by such a mechanism. In Experiment 3, we
show that reflexive prosaccades to texture targets are most
likely to be based on an early border detection signal, and
not on a longer-latency signal, whereas antisaccades are
based on long-latency visual processes. This fits with the
idea that task demands also play a role in selecting what
information to use in perceptual decision-making. Reflex-
ive saccades are evoked by suddenly appearing targets that
may be highly relevant for an organism; the motor pro-
gram to execute such a saccade can be programmed and
executed quickly. A false alarm in such a case may not be
too harmful: An eye movement has no consequences in
the “external world.”
However, the relative harmlessness of an erroneous eye
movement may not be sufficient to explain the pattern of
results concerning the effects of unconscious visual rep-
resentations on visually guided behavior observed in the
literature. In several studies, Schmidt and Schmidt (2009)
and Schmidt, Niehaus, and Nagel (2006) investigated the
effects of unconsciously (or preconsciously) processed
information on pointing movements: They found that
the initial trajectory of a pointing movement is guided by
feedforward-processed visual information. Interestingly,
both the results obtained by Schmidt and coworkers, and
the results we obtained in Experiment 3 share one critical
feature: Both prosaccades and pointing movements do
not require a spatial transformation or remapping of a re-
sponse, and are completely stimulus-driven.
A manual response on a response box, or even more
complex, cognitively guided behavior, such as antisaccades
on the other hand, requires such a spatial transformation.
Pressing a button on a response box when you see a stim-
ulus does require a specific stimulus–response mapping
that bears no direct relation with the stimulus, contrary
to a prosaccade or a pointing movement. It is well possi-
ble that such a response remapping requires cognitive ef-
fort and deeper visual processing of a stimulus to avoid
errors that may induce slowing in the cognitive control
system (Ridderinkhof, Van Den Wildenberg, Segalowitz,
& Carter, 2004). However, this hypothesis needs to be
experimentally verified.
Summarizing, we believe that our results are in line with
the hypothesis that the normal modus operandi of human
observers is to base a visually guided response on a more
elaborated, and most likely conscious, representation of
the environment, even though response generation on
the basis of quickly formed unconscious representations
is very well possible. However, these latter representations
are only used to guide behavior when necessary, for ex-
ample, when a fast response is required (e.g., in fight–flight
Jolij et al.
3743
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
2
-
3
p
7
d
3
f
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
o
3
c
4
n
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
3
0
4
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
t
f
.
o
n
1
8
M
a
y
2
0
2
1
situations or in reflexive saccades), or when a conscious
representation is not available as a result of a lesion, or in
experimental manipulations such as TMS or masking.
Acknowledgments
We thank Danielle Huisman and Renske Bijl for help with data col-
lection and two anonymous referees for their helpful comments.
J. J. designed the experiments, collected and analyzed the data,
and wrote the article; H. S. S. analyzed the data; S. v. G. collected
the data; T. L. H. designed the experiments and analyzed the
data; V. A. F. L. designed the experiments and wrote the article.
Part of this research was funded by NWO (Dutch Organization
for Scientific Research) grant 425-20-307 to V. A. F. L.
Reprint requests should be sent to Jacob Jolij, Department of
Experimental Psychology, Faculty of Behavioural and Social
Sciences, University of Groningen, Grote Kruisstraat 2/1, 9712
TS Groningen, The Netherlands, or via e-mail: j.jolij@rug.nl.
REFERENCES
Bandt, C., Weymar, M., Samaga, D., & Hamm, A. O. (2009).
A simple classification tool for single-trial analysis of ERP
components. Psychophysiology, 46, 747–757.
Boyer, J. L., Harrison, S., & Ro, T. (2005). Unconscious
processing of orientation and color without primary visual
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 102, 16875–16879.
Caputo, G., & Casco, C. (1999). A visual evoked potential
correlate of global figure–ground segregation. Vision
Research, 39, 1597–1610.
Fahrenfort, J. J., Scholte, H. S., & Lamme, V. A. F. (2007).
Masking disrupts reentrant processing in human
visual cortex. Journal of Cognitive Neuroscience,
19, 1488–1497.
Goodale, M. A., & Milner, A. D. (1992). Separate visual pathways
for perception and action. Trends in Neurosciences, 15,
20–25.
Heinen, K., Jolij, J., & Lamme, V. A. F. (2005). Figure–ground
segregation requires two distinct periods of activity in V1:
A transcranial magnetic stimulation study. NeuroReport,
16, 1483–1487.
Hochstein, S., & Ahissar, M. (2002). View from the top:
Hierarchies and reverse hierarchies in the visual system.
Neuron, 36, 791–804.
Jaskowski, P. (1996). Simple reaction time and perception
of temporal order: Dissociations and hypotheses.
Perceptual and Motor Skills, 82, 707–730.
Jolij, J., & Lamme, V. A. F. (2005). Repression of unconscious
information by conscious processing: Evidence from affective
blindsight induced by transcranial magnetic stimulation.
Proceedings of the National Academy of Sciences, U.S.A.,
102, 10747–10751.
Kayser, A. S., Buchsbaum, B. R., Erickson, D. T., & DʼEsposito,
M. (2010). The functional anatomy of a perceptual decision
in the human brain. Journal of Neurophysiology, 103,
1179–1194.
Lamme, V. A. F. (1995). The neurophysiology of figure–ground
segregation in primary visual cortex. Journal of
Neuroscience, 15, 1605–1615.
Lamme, V. A. F. (2003). Why visual attention and awareness
are different. Trends in Cognitive Sciences, 7, 12–18.
Lamme, V. A. F., & Roelfsema, P. R. (2000). The different
modes of vision offered by feedforward and recurrent
processing. Trends in Neurosciences, 23, 571–579.
Lamme, V. A. F., Van Dijk, B. W., & Spekreijse, H. (1992).
Texture segregation is processed by primary visual cortex
in man and monkey: Evidence from VEP experiments.
Vision Research, 32, 797–807.
Lamme, V. A. F., Zipser, K., & Spekreijse, H. (2002).
Masking interrupts figure-ground signals in V1.
Journal of Cognitive Neuroscience, 14, 1044–1053.
Ratcliff, R., & McKoon, G. (2008). The diffusion decision
model: Theory and data for two-choice decision tasks.
Neural Computation, 20, 873–922.
Ratcliff, R., & Smith, P. M. (2010). Perceptual discrimination
in static and dynamic noise: The temporal relation
between perceptual encoding and decision making.
Journal of Experimental Psychology: General, 139,
70–94.
Ridderinkhof, K. R., Van Den Wildenberg, W. P., Segalowitz,
S. J., & Carter, C. S. (2004). Neurocognitive mechanisms
of cognitive control: The role of prefrontal cortex in action
selection, response inhibition, performance monitoring,
and reward-based learning. Brain and Cognition, 56,
129–140.
Roelfsema, P. R., Lamme, V. A. F., & Spekreijse, H. (1998).
Object-based attention in the primary visual cortex of
the macaque monkey. Nature, 395, 376–381.
Roelfsema, P. R., Lamme, V. A. F., Spekreijse, H., & Bosch, H.
(2002). Figure–ground segregation in a recurrent network
architecture. Journal of Cognitive Neuroscience, 14,
525–537.
Scharnowski, F., Rüter, J., Jolij, J., Hermens, F., Kammer, T.,
& Herzog, M. H. (2009). Long-lasting modulation of feature
integration by transcranial magnetic stimulation. Journal
of Vision, 9, 1–10.
Schmidt, T., Niehaus, S., & Nagel, A. (2006). Primes and targets
in rapid chases: Tracing sequential waves of motor activation.
Behavioral Neuroscience, 120, 1005–1016.
Jehee, J. F., Lamme, V. A. F., & Roelfsema, P. R. (2007).
Schmidt, T., & Schmidt, F. (2009). Processing of natural
Boundary assignment in a recurrent network architecture.
Vision Research, 47, 1153–1165.
Jehee, J. F., Roelfsema, P. R., Deco, G., Murre, J. M., &
Lamme, V. A. F. (2007). Interactions between higher and
lower visual areas improve shape selectivity of higher
level neurons-explaining crowding phenomena.
Brain Research, 157, 167–176.
Jolij, J. (2008). From affective blindsight to affective blindness:
When cortical processing suppresses subcortical information.
In F. Columbus (Ed.), Neural pathways: New research
(pp. 205–208). New York, NY: Nova Science Publishers.
Jolij, J., Huisman, D., Scholte, H. S., Hamel, R., Kemner, C.,
& Lamme, V. A. F. (2007). Processing speed in recurrent
networks correlates with general intelligence. NeuroReport,
18, 39–43.
images is feedforward: A simple behavioral test. Attention,
Perception, and Psychophysics, 71, 594–606.
Scholte, H. S., Ghebreab, S., Waldorp, L., Smeulders, A. W., &
Lamme, V. A. F. (2009). Brain responses strongly correlate
with Weibull image statistics when processing natural images.
Journal of Vision, 9, 1–15.
Scholte, H. S., Jolij, J., Fahrenfort, J. J., & Lamme, V. A. F.
(2008). Feedforward and recurrent processing in scene
segmentation: Electroencephalography and functional
magnetic resonance imaging. Journal of Cognitive
Neuroscience, 20, 2097–2109.
Scholte, H. S., Witteveen, S. C., Spekreijse, H., & Lamme,
V. A. F. (2006). The influence of inattention on the neural
correlates of scene segmentation. Brain Research, 1076,
106–115.
3744
Journal of Cognitive Neuroscience
Volume 23, Number 12
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
2
-
3
p
7
d
3
f
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
o
3
c
4
n
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
3
0
4
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
f
.
/
o
n
1
8
M
a
y
2
0
2
1
Schubö, A., Meinecke, C., & Schröger, E. (2001). Automaticity
and attention: Investigating automatic processing in texture
segmentation with event-related brain potentials. Brain
Research, Cognitive Brain Research, 11, 341–361.
Sugase, Y., Yamane, S., Ueno, S., & Kawano, K. (1999). Global
and fine information coded by single neurons in the temporal
visual cortex. Nature, 400, 869–873.
Supèr, H., Lamme, V. A. F., & Spekreijse, H. (2001). Two
distinct modes of sensory processing observed in monkey
primary visual cortex ( V1). Nature Neuroscience, 4, 304–310.
Thorpe, S., Fize, D., & Marlot, C. (1996). Speed of processing
in the human visual system. Nature, 381, 520–522.
Vandenbroucke, M. W., Scholte, H. S., van Engeland, H.,
Lamme, V. A. F., & Kemner, C. (2008). A neural substrate
for atypical low-level visual processing in autism spectrum
disorder. Brain, 131(Pt 4), 1013–1024.
VanRullen, R., & Koch, C. (2003). Visual selective behavior
can be triggered by a feedforward process. Journal of
Cognitive Neuroscience, 15, 209–217.
VanRullen, R., Reddy, L., & Koch, C. (2004). Visual search
and dual tasks reveal two distinct attentional resources.
Journal of Cognitive Neuroscience, 16, 4–14.
Weiskrantz, L. (1996). Blindsight revisited. Current Opinion
in Neurobiology, 6, 215–220.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
2
-
3
p
7
d
3
f
4
/
1
2
9
3
4
/
2
1
6
2
0
/
9
3
7
o
3
c
4
n
_
/
a
1
_
7
0
7
0
7
0
1
3
0
4
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
3
0
4
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
1
8
M
a
y
2
0
2
1
Jolij et al.
3745