Asimetría hemisférica en la remapeo y
Maintenance of Visual Saliency Maps: A TMS Study
Martijn Gerbrand van Koningsbruggen1, Shai Gabay2, Ayelet Sapir1,
Avishai Henik2, and Robert D. Rafal1
Abstracto
■ Parietal cortex has been implicated in the updating, after eye
movimientos, of a salience map that is required for coherent visual
experience and for the control of visually guided behavior. El
current experiment investigated whether TMS over anterior intra-
parietal cortex (AIPCx), just after a saccade, would affect the ability
to update and maintain a salience map. In order to generate a
salience map, we employed a paradigm in which an uninforma-
tive cue was presented at one object in a display to generate in-
hibition of return (IOR)—an inhibitory tag that renders the cued
object less salient than others in the display, and that slows sub-
sequent responses to visual transients at its location. Following
the cue, participants made a saccade to either left or right, y
we then probed for updating of the location of IOR by measuring
manual reaction time to targets appearing at cued location of the
cued compared to an uncued object. Between the time of saccade
initiation and target appearance, dual-pulse TMS was targeted
over right (Experimento 1) or left AIPCx (Experimento 2), y un
vertex control side. Updating of the location of IOR was elimi-
nated by TMS over right, but not the left, AIPCx, suggesting that
right parietal cortex is involved in the remapping of IOR. Re-
mapping was eliminated by right AIPCx, independientemente de si
the saccade was made to the left (contralateral), or right (ipsi-
lateral) visual field, and regardless of which field the target ap-
peared in. We conclude that right AIPCx is the neural substrate
for maintaining a salience map across saccades, and not simply
for propagating an efference copy of saccade commands. ■
INTRODUCCIÓN
Although the retinal input changes dramatically with every
eye movement, our visual experience is coherent. Este
consistency is hypothesized to be achieved by a remapping
mechanism that uses corollary discharge as an “extra-retinal
signal” to compensate for each saccade (verano & Wurtz,
2008). This remapping mechanism integrates information
across successive fixations into a spatial consistent percept.
The current experiments investigated the role of human
parietal cortex in remapping visual saliency maps by apply-
ing TMS just after an eye movement.
Duhamel, Colby, and Goldberg (1992) reported the first
neurophysiological evidence of neurons in monkey lateral
intraparietal (LIP) cortex that remapped their receptive
fields either before or after eye movements. During a task
requiring continuous fixation, neurons in LIP only respond
to visual stimuli presented within their retinotopic recep-
tive field. Sin embargo, in an experiment that involved eye
movimientos, in which a stimulus was presented outside
a neuronʼs receptive field, and in which the monkey was
instructed to make a saccade which would bring the stimu-
lus into the receptive field, a subset of the LIP neurons
responded to stimuli at the location of the future receptive
1Bangor University, Reino Unido, 2Ben-Gurion University of the Negev,
Beer-Sheva, Israel
field before saccade initiation. Subsequent studies have
reported neurons with similar properties in monkey su-
perior colliculus (Caminante, Fitzgibbon, & Goldberg, 1995),
frontal eye field (FEF; Umeno & Goldberg, 1997, 2001),
striate, and extrastriate cortex (Nakamura & Colby, 2002).
More important to the current investigation, Duhamel,
Colby, et al. (1992) and Duhamel, Goldberg, Fitzgibbon,
Sirigu, and Grafman (1992) also observed LIP neurons
that responded to “remembered” targets. When a briefly
flashed stimulus was presented outside their receptive
fields before a saccade, the neurons responded after the
saccade brought this location into their receptive field, incluso
though this location no longer contained the stimulus.
Duhamel et al. concluded that visual memory has a retino-
topic representation, which is updated after every saccade.
More recently, it has been shown that LIP neurons do not
simply remap visual stimuli but, more specifically, remap
the saliency of the visual stimulus (Gottlieb, Kusunoki, &
Goldberg, 1998).
There is converging evidence implicating human parietal
cortex in saccade updating from neuropsychological, TMS,
and fMRI investigations. Patients with lesions in parietal
cortex are impaired when executing the second saccade
of a double-step saccade task. en esta tarea, two saccades
are made to sequentially flashed targets, each of which dis-
appears before the first eye movement. The first saccade can
be made on the basis of retinotopic coordinates. Sin embargo,
© 2009 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 22:8, páginas. 1730–1738
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
8
mi
–
1
pag
7
d
3
F
0
/
1
2
9
2
3
/
9
8
4
/
8
1
9
7
oh
3
C
0
norte
/
1
2
0
7
0
7
9
0
2
2
1
1
4
3
5
/
6
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
3
5
0
6
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
t
.
.
.
F
/
oh
norte
1
8
METRO
a
y
2
0
2
1
an accurate second saccade requires updating of the loca-
tion of the second target based on the motor vector of the
first saccade. Failure to update the location faithfully results
in inaccurate saccades to the second target or, if no extra-
retinal signal is generated at all, the second saccade either
cannot be executed at all, or will be made to the retinal
location of the target. In a task in which the two targets
were presented in opposite visual fields, patients with
both left and right parietal lesions were impaired in the
second saccade when the first was directed contralesionally
(Heide, Blankenburg, Zimmermann, & Kompf, 1995). Re-
cent fMRI studies have confirmed and extended the involve-
ment of parietal cortex in spatial updating (Medendorp,
Goltz, & Vilis, 2005, 2006; Medendorp, Goltz, Vilis, &
Crawford, 2003; Merriam, Genovese, & Colby, 2003).
Several TMS studies have investigated the role of parietal
cortex in remapping. Because the current study uses TMS,
these studies are discussed in more detail below. camioneta
Donkelaar and Muri (2002) found that right parietal TMS
150 mseg, but not earlier, after the onset of the first saccade
of a double-step paradigm impaired accuracy of the second
saccade. Right TMS only impaired performance if the first
saccade was to the left and the second saccade to the right.
morris, Chambers, and Mattingley (2007) used a more focal
figure-of-eight coil and found that a posterior part of the
intraparietal sulcus (IPS), close to the transverse occipital
sulcus, and not a more anterior part, was involved in updat-
ing in a variant of the double-step paradigm. Other studies
have found that TMS over parietal cortex also impairs the
detection of displacement of visual targets that moved
during a saccade (Chang & Ro, 2007) and transsaccadic
memory of visual feature (Prime, Vesia, & Crawford, 2008).
In an experiment motivating the current research, Sapir,
Hayes, Henik, Danziger, and Rafal (2004) demonstrated
that human parietal cortex is also involved in updating
visual saliency maps across eye movements. To generate
a salience map, the experiment employed an paradigm in
which an exogenous cue engendered an inhibitory tag
at the location of the cue, resulting in slower responses
to targets presented at the previously cued location (inhibi-
tion of return, IOR; posner, Rafal, Choate, & Vaughn, 1985).
IOR has been hypothesized to contribute to the elaboration
of a salience map that can guide efficient visual exploration
by favoring novel locations (Klein, 1988, 2000; Posner et al.,
1985). Sapir et al. (2004) exploited the fact that the location
of this inhibitory tag is updated after a saccade (Tipper,
Grison, & Kessler, 2003; Danziger, Fendrich, & Rafal, 1997;
Maylor & Hockey, 1985; posner & cohen, 1984).
In the IOR paradigm employed by Sapir et al. (2004), uno
of four boxes was briefly cued to generate an inhibitory tag
y, after a saccade was made to a new location, a target
requiring a manual detection response was presented at
either the retinal location of the cue, the environmental
location of the cue, or at corresponding uncued locations.
They tested five patients with a unilateral lesion involving
superior parietal cortex and healthy controls. Healthy con-
trol participants showed inhibitory tagging (IOR) at the re-
mapped, environmental location of the cue, as well as a
smaller inhibitory effect at the retinal location. The patientsʼ
results revealed no evidence of updating the location of the
inhibitory tag, eso es, IOR was observed only at the retinal
location of the cue.
In contrast to the results of double-step saccade para-
digms summarized above, the deficit in remapping was bi-
lateral; eso es, it occurred for targets in both the ipsilesional
and contralesional visual field, and after both ipsilesional
and contralesional saccades. Curiosamente, abolished re-
mapping of the inhibitory tag was found in the three pa-
tients with a right hemisphere lesion but not in the two
left hemisphere patients. Sin embargo, due to the small sam-
ple size, this was not statistically tested. Sapir et al. (2004)
interpreted their results to indicate that parietal cortex
was not simply the source of the corollary discharge that
provides the extra-retinal signal for saccade remapping,
but that it may also provide the neural substrate for main-
taining a salience map across saccades.
The patients studied by Sapir et al. (2004) all had chronic
lesiones. It is not clear whether the effects reported in those
patients reflect the normal function of parietal cortex, o
are the consequence of brain reorganization. Además,
Sapir et al. studied only two patients with left parietal
lesions and three with right parietal lesions, and therefore,
could not draw definitive conclusions about possible hemi-
spheric asymmetries for maintaining salience maps. Aquí
we employed dual-pulse TMS to transiently disrupt the
function of parietal cortex, and to compare the effects of
right and left parietal TMS in order to test for a hemispheric
asymmetry in two experiments: In Experiment 1, TMS was
applied over right parietal cortex, whereas TMS was given
over left parietal cortex in Experiment 2. The parietal stimu-
lation site, over superior parietal cortex, corresponded to
the area of lesion overlap in the patients studied by Sapir
et al. A TMS vertex control site was also stimulated in
each experiment. The timing of the TMS pulses, 150 y
250 msec after saccade onset, was based on the observa-
tions of van Donkelaar and Muri (2002).
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
8
mi
–
1
pag
7
d
3
F
0
/
1
2
9
2
3
/
9
8
4
/
8
1
9
7
oh
3
C
0
norte
/
1
2
0
7
0
7
9
0
2
2
1
1
4
3
5
/
6
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
3
5
0
6
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
/
.
t
.
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
MÉTODOS
Participantes
Fourteen subjects (9 women) participated in Experiment 1
in which TMS was applied over either right parietal cortex,
or the vertex. A different group of 14 subjects (7 women;
mean age) participated in Experiment 2 in which TMS
was applied over either left parietal cortex, or the vertex. Six
subjects of (3 of whom had participated in Experiment 1,
the other 3 participated in Experiment 2) participó
also in a control experiment in which inhibitory tagging
without eye movement was tested. Written informed con-
sent was obtained from each participant. Además, sub-
jects filled in a safety screening questionnaire for TMS
(Keel, Herrero, & Wassermann, 2001). Ethics approval was
van Koningsbruggen et al.
1731
obtained from the School of Psychology at the Bangor
Universidad, Reino Unido. Participants received £10/hour for their
participación.
Apparatus
A limbus tracker (ASL 210, Bedford, MAMÁ) was used to
monitor horizontal eye position at a rate of 1000 Hz.
The eye movement recording device was calibrated by
a three-point calibration every 20 ensayos. A chin and cheek
rest was used to reduce head movements. The analogue
output of the eye tracker was processed on-line to deter-
mine the onset of saccades. When the velocity of saccades
reached 50°/sec, a TTL pulse was sent to stimulus PC which
recorded the saccadic latency and direction. Próximo, the stim-
ulus PC send out two TTL pulses to the TMS stimulator to
trigger the dual TMS pulse. On each trial, two TMS pulses
were given, 150 y 250 msec after the onset of the eye
movimiento. Presentation software (Neurobehavioral Sys-
tems, Albany, California) was used for stimulus presentation
and triggering of the TMS machine. Stimuli were presented
on an IIyama vision master pro 512 monitor (200 Hz). A
response device connected to the game port was used to
record manual reaction times (RTs).
Transcranial Magnetic Stimulation
A MagStim Super Rapid with a 70-mm figure-eight coil was
used for the TMS. The hand area of motor cortex was first
localized in the left hemisphere. The motor threshold then
was determined by finding the minimum amount of TMS
intensity that was required to elicit a visible hand twitch in
the relaxed right hand. Stimulation was set to 120% del
MONTE. All subjects participated in two sessions, separated by
al menos 1 week. In Experiment 1, subjects received TMS
over either the vertex (control site) or a right parietal loca-
tion that was 3 cm to the right and 4 cm posterior relative
to the vertex, with the order of TMS location counter-
balanced across subjects. In Experiment 2, subjects re-
ceived TMS over either the vertex, or a left parietal site
that was 3 cm to the left and 4 cm posterior relative to the
vertex. A similar criterion for parietal cortex stimulation
has been used in previous investigations (Chang & Ro,
2007; Kapoula, Cual, Coubard, Daunys, & Orssaud, 2004,
2005; van Donkelaar & Muri, 2002). All subjects were naive
to the purpose of the study and location of stimulation.
Vertex stimulation and parietal stimulation locations were
relatively close to each other, and only one location was
stimulated in each session, subjects did not report feeling
any differences between sessions.
Procedimiento
The experiment was conducted in a dimly lit room. The dis-
tance between the monitor and the subjects was 57 cm. El
stimulus display consisted of three small white fixation
puntos (0.1° × 0.1°) on a black background, one presented
in the center, and the other two presented 10° to the right
or left of the center. A white unfilled box (3° × 3°) was pre-
sented 5° above and below each fixation point. The six
boxes and the two peripheral fixation points were pre-
sented throughout the experiment. The stimulus display
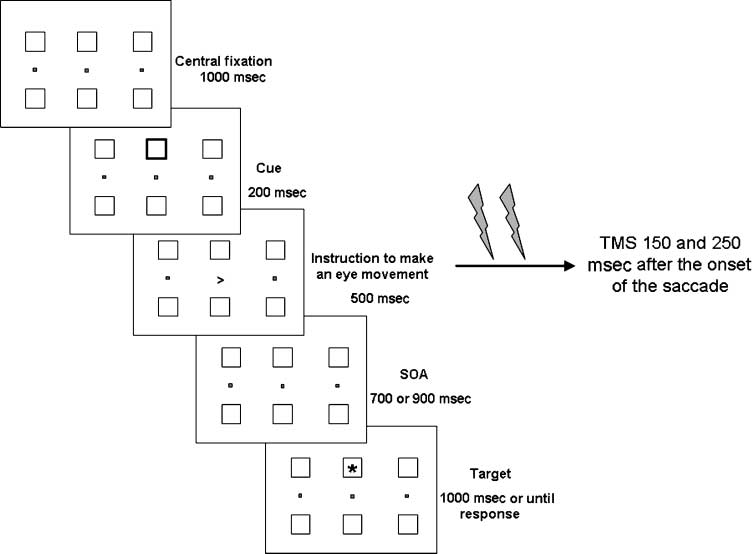
and the trial structure are shown in Figure 1.
Each trial began with the onset of a central fixation
punto. If the subject did not fixate at the central fixation
point within 250 mseg, the trial was aborted and an error
sound was presented. Después 1000 mseg, a noninformative
cue was presented in one of the midline boxes, either
above or below the central fixation. The cue was a thicken-
ing of the line for 200 mseg. A right or left arrow (1°) era
presented at central fixation 300 msec after cue offset. El
arrow was presented for 200 mseg. Subjects were in-
structed to move their eyes as fast as possible in the direc-
tion of the arrow toward either the left or right peripheral
fixation point. If subjects made a saccade in the wrong
direction, or did not make a saccade within 500 mseg,
the trial was aborted and an error sound was presented.
Following the eye movement, subjects were required to
keep fixation at the indicated peripheral fixation point.
Después 700 o 900 mseg, a target was presented either above
or below the central fixation point (es decir., a SOA of either
700 o 900 mseg). The target was presented until the sub-
ject responded by pressing a button with their right index
finger, or for 1000 mseg (ver figura 1 for a graphical illus-
tration of the trial structure). Although both the cue and
targets were always presented in the box above, or below
initial central fixation, there were an additional four boxes.
The additional boxes were presented because pilot testing
revealed that this led to a reliable remapped IOR. Following
a training session of 20 ensayos, a total of 176 trials were pre-
enviado, con 10% catch trials. Catch trials were exactly the
same as the other trials (es decir., including TMS), except that no
target was presented.
All subjects participated in two sessions: one session
with either right parietal TMS (Experimento 1) or left parie-
tal TMS (Experimento 2), and one with TMS over the vertex.
Each session took around 60 mín., and the order of sessions
was counterbalanced across subjects. The intertrial interval
was set to 4000 mseg, in order to make sure that the time
between two successive TMS trains was never shorter than
5000 msec to conform with safety guidelines (Wassermann,
1998; Chen et al., 1997). Sapir et al. (2004) probed for IOR
at both retinal and environmental locations. Sin embargo,
because of the long intertrial interval required, in the cur-
rent study the TMS sessions were 60 min long, and it was
not practical to test for both retinal and environmental
IOR. Por lo tanto, we only tested environmental IOR in this
experimento.
RESULTADOS
TMS Location
Because the stimulation site was based on skull landmarks
(3 cm lateral and 4 cm posterior to the vertex, we acquired
1732
Revista de neurociencia cognitiva
Volumen 22, Número 8
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
8
mi
–
1
pag
7
d
3
F
0
/
1
2
9
2
3
/
9
8
4
/
8
1
9
7
oh
3
C
0
norte
/
1
2
0
7
0
7
9
0
2
2
1
1
4
3
5
/
6
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
3
5
0
6
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
t
/
.
F
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Cifra 1. Trial structure
and stimulation sequence
for a cued target with a saccade
to the right between the
cue and target presentation.
Note that the colors are
inverted for illustration
purposes.
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
8
mi
–
1
pag
7
d
3
F
0
/
1
2
9
2
3
/
9
8
4
/
8
1
9
7
oh
3
C
0
norte
/
1
2
0
7
0
7
9
0
2
2
1
1
4
3
5
/
6
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
3
5
0
6
7
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
.
.
t
/
F
oh
norte
1
8
METRO
a
y
2
0
2
1
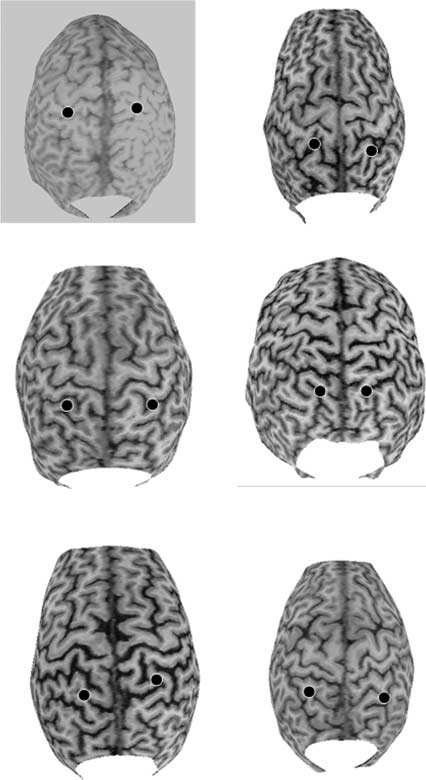
a T1-weighted anatomical MRI scan of six subjects (4 de
Experimento 1) to more precisely specify the anatomical
location target with TMS in a sample of our subjects. Irre-
spective of the experiment in which the subject had partic-
ipated, a marker (vitamin E capsule) was placed over both
the right and left parietal stimulation side prior to the MRI
scan. Brainsight Software was used to process the MRI
scans. Cifra 2 shows that the parietal TMS was over the
rostral superior parietal lobule, including the anterior part
of the IPS. This is in accordance with region of maximum
overlap in the Sapir et al. (2004) estudiar.
Results Experiment 1
Errores
There were two different types of errors subjects could
make: an eye movement error, or a manual keypress error.
A failure to keep fixation at the center, a blink, an eye move-
ment which was not in the right direction or was too slow
(>500 msec), or a failure to keep fixation at the peripheral
fixation point after a successful eye movement were
all classified as eye movement errors. We used such a strict
criterion as TMS was given relative to the saccade onset.
Subject made an average of 6.5% eye movement errors,
which did not differ between TMS conditions (right parietal,
or vertex), saccade direction ( left or right), or the inter-
action between these two (F < 1). All trials with a saccade
error were aborted. Responses faster than 90 msec or
slower than 750 msec were all classified as keypress errors,
and omitted from the analyses. Subjects made a very small
number of manual keypress errors (1.5%), and were there-
fore not further analyzed. Catch trials were omitted from
the analyses as well.
Saccadic Reaction Times
The overall mean saccadic RT was 306 msec (SD = 24 msec).
Thus, on average, the two TMS pulses were given 456 msec
and 556 msec after the onset of the arrow (i.e., 150 and
250 msec after the saccade onset). Mean saccade RTs were
computed for each participant in each condition and were
analyzed with a repeated measures analysis of variance
(ANOVA) with TMS condition (right parietal or vertex)
and saccade direction (left or right) as factors. TMS did
not affect saccade latencies [F(1, 13) < 1]. The interaction
between TMS condition and saccade direction was also not
significant [F(1, 13) < 1]. However, the leftward directed
saccades were significantly faster (302 msec) than rightward
directed saccades (309 msec) [F(1, 13) = 7.48, p < .05,
ηp
2 = .37]. Although the difference was statistically reliable,
it was only 7 msec, that is, the TMS pulses were almost given
at the same time for left and right saccades.
Manual Reaction Times
Anticipatory responses (faster than 90 msec) and slow re-
sponses (slower than 750 msec) were excluded from the
analysis. Note, that if subjects did not execute the saccade
correctly, the trial would have been aborted. Mean RT for
each subject in each condition was computed and analyzed
in a repeated measures ANOVA with TMS condition (right
parietal or vertex), saccade direction (left or right), cue (pre-
viously cued or uncued box), and SOA (700 or 900 msec)
van Koningsbruggen et al.
1733
as within-subject factors. There were only two significant
effects. RTs were slower [F(1, 13) = 7.24, p < .05, ηp
2 =
.36] for cued targets (296 msec) than for uncued targets
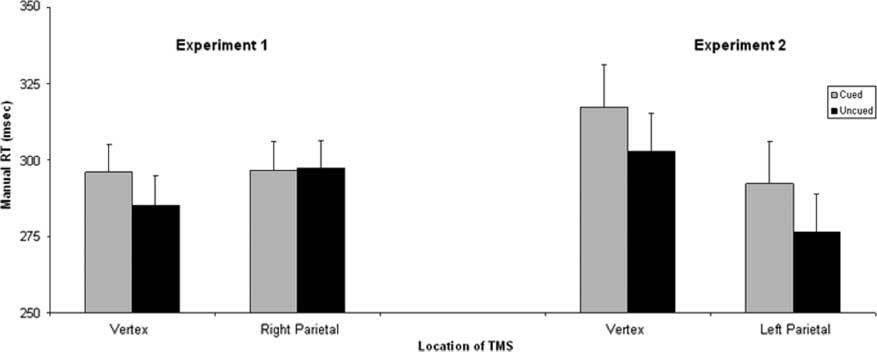
(291 msec), that is, there was a significant IOR. As shown
in Figure 3, the interaction between cue and TMS location
was also significant [F(1, 13) = 24.72, p < .01, ηp
2 = .66], in-
dicating that the site of TMS affected IOR.
Two pairwise comparisons were performed to investi-
gate the interaction between cue (previously cued or un-
cued) and TMS location (vertex or right parietal). There
was a significant effect of cue on RTs during vertex stimula-
tion [t(13) = 5.64, p < .001], that is, there was a significant
IOR of 11 msec. However, when TMS was administered
over right parietal cortex, there was no significant differ-
ence in RT for cued and uncued targets (t < 1).
No other effects were significant, including the inter-
action between TMS × Cue × Saccade direction (F < 1),
indicating that the remapping impairment induced by right
parietal stimulation was independent of the direction of the
eye movement.
Results Experiment 2
Errors
Subject made an average of 6.7% eye movement errors,
which did not differ between TMS condition, saccade direc-
tion, or the interaction between these two (F < 1). Subjects
made a very small number of manual keypress errors
(1.9%), and were therefore not further analyzed.
Saccadic Reaction Times
Figure 2. The 3-D-rendered structural MRI scans of the six scanned
subjects. On each scan, the two possible TMS locations are marked:
4 cm posterior and 3 cm to the right and left relative to the vertex.
The overall mean saccadic RT was 335 msec (SD = 39 msec).
Thus, on average, the two TMS pulses were given 485
and 585 msec after the onset of the arrow, that is, 150
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
8
e
-
1
p
7
d
3
f
0
/
1
2
9
2
3
/
9
8
4
/
8
1
9
7
o
3
c
0
n
/
1
2
0
7
0
7
9
0
2
2
1
1
4
3
5
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
f
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 3. The mean manual RT for cued (gray) and uncued (black) targets for each TMS location. The data for Experiment 1 (right parietal
and vertex TMS) are on the left. The data for Experiment 2 (left parietal and vertex TMS) are on the right. Error bars reflect within-subjects
standard error of the mean (Loftus & Masson, 1994) calculated for each experiment separately.
1734
Journal of Cognitive Neuroscience
Volume 22, Number 8
and 250 msec after the saccade onset. Mean saccade RT
was computed for each subject in each condition and ana-
lyzed with a repeated measures ANOVA with TMS (left pari-
etal or vertex) and saccade direction (left or right) as
within-subject factors. There were no significant effects
(F < 1 in all cases).
Manual Reaction Times
Mean manual RTs for each subject in each condition were
computed and analyzed in a repeated measures ANOVA,
with TMS (left parietal or vertex), saccade direction (left
or right), cue (previously cued or uncued box), and SOA
(700 or 900 msec) as within-subject factors performed
to study the effects of left parietal TMS. RTs were slower
RTs for cued targets (305 msec) than for uncued targets
(290 msec), indicating that there was a significant IOR of
15 msec [F(1, 13) = 19.45, p < .01, ηp
2 = .6]. IOR was larger
at the short SOA (21 msec) than at the long SOA (9 msec)
[F(1, 13) = 7.01, p < .05, ηp
2 = .35].
There was also a significant interaction between Sac-
cade direction × TMS [F(1, 13) = 5.61, p < .05, ηp
2 =
.30]. In order to study this interaction, four pairwise com-
parisons were conducted. RTs were significantly faster
after a saccade to the left (280 msec) than after a saccade
to the right (289 msec) when left parietal TMS was admin-
istered [t(13) = 5.96, p < .01]. There were no other sig-
nificant effects. Most important, the interaction between
TMS × Cue was not significant (F < 1), suggesting that
TMS to the left parietal and TMS to the vertex have simi-
lar effects. Figure 3 shows that there is a significant IOR
after left parietal TMS, which is comparable to that ob-
served after control stimulation at the vertex; that is, left
parietal TMS does not influence the remapping of the
inhibitory tag.
Comparing the Results of Experiments 1 and 2
Two different ANOVAs were performed to compare the
effects of right and left PPC TMS, with experiment as a
between-subjects factor. One ANOVA compared the effect
of vertex TMS for both experiments, whereas the other
ANOVA compared the effect of parietal TMS for both ex-
periments. The reason for comparing the vertex stimula-
tion between experiments was to determine whether
there is a baseline difference between the groups. The
mixed effect repeated measures ANOVA with experiment
(1 or 2) as a between-subjects factor, and saccade direction
(left or right), cue (previously cued or uncued box), and
SOA (700 or 900 msec) as within-subject factors revealed
only a significant effect of cue [F(1, 26) = 20.91, p < .01,
ηp
2 = .45], reflecting longer latencies for targets at cued
(307 msec) than at uncued (294 msec) locations. There
were no other significant effects, including no main effect
of experiment, or interactions with the experiment factor.
Thus, the groups in Experiments 1 and 2 did not differ
from each other in the vertex condition.
The same ANOVA was performed for parietal TMS.
There was a significant effect of cue [F(1, 26) = 8.10, p <
.01, ηp
2 = .24] and a significant Cue × Experiment inter-
action [F(1, 26) = 9.70, p < .01, ηp
2 = .27]. The source of
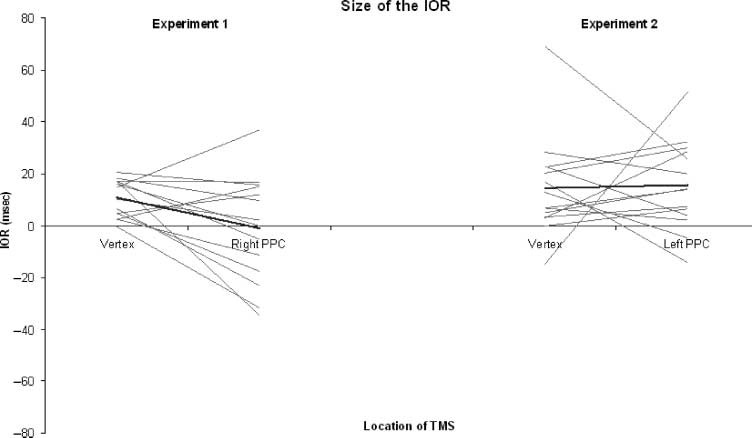
this interaction was examined by comparing the size of
IOR for both experiments. As expected, the size of the
IOR was significantly larger in Experiment 2 ( left parietal
TMS; 16 msec) than in Experiment 1 (right TMS; −1 msec)
[t(26) = 3.11, p < .01]. The effect of saccade direction
was also significant [F(1, 26) = 5.05, p < .05, ηp
2 = .16],
as was the interaction between saccade direction and ex-
periment [F(1, 26) = 6.52, p < .05, ηp
2 = .20]. Follow-up
t test found that there was no effect of saccade direction
in Experiment 1 (right parietal TMS; RT was 297 msec inde-
pendent of saccade direction). However, in Experiment 2
Figure 4. The mean size
of IOR (thick black line)
and individual subject data
(thin gray lines). The data
for Experiment 1 are on the
left, and for Experiment 2
on the right.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
8
e
-
1
p
7
d
3
f
0
/
1
2
9
2
3
/
9
8
4
/
8
1
9
7
o
3
c
0
n
/
1
2
0
7
0
7
9
0
2
2
1
1
4
3
5
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
/
.
.
f
o
n
1
8
M
a
y
2
0
2
1
van Koningsbruggen et al.
1735
(left parietal TMS), RTs were faster after a saccade to the left
(280 msec) than after a saccade to the right (289 msec)
[t(13) = 5.96, p < .01]. No other effects were significant. The
individual and mean sizes of IOR are displayed in Figure 4.
Control Experiment
The current result suggests that right parietal TMS impairs
remapping of the inhibitory tag regardless of the direction
of eye movements. However, the conditions in this study
do not allow us to rule out the possibility that right parietal
TMS abolishes IOR in general and not only the remapping
of the inhibitory tag. Sapir et al. (2004) also presented tar-
gets at retinal cued locations, for which no remapping was
required. Like the healthy controls, the patientsʼ RTs were
slower for the cued retinal location. This finding demon-
strated that patients had a normal IOR, but that this IOR
was lost when they were required to remap the inhibitory
tag. Because there were no such targets presented at the
retinal location of the cue in the current experiment (i.e.,
subjects were always required to remap the inhibitory
tag), we performed a control experiment in a few partici-
pants, in which no remapping was required.
The procedure was identical, except that subjects were
not required to make an eye movement. Instead of an
arrow, an equal sign of the same size was presented, and
subjects were instructed to maintain central fixation. The
TMS pulses were given relative to their mean saccadic RTs
of the previous experiment. We recruited six subjects who
had participated in either Experiment (3 had participated
in Experiment 1 and the other 3 in Experiment 2). They par-
ticipated in one vertex and one right parietal TMS session.
RTs were subject to a 2 (TMS) × 2 (cue) × 2 (SOA) re-
peated measures ANOVA. The main effect of cue was signifi-
cant [F(1, 5) = 16.43, p = .01, ηp
2 = .77], reflecting the slow
RTs to valid than for invalid trials (i.e., IOR). There was no
interaction between TMS site and cue, and a paired-samples
t test confirmed that there was no significant difference in
the size of IOR during vertex stimulation (11 msec) and
parietal stimulation (10 msec) [t(5) = 0.16, p = .88]. This
result confirms that right parietal TMS does not influence
IOR when there is no need to update the saliency map.
DISCUSSION
The results of these experiments converge with those re-
ported in neurological patients (Sapir et al., 2004), implicat-
ing the rostral superior parietal lobule in updating saliency
maps after eye movements. They also demonstrate a hemi-
spheric asymmetry in representing salience maps. TMS
over the right, but not the left, rostral superior parietal
lobule prevented remapping of the inhibitory tag after
either left or right saccades. This remapping deficit was
regardless of whether the target appeared in the field ipsi-
lateral or contralateral to the right parietal stimulation site.
Our results also converge with another recent TMS study
reporting that stimulation over the right, but not left, hemi-
sphere, at a more dorsal site over the IPS than the one used
here (i.e., at the P3 electrode site), disrupted transsaccadic
working memory (Prime et al., 2008).
As was the case in the case of the patients studied by
Sapir et al. (2004), the absence of environmental IOR was
a result of a slower detection RT at invalidly cued locations.
There was no difference between right parietal cued, right
parietal uncued, and vertex cued targets. One possible
explanation for this effect is that the cued location has a
reduced saliency, and that TMS impaired the updating of
the whole saliency map, resulting in lower saliency and
longer RTs for all possible target locations. Another possi-
bility is that IOR occurs because the other target location
becomes more salient, and that this benefit has been
disrupted.
Chang and Ro (2007) showed that TMS over right parie-
tal cortex impaired perisaccadic displacement detection.
Performance was only affected when the probe was pre-
sented in the left visual field, subjects were required to
make a contralateral saccade, and TMS was given just
before the saccade. They hypothesized that the reason that
TMS affects perception is that TMS introduces external
noise into the PPC representation. In the current study,
the absence of a remapped inhibitory tag might be caused
by that TMS-introduced noise into the parietal representa-
tion as Chang and Ro (2007) hypothesized. When subjects
were not required to make eye movements (control experi-
ment), or TMS was applied over left PPC, the inhibitory tag
was remapped, making it less likely that TMS introduces
just noise. To further investigate this issue, it would be in-
teresting to study whether right PPC TMS affects retinal
IOR. However, patients with a parietal lobe lesion show
preserved retinal IOR. This further supports the idea that
parietal lobe disruption interferes with remapping rather
than just degrading location information.
The TMS pulses were given 150 and 250 msec after the
onset of the eye movement. This time interval was chosen
based on previous research indicating that this is the critical
time of spatial updating in an ERP study (Bellebaum,
Hoffmann, & Daum, 2005), in single-unit recordings (Gottlieb
et al., 1998; Duhamel et al., 1992), and in previous studies
using the double-step saccade paradigm (Morris et al., 2007;
van Donkelaar & Muri, 2002). Although, no other time
points were tested, it is interesting to note that there was
no effect of SOA (700 or 900 msec) in right parietal TMS
group. This indicates that once the representation of the
inhibitory tag is affected by TMS, it cannot be regained.
Further research is needed to definitively specify the critical
time that TMS must be applied to disrupt remapping.
Like the patient study of Sapir et al. (2004), but unlike
previous patient (Heide & Kompf, 1998; Heide et al., 1995;
Duhamel et al., 1992) and TMS (Morris et al., 2007; van
Donkelaar & Muri, 2002) studies employing the double-
step saccade paradigm, disruption of remapping occurred
when saccades were directed toward the ipsilateral as well
as contralateral fields. In double-step saccade studies, the
deficit has been observed only when saccades were directed
1736
Journal of Cognitive Neuroscience
Volume 22, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
8
e
-
1
p
7
d
3
f
0
/
1
2
9
2
3
/
9
8
4
/
8
1
9
7
o
3
c
0
n
/
1
2
0
7
0
7
9
0
2
2
1
1
4
3
5
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
/
.
f
.
.
o
n
1
8
M
a
y
2
0
2
1
contralateral to the disrupted cortex. The current results
suggest that parietal cortex is not simply responsible for
generating an extra-retinal signal necessary for updating a
salience map of the visual field, but that right parietal cortex
is critical for maintaining a durable, transsaccadic representa-
tion of that map across the visual field.
This interpretation of the current results is consistent with
observations from neurophysiological recording in LIP of
nonhuman primates, demonstrating that this area is con-
cerned specifically with remapping of objects that are salient
by virtue of either a recent appearance or because they are
designated as targets of visual search (Gottlieb et al., 1998).
Moreover, updating of the receptive fields of LIP neurons
is independent of whether the saccade is directed ipsi-
lateral or contralateral to the neuronʼs receptive field, and
whether the object is updated within or between hemi-
spheres (Heiser & Colby, 2006).
It is possible that separate regions of the IPS may be
responsible for generating the extra-retinal signal and
for maintaining a remapped representation. A recent TMS
study (Morris et al., 2007), using an adaptation of the
double-step saccade paradigm, reported inaccurate second
saccades after stimulation of a posterior area of right pa-
rietal cortex, but not after TMS of a more rostral site that
approximated the region stimulated in the current investi-
gation. We might speculate that the more posterior part of
parietal cortex is necessary for generating an extra-retinal
signal, such that its inactivation only affects performance
after contralateral saccades, whereas more anterior parts
of right parietal cortex maintain durable representation of
the remapped salience map after a saccade in either direc-
tion. However, it is notable that the effect specific to PPC
in the Morris et al. (2007) experiment was an increase in
variability in the second saccade end point, suggestive
perhaps of a degraded representation of the location of
the second target. There was no evidence that TMS of this
site resulted in saccades to the retinal location of the
second target, as might be expected if TMS prevented the
generation of a critical extra-retinal signal. Further TMS
experiments, over the more posterior site examined by
Morris et al., using the kind of paradigm used here, or the
transsaccadic memory paradigm employed by Prime et al.
(2008), may seek further evidence for a dissociation of
function along parietal cortex that may contribute to up-
dating and maintaining salience maps across saccades.
A TMS study targeting FEF that examines the remapping
of IOR may also be informative. Lesions of the FEF have
been reported to not disrupt performance on double-step
saccades (Heide et al., 1995). However, parietal cortex
receives remapping signals from the colliculo-thalamic-
FEF circuit elucidated by Sommer and Wurtz (2006), and
these signals may be critical for the encoding of a durable
salience map.
All previous TMS studies using the double-step saccade
paradigm have, to our knowledge, only stimulated the right
parietal lobe (Morris et al., 2007; van Donkelaar & Muri,
2002), and further research is needed to clarify whether
there may be hemispheric asymmetries in saccade re-
mapping in this paradigm.
The patient research using the double-step saccade
paradigm, reported by Heide et al.’s (1995) lab, does
suggest that the left parietal lobe participates in saccade
remapping. In a double-step saccade task, in which the
first and second targets occurred in opposite visual fields,
patients with left parietal lesions did show a deficit,
although patients with right parietal lesions were more
impaired. fMRI studies have not revealed hemispheric
updating asymmetries (Medendorp et al., 2003; Merriam
et al., 2003): Representations of stimuli presented to the
right hemisphere are remapped to the left hemisphere
after left saccades, and representations of stimuli pre-
sented to the left hemisphere are remapped to the right
hemisphere after right saccades.
Although there is, then, evidence for a role of both hemi-
spheres in saccade remapping, there is also evidence that
their contributions may differ. Heide et al. (1995) also
tested parietal lesioned patients on a within-hemifield
double-step saccade task, in which both targets were pre-
sented in the same visual field. In this task, in which no
between-hemispheric spatial updating was necessary, an
asymmetric effect of right and left parietal lesions was
observed. In addition to impaired performance on the
between-hemifield task, patients with right parietal lesions
also had an impairment in the within-hemifield condi-
tion in the left visual field. An ERP study by Bellebaum
et al. (2005) also provided evidence for different contri-
butions of left and right hemisphere in saccade remapping.
Bellebaum et al. reported a larger slow positive wave
when remapping was required, starting between 150 and
200 msec after first saccade onset. Source analysis showed
that whereas the source was restricted to right PPC in trials
with leftward first saccades, left and right PPC were both
involved in rightward trials.
Further research is needed to specify the circumstances
under which left parietal cortex may be involved in up-
dating salience maps. Future studies, using the paradigm
employed here, will test for hemispheric asymmetries in
salience updating when the stimuli to be updated are pre-
sented to either the left or right visual field, when vertical
saccades are made, and when stimuli must be remapped
either within or between hemispheres.
In conclusion, these observations converge with those
made in neurological patients with chronic lesions of pari-
etal cortex implicating this region as a neural substrate for
maintaining the spatial constancy necessary for a coherent
continuity of visual experience. They also suggest a special
role for the right parietal lobe, at least under the condi-
tions of the current experiments. The observation that re-
mapping was disrupted when saccades were executed
toward the field ipsilateral as well as contralateral to cortical
disruption suggests that parietal cortex is not involved
simply in generating the corollary discharge that provides
the extra-retinal signal needed for remapping the visual
scene. Rather, the results implicate parietal cortex as a
van Koningsbruggen et al.
1737
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
8
e
-
1
p
7
d
3
f
0
/
1
2
9
2
3
/
9
8
4
/
8
1
9
7
o
3
c
0
n
/
1
2
0
7
0
7
9
0
2
2
1
1
4
3
5
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
/
.
.
t
.
o
n
1
8
M
a
y
2
0
2
1
neural substrate that uses the extra-retinal signal to main-
tain a continuous salience map across saccades.
Acknowledgments
This work was supported by the Wellcome Trust (grant 079886).
Reprint requests should be sent to Martijn Gerbrand van
Koningsbruggen, School of Psychology, Brigantia Building, Penrallt
Road, Bangor, Gwynedd LL572AS, UK, or via e-mail: Martijn_van_
koningsbruggen@hotmail.com.
REFERENCES
Bellebaum, C., Hoffmann, K. P., & Daum, I. (2005). Post-
saccadic updating of visual space in the posterior parietal
cortex in humans. Behavioural Brain Research, 163, 194–203.
Chang, E., & Ro, T. (2007). Maintenance of visual stability in
the human posterior parietal cortex. Journal of Cognitive
Neuroscience, 19, 266–274.
Chen, R., Gerloff, C., Classen, J., Wassermann, E. M., Hallett, M.,
& Cohen, L. G. (1997). Safety of different inter-train
intervals for repetitive transcranial magnetic stimulation
and recommendations for safe ranges of stimulation
parameters. Electroencephalography and Clinical
Neurophysiology, 105, 415–421.
Danziger, S., Fendrich, R., & Rafal, R. D. (1997). Inhibitory
tagging of locations in the blind field of hemianopic
patients. Consciousness and Cognition, 6, 291–307.
Duhamel, J. R., Colby, C. L., & Goldberg, M. E. (1992). The
updating of the representation of visual space in parietal
cortex by intended eye movements. Science, 255, 90–92.
Duhamel, J. R., Goldberg, M. E., Fitzgibbon, E. J., Sirigu, A.,
& Grafman, J. (1992). Saccadic dysmetria in a patient with
a right frontoparietal lesion. The importance of corollary
discharge for accurate spatial behaviour. Brain, 115, 1387–1402.
Gottlieb, J. P., Kusunoki, M., & Goldberg, M. E. (1998). The
representation of visual salience in monkey parietal
cortex. Nature, 391, 481–484.
Heide, W., Blankenburg, M., Zimmermann, E., & Kompf, D.
(1995). Cortical control of double-step saccades: Implications
for spatial orientation. Annals of Neurology, 38, 739–748.
Heide, W., & Kompf, D. (1998). Combined deficits of
saccades and visuo-spatial orientation after cortical lesions.
Experimental Brain Research, 123, 164–171.
Heiser, L. M., & Colby, C. L. (2006). Spatial updating in area
LIP is independent of saccade direction. Journal of
Neurophysiology, 95, 2751–2767.
Kapoula, Z., Yang, Q., Coubard, O., Daunys, G., & Orssaud, C.
(2004). Transcranial magnetic stimulation of the posterior
parietal cortex delays the latency of both isolated and
combined vergence–saccade movements in humans.
Neuroscience Letters, 360, 95–99.
Kapoula, Z., Yang, Q., Coubard, O., Daunys, G., & Orssaud, C.
(2005). Role of the posterior parietal cortex in the
initiation of saccades and vergence: Right/left functional
asymmetry. Annals of the New York Academy of
Sciences, 1039, 184–197.
Keel, J. C., Smith, M. J., & Wassermann, E. M. (2001). A
safety screening questionnaire for transcranial magnetic
stimulation. Clinical Neurophysiology, 112, 720.
Klein, R. (1988). Inhibitory tagging system facilitates visual
search. Nature, 334, 430–431.
Klein, R. (2000). Inhibition of return. Trends in Cognitive
Sciences, 4, 138–147.
Loftus, G. R., & Masson, M. E. J. (1994). Using confidence
intervals in within-subject designs. Psychonomic Bulletin
& Review, 1, 476–490.
Maylor, E. A., & Hockey, R. (1985). Inhibitory component
of externally controlled covert orienting in visual space.
Journal of Experimental Psychology: Human Perception
and Performance, 11, 777–787.
Medendorp, W. P., Goltz, H. C., & Vilis, T. (2005). Remapping
the remembered target location for anti-saccades in human
posterior parietal cortex. Journal of Neurophysiology,
94, 734–740.
Medendorp, W. P., Goltz, H. C., & Vilis, T. (2006). Directional
selectivity of BOLD activity in human posterior parietal
cortex for memory-guided double-step saccades. Journal
of Neurophysiology, 95, 1645–1655.
Medendorp, W. P., Goltz, H. C., Vilis, T., & Crawford, J. D. (2003).
Gaze-centered updating of visual space in human parietal
cortex. Journal of Neuroscience, 23, 6209–6214.
Merriam, E. P., Genovese, C. R., & Colby, C. L. (2003). Spatial
updating in human parietal cortex. Neuron, 39, 361–373.
Morris, A. P., Chambers, C. D., & Mattingley, J. B. (2007).
Parietal stimulation destabilizes spatial updating across
saccadic eye movements. Proceedings of the National
Academy of Sciences, U.S.A., 104, 9069–9074.
Nakamura, K., & Colby, C. L. (2002). Updating of the visual
representation in monkey striate and extrastriate cortex
during saccades. Proceedings of the National Academy
of Sciences, U.S.A., 99, 4026–4031.
Posner, M. I., & Cohen, Y. (1984). Components of visual
orienting. London: Erlbaum.
Posner, M. I., Rafal, R. D., Choate, L., & Vaughn, J. (1985).
Inhibition of return: Neural basis and function. Cognitive
Neuropsychology, 2, 211–228.
Prime, S. L., Vesia, M., & Crawford, J. D. (2008). Transcranial
magnetic stimulation over posterior parietal cortex disrupts
transsaccadic memory of multiple objects. Journal of
Neuroscience, 28, 6938–6949.
Sapir, A., Hayes, A., Henik, A., Danziger, S., & Rafal, R. (2004).
Parietal lobe lesions disrupt saccadic remapping of
inhibitory location tagging. Journal of Cognitive
Neuroscience, 16, 503–509.
Sommer, M. A., & Wurtz, R. H. (2006). Influence of the
thalamus on spatial visual processing in frontal cortex.
Nature, 444, 374–377.
Sommer, M. A., & Wurtz, R. H. (2008). Brain circuits for
the internal monitoring of movements. Annual Review
of Neuroscience, 31, 317–338.
Tipper, S. P., Grison, S., & Kessler, K. (2003). Long-term inhibition
of return of attention. Psychological Science, 14, 19–25.
Umeno, M. M., & Goldberg, M. E. (1997). Spatial processing
in the monkey frontal eye field: I. Predictive visual responses.
Journal of Neurophysiology, 78, 1373–1383.
Umeno, M. M., & Goldberg, M. E. (2001). Spatial processing
in the monkey frontal eye field: II. Memory responses.
Journal of Neurophysiology, 86, 2344–2352.
van Donkelaar, P., & Muri, R. (2002). Craniotopic updating
of visual space across saccades in the human posterior
parietal cortex. Proceedings of the Royal Society of
London: Series B, Biological Sciences, 269, 735–739.
Walker, M. F., Fitzgibbon, E. J., & Goldberg, M. E. (1995).
Neurons in the monkey superior colliculus predict the
visual result of impending saccadic eye movements.
Journal of Neurophysiology, 73, 1988–2003.
Wassermann, E. M. (1998). Risk and safety of repetitive
transcranial magnetic stimulation: Report and suggested
guidelines from the International Workshop on the
Safety of Repetitive Transcranial Magnetic Stimulation,
June 5–7, 1996. Electroencephalography and Clinical
Neurophysiology, 108, 1–16.
1738
Journal of Cognitive Neuroscience
Volume 22, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
8
e
-
1
p
7
d
3
f
0
/
1
2
9
2
3
/
9
8
4
/
8
1
9
7
o
3
c
0
n
/
1
2
0
7
0
7
9
0
2
2
1
1
4
3
5
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
6
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
.
f
t
/
o
n
1
8
M
a
y
2
0
2
1