Shared Neural Circuits for Mentalizing about the
Self and Others
Michael V. Lombardo1, Bhismadev Chakrabarti1,2,
Edward T. Bullmore1, Sally J. Wheelwright1, Susan A. Sadek1,
John Suckling1, MRC AIMS Consortium*, and Simon Baron-Cohen1
D
oh
w
norte
yo
oh
a
d
mi
d
Abstracto
■ Although many examples exist for shared neural represen-
tations of self and other, it is unknown how such shared repre-
sentations interact with the rest of the brain. Además, hacer
high-level inference-based shared mentalizing representations
interact with lower level embodied/simulation-based shared
representaciones? We used functional neuroimaging (resonancia magnética funcional) y
a functional connectivity approach to assess these questions
during high-level inference-based mentalizing. Shared mental-
izing representations in ventromedial prefrontal cortex, poste-
rior cingulate/precuneus, and temporo-parietal junction (TPJ)
all exhibited identical functional connectivity patterns during
mentalizing of both self and other. Connectivity patterns were
distributed across low-level embodied neural systems such as the
frontal operculum/ventral premotor cortex, the anterior insula,
the primary sensorimotor cortex, and the presupplementary
motor area. These results demonstrate that identical neural
circuits are implementing processes involved in mentalizing
of both self and other and that the nature of such processes
may be the integration of low-level embodied processes within
higher level inference-based mentalizing. ■
INTRODUCCIÓN
“Know thyself ”… And know others? This ancient Greek
aphorism lies at the heart of centuries of scholarly inquiry
in many fields such as psychology, philosophy, and most
recently, social neuroscience (Decety & Grezes, 2006).
One theoretical framework providing the basis for such
inquiry is the simulationist accounts of social cognition
(hombre de oro, 2006). Simulationist accounts posit that we
gain insight about anotherʼs mental experience through
the use of privileged access to our own mental states,
sensaciones, emotions, and other embodied representa-
ciones. One of the main premises forming the basis of
simulationist accounts is the simple prediction that the
1University of Cambridge, Reino Unido, 2Universidad de
Reading, Reino Unido
*The MRC AIMS Consortium is a UK collaboration of autism
research centers including the Institute of Psychiatry at Kings
College, Londres, the Autism Research Centre at the University
de Cambridge, and the Autism Research Group at the Univer-
sity of Oxford. It is funded by the Medical Research Council
(MRC) UK and headed by the Section of Brain Maturation, Insti-
tute of Psychiatry. The Consortium members are, in alphabet-
ical order, Bailey, A. J., Baron-Cohen, S., Bolton, PAG. F., bullmore,
mi. T., Carrington, S., Chakrabarti, B., Daly, mi. METRO., Deoni, S. C.,
Ecker, C., Happé, F., Henty, J., Jezzard, PAG., Johnston, PAG., jones,
D. K., Lombardo, METRO. v., Madden, A., Mullins, D., Murphy, C.,
Murphy, D. GRAMO., Pasco, GRAMO., Sadek, S. A., España, D., Stewart, r.,
Suckling, j. S., Wheelwright, S. J., and Williams, S. C.
brain engages the same neural systems for self and other
referential cognitive processes. Findings supporting this
premise, sin embargo, are divided across two important neural
sistemas. The first system, coding for low-level embodied/
simulative representations, exists within frontal operculum/
ventral premotor cortex (FO/PMv) (Gazzola & Keyser,
2009; Gazzola, Aziz-Zadeh, & Keyser, 2006; Iacoboni &
Dapretto, 2006; Rizzolatti & Craighero, 2004), somatosen-
sory cortices (SI/SII) (Blakemore, Bristow, Bird, Frith, &
Ward, 2005; Keysers et al., 2004), anterior insula (AI), y
caudal ACC (cACC) extending into the presupplementary
motor area (pre-SMA; Critchley, Wiens, Rotshtein, Ohman,
& Dolan, 2004; Singer et al., 2004; Carr, Iacoboni, Dubeau,
Mazziotta, & Lenzi, 2003; Wicker et al., 2003). El segundo
neural system, dealing with more high-level inference-
based mentalizing about both self and other, comprises
an independent set of neural regions within the me-
dial prefrontal cortex (MPFC), posterior cingulate cortex/
precuneus (PCC), and TPJ (Amodio & Frith, 2006; mitchell,
Macrae, & Banaji, 2006; sajonia, Moran, Scholz, & Gabrieli,
2006; Ochsner et al., 2005). Given the importance of both
shared neural systems in social cognition, one puzzling
question is why are these two neural systems consistently
observed independently of one another? And second, si
they are not independent, how do these two crucial shared
neural systems interact during social cognitive processes
such as mentalizing (Keyser & Gazzola, 2007; Uddin,
Iacoboni, Lange, & Keenan, 2007)? These central questions
pose key challenges for explaining how we navigate and
© 2009 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 22:7, páginas. 1623–1635
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
t
F
.
/
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
interact with the social world around us. Comprensión
cómo, en todo caso, such shared neural systems interact will help
clarify the nature of the underlying processes implemented
during mentalizing and canalize further theoretical refine-
ment and empirical work on the topic.
One method for gaining insight into these questions is
through functional or effective connectivity approaches
(Friston et al., 1997). Functional connectivity can provide
a more precise test of whether shared neural representa-
tions for mentalizing about the self and other are in fact
“shared” in how they interact with the rest of the brain. Pre-
vious examples in the domain of shared pain and disgust
processing suggest that rather than converging across
similar neural circuits, these shared representations actu-
ally diverge into distinct functionally connected neural cir-
cuits ( Jabbi, Bastiaansen, & Keyser, 2008; Zaki, Ochsner,
Hanelin, Apostar, & Mackey, 2007). These examples are
prime illustrations of how functional connectivity can
provide additional insight into the nature of the pro-
cesses underlying shared representations. Extending this
idea into the domain of higher level inference-based
mentalizing, if functional connectivity patterns are similar
for mentalizing about both self and other, this would sup-
port the idea that shared mentalizing representations im-
plement similar underlying processes for both self and
otro. Sin embargo, the alternative hypothesis would predict
that the implementation of processes underlying shared
mentalizing representations (indexed by functional con-
conectividad) actually diverge into distinct neural circuits for
self and other. This alternative would support the claim
that the processes underlying self-mentalizing are distinct
from those underlying other-mentalizing.
Segundo, by identifying the neural space through which
shared mentalizing processes are implemented, func-
tional connectivity approaches can also constrain ideas
about the psychological/cognitive significance of such
underlying processes. Historically speaking, scholars in
the field have contrasted high-level inference-based men-
talizing processes as an opposite of low-level simulation-
based processes (hombre de oro, 2006; gopnik & Wellman,
1992; gordon, 1992). Sin embargo, a middle ground may
exist within the idea that some aspects of both high-level
and low-level social cognitive processes are grounded
within the framework of embodied cognition (hombre de oro
& de Vignemont, 2009; Barsalou, 1999, 2008). Based on
theories of embodied cognition (Barsalou, 1999, 2008), él
would be predicted that high-level social-cognitive con-
ceptual representations such as those occurring through
explicit mentalizing are built upon through their interac-
tions with low-level embodied/simulative representations.
A recent model by Keysers and Gazzola (2007) proposes
that this may be implemented in the brain through the
integration of information from neural systems for low-
level embodied/simulation-based processes (p.ej., action–
perception mirroring, somatosensory or other embodied
representaciones) and high-level inference-based mental-
izing. The integration of these two neural systems makes
the prediction that during high-level inference-based
mentalizing, neural systems such as MPFC, PCC, and TPJ
should be functionally connected to lower level embodied/
simulation-based neural systems like FO/PMv, SI/SII, AI,
and cACC/pre-SMA. Such a finding would suggest that
during high-level inference-based mentalizing, we use our
own lower level embodied/simulated shared representa-
tions as the building blocks for making inferences about
our own mind as well as otherʼs minds.
We used fMRI and a functional connectivity approach
to examine these interactions within 33 healthy male
Participantes. Participants were scanned in a 2 × 2 facto-
rially designed fMRI experiment where participants either
made a reflective judgment about the self or a familiar non-
close other (the British Queen) in a mentalistic way (self-
mentalizing [SM], other-mentalizing [OM]) or physical
way (self-physical [SP], other-physical [OP]; see Methods).
MÉTODOS
Participantes
Thirty-three healthy male participants were included in
este estudio (edad media = 27.97 años, DE = 6.10 años,
range = 18–42 years). Informed consent was obtained
for all participants in accord with procedures approved
by the Suffolk Local Research Ethics Committee. All par-
ticipants were right-handed native English speakers with
normal or corrected-to-normal vision. Participants reported
no history of psychiatric or neurological conditions and
were not currently taking any medication.
Task Design
The study design was a 2 × 2 within-subjects factorial
block design where participants were asked to make
either reflective “mentalizing” or “physical” judgments
about two target individuals: the “self” or a familiar non-
close “other” (the British Queen). For SM blocks, participar-
pants judged on a scale from 1 (not at all likely) a 4
(very likely) how likely they themselves would personally
agree with opinion questions that focused on mental
características (p.ej., “How likely are You to think that
keeping a diary is important”). On OM blocks, lo mismo
mentalizing judgments were made, except this time it
was in reference to how likely the British Queen would
agree with the opinion questions (p.ej., “How likely is the
Queen to think that keeping a diary is important”). During
SP blocks, participants judged how likely they would
personally agree to questions about their own physical
características (p.ej., “How likely are You to sneeze when
a cat is nearby”). En cambio, the same physical judg-
ments were made during OP blocks, except that partici-
pants rated these questions with the Queen as the
target person (p.ej., “How likely is the Queen to sneeze
when a cat is nearby”).
1624
Revista de neurociencia cognitiva
Volumen 22, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
F
t
.
/
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
All participants completed one scanning session with
one functional imaging run. Within this run, Había
20 trials within each condition and five blocks per con-
condición. Each trial type was presented in blocks of four
ensayos, and the duration of each trial was 4 segundo (16 segundo
per block). After each block, there was a rest period of
16 sec where participants fixated on a cross in the middle
of the screen. All trials within blocks and all blocks
throughout the functional run were presented in pseu-
dorandom order. All opinion questions were acquired
from Jason Mitchellʼs laboratory and have been used in
previous studies on reflective judgments about the self
y otros (Jenkins, Macrae, & mitchell, 2008; mitchell
et al., 2006). Stimulus presentation was implemented
with the DMDX software, and the stimulus presentation
computer was synchronized with the onset of the func-
tional run to ensure accuracy of event timing.
fMRI Acquisition
Imaging was performed using a 3-T GE Signa Scanner
(Sistemas médicos eléctricos generales, Milwaukee, Wisconsin) en
the Cambridge Magnetic Resonance Imaging and Spec-
troscopy Unit (MRIS Unit). Our functional imaging run
consistió en 325 whole-brain functional T2*-weighted
EPIs (slice thickness = 3 mm; 0.8 mm skip; 33 axial slices;
repetition time = 2000 mseg; echo time = 30 mseg; flip
angle = 90°; matrix = 64 × 64; campo de visión = 240 mm;
sequential slice acquisition). The first five time points
of the run were discarded to allow for T2 stabilization
efectos. Además, a high-resolution three-dimensional
spoiled gradient (SPGR) anatomical image was acquired
for each participant for registration purposes.
Análisis de los datos
Behavioral RT data were analyzed with a repeated mea-
sures ANOVA in SPSS 16 (http://www.spss.com). resonancia magnética funcional
data preprocessing and statistics were implemented
using SPM5 ( Wellcome Trust Centre for Neuroimaging,
http://www.fil.ion.ucl.ac.uk/spm). The preprocessing
steps were conducted in the following manner: Func-
tional data were slice timing corrected and realigned to
the mean functional image. Próximo, the realigned and
slice-timing-corrected functional data were coregistered
to the high-resolution SPGR. The high-resolution SPGR
was then segmented into cerebrospinal fluid, gray, y
white matter. The normalization transformation matrix
from the segmentation step was then applied to the func-
tional and structural images, thus transforming it into
standard anatomical space based on the ICBM 152 cerebro
template (Instituto Neurológico de Montreal [MNI]) at a reso-
lution of 2 mm of isotropic voxels. Normalized functional
data were then smoothed with a FWHM of 8 mm.
Whole-brain statistical analysis was performed using the
general linear model in SPM5. Each trial was convolved
with the canonical hemodynamic response function. High-
pass temporal filtering with a cutoff of 128 sec was applied
to remove low-frequency drift in the time series, and global
changes were removed by proportional linear scaling.
Serial autocorrelations were estimated with a restricted
maximum likelihood algorithm with an autoregressive
model of order 1. Factorial contrast images were outputted
automatically in the first-level single subject analysis.
At the group level of analysis, three conjunction ana-
lyses were performed. All conjunction analyses use a log-
ical “AND” masking procedure whereby a map of false
discovery rate (FDR)-corrected suprathreshold voxels in
contrast A is overlaid on a map of FDR-corrected supra-
threshold voxels in orthogonal contrast B and only the
voxels that overlap across contrast A “AND” B are ex-
tracted. The first conjunction analysis identified overlap
among mentalizing representations for self and other
(SM > SP “AND” OM > OP). The second and the third
conjunction analyses identified general self-biases (SM >
OM “AND” SP > OP) or other biases (OM > SM “AND”
OP > SP). To test for whole-brain interaction effects
among the four conditions, we conducted a paired sam-
ples t test comparing the contrasts of SM > SP to OM >
OP (es decir., [SM > SP] > [OM > OP]). This analysis was
thresholded at p < .05, FDR corrected.
Functional connectivity analyses were implemented
with psychophysiological interaction (PPI) analyses with-
in SPM5 (Friston et al., 1997). Three seed regions were
defined for the PPI analyses. These seeds were the ventro-
medial prefrontal cortex (vMPFC), the PCC, and the right
TPJ (RTPJ) and were each functionally defined as the entire
cluster of suprathreshold voxels in each respective region
identified from the group-level conjunction analysis as
shared mentalizing representations under both SM > SP
“AND” OM > OP contrasts (see the first three clusters in
Mesa 1). Time courses from each seed region were ex-
tracted and multiplied by a condition vector of 0, 1, o
−1, where mentalizing trials (p.ej., in SM > SP, this is the
SM condition) were coded as 1, physical trials were coded
as −1 (p.ej., in SM > SP, this is the SP condition), and all
other events were coded as 0. This product vector of [tiempo
courses × condition vector] was our PPI vector. The time
curso, the condition vector, and the PPI vector were
entered as regressors into single subject analyses, y estafa-
trast maps were computed for the PPI regressor. Single
subject PPI contrast maps were then entered into a second-
level group analysis.
A nivel de grupo, one-sample t tests were computed
on the PPI contrast images and thresholded at p < .05,
FDR corrected. These thresholded maps were then used
in conjunction analyses (using the aforementioned log-
ical “AND” masking procedure) to identify areas of com-
mon functional connectivity to both SM > SP “AND” OM >
OP. These conjunction analyses were run on each seed
region independently. The resulting conjunction maps for
each seed region were then used to compute a final con-
junction map that isolated shared functional connectivity
for SM and OM that was common to every seed region.
Lombardo et al.
1625
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
F
.
t
.
.
/
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 1. Shared Neural Mentalizing Representations for Self AND Other
Hemisphere
BA
MNI (X,y,z)
t
Cluster Size
BA
MNI (X,y,z)
SM > SP
OM > OP
Region
vMPFC
PCC
TPJ
TPJ
ATL
SI/MI
Cerebellum
l
l
B
R
R
l
l
l
l
l
l
R
10, 11
−6, 54, −8
30
30
19
37
39
20
21
21
4
4
−4, −56, 12
6, −54, 10
16, −50, −2
54, −58, 14
−44, −66, 18
−56, −10, −18
−54, 4, −28
−54, −6, −26
−34, −34, 54
−32, −22, 58
26, −38, −24
4.39
10.65
56
2285
7.64
5.95
6.29
4.8
3.37
3.08
2.91
3.76
3.54
4.93
333
247
46
88
93
10, 11
−4, 50, −8
19
30
19
37
39
21
21
20
3
−16, −52, −2
−6, −58, 16
16, −52, −4
52, −60, 14
−46, −64, 18
−56, −4, −26
−54, 4, −28
−58, −12, −18
−32, −32, 58
t
4.73
8.25
7.33
6.67
7.27
5.23
4.68
3.94
3.91
3.96
28, −40, −20
4.56
Shared neural representations for self and other during mentalizing judgments. All coordinates are reported from the conjunction analysis of overlap
between the SM > SP contrast and the OM > OP contrast ( pag < .05, FDR-corrected threshold for each contrast).
SM = self-mentalizing; SP = self-physical; OM = other-mentalizing; OP = other-physical; vMPFC = ventromedial prefrontal cortex; PCC = posterior
cingulate cortex/precuneus; ATL = anterior temporal lobe; SI/MI = primary sensorimotor cortex.
To identify areas of functional connectivity specific to
SM or OM, interaction effect analyses were conducted on
the PPI contrasts using paired samples t tests. These paired
samples t test compared the PPI contrasts of SM > SP to
OM > OP and were done individually on all seed regions.
All paired samples t tests were thresholded at p < .05, FDR
corrected.
To corroborate the direction of the relationships iden-
tified with PPI, we used the following procedure. Briefly,
the raw time courses of the seed and target ROIs (target
ROIs of FO, PMv, pre-SMA, and SI/MI were defined from
the final conjunction map of shared connectivity among
all seed regions) were extracted and multiplied by an
HRF-convolved task vector of each condition. Correlations
were computed separately for each condition among the
seed and target ROIs. Correlations coefficients were con-
verted into Fisherʼs z scores (Steiger, 1980) for the pur-
poses of visual comparisons of correlations between
conditions.
RESULTS
Behavioral Data
RTs for all other conditions were statistically equivalent to
one another ( p > .05).
Shared Neural Representations
Using conjunction analyses, we defined shared mentaliz-
ing representations as the overlap between the SM > SP
contrast and the OM > OP contrast (each thresholded at
pag < .05, FDR corrected). These areas of overlap consisted
of the vMPFC (Brodmannʼs area [BA] 10/11), PCC (BA 30/
19), and bilateral TPJ (BA 37/39) as well as the left ante-
rior temporal lobe (ATL) along the middle temporal
gyrus (BA 20/21), left primary sensorimotor cortex (SI/
MI; BA 3/4), and cerebellum (see Figure 1A and Table 1).
To identify any interaction effects, we examined the
paired samples t test comparing the SM > SP contrast
images to the OM > OP contrast images ([SM > SP] >
[OM > OP]). This analysis revealed no significant results.
The absence of any interactions within our factorial design
combined with the results of the conjunction analyses sig-
nals that mentalizing representations about the self and
others are largely recruiting identical neural circuitry.
Participants were significantly faster to make judgments
about the self in a mentalizing context (SM, mean =
2413.65, DE = 56.72) compared with all other conditions
(OM, mean = 2564.49, DE = 58.87; SP, mean = 2523.14,
DE = 52.31; OP, mean = 2565.45, DE = 57.63; pag < .001).
Self–Other Distinction
Although mentalizing representations shared the same
neural space for self and other, three of the shared menta-
lizing regions (vMPFC, PCC, and RTPJ) were also identified
1626
Journal of Cognitive Neuroscience
Volume 22, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
7
e
-
1
p
6
d
2
f
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
o
2
c
3
n
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
2
8
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
/
.
f
.
o
n
1
8
M
a
y
2
0
2
1
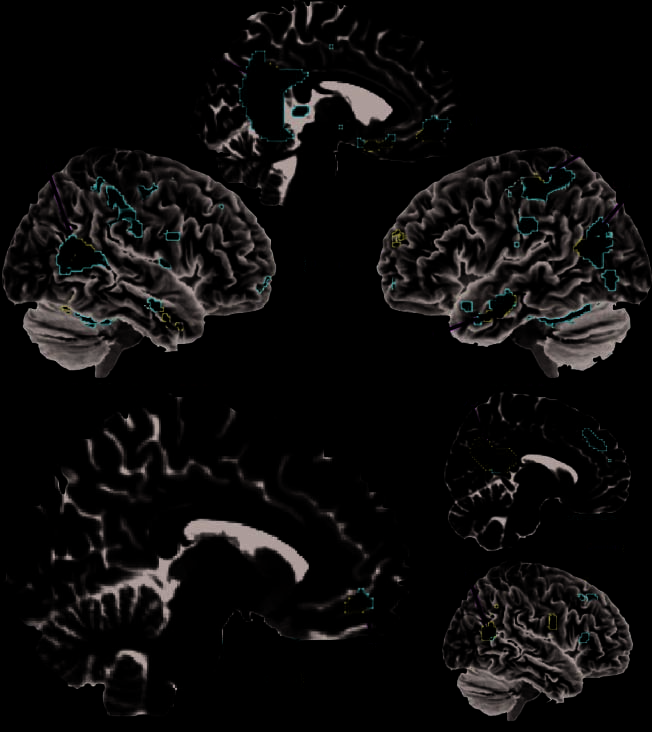
Figure 1. Conjunction analyses
activation results. This figure
plots the activation results for
the conjunction analysis of
(A) SM compared with SP
judgments (red voxels)
superimposed on the results
of the OM compared with
OP judgments (blue voxels).
The white voxels show the
overlap in mentalizing
representations for both
self and other. (B) Activation
results for the self-conjunction
analysis of SM judgments
compared with OM judgments
(red voxels) superimposed
on the results of SP judgments
compared with OP judgments
(blue voxels). Again, white
voxels denote the overlap for
self-judgments compared
with other-judgments. (C)
Results of the other-conjunction
analysis for OM compared with
SM judgments (red voxels)
superimposed on the contrast
of OP compared with SP
judgments (blue voxels).
The white voxels denote the
overlap for other judgments.
Each contrast is thresholded
independently at p < .05,
FDR corrected.
as biased toward target-specific information processing.
In another set of whole-brain conjunction analyses of
the SM > OM and SP > OP contrasts (thresholded at p <
.05, FDR corrected), vMPFC (BA 10/11) was the only region
identified as having such a self > other bias (ver Figura 1B
and Table 2A). A diferencia de, the conjunction of OM > SM
and OP > SP contrasts revealed a general other > self bias
in both right PCC (BA 23/29/30) and RTPJ (BA 37; ver
Figure 1C and Table 2B).
Shared Functional Connectivity
Próximo, we ran PPI analyses to identify areas that are more
functionally connected during SM and OM compared
with connectivity during physical judgments. Three seed
regions were specified in vMPFC, PCC, and RTPJ. Estos
seed regions consisted of all suprathreshold voxels in
each respective region that were identified from the SM >
SP and OM > OP group-level conjunction analysis. Para
each seed, two separate PPI analyses were conducted;
one identified increased functional connectivity during
SM > SP, whereas the second identified increased func-
tional connectivity during OM > OP. Each PPI analysis
was thresholded at p < .05, FDR corrected. Conjunction
maps of the overlap between the PPI results for SM > SP
and OM > OP were examined to identify connectivity
common to both SM and OM within each seed region.
To test for any distinct connectivity patterns as a function
of either SM or OM, paired sample t tests ([SM > SP] >
[OM > OP]) for each seed region were computed and
thresholded at p < .05, FDR corrected.
Within each of the three shared mentalizing seed re-
gions (vMPFC, PCC, and RTPJ), robust increased func-
tional connectivity was observed during SM > SP and
OM > OP in a widely distributed neural system consist-
ing of the medial pre-SMA extending into SMA and
cACC (BA 6/24/32), left anterior temporal pole extend-
ing into AI, ventrolateral prefrontal cortex, and lateral
left FO and dorsal premotor cortex
OFC (BA 47 ),
(PMd) and PMv (BA 6/44/48), left SI/MI (BA 3/4), y
adjacent left anterior intraparietal sulcus (BA 19) como
well as visual cortex (BA 18) extending into the cere-
bellum. Hippocampal formation connectivity was also in-
creased during SM > SP, but only for the PCC and RTPJ
seeds (see Figure 2A and Table 3A–C). Curiosamente, estafa-
nectivity within AI and hippocampal formation during
OM > OP was below the FDR threshold among all three
seeds.
Lombardo et al.
1627
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
/
t
.
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 2. Biases for Self or Other
(A) Self-bias [SM > OM] y [SP > OP]
Region
vMPFC
Hemisphere
BA
l
10, 11
(B) Other-bias [OM > SM] y [OP > SP]
Region
Hemisphere
PCC
TPJ
R
R
R
R
BA
29
23
37
SM > OM
MNI (X,y,z)
−8, 52, −8
OM > SM
MNI (X,y,z)
8, −44, 14
8, −58, 28
t
5.9
Cluster Size
BA
13
10, 11
Cluster Size
196
t
4.56
4.43
BA
23
30
29
37
50, −60, 14
4.89
55
SP > OP
MNI (X,y,z)
−6, 50, −8
OP > SP
MNI (X,y,z)
10, −60, 32
8, −54, 20
6, −42, 14
52, −62, 14
t
5.75
t
6.51
4.86
4.76
4.35
Target-specific biases in processing for self (A) or other (B). All coordinates are reported from the conjunction analysis of the overlap between the
SM > OM contrast and the SP > OP contrast (Table 2A) or the conjunction of the OM > SM contrast and OP > SP contrast (Table 2B). Each contrast
was thresholded at p < .05, FDR corrected.
SM = self-mentalizing; SP = self-physical; OM = other-mentalizing; OP = other-physical; vMPFC = ventromedial prefrontal cortex; PCC = posterior
cingulate cortex/precuneus.
Corroborating the idea that the difference in functional
connectivity during SM > SP was statistically indistin-
guishable across the whole brain to the difference in
functional connectivity during OM > OP, a paired sample
t test of [SM > SP] > [OM > OP] revealed no significant
resultados.
Finalmente, we computed another conjunction map to
identify shared mentalizing connectivity that is common
to all three seed regions. This was done by first comput-
ing a conjunction map of overlap among all seed regions
within the SM > SP PPI analyses. The same procedure
was carried out for OM > OP PPI analyses. A final con-
junction map extracted the overlap between the SM >
SP and the OM > OP PPI conjunction maps. De este modo, qué
this final conjunction map reflects is the shared connec-
tivity for both SM and OM that is common to all three
seed regions. This conjunction map highlighted pre-
SMA, FO, PMv, SI/MI, anterior intraparietal sulcus, y
visual cortex/cerebellum as convergent areas of increased
functional connectivity during both SM and OM that are
common to all three seed regions (see Figure 2B).
Given that the PPI results are a measure of connectivity
change between two experimental conditions, a positive
PPI result could have been the result of a positive corre-
lation between the seed and a target region for mentaliz-
ing whereas the physical condition exhibits a negative or a
nonsignificant correlation. Sin embargo, a positive PPI result
could have also resulted from a strong negative correla-
tion in the physical condition and a nonsignificant or
less negative correlation in the mentalizing condition.
To disambiguate the PPI results, we followed the PPI
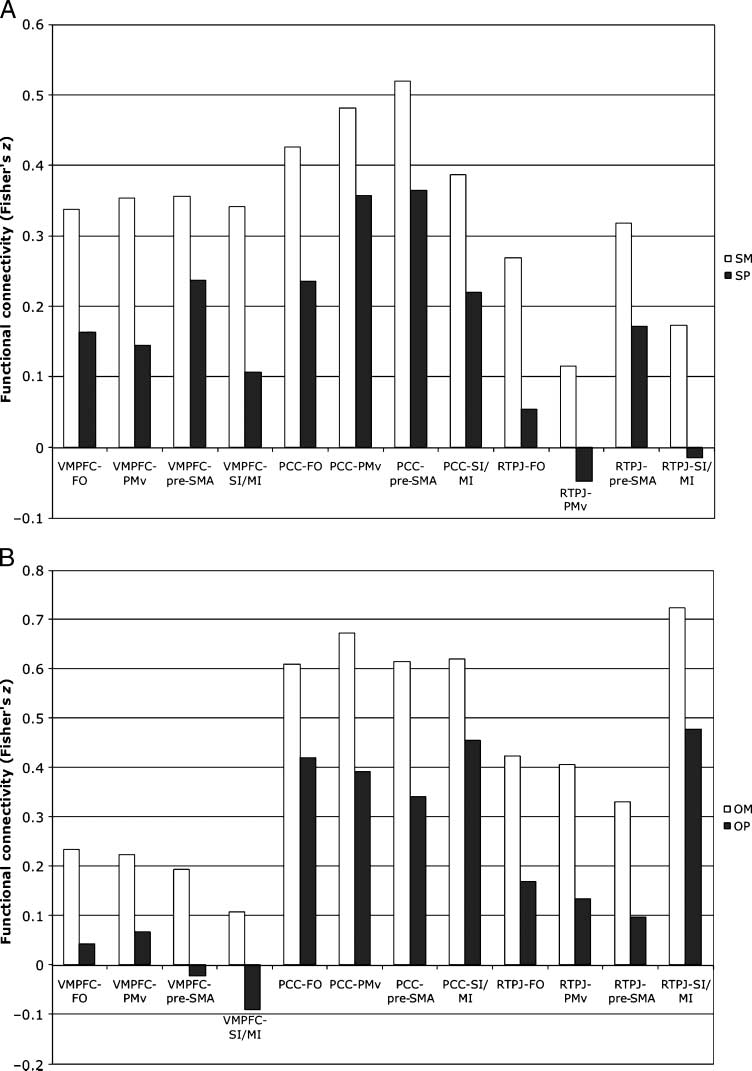
analyses by extracting the correlations for the seed
and the target ROIs for each condition separately (ver
Métodos). Visual examination of the correlations sepa-
rately for each condition confirmed that each correla-
tion between seed and target ROIs was always more
positive in the mentalizing condition compared with
the physical condition (see Figure 3A and B).
DISCUSIÓN
Previous research has emphasized the crucial impor-
tance of shared neural representations in demonstrating
how we understand others via ourselves (Gazzola &
Keyser, 2009; Amodio & Frith, 2006; Decety & Grezes,
2006; Gazzola et al., 2006; hombre de oro, 2006; Iacoboni &
Dapretto, 2006; Mitchell et al., 2006; Saxe et al., 2006;
Blakemore et al., 2005; Ochsner et al., 2005; Keyser
et al., 2004; Rizzolatti & Craighero, 2004; Singer et al.,
2004; Carr et al., 2003; Wicker et al., 2003). The current
study provides independent replication of this and also
provides many substantial new insights. Primero, with our
factorial design, we were able to test the interaction
effect of whether mentalizing or physical representations
recruit distinct regions for the self or other. The absence
of such an effect in our large sample suggests that men-
talizing representations about the self or others are not
recruiting distinct and independent neural systems. En-
lugar, we found robust evidence that mentalizing repre-
sentations are distributed across similar neural systems
with respect to self and other.
In the domain of high-level inference-based mentaliz-
En g, many of the areas identified in the current study have
1628
Revista de neurociencia cognitiva
Volumen 22, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
/
.
F
.
.
t
oh
norte
1
8
METRO
a
y
2
0
2
1
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
/
.
t
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
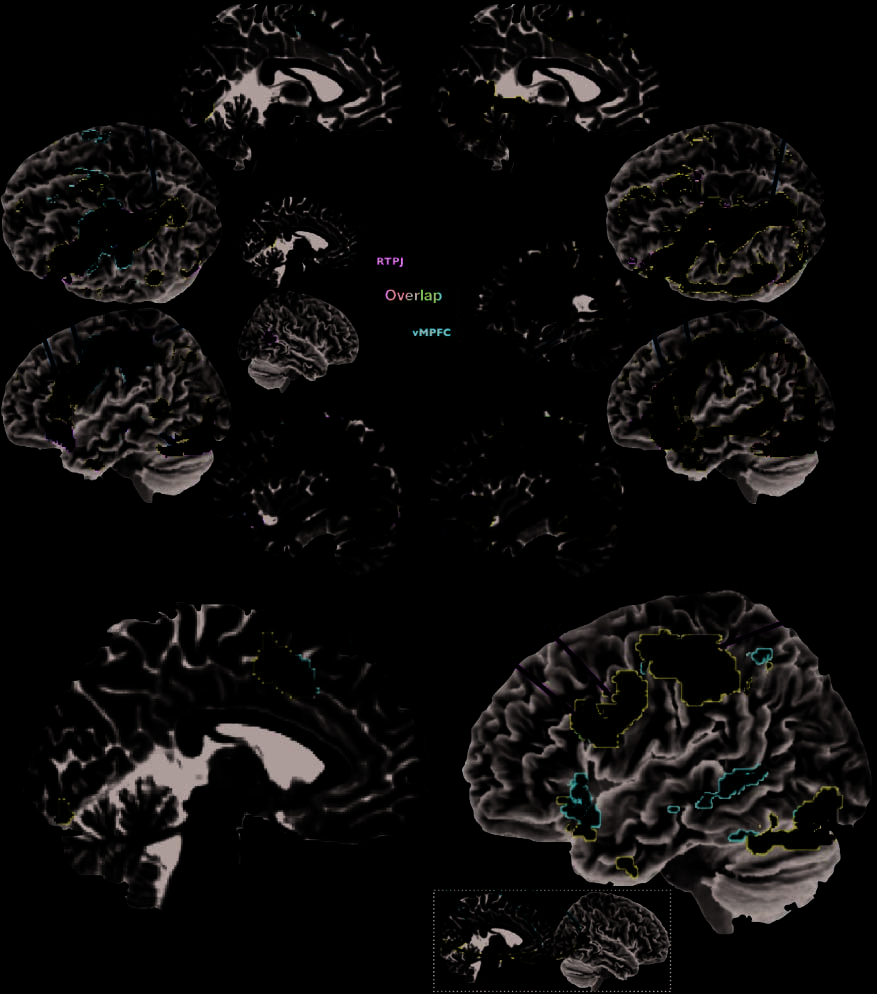
Cifra 2. Shared mentalizing connectivity from vMPFC, PCC, and RTPJ. (A, left side) Increases in functional connectivity during OM judgments
compared with connectivity during OP judgments (OM > OP) from vMPFC (red voxels), PCC (blue voxels), and RTPJ (green voxels). (A, right side)
Same results, but for when connectivity is increased during SM judgments compared with connectivity during SP judgments (SM > SP). Purple
and turquoise voxels denote overlapping connectivity among two of three seed regions. White voxels denote connectivity patterns that are
common to all three seed regions. (B) Increases in functional connectivity common to all three seed regions for SM compared with connectivity
during SP judgments (SM > SP; red voxels) superimposed on top of connectivity increases common to all three seed regions under OM compared
with connectivity during OP judgments (OM > OP; blue voxels). White voxels are areas where functional connectivity converges among all
three seeds for both SM and OM.
been previously identified as crucial for mentalizing,
a saber, the vMPFC, PCC, RTPJ, and ATL (Amodio & Frith,
2006; Mitchell et al., 2006; sajonia & Powell, 2006; sajonia
et al., 2006; Ochsner et al., 2005; Frith & Frith, 2003; sajonia
& Kanwisher, 2003). Sin embargo, what was particularly
interesting and new was the observation that left SI/MI
was also sensitive to mentalizing about both the self
and other. The role of somatosensory cortex in low-level
shared representations of touch (Blakemore et al., 2005;
Keysers et al., 2004), self-experienced pain (Singer et al.,
2004), and action–perception mirroring (Gazzola &
Keyser, 2009; Gazzola et al., 2006) is well established,
Lombardo et al.
1629
Mesa 3. Shared Mentalizing Connectivity
Region
Hemisphere
BA
MNI (X,y,z)
t
Cluster Size
BA
MNI (X,y,z)
t
SM > SP
OM > OP
(A) vMPFC Connectivity [SM > SP] y [OM > OP]
Pre-SMA/SMA/cACC
SI/MI
FO
PMv
PMd
TP
Visual cortex/cerebellum
Visual cortex/cerebellum
IPS
B
B
B
l
l
l
l
l
l
l
l
l
l
l
l
R
R
R
l
32
32
6
3
3
3
48
6
6
38
38
18
19
19
18
19
7
−8, 14, 42
8, 18, 40
10, 8, 48
−36, −28, 52
−48, −24, 54
−40, −30, 60
−44, 16, 22
−56, 2, 32
−32, −6, 46
−40, 22, −18
−50, 26, −14
−28, −84, −14
−42, −78, −14
32, −82, −8
22, −84, −10
32, −84, −18
−30, −56, 52
(B) PCC Connectivity [SM > SP] y [OM > OP]
Pre-SMA/SMA/cACC
SMA
SI/MI/IPS/PMd/PMv/FO/AI
TP
ATL
Visual cortex/cerebellum
MTG
IPS
dMPFC
B
B
B
l
l
l
l
l
l
l
B
B
B
l
R
l
32
32
−2, 14, 46
10, 10, 44
6/24
−4, −6, 50
6
3
6
6
38
20
20
18
18
18
21
19
9
−4, −8, 66
−44, −28, 52
−52, 4, 36
−44, −4, 50
−44, 18, −18
−44, 0, −40
−48, 6, −34
14, −86, −10
−16, −90, 4
−8, −88, −10
−54, −44, 0
30, −66, 36
−10, 56, 30
4.47
3.5
3.21
3.22
3.08
3.07
3.02
2.85
3.57
3.66
3.28
3.08
2.71
3.02
2.85
2.57
3.24
5.69
4.04
3.92
2.67
7.51
6.79
5.9
4.15
3.71
3.17
7.56
6.85
6.66
6.22
3.99
2.84
431
32
−8, 14, 48
4.9
441
46
38
91
122
383
93
13
3
4
3
−46, −22, 52
−38, −22, 60
−38, −34, 50
44
−46, 10, 26
4
6
6
38
38
19
19
18
18
18
19
7
−56, 0, 30
−48, 0, 32
−36, −4, 52
−42, 26, −18
−52, 28, −12
−34, −84, −22
−34, −78, −14
−22, −88, −12
20, −84, −12
26, −86, −18
36, −82, −18
−32, −54, 48
5.3
4.86
4.63
4.67
3.77
3.7
4.43
4.68
4.45
4.02
3.93
3.81
4.39
4.2
3.74
3.25
971
32
−4, 8, 48
5.63
18
4959
42
52
3691
152
66
12
6
3
6
6
38
20
18
19
19
21
19
10
−6, −6, 66
−38, −34, 48
−36, −8, 58
−44, 0, 42
−40, 22, −22
−46, 0, −36
−26, −90, −8
−34, −86, −14
−40, −80, 14
−58, −52, 4
34, −66, 36
−6, 56, 28
4.79
6.52
6.31
6.25
3.58
4.21
6.26
6.08
6.05
4.94
3.35
3.2
1630
Revista de neurociencia cognitiva
Volumen 22, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
F
/
.
.
t
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 3. (continued )
Region
Hemisphere
BA
MNI (X,y,z)
t
Cluster Size
BA
MNI (X,y,z)
t
SM > SP
OM > OP
(C) RTPJ Connectivity [SM > SP] y [OM > OP]
Pre-SMA/SMA/MCC
SI/MI/PMd/PMv/FO
TP/AI/ VLPFC
Visual cortex/cerebellum/IPS
MTG
B
l
l
l
l
l
R
l
l
l
32
3
2
6
38
47
18
19
18
21
−4, 12, 48
−40, −30, 58
−48, −32, 50
−54, 6, 34
−44, 20, −12
−42, 26, −6
10, −86, −6
−34, −82, −20
−14, −88, −18
−52, −44, −2
5.57
5.62
4.78
4.78
4.25
3.95
7.36
6.18
5.85
4.84
454
1786
496
5002
269
6/32
−4, 2, 52
2
48
40
38
45
38
18
18
18
21
−46, −34, 50
−48, 16, 28
−38, −34, 46
−36, 22, −22
−52, 26, 0
−46, 30, −12
−8, −88, −12
−20, −88, −16
−26, −90, −10
−60, −52, 4
3.68
4.9
4.84
4.62
4.15
3.15
2.79
4.93
4.78
4.76
2.94
Shared functional connectivity for mentalizing about the self and other for (A) vMPFC, (B) PCC, y (C) RTPJ seed regions. All coordinates are
reported from the conjunction analysis of the overlap between the SM > SP PPI contrast and the OM > OP PPI contrast ( p > .05, FDR-corrected
threshold for each contrast).
SM = self-mentalizing; SP = self-physical; OM = other-mentalizing; OP = other-physical; vMPFC = ventromedial prefrontal cortex; PCC = posterior
cingulate cortex/precuneus; pre-SMA = presupplementary motor area; cACC = caudal ACC; dMPFC = dorsomedial prefrontal cortex; SI/ MI =
primary sensorimotor cortex; VLPFC = ventrolateral prefrontal cortex; FO = frontal operculum; AI = anterior insula; PMv = ventral premotor
corteza; PMd = dorsal premotor cortex; ATL = anterior temporal lobe; TP = temporal pole; IPS = intraparietal sulcus; MTG = middle temporal
gyrus.
and the disruption of somatosensory cortex (via trans-
cranial magnetic stimulation or lesion studies) impairs
emotion recognition (Pitcher, Garrido, Walsh, & duchaína,
2008; Adolphs, Damasio, Tranel, Cooper, & Damasio, 2000)
as well as automatic motor- and somatosensory-evoked
potentials when viewing actions or others in pain (Avenanti,
Bolognini, Maravita, & Aglioti, 2007; Bufalari, Aprile,
Avenanti, Di Russo, & Aglioti, 2007; Avenanti, Bueti, Galati,
& Aglioti, 2005). Además, in mirror-touch synesthe-
sia, shared representations of touch are heightened in
SI/SII (Blakemore et al., 2005), and such individuals are
higher on measures of empathy (Banissy & Ward, 2007).
De este modo, the current observation that SI/MI is also recruited
for mentalizing about self and other suggests that such
low-level embodied/simulative shared representations
computed by SI/MI are also important for the processes
underlying more inference-based mentalizing when com-
pared with reflecting on physical characteristics.
Although the neural implementation of both SM and
OM was identical, it cannot be said that the brain is ag-
nostic with regard to distinguishing self from other. On
the contrary, the brain does make a general self–other dis-
tinction and does so within neural circuitry that is also
biased toward mentalizing. Replicating past research (kelly
et al., 2002), we found that the vMPFC was biased for self-
referential processing in general, whereas the PCC and the
RTPJ were biased for other-referential processing (Pfeifer,
Lieberman, & Dapretto, 2007; Saxe et al., 2006; Ruby &
Decety, 2001). Sin embargo, regarding the other > self effect
in the PCC, we note that other studies find the opposite
effect of self > other in PCC (DʼArgembeau et al., 2008;
Kelley et al., 2002). One explanation for these opposing
observations may be due to the different task demands
from studies that find opposing effects. Studies that find
an other > self effect (including the current study) probe
event-specific judgments about the self and other (Pfeifer
et al., 2007; Ruby & Decety, 2001), whereas more general-
ized trait inferences about self and other are probed in
studies that find a self > other effect (DʼArgembeau et al.,
2008; Kelley et al., 2002). These subtle differences in task
demands may alter the underlying memory retrieval pro-
cesses that are integral to PCC functioning (Cavanna &
Trimble, 2006; Maddock, Garrett, & Buonocore, 2001;
Henson, Rugg, Shallice, Josephs, & Dolan, 1999). Future
work should systematically explore this interaction be-
tween the possible deployment of different retrieval pro-
cesses in PCC during specific versus general information
processing about the self and others.
Although the current study isolated several regions
coding for shared mentalizing via standard fMRI contrast
analiza, one of the main contributions of the current
study is in demonstrating that results from standard
fMRI contrast analyses are only the beginning of making
such an inference. We have demonstrated that functional
Lombardo et al.
1631
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
t
F
/
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Cifra 3. Direction of PPI
efectos. This figure depicts
the relationships among the
seeds and each target region
shown in Figure 2B for the
PPI of (A) SM compared with
SP judgments (SM > SP) y
(B) OM compared with OP
judgments (OM > OP). Visual
inspection shows that the
mentalizing conditions (white
barras) are always greater positive
correlations compared with
the physical conditions (gray
barras). Error bars are not
plotted here because statistical
inference on these comparisons
has already being established
from the whole-brain PPI
analiza.
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
t
/
.
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
connectivity approaches are necessary for fully qualifying
statements that shared neural representations for self
and other are in fact participating in the same underlying
processes via a similar neural implementation. It is clear
from our connectivity analyses that high-level inference-
based mentalizing areas such as vMPFC, PCC, and RTPJ
are participating in identical neural circuits for both
reflecting on oneʼs own mental states and reflecting on
anotherʼs mental states. Without such insights from
connectivity approaches, shared neural representations,
while indexing some kind of similarity (p.ej., a convergence
zone), might also be participating in functionally distinct
neural circuits that may be performing very different types
of processes (Jabbi et al., 2008; Zaki et al., 2007).
In addition to demonstrating an identical neural imple-
mentation for mentalizing about the self and other, el
distribution of where such implementation arose has
substantial theoretical implications about the debate
between contrasting high-level inference-based models
of mentalizing with lower level embodied/simulationist
accounts. Históricamente, scholars have pinned high-level
inference-based accounts and low-level embodied/
1632
Revista de neurociencia cognitiva
Volumen 22, Número 7
simulationist accounts as opposites of each other (gopnik
& Wellman, 1992; gordon, 1992). Sin embargo, as recent
scholars have pointed out (Keyser & Gazzola, 2007; Uddin
et al., 2007; hombre de oro, 2006), perhaps such arguments are
missing a third alternative; eso es, perhaps higher level
inference-based processes are grounded in their inter-
actions with lower level embodied/simulation-based
procesos (hombre de oro & de Vignemont, 2009; Barsalou,
1999, 2008). The prediction here would be that high-
level inference-based mentalizing systems (p.ej., vMPFC,
PCC, RTPJ) are integrating their signal with lower level
embodied/simulation-based systems (p.ej., FO/PMv, IPL,
AI, SI/MI, pre-SMA; Keyser & Gazzola, 2007). One piece
of evidence supporting this integration hypothesis is
based on the earlier observation of the shared activation
of SI/MI for SM and OM. Además, the connectivity
analyses went further to directly test the prediction that
these two systems were specifically linked during men-
talizing more than during physical judgments. The pat-
terns of connectivity strikingly map onto this prediction.
Even more striking was that such patterns of connectivity
were apparent for both SM and OM. De este modo, the patterns of
connectivity combined with the identical neural imple-
mentation for both SM and OM are the first formal obser-
vations of such an integration between two paramount
neural systems for social cognition. These results support
the idea that during high-level mentalizing, there is a mid-
dle ground where both inference-based processes merge
with lower level embodied/simulative processes.
Finalmente, one important point for which the current re-
sults have some bearing is on the link between the litera-
ture on the default mode network (DMN) of intrinsic
functional brain organization (Fox & Raichle, 2007).
Although it is tempting to link the activation maps of
shared mentalizing results with the DMN, we would sug-
gest two reasons for resisting interpretation of the cur-
rent findings in relation to the DMN literature. Primero, como
Morcom and Fletcher (2007) point out, the cognitive/
psychological nature of what happens in the DMN at rest
is unknown, and thus interpretations of well-controlled
task-specific differences in signal change (as in the current
estudiar) stand relatively independently of what they mean
in relation to the unconstrained and unknown cognitive/
psychological nature of intrinsic DMN organization.
We also have empirical reasons to resist interpreting
the current findings as simply an extension of intrinsic
DMN resting state organization. Fox et al. (2005) elegantly
showed that the intrinsic connectivity of the DMN is like
an internal feedback loop; eso es, the time course of
signal in MPFC, PCC, and lateral parietal regions is posi-
tively correlated with each other. Sin embargo, Fox et al.
extended this by also showing that MPFC, PCC, y
lateral parietal regions are negatively correlated with re-
gions similar to those found in our connectivity maps:
lateral PMd and PMv, medial SMA/pre-SMA, FO, AI,
and intraparietal sulcus. The striking difference here is
that whereas the current studyʼs task-specific connectiv-
ity analyses observed this same network, Encontramos eso
connectivity was in the opposite direction (es decir., positivo
correlations) of what Fox et al. found (es decir., negative cor-
relaciones). We found that vMPFC, PCC, and RTPJ were
positively correlated with a similar network to what
Fox et al. found was negatively correlated at rest with
MPFC, PCC, and a lateral parietal region near TPJ. If the
task-specific shared mentalizing circuits were simply the
same network involved in intrinsic resting state organi-
zación (with the same kinds of cognitive/psychological
interpretations associated with it), we should have ob-
served task-specific increases in mentalizing connectivity
from vMPFC, PCC, and RTPJ similar to an internal feedback
loop; eso es, vMPFC, PCC, and RTPJ should have been posi-
tively correlated with each other during SM and OM. Given
the observation that this network shows opposing patterns
of connectivity between rest and mentalizing, we suggest
that the intrinsic mode of functional brain organization is
exactly the opposite of what occurs during task-specific
shared mentalizing processes for both self and other. De este modo,
if there are any interpretations to be made about the cur-
rent findings in relation to the DMN literature, it would be
that during mentalizing about the self and other, hay
possibly an adaptive reconfiguration of dynamic functional
organization from how the brain is naturally functionally
organizado (bassett, Meyer-Lindenberg, Achard, Duke, &
bullmore, 2006).
En resumen, we used fMRI and a functional con-
nectivity approach to test whether the neural imple-
mentation of high-level inference-based mentalizing
processes was similar or different with respect to the self
and other. Our findings show that such neural imple-
mentation is indeed identical across activation contrasts
and functional connectivity for SM and OM. Además,
the tight link between high-level inference-based men-
talizing systems and low-level embodied/simulation-
based systems suggests that these two neural systems
for social cognition are integrated in a task-specific
manner for mentalizing about both self and other. Estos
observations provide a first glimpse at how such an inte-
gration takes place and provides the groundwork for
further theoretical refinement and empirical work as well
as translational work into how such systems may be dis-
rupted, as is the case for neurodevelopmental conditions
such as autism (Lombardo, Barnes, Wheelwright, & Baron-
cohen, 2007; Dapretto et al., 2006).
Expresiones de gratitud
The authors thank Jason Mitchell and Adrianna Jenkins for
generously letting us use their stimuli and Mike Cohen,
Matthew Belmonte, Caroline Robertson, Teresa Tavassoli, y
anonymous reviewers for their valuable discussion and com-
mentos. They also acknowledge the generous funding of the
Shirley Foundation, the Cambridge Overseas Trust, y el
Medical Research Council (MRC) as well as the support from
the MRC Autism Imaging Multi-Centre Study (AIMS) Consorcio.
Lombardo et al.
1633
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
/
F
t
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Competing Interests Statement
mi. t. B. is employed half-time by the University of Cambridge
and half-time by GlaxoSmithKline plc. None of the other authors
have any other biomedical financial interests or potential con-
flicts of interest. This work was conducted in association with
the NIHR-CLAHRC for Cambridgeshire and Peterborough NHS
Mental Health Trust.
Reprint requests should be sent to Michael V. Lombardo, Au-
tism Research Centre, Department of Psychiatry, Universidad de
Cambridge, Douglas House, 18B Trumpington Road, Cambridge,
CB2 8AH, Reino Unido, o por correo electrónico: ml437@cam.ac.uk.
REFERENCIAS
Adolphs, r., Damasio, h., Tranel, D., Cooper, GRAMO., & Damasio,
A. R. (2000). A role for somatosensory cortices in the visual
recognition of emotion as revealed by three-dimensional
lesion mapping. Revista de neurociencia, 20, 2683–2690.
Amodio, D. METRO., & Frith, C. D. (2006). Meeting of minds: El
medial frontal cortex and social cognition. Reseñas de naturaleza
Neurociencia, 7, 268–277.
Avenanti, A., Bolognini, NORTE., Maravita, A., & Aglioti, S. METRO. (2007).
Somatic and motor components of action simulation.
Biología actual, 17, 2129–2135.
Avenanti, A., Bueti, D., Galati, GRAMO., & Aglioti, S. METRO. (2005).
Transcranial magnetic stimulation highlights the
sensorimotor side of empathy for pain. Naturaleza
Neurociencia, 8, 955–960.
Banissy, METRO. J., & Ward, j. (2007). Mirror-touch synesthesia is
linked with empathy. Neurociencia de la naturaleza, 10, 815–816.
Barsalou, l. W.. (1999). Perceptual symbol systems. conductual
and Brain Sciences, 22, 577–609.
Barsalou, l. W.. (2008). Grounded cognition. Annual Reviews of
Psicología, 59, 617–645.
bassett, D. S., Meyer-Lindenberg, A., Achard, S., Duke, T., &
bullmore, mi. (2006). Adaptive reconfiguration of fractal
small-world human brain functional networks. Actas de
the National Academy of Sciences, EE.UU., 103, 19518–19523.
Blakemore, S. J., Bristow, D., Bird, GRAMO., Frith, C., & Ward, j.
(2005). Somatosensory activations during the observation
of touch and a case of vision–touch synaesthesia. Cerebro, 128,
1571–1583.
Bufalari, I., Aprile, T., Avenanti, A., Di Russo, F., & Aglioti, S. METRO.
(2007). Empathy for pain and touch in human somatosensory
corteza. Corteza cerebral, 17, 2553–2561.
Carr, l., Iacoboni, METRO., Dubeau, METRO. C., Mazziotta, j. C., & Lenzi,
GRAMO. l. (2003). Neural mechanisms of empathy in humans: A
relay from neural systems for imitation to limbic areas.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
100, 5497–5502.
Cavanna, A. MI., & Trimble, METRO. R. (2006). The precuneus: A
review of its functional anatomy and behavioural correlates.
Cerebro, 129, 564–583.
Critchley, h. D., Wiens, S., Rotshtein, PAG., Ohman, A., & Dolan,
R. j. (2004). Neural systems supporting interoceptive
awareness. Neurociencia de la naturaleza, 7, 189–195.
Dapretto, METRO., Davies, METRO. S., Pfeifer, j. h., Scott, A. A., Sigman,
METRO., Bookheimer, S. y., et al. (2006). Understanding emotions
in others: Mirror neuron dysfunction in children with autism
spectrum disorders. Neurociencia de la naturaleza, 9, 28–30.
DʼArgembeau, A., Feyers, D., Majerus, S., Collette, F., Van der
Linden, METRO., Maquet, PAG., et al. (2008). Self-reflection across
tiempo: Cortical midline structures differentiate between
present and past selves. Social Cognitive and Affective
Neurociencia, 3, 244–252.
Decety, J., & Grezes, j. (2006). The power of simulation:
Imagining oneʼs own and otherʼs behavior. Brain Research,
1079, 4–14.
Fox, METRO. D., & Raichle, METRO. mi. (2007). Spontaneous fluctuations in
brain activity observed with functional magnetic resonance
imaging. Naturaleza Reseñas Neurociencia, 8, 700–711.
Fox, METRO. D., Snyder, A. Z., Vincent, j. l., Corbetta, METRO., VanEssen,
D. C., & Raichle, METRO. mi. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
102, 9673–9678.
Friston, k. J., Buchel, C., Fink, GRAMO. r., morris, J., Rolls, MI., &
Dolan, R. j. (1997). Psychophysiological and modulatory
interactions in neuroimaging. Neuroimagen, 6, 218–229.
Frith, Ud., & Frith, C. D. (2003). Development and
neurophysiology of mentalizing. Philosophical Transactions
of the Royal Society of London, Serie B, Ciencias Biologicas,
358, 459–473.
Gazzola, v., Aziz-Zadeh, l., & Keyser, C. (2006). Empathy and
the somatotopic auditory mirror system in humans. Actual
Biología, 16, 1824–1829.
Gazzola, v., & Keyser, C. (2009). The observation and
execution of actions share motor and somatosensory voxels
in all tested subjects: Single-subject analyses of unsmoothed
fMRI data. Corteza cerebral, 19, 1239–1255.
hombre de oro, A. (2006). Simulating minds: The philosophy,
psicología, and neuroscience of mind reading. Nueva York:
prensa de la Universidad de Oxford.
hombre de oro, A., & de Vignemont, F. (2009). Is social cognition
embodied? Tendencias en Ciencias Cognitivas, 13, 154–159.
gopnik, A., & Wellman, h. (1992). Why the childʼs theory
of mind really is just a theory. Mind and Language, 7,
145–171.
gordon, R. METRO. (1992). The simulation theory: Objections and
misconceptions. Mind and Language, 7, 11–34.
Henson, R. norte. A., Rugg, METRO. D., Shallice, T., Josephs, o., &
Dolan, R. j. (1999). Recollection and familiarity in
recognition memory: An event-related functional magnetic
resonance imaging study. Revista de neurociencia, 19,
3962–3972.
Iacoboni, METRO., & Dapretto, METRO. (2006). The mirror neuron system
and the consequences of its dysfunction. Reseñas de naturaleza
Neurociencia, 7, 942–951.
Jabbi, METRO., Bastiaansen, J., & Keyser, C. (2008). A common
anterior insula representation of disgust observation,
experiencia, and imagination shows divergent functional
connectivity pathways. Más uno, 3, e2939.
Jenkins, A. C., Macrae, C. NORTE., & mitchell, j. PAG. (2008). Repetition
suppression of ventromedial prefrontal activity during
judgments of self and others. Actas del Nacional
Academia de Ciencias, EE.UU., 105, 4507–4512.
kelly, W.. METRO., Macrae, C. NORTE., Wyland, C. l., Caglar, S., Inati, S.,
& Heatherton, t. F. (2002). Finding the self ? An event-related
fMRI study. Revista de neurociencia cognitiva, 14, 785–794.
Keyser, C., & Gazzola, V. (2007). Integrating simulation and
theory of mind: From self to social cognition. Tendencias en
Cognitive Sciences, 11, 194–196.
Keyser, C., Wicker, B., Gazzola, v., Anton, j. l., Fogassi, l.,
& Gallese, V. (2004). A touching sight: SII/PV activation
during the observation and experience of touch. Neurona,
42, 335–346.
Lombardo, METRO. v., Barnes, j. l., Wheelwright, S. J., &
Baron-Cohen, S. (2007). Self-referential cognition and
empathy in autism. Más uno, 2, e883.
Maddock, R. J., Garrett, A. S., & Buonocore, METRO. h. (2001).
Remembering familiar people: The posterior cingulate cortex
and autobiographical memory retrieval. Neurociencia, 104,
667–676.
1634
Revista de neurociencia cognitiva
Volumen 22, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
F
.
.
/
t
oh
norte
1
8
METRO
a
y
2
0
2
1
mitchell, j. PAG., Macrae, C. NORTE., & Banaji, METRO. R. (2006). Disociable
medial prefrontal contributions to judgments of similar
and dissimilar others. Neurona, 50, 655–663.
Morcom, A. METRO., & Fletcher, PAG. C. (2007). Does the brain have
a baseline? Why we should be resisting a rest. Neuroimagen,
37, 1073–1082.
Ochsner, k. NORTE., Beer, j. S., Robertson, mi. r., Cooper, j. C.,
Gabrieli, j. D., Kihsltrom, j. F., et al. (2005). el neural
correlates of direct and reflected self-knowledge.
Neuroimagen, 28, 797–814.
Pfeifer, j. h., Lieberman, METRO. D., & Dapretto, METRO. (2007).
“I know what you are but what am I?!": Neural bases of
self- and social knowledge retrieval in children and adults.
Revista de neurociencia cognitiva, 19, 1323–1337.
Pitcher, D., Garrido, l., Walsh, v., & duchaína, B. C. (2008).
Transcranial magnetic stimulation disrupts the perception
and embodiment of facial expressions. Diario de
Neurociencia, 28, 8929–8933.
Rizzolatti, GRAMO., & Craighero, l. (2004). The mirror-neuron
sistema. Annual Reviews of Neuroscience, 27, 169–192.
Ruby, PAG., & Decety, j. (2001). Effect of subjective perspective
taking during simulation of action: A PET investigation of
agency. Neurociencia de la naturaleza, 4, 546–550.
sajonia, r., & Kanwisher, norte. (2003). People thinking about
thinking people. The role of the temporo-parietal junction
in “theory of mind”. Neuroimagen, 19, 1835–1842.
sajonia, r., Moran, j. METRO., Scholz, J., & Gabrieli, j. (2006).
Overlapping and non-overlapping brain regions for
theory of mind and self reflection in individual subjects.
Social Cognitive and Affective Neuroscience, 1, 229–234.
sajonia, r., & Powell, l. j. (2006). Itʼs the thought that counts:
Specific brain regions for one component of theory of
mente. ciencia psicológica, 17, 692–699.
Cantante, T., Seymour, B., OʼDoherty, J., Kaube, h., Dolan, R. J.,
& Frith, C. D. (2004). Empathy for pain involves the
affective but not sensory components of pain. Ciencia, 303,
1157–1162.
Steiger, j. h. (1980). Tests for comparing elements of a
correlation matrix. Boletín Psicológico, 87, 245–251.
Uddin, l. P., Iacoboni, METRO., Lange, C., & Keenan, j. PAG. (2007).
The self and social cognition: The role of cortical midline
structures and mirror neurons. Tendencias en Ciencias Cognitivas,
11, 153–157.
Wicker, B., Keyser, C., Plailly, J., Royet, j. PAG., Gallese, v., &
Rizzolatti, GRAMO. (2003). Both of us disgusted in My insula: El
common neural basis of seeing and feeling disgust. Neurona,
40, 655–664.
Zaki, J., Ochsner, k. NORTE., Hanelin, J., Apostar, t. D., & Mackey,
S. C. (2007). Different circuits for different pain: Patrones
of functional connectivity reveal distinct networks for
processing pain in the self and others. Social Neuroscience,
2, 276–291.
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
7
mi
–
1
pag
6
d
2
F
3
/
1
2
9
2
3
/
9
7
1
/
7
1
6
6
oh
2
C
3
norte
/
1
2
0
7
0
6
9
9
9
2
8
1
8
2
8
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
8
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
F
.
.
t
/
oh
norte
1
8
METRO
a
y
2
0
2
1
Lombardo et al.
1635