Neural Correlates of Positive and Negative
Emotion Regulation
Sang Hee Kim and Stephan Hamann
Abstracto
& The ability to cope adaptively with emotional events by voli-
tionally altering one’s emotional reactions is important for psy-
interacción.
chological and physical health as well as social
Cognitive regulation of emotional responses to aversive events
engages prefrontal regions that modulate activity in emotion-
processing regions such as the amygdala. Sin embargo, the neural
correlates of the regulation of positive emotions remain largely
unexplored. We used event-related functional magnetic reso-
nance imaging to examine the neural correlates of cognitively
increasing and decreasing emotional reactions to positive and
negative stimuli. Participants viewed negative, positivo, and neu-
tral pictures while attempting to increase, decrease, or not alter
their emotional reactions. Subjective reactions were assessed
via on-line ratings. Consistent with previous studies, increasing
negative and positive emotion engaged primarily left-lateralized
prefrontal regions, whereas decreasing emotion activated bi-
lateral prefrontal regions. Different activations unique to in-
creasing versus decreasing emotion were observed for positive
and negative stimuli: Unique increase-related activations were
observed only for positive stimuli, whereas unique decrease-
related activations were observed only for negative stimuli.
Regulation also modulated activity in the amygdala, a key
emotion-processing region. Regulation effects on amygdala ac-
tivity were larger for positive than for negative stimuli, después-
tentially reflecting a greater malleability of positive emotional
reactions. Increasing and decreasing positive and negative emo-
tion can thus increase and decrease subjective reactions and
associated amygdala activity in line with regulatory goals, and is
associated with different patterns of prefrontal activation as a
function of emotional valence and regulatory goal. &
INTRODUCCIÓN
Individuals can cognitively regulate their emotional re-
sponses to events, increasing or decreasing their emo-
tional reactions in line with their behavioral goals. Este
process of regulating emotional responses by changing
the cognitive representation of events, often referred
to as reappraisal, is important for mental and physical
interacción. When successful,
health as well as social
emotion regulation allows us to adaptively cope with
aversive situations by minimizing negative, distressing
emotions, or alternatively, by maximizing the positive
aspects of situations (Bruto, 1998). Emotion regulation
can alter both psychological and physiological reactions
to emotional stimuli ( Jackson, Malmstadt, larson, &
Davidson, 2000). Impaired emotion regulation is asso-
ciated with affective disorders and a variety of other
maladaptive psychological conditions. Compared to
other types of emotion regulation strategies such as sup-
pression of behavioral expressions, reappraisal has been
proposed to be more effective because its influence be-
gins at an early stage of emotion generation, before emo-
tional reactions have fully unfolded (Richards & Bruto,
Emory University
2000). In support of this notion, recent studies of the
neural correlates of reappraisal have found that volun-
tary reappraisal can modulate the activity in the amyg-
dala, a subcortical structure which plays a critical role
in detecting and evaluating emotional significance of
estímulos (Ochsner et al., 2004; Ochsner, Bunge, Bruto,
& Gabrieli, 2002; Schaefer et al., 2002; Beauregard,
Levesque, & Bourgouin, 2001).
Although only a few studies have examined the neu-
ral bases of emotion regulation to date, a general pat-
tern has emerged in which prefrontal and anterior
cingulate regions involved in cognitive control show
increased activity during active attempts to regulate
emotion, together with modulation of activity in regions
involved in emotion processing such as the amygdala
(Ochsner & Bruto, 2005). Por ejemplo, in a functional
magnetic resonance imaging (resonancia magnética funcional) estudiar, Ochsner
et al. (2004) examined regions associated with reap-
praisal of negative scenes to decrease or increase their
emotional significance. Both up-regulation and down-
regulation of emotion was associated with increased
activity in the prefrontal cortex (PFC) and the anterior
cingulate, and emotion-related activity in the amygdala
increased or decreased in accordance with the regula-
tory goal.
D 2007 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 19:5, páginas. 776–798
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
.
.
.
t
.
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
These neuroimaging studies of emotion regulation
have focused on negative emotion. The neural corre-
lates of regulating positive emotion remain largely un-
conocido, sin embargo, with the exception of one study that
examined down-regulation of sexual arousal to erotic
in men (Beauregard et al., 2001). A pesar de
estímulos
regulation of emotion likely recruits common regions
regardless of the specific emotion involved, the neural
correlates of regulation for positive and negative emo-
tional stimuli may also differ in important respects.
To explore this issue, in the current study, we investi-
gated the neural correlates of regulating positive and
negative emotional reactions. Given the involvement
of the amygdala in responses to both positive and
negative emotionally arousing stimuli (Anderson et al.,
2003; Hamann, Ely, Hoffman, & Kilts, 2002; Dolan,
carril, Chua, & Fletcher, 2000; Hamann, Ely, Grafton, &
Kilts, 1999), we predicted that both positive and nega-
tive stimuli would elicit amygdala activity relative to
neutral stimuli, and that regulation-related changes in
emotional arousal would be reflected in increased ac-
tivity during successful attempts to increase emotion
and decreased activity when decreasing emotion. In ad-
condición, because activity in the ventral striatum has been
linked specifically to reward and appetitive processing
(Hamann & Mao, 2002; montesco & Berns, 2002), nosotros
also examined whether attempts to increase and de-
crease responses to positive emotional pictures would
result in corresponding increases and decreases in ven-
tral striatal activity.
En el estudio actual, we used event-related fMRI to
examine the neural correlates of cognitively increasing
and decreasing emotional reactions to affectively posi-
tive and negative stimuli. Participants viewed pleasant,
unpleasant, and neutral pictures while attempting to
increase, decrease, or not alter their emotional reac-
ciones. Subjective reactions were assessed on-line. We pre-
dicted that reappraisal processes involved in increasing
and decreasing emotion would recruit prefrontal re-
gions generally implicated in cognitive control, también
as regions involved specifically in increasing or decreas-
ing emotional responses. We hypothesized that emotion
regulation would alter activity in emotion-processing
regions such as the amygdala, in line with the regulatory
meta, with increased activity associated with successful
efforts to increase emotion and decreased activity asso-
ciated with decreasing emotion.
MÉTODOS
Participantes
Ten healthy right-handed female volunteers (ages 18–29,
m = 20.7) were recruited from the Emory University
community and monetarily compensated for their par-
ticipation. Written informed consent was obtained from
all participants prior to the study, and the study was
approved by the local human participants protection
committee.
Stimuli and Task
Three sets of 16 negative and 16 positive color pictures
and one set of 16 neutral color pictures were selected
from the International Affective Picture System (Lang,
Bradley, & Cuthbert, 1995) for the regulation task.
Negative pictures depicted a variety of aversive stimuli
(p.ej., traffic accidents, vermin, domestic violence, y
bodily injury). Positive pictures depicted a variety of
pleasant stimuli (p.ej., celebrations, domestic pets, sport-
ing events, and romantic couples). Each set of nega-
tive and positive pictures was matched on normative
ratings of arousal and valence (because valence is as-
sessed with a bipolar scale, the absolute value of the
mean difference from neutral valence was used for
matching positive and negative stimulus sets) (Lang
et al., 1995), and was assigned to either the increase,
decrease, or watch condition, with the assignment
counterbalanced across participants. Neutral pictures
were always assigned to the watch condition, as our
primary interest was in characterizing the effects of
emotion regulation on emotional stimuli, and prelimi-
nary work indicated that participants found it confusing
to attempt to increase or decrease emotional reactions
for stimuli that had little intrinsic emotional content.
An additional 18 pictures were selected for use in a
practice task to familiarize participants with the experi-
mental procedure prior to scanning.
In the regulation task, participants were instructed
to either increase or decrease their emotional reactions
to each picture. In the increase condition, Participantes
were instructed to think about the negative or positive
pictures in such a way that they felt the emotion elic-
ited by the presented picture more intensely. In the de-
crease condition, participants were instructed to think
about the negative or positive picture in such a way
that they felt the emotion elicited by the presented
picture less intensely. For the increase and decrease
condiciones, participants were specifically instructed not
to regulate their emotions by attempting to generate
an emotion opposite in valence to the one they would
normally experience (es decir., substituting a positive emo-
tion for a negative emotion). In the watch condition,
participants were instructed to view the picture in a
natural way and not to try to change the emotion elic-
ited by the picture. To assist participants in regulating
their emotions, example strategies identified by partic-
ipants from an earlier study (Jackson et al., 2000) y
from a behavioral pilot study were suggested. Estos
strategies included imagining the scenes as more per-
sonally relevant (p.ej., associating the main figure of
the scene to themselves or their close family members/
amigos) or less personally relevant (p.ej., dissociating
themselves from the main figures), imagining the scenes
Kim and Hamann
777
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
.
.
F
.
t
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
as unreal, and imagining the scenes as physically closer
or farther away from themselves. Participants were en-
couraged to use the strategies they found most effective,
although most participants used the suggested strate-
gies, as indicated by postscan interviews (see Results).
Procedimiento
Prior to scanning, participants received instructions on
the regulation task and performed a practice task
con 18 pictures depicting similar contents to those
presented during scanning. In the practice trials, a
regulation instruction (watch, decrease, increase) era
presented on a blank screen for 2 segundo, followed by a
picture for 8 segundo. Próximo, a Likert-type rating scale ranging
de 1 (weak) a 4 (strong) was presented, and partic-
ipants were asked to rate the strength of the emotion
they were currently feeling. Participants were asked to
verbally report the strategies they used while performing
the regulation task. To help insure that only participants
who could successfully regulate their emotions were
scanned, participants were required to pass a criterion
such that their mean subjective emotion ratings for the
decrease condition were less than 3 and their mean
ratings for the increase condition were greater than 2. Todo
participants met these inclusion criteria and could per-
form the regulation task as instructed.
In the scanner, participants again completed six prac-
tice trials with a subset of the same pictures that they
had practiced outside of the scanner to ensure that they
were comfortable with performing the task inside the
scanner. Each instruction appeared on the screen for
2 segundo (Cifra 1), followed by a picture for 8 segundo. Follow-
ing the picture, the 1–4 rating scale was presented for
4 sec and participants were asked to rate the strength
of the emotion they were currently feeling by pressing
a button on an MRI-compatible four-button fiber-optic
response box. Próximo, a fixation cross in a blank screen was
presented for 2 sec while participants were instructed
to rest. Images were rear-projected onto a projection
screen positioned at the head end of the MRI scanner
Cifra 1. Design of
experimental trials.
bore, controlled by a PC laptop computer using the
Presentation software program (versión 0.50, Neurobe-
havioral Systems, 2002; http://nbs.neuro-bs.com/). Par-
ticipants viewed the screen through a mirror mounted
on the head coil. Once the practice trials were complete,
the experimental task began.
Un total de 112 trials were completed over four separate
carreras. Each run consisted of four repetitions of seven
condiciones (es decir., decrease negative, watch negative, en-
crease negative, decrease positive, watch positive,
en-
crease positive, and watch neutral). The conditions were
presented in pseudorandom order in each run such that
no more than two identical regulation conditions nor
conditions with the same emotional valence (positivo,
negative, or neutral) were presented consecutively. No
picture stimuli were repeated during scanning. El
order of conditions was counterbalanced across runs.
After scanning had completed, participants exited the
scanner and were asked to write down brief descriptions
of the typical strategies they used in each regulation
condition and to provide a brief example of how each
strategy was used. Participants also indicated how suc-
cessful they believed they were in accomplishing each
regulation task, separately for negative and positive pic-
turas. They were given a 5-point Likert-type scale and
were asked to indicate their degree of success at car-
rying out each regulation task from 1 a 5 (1 = not
successful at all; 3 = moderately successful; 5 = very
successful).
Participants returned after 1 week and viewed all pic-
tures they had seen previously in the scanner and rated
them on a 7-point Likert-type scale (1 = not arousing;
4 = moderately arousing; 7 = highly arousing) to in-
dicate the level of arousal elicited by each picture in the
absence of active regulation attempts. For this rating
tarea, two pseudorandomly ordered picture lists were
created and administered in a counterbalanced manner
across participants. Próximo, participants were debriefed
and encouraged to ask any questions they had about
el estudio. Finalmente, participants were thanked for their
participation and excused.
778
Revista de neurociencia cognitiva
Volumen 19, Número 5
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
F
.
.
t
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Data Acquisition
All imaging data were acquired using a Siemens 3.0-Tesla
MRI scanner. Brain imaging involved acquisition of 30
axial slices of 3 mm thickness acquired parallel to the
AC–PC line. Functional scans were acquired using T2*-
weighted gradient-echo, echo-planar pulse sequences
(TR = 2516 mseg, TE = 30 mseg, 64 (cid:1) 64 matrix, 3 (cid:1)
3 (cid:1) 3 mm voxel size). Un total de 185 scans were acquired
in each of four runs. Structural images were acquired
using a gradient-echo, T1-weighted pulse sequence
(TR = 500 mseg, TE = 20 mseg, 256 (cid:1) 256 matrix, 1 (cid:1)
1 (cid:1) 1 mm voxel size).
Análisis de los datos
Data were analyzed using Statistical Parametric Mapping
software (SPM99, Wellcome Department of Cognitive
Neurología; www.fil.ion.ucl.ac.uk; Friston et al., 1995).
Funcional
images were realigned and spatially nor-
malized (voxel size 3 (cid:1) 3 (cid:1) 3 mm) to the Montreal
Neurological Institute (MNI) template (Ashburner &
Friston, 1999). Normalization parameters were gener-
ated from the mean realigned EPI image. Images were
smoothed using a 6-mm Gaussian kernel. Low-frequency
noise was removed using a high-pass filter (holmes,
Josephs, Buchel, & Friston, 1997). Individual partici-
pants’ data were analyzed using a fixed-effects model
(Friston, Jezzard, & Tornero, 2004). Group data were
analyzed using a random-effects model (holmes &
Friston, 1998).
Condition effects were modeled using a box-car regres-
sor convolved with a canonical hemodynamic response
function corresponding to the picture presentation inter-
vale (8 segundo). To ensure that regions of lower signal were
not excluded from statistical analysis, the more inclusive
threshold for including gray matter used in SPM2 was used
rather than the stricter SPM99 criterion. Inspection of
regions of signal dropout in individual subjects confirmed
that signal dropout was minimal near the amygdala, Alabama-
though signal attenuation was present in some parts of
the orbito-frontal cortex (OFC), a common occurrence
particularly at high field strengths. Voxel values for each
contrast yielded a statistical parametric map of the
t statistic, subsequently transformed to the unit normal
distribución. For the group analysis, one-sample t tests
were conducted on participants’ contrast images to create
statistical maps depicting differences in brain activation
between conditions across participants.
The amygdala regions of interest (ROI) were spheri-
cally defined using an automated algorithm (SPM ROI
Toolbox; http://spm-toolbox.sourceforge.net) with cen-
tral coordinates at x = ±18, y = (cid:2)3, z = (cid:2)18 en
Instituto Neurológico de Montreal (MNI) espacio, an ap-
proximation of Talairach space (Talairach & Tournoux,
1988), and with a radius of 8 mm. Para cada participante,
the average percent signal change across the peristimu-
lus time interval for each trial type was estimated. El
peak responses for each condition, defined as the peri-
stimulus latency between 5.03 y 12.58 segundo (corre-
sponding to TRs 3–5 of the peristimulus interval),
where the group average was maximal, were selected
for statistical analysis. Planned t tests (una cola, given
the directional nature of the hypothesized effects) eran
subsequently performed for statistical
inferencia. Ser-
cause this ROI analysis averaged over all voxels in the
amygdala ROIs, it was possible that this analysis might
fail to detect portions of the amygdala exhibiting oppo-
site responses (p.ej., increasing and decreasing activity
in separate subregions). To address this possibility, nosotros
conducted a follow-up small-volume-corrected voxel-
wise analysis (SVC, pag < .05, corrected) using the center
coordinates and radius used in defining the amygdala
ROIs, to probe the amygdala for activated voxels clusters
that varied as a function of emotion regulation.
To investigate modulation of activity in the ventral
striatum during regulation of positive emotion, left and
right ventral striatum ROIs were spherically defined
using the same methods used for the amygdala ROIs,
with central coordinates located at x = ±15, y = 12,
z = (cid:2)6. ROI analyses were conducted using the same
methods used for the amygdala ROIs.
A priori regions were defined on the basis of previous
neuroimaging studies on emotion regulation (Ochsner
et al., 2002, 2004; Beauregard et al., 2001). These
a priori regions included the PFC, the anterior cingulate,
and the amygdala. For group contrasts and regression
analysis, a threshold of p < .005 was applied for a priori
regions, p < .001 for all other regions, with an ex-
tent threshold of 5 contiguous voxels. Coordinates of
activated regions are reported in MNI space, an ap-
proximation of Talairach space (Talairach & Tournoux,
1988).
RESULTS
Behavioral Results
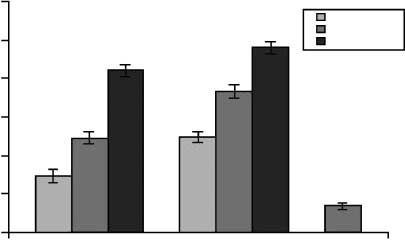
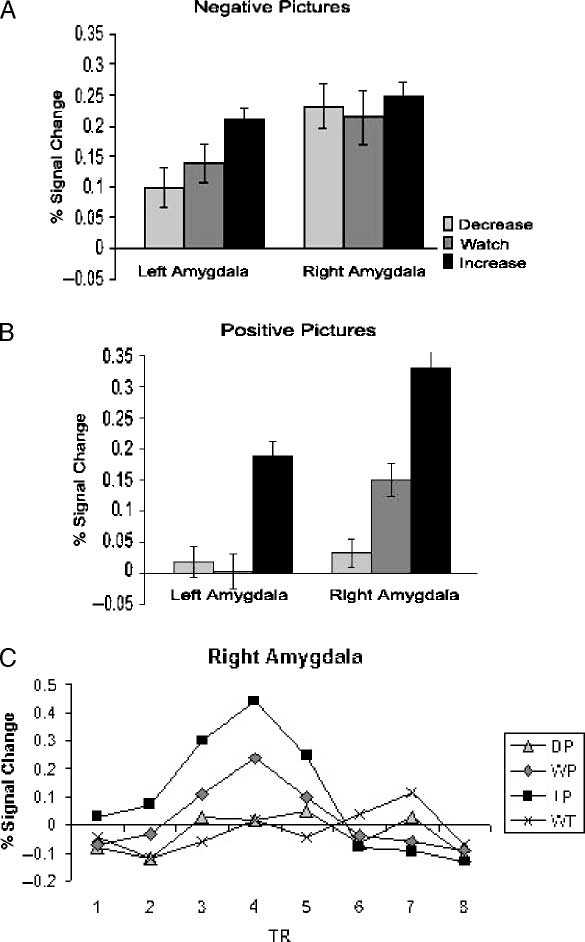
Self-ratings of Emotional Arousal
A 2 (emotion: positive, negative) (cid:1) 3 (regulation: de-
increase) repeated-measures analysis of
crease, watch,
variance (ANOVA) on on-line ratings for emotional arous-
al was conducted. Significant main effects of emotion
[F(1, 9) = 27.10, p < .001] and regulation [F(2, 18) =
65.07, p < .000] were found. No significant interaction
was found. Consistent with the predicted effect of regu-
lation, participants reported experiencing greater arousal
during the increase condition than the watch condition
[t(9) = 6.45, p < .000], and lower arousal during the
decrease condition than the watch condition [t(9) = 5.78,
p < .001]. Although negative and positive pictures had
been matched on normative ratings of arousal and va-
lence, participants reported greater arousal while viewing
negative pictures than positive pictures (Figure 2).
Kim and Hamann
779
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
.
.
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Strategy Use and Self-ratings of Regulation Success
Postscan ratings of how successful the participants be-
lieved they were in carrying out the instructions asso-
ciated with each regulation condition were compared
in a 2 (emotion: positive, negative) (cid:1) 3 (regulation:
increase) repeated-measures ANOVA
decrease, watch,
(Table 2). There was a significant main effect of regula-
tion [F(2,18) = 6.11, p < .01]. Overall, participants rated
their performance in the decrease condition as less suc-
cessful than in the control (watch) condition [t(9) =
2.37, p < .05], and rated their performance in the in-
crease condition as more successful than in the decrease
condition [t(9) = 3.35, p < .01]. Success ratings for the
watch condition reflected success in refraining from
active regulation and experiencing natural emotional
reactions to the stimuli. The effect of emotion on reg-
ulation success was marginally significant, with partic-
ipants rating regulating positive emotion as being easier
than regulating negative emotion [F(1, 9) = 1.10, p <
.09]. No interaction was found between emotion and
regulation type.
To determine whether participants’ success ratings cor-
related with their on-line arousal ratings, we conducted
correlation analyses between post hoc regulation success
ratings and mean on-line arousal ratings in each condi-
tion. Participants who reported being more successful
in decreasing negative emotion rated stimulus-elicited
arousal
lower during the decrease-negative condition
(r = (cid:2).552, two-tailed, p = .098), and participants who
reported being more successful
in increasing negative
emotion reported higher arousal during the increase-
negative condition (r = .695, p = .026). Correlations in
other conditions were not significant (all p values >.3).
We also assessed whether regulation success as indexed
by differences in on-line arousal ratings between each reg-
ulation condition and the watch condition (es decir., increase
positivo (cid:2) watch positive) correlated with corresponding
self-reported success ratings (es decir., self-reported success in
increasing positive emotion). No significant correlations
were found (all p values >.3).
Próximo, we examined the possible role of differences
in the specific regulation strategies used for regulating
positive and negative emotion. For increasing emotional
Positive
Negative
Neutral
Cifra 2. Mean on-line ratings of emotional arousal during each
regulation task for positive, negative, and neutral pictures.
To confirm that subjective arousal differed across stimu-
lus types when participants were not actively attempting
to regulate their emotional responses, an ANOVA was
conducted on on-line arousal ratings for positive, nega-
tivo, and neutral pictures that had been presented
during the watch condition. A main effect of emotion
was observed [F(2, 18) = 59.36, pag < .000]. Follow-up
comparisons confirmed that positive pictures [t(9) =
5.83, p < .000] and negative pictures [t(9) = 13.32, p <
.000] were rated higher than neutral pictures on arousal.
Moreover, negative pictures were rated higher than pos-
itive pictures on arousal [t(1,9) = 4.15, p < .002], al-
though they had been matched pre-experimentally on
normative arousal (Lang et al., 1995).
An ANOVA conducted on arousal ratings assessed
1 week later for all of the pictures used during the
scanning verified that positive and negative pictures were
more arousing than neutral pictures (Table 1). A main
effect of emotion was found [F(2, 18) = 57.48, p < .000].
Further comparisons confirmed that positive pictures
[t(9) = 6.22, p < .000] and negative pictures [t(9) =
10.36, p < .000] were rated higher in arousal than in
neutral pictures. Consistent with on-line ratings assessed
during scanning, negative pictures were rated higher in
arousal than positive pictures [t(9) = 3.91, p < .003].
Table 1. Postscanning Arousal Ratings for Positive, Negative,
and Neutral Stimuli
Decrease
Watch
Increase
Table 2. Self-ratings of Regulation Success across
Regulation Conditions
Mean
SEM
Mean
SEM
Mean
SEM
Decrease
Watch
Increase
Positive
Negative
Neutral
4.10
5.06
0.40
0.21
4.07
4.88
2.03
0.32
0.27
0.16
4.20
5.03
0.37
0.27
Mean
SEM
Mean
SEM
Mean
SEM
Positive
Negative
3.7
3.2
0.21
0.13
4.2
3.7
0.25
0.37
4.3
4.5
0.30
0.30
Arousal ratings were made on a scale that ranged from 1 (not arous-
ing) to 7 (highly arousing). SEM = standard error of the mean. For
positive and negative stimuli, assignment of stimulus sets was counter-
balanced across conditions across participants.
Ratings were made on a scale that ranged from 1 (not successful at all )
to 5 (very successful). Success ratings for the watch condition reflected
success in refraining from active regulation and experiencing natural
emotional reactions to stimuli. SEM = standard error of the mean.
780
Journal of Cognitive Neuroscience
Volume 19, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
f
.
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
reactions, all participants used the suggested strategy
of increasing personal relevance to increase both posi-
tive and negative emotional reactions. For decreasing
emotional reactions to negative pictures, all but one
participant reported pretending that the scene was
unreal. For decreasing emotional reactions to positive
pictures, four participants reported that they used the
‘‘pretend unreal’’ strategy and four participants attempt-
ed to focus on less positive elements of the scene. Two
participants did not specify their strategies in this con-
dition. To further investigate whether these two differ-
ent types of strategy employed for decreasing positive
emotion resulted in different outcomes in on-line arous-
al ratings and post hoc success ratings, we compared
the two strategy groups on on-line arousal ratings and
post hoc success ratings. There were no differences
in on-line arousal ratings [‘‘pretend unreal’’ M = 1.98;
‘‘less positive elements’’ M = 1.70, t(6) = 0.60, p >
.5] and post hoc success ratings [‘‘pretend unreal’’ M =
3.50; ‘‘less positive elements’’ M = 3.50, t(6) = 0] ser-
tween these two strategies, suggesting that both re-
sulted in similar levels of regulation efficacy as indexed
by these measures.
Brain Imaging Results
Brain Regions Activated by Voluntarily Increasing
and Decreasing Emotion
Activation by decreasing emotion. Brain regions asso-
ciated with decreasing negative and positive emotion
were identified by comparing activations in the decrease
condition and the watch condition (es decir., the decrease (cid:2)
watch contrast), separately for positive and negative pic-
turas. Based on previous neuroimaging studies on emo-
tion regulation (Ochsner et al., 2002, 2004; Beauregard
et al., 2001), a priori regions were defined, incluido
the PFC, the anterior cingulate cortex, and the amygdala.
A threshold of p < .005 was applied for a priori regions,
p < .001 for all other regions. For correlations between
brain activations and behavioral variables, a threshold
of p < .05 was used. In addition, an extent threshold
of five contiguous voxels was used for all whole-brain
analyses. All activation images are displayed in neuro-
logical format overlaid on a structural image normalized
to MNI space from a single subject from the SPM99
standard image library (www.fil.ion.ucl.ac.uk/spm), with
the left hemisphere on the left side of the image.
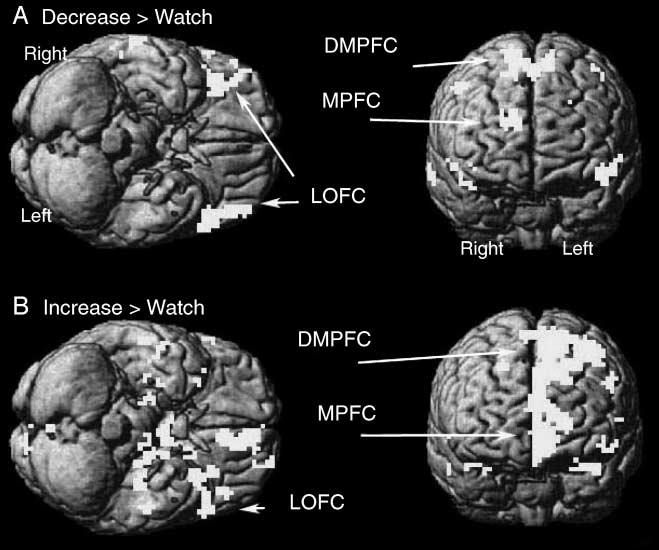
Positive pictures. Brain regions showing greater activa-
tion in the decrease condition than in the watch condi-
tion included the right LPFC (BA 9, 46), DMPFC (BA 6),
MPFC (BA 10), and bilateral LOFC (BA 47) (Table 4,
Figure 4).
Differences between decreasing negative and decreas-
ing positive emotion. We directly compared brain ac-
tivity associated with decreasing emotional reactions
to negative and positive pictures. Because participants
rated the negative pictures higher in mean arousal than
the positive pictures, we controlled for this factor by
including mean arousal ratings as a covariate in the
random effects analyses. To identify brain regions more
responsible for decreasing negative emotion than for
decreasing positive emotion, brain activity associated
with decreasing negative emotion (vs. the watch nega-
tive condition) was contrasted with brain activity associ-
ated with decreasing positive emotion (vs. the watch
positive condition) [i.e., (decrease negative (cid:2) watch
negative) (cid:2) (decrease positive (cid:2) watch positive)]. The
bilateral LPFC (BA 46, 10), DMPFC (BA 8/32), bilateral
MOFC (BA 11), right LOFC (BA 47), and bilateral ante-
rior cingulate were activated in this contrast (Table 5).
When post hoc success ratings were entered as a co-
variate instead of arousal ratings, the activation was the
same as for the analysis with arousal ratings as a co-
variate. When no covariates were entered, the pattern
was highly similar, with the exception of the activation
in the left OFC (BA 11), which was eliminated.
The reverse contrast [i.e., (decrease positive (cid:2) watch
positive) (cid:2) (decrease negative (cid:2) watch negative)] was
also conducted to identify brain regions more responsi-
ble for decreasing positive emotion than negative emo-
tion, with arousal ratings entered as a covariate. No areas
were identified that showed greater activation for de-
creasing positive emotion than decreasing negative
emotion; the same result was found when post hoc
success ratings were entered and when no covariates
were entered.
Activation by increasing emotion. Brain regions asso-
ciated with increasing negative and positive emotion
were identified by greater activation during the in-
crease condition than the watch condition (i.e., the
increase (cid:2) watch contrast), separately for positive and
negative pictures.
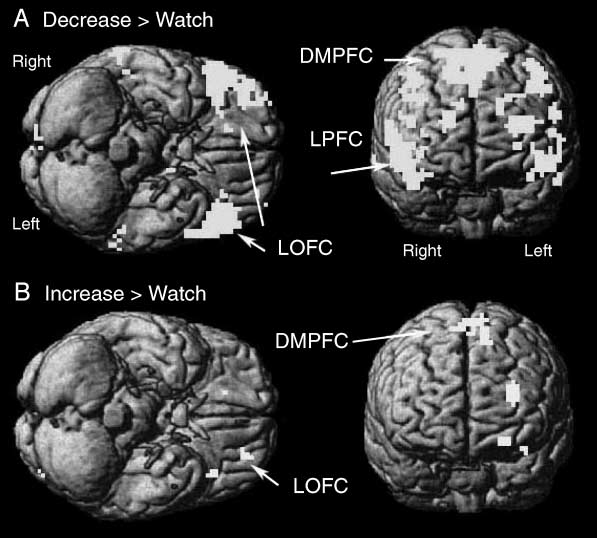
Negative pictures. Brain regions showing greater acti-
vation in the decrease condition than in the watch con-
dition included the bilateral lateral PFC (LPFC, BA 9, 10,
45), dorsomedial PFC (DMPFC, BA 6/32), medial PFC
(MPFC, BA 9/10), bilateral lateral OFC (LOFC, BA 47),
and anterior cingulate (BA 24/32) (Table 3, Figure 3). In
addition, the middle temporal gyrus and the pallidum
were also activated.
Negative pictures. Brain regions showing greater ac-
tivation in the increase condition than in the watch
condition included the left LPFC (BA 46/10), DMPFC
(BA 8), left LOFC (BA 47), left MOFC (BA 11), and the
anterior cingulate (BA 24/32) (Table 6, Figure 3). Ad-
ditional activations were observed in the lingual gyrus
and caudate.
Kim and Hamann
781
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
t
.
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Table 3. Brain Areas Activated for the Decrease > Watch Contrast for Negative Pictures
Coordinates (MNI )
HEM
BA
inf. Orbito-frontal G.
inf. Orbito-frontal G.
Sup. Temporal pole
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
inf. Frontal G.
inf. Orbito-frontal G.
inf. Orbito-frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
Sup. Med. Frontal G.
pre-SMA
Sup. Med. Frontal G.
Sup. Med. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Ant. Cingulate
Ant. Cingulate
Ant. Cingulate
Mid. Temporal G.
Mid. Temporal G.
Mid. Temporal G.
Pallidum
l
l
l
l
l
l
l
l
l
R
R
R
R
R
R
R
l
l
l
l
R
R
l
l
l
l
R
l
l
l
l
R
l
l
l
l
R
l
47
47
38
9
46
46
10
46
45
44
46
9
10
45
47
47
44
44
45
32
6
8
9
10
10
9
9
10
10
10
9
9
10
24
32
32
22
21
21
X
(cid:2)42
(cid:2)48
(cid:2)42
(cid:2)45
(cid:2)39
(cid:2)27
(cid:2)27
(cid:2)36
(cid:2)42
36
39
42
36
57
48
60
(cid:2)54
(cid:2)54
(cid:2)51
0
0
(cid:2)3
12
18
(cid:2)18
(cid:2)24
(cid:2)15
(cid:2)27
18
(cid:2)18
(cid:2)24
(cid:2)15
(cid:2)27
9
(cid:2)6
(cid:2)12
(cid:2)57
(cid:2)57
60
(cid:2)15
y
36
30
21
15
21
21
54
48
39
9
21
12
60
27
30
27
21
18
39
30
12
30
60
63
57
51
48
54
63
57
51
48
54
21
18
27
(cid:2)45
(cid:2)30
(cid:2)39
0
z
(cid:2)12
(cid:2)9
(cid:2)15
39
42
33
12
15
18
39
45
48
9
9
(cid:2)12
0
30
21
12
39
63
51
30
21
27
36
33
3
21
27
36
33
3
21
27
24
3
(cid:2)3
(cid:2)3
0
k (volumen)
290
(LM )
(LM )
161
(LM )
(LM )
108
(LM )
(LM )
169
(LM )
(LM )
5
364
(LM )
(LM )
20
(LM )
5
570
(LM )
(LM )
6
18
12
9
(LM )
5
18
12
9
(LM )
5
39
11
10
143
(LM )
104
24
z
5.23
4.17
3.63
4.72
3.94
3.41
4.69
3.84
2.81
4.35
4.09
4.04
3.12
4.42
4.25
4.12
3.02
3.00
2.67
4.39
4.24
3.97
3.32
3.51
3.26
3.07
2.87
2.84
3.51
3.26
3.07
2.87
2.84
4.50
2.94
2.82
4.98
4.05
3.70
3.69
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions, uncorrected, son
reported. Local maxima for these clusters are denoted with (LM ). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right; k = volume in voxel units; Z =
maximal Z score for contrast; inf. = inferior; Sup. = superior; Mid. = middle; Med. = medial; Ant. = anterior; GRAMO. = gyrus; SMA = supplementary motor area.
782
Revista de neurociencia cognitiva
Volumen 19, Número 5
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
.
.
.
.
F
t
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Cifra 3. Activated brain
regions for the contrasts of
(A) decrease > watch and
(B) increase > watch for
negative pictures.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
.
.
F
.
t
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 4. Brain Areas Activated for the Decrease > Watch Contrast for Positive Pictures
Coordinates
HEM
BA
inf. Orbito-frontal G.
Sup. Temporal Pole
inf. Orbito-frontal G.
inf. Orbito-frontal G.
inf. Orbito-frontal G.
inf. Orbito-frontal G.
inf. Orbito-frontal G.
inf. Orbito-frontal G.
Sup. Frontal G./Pre-SMA
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Sup. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
inf. Frontal G.
l
l
l
R
R
R
R
R
R
R
R
R
R
R
l
l
R
R
47
47
47
47
47
47
47
47
6
8
8
10
10
9
9
9
46
47
X
(cid:2)57
(cid:2)51
(cid:2)45
39
48
33
54
42
9
15
21
15
21
51
(cid:2)18
(cid:2)24
45
60
y
21
21
39
36
21
27
42
24
15
12
15
66
63
12
30
33
42
24
z
(cid:2)3
(cid:2)12
(cid:2)12
(cid:2)9
(cid:2)9
(cid:2)12
(cid:2)9
(cid:2)21
63
69
54
21
30
45
30
36
(cid:2)21
0
k (volumen)
97
(LM)
(LM)
50
(LM)
(LM)
6
5
149
(LM)
(LM)
29
(LM)
22
7
(LM)
7
7
z
4.00
3.45
3.44
3.55
3.41
3.38
3.28
3.20
3.74
3.50
3.32
3.31
2.74
3.74
3.05
2.59
3.02
3.62
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions, y-
corregido, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right; k =
volume in voxel units; Z = maximal Z score for contrast; inf. = inferior; Sup. = superior; Mid. = middle; GRAMO. = gyrus; SMA = supplementary motor area.
Kim and Hamann
783
Cifra 4. Activated brain
regions for the contrast of
(A) decrease > watch and
(B) increase > watch for
positive pictures.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
.
.
t
.
.
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 5. Brain Areas More Active for Decreasing Negative Emotion than for Decreasing Positive Emotion, Controlling
for On-line Arousal Ratings
Coordinates (MNI)
HEM
BA
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Sup. Frontal G.
Sup. Med. Frontal G.
Sup. Med. Frontal G.
Mid. Orbito-frontal G.
Sup. Orbito-frontal G.
Mid. Orbito-frontal G.
inf. Orbito-frontal G.
Ant. Cingulate
Ant. Cingulate
Ant. Cingulate
Ant. Cingulate
R
R
l
R
R
l
l
l
R
R
R
l
R
l
47
46
46
10
8./32
32
11
11
11
47
24
24
24
24
X
36
42
(cid:2)36
30
6
(cid:2)3
(cid:2)3
(cid:2)21
3
48
9
(cid:2)3
6
(cid:2)6
y
45
57
57
63
27
21
36
39
48
30
24
21
33
30
z
6
9
15
12
45
39
(cid:2)12
(cid:2)18
(cid:2)6
(cid:2)9
24
24
18
18
k (volumen)
49
(LM)
8
13
14
(LM)
14
5
7
12
41
(LM)
(LM)
5
z
4.46
3.63
3.05
2.97
2.89
2.8
3.62
3.53
3.52
3.48
4.11
3.48
2.88
3.05
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other re-
gions, uncorrected, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R =
bien; k = volume in voxel units; Z = maximal Z score for contrast; inf. = inferior; Sup. = superior; Mid. = middle; Med. = medial; Ant. = anterior;
GRAMO. = gyrus.
784
Revista de neurociencia cognitiva
Volumen 19, Número 5
Mesa 6. Brain Areas Activated for the Increase > Watch Contrast for Negative Pictures
Coordinates (MNI)
Sup. Frontal G./Pre-SMA
Sup. Med. Frontal G.
Sup. Frontal G./Pre-SMA
inf. Frontal G.
Mid. Frontal G.
inf. Orbito-frontal G.
Sup. Orbito-frontal G.
Ant. Cingulate
Ant. Cingulate
Mid. Cingulate
Ant. Cingulate
Lingual G.
Caudate
Caudate
Caudate
BA
8
8
6./8
13
46./10
47
11
32
24
24
32
19
HEM
R
l
l
l
l
l
l
l
l
R
R
R
l
l
X
3
(cid:2)9
(cid:2)3
(cid:2)39
(cid:2)30
(cid:2)36
(cid:2)21
(cid:2)15
(cid:2)3
0
15
30
21
(cid:2)18
(cid:2)9
y
24
27
18
15
51
27
48
24
33
18
33
(cid:2)54
9
9
0
z
66
57
69
12
18
(cid:2)18
(cid:2)15
36
21
39
(cid:2)6
0
18
21
24
k (volumen)
93
(LM)
(LM)
5
27
15
8
47
(LM)
(LM)
6
10
6
20
(LM)
z
4.49
3.69
3.69
3.37
3.28
3.14
3.09
3.84
3.27
2.91
3.08
4.30
4.12
3.82
3.72
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions,
uncorrected, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right;
k = volume in voxel units; Z = maximal Z score for contrast; inf. = inferior; Sup. = superior; Mid. = middle; Med. = medial; Ant. = anterior; GRAMO. =
gyrus; SMA = supplementary motor area.
Positive pictures. Brain regions showing greater activa-
tion in the increase condition than in the watch condi-
tion included the left LPFC (BA 8), DMPFC (BA 6), MPFC
(BA 10), MOFC (BA 11), and left LOFC (BA 47) (Mesa 7,
Cifra 4). Additional activations were observed in the
middle temporal gyrus, primary visual areas, the cau-
fecha, the putamen, and the pallidum.
Differences between increasing negative and increasing
positive emotion. Próximo, we compared brain activity as-
sociated with increasing emotional reactions to negative
and positive pictures. Específicamente, brain activity associated
with increasing negative emotion was contrasted with
brain activity associated with increasing positive emotion
[es decir., (increase negative (cid:2) watch negative) (cid:2) (increase
positivo (cid:2) watch positive)], controlling for differences in
on-line arousal ratings. Only one small cluster in the supe-
rior frontal gyrus (DMPFC, BA 8) (size = 5 vóxeles, z = 3.32)
showed greater activation for increasing negative emotion
than increasing positive emotion. When post hoc success
ratings were controlled for instead of on-line arousal
ratings, the results showed the same areas of activation.
When no covariates were entered, the LPFC (BA 46) era
activated in addition to the DMPFC activation identified
in the analysis controlling for on-line arousal ratings.
To identify brain regions more responsible for increas-
ing positive emotion than for increasing negative emo-
ción, brain activity associated with increasing positive
emotion was contrasted with brain activity associated
with increasing negative emotion [es decir., (increase positive
(cid:2) watch positive) (cid:2) (increase negative (cid:2) watch nega-
tivo)], controlling for differences in on-line arousal
left LPFC
ratings. The left rostromedial PFC (BA 10),
(BA 9), right LPFC (BA 45, 48), and the left amygdala
were activated (Mesa 8). When success ratings were
controlled instead of on-line arousal ratings, the anterior
cingulate (BA 32) and the precuneus (BA 30) eran
activated in addition to the regions noted above for
the analysis with on-line arousal as a covariate. When no
covariates were entered, the DMPFC (BA 6), the right
OFC (BA 11/47), the precuneus (BA 23, 30), the hippo-
campus, as well as left occipital and temporal regions,
were activated in addition to the regions noted above for
the analysis with on-line arousal as a covariate.
Similarities between regions involved in increasing
and decreasing emotion. Brain areas involved in both
increasing and decreasing emotion were characterized
by identifying regions that commonly activated for the
increase (cid:2) watch contrast and the decrease (cid:2) watch
Kim and Hamann
785
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
F
.
.
.
.
t
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 7. Brain Areas Activated for the Increase > Watch Contrast for Positive Pictures
Coordinates (MNI)
HEM
BA
8
6
32
10
32
9
47
47
38
47
9
44./45
45
47
47
21
21
21
18
Mid. Frontal G.
SMA
Sup. Frontal G.
Mid. Orbito-frontal G.
Ant. Cingulate
Sup. Med. Frontal G.
inf. Orbito-frontal G.
inf. Orbito-frontal G.
Sup. Temporal Pole
inf. Frontal G.
inf. Frontal G.
Mid. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Orbito-frontal G.
Mid. Temporal G.
Mid. Temporal G.
Mid. Temporal G.
Calcarine Sulcus
Amygdala
Hippocampus
Thalamus
Correo. Cingulate
Correo. Cingulate
Precuneus
Precuneus
Putamen
Putamen
Caudate
Caudate
Caudate
Caudate
Caudate
Putamen
Pallidum
Pallidum
Thalamus
Thalamus
l
l
l
l
l
l
l
l
R
l
l
l
l
l
R
R
R
R
R
l
l
l
l
l
l
l
l
l
l
l
l
l
l
R
R
l
l
X
(cid:2)27
(cid:2)9
(cid:2)15
(cid:2)3
0
(cid:2)3
(cid:2)30
(cid:2)42
(cid:2)39
60
(cid:2)51
(cid:2)45
(cid:2)54
(cid:2)39
(cid:2)42
48
45
54
6
18
(cid:2)30
(cid:2)6
(cid:2)6
(cid:2)6
(cid:2)9
(cid:2)6
(cid:2)21
(cid:2)18
(cid:2)6
(cid:2)15
(cid:2)12
(cid:2)15
(cid:2)15
(cid:2)24
18
18
(cid:2)6
(cid:2)9
y
24
9
27
48
51
57
15
24
21
24
18
24
21
30
36
(cid:2)45
(cid:2)36
(cid:2)6
(cid:2)87
3
(cid:2)18
(cid:2)6
(cid:2)48
(cid:2)48
(cid:2)57
(cid:2)60
15
6
12
(cid:2)12
9
0
9
9
12
12
(cid:2)6
(cid:2)27
z
48
60
39
(cid:2)6
15
39
(cid:2)21
(cid:2)9
(cid:2)24
(cid:2)3
33
33
9
(cid:2)3
(cid:2)9
3
6
(cid:2)18
9
(cid:2)18
(cid:2)18
0
30
30
24
51
3
(cid:2)9
15
(cid:2)18
24
18
12
12
6
15
0
18
k (volumen)
512
(LM)
(LM)
401
(LM)
(LM)
84
(LM)
(LM)
7
24
(LM)
5
15
(LM)
40
(LM)
43
32
9
946
(LM)
(LM)
69
(LM)
(LM)
24
(LM)
11
6
6
(LM)
5
(LM)
12
(LM)
15
6
z
4.75
3.91
3.74
4.38
3.76
3.76
4.00
3.42
3.34
3.28
3.07
2.67
3.06
3.02
2.94
4.23
3.42
3.88
4.04
3.44
4.39
4.10
4.09
4.09
3.77
3.51
3.91
3.32
3.80
3.48
3.30
3.26
3.20
3.15
3.51
3.38
4.10
3.50
786
Revista de neurociencia cognitiva
Volumen 19, Número 5
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
F
.
.
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 7. (continued )
HEM
BA
Cerebellum
Cerebellum
Cerebellum
Cerebellum
Cerebellum
Cerebellum
R
R
R
l
l
Coordinates (MNI)
X
6
0
27
18
(cid:2)6
(cid:2)3
y
(cid:2)81
(cid:2)78
(cid:2)84
(cid:2)84
(cid:2)36
(cid:2)36
z
(cid:2)18
(cid:2)12
(cid:2)24
(cid:2)21
(cid:2)6
(cid:2)6
k (volumen)
12
(LM)
6
(LM)
16
(LM)
z
4.34
3.29
3.49
3.25
3.77
3.75
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions,
uncorrected, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right; k =
volume in voxel units; Z = maximal Z score for contrast; inf. = inferior; Sup. = superior; Mid. = middle; Med. = medial; Ant. = anterior; Correo. =
posterior; GRAMO. = gyrus; SMA = supplementary motor area.
contrast, using inclusive masking procedures at the
group level (with a threshold level of .005, uncorrected).
Negative pictures. The DMPFC (BA 6, 8), the LPFC, el
anterior cingulate (BA 24/32), and the left OFC (BA 47)
were commonly activated for regulating negative emo-
ción (Mesa 9).
Positive pictures. The DMPFC (BA 6, 8) and the left
OFC (BA 47) were commonly activated for regulating
positive emotion (Mesa 9).
Differences between regions involved in increasing and
decreasing emotion. Brain areas more involved in in-
creasing emotion than in decreasing emotion were char-
acterized by identifying regions that showed greater
activation during the increase condition than the de-
crease condition (es decir., the increase (cid:2) decrease contrast),
within the regions that showed greater activation in the
increase (cid:2) watch contrast. Eso es, the increase (cid:2) de-
crease contrast was inclusively masked by the increase (cid:2)
watch contrast at the threshold level of p < .005, uncor-
rected. Conversely, brain areas more involved in decreas-
ing emotion than increasing emotion were identified by
greater activation during the decrease condition than the
increase condition, within the regions that showed great-
er activation in the decrease (cid:2) watch contrast. That is, the
decrease (cid:2) increase contrast was inclusively masked by
the decrease (cid:2) watch contrast at the threshold level of
p < .005, uncorrected.
Negative pictures. No areas were identified that ex-
hibited significantly greater activation during the in-
crease condition than the decrease condition for
negative pictures. Areas more activated for decreasing
than for increasing negative emotion included the right
Table 8. Brain Areas More Active for Increasing Positive Emotion than for Increasing Negative Emotion, Controlling
for On-line Arousal Ratings
Coordinates (MNI)
Sup. Frontal G.
Sup. Frontal G.
Mid. Frontal G.
Inf. Frontal G.
Inf. Frontal G.
Inf. Frontal G.
Sup. Med. Frontal G.
Amygdala
HEM
L
L
L
R
R
R
L
L
BA
10
10
9
45
45
48
9
x
(cid:2)24
(cid:2)12
(cid:2)33
54
54
39
(cid:2)6
(cid:2)27
y
63
60
15
33
39
15
54
0
z
6
12
54
24
18
21
39
(cid:2)21
k (volume)
41
(LM)
5
22
(LM)
5
8
5
Z
3.75
3.58
3.24
3.57
3.02
3.01
3.17
2.72
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions,
uncorrected, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right;
k = volume in voxel units; Z = maximal Z score for contrast; Inf. = inferior; Sup. = superior; Mid. = middle; Med. = medial; G. = gyrus.
Kim and Hamann
787
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
t
.
f
o
n
1
8
M
a
y
2
0
2
1
Table 9. Brain Areas Commonly Recruited for Both Increasing and Decreasing Emotion
Coordinates (MNI)
Negative pictures
Sup. Frontal G./Pre-SMA
Sup. Frontal G./Pre-SMA
Frontal Sup. Med. G.
Frontal Sup. Med. G.
Mid. Frontal G.
Ant. Cingulate
Ant. Cingulate
Inf. Orbito-frontal G.
Positive pictures
Sup. Frontal G./Pre-SMA
Sup. Med. Frontal G.
Sup. Frontal G./Pre-SMA
Inf. Orbito-frontal G.
Inf. Orbito-frontal G.
HEM
BA
R
R
L
L
L
L
L
L
L
L
L
L
8
6
8
8
46
24./32
47
6
8
6
47
47
x
3
(cid:2)3
(cid:2)9
(cid:2)9
(cid:2)30
(cid:2)9
0
(cid:2)36
(cid:2)9
(cid:2)9
(cid:2)9
(cid:2)42
(cid:2)51
y
24
6
27
21
51
24
21
27
12
24
15
24
24
z
k (volume)
Z
66
69
57
66
18
36
39
(cid:2)18
57
63
66
(cid:2)9
(cid:2)3
40
(LM)
16
(LM)
18
10
(LM)
8
29
(LM)
(LM)
23
(LM)
4.49
3.08
3.69
2.93
3.28
3.19
2.71
3.14
3.85
3.34
3.18
3.42
2.65
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions,
uncorrected, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right;
k = volume in voxel units; Z = maximal Z score for contrast; Inf. = inferior; Sup. = superior; Mid. = middle; Med. = medial; Ant. = anterior; G. =
gyrus; SMA = supplementary motor area.
LPFC, the MPFC, the bilateral LOFC, and the supra-
marginal gyrus (Table 10).
Positive pictures. Areas more activated for increasing
than for decreasing emotion included the anterior re-
gion of the MPFC, the MOFC, the anterior cingulate, the
precuneus, the caudate, and the thalamus. No areas
were more activated for decreasing than for increasing
positive emotion (Table 11).
These findings are broadly consistent with those of
the analyses described above that directly contrasted
regulation-related activations for positive versus negative
stimuli (Tables 5 and 8), particularly with respect to the
substantially greater number of brain regions involved in
down-regulation of negative emotion relative to positive
emotion, and the greater number of brain regions
involved in up-regulation of positive emotion relative
to negative emotion.
sentative peristimulus timecourse of amygdala activity is
shown in Figure 5C. For comparisons between ROI activ-
ity, planned one-tailed t tests were used in line with the
directional nature of the predicted effects. Consistent with
previous findings of greater amygdala activity for emotion-
ally arousing stimuli (Hamann et al., 1999, 2002; Dolan
et al., 2000), we found greater activity in both the left
[t(9) = 2.25, p < .03] and the right [t(9) = 2.46, p < .02]
amygdala in response to the negative pictures, and in the
right [t(9) = 2.5, p < .02] amygdala in response to the
positive pictures, compared to the neutral pictures.
Negative pictures. A t test on the means of the increase
and decrease conditions showed that the difference
between the two conditions was marginally significant
[t(9) = 1.6, p < .07; Figure 5A]. Differences between
each of these two conditions and the watch condition
were not statistically significant, however.
Effects of Emotion Regulation on Activity
in Emotion Processing
Amygdala ROI analysis. To determine whether emo-
tion regulation modulated activity in the amygdala elic-
ited by emotional pictures, fMRI signal change within
the left and right amygdala ROIs was analyzed. A repre-
Positive pictures. Activity in the left [t(9) = 4,2, p <
.001] and right [t(9) = 4.42, p < .001] amygdala ROIs
during the increase condition was greater than during
the watch condition, p < .001 for both comparisons.
Activity in the right amygdala ROI was also lower during
the decrease condition than during the watch condition
[t(9) = 3.9, p < .002; Figure 5B].
788
Journal of Cognitive Neuroscience
Volume 19, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
f
.
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Table 10. Brain Areas Differentially Recruited by Increasing or Decreasing Negative Emotion
Coordinates (MNI)
HEM
BA
x
y
z
k (volume)
Z
Increase > Decrease
No activations
Decrease > Increase
inf. Orbito-frontal G.
Mid. Orbito-frontal G.
Mid. Orbito-frontal G.
inf. Orbito-frontal G.
inf. Orbito-frontal G.
Frontal. Sup. Med. GRAMO.
Frontal. Sup. Med. GRAMO.
Frontal. Sup. Med. GRAMO.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Sup. Frontal G.
Frontal. Sup. Med. GRAMO.
Sup. Frontal G.
inf. Frontal G.
inf. Frontal G.
Supramarginal G.
Angular G.
inf. Parietal G.
Mid. Temporal G.
R
R
R
l
l
R
l
R
R
R
R
R
R
R
R
R
R
R
R
R
R
47
47
47
47
47
32./8
32
8
9
45
45
45
8
8
10
45./47
45
40
40
40
21
39
42
39
(cid:2)51
(cid:2)39
3
(cid:2)9
6
42
54
45
42
15
6
21
51
57
57
57
42
60
45
51
60
36
39
36
27
42
18
39
36
33
27
30
63
36
33
(cid:2)42
(cid:2)51
(cid:2)51
(cid:2)39
(cid:2)18
(cid:2)12
(cid:2)6
(cid:2)9
(cid:2)9
36
42
45
48
21
21
36
57
63
18
(cid:2)3
9
39
36
54
(cid:2)6
36
(LM)
(LM)
47
(LM)
37
(LM)
16
55
55
(LM)
(LM)
32
(LM)
9
26
5
17
(LM)
(LM)
8
4.52
4.40
3.88
3.79
3.17
4.5
3.09
4.31
4.08
3.85
3.73
3.70
3.96
3.20
3.24
3.56
2.92
4.04
3.89
3.56
3.71
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions,
uncorrected, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right;
k = volume in voxel units; Z = maximal Z score for contrast; inf. = inferior; Sup. = superior; Mid. = middle; Med. = medial; GRAMO. = gyrus.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
F
.
.
.
Voxelwise small-volume-corrected amygdala analysis.
To examine whether subregions within the left and right
amygdala ROIs exhibited different responses not de-
tected in the primary ROI analysis that averaged signal
change across all voxels in each ROI, we used an SVC
voxelwise analysis ( pag < .05, corrected for multiple com-
parisons) using the center coordinates and radius used
in defining the amygdala ROIs. The results of this analy-
sis matched the results of the primary ROI analysis.
No additional activation clusters with responses differing
from the primary ROI analysis were observed.
Correlations between arousal and amygdala activity.
To investigate whether changes in on-line arousal ratings
correlated with changes in amygdala activity, correlation
analyses were conducted between changes in arousal for
each regulation condition relative to the watch condi-
tion (e.g., increase (cid:2) watch) and changes in amygdala
activity for the corresponding conditions. On-line arous-
al ratings associated with increasing positive emotion
showed a marginal correlation with corresponding in-
creases in activity in the left amygdala ROI, r = .57,
p < .086. All other correlations were not statistically
significant ( ps > .15).
Ventral striatum ROI analysis. To investigate the ef-
fect of emotion regulation on the activity in the ventral
striatum during viewing of positive emotional pictures,
oh
norte
1
8
METRO
a
y
2
0
2
1
Kim and Hamann
789
Mesa 11. Brain Areas Differentially Recruited by Increasing versus Decreasing Positive Emotion
Coordinates (MNI)
HEM
BA
10
10
10
9
9
11
9
25
23
l
l
l
l
l
l
l
l
l
R
l
l
X
(cid:2)3
(cid:2)9
(cid:2)9
0
(cid:2)33
(cid:2)3
(cid:2)21
(cid:2)3
(cid:2)6
(cid:2)3
21
(cid:2)3
(cid:2)6
y
54
57
60
54
9
57
27
30
(cid:2)51
(cid:2)9
(cid:2)27
(cid:2)45
(cid:2)54
z
15
0
9
39
48
(cid:2)15
42
12
33
0
21
3
15
k (volumen)
z
189
(LM)
(LM)
10
5
5
33
6
17
11
6
21
(LM)
4.11
4
3.94
3.27
3.14
3.45
3.32
3.62
4.08
4.26
3.60
3.53
3.28
Increase > Decrease
Frontal Sup. Med. GRAMO.
Frontal Sup. Med. GRAMO.
Frontal Sup. Med. GRAMO.
Frontal Sup. Med G.
Mid. Frontal G.
Mid. Orbito-frontal G.
Sup. Frontal G.
Ant. Cingulate
Precuneus
Thalamus
Caudate
Vermis
Decrease > Increase
No activations
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions,
uncorrected, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right;
k = volume in voxel units; Z = maximal Z score for contrast; Sup. = superior; Mid. = middle; Med. = medial; Ant. = anterior; GRAMO. = Gyrus.
signal change within the left and right ventral striatum
ROIs was analyzed as a function of regulation condition.
The mean peak responses for each condition are sum-
marized in Table 12. Two-tailed t tests were conducted to
examine increase and decrease in activity of ventral stria-
tum due to up- and down-regulation of emotion.
Consistent with the role of the ventral striatum in posi-
tive emotion, activity in the left [t(9) = 3.86, pag < .004]
and right [t(9) = 3.00, p < .015] ventral striatum ROI
was greater when subjects attempted to up-regulate pos-
itive emotion, relative to the watch condition. Activity
in the left [t(9) = 3.09, p < .013] and right [t(9) = 2.52,
p < .033] ventral striatum ROI was also greater during
the increase condition than the decrease condition.
However, activity in the left and right ventral striatum
ROIs did not differ significantly between the decrease
and the watch conditions, ps > .3.
Correlation between arousal and whole-brain activity.
To investigate brain areas associated with changes in
emotional arousal during up- and down-regulation of
emotion, whole-brain correlation analyses were con-
ducted for each regulation contrast (p.ej., increase neg-
ative (cid:2) watch negative) with corresponding on-line
arousal changes.
Negative pictures. Brain areas whose activity was pos-
itively correlated with changes in on-line arousal ratings
due to decreasing negative emotion included the bilat-
eral DMPFC (BA 6, 8, 32), the MPFC (BA 10), la izquierda
LPFC (BA 44, 45), the right LOFC (BA 47), the bilateral
amygdala, the right SMG (BA 40/48), and the right
angular gyrus (Mesa 13). Sin embargo, no areas were
correlated with changes in on-line arousal ratings due
to increasing negative emotion.
Positive pictures. Activity in the right LOFC (BA 47)
(MNI coordinates at 51 30 (cid:2)9, z = 3.18, k = 5) was pos-
itively correlated with changes in on-line arousal ratings
due to decreasing positive emotion. Brain activity in the
areas including the left DMPFC (BA 6), the bilateral LPFC
(BA 44), and the right LOFC (BA47) (Mesa 14) presentado
positive correlation with changes in on-line arousal
ratings due to increasing positive emotion.
DISCUSIÓN
En el estudio actual, we investigated the neural correlates
of emotion regulation for negative and positive emotional
estímulos. Consistent with prior findings from studies ex-
790
Revista de neurociencia cognitiva
Volumen 19, Número 5
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
.
.
.
F
.
t
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Cifra 5. Activity in the left
and right amygdala ROIs
averaged across participants
during each regulation
condition for (A) negative
pictures and (B) positivo
pictures. Mean percent signal
change relative to the global
baseline mean for neutral
pictures was (cid:2)0.09 (SEM =
0.07) for the left amygdala
y (cid:2)0.03 (SEM = 0.04)
for the right amygdala. (C)
Representative peristimulus
time-series representing
percent signal change in the
right amygdala ROI averaged
across participants (DP =
decrease positive, WP =
watch positive, IP = increase
positivo, WT = watch neutral).
amining the regulation of negative emotion, activity in the
amygdala was modulated for emotional stimuli in line
with participants’ regulatory goals. Regulation-related
activity was also observed in several prefrontal and cingu-
late regions previously implicated in emotion regulation
Mesa 12. Mean and Standard Deviation of Activity in
the Left and Right Ventral Striatum ROIs as a Function
of Regulation Condition for Positive Emotion
Decrease
Watch
Increase
Significar
SEM
Significar
SEM
Significar
SEM
Left
Right
0.02
0.05
0.05
0.03
0.05
0.05
0.04
0.03
0.17
0.14
0.03
0.03
y control cognitivo (Ochsner et al., 2002, 2004). El
current results are consistent with appraisal theories of
emotion (Ochsner & Bruto, 2005; Lazarus, 1991; Frijda,
1986), with reappraisal processes mediated primarily by
prefrontal and cingulate regions which, Sucesivamente, modulate
regions involved in the representation of emotional
estados, including the amygdala. The inclusion of both
increase and decrease regulation instructions, together
with positive and negative emotional stimuli, allowed us
to compare the neural correlates of up-regulation and
down-regulation, and to assess whether these correlates
were similar for regulation of positive and negative affec-
tive responses.
On-line behavioral ratings of emotional arousal con-
firmed that participants were successful in using regula-
tion to alter their subjective emotional reactions, para
Kim and Hamann
791
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
.
t
.
.
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 13. Regional Activity Correlated with Changes in Emotional Arousal during Decreasing Negative Emotion, Relativo
to the Watch Negative Condition
Coordinates (MNI)
HEM
BA
Sup. Frontal G./SMA
Sup. Frontal G.
Sup. Med. Frontal G.
Sup. Frontal G./SMA
Sup. Frontal G./SMA
Sup. Med. Frontal G.
Sup. Med. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
Mid. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Frontal G.
inf. Orbito-frontal G.
inf. Orbito-frontal G.
Mid. Orbito-frontal G.
Precentral G.
Precentral G.
Precentral G.
Sup. Temporal Pole
Sup. Temporal Pole
Sup. Temporal Pole
Mid. Temporal G.
inf. Temporal G.
Parahippocampal G.
Sup. Parietal G.
R
R
l
l
l
l
R
R
l
l
l
R
R
R
l
l
l
R
R
R
l
R
R
R
R
R
l
l
l
R
R
l
l
R
R
R
32
32
8
6
6
10
10
6
6
8
8
8
47
44
48
45
15
45
45
45
45
48
48
44./45
47
47
11
6
6
6
38
38
38
48
37
30
7
X
9
18
(cid:2)3
(cid:2)6
(cid:2)9
(cid:2)6
0
36
36
(cid:2)27
(cid:2)27
(cid:2)27
48
57
39
(cid:2)51
(cid:2)48
(cid:2)42
51
60
54
(cid:2)33
39
45
33
30
33
(cid:2)33
(cid:2)42
(cid:2)42
42
36
(cid:2)27
(cid:2)45
39
27
24
y
21
12
30
3
15
66
63
6
9
24
18
9
33
18
27
33
42
33
36
24
27
30
12
12
30
27
45
(cid:2)3
(cid:2)3
(cid:2)3
15
9
6
(cid:2)12
(cid:2)51
(cid:2)33
(cid:2)63
z
45
45
51
66
57
15
21
54
42
51
42
63
(cid:2)9
15
15
12
12
6
18
21
12
12
18
24
(cid:2)21
(cid:2)9
(cid:2)15
36
39
48
(cid:2)24
(cid:2)24
(cid:2)24
(cid:2)21
(cid:2)12
(cid:2)15
60
k (volumen)
41
(LM)
(LM)
8
15
23
(LM)
28
(LM)
21
(LM)
5
33
9
14
37
(LM)
(LM)
18
13
(LM)
16
19
(LM)
53
(LM)
(LM)
29
(LM)
(LM)
9
(LM)
11
5
5
14
8
z
3.36
3.1
3.01
3.31
2.91
3.67
3.42
3.94
2.6
3.78
3.15
2.86
3.77
3.59
3.59
3.46
3.36
3.26
3.44
3.13
2.79
3.12
3.03
2.91
3.95
3.5
3.31
4.08
3.47
3.28
4.38
3.65
4.03
3.4
4.18
4.12
3.67
792
Revista de neurociencia cognitiva
Volumen 19, Número 5
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
oh
t
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
r
pag
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
metro
a
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
mi
a
–
r
pag
t
d
i
C
1
yo
9
mi
5
–
7
pag
7
d
6
F
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
oh
6
C
/
norte
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
oh
7
C
6
norte
pag
.
d
2
0
b
0
y
7
gramo
.
tu
1
mi
9
s
.
t
5
oh
.
norte
7
0
7
8
6
S
.
pag
mi
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
.
.
t
.
.
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 13. (continued )
Coordinates (MNI)
Supramarginal G.
Lingual G.
Angular G.
Cuneus
Calcarine
Sup. Occipital G.
Sup. Occipital G.
Hippocampus
Hippocampus
Fusiform G.
Insula
Insula
Insula
Caudate
BA
48./40
37
7
7
17
19
48./20
HEM
R
R
R
R
R
R
l
R
l
l
l
R
R
l
X
45
33
30
21
24
21
(cid:2)24
36
(cid:2)36
(cid:2)36
(cid:2)30
33
39
(cid:2)12
y
(cid:2)33
(cid:2)57
(cid:2)60
(cid:2)57
(cid:2)66
(cid:2)72
(cid:2)84
(cid:2)3
(cid:2)18
(cid:2)15
24
(cid:2)12
6
12
z
33
3
42
33
12
18
30
(cid:2)18
(cid:2)18
(cid:2)27
(cid:2)6
24
(cid:2)9
18
k (volumen)
94
14
22
8
31
(LM)
7
12
12
(LM)
5
7
14
6
z
4.46
4.32
4.23
4.06
4.06
3.47
3.28
3.67
3.58
3.53
3.53
3.47
3.42
3.37
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions,
uncorrected, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right;
k = volume in voxel units; Z = maximal Z score for contrast; inf. = inferior; Sup. = superior; Mid. = middle; Med. = medial; GRAMO. = gyrus; SMA =
supplementary motor area.
both positive and negative stimuli. Regulation success
ratings indicated that participants reported down-
regulation as more difficult than up-regulation, en línea
with similar results from a previous regulation study
(Ochsner et al., 2004),
in which it was noted that
reversing an emotional reaction is typically more dif-
ficult than amplifying an emotional reaction. A trend
( pag < .09) was found suggesting that regulation was
Table 14. Regional Activity Correlated with Changes in Emotional Arousal during Increasing Positive Emotion, Relative
to the Watch Positive Condition
Sup. Frontal G./SMA
SMA
Sup. Frontal G./SMA
Mid. Frontal G.
Mid. Frontal G.
Inf. Orbito-frontal G.
Inf. Orbito-frontal G.
Mid. Temporal G.
Mid. Temporal G.
Coordinates (MNI)
HEM
BA
L
L
L
R
L
R
R
R
R
6
6
6
44./6
44./6
47
47
21
21
x
(cid:2)9
(cid:2)15
(cid:2)12
42
(cid:2)27
39
36
69
63
y
3
(cid:2)3
(cid:2)6
9
6
33
39
(cid:2)27
(cid:2)27
z
51
48
75
39
36
(cid:2)15
(cid:2)9
(cid:2)6
0
k (volume)
8
(LM)
6
25
8
14
(LM)
5
(LM)
Z
3.51
3.46
3.24
4.27
3.21
3.39
2.80
3.34
3.23
Clusters of 5 contiguous voxels whose global maxima meet a p threshold of .005, for a priori regions, and p threshold of .001, for any other regions,
uncorrected, are reported. Local maxima for these clusters are denoted with (LM). BA = Brodmann’s area; HEM = hemisphere; L = left; R = right;
k = volume in voxel units; Z = maximal Z score for contrast; Inf. = inferior; Sup. = superior; Mid. = middle; G. = gyrus; SMA = supplementary
motor area.
Kim and Hamann
793
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
t
.
.
f
.
o
n
1
8
M
a
y
2
0
2
1
subjectively easier for positive emotion than for nega-
tive emotion. Although positive and negative pictures
had been matched on normative arousal ratings, nega-
tive pictures were, nevertheless, rated by participants
as being more arousing than positive pictures, possibly
contributing to greater difficulty in regulating reactions
to negative pictures. Alternately, reactions to positive
emotion stimuli may be intrinsically more malleable
than reactions to negative emotion stimuli,
indepen-
dent of differences in arousal. The finding of greater
regulation-related modulation in amygdala activity for
positive stimuli relative to negative stimuli is also con-
sistent with a greater malleability of positive emotional
reactions.
Similarities between Up-regulation
and Down-regulation
Increasing and decreasing negative emotion recruited
similar activations in the DMPFC, left LPFC, anterior
cingulate, and left OFC, consistent with previous results
(Ochsner et al., 2004). Both increasing and decreasing
negative emotion recruited regions in the anterior cingu-
late and the PFC that have been linked to emotion regu-
lation, working memory, and cognitive control. As with
negative emotion, regulating positive emotion also re-
cruited prefrontal regions that have been previously im-
plicated in cognitive regulation of emotional responses,
including the DMPFC and the left OFC. This overlap in
activations is consistent with previous proposals that emo-
tion regulation recruits certain core processes involved in
cognitively reinterpreting an event, regardless of the re-
appraisal goal (Ochsner et al., 2004). The dorsal sector of
the MPFC, the superior prefrontal gyrus (Ochsner et al.,
2004) or pre-SMA (Picard & Strick, 1996), has been im-
plicated in maintaining spatial (D’Esposito et al., 1998;
Petit, Courtney, Ungerleider, & Haxby, 1998; Goldberg,
Berman, Randolph, Gold, & Weinberg, 1996) and non-
spatial
information during the delay for a response
(D’Esposito et al., 1998; Petit et al., 1998; Braver et al.,
1997; Jonides et al., 1997; Goldberg et al., 1996; Smith,
Jonides, & Koeppe, 1996). Accordingly, activation in this
region during active regulation may reflect the mainte-
nance of regulation strategies throughout each trial. The
cognitive division of the anterior cingulate (Bush, Luu, &
Posner, 2000) was activated for both increasing and de-
creasing negative emotion. This region has been implicat-
ed in the monitoring of ongoing responses (Botvinick,
Braver, Barch, Carter, & Cohen, 2001), suggesting that
activity in this region may reflect monitoring internal and
external emotional responses required for accurate feed-
back relevant to current regulatory goals.
The left orbito-frontal region was also commonly
activated for both increasing and decreasing emotion.
The OFC has been implicated in the down-regulation
of negative emotion such as aggression and violence
(Davidson et al., 2002; Gur, Gunning-Dixon, Biker, &
Gur, 2002; Davidson, Putnam, & Larson, 2000), and in-
creased right lateral OFC activity has been associated
with down-regulating rather than up-regulating negative
affect (Ochsner et al., 2004). The left lateral prefrontal
area (BA 46) was also commonly activated for both
decreasing and increasing negative emotion. Activity in
this region has been reported to be inversely correlated
with the activity in the emotion-processing regions such
as the amygdala and the MOFC (Ochsner et al., 2002),
suggesting that this region has a modulatory role in
down-regulating emotion.
Differences between Up-regulation
and Down-regulation
Ochsner et al. (2004) found that up-regulation of nega-
tive emotion engaged primarily left-lateralized prefrontal
regions, whereas down-regulation engaged primarily
bilateral prefrontal regions. This was interpreted as most
likely reflecting greater self-generation of affective de-
scriptors to increase emotion and greater recruitment
during down-regulation of right prefrontal regions in-
volved in mediating interference between prepotent re-
sponses and cognitive reappraisals. The current results
for regulation of negative and positive emotion are
generally consistent with these prior findings. Relative
to the watch condition, up-regulation for both nega-
tive and positive stimuli was associated with primarily
left prefrontal activation, whereas down-regulation
was associated with a bilateral pattern of activation.
This consistent pattern suggests that up-regulation and
down-regulation recruit specific common cognitive pro-
cesses independent from the type of emotion that is
regulated.
A consistent pattern was observed when activations
unique to up-regulation versus activations unique to
down-regulation were compared. For negative emotion,
although up-regulation activated several regions when
compared to the watch condition (Figure 3, Table 6),
none of these regions were significantly more active
during up-regulation than down-regulation. However,
several regions were uniquely activated during down-
regulation for negative emotion, including bilateral OFC
and other right PFC and right parietal regions (Table 10).
The opposite pattern was observed for positive emo-
tion. No regions were significantly more active during
down-regulation than up-regulation, whereas several
regions were uniquely activated during up-regulation
for positive emotion, including the medial orbito-frontal
gyrus, additional medial and left prefrontal regions, as
well as the anterior cingulate, thalamus, and caudate
(Table 11).
These patterns were similar to those found in the
direct comparison between activations associated with
regulation of positive and negative emotion (Tables 5
and 8). Several regions were identified that were more
active during the down-regulation of negative emotion
794
Journal of Cognitive Neuroscience
Volume 19, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
.
t
f
o
n
1
8
M
a
y
2
0
2
1
versus the down-regulation of positive emotion, but no
regions were significantly more active for positive emo-
tion in the reverse contrast. Similarly, several regions
were more active during the up-regulation of positive
emotion versus the up-regulation of negative emotion,
but only a single, small cluster in the PFC was more
active in the reverse contrast. This same pattern was
observed when possible confounds between positive
and negative stimuli, including arousal ratings and suc-
cess ratings, were included as covariates in the analysis.
Activations unique to positive up-regulation were all
left-lateralized or medial, with the exception of the right
caudate, whereas activations unique to negative down-
regulation were all right-lateralized, with the exception
of OFC activation, which was bilateral. This lateralization
may reflect the overall pattern for left-lateralized activa-
tion for up-regulation and right or bilateral activation for
down-regulation, although it is also consistent with
proposed hemispheric asymmetries in emotion process-
ing that posit a greater role for the right hemisphere in
processing of negative affect (Davidson et al., 2002;
Heilman & Gilmore, 1998; Davidson, 1995). This same pat-
tern of left-lateralized activations unique to up-regulation
and right-lateralized activation unique to down-regulation
was also observed by Ochsner et al. (2004) for negative
emotion regulation, further suggesting that the laterali-
zation reflects up- versus down-regulation rather than
valence-specific effects. This previous study observed
increase-specific activations for negative emotion in the
left rostral medial prefrontal (BA 9/10) and posterior
cingulate cortex (BA 23). Both of these increase-specific
regions were identified for positive emotion in the cur-
rent study, although increase-specific activations for neg-
ative emotion were not observed. The lack of unique
activation in these regions for negative emotion was
attributable here to left rostral PFC activation in both
the increase > watch and the decrease > watch contrasts
and the absence of posterior cingulate activation in
either contrast.
Whereas the dorsal sector of the MPFC was activated
commonly for all types of regulation tasks regardless of
emotion type, the ventral part of the MPFC (BA 10), a
region associated with self-referential processing and
evaluation of internally generated information (Kelly,
Macrae, Wyland, Inati, & Heatherton, 2002; Craik et al.,
1999), and perceived similarity between self and others
(mitchell, Banaji, & Macrae, 2005), was uniquely activat-
ed while increasing positive emotion but not for any
other types of regulation. This activation may reflect
increased self-referential processing while subjects up-
regulated positive emotion by imagining the scenes as
more personally relevant.
In addition to regions exhibiting differential activity as
a function of regulation condition, correlations between
changes in on-line arousal ratings and regional brain
activity revealed several regions whose activity tracked
changes in subjective arousal (Tables 13 y 14), incluir-
ing prefrontal regions previously implicated in emotion
regulación (Ochsner et al., 2004). Notablemente, activity in
the insula was correlated with decreasing negative af-
fect but not with changes in positive affect, consistent
with the predicted specific relation of this region to
negative affect (Mesa 13). Activity in the right LOFC
(BA 47) was correlated with changes in emotional in-
tensity due to voluntary emotion regulation for all
contrasts where significant neural correlates were ob-
served (no significant correlations were observed for
the increase negative (cid:2) watch negative contrast). El
regulation-related activity observed in the right OFC
related to changes in self-reported on-line arousal may
reflect subjective awareness of emotional states and
their intensity (Schirmer & Kotz, 2006).
Modulation of Amygdala Activity by Regulation
As predicted, voluntary emotion regulation modulated
activity in the amygdala elicited by both negative and
positive pictures, indicating that emotion regulation not
only affected the subjectively reported experience of
emotion but also activity in a key region mediating
emotion processing. This pattern was more pronounced
for positive pictures than for negative pictures. Para
positive pictures, activity in both the left and the right
amygdala ROIs increased during the increase condition,
and activity in the right amygdala ROI decreased during
the decrease condition, compared to the watch condi-
ción. This is the first demonstration, a nuestro conocimiento,
that emotion regulation can both increase and decrease
activity in the amygdala for positive emotional stimuli,
paralleling previous findings for regulation-related mod-
ulation of amygdala activity elicited by negative emotion
estímulos (Ochsner et al., 2004).
The current findings are consistent with those of
Beauregard et al. (2001) who reported that down-
regulation of sexual arousal to visual sexually arousing
stimuli was associated with decreased activity in the
right amygdala in men, relative to viewing these stimuli
without engaging in voluntary emotion regulation. El
ROI results extend these previous findings by demon-
strating that down-regulation decreases right amygdala
activity for a wide variety of affectively positive visual
estímulos, and that up-regulation can increase activity in
this structure. The lack of a decrease in left amygdala
activity for positive pictures associated with down-
regulation in the current study is likely related to the
absence of significant activity in the left amygdala for
positive stimuli during the watch condition. Activity in
the left amygdala ROI showed a trend ( pag < .07) of
increasing from the decrease condition to the increase
condition for negative pictures. Although the modula-
tion effect for negative pictures was smaller in magni-
tude than that observed by Ochsner et al. (2004), the
left-lateralization of the current finding is consistent with
the increase in left amygdala activation associated with
Kim and Hamann
795
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
t
.
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
up-regulation and a left-lateralized decrease in amygdala
activation during down-regulation. As previously noted,
negative pictures were rated higher on arousal than
positive pictures, possibly contributing to greater diffi-
culty in regulating reactions to negative pictures and
weaker modulation effects in the amygdala. In addition,
reactions to positive emotion stimuli may be intrinsically
more malleable than reactions to negative emotion
stimuli, possibly because negative emotion stimuli are
typically associated with a greater degree of automatic
processing (Morris, Ohman, & Dolan, 1999; Whalen
et al., 1998).
Modulation of Ventral Striatum Activity
by Positive Emotion Regulation
Consistent with previous reports demonstrating a role
of the ventral striatum in reward processes (Montague
& Berns, 2002) and in experiencing positive emotions
(Hamann & Mao, 2002), activity in the ventral striatum
showed a pattern of modulation consistent with regu-
lation goals for positive emotion. When regulating pos-
itive emotion, up-regulation resulted in an increase in
left and right ventral striatum activity, whereas down-
regulation resulted in a nonsignificant decrease in activ-
ity in the left ventral striatum ROI, compared to the
watch condition. When activity during up- and down-
regulation of positive emotion was directly compared,
activity in the ventral striatum was significantly greater
during up-regulation. These results suggest that the ven-
tral striatum is a modulatory target for the up-regulation
of positive emotion, but leave open the question of
whether down-regulation of positive emotion can mod-
ulate activity in the ventral striatum.
Future Directions and Conclusion
Only women were included in the current study because
of evidence that women typically experience emotion
more intensely (Fujita, Diener, & Sandvik, 1991), to
increase comparability with the results from previous
studies that have studied only women, and to avoid
potential sex differences that might have tended to
obscure the valence and regulation effects that were of
primary interest. Additional study is needed to charac-
terize the extent to which the results observed here also
extend to men. Individual differences in genotype,
personality, and other factors are increasingly being rec-
ognized as important influences on neural reactivity to
emotional stimuli (Hamann & Canli, 2004). Correlations
between individual difference measures and the brain
responses of subjects in this study will be detailed in
a separate report (Kim & Hamann,
in preparation).
Regarding clinical implications, a study of how the reg-
ulatory mechanisms examined here are affected in psy-
chological disorders, such as depression and anxiety, can
potentially further our understanding of the basic mech-
anisms underlying these disorders.
Although subjects were given examples of typical
regulation strategies, and most subjects, in fact, used
broadly similar strategies as indicated by postscan re-
ports (e.g., increasing personal relevance), to increase
the ecological validity and to maximize the effectiveness
of the regulation manipulation, we allowed subjects to
use whatever strategy worked best for them. Variability
in strategies used may have influenced the current re-
sults, particularly for the down-regulation of positive
emotion, where the most variability in strategy use was
observed. Although self-rated regulation success and
emotional arousal did not differ as function of specific
strategy employed, it remains possible that this factor
influenced patterns of brain activation. Further exami-
nation of how specific strategic factors alter the neural
correlates of up-regulation and down-regulation will help
to better characterize the regions that are commonly
recruited by each type of regulation.
Because subjects were instructed not to increase or
decrease their emotional responses by substituting an
opposite emotion, regulation-related decreases and in-
creases in emotional response most likely reflect de-
creases and increases in emotional arousal relative to
a neutral baseline, rather than a reversal of emotional
valence (e.g., from negative to positive affect). Further
study will be needed to investigate the neural corre-
lates associated with regulation strategies involving re-
appraisal by substituting opposing affective reactions.
We also did not contrast regulation effects between
different subcategories of emotion stimuli within a
valence category, for example, between stimuli that
primarily elicited disgust versus fear. A recent study re-
ported that the neural correlates of down-regulating
negative emotion differed depending on whether the
stimuli depicted violations of moral rules, an effect
that was observed even after arousal and other relevant
stimulus factors had been equated (Harenski & Hamann,
2006). These findings suggest that the neural corre-
lates of emotion regulation may differ in important
ways according to other salient dimensions of emotional
experience.
In conclusion, our findings indicate that voluntary up-
regulation and down-regulation of positive and negative
emotions engages prefrontal and cingulate regions in-
volved in cognitive control and modulates activity in the
amygdala, a key substrate for emotion processing. Regions
involved in regulation of positive and negative emotion
overlapped, but substantial differences were also found in
cortical and subcortical responses during regulation, both
between negative and positive emotion and between up-
regulation and down-regulation. In summary, these find-
ings help contribute to our understanding of the neural
mechanisms underlying how individuals manage their
own emotions, as they strive to accentuate the positive
and eliminate the negative in their lives.
796
Journal of Cognitive Neuroscience
Volume 19, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
t
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Reprint requests should be sent
to Stephan Hamann,
Psychology Department, 532 Kilgo Circle, NE, Emory Univer-
sity, Atlanta, GA 30322, or via e-mail: shamann@emory.edu.
An integrative review. Review of General Psychology, 2,
271–290.
Gur, R. C., Gunning-Dixon, F., Biker, W. B., & Gur, R. E.
REFERENCES
Anderson, A. K., Christoff, K., Stappen, I., Panitz, D.,
Ghahremani, D. G., Glover, G., et al. (2003). Dissociated
neural representations of intensity and valence in human
olfaction. Nature Neuroscience, 6, 196–202.
Ashburner, J., & Friston, K. J. (1999). Nonlinear spatial
normalization using basis functions. Human Brain
Mapping, 7, 254–266.
Beauregard, M., Levesque, J., & Bourgouin, P. (2001).
Neural correlates of conscious self-regulation of emotion.
Journal of Neuroscience, 21, 1–6.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S.,
& Cohen, J. D. (2001). Conflict monitoring and cognitive
control. Psychological Review, 108, 624–652.
Braver, T. S., Cohen, J. D., Nystrom, L. E., Jonides, J., Smith,
E. E., & Noll, D. C. (1997). A parametric study of prefrontal
cortex involvement in human working memory.
Neuroimage, 5, 49–62.
Bush, G., Luu, P., & Posner, M. I. (2000). Cognitive and
emotional influences in anterior cingulate cortex. Trends
in Cognitive Sciences, 4, 215–222.
(2002). Sex differences in temporo-limbic and frontal brain
volumes of healthy adults. Cerebral Cortex, 12, 998–1003.
Hamann, S., & Canli, T. (2004). Individual differences in
emotion processing. Current Opinion in Neurobiology,
14, 233–238.
Hamann, S., Ely, T. D., Grafton, S. T., & Kilts, C. D. (1999).
Amygdala activity related to enhanced memory for pleasant
and aversive stimuli. Nature Neuroscience, 2, 289–293.
Hamann, S. B., Ely, T. D., Hoffman, J. M., & Kilts, C. D.
(2002). Ecstasy and agony: Activation of the human
amygdala in positive and negative emotion. Psychological
Science, 13, 135–141.
Hamann, S., & Mao, H. (2002). Positive and negative
emotional verbal stimuli elicit activity in the left amygdala.
Psychological Science, 13, 15–19.
Harenski, C. N., & Hamann, S. (2006). Neural correlates of
regulating negative emotions related to moral violations.
Neuroimage, 313–324.
Heilman, K. M., & Gilmore, R. L. (1998). Cortical influences
in emotion. Journal of Clinical Neurophysiology, 15,
409–423.
Holmes, A. P., & Friston, K. J. (1998). Generalisability, random
effects, and population inference. Neuroimage: Abstracts
of the Fourth International Conference on Functional
Mapping of the Human Brain, 7, S754.
Craik, F. I., Moroz, T. M., Moscovitch, M., Stuss, D. T.,
Holmes, A. P., Josephs, O., Buchel, C., & Friston, K. J.
Winocur, G., Tulving, E., et al. (1999). In search of the
self: A positron emission tomography study. Physiological
Science, 10, 26–34.
Davidson, R. J. (1995). Cerebral asymmetry, emotion, and
affective style. In R. J. Davidson & K. Hugdahl (Eds.),
Brain asymmetry (pp. 361–387). Cambridge: MIT Press.
Davidson, R. J., Lewis, D. A., Alloy, L. B., Amaral, D. G.,
Bush, G., Cohen, J. D., et al. (2002). Neural and behavioral
substrates of mood and mood regulation. Biological
Psychiatry, 52, 478–502.
Davidson, R. J., Putnam, K. M., & Larson, C. L. (2000).
Dysfunction in the neural circuitry of emotion regulation—
A possible prelude to violence. Science, 289, 591–594.
D’Esposito, M., Aguirre, G. K., Zarahn, E., Ballard, D.,
Shin, R. K., & Lease, J. (1998). Functional MRI studies of
spatial and nonspatial-working memory. Cognitive Brain
Research, 7, 1–13.
Dolan, R. J., Lane, R. D., Chua, P., & Fletcher, P. (2000).
Dissociable temporal lobe activations during emotional
episodic memory retrieval. Neuroimage, 11, 203–209.
Frijda, N. H. (1986). The emotions. Cambridge, England:
Cambridge University Press.
Friston, K., Holmes, A., Worsley, K., Poline, J., Frith, C.,
& Frackowiak, R. (1995). Statistical parametric maps in
functional imaging: A general linear approach. Human
Brain Mapping, 2, 189–210.
Friston, K. J., Jezzard, P., & Turner, R. (2004). Analysis of
functional MRI time-series. Human Brain Mapping, 1,
153–171.
Fujita, F., Diener, E., & Sandvik, E. (1991). Gender differences
in negative affect and well-being: The case for emotional
intensity. Journal of Personality and Social Psychology,
61, 427–434.
Goldberg, T. E., Berman, K. F., Randolph, C., Gold, J. M.,
& Weinberg, D. R. (1996). Isolating the mnemonic
component in spatial delayed response: A controlled PET
15O-labeled water regional cerebral blood flow study in
normal humans. Neuroimage, 3, 69–78.
Gross, J. J. (1998). The emerging field of emotion regulation:
(1997). Statistical modelling of low frequency confounds
in fMRI. Neuroimage, 5, S480.
Jackson, D., Malmstadt, J., Larson, C. L., & Davidson, R. J.
(2000). Suppression and enhancement of emotional
responses to unpleasant pictures. Psychophysiology, 37,
515–522.
Jonides, J., Schumacher, E. H., Smith, E. E., Lauber, E. J.,
Awh, E., Minoshima, S., et al. (1997). Verbal working
memory load affects regional brain activation as
measured by PET. Journal of Cognitive Neuroscience,
9, 462–475.
Kelly, W. M., Macrae, C. N., Wyland, C. L., Inati, C. S.,
& Heatherton, T. F. (2002). Finding the self? An
event-related fMRI study. Journal of Cognitive
Neuroscience, 14, 785–794.
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (1995).
International Affective Picture System (IAPS): Technical
manual and affective ratings. Gainesville, FL: The Center
for Research in Psychophysiology, University of Florida.
Lazarus, R. S. (1991). Emotion and adaptation. New York:
Oxford University Press.
Mitchell, J. P., Banaji, M. R., & Macrae, C. N. (2005). The
link between social cognition and self-referential thought
in the medial prefrontal cortex. Journal of Cognitive
Neuroscience, 17, 1306–1315.
Montague, P., & Berns, G. (2002). Neural economics and the
biological substrates of valuation. Neuron, 36, 265–284.
Morris, J. S., Ohman, A., & Dolan, R. J. (1999). A subcortical
pathway to the right amygdala mediating ‘‘unseen’’ fear.
Proceedings of the National Academy of Sciences, U.S.A.,
96, 1680–1685.
Ochsner, K. N., Bunge, S. A., Gross, J. J., & Gabrieli, J. D. E.
(2002). Rethinking feelings: An fMRI study of the cognitive
regulation of emotion. Journal of Cognitive Neuroscience,
14, 1215–1229.
Ochsner, K. N., & Gross, J. J. (2005). The cognitive control
of emotion. Trends in Cognitive Sciences, 9, 242–249.
Ochsner, K. N., Ray, R. D., Cooper, J., Robertson, E. R.,
Chopra, S., Gabrieli, J. D. E., et al. (2004). For better
Kim and Hamann
797
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
or for worse: Neural systems supporting the cognitive
down- and up-regulation of negative emotion. Neuroimage,
23, 483–499.
Petit, L., Courtney, S. M., Ungerleider, L. G., & Haxby, J. V.
(1998). Sustained activity in the medial wall during working
memory delays. Journal of Neuroscience, 18, 9429–9437.
Picard, N., & Strick, P. L. (1996). Motor areas of the medial
wall: A review of their location and functional activation.
Cerebral Cortex, 6, 342–353.
Richards, J. M., & Gross, J. J. (2000). Emotion regulation
and memory: The cognitive costs of keeping one’s cool.
Journal of Personality and Social Psychology, 79, 410–424.
Schaefer, S. M., Jackson, D. C., Davidson, R. J., Aguirre,
G. K., Kimberg, D. Y., & Thompson-Schill, S. L. (2002).
Modulation of amygdala activity by the conscious regulation
of negative emotion. Journal of Cognitive Neuroscience,
14, 913–912.
Schirmer, A., & Kotz, S. (2006). Beyond the right hemisphere:
Brain mechanisms mediating vocal emotional processing.
Trends in Cognitive Sciences, 10, 24–30.
Smith, E. E., Jonides, J., & Koeppe, R. A. (1996). Dissociating
verbal and spatial working memory using PET. Cerebral
Cortex, 6, 11–20.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
atlas of the human brain. New York: Thieme.
Whalen, P. J., Rauch, S. L., Etcoff, N. L., McInerney, S. C.,
Lee, M. B., & Jenike, M. A. (1998). Masked presentations
of emotional facial expressions modulate amygdala activity
without explicit knowledge. Journal of Neuroscience,
105, 141–153.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
5
-
7
p
7
d
6
f
/
1
1
9
9
3
6
/
3
5
4
/
2
7
7
o
6
c
/
n
1
2
7
0
5
0
6
7
6
1
8
9
6
/
5
j
7
o
7
c
6
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
5
o
.
n
7
0
7
8
6
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
f
.
.
t
.
.
o
n
1
8
M
a
y
2
0
2
1
798
Journal of Cognitive Neuroscience
Volume 19, Number 5